
Unequally distributed electrolytes Role of alkaline and alkaline earth metal ions.
29
Unequally distributed electrolytes Role of alkaline and alkaline earth metal ions
-
Upload
suzanna-may -
Category
Documents
-
view
237 -
download
0
Transcript of Unequally distributed electrolytes Role of alkaline and alkaline earth metal ions.

Unequally distributed electrolytes
Role of alkaline and alkaline earth metal ions

Unequally distributed electrolytes: Function and transport of alkaline and alkaline earth metal cations
Concentration gradient between the cells and their environments:
This gradient can be maintained only by significant energy consumption
This differs from their thermodynamic equilibrium state
Average extracellular and intracellular ion concentrations of mammals in mM
Na+ K+ Mg2+ Ca2+ Cl- HCO3-
HPO42-/
H2PO4-
Inside the cellOutside the cell
150100
5140
22.5
20.0001
11010
3010
44

Chemical characteristics of the ions
Do not form strong (corrdinative) bonds.
They are present in hydrated form - diffusion.
They are present in high concentration.
Physical data of the biologically important alkaline and alkaline-earth metal ions
Na+ K+ Mg2+ Ca2+
Ionic radius (Å, N=6)Relative surface charge densityCoordination numberCoordination geometryPreferred donor atomMobility in biological systemsWater exchange rate (s-1)
1.020.256-8
variousO
highly mobile10-10
1.380.146-8
variousO
highly mobile10-10
0.721.00
6octahedron
O-,Nslightly mobile
10-6
1.000.526-8
variousO,O-
mobile10-9

Functions – alkaline metal ions
(i) Form the necessary osmotic pressure, providing stability for the cells and cell
organells.
(ii) Provide charge compensation and stabilisation of structure. Most of the
bio(macro)molecules in the cells have negatively charged groups (phosphate,
carboxylate, sulphonate), which should be neutralised. Furthermore, as the
counter ions of the polyelectrolyte macromolecules (proteins, DNA, RNA,
polysacharydes), or constituents of cells (e.g. phospholipid membranes), actively
participate in stabilisation of their structure. It can be understand easily, if we think
that a single DNA strain should have a few thousand negative charge, and thus a
double helical strain should not be formed without the presence of the counter
ions.
(iii) Information transfer among the cells (action potential/nerve conduction). The
most basic form of the biological control is the electrolytic nerve conduction.
Regulation of the different functions within the cells, even the communication
among the different organs may happen via ion currents, action
potentials/currents.

Physical and chemical characteristics of Mg2+ and Ca2+ ions
The Mg2+ ion is smaller, occurs in regular octahedral geometry, surrounded by water or negatively charged O donors.
The Ca2+ ion is larger, occurs in irregular geometry with high coordination number, and prefers interactions also with neutral O donors.
Due to their two positive charge and higher surface charge density, these ions form mostly ionic, medium strong interactions with bioligands.
The strengths of the coordinative bonds of Mg2+ are about the same as that of Ca2+ ion, but the rate of its reactions is about 3 orders of magnitude slower.
Among the essential divalent metal ions Ca2+-ion is the most mobile.

Functions of Mg2+ and Ca2+ ions
The extracellular Ca2+ stabilises the cell wall and tissue.
In the cell, because of the very low concentration of calcium, Mg2+ ion
have a similar role.
Because of the dipositive charge and their relatively high surface charge
density they form mostly ionic, medium strong interactions with
biomolecules.
Ca2+ - information transfer regulated by coordinative bonds.

