
Two epsilon glutathione S-transferase cDNAs from the common cutworm, Spodoptera litura:...
10
Two epsilon glutathione S-transferase cDNAs from the common cutworm, Spodoptera litura: Characterization and developmental and induced expression by insecticides Huimin Deng, Yufen Huang, Qili Feng, Sichun Zheng * Guangdong Provincial Key Lab of Biotechnology for Plant Development, College of Life Sciences, South China Normal University, Guangzhou, 510631, China 1. Introduction Glutathione S-transferase (GST; E.C. 2.5.1.18) belongs to a protein family involved in detoxification of xenobiotics, protection from oxidative damage and intracellular transport of hormones, endogenous metabolites and exogenous chemicals. Interests in insect GSTs are primarily because of their roles in insecticide resistance. It is known that GSTs confer resistance to organo- chlorine (Clark and Shamaan, 1984; Tang and Tu, 1994), organophosphorous (Lewis and Sawicki, 1971; Huang et al., 1998) and pyrethroid insecticides (Vontas et al., 2001, 2002). GSTs catalyze the conjugation of reduced glutathione (GSH) with exogenous and endogenous toxic compounds or their metabolites, rendering them more water soluble, less toxic, and easier to excrete (Hayes and Pulford, 1995). In addition, some GSTs catalyze the dehydrochlorination of DDT to the noninsecticidal metabolite DDE by using reduced glutathione as a cofactor (Clark and Shamaan, 1984; Ranson et al., 2001). Resistant strains of various insect species often show an increase in GST expression and/or GST activity against insecticides (Ortelli et al., 2003). Insect GSTs have been classified into six classes: delta, epsilon, omega, sigma, theta and zeta (Chelvanayagam et al., 2001; Ranson et al., 2001). The delta and epsilon classes of insect GSTs are insect- specific and exist as large gene clusters in insect genomes. Most of the GSTs that have been implicated to play roles in insecticide resistance belong to the delta and epsilon classes (Tang and Tu, 1994; Ranson et al., 2001; Ding et al., 2003). In Lepidoptera, the GST-caused insecticide resistance and the mechanisms have been studied, although not as well as in dipteran species. In the diamondback moth Plutella xylostella, Pxgste1 is responsible for organophosphorous (OP) and benzoylphenylureas (chlorflua- zuron) resistance (Huang et al., 1998; Sonoda and Tsumuki, 2005). Enzyme activity of an omega GST from the silkworm Bombyx mori is induced by OP (Yamamoto et al., 2008). CfGSTs1, CfGSTd1 and CfGSTd2 from Choristoneura fumiferana are induced by tebufenozide (RH5992) (Feng et al., 2001; Zheng et al., 2007). CfGSTs1 was also induced at both mRNA and protein levels by balsam fir foliage and the insecticides (e.g. Bt delta-endotoxin and permethrin) (Feng et al., 2001). In the common cutworm Spodoptera litura, a GST cDNA has been reported whose mRNA level is increased by pyriproxyfen treatment (Wu and Lu, 2008). S. litura is one of the most damaging agricultural pests in the tropical and subtropical areas around the world. This insect feeds on more than 90 families of plants, in particular cruciferous vegetables, and causes enormous losses in many economically Journal of Insect Physiology 55 (2009) 1174–1183 ARTICLE INFO Article history: Received 7 July 2009 Received in revised form 18 August 2009 Accepted 18 August 2009 Keywords: Glutathione S-transferases Detoxification Gene expression Spodoptera litura ABSTRACT Two Spodoptera litura glutathione S-transferase cDNAs (Slgste2 and Slgste3) which were cloned from a midgut cDNA, encoded two structurally distinct proteins with a predicted molecular mass of 25 and 24 kDa, respectively. Slgste2 and Slgste3 were single-copy genes in the S. litura genome and there was no intron within the genes. The transcripts and proteins of Slgste2 and Slgste3 were predominately present in the midgut of the 5th and 6th instar larvae. The apparent Vmax of the purified SlGSTE2 and SlGSTE3 recombinant proteins towards the substrates glutathione and 1-chloro-2,4-dinitrobenezene (CDNB) were similar. Slgste2 expression was up-regulated by 1-naphthyl methylcarbamate (carbaryl), 1,1,1- trichloro-2,2-bis-(p-chlorophenyl) ethane (DDT), deltamethrin, tebufenozide (RH5992) and Bacillus thuringiensis (Bt), but not affected by malathion, while Slgste3 expression was slightly up-regulated by carbaryl, Bt and DDT, but not affected by RH5992, malathion and deltamethrin. The results suggest that Slgste2 and Slgste3 may play roles in detoxifying various insecticides in S. litura. ß 2009 Elsevier Ltd. All rights reserved. * Corresponding author. E-mail address: [email protected] (S. Zheng). Contents lists available at ScienceDirect Journal of Insect Physiology journal homepage: www.elsevier.com/locate/jinsphys 0022-1910/$ – see front matter ß 2009 Elsevier Ltd. All rights reserved. doi:10.1016/j.jinsphys.2009.08.017
-
Upload
huimin-deng -
Category
Documents
-
view
215 -
download
1
Transcript of Two epsilon glutathione S-transferase cDNAs from the common cutworm, Spodoptera litura:...

Two epsilon glutathione S-transferase cDNAs from the common cutworm,Spodoptera litura: Characterization and developmental and induced expressionby insecticides
Huimin Deng, Yufen Huang, Qili Feng, Sichun Zheng *
Guangdong Provincial Key Lab of Biotechnology for Plant Development, College of Life Sciences, South China Normal University, Guangzhou, 510631, China
Journal of Insect Physiology 55 (2009) 1174–1183
A R T I C L E I N F O
Article history:
Received 7 July 2009
Received in revised form 18 August 2009
Accepted 18 August 2009
Keywords:
Glutathione S-transferases
Detoxification
Gene expression
Spodoptera litura
A B S T R A C T
Two Spodoptera litura glutathione S-transferase cDNAs (Slgste2 and Slgste3) which were cloned from a
midgut cDNA, encoded two structurally distinct proteins with a predicted molecular mass of 25 and
24 kDa, respectively. Slgste2 and Slgste3 were single-copy genes in the S. litura genome and there was no
intron within the genes. The transcripts and proteins of Slgste2 and Slgste3 were predominately present
in the midgut of the 5th and 6th instar larvae. The apparent Vmax of the purified SlGSTE2 and SlGSTE3
recombinant proteins towards the substrates glutathione and 1-chloro-2,4-dinitrobenezene (CDNB)
were similar. Slgste2 expression was up-regulated by 1-naphthyl methylcarbamate (carbaryl), 1,1,1-
trichloro-2,2-bis-(p-chlorophenyl) ethane (DDT), deltamethrin, tebufenozide (RH5992) and Bacillus
thuringiensis (Bt), but not affected by malathion, while Slgste3 expression was slightly up-regulated by
carbaryl, Bt and DDT, but not affected by RH5992, malathion and deltamethrin. The results suggest that
Slgste2 and Slgste3 may play roles in detoxifying various insecticides in S. litura.
