
UGT Structure/Function Studies (Part 2) - Strona główna · UGT Structure/Function Studies (Part...
39
UGT Structure/Function Studies (Part 2) Anna Radominska-Pandya Department of Biochemistry and Molecular Biology University of Arkansas for Medical Sciences Little Rock, Arkansas, US October 2010; Gdansk University of Technology
Transcript of UGT Structure/Function Studies (Part 2) - Strona główna · UGT Structure/Function Studies (Part...

UGT Structure/Function Studies(Part 2)
Anna Radominska-Pandya
Department of Biochemistry and Molecular Biology
University of Arkansas for Medical Sciences
Little Rock, Arkansas, US
October 2010; Gdansk University of Technology

Identification of Substrate Binding Sites of UGT1A10
Photoaffinity Labeling, MS Analysis, Sequence Alignment, and Mutagenesis
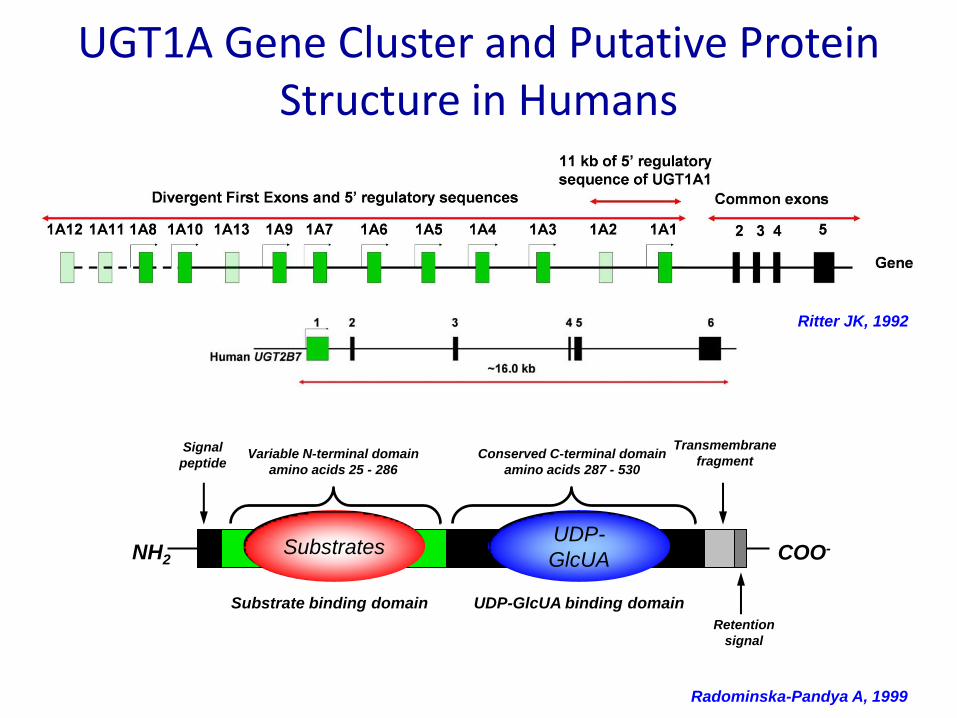
Ritter JK, 1992
UGT1A Gene Cluster and Putative Protein Structure in Humans
Radominska-Pandya A, 1999
Transmembrane
fragment
Substrate binding domain UDP-GlcUA binding domain
Variable N-terminal domain
amino acids 25 - 286
NH2 COO-
Retention
signal
Signal
peptideConserved C-terminal domain
amino acids 287 - 530
UDP-GlcUASubstratesUDP-
GlcUA

Hypotheses
• We hypothesize that the N-terminal domain of UGTs consists of distinct substrate recognition sequences that display specificity for classes of UGT substrates and, thus, determine the catalytic mechanism and substrate specificity of individual UGT isoforms.
• Therefore, we postulate that it is unlikely that a single binding motif could accommodate all substrates specific for each isoform. – Instead, each of these substrate categories may bind to its own
amino acid motif within the larger variable N-terminal “substrate binding region”.
– The proximity of these motifs could have major implications for binding, kinetics, and substrate specificity in the presence of multiple substrates.
• Separate• Overlapping

