
Unit IV- Animal Physiology 1. Comparative muscle structure ...
24
Unit IV- Animal Physiology 1. Comparative muscle structure and function Different Types, Different Functions Muscle tissue is a soft tissue, and is one of the four fundamental types of tissue present in animals. There are three types of muscle tissue recognized in vertebrates. Skeletal muscle, or voluntary muscle, is anchored to bone by tendons, or by aponeuroses at a few places, and is used to effect skeletal movement in activities such as locomotion and maintaining posture. Though this postural control is generally maintained as an unconscious reflex, the muscles responsible react to conscious control like non-postural muscles. An average adult male is made up of 42% of skeletal muscle and an average adult female is made up of 36%, expressed as a percentage of body mass. Figure 1: Muscle Types. Cardiac and skeletal muscle are both striated in appearance, while smooth muscle is not. Both cardiac and smooth muscle are involuntary while skeletal muscle is voluntary. Smooth muscle, or involuntary muscle, is found within the walls of organs and structures such as the esophagus, stomach, intestines, bronchi, uterus, urethra, bladder, blood vessels, and the arrector pili in the skin, in which it controls the erection of body hair. Unlike skeletal muscle, smooth muscle is not under conscious control. Cardiac muscle is also an involuntary muscle but is more akin in structure to skeletal muscle, and is found only in the heart. Cardiac and skeletal muscles are striated, in that they contain sarcomeres and are packed into highly regular, repeating arrangements of bundles; smooth muscle has neither attribute. While skeletal muscles are arranged in regular, parallel bundles, cardiac muscle connects at branching, irregular angles, called intercalated discs. Striated muscle contracts and relaxes in short, intense bursts, whereas smooth muscle sustains longer or even near-permanent contractions. The density of mammalian skeletal muscle tissue is about 1.06 kg/liter. This can be contrasted with the density of adipose tissue (fat), which is 0.9196 kg/liter. This makes muscle tissue approximately 15% denser than fat tissue. Skeletal muscles are voluntary, striated muscles that allow movement of an organism by the deliberate generation of force . The action a skeletal muscle generates is determined by the origin and insertion locations. The cross-sectional area of a muscle (rather than volume or length) determines the amount of force it can generate by defining the number of sarcomeres which can operate in parallel. The amount of force applied to the external environment is
Transcript of Unit IV- Animal Physiology 1. Comparative muscle structure ...

Unit IV- Animal Physiology
1. Comparative muscle structure and function
Different Types, Different Functions
Muscle tissue is a soft tissue, and is one of the four fundamental types of tissue present in
animals. There are three types of muscle tissue recognized in vertebrates. Skeletal
muscle, or voluntary muscle, is anchored to bone by tendons, or by aponeuroses at a few
places, and is used to effect skeletal movement in activities such as locomotion and maintaining
posture. Though this postural control is generally maintained as an unconscious reflex, the
muscles responsible react to conscious control like non-postural muscles. An average adult
male is made up of 42% of skeletal muscle and an average adult female is made up of 36%,
expressed as a percentage of body mass.
Figure 1: Muscle Types. Cardiac and skeletal muscle are both striated in appearance, while
smooth muscle is not. Both cardiac and smooth muscle are involuntary while skeletal muscle
is voluntary.
Smooth muscle, or involuntary muscle, is found within the walls of organs and structures
such as the esophagus, stomach, intestines, bronchi, uterus, urethra, bladder, blood vessels, and
the arrector pili in the skin, in which it controls the erection of body hair. Unlike skeletal
muscle, smooth muscle is not under conscious control. Cardiac muscle is also an involuntary
muscle but is more akin in structure to skeletal muscle, and is found only in the heart.
Cardiac and skeletal muscles are striated, in that they contain sarcomeres and are packed into
highly regular, repeating arrangements of bundles; smooth muscle has neither attribute. While
skeletal muscles are arranged in regular, parallel bundles, cardiac muscle connects at
branching, irregular angles, called intercalated discs. Striated muscle contracts and relaxes in
short, intense bursts, whereas smooth muscle sustains longer or even near-permanent
contractions.
The density of mammalian skeletal muscle tissue is about 1.06 kg/liter. This can be contrasted
with the density of adipose tissue (fat), which is 0.9196 kg/liter. This makes muscle tissue
approximately 15% denser than fat tissue.
Skeletal muscles are voluntary, striated muscles that allow movement of an organism by the
deliberate generation of force . The action a skeletal muscle generates is determined by the
origin and insertion locations. The cross-sectional area of a muscle (rather than volume or
length) determines the amount of force it can generate by defining the number of sarcomeres
which can operate in parallel. The amount of force applied to the external environment is

determined by lever mechanics, specifically the ratio of in lever to out lever. For example,
moving the insertion point of the biceps more distally on the radius (farther from the joint of
rotation) would increase the force generated during flexion (and, as a result, the maximum
weight lifted in this movement), but decrease the maximum speed of flexion. Moving the
insertion point proximally (closer to the joint of rotation) would result in decreased force but
increased velocity. This can be most easily seen by comparing the limb of a mole to a horse:
In the former, the insertion point is positioned to maximize force (for digging), while in the
latter, the insertion point is positioned to maximize speed (for running).
In addition to voluntary contractions of skeletal muscle, involuntary muscle also contracts in a
similar fashion but does so involuntarily. Smooth muscle is responsible for movement of food
through the digestive system via peristalsis and regulates the diameter of blood vessels,
determining how much blood flows through the vessels. Cardiac muscle is responsible for
contraction of the heart . The contraction of cardiac muscle of the heart is coordinated such that
the entire heart beats in a controlled, uniform manner, ensuring blood is efficiently pumped
from the chambers. Cardiac and smooth muscle contraction occurs without conscious thought
and is necessary for survival.