� 2009 Elsevier Ltd. All rights reserved.
Contents lists available at ScienceDirect
Journal of Insect Physiology
journa l homepage: www.e lsev ier .com/ locate / j insphys
1. Introduction
Glutathione S-transferase (GST; E.C. 2.5.1.18) belongs to aprotein family involved in detoxification of xenobiotics, protectionfrom oxidative damage and intracellular transport of hormones,endogenous metabolites and exogenous chemicals. Interests ininsect GSTs are primarily because of their roles in insecticideresistance. It is known that GSTs confer resistance to organo-chlorine (Clark and Shamaan, 1984; Tang and Tu, 1994),organophosphorous (Lewis and Sawicki, 1971; Huang et al.,1998) and pyrethroid insecticides (Vontas et al., 2001, 2002).GSTs catalyze the conjugation of reduced glutathione (GSH) withexogenous and endogenous toxic compounds or their metabolites,rendering them more water soluble, less toxic, and easier toexcrete (Hayes and Pulford, 1995). In addition, some GSTs catalyzethe dehydrochlorination of DDT to the noninsecticidal metaboliteDDE by using reduced glutathione as a cofactor (Clark andShamaan, 1984; Ranson et al., 2001). Resistant strains of variousinsect species often show an increase in GST expression and/or GSTactivity against insecticides (Ortelli et al., 2003).
* Corresponding author.
E-mail address: [email protected] (S. Zheng).
0022-1910/$ – see front matter � 2009 Elsevier Ltd. All rights reserved.
doi:10.1016/j.jinsphys.2009.08.017
Insect GSTs have been classified into six classes: delta, epsilon,omega, sigma, theta and zeta (Chelvanayagam et al., 2001; Ransonet al., 2001). The delta and epsilon classes of insect GSTs are insect-specific and exist as large gene clusters in insect genomes. Most ofthe GSTs that have been implicated to play roles in insecticideresistance belong to the delta and epsilon classes (Tang and Tu,1994; Ranson et al., 2001; Ding et al., 2003). In Lepidoptera, theGST-caused insecticide resistance and the mechanisms have beenstudied, although not as well as in dipteran species. In thediamondback moth Plutella xylostella, Pxgste1 is responsible fororganophosphorous (OP) and benzoylphenylureas (chlorflua-zuron) resistance (Huang et al., 1998; Sonoda and Tsumuki,2005). Enzyme activity of an omega GST from the silkwormBombyx mori is induced by OP (Yamamoto et al., 2008). CfGSTs1,CfGSTd1 and CfGSTd2 from Choristoneura fumiferana are inducedby tebufenozide (RH5992) (Feng et al., 2001; Zheng et al., 2007).CfGSTs1 was also induced at both mRNA and protein levels bybalsam fir foliage and the insecticides (e.g. Bt delta-endotoxin andpermethrin) (Feng et al., 2001). In the common cutwormSpodoptera litura, a GST cDNA has been reported whose mRNAlevel is increased by pyriproxyfen treatment (Wu and Lu, 2008).
S. litura is one of the most damaging agricultural pests in thetropical and subtropical areas around the world. This insect feedson more than 90 families of plants, in particular cruciferousvegetables, and causes enormous losses in many economically

H. Deng et al. / Journal of Insect Physiology 55 (2009) 1174–1183 1175
important crops. Significant resistance against various insecticideshas been developed and reported (Armes et al., 1997). In thepresent paper, two GST cDNAs (Slgste2 and Slgste3) of the epsilonclass were identified from S. litura. Spatial and temporal expressionpatterns of mRNA and proteins in the epidermis, fat body, andmidgut during the larval development were analyzed. Inducedexpression by different insecticides was also determined. The datasuggest that Slgste2 and Slgste3 may play a role in detoxification ofvarious insecticides in S. litura.
2. Materials and methods
2.1. Experimental insects and treatments
Larvae of Spodoptera litura (Lepidoptera:Noctuidae) werereared on artificial diet at 25 8C, 70% relative humidity and undera photoperiod of 12 h light and 12 h dark, and until they reachedthe pupal stage.
Nonsteroidal ecdysone agonist, tebufenozide (RH5992), 1-naphthyl methylcarbamate (carbaryl), dichloro-diphenyl-trichlor-oethane (DDT), malathion, deltamethrin and Bacillus thuringiensis
(Bt) were applied to one-day-old 3rd instar larvae. One microliterof 70 ng/mL RH5992, 4.09 mg/mL carbaryl, 0.7 mg/mL DDT,2.43 mg/mL malathion, or 0.03 mg/mL deltamethrin, in imbibingsolution (1% sucrose) on a small diet plug were fed to 3rd instarlarvae. The controls were treated with the imbibing solution alone.After consumption of the insecticide-containing diet, the larvaewere transferred to regular diet. The insects were collected at 6, 12,24, 48 and 72 h post-treatment for RNA isolation. All the sampleswere immediately frozen with liquid nitrogen and stored at�80 8Cuntil they were used for RNA extraction.
2.2. Cloning and sequencing of Slgst cDNAs
Partial sequences of putative GST cDNAs were identified from anexpressed sequence tag (EST) set, which were isolated from a cDNAlibrary that was constructed in the Uni-ZAP XR vector by using theZAP cDNA Gigapack II Gold Cloning Kit (Stratagene, La Jolla, CA) withmRNA isolated from the midgut of 6th larvae of S. litura. The partialsequences were then assembled into 8 full-length cDNA sequencesby using the CAP3 program (Huang and Maddan, 1999) andconfirmed by sequencing the longest target cDNA clones selectedfrom the cDNA library. DNA sequencing was conducted using ABI377 capillary automatic sequencer. Annotation and comparisonalignment of sequences were performed using the National Centerfor Biotechnology Information BLAST search services (Altschul et al.,1990). Multiple sequence alignment and phylogenetic tree analysiswere conducted using the Neighbor-Joining method in MEGA 4software with 1000 bootstrap replicates.
2.3. Genomic DNA isolation and PCR analysis of Slgst genes
Genomic DNA was prepared from larvae of S. litura using amodified phenol extraction method as described by Sambrooket al. (1989). Primers used for gene amplification from the genomicDNA were designed based on sequences of the cloned Slgste2 andSlgste3 cDNAs. The primers used for amplification of Slgste2 genewere: forward primer: 50-ATCAGGTGATCCTTCTGC-30; reverseprimer: 50-ACAATCTACTTCTTCTTG TTC-30; the primers used forSlgste3 gene were: forward primer: 50-CCCGGAACAG TCTGTCAA-CAT-30, reverse primer: 50-TTATTGATTATTTCTAGCAGT-30. Geno-mic DNA was denatured at 94 8C for 3 min, followed by 30-cycleamplification (94 8C for 30 s; 55 8C for 30 s; 72 8C for 30 s), PCRproducts were analyzed using 0.8% agarose gel electrophoresis.The PCR products were cloned into pMD18T vector (TaKaRa, China)for sequencing.