Mg2+ ion is essential for thermodynamic stabilization of RNA tertiary structures
Functions of Mg2+ and Ca2+ ions

Selective recognition of the Na+, K+, Ca2+ and Mg2+ ions
The Mg2+ ion is small and prefers the regular octahedral environment,
while the Ca2+ ion is larger and forms easily irregular, fluctuating
coordination sphere with the participation of six-eight donor atoms.
This difference in geometry may results in even 4-5 orders of magnitude
difference in the metal binding strength of thne individual sites, resulting in
complete selectivity between the two metal ions.
However, Na+ and K+ show much higher similarities.
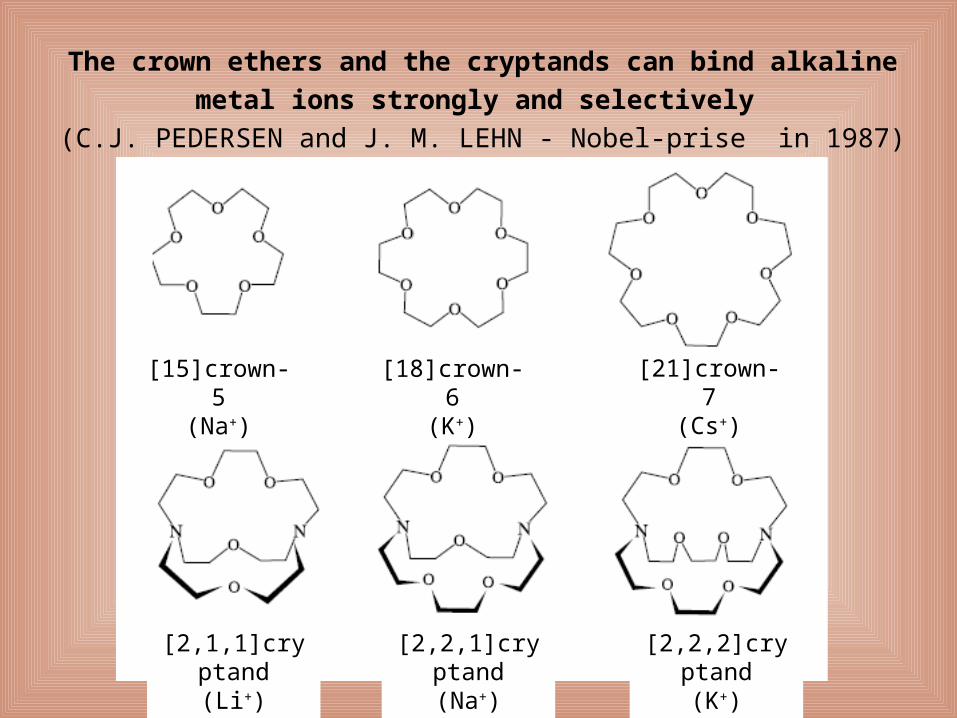
The crown ethers and the cryptands can bind alkaline metal ions
strongly and selectively
(C.J. PEDERSEN and J. M. LEHN - Nobel-prise in 1987)
[15]crown-5(Na+)
[18]crown-6(K+)
[21]crown-7(Cs+)
[2,1,1]cryptand(Li+)
[2,2,1]cryptand(Na+)
[2,2,2]cryptand(K+)

Stability of the K+ complex is ~ 17000 x higher than that of its Na+ complex.
Structure of valynomicin and its potassium complex

Membrane transport processes
Schematic picture of the cell membrane
Outside of cell
Inside of cell
Phospholipid bilayerthickness 30-50 nm
Cholesterol
Peripheral protein
Polysaccharide
Glycoprotein
Transmembrane protein

Membrane transport processes
Two basic membrane transport processes can be distinguished:
Passive transport – in the direction of a concentration gradient
Via ionophores, or channel forming transport proteins
Active transport – opposite to the direction of the concentration gradientVia pumps (special transport proteins)

Ionophores
Mobile ionophores (siderophores, valynomicine)
Channel forming ionophores (gramicidin A)

Gramicidin A:
HCONH-(L)-Val-Gly-(L)-Ala-(D)-Leu-(L)-Ala-(D)-Val-(L)-Val-(D)-Val-(L)-
Trp-(D)-Leu-(L)-Trp-(D)-Leu-(L)-Trp-CONHCH2-CH2-OH
Ionophores