Cardiac muscle is striated, similar to skeletal muscle, but beats involuntarily. The cells
beat in unison as a result of unique gap junctions between the muscle cells.
Diagram of contraction of smooth muscle fiber
showing peristalsis
Skeletal muscle fibers can be characterized by their metabolic processes and corresponding
physiological traits.
Signaling Pathways that Regulate Skeletal Muscle Fiber-type Phenotype
Exercise-induced signaling pathways in skeletal muscle that determine specialized
characteristics of slow-twitch and fast-twitch muscle fibers.
Oxidative fibers rely on aerobic respiration to fuel muscle contractions, and consist of slow-
twitch (Type I) fibers, which are characterized as muscles with long contraction duration,
associated with endurance. Slow-twitch fibers are used to maintain posture. They are usually
found in red muscles, indicative of the large concentration of myoglobin providing a steady
supply of oxygen to them. The red muscles use oxidative phosphorylation to obtain ATP.
Oxidative phosphorylation occurs in the red muscles as the process requires a lot of oxygen,
and the red muscles contain high amounts of myoglobin. The process is slower than glycolysis,
but much more efficient, which is why slow-twitch muscles do not tire easily. Also, slow-
twitch fibers contain less sarcoplasmic reticulum, facilitating a slower release of calcium,
regulating muscle contraction at slower rates.