2.4. RNA isolation and reverse-transcription PCR (RT-PCR)
Total RNA was isolated from larval tissues using the guanidi-nium thiocyanate method (Chomyczynski and Sacchi, 1987). ForRT-PCR, first strand DNA synthesis was performed using Super-script First Strand Synthesis System for RT-PCR from TaKaRaaccording to the manufacturer’s instruction. The primers foramplifying Slgste2 were: forward primer: 50-ACATCATCGGGCTG-GAACTC-30, reverse primer: 50-AGTTTATGCAAAGGTTGCAT-30; theprimers for amplifying Slgste3 were: forward primer: 50-TCACAT-CATTGGACTCAAACT-30, reverse primer: 50-TGTGCAGAACCTGTG-CAGCT-30. Template DNA was denatured at 94 8C for 3 min,followed by 26 cycles of 94 8C for 30 s, 58 8C for 30 s, and 72 8C for30 s for each cycle. PCR products were separated in 0.8% agarosegels and stained with ethidium bromide.
2.5. Southern and northern hybridization
Aliquots (20 mg) of genomic DNA were digested with Xho I, Hind
III or EcoR I, fractionated on a 1% agarose gel, transferred to aHybond N+ nylon membrane (Amersham Co. USA) and hybridizedusing the 32P-labelled probes made with Slgste2 and Slgste3 full-length cDNA and a Random Primer DNA Labeling Kit Ver. 2.0(TaKaRa, China). Hybridization was carried out at 42 8C for 16 h inhybridization buffer [5 � Denhart’s solution, 5 � SSPE (1 � SSPE =180 mM NaCl, 10 mM sodium phosphate, pH 7.7, 1 mM EDTA),0.1% (w/v) SDS, 0.1% (w/v) sodium pyrophosphate, 5% (w/v)polyethylene glycol and 100 mg/mL boiled and sheared herringsperm DNA]. The blot was washed at 65 8C in 5 � SSPE for 30 minand then in 1 � SSPE/0.1% SDS for 30 min. A final wash wasconducted at high stringency (0.1 � SSPE/0.1% SDS at 65 8C) for30 min. The membranes were scanned and photographs weretaken in Typhon-TRIO (Amersham Biosciences, USA).
For northern blot analysis, 10 mg of total RNA from theepidermis, fat body and midgut of larvae was separated on aformaldehyde–agarose (1%) gel according to Lehrach et al. (1977).The RNA was visualized by staining with ethidium bromide andphotographed under UV light. RNA was transferred from the gels toHybond N+ membranes. Hybridization, washing and scanningwere performed as described for southern blot analysis.
2.6. Protein isolation and western blot analysis
The whole larvae or larval tissues of S. litura were homogenizedin a homogenization buffer (5 mL/g tissue; 50 mM Tris, 10 mMEDTA, 15% glycerol, 0.005% phenylthiourea, pH 7.8) using a motor-driven Teflon pestle in a 1.5 mL polypropylene microcentrifugetube. The homogenate was centrifuged at 10,000 � g for 5 min andthe supernatant was re-centrifuged under the same condition. Thefinal supernatant was used for protein analysis. Protein concen-tration was estimated using the method of Bradford (1976).Absorbance was measured at 595 nm and the concentration wascalculated according to a bovine serum albumin standard curve.
Protein samples were denatured at 100 8C for 5 min in an equalvolume of 2� protein loading buffer (0.1 M Tris buffer, pH 6.8, 4%SDS, 0.2% bmercaptoethanol, 40% glycerol, and 0.002% bromophe-nol blue). SDS-PAGE was performed using 12% acrylamide gels inTris–glycine–SDS buffer (10 mM Tris, 50 mM glycine, 0.1% SDS, pH8.0). The gel was stained with Coomassie Blue R-250. Themolecular mass of the proteins was calculated using the ProteinMolecular Weight Marker (low) (TaKaRa, China).
For western blot analysis, proteins were transferred afterelectrophoresis from the acrylamide gel to nitrocellulose mem-brane. The membrane was blocked with 3% BSA in 1 � PBS bufferfor 2 h at room temperature, and then incubated with the primaryantibodies, anti-SlGSTE2 or anti-SlGSTE3 antibody, (1:1000) at

H. Deng et al. / Journal of Insect Physiology 55 (2009) 1174–11831176
room temperature for 1 h. Goat anti-rabbit IgG (Dingguo, China)conjugated with alkaline phosphatase was used as the secondaryantibody at a dilution of 1:1000. Nitroblue tetrazolium and 5-bromo-4-chloro-3-indolyl phosphate were used as substrates forcolor development.
2.7. In vitro expression and purification of recombinant SlGST proteins
Open reading frames (ORF) of the target cDNAs were clonedinto the pPROEX HTb expression vector (Life Technologies,Burlington, Canada), generating the recombinant expressionvectors, pPROEXb-Slgste2 and pPROEXb-Slgste3. E. coli cells(DH5a) were transformed with the recombinant plasmid DNAsand were grown at 37 8C in Luria–Bertani (LB) media containing100 mg/mL ampicillin. Protein expression was induced by addingisopropyl-b-D-thiogalactopyranoside (IPTG) at a final concentra-tion of 1 mM. The cells were further cultured for 3 h beforecollection by centrifugation. The resulting cell pellets weresuspended in 20 mM Tris–HCl buffer (pH 8.0) containing 0.5 MNaCl, 4 mg/mL of lysozyme, and 1 mM PMSF (phenymethanesul-fonyl fluoride) and then lysed by sonication. The suspension wascentrifuged at 10,000 � g at 4 8C for 5 min. The supernatant andprecipitant were collected for protein analysis.
Protein purification was conducted using a GST-glutathioneaffinity chromatography system from Amersham PharmaciaBiotech (Piscataway, NJ, USA) following the manufacturer’sprotocol. To each milliliter of the protein extracts, 100 mL of 50%slurry of glutathione-sepharose 4B was added. The mixture wasincubated with gentle agitation for 30 min at room temperature.The suspension was then centrifuged at 500 � g for 5 min. Theagarose matrix precipitate was collected and washed three timeswith 10 bed volumes of phosphate buffered saline (140 mM NaCl,2.7 mM KCl, 10 mM Na2HPO4, 1.8 mM KH2PO4, pH 7.5) each time.After the final wash, the matrix was suspended in 1 mL ofglutathione elution buffer (10 mM reduced glutathione in 50 mMTris–HCl, pH 8.0) for each 1 mL bed volume of the sedimentedmatrix. The suspension was incubated for 20 min at roomtemperature and then centrifuged at 500 � g for 5 min.