The differences in the size is differentiated further by the charge,
as the charge/ion radius ratio determines the extent of hydration (the
higher this ratio the larger the number the strongly bound water molecule
surrounded the metal ion). This relatively simple control mechanism
works fairly well. Differentiation between anions and cations is made by
charged groups located at the openings of the channel, electrostatic
interactions between the given ions perform this task perfectly. These
channels are usually in a closed state, their openings are covered by a
part of the protein. Opening of the channel is initiated by some stimulus,
which results in conformation change. Depending on the type of the
stimulus channels can be classified as potential or receptor dependent
ion channels.
Channel forming proteins

a b c
K-channel forming protein

K-channel forming protein
In the open state of the channel, the opposite direction flow of the Na+ ions is prevented by a filter. As in its inner hole the dehydrated Na+ ion may be bound only „loosely”, through several carbonyl oxygens; this energy is not enough to cover the dehydration energy of the sodium ion. Besides, the hydrated Na+ ion is too large to go across the filter.

I.
Foszforiláció éskonformáció változás
A nátriumionok távozásaa sejten kívüli térbe
A káliumionokmegkötése
Defoszforiláció éskonformáció változás
A káliumionoktávozása a sejtbelsejébe
A nátriumionokmegkötése
I.
Binding of Na+ ion
Protein Phosphorylation and conformational change
Release of Na+ outside of the cell
Binding of K+ ion
Dephosphorylation and structral change in protein
Release of K+ to the inside of the cell
During a cycle of the Na+-K+ATPase 3 Na+ ion is transported out of the cell and 2 K+ ion into the cell. Both ions are transported against the concentration gradient; the energy need of these processes is covered by the hydrolysis of an ATP molecule.

Nerve conduction/Action potential
In resting state number of the positive and negative ions little differs on the
two sides of the nerve membrane (inside the cell there is a little excess of
the negative charge), and this results in some ptential in the two sides of
the membrane, this is the so called membrane potential. As the membrane
is permeable for the K+ ions, the membrane potential in the resting state is
close to the equilibrium potential of K+ (-70mV).
Nerve conduction, i.e. spreading the action potential along the axon, is
caused by the molecular work of the potential-dependent Na+ and K+
channels located in the nerve membranes.

Mem
bran
e po
tent
ial (
mV
)
Time (s)
Action potential
Na+ ions flow in thecell, K+ ions flow out.
Phase K+-channel Na+-channel
1. Resting state A few are opened All are closed
2. Stimulation A few are opened All become open
3. Depolarization All become open open
4. Max depolarization Open All become inactive
5. Repolarization Open All are inactivated
6. Refractory period Most of them close
inactive - closed transition
7. Resting state A few are opened All are closed

Nerve conduction
When this wave reaches the end of the axon the so called
synapse, due to depolarisation of the membrane through the potential-
dependent Ca2+ channel Ca2+ ionok flow in the synapse, and
neurotransmitter molecules (e.g. acethyl-cholin) are released. These
cross the synaptic cleft and initiate a new action potential in a
neighbouring nerve by chemical stimulation. This new action potential
goes on its way through the other neuron. The neurotransmitter
decomposes soon and the stimulus ceased. The stimulus is conducted
at high speed, it can reach even 100 m/s (360 km/h) value.

Muscle contraction
Components of the thin filament and
their connection to myosine
Izom
Izomrostokkötege
IzomrostMiofibrillum Myofibrill
Muscle fibre
Fascicle
Muscle
TropomiozinMiozin
AktinCa2+
Troponin
A miozin fejrésze
Troponin
Tropomyosin
Actin
Myosin head Myosin

I. In the resting state of the muscle cell actin and myosine are not connected.
Muscle contraction

Muscle contraction
II. Ca2+ ion is bound to one component to the troponin complex (troponin C), and undergoes a con-formational change; a phosphate dissociate from the myosin binding site and the actin binds at the head space of myosin.

Muscle contraction
III. As a result of this, the Mg2+-ADP adduct cleaved of the head space of myosin, its conformation changes and the actin can move as compared to myosin.

Muscle contraction
IV. In the next step Mg2+-ATP binds to the head space of myosin. Meanwhile concentration of Ca2+ decreases in the sarcoplasma due to the function of the Ca2+ pump.


Zoltán Kerényi



