Purification of His-tagged Recombinant UGT1A10 by IMAC
A. Coomassie blue staining
B. Western blot:Lane 1, 2,3: UGT1A10 eluted in 300 mM imidazole fractions 1, 2 and 3, respectively.
• Solubilization:– 25 mM Tris, pH 7.5, 500 mM NaCl, 2%
Triton X-100
– 16 hr at 4ºC
– Centrifuged at 41,000 x g for 1.5 h.
• Elution buffer:– 50-400mM imidazole buffer, in the
presence of NaCl and Triton X-100.

Photoactivable group
Protein Photoaffinity probe
Irradiation
Proteolytic digestion
Amino acid composition
around the covalent
attachment point
Sequencing
Photoaffinity Labeling
Adapted from Trends Biotechnol. 2000,18:64-77.
UGT1A10 + 4-AzHBA UGT1A10― 4-AzHBA 365nM
25min
UGT1A10 + AzMc UGT1A10―AzMc254nM
90sec
N3N :
h
azide nitrene

Lanes: 1) DMSO; 2-5) Photolabeled with 0.1, 0.15, 0.25, and 1.0 mM AzMC; 6) Labeled is 0.15 mM AzMC in absence of UV.
Lanes: 1) DMSO; 2-6) Photolabeled with 0.1, 0.15, 0.25, 0.75 and 2.0 mM AzMC; 7) Labeled is 0.15 mM AzMC in absence of UV.
Lanes: 1) DMSO; 2-6) Photolabeled with 0.1, 0.15, 0.25, 0.75 and 2.0 mM AzMC; 7) Labeled is 0.15 mM AzMC in absence of UV.
Photoaffinity Labeling of Membrane UGTs with AzMc

Peptide Fragmentation
CH2N
R1
H
C
O
N C
H
R2
C
H
O
N
H
C
R3
H
C
O
N C
H
R4
C
H OH
O
B1
Y3 Y2 Y1
B2 B3
N-terminal ions
C-terminal ions
b―N-terminal side
y―C-terminal side

Peptide Fragmentation of 4-AzHBA-Modified Peptide Identified by MALDI-TOF Analysis
E-F-M-V-F-A-H-A-Q-W-K
Mass+ b - ions y- ions Mass+
E
EF
EFM
EFMV
EFMVF
EFMVFA
EFMVFAH
EFMVFAHA
EFMVFAHAQ
EFMVFAHAQW
FMVFAHAQWK
MVFAHAQWK
VFAHAQWK
FAHAQWK
AHAQWK
HAQWK
AQWK
QWK
WK
K
130.1
277.1
406.2
507.2
654.2
725.3
862.4
933.4
1061.5
1247.6
1264.6
1117.6
986.5
887.5
740.4
669.3
532.3
461.3
333.2
147.1
#
1
2
3
4
5
6
7
8
9
10
#
10
9
8
7
6
5
4
3
2
1
284.1
431.1
562.2
661.2
808.3
879.3
1016.4
1087.4
1215.5
1401.6
b – ions (+154) y ion + 154
1418.6
1271.6
1140.5
1041.5
894.4
823.3
686.3
615.3
487.2
301.1

Fragmentation of the 4-AzHBA-crosslinked Peptide to Identify Phe90, Val92, and Phe93 as Potential Sites for 4-AzHBA
Binding
1 2 3 4 5 6 7 8 9 10 11
11 10 9 8 7 6 5 4 3 2 1