For the insoluble recombinant protein that was expressed asocclusion body, the insoluble protein occlusion bodies werecollected by centrifugation and was washed twice using 50 mMTris–HCl buffer (pH 7.5) containing 10 mM EDTA, 5 mM DDT, 2%TritonX-100, 500 mM NaCl, followed by two washes using 50 mMTris–HCl buffer (pH 7.5) containing 10 mM EDTA. The pellet wascollected by centrifugation at 15,000 � g for 20 min at 4 8C andsuspended in 50 mM Tris–HCl buffer (pH 8.0) containing 8 M ureaat 4 8C for overnight. It was centrifuged at 7500 � g for 20 min. Thesupernatant was dialyzed against 100 volumes of 0.15 M NaCl at4 8C for overnight. The samples were finally centrifuged at10,000 � g for 20 min 4 8C and the supernatants were collected.The purification was done following the procedure describedabove.
2.8. Antibody production and immunohistochemistry
The GST affinity column purified recombinant GST proteinswere used as antigens. Polyclonal antiserum was made in rabbitsby three booster injections, each with 500 ng protein in Freund’sadjuvant. Antiserum was collected after the booster injections andthe pre-immune serum that was collected from the same rabbitprior to immunization was used as a control.
Immunohistochemistry localization of SlGSTE2 and SlGSTE3proteins were performed as described in Feng et al. (2001). One-day-old 5th instar larvae were fixed in 4% formaldehyde at 4 8Cfor overnight. The larvae were the embedded in paraffin and 8-mm-thick section were made for immunostaining. The sections
were stained with the primary antibody, anti-SlGSTE2 or anti-SlGSTE3 antibody, at a dilution of 1:200. The secondary antibody,sheep anti-rabbit IgG conjugated with SABC-FITC (StreptAvidin–Biotin Complex-fluorescein isothiocyanate) (Boster, Wuhan,China) was used at a dilution of 1:1000. The sections werecounter-stained with 40,6-diamidine-20-phenylindole dihy-drochloride (DAPI) at a dilution of 1:1000 for 30 min andexamined under a fluorescence microscope (DMI4000B). Thephotographs were taken as double exposures using fluoresceinand DAPI filters.
2.9. Enzyme activity assay for SlGSTs
Measurement of enzymic activity SlGSTE2 and SlGSTE3recombinant proteins were conducted as described by Grantet al. (1989). Three micrograms of protein was used in a totalvolume of 300 mL of a reaction mixture. The two substrates for GST,1-chloro-2,4-dinitrobenzene (CDNB) and reduced glutathione,were added to the reaction wells. The change in absorbance ofCDNB conjugate for the first minute was measured at 340 nm and25 8C using DU800 spectrometer (Beckman coulter Instruments,Inc., Winooski, VT, USA). Protein concentrations were measuredusing the Bio-Rad protein reagent and bovine serum albumin asstandard. Enzyme activity is represented as mmol of CDNBconjugated per min per mg protein. The apparent Km and Vmaxwere determined using the double reciprocal plot analysis. Eachdata point represents the average of 6 measurements.
Initial velocities for the GST-catalyzed reaction were measuredat 25 8C in a total volume of 300 mL containing 100 mM potassiumphosphate buffer, pH 7.3, 2 mM CDNB, 3 mM GSH in the presenceor absence of 100 mM of a insecticidal compound.
3. Results
3.1. Cloning of S. litura GST cDNAs
Out of 6815 ESTs generated from S. litura midgut cDNA libraries(Qili Feng, unpublished data), eight ESTs encoding different GSTswere identified. All of these GSTs differed from SlGST1 clonedpreviously identified from the same species by Wu and Lu (2008).These ESTs were assembled into two epsilon GSTs and were namedSlgste2 and Slgste3 (Fig. 1A). Slgste2 was 826 nucleotides in lengthand encoded a 223-amino acid protein with a predicted molecularmass of 25 kDa and a pI of 6.24 (GenBank accession number:GQ131803). Slgste3 was 721 nucleotides in length and encoded a217-amino acid protein with a predicted molecular mass of 24 kDaand a pI of 7.10 (GenBank accession numbers GQ131804). Both thetwo sequences had a glutathione binding region (G-site) and aelectrophilic substrate binding region (H-site), which are typicalfeatures of GSTs (Fig. 1A). No potential N-glycosylation sites weredetected in the deduced amino acid sequences of SlGSTE2 andSlGSTE3, when predication was made by using NetNGlyc 1.0software (http://www.cbs.dtu.dk/services/NetNGlyc). YinOYangprogram (http://www.cbs.dtu.dk/services/YinOYang) predicted apotential O-b-glycosylation site (Ser5) in SlGSTE3 but not inSlGSTE2. A serine residue, which is suggested to be necessary forthe GSH catalytic activity (Sheehan et al., 2001), was found at theresidue position 11 and the position 5 of the N-terminal domains ofSlGSTE2 and SlGSTE3, respectively (Fig. 1A).
3.2. Comparison and phylogenetic analysis of the S. litura SlGSTE2 and
SlGSTE3 with other insect GSTs
Sequence comparison revealed that the deduced amino acidsequences of SlGSTE2 and SlGSTE3 shared an identity of 49.8%. TheN-terminal catalytic domains of SlGSTE2 and SlGSTE3 are more

Fig. 1. (A) Sequence alignment of the three S. litura epsilon GSTs. The putative glutathione binding region (G-site) and the electrophilic substrate binding region (H-site) are
indicated by lines at the top of the panels. The characteristic motif region of epsilon GSTs is boxed with dash line and the feature motif for GSTs is boxed with solid line. The
residues that are identical in all of the three sequences are indicated in white letters with black background. The residues that are conserved in two out of three sequences are
labeled with grey background. The serine residue, which is suggested to be necessary for the GSH catalytic activity (Sheehan et al., 2001), is indicated by an arrowhead. (B)
Phyologenetic tree analysis of SlGSTE2 and SlGSTE3 with other 34 insect GSTs from different species. The analysis was carried out using the Neighbor-Joining method in MEGA
4 software. The numbers on the forks indicate the percentage values of bootstrap at 1000 replicates.
H. Deng et al. / Journal of Insect Physiology 55 (2009) 1174–1183 1177
conserved than the G-site region (substrate binding domain) ofthese proteins. SlGSTE2 and SlGSTE3 showed 36.7% and 32.7%amino acid identities, respectively, with SlGSTE1 cloned from thesame species (called GST1 by Wu and Lu, 2008) [AAS7989].Phylogenetic analysis of insect GST showed that the SlGSTE2 andSlGSTE3 were clustered closely with the GSTs from other insectsincluding Bombyx mori GSTe6 (NP_001108465), C. fumiferana
GSTd1 (ABQ53630), S. litura GSTe1 (AAS7989), P. xylostella GSTe1(AAC35245) and Helicoverpa armigera GST (AAP75792) (Fig. 1B),which are the members of the epsilon class. Therefore, the SlGSTE2and SlGSTE3 were classified into the epsilon group. According tothe GST nomenclature system suggested by Chelvanayagam et al.(2001), these two S. litura GSTs genes are named Slgste2 and Slgste3,respectively. A feature motif, RAVELTAK, for epsilon GSTs in A.
gambiae was suggested by Ortelli et al. (2003). However, in
SlGSTE1, SlGSTE2 and SlGSTE3 this motif is various, with only thesecond and third amino acid residues (Ala and Val) conserved(Fig. 1A).