Glycolytic fibers rely on glycolysis to fuel muscle contractions and consist of fast-twitch (Type
II) fibers, which are characterized by fast muscle contractions of short duration. Fast-twitch
fibers are constituents of white muscles and have less myoglobin due to their primary reliance
on glycolysis (anaerobic respiration) to fuel muscle contractions. Although glycolysis is very
quick, it is also inefficient at producing ATP. Glycolysis produces lactic acid as a byproduct,
which leads to fatigue. The use of the glycogen cycle is the reason why fast-twitch muscles tire
out quickly.
There is some evidence that the proportion of fast-twitch versus slow-twitch muscles of an
individual is partly genetic in nature. That is, we are born with a unique proportion of such
muscles that suit us to particular types of physical activity. This is not without debate, however.
Regardless, concentrated exercise that prioritizes one type of muscle fiber use over the other,
can lead to muscle hypertrophy (increase in size), improving an individual’s ability to perform
related physical activities.
Muscle fiber generates tension through the action of actin and myosin cross-bridge cycling.
While under tension, the muscle may lengthen, shorten, or remain the same. Although the term
contraction implies shortening, when referring to the muscular system, it means muscle fibers
generating tension with the help of motor neurons. Several types of muscle contractions occur
and they are defined by the changes in the length of the muscle during contraction.
Isotonic Contractions
Isotonic contractions maintain constant tension in the muscle as the muscle changes length.
This can occur only when a muscle’s maximal force of contraction exceeds the total load on
the muscle. Isotonic muscle contractions can be either concentric (muscle shortens) or eccentric
(muscle lengthens).
Concentric Contractions
A concentric contraction is a type of muscle contraction in which the muscles shorten while
generating force. This is typical of muscles that contract due to the sliding filament mechanism,
and it occurs throughout the muscle. Such contractions also alter the angle of the joints to which
the muscles are attached, as they are stimulated to contract according to the sliding filament
mechanism.
This occurs throughout the length of the muscle, generating force at the musculo-tendinous
junction; causing the muscle to shorten and the angle of the joint to change. For instance, a
concentric contraction of the biceps would cause the arm to bend at the elbow as the hand
moves from near to the leg to close to the shoulder (a biceps curl). A concentric contraction of
the triceps would change the angle of the joint in the opposite direction, straightening the arm
and moving the hand toward the leg.
Eccentric Contractions
An eccentric contraction results in the elongation of a muscle. Such contractions decelerate the
muscle joints (acting as “brakes” to concentric contractions) and can alter the position of the
load force. These contractions can be both voluntary and involuntary. During an eccentric
contraction, the muscle elongates while under tension due to an opposing force which is greater
than the force generated by the muscle. Rather than working to pull a joint in the direction of
the muscle contraction, the muscle acts to decelerate the joint at the end of a movement or
otherwise control the repositioning of a load.
This can occur involuntarily (when attempting to move a weight too heavy for the muscle to
lift) or voluntarily (when the muscle is “smoothing out” a movement). Over the short-term,
strength training involving both eccentric and concentric contractions appear to increase
muscular strength more than training with concentric contractions alone.
Isometric Contractions
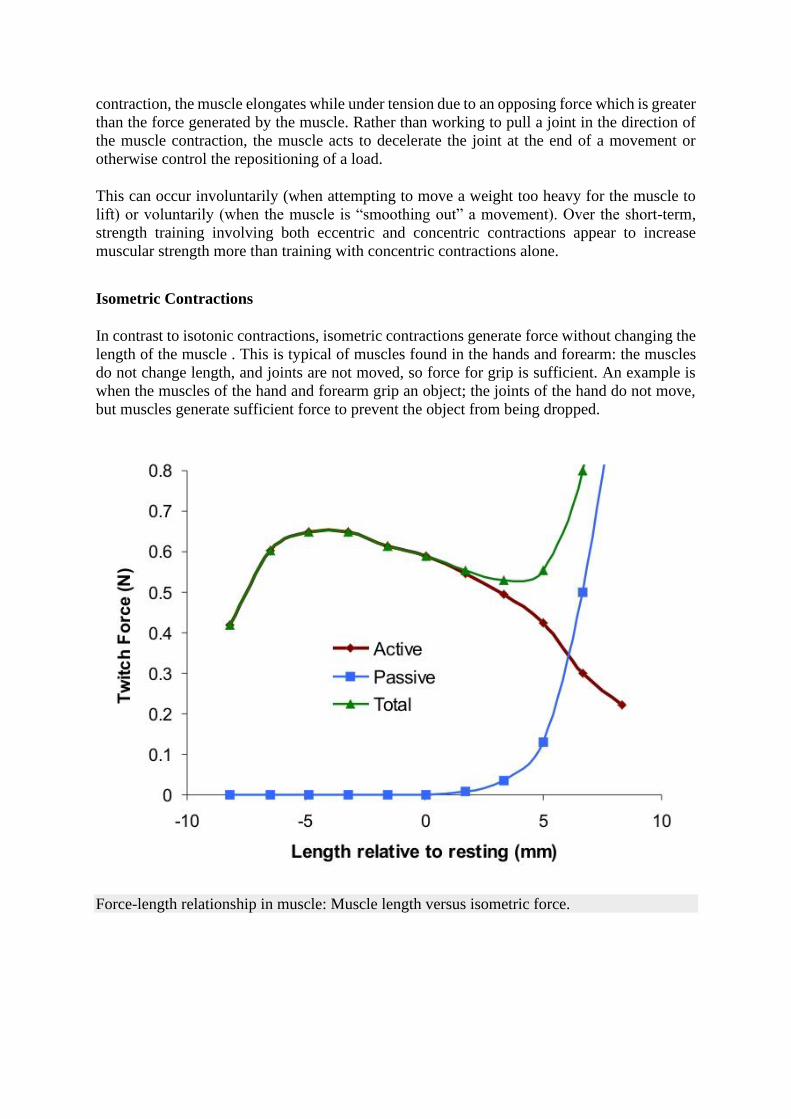
In contrast to isotonic contractions, isometric contractions generate force without changing the
length of the muscle . This is typical of muscles found in the hands and forearm: the muscles
do not change length, and joints are not moved, so force for grip is sufficient. An example is
when the muscles of the hand and forearm grip an object; the joints of the hand do not move,
but muscles generate sufficient force to prevent the object from being dropped.
Force-length relationship in muscle: Muscle length versus isometric force.

2. Contraction of Muscles
Skeletal muscles contract and relax to mechanically move the body. Messages from the nervous
system cause these muscle contractions. The whole process is called the mechanism of muscle
contraction and it can be summarized in three steps:
(1) A message travels from the nervous system to the muscular system, triggering chemical
reactions.
(2) The chemical reactions lead to the muscle fibers reorganizing themselves in a way that
shortens the muscle--that’s the contraction.
(3) When the nervous system signal is no longer present, the chemical process reverses, and
the muscle fibers rearrange again and the muscle relaxes.
Let’s look a little more closely at the steps in the mechanism of muscle contraction.
1. A Muscle Contraction Is Triggered When an Action Potential Travels Along the Nerves
to the Muscles
Muscle contraction begins when the nervous system generates a signal. The signal, an impulse
called an action potential, travels through a type of nerve cell called a motor neuron. The
neuromuscular junction is the name of the place where the motor neuron reaches a muscle cell.
Skeletal muscle tissue is composed of cells called muscle fibers. When the nervous system
signal reaches the neuromuscular junction a chemical message is released by the motor neuron.
The chemical message, a neurotransmitter called acetylcholine, binds to receptors on the
outside of the muscle fiber. That starts a chemical reaction within the muscle.