Photoaffinity Labeling Reveals Aglycon Binding Motif for UGT1A10
• Motif was identified using– Photoaffinity labeling
– Proteolytic digestion
– MALDI and LCMS/MS sequencing
– Site-directed mutagenesis
UGT1A1 65 SLYIRDGAFYTLKTYPVPFQREDVKESFVSLGHNVFEN--DSFLQRVIKT 112
UGT1A6 64 NLLLKESKYYTRKIYPVPYDQEELKNRYQSFGNNHFAE--RSFLTAPQTE 111
UGT1A7 63 SWQLGRSLNCTVKTYSTSYTLEDQDREFMVFADARWTAPLRSAFSLLTSS 112
UGT1A8 63 SWQLGKSLNCTVKTYSTSYTLEDLDREFMDFADAQWKAQVRSLFSLFLSS 112
UGT1A9 63 SWQLGRSLNCTVKTYSTSYTLEDLDREFKAFAHAQWKAQVRSIYSLLMGS 112
UGT1A10 63 SWQLERSLNCTVKTYSTSYTLEDQNREFMVFAHAQWKAQAQSIFSLLMSS 112
90FxVF93
Xiong, et al., Biochemistry 2006
4-AzHBA
OH
OOH
NN
+
N-

Conclusions: Phenol Binding Site of UGT1A10
• As identified by MS analysis of 4-AzHBA photolabeled UGT1A10, the N-terminal motif F90-M-V92-F93 is the binding site for probe
• Site-directed mutagenesis of F90-M-V92-F93 motif demonstrated that:
– Phe90 plays a crucial role in binding substrate
– Phe93 is also important for binding phenol substrates
– Val92 is not essential for binding phenol substrates
• Motif F90-M-V92-F93 of UGT1A10 is involved in binding with simple phenol, pNP, and 4-MU

Site-Directed Mutagenesis of UGT1A10 Binding Motif Alters Substrate Specificity
Autoradiograph of TLC separated glucuronide products
4-OH-E1
4-OH-E2
Formononetin
WT V92A
Scopoletin
TAO
TAB
E2
TCC
pNP
4MU
4-OH-E2
TAO
Cl Cl
OH
N3
OH
N3
O2N
TAB

Kinetic profiles for mutant UGT1A10s indicate roles in binding, stoichiometry, and catalysis