3.3. Gene structure and copy number of Slgste2 and Slgste3
To examine whether or not the Slgste2 and Slgste3 genes haveintrons within the ORFs, PCR amplification was performed withgenomic DNA as a template using the primers designed based onthe sequences of Slgste2 and Slgste3 cDNAs. The results ofsequencing showed that the genomic PCR products are identicalto Slgste2 and Slgste3 cDNAs, respectively, and no intron was foundin the coding regions of Slgste2 and Slgste3. Southern blot analysiswith genomic DNA digested with restriction enzymes EcoR I, Hind
III and Xho I, respectively, showed a single positive band for both
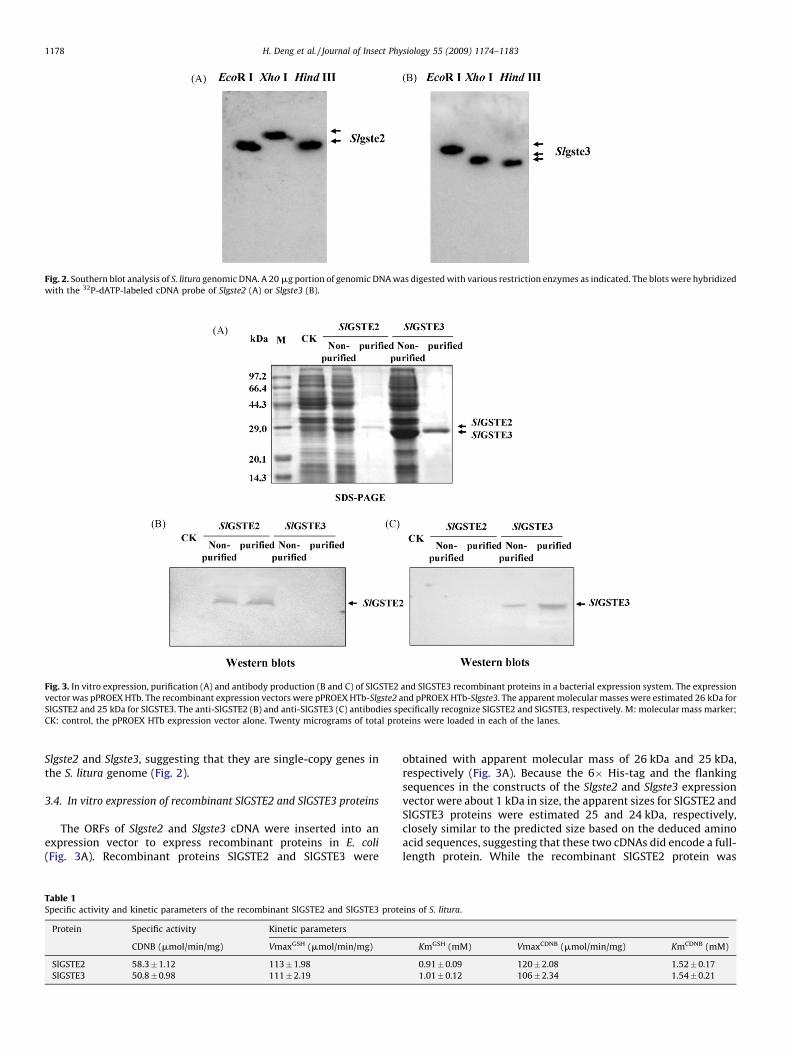
Fig. 2. Southern blot analysis of S. litura genomic DNA. A 20 mg portion of genomic DNA was digested with various restriction enzymes as indicated. The blots were hybridized
with the 32P-dATP-labeled cDNA probe of Slgste2 (A) or Slgste3 (B).
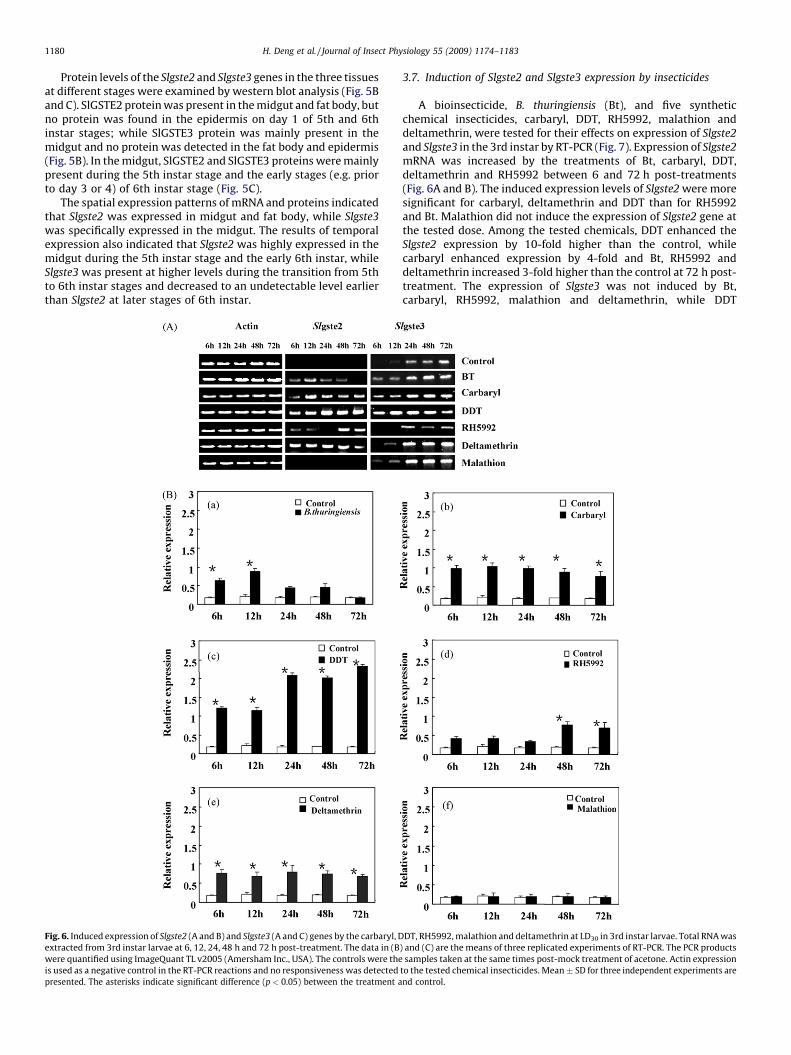
Fig. 3. In vitro expression, purification (A) and antibody production (B and C) of SlGSTE2 and SlGSTE3 recombinant proteins in a bacterial expression system. The expression
vector was pPROEX HTb. The recombinant expression vectors were pPROEX HTb-Slgste2 and pPROEX HTb-Slgste3. The apparent molecular masses were estimated 26 kDa for
SlGSTE2 and 25 kDa for SlGSTE3. The anti-SlGSTE2 (B) and anti-SlGSTE3 (C) antibodies specifically recognize SlGSTE2 and SlGSTE3, respectively. M: molecular mass marker;
CK: control, the pPROEX HTb expression vector alone. Twenty micrograms of total proteins were loaded in each of the lanes.