2. Acetylcholine Is Released and Binds to Receptors on the Muscle Membrane
A multistep molecular process within the muscle fiber begins when acetylcholine binds to
receptors on the muscle fiber membrane. The proteins inside muscle fibers are organized into
long chains that can interact with each other, reorganizing to shorten and relax. When
acetylcholine reaches receptors on the membranes of muscle fibers, membrane channels open
and the process that contracts a relaxed muscle fibers begins:
• Open channels allow an influx of sodium ions into the cytoplasm of the muscle fiber.
• The sodium influx also sends a message within the muscle fiber to trigger the release
of stored calcium ions.
• The calcium ions diffuse into the muscle fiber.
• The relationship between the chains of proteins within the muscle cells changes, leading
to the contraction.
3. Muscle Fibers Relax When the Nervous System Signal Is No Longer Present

When the stimulation of the motor neuron providing the impulse to the muscle fibers stops, the
chemical reaction that causes the rearrangement of the muscle fibers' proteins is stopped. This
reverses the chemical processes in the muscle fibers and the muscle relaxes.

3. Sliding Filament Theory of Muscle Contraction
The mechanism of muscle contraction is explained by sliding filament model. This theory was
proposed by H.E Huxley and J. Hanson, and A. F. Huxley and R. Niedergerke in 1954.
The arrangement of actin and myosin myofilament within a sarcomere is crucial in the
mechanism of muscle contraction. It is proposed that muscle contracts by the actin and myosin
filaments sliding past each other. For analogy, muscle contraction by sliding filament model is
equivalent to interlocking fingers, pushing them together shortens the distance.
As sarcomere is the unit of muscle contraction, its length contracts resulting in whole muscle
contraction. During contraction, length of A-band (Dark band) remains same whereas length
of I-band (Light band) and H-zone gets shorter.
Actin myofilament:
▪ An actin myofilament is made up of actin molecule, tropomyosin and troponin
complex. Troponin is composed of three sub-units (troponin I, T and
C). Tropomyosin form two helical strand which are wrapped around actin
molecules (G-actins) longitudinally in thin twisted stranded form.
▪ Each G-actin is attached with an ATP molecule. The whole assembly of actin
molecules is known as F-actin (Fibrous actin).
▪ Tropomyosin switches ON or OFF the muscle contraction mechanism.
▪ Troponin complex is a globular protein which binds to tropomyosin and calcium
ions.
Myosin myofilament:
▪ A myosin myofilament consists of two distinct region, a long rod-shaped tail
called myosin rod and two globular intertwined myosin head.
▪ The globular head appear at interval along the myosin myofilament, projecting
from the sides of the filament.
▪ The myosin head can attach to the neighboring acting filament where actin and
myosin filaments overlaps.

source:www.crossfitinvictus.com
Figure: Actin and Myosin myofilament
Mechanism of Muscle contraction:
▪ When the nerve impulse from brain and spinal cord are carried along motor
neuron to neuromuscular junction, Ca++ ions are released in the terminal axon.
Increases calcium ion concentration stimulates the release of neurotransmitter
(Acetylcholine) in the synaptic cleft. The neurotransmitter binds to the receptor
on the sarcolemma and depolarization and generate action potential across
muscle fiber for muscle contraction. The action potential propagates over entire
muscle fiber and move to the adjacent fibers along transverse tubules. The action
potential in transverse tubules causes the release of calcium ion from
sarcoplasmic reticulum, which stimulate for muscle contraction.
The sequences of muscle contraction explained by sliding filament theory are as follows
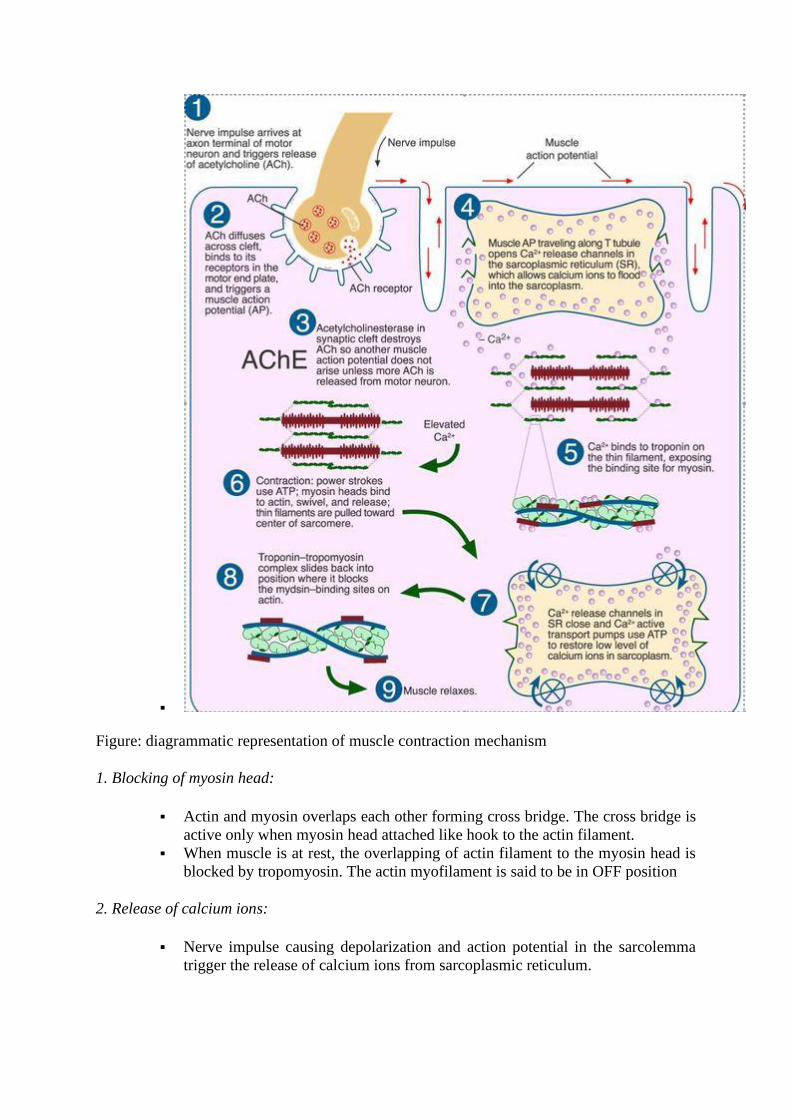
▪
Figure: diagrammatic representation of muscle contraction mechanism
1. Blocking of myosin head:
▪ Actin and myosin overlaps each other forming cross bridge. The cross bridge is
active only when myosin head attached like hook to the actin filament.
▪ When muscle is at rest, the overlapping of actin filament to the myosin head is
blocked by tropomyosin. The actin myofilament is said to be in OFF position
2. Release of calcium ions:
▪ Nerve impulse causing depolarization and action potential in the sarcolemma
trigger the release of calcium ions from sarcoplasmic reticulum.