Kinetic profiles for mutant UGT1A10s indicate roles in binding, stoichiometry, and catalysis
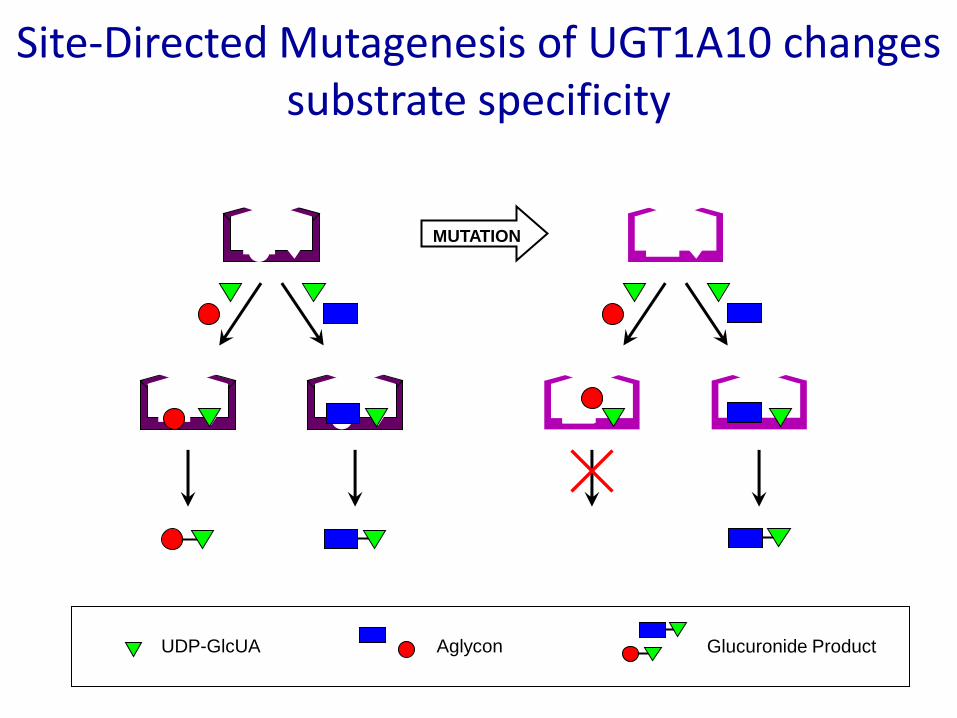
Site-Directed Mutagenesis of UGT1A10 changes substrate specificity
MUTATION
UDP-GlcUA Aglycon Glucuronide Product
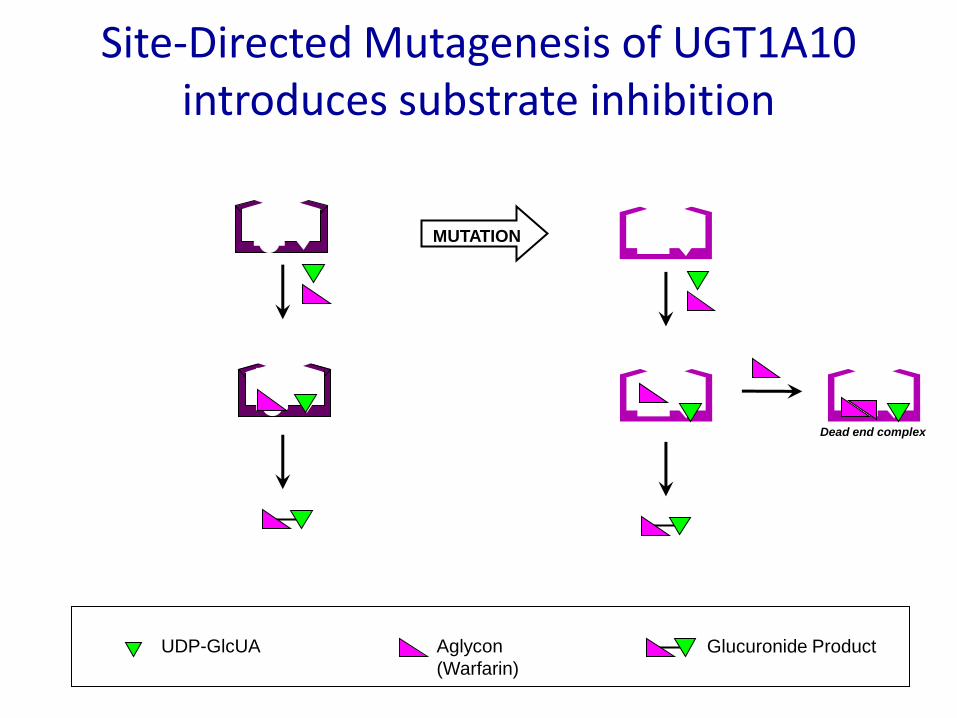
Site-Directed Mutagenesis of UGT1A10 introduces substrate inhibition
Dead end complex
MUTATION
UDP-GlcUA Aglycon
(Warfarin)
Glucuronide Product

Application of Structural Studies
• Inhibition Studies
– Synthesis of UDP-GlcUA binding site-directed inhibitors
– Analysis of inhibition kinetics of UGT1A10
– Homology modeling of UDP-GlcUA binding site in UGT1A10
– Docking of the inhibitor to the active center of UGT1A10 guided by the co-substrate
• Determination of the molecular mechanism of glucuronidation

Bi-dentate UGT Inhibitors
Inhibition of UGT1A10

Structures of PP Inhibitors
• N-acyl phenylaminoalcohol derivative, linked to uridinevia different spacers have been found to be inhibitors of membrane-bound UGTs
• Why “PP” inhibitors?– Named after chemist Pascal Paul – First author of the paper
describing their synthesis.

Effect of PP inhibitors on the glucuronidation of 4-MU
• UGT1A6, 1A8, and 1A10 were incubated with:
– 250 μM 4-MU
– 0, 50, or 200 μM concentrations of each inhibitor.
• Activities are normalized to percent maximal activity.

IC50 calculations
• UGT1A10 was incubated with:– Constant concentration of
substrate and co-substrate
– Increasing concentrations of PP55B.
• The IC50 values were calculated and are shown in μM.
• The inhibition was assumed to be 100% unless otherwise indicated.

Ki Data
• UGT1A10 was incubated with:– Increasing
concentration of substrate at
– Fixed concentrations of inhibitor
• The data was graphed as 1/v against 1/[s].