H. Deng et al. / Journal of Insect Physiology 55 (2009) 1174–11831178
Slgste2 and Slgste3, suggesting that they are single-copy genes inthe S. litura genome (Fig. 2).
3.4. In vitro expression of recombinant SlGSTE2 and SlGSTE3 proteins
The ORFs of Slgste2 and Slgste3 cDNA were inserted into anexpression vector to express recombinant proteins in E. coli
(Fig. 3A). Recombinant proteins SlGSTE2 and SlGSTE3 were
Table 1Specific activity and kinetic parameters of the recombinant SlGSTE2 and SlGSTE3 prot
Protein Specific activity Kinetic parameters
CDNB (mmol/min/mg) VmaxGSH (mmol/min/mg)
SlGSTE2 58.3�1.12 113�1.98
SlGSTE3 50.8�0.98 111�2.19
obtained with apparent molecular mass of 26 kDa and 25 kDa,respectively (Fig. 3A). Because the 6� His-tag and the flankingsequences in the constructs of the Slgste2 and Slgste3 expressionvector were about 1 kDa in size, the apparent sizes for SlGSTE2 andSlGSTE3 proteins were estimated 25 and 24 kDa, respectively,closely similar to the predicted size based on the deduced aminoacid sequences, suggesting that these two cDNAs did encode a full-length protein. While the recombinant SlGSTE2 protein was
eins of S. litura.
KmGSH (mM) VmaxCDNB (mmol/min/mg) KmCDNB (mM)
0.91�0.09 120�2.08 1.52� 0.17
1.01�0.12 106�2.34 1.54� 0.21

Fig. 4. Inhibition of the SlGSTE2 and SlGSTE3 proteins by insecticides at a
concentration of 100 mM. Initial velocities for the GST-catalyzed reaction were
measured at 25 8C in a total 238 volume of 300 ml containing 100 mM potassium
phosphate buffer, pH 7.3, 2 mM 239 CDNB, 3 mM GSH in the presence or absence of
100 mM of a insecticidal compound.
H. Deng et al. / Journal of Insect Physiology 55 (2009) 1174–1183 1179
expressed as soluble protein, the recombinant SlGSTE3 protein wasexpressed as occlusion bodies. After denaturing and refolding, thetwo proteins were purified using GST affinity columns and used forantibody production, as well as enzyme activity assay. Antiseragenerated against the proteins specifically recognized the recom-binant SlGSTE2 and SlGSTE3, respectively (Fig. 3B and C).
3.5. Kinetic properties of SlGSTE2 and SlGSTE3 recombinant proteins
Enzyme kinetics studies revealed that Vmax and Km values ofthe recombinant SlGSTE2 were 113 � 1.98 mmol/mg/min and0.91 � 0.09 mM, respectively, when the concentration of GSH wasfixed at 5 mM, and were 122 � 2.08 mmol/mg/min and1.52 � 0.17 mM, respectively, when the CDNB concentration fixedat 3 mM; for the SlGSTE3 recombinant protein, Vmax and Km valueswere 111 � 2.19 mmol/mg/min and 1.01 � 0.1 mM, respectively,when the concentration of GSH was fixed at 5 mM, and were106 � 2.34 mmol/mg/min and 1.54 � 0.21 mM, respectively, when
Fig. 5. Northern blot (A) and western blot (B and C) analyses of temporal and spatial expre
tissues at different stages was used in each of the lanes. The blots were hybridized with
extracts were plotted with anti-SlGSTE2 and anti-SlGSTE3 antibody (B and C). Twenty
midgut; E: epidermis; F: fatty body; L: larvae; D: day; H: hour; MG: midgut; EP: epidermi
the lanes in the western blots.
the CDNB concentration fixed at 3 mM (Table 1). The apparent Vmaxvalues for glutathione and CDNB appeared to be similar, while theapparent Km value for glutathione was lower than that for CDNB forboth of the two recombinant proteins. These results suggest thatSlGSTE2 and SlGSTE3 had higher affinities for CDNB than forglutathione. This result was consistent with the observations onother insect GSTs (Clark, 1989).
The insecticides DDT and deltamethrin, were tested for theirability to competitively inhibit the enzyme activity of SlGSTE2and SlGSTE3 towards CDNB. The assays showed that at 100 mMconcentration, both DDT and deltamethrin significantly inhibitedenzyme activity of SlGSTE2 against CDNB at 2 mM; while onlyDDT inhibited the activity of SlGSTE3 against CDNB (Fig. 4).These data indicated that the SlGSTE2 recombinant proteinpreferably bound to the insecticides DDT and deltamethrin thatwere at a much lower concentration than the common substrateCDNB.
3.6. Developmental expression of Slgste2 and Slgste3
Developmental expression of Slgste2 and Slgste3 genes indifferent tissues during larval stages was examined using northernblot analysis. Single mRNA transcript was detected for Slgste2 orSlgste3 and the sizes of the transcripts were consistent with the826-nucleotide Slgste2 cDNA and the 726-nucleotide Slgste3 cDNA,respectively. The mRNAs of both of Slgste2 and Slgste3 weredetected in the midgut of 5th instar and of the 6th instar prior toprepupal stage, e.g. L6D4 (Fig. 5A), after which the mRNA levels ofSlgste2 decreased and were not detectable during the pupae stage.Similarly, the mRNA of Slgste3 was not detected after 3 days afterecdysis into 6th instar stage. Low levels of Slgste2 mRNA weredetected in the fat body during the 5th and the early stages of 6thinstar (Fig. 5A). No mRNA of Slgste3 was detected in the fat bodythroughout the 5th and 6th instar stages. In the epidermis, nomRNA of Slgste2 and Slgste3 were detected during all of the stagestested (Fig. 5A).
ssion of Slgste2 and Slgste3. For northern blot, 10 mg aliquot of total RNA from various32P-dATP-labeled cDNA probe of Slgste2 or Slgste3. In the western blots, the protein
grams were loaded per lane on 12% SDS-PAGE gels. L: larvae; D: day; H: hour; M:
s; FB: fat body; P: pupae. Twenty micrograms of total proteins were loaded in each of
H. Deng et al. / Journal of Insect Physiology 55 (2009) 1174–11831180
Protein levels of the Slgste2 and Slgste3 genes in the three tissuesat different stages were examined by western blot analysis (Fig. 5Band C). SlGSTE2 protein was present in the midgut and fat body, butno protein was found in the epidermis on day 1 of 5th and 6thinstar stages; while SlGSTE3 protein was mainly present in themidgut and no protein was detected in the fat body and epidermis(Fig. 5B). In the midgut, SlGSTE2 and SlGSTE3 proteins were mainlypresent during the 5th instar stage and the early stages (e.g. priorto day 3 or 4) of 6th instar stage (Fig. 5C).