▪ The calcium ion then binds with the troponin complex on the actin myofilament
causing displacement of troponin complex and tropomyosin from its blocking
site exposing myosin binding site.
▪ As soon as the myosin binding site is exposed, myosin head cross bridge with
actin filament. Now, the actin myofilament is said to be in ON position.
3. Active Cross-bridge formation:
▪ When myosin head attached like hooks to the neighboring actin filament, active
cross bridge is formed. The cross bridge between actin and myosin filament acts
as an enzyme (Myosin ATPase).
▪ The enzyme Myosin ATpase hydrolyses ATP stored into ADP and inorganic
phosphate and release energy. This released energy is used for movement of
myosin head toward actin filament. The myosin head tilts and pull actin filament
along so that myosin and actin filament slide each other. The opposite end of
actin myofilament within a sarcomere move toward each other, resulting in
muscle contraction.
▪ After sliding the cross bridge detached and the actin and myosin filament come
back to original position. The active cross bridge form and reform for 50-100
time within a second using ATP in rapid fashion. Therefore, muscle fiber consists
of numerous mitochondria.
▪ In muscle contraction, sarcomere can contracts by 30-60% of its length

4. Significance of Membrane potentials
Membrane potential (also transmembrane potential or membrane voltage) is the
difference in electric potential between the interior and the exterior of a biological cell. For the
exterior of the cell, typical values of membrane potential, normally given in units
of millivolts and denoted as mV, range from –40 mV to –80 mV.
All animal cells are surrounded by a membrane composed of a lipid
bilayer with proteins embedded in it. The membrane serves as both an insulator and a diffusion
barrier to the movement of ions. Transmembrane proteins, also known as ion transporter or ion
pump proteins, actively push ions across the membrane and establish concentration gradients
across the membrane, and ion channels allow ions to move across the membrane down those
concentration gradients. Ion pumps and ion channels are electrically equivalent to a set
of batteries and resistors inserted in the membrane, and therefore create a voltage between the
two sides of the membrane.
Almost all plasma membranes have an electrical potential across them, with the inside usually
negative with respect to the outside.[1] The membrane potential has two basic functions. First,
it allows a cell to function as a battery, providing power to operate a variety of "molecular
devices" embedded in the membrane.[2] Second, in electrically excitable cells such
as neurons and muscle cells, it is used for transmitting signals between different parts of a cell.
Signals are generated by opening or closing of ion channels at one point in the membrane,
producing a local change in the membrane potential. This change in the electric field can be
quickly affected by either adjacent or more distant ion channels in the membrane. Those ion
channels can then open or close as a result of the potential change, reproducing the signal.
In non-excitable cells, and in excitable cells in their baseline states, the membrane potential is
held at a relatively stable value, called the resting potential. For neurons, typical values of the
resting potential range from –70 to –80 millivolts; that is, the interior of a cell has a negative
baseline voltage of a bit less than one-tenth of a volt. The opening and closing of ion channels
can induce a departure from the resting potential. This is called a depolarization if the interior
voltage becomes less negative (say from –70 mV to –60 mV), or a hyperpolarization if the
interior voltage becomes more negative (say from –70 mV to –80 mV). In excitable cells, a
sufficiently large depolarization can evoke an action potential, in which the membrane
potential changes rapidly and significantly for a short time (on the order of 1 to 100
milliseconds), often reversing its polarity. Action potentials are generated by the activation of
certain voltage-gated ion channels.
In neurons, the factors that influence the membrane potential are diverse. They include
numerous types of ion channels, some of which are chemically gated and some of which are
voltage-gated. Because voltage-gated ion channels are controlled by the membrane potential,
while the membrane potential itself is influenced by these same ion channels, feedback loops
that allow for complex temporal dynamics arise, including oscillations and regenerative events
such as action potentials.