Inhibition of UGT1A10 by PP55B
• Km, IC50, and Ki values as well as inhibition type for the inhibition of UGT1A10 glucuronidation activity towards selected substrates by PP55B.
“—“ indicates not calculated
Substrate
Substrate UDP-GlcUA
Km
(µM)
IC50
(µM)
Kiapp
(µM)
Inhibition
Type
Km
(µM)
IC50
(µM)
Kiapp
(µM)
Inhibition
Type
7-OH Warfarin 500 ± 60 11 ± 2 8.7 Competitive 990 ± 90 4.9 ± 0.2 -- Mixed
8-OH Warfarin 520 ± 20 35 ± 4 16 Competitive 790 ± 130 14 ± 1 26 Competitive
4-MU 90 ± 20 8 ± 1 16 Competitive 250 ± 90 15 ± 2 5.5 Competitive
E2 48 ± 9 -- 42 Uncompetitive 120 ± 40 39 ± 11 -- Mixed

Conclusions
• Competitive inhibition indicates that PP55B binds to the site for the molecule binding varied in the assay but NOT the other site. – In the case for 4-MU and 8-hydroxywarfarin, the inhibitor only
binds to a SINGLE site in the presence of substrate and inhibitor
• Uncompetitive inhibition indicates PP55B binds to the site of the molecule that is NOT being varied, and thus titrates out active enzyme. – This mechanism may apply to E2 turnover indicating that
binding of E2 favors the association of PP55B to the cofactor binding site
• Mixed inhibition posits that PP55B is binding to both substrate and cofactor sites during turnover. – Whether this process can occur simultaneously is unclear at this
point.

Homology Modeling of UGT1A10
• A model of the mature UGT1A10, residues 25-529, was built with the program Modeller, version 9v4
• The template proteins (2o6l, 2vce and 2iyf) were aligned structurally, and the UGT sequences were added stepwise to the structural alignment with the program ClustalW v2.0.5
• The C-terminal helix missing from the templates was built de novo
• The uridinyl-sugar moieties of the templates were transferred to the UGT protein at construction
• The model was optimimized within Modeller with CHARMM forcefield

Homology Modeling of UGT1A10
• A new model of UGT1A10 was built, based on sequence homology to available structures of plant glucosyltransferases with the GT-B folding motif.
• The model highlights
– Proposed location of His37 that probably participates in the activation of the aglycon substrate
– Conserved aspartic acid residue, Asp148, that may stabilize His37
– Binding site of the sugar donor, UDP-glucuronic acid (UDP-GlcUA)
• Within the C-terminal domain of the enzyme, but in close proximity to His37 within the N-terminal domain