The spatial expression patterns of mRNA and proteins indicatedthat Slgste2 was expressed in midgut and fat body, while Slgste3
was specifically expressed in the midgut. The results of temporalexpression also indicated that Slgste2 was highly expressed in themidgut during the 5th instar stage and the early 6th instar, whileSlgste3 was present at higher levels during the transition from 5thto 6th instar stages and decreased to an undetectable level earlierthan Slgste2 at later stages of 6th instar.
Fig. 6. Induced expression of Slgste2 (A and B) and Slgste3 (A and C) genes by the carbaryl, D
extracted from 3rd instar larvae at 6, 12, 24, 48 h and 72 h post-treatment. The data in (B
were quantified using ImageQuant TL v2005 (Amersham Inc., USA). The controls were the
is used as a negative control in the RT-PCR reactions and no responsiveness was detected
presented. The asterisks indicate significant difference (p < 0.05) between the treatment a
3.7. Induction of Slgste2 and Slgste3 expression by insecticides
A bioinsecticide, B. thuringiensis (Bt), and five syntheticchemical insecticides, carbaryl, DDT, RH5992, malathion anddeltamethrin, were tested for their effects on expression of Slgste2
and Slgste3 in the 3rd instar by RT-PCR (Fig. 7). Expression of Slgste2
mRNA was increased by the treatments of Bt, carbaryl, DDT,deltamethrin and RH5992 between 6 and 72 h post-treatments(Fig. 6A and B). The induced expression levels of Slgste2 were moresignificant for carbaryl, deltamethrin and DDT than for RH5992and Bt. Malathion did not induce the expression of Slgste2 gene atthe tested dose. Among the tested chemicals, DDT enhanced theSlgste2 expression by 10-fold higher than the control, whilecarbaryl enhanced expression by 4-fold and Bt, RH5992 anddeltamethrin increased 3-fold higher than the control at 72 h post-treatment. The expression of Slgste3 was not induced by Bt,carbaryl, RH5992, malathion and deltamethrin, while DDT
DT, RH5992, malathion and deltamethrin at LD30 in 3rd instar larvae. Total RNA was
) and (C) are the means of three replicated experiments of RT-PCR. The PCR products
samples taken at the same times post-mock treatment of acetone. Actin expression
to the tested chemical insecticides. Mean � SD for three independent experiments are
nd control.

Fig. 6. (Continued ).
H. Deng et al. / Journal of Insect Physiology 55 (2009) 1174–1183 1181
treatment induced the Slgste3 expression to some extent (Fig. 6Aand C). These data suggest that Slgste2 was more sensitive to theinsecticide treatments than Slgste3.
3.8. Immunohistochemistry localization of SlGSTE2 and SlGSTE3
proteins
Immunohistochemistry localization analysis of SlGSTE2 andSlGSTE3 in one-day-old 5th instar larvae showed that both of theproteins were predominately present in the midgut (Fig. 7). Thisresult was consistent with the data obtained by northern andwestern blot analyses. The SlGSTE2 protein was also found in thefat body and tracheal system, while the SlGSTE3 protein was alsopresent in the tracheal system.
4. Discussion
In this study, two epsilon class GST cDNAs, Slgste2 and Slgste3
were cloned from S. litura and characterized in terms of structure,developmental and induced expression and localization. Severalnovel findings are reported. First, the SlGSTE2 and SlGSTE3proteins are identified as members of the typical epsilon class ofGSTs based on their structure features and relationship with otherepsilon insect GSTs. A serine residue which is catalytically essentialfor the activity of GSTs as reported by Sheehan et al. (2001), wasfound in the N-terminus regions of SlGSTE2 (Ser11) and SlGSTE3(Ser5). This result indicates that there is more than one epsilonGSTs in S. litura.
Secondly, both the SlGSTE2 and SlGSTE3 recombinant proteinsprocessed enzymatic activity against the standard substrate CDNB.DDT and deltamethrin competitively inhibited the enzymaticactivity of SlGSTE2 toward CDNB by 65% and 34%, respectively.There was a slight effect of DDT and no effect of deltamethrin on
SlGSTE3 activity toward CDNB. It was reported that most of thecharacterized epsilon GSTs have low levels of activity towards themodel substrate, CDNB (Ranson and Hemingway, 2005). Five out ofthe eight epsilon classes of GSTs in a DDT resistant strain of A.
gambiae have elevated expression levels of the GSTs. AgGSTe2 isthe only member of the epsilon class with confirmed DDT-metabolizing activity and it is considered the most important GSTin conferring DDT resistance in A. gambiae (Ding et al., 2003; Ortelliet al., 2003). DDT-dehydrochlorinase activity catalyzed by GST isthe major mechanism responsible for DDT resistance in insects(Brown, 1986). The contributing residues for putative DDT-bindingincludes Leu9, Leu11, Ser12, Pro13, Pro14, Leu36, Leu37, His41,Ile55, Phe108, Met111, Phe115, Leu119, Phe120, Leu207 andPhe210, most of which are hydrophobic in nature. Three residues,Glu116, Phe120, and Arg112 are proposed as the key players ofcreating a DDT-binding pocket with high affinity for DDT (Wanget al., 2008). Compared with AgGSTE2, both SlGSTE2 and SlGSTE3had seven amino acid residues in the putative DDT-binding.SlGSTE2 shared one key residue, Phe120, at the correspondingposition in the DDT-binding pocket (Wang et al., 2008), whereasSlGSTE3 did not contain any of these key residues. These datasuggest that SlGSTE2 has higher DDT-metabolizing activity thanSlGSTe3 and more likely plays a role in conferring insecticidedetoxification.
Thirdly, several lines of evidence indicate that Slgste2 probablyplays more important roles than Slgste3 in the insecticideresistance in S. litura. Slgste2 appeared to have higher expressionlevels during larval development and more sensitive to insecticidetreatments than Slgste3. Six insecticides were used at a LC30 doseto induce Slgste2 and Slgste3 expression in order to examine theresponses of these two genes to the insecticide treatments. Theseinsecticides are quite different in structure, chemical properties,toxicity and mode of action. DDT induced Slgste2 expression about

Fig. 7. Immunohistochemistry localization of SlGSTE2 (A and B) and SlGSTE3 (E and F) in 1-day-old 5th instar larvae. Five-micron cross-sections were immunostained with
anti-SlGSTE2 (A and B) or anti-SlGSTE3 (E and F), or pre-immune serum from the same rabbit prior to immunization as negative controls (C and D, G and H), followed by goat
anti-rabbit IgG conjugated with SABC-FITC and counter-stained with DAPI. The sections were observed by fluorescence microscopy and the photographs were taken as double
exposure with fluorescein (A, C, E and G) and DAPI filters (B, D, F and H). FB: fat body; MG: midgut; EP: epidermis; CT: cuticle; TC: tracheal. The bars represent 30 mm.