5. Nerve impulse Propagation
Definition of Nerve Impulse:
The electrochemical wave that travels along nerve fibre and stimulates muscles, glands or other
nerve cells is called nerve impulse.
Typical structure of neuron:
Neuron is the structural and functional unit of nervous system. It consists of a nerve cell body
or soma and two types of processes-axon and dendrite (Fig. 8.29).
Soma:
It is an irregular-shaped structure in the centre of which there lies a spherical nucleus with
prominent nucleolus and Nissl granules. It also contains mitochondria, Golgi body, ribosome
and ER etc.
Dendrite:
It is the process of the cell body that carries impulse towards the cell body. It is usually short
with many branches and contains Nissl granules.
Axon:
It is the process of a nerve cell body that carries impulse away from it. It is usually single, long
slender process and sometimes branched and contains axoplasm, neuro-fibril, etc. It terminates
into branches with terminal buttons.
Classification of stimulus:
(a) Subliminal stimulus:
Which can produce the local excitatory state (LES) only?
(b) Threshold value of stimulus:
Which can produce or transform the LES to action potential?

Propagation of Nerve Impulse:
The propagation of nerve impulse involves two major parts – A. Origin/stimulation of nerve
impulse, B. Propagation/ travelling of nerve impulse.
A. Origin of Nerve impulse:
In resting nerve cells, the surface is positively charged and the interior is negatively charged.
When the surface is stimulated the stimulated point becomes negative. The fluids both inside
and outside the cell are electrolytic solution containing 150-160m Eq/litre.
Positive ions and negative ions are accumulated along the outer and inner surface of the cell
membrane, respectively. This is achieved by Na+ outside and K+ inside the cell membrane, and
because Na+ is placed above the K+ in the electrochemical series.
Development of local excitatory state and Development of action potential:
When stimulation is applied on the nerve cell, external Na+ rushes inside the cell making the
inner surface positively charged. The amount of Na+ is not sufficient to generate action
potential and returns to the outside immediately, causing a closed circuit.
Resting potential:
In resting state the nerve fibres remain in polarised state and the membrane potential lies within
-70 mv. Na+ concentration outside the membrane is higher than that of inside and
K+ concentration inside the membrane is higher than that of outside. K+ can permeate through
the membrane at resting condition but the Na+ cannot permeate.
State of De-polarisation:
Permeability of Na+ to membrane is increased only after stimulation; causing de-polarisation.
The tremendous increase in Na+ conductance during this period is known as activation of
membrane. After an initial slow rise, de-polarisation wave overshoots rapidly and can reach
the iso-potential line (above zero line) to approximately +35 mv (Fig. 8.30).

State of re-polarisation:
After reaching the iso-potential line K+ begins to come out from inside the membrane, causing
outside to be positive again. This is called re-polarisation.
Spike potential:
The periodic rise of de-polarisation wave and rapid fall of re-polarisation wave are known as
spike potential.
Negative after potential:
At approximately 2/3rd of the polarisation, the rate of fall is being abruptly slowed down. This
slower fall is known as negative after potential.
Positive after potential:
With the disappearance of the negative after potential, although the rising membrane potential
is achieved, yet the resting ionic status is not established. It is achieved by the active Na+ pump
mechanism, which causes the positive after potential. At the same time K+ travels back to the
inside of the membrane.
B. Propagation/travelling of nerve impulse:
1. Propagation on non-medulated nerve fibre:
According to the membrane theory, nerve impulse is a propagated wave of de-polarisation.
i) When the fibre is excited at a point, the polarity is reversed. This reversed polarity is due to
increased permeability of Na+ to the membrane, which develops de-polarisation wave.
ii) A local circuit current flows between the de-polarised membrane and the resting membrane
areas.
iii) Positive current flows inward through the de-polarised membrane and outward through the
resting membrane and in this way circuit is completed.

iv) The local de-polarisation current then exits the adjacent portion of the membrane
progressively more and more de-polarisation.
v) The de-polarisation wave travels in all direction along the entire length of the nerve fibre.
2. Propagation in myelinated nerve fibre: Salutatory conduction:
In the myelinated nerve fibre conduction depends upon the similar pattern of circular current
flow. Myelin sheath is an effective insulator. Ions cannot pass along the myelin sheaths but
nodes of Ranvier permeate ions through it more easily. For this reason the impulse is
transmitted from node to node rather than continuously along the entire of the nerve fibre (Fig.
8.32).
The de-polarisation in myelinated action jumps from one node of Ranvier to the next. This
jumping or leaping of de-polarisation from node to node is known as saltatory conduction.

Rate of Conduction of Nerve Impulse:
The basic principle of origin and propagation of nerve impulse is same, both in non-medulated
and myelinated nerve fibres but the saltatory mechanism of conduction in myelinated nerve
fibre increases the velocity of conduction more than 500 times.
The rate of conduction of a nerve impulse increases with an increase in the cross sectional
diameter of the neuron and with increasing thickness of the myelin sheath. The rate of trans-
mission for a given neuron is a constant. Table 8.8 gives an idea about rate of nerve impulse
conduction through different nerves of various animals.