UDP-GlcUA
N-Terminal DomainC-Terminal Domain
Active site of human UGT1A10
• The active site of human UGT1A10 is located between the N- and C-terminal domains. – Helixes are shown as
ice blue cylinders
– Beta-strands as red arrows
– Catalytically important residues His37 and Asp148 are shown in detail
– UDP-GlcUA is shown in detail
H37
D148
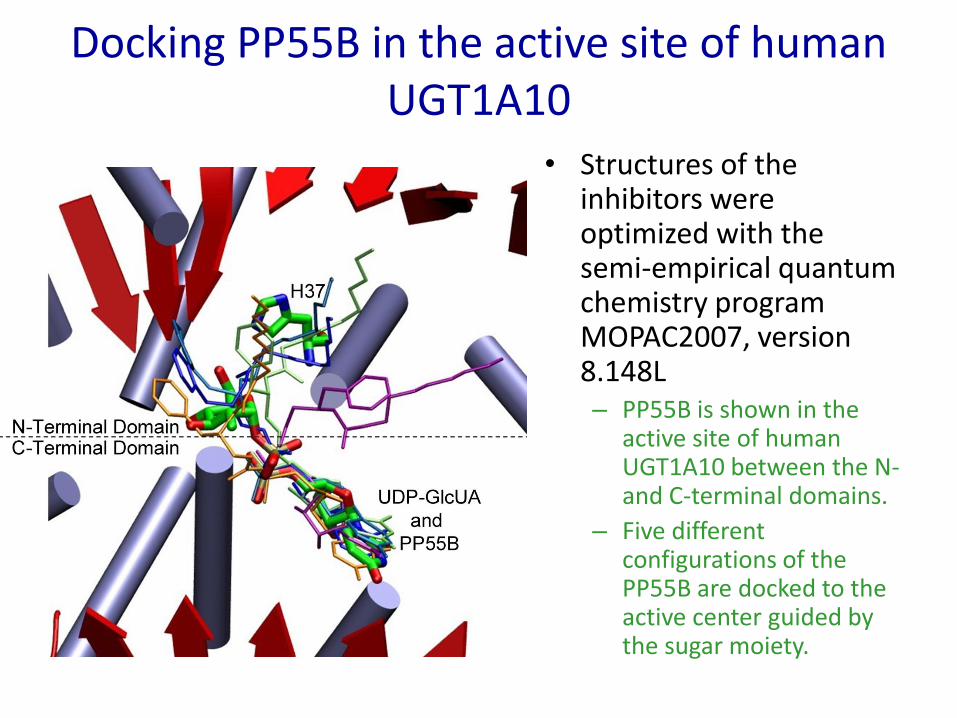
Docking PP55B in the active site of human UGT1A10
• Structures of the inhibitors were optimized with the semi-empirical quantum chemistry program MOPAC2007, version 8.148L– PP55B is shown in the
active site of human UGT1A10 between the N-and C-terminal domains.
– Five different configurations of the PP55B are docked to the active center guided by the sugar moiety.

Molecular Mechanism of Glucuronidation

Homology Model of 2B7 Active Site

Putative Catalytic Mechanism of Phenol Glucuronidation by UGT1A10
O N
O
OH
O
OH
OP
NH
O
O
OH
PO
OH
O
O
OHOH
OH
O
OH
OH
O
OH
Asp148
His37
NHN
Phe90
OH
OAsp393O
Gln394
NH2
“Ring Stacking”
Interactions
Interactions with
O2’,O3’, and O4’
Of GlcUA
N-terminal domain
C-terminal domain

Putative Catalytic Mechanism of Phenol Glucuronidation by UGT1A10
NO
O
OH
O
OH
OP
NH
O
O
OH
PO
OH
O
O
OHOH
OH
O
OH
OH
O-
Asp148
OH
His37
NHN
Phe90
OH
OAsp393O
Gln394
NH2
N-terminal domain
C-terminal domain

Putative Catalytic Mechanism of Phenol Glucuronidation by UGT1A10
O-
NO
O
OH
O
OH
OP
NH
O
O
OH
PO
OH
O
O
OHOH
OH
O
OH
OH
O-
Asp148
His37
NHN+
Phe90
OH
OAsp393O
Gln394
NH2
H
H
N-terminal domain
C-terminal domain

Putative Catalytic Mechanism of Phenol Glucuronidation by UGT1A10
O
-
NO
O
OH
O
OH
OP
NH
O
O
OH
PO
OH
O
O
OHOH
OH
O
OH
OH
O-
Asp148
His37
NHN+
Phe90
OH
OAsp393O
Gln394
NH2
H
-
N-terminal domain
C-terminal domain

Putative Catalytic Mechanism of Phenol Glucuronidation by UGT1A10
OH
O-
Asp148
His37
NHNH+
O
O
OHOH
OH
O
OH
NO
O
OH
O
OH
OP
NH
O
O
OH
PO
OH
O-
Phe90
OH
OAsp393O
Gln394
NH2
N-terminal domain
C-terminal domain

Conclusions
• The signature of the UGT is its specific interaction with the carboxylic acid of glucuronic acid
• Significant differences in catalysis are observed between the 1A and 2B families– Amino acids involved in binding donor and acceptor
– Role of metal in catalysis
• Cannot consider the C-terminal and N-terminal ends separately. We must view the protein as a whole with both domains contributing to interactions with both donor and acceptor.– Active site in “cleft” between two domains
– Interactions between domains are essential
• First molecular mechanism of phenols for UGT1A10 was postulated

Practice Questions

UGT1A1
UGT1A10


