6. Synoptic transmission Introduction
Humans and other vertebrates have developed a highly efficient system of communication, the
nervous system. It is a network of interconnected neurons that are capable of generating and
transmitting nerve impulses. The communication process is based on the successful
transmission of nerve impulses from one neuron to the other.
Neurons are connected via specialized structures called synapses. The transmission of a nerve
impulse or action potential from one neuron to another neuron or non-neuron cell, across the
synapse, is called synaptic transmission.
The process of synaptic transmission can be easily understood after studying the structure and
the type of synapses involved. In this article, we will discuss the structure and types of
synapses, the process of synaptic transmission, the role of neurotransmitters, the effects of
drugs, and clinical conditions associated with the synaptic transmission.
Types of Synapses
Not all the synapses found in the body are the same. Based on the mode of synaptic
transmission, the synapses found in the human body are divided into two types; chemical
synapses and electrical synapses.
Chemical Synapses
These are the most abundant synapses found in the body. The two neurons are joined via a
synaptic cleft. One of the two cells secretes neurotransmitters into the cleft. These chemical
messengers diffuse through the cleft and act on the receptors present on the other cell.
As chemical messengers i.e. neurotransmitters are used for transmission of nerve impulses
across a synapse, such synapses are called chemical synapses.
The detailed structure of such synapses will be discussed in detail.
Electrical Synapses
In some cases, the two cells are connected via gap junctions. The cytoplasm of these cells is
connected in such a way that ions can freely diffuse among the cells. the action potential
generated in one cell is transmitted to the next cell by the flow of ions.
As ions or electric currents are used for the transmission of action potential among the cells,
such synapses are called electrical synapses. They are found among the cardiac muscles and
some smooth muscle cells.
Structure of a Chemical Synapse

Understanding the structure of a chemical synapse will help us grab the concept of synaptic
transmission.
A chemical synapse has three major components; pre-synaptic terminal, synaptic cleft, and
post-synaptic terminal.
Pre-synaptic Terminal
It is the axon terminal of the pre-synaptic neuron. The axon terminal makes a dilation
called axon bouton. It is connected to the post-synaptic neuron or cell via the synaptic cleft.
The pre-synaptic terminal has multiple adaptations to release neurotransmitter when the action
potential reaches the axon bouton. Neurotransmitters are kept stored in the axonal terminal in
the form of vesicles. Certain calcium channels are present in the plasma membrane. Certain
channels for the release and uptake of neurotransmitters are also present.
Synaptic Cleft
It is the space between the pre-synaptic and post-synaptic cells. the neurotransmitters released
by the pre-synaptic terminal diffuse through this space to act on the post-synaptic cells.
The size of the synaptic cleft is of the order of 20 nm. This small size allows the
neurotransmitters to rapidly pile up and diffuse through the cleft.
This space between the two cells also has certain enzymes that can cause degradation of
neurotransmitters. It helps remove the neurotransmitters when they have done their action.
Post-synaptic Terminal
The post-synaptic terminal may be a neuron or non-neuronal cell. It contains receptors for
neurotransmitters that are linked to some ion channels. The binding of neurotransmitters to
these receptors results in the opening of ion channels, the ion diffuse across the cell, and an
action potential is generated in the post-synaptic cell.
The post-synaptic receptors are usually located in an invagination of the cell membrane called
synaptic gutter or junctional folds.
Process of Synaptic Transmission
The process of synaptic transmission involves two steps; release of neurotransmitter from pre-
synaptic cell and generation of an action potential in the post-synaptic cell.
Release of Neurotransmitter
Neurotransmitters are released by the pre-synaptic cell when an action potential reaches the
terminal. The process is as follows;

• Specialized voltage-gated calcium channels are located in the pre-synaptic
terminal.
• When the action potential reaches the axon terminal, the depolarization of
axolemma (plasma membrane of axons) causes the opening of calcium
channels.
• Calcium ions are present in higher concentrations in the extracellular fluid
surrounding the terminal. The opening of calcium channels causes these ions to
diffuse into the axonal fiber.
• Once inside the axon terminal, calcium ions bind to some specialized proteins
located on the inner surface of the membrane called release sites.
• Binding of calcium ions to these release sites causes the synaptic vesicles to
diffuse with the terminal membrane.
• Synaptic vesicles diffuse and the neurotransmitter molecules present in them
are released into the synaptic cleft.
The number of neurotransmitters released into the cleft is proportional to the number of calcium
ions diffusing into the pre-synaptic terminal.
Generation of Action Potential
The process of synaptic transmission is completed when an action potential is generated in the
post-synaptic cell. It involves the following steps.
• Neurotransmitters diffuse across the synaptic cleft and reach the junctional folds
on the post-synaptic cell.
• Here, the neurotransmitters bind to the membrane receptors and activate them.
• The activation of receptors causes the opening of ion channels.
• Sodium ions present abundantly in the surrounding fluid rapidly enter the cell
down the concentration gradient.
• The diffusion of sodium ions into the cell causes depolarization and a receptor
potential is generated.
• If the receptor potential is greater than the threshold, an action potential is
generated in the post-synaptic cell.
The strength of receptor potential depends on the number of neurotransmitters binding to the
receptors.
Removal of Neurotransmitters
In most of the cases, the number of neurotransmitters released during synaptic transmission is
greater than the requirements. Once an action potential is generated, the excess
neurotransmitters must be removed from the cell. If the extra neurotransmitters are not
removed, they will cause continuous excitation of the post-synaptic cell.
There are two ways to remove extra neurotransmitters;
• Breakdown enzymes are present in the synaptic cleft. They cleave the
neurotransmitters to rapidly decrease their concentration in the synapse. The
breakdown products may be taken up by the pre-synaptic neuron to make new
neurotransmitters.

• Certain protein channels are present in the pre-synaptic terminal. They use ATP
to actively pump the neurotransmitters back into the axon. This also helps to
rapidly decrease the concentration of neurotransmitters.
The reabsorbed neurotransmitters are again packed into the synaptic vesicle and are ready to
be released again during the next cycle.
Neurotransmitters
These are the chemical messengers that are involved in the process of synaptic transmission in
chemical synapses. Their study is necessary to completely understand the synaptic
transmission.
Here we will discuss briefly the synthesis and types of neurotransmitters.
Synthesis
Neurotransmitters are either amino acids or peptides in nature. They are made by the rough
endoplasmic reticulum present in the cell body of the pre-synaptic neuron.
The neurotransmitters are then packed into excretory vesicles called synaptic vesicles. The
synaptic vesicles are formed by the Golgi bodies and are stored in the pre-synaptic terminal.
Types
Not every neurotransmitter released into the synapse causes an action potential in the post-
synaptic cell. Sometimes, they can also block nervous transmission.
Based on their function, there are two types of neurotransmitters.
Excitatory Neurotransmitters
These can cause the excitation of the post-synaptic cells. Binding of such neurotransmitters to
the receptors causes;
• Increased conduction through the sodium channels by opening them
• Decreased conduction through the potassium channels by blocking them
Both these changes cause depolarization of the cell and an action potential is generated.
Examples of excitatory neurotransmitters include norepinephrine, acetylcholine, and
dopamine, etc.
Inhibitory Neurotransmitters
These serve to block synaptic transmission. The following changes take place in a post-synaptic
cell when an inhibitory neurotransmitter binds to its receptors.

• Increased conduction through the potassium channels so that potassium ions
diffuse out of the cell
• Decreased conduction through the sodium ions so that no sodium ion enters the
cell
All these changes make the cell hyperpolarized and no action potential is generated. As a result,
synaptic transmission is blocked.
Examples of inhibitory transmitters include GABA, serotonin, and dopamine in some cases.
Drugs Action
Sometimes, there is a medical need to enhance or suppress synaptic transmission. For example,
in chronic pain conditions, synaptic transmission is blocked to treat pain. Certain drugs act on
synapse and are responsible for modifications in synaptic transmission. Some of these drugs
are as mentioned below.
Curare Drugs
These drugs are used for complete blockage of action potential transmission at some synapses.
They are the acetylcholine antagonists. Curare drugs bind to acetylcholine receptors and block
their activation. The receptors are not stimulated even when abundant acetylcholine is present
in the synaptic cleft.
Some examples of curare drugs are atracurium, pancuronium, and vecuronium, etc. These are
the non-depolarizing muscle reactants, as they prevent the depolarization of post-synaptic cells.
Atracurium is used as a skeletal muscle relaxant during surgery or mechanical ventilation.
Morphine
It is a powerful pain-killer used in extreme situations. Morphine inhibits synaptic transmission
of pain signals by activating meu-receptors.
Meu-receptors cause increased potassium efflux out of the cell and decreased influx of calcium
and sodium ions. The cell becomes hyperpolarized and the synaptic transmission is blocked.
Acetylcholine esterase Inhibitors
Acetylcholine esterase is an enzyme present in the synaptic cleft that cleaves acetylcholine into
choline and acetic acid. The action of this enzyme is necessary to remove excess acetylcholine
and end the transmission process after one cycle.
The inhibitors of this enzyme are used to increase the concentration of acetylcholine in the
synaptic cleft. These drugs are used in Myasthenia gravis, glaucoma, and to increase the
motility of digestive and urinary systems.
Examples of these drugs include physostigmine, neostigmine, rivastigmine, etc.

Strychnine
It is a poisonous drug that blocks synaptic transmission at the motor endplate (synapse between
skeletal muscle fiber and a motor neuron). It blocks the glycine receptors on alpha motor
neurons in the spinal cord. Thus, there is no more inhibitory effect of glycine and uncontrolled
muscle contractions occur. This leads to muscle spasm.
Alcohol
Alcohol plays a role in the transmission of inhibitory synaptic signals. It mimics the action of
inhibitory neurotransmitter GABA, bi binding to GABAA receptors. As a result, the inhibitory
effect of GABA is enhanced. The post-synaptic neuron becomes hyperp[olarized due to this
action.


