H. Deng et al. / Journal of Insect Physiology 55 (2009) 1174–11831182
10-fold over the control, while carbaryl, RH59924, deltamethrinand Bt increased the expression by 3–4-fold over the control.Slgste3 expression was slightly increased by carbaryl, Bt and DDT,but not affected by RH5992, malathion and deltamethrin. Inaddition, as described above, the recombinant SlGSTE2 protein hadhigher affinity to DDT and deltamethrin than SlGSTE3. Althoughhighly induced expression of GST genes is not always related toresistance, but most studies propose a positive relationshipbetween induced over-expression of GST and insecticide resistancein most species. Tang and Tu (1994) found that DmGSTD1 inducedover-expression is a major mechanism underlying DDT resistancein the PSU-R strain of Drosophila melanogaster. There are four GSTgenes (PxGSTe1, AgGSTe2, AaGSTd1 and AaGSTe2) that have beenshown to be involved in insecticide resistance and their inducedover-expression is the major mechanism for the resistant strains(Sonoda and Tsumuki, 2005; Prapanthadara et al., 1998; Grant andHammock, 1992). Most of GSTs that play roles in insecticideresistance belong to the insect-specific delta or epsilon class (Liet al., 2007), although not all delta or epsilon GSTs are shown to beinvolved in insecticide resistance.
Fourthly, the spatial and temporal expression of Slgste2 andSlgste3 by analyses of northern blot, western blot and immuno-histochemistry revealed that the Slgste2 mRNA and protein mainlypresent in the midgut, as well as the fat body in a relatively low
level; Slgste3 was only present in the midgut. This is consistentwith the results for other lepidopteran GSTs, in which GSTs aredetected mainly in the midgut and the fat body for detoxification ofxenobiotics (Feng et al., 1999; Snyder et al., 1995). Constitutive andhigh-level expressions of individual genes are often detected inresistant strains of many species. However, high levels ofconstitutive expression of GST genes are not always associatedwith insecticide resistance. For example, in B. mori, 23 GST geneshave been found and most of the noninsect-specific GSTs showedhigh constitutive expression in various tissues, whereas the insect-specific delta and epsilon GSTs showed little or no constitutiveexpression in the tissues, such as the fat body, but the expression ofthese genes was induced by insecticides and played a role ininsecticide resistance (Yu et al., 2008). In addition, in this study,Slgste2 mRNA and protein were present in the fat body at lowlevels, but were induced after exposure to DDT, carbaryl, RH59924,deltamethrin and Bt, implying that it may need to be induced toplay a role in detoxification of these insecticides.
Fifthly, no introns for GST genes have been reported in manyspecies, for example, BmGSTe2 and the D. melanogaster epsilon GSTgenes were intronless (Yu et al., 2008; Sawicki et al., 2003). Theintronless feature may contribute to selective advantage understress condition such as heat shock (D’Onofrio et al., 1991). P.
xylostella GSTe1 was a single-copy gene in the genome, which is

H. Deng et al. / Journal of Insect Physiology 55 (2009) 1174–1183 1183
over-expressed in the OP resistant strain (Huang et al., 1998).Slgste2 and Slgste3 are both intronless and represented by a single-copy in the genome. This is consistent with the observations onmost of insect GST genes in other species.
In summary, two epsilon GSTs cDNA Slgste2 and Slgste3 werecloned from S. litura and characterized in terms of sequencestructure and comparison, developmental and induced expression,as well as localization. The results suggest that the expression ofthe Slgste2 and Slgste3 genes was induced by some of theinsecticides tested and they may be associated with detoxificationof these insecticides. Further investigation is necessary to studytheir mechanisms in insecticide resistance.
Acknowledgments
This research was supported by the Chinese National NaturalScience Foundation (Grant no.: 30671408) and Guangdong ProvinceNatural Science Foundation of (Grant nos.:9151063101000009and 06105204).
References
Altschul, S.F., Warren, G., Webb, M., Myers, E.W., Ligman, D.J., 1990. Basic localalignment search tool. Journal of Molecular Biology 215, 403–410.
Armes, N.J., Wightman, J.A., Jadhav, D.R., Ranga Rao, G.V., 1997. Status of insecticideresistance in Spodoptera litura in Andhra Pradesh, India. Pesticide Science 50,240–248.
Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of micro-gram quantities of protein utilizing the principle of protein-dye binding.Analytical Biochemistry 72, 248–254.
Brown, A.W., 1986. Insecticide resistance in mosquitoes: a pragmatic review.Journal of the American Mosquito Control Association 2, 123–140.
Chelvanayagam, G., Parker, M.W., Board, P.G., 2001. Fly fishing for GSTs: a unifiednomenclature for mammalian and insect glutathione transferases. Chemico-Biological Interactions 133, 256–260.
Chomyczynski, P., Sacchi, N., 1987. Methods of RNA isolation by acid guanidiniumthiocyanate–phenol–chloroform extraction. Analytical Biochemistry 162, 156–159.
Clark, A.G., 1989. The comparative enzymology of the glutathione S-transferasesfrom non-vertebrate organisms. Comparative Biochemistry and Physiology B92, 419–446.
Clark, A.G., Shamaan, N.A., 1984. Evidence that DDT dehydrochlorinase from thehouse fly is a glutathione S-transferase. Pesticide Biochemistry and Physiology22, 249–261.
D’Onofrio, M., Lee, M.D., Starr, C.M., Miller, M., Hanover, J.A., 1991. The geneencoding rat nuclear pore glycoprotein p62 is intronless. Journal of BiologicalChemistry 266, 664–666.
Ding, Y., Ortelli, F., Rossiter, L.C., Hemingway, J., Ranson, H., 2003. The Anophelesgambiae glutathione transferase supergene family: annotation, phylogeny andexpression profiles. BMC Genomics 4, 35–50.
Feng, Q.L., Davey, K.G., Pang, A.S., Primavera, M., Ladd, T.R., Zheng, S.C., Retnakaran, A.,Palli, S.R., 1999. Glutathione S-transferase from the spruce budworm, Choristo-neura fumiferana: identification, characterization, localization, cDNA cloning andexpression. Insect Biochemistry and Molecular Biology 29, 779–793.
Feng, Q., Davey, K.G., Pang, A.S., Ladd, T.R., Retnakaran, A., Tomkins, B.L., Zheng, S.C.,Palli, S.R., 2001. Developmental expression and stress induction of glutathioneS-transferase in the spruce budworm, Choristoneura fumiferana. Journal ofInsect Physiology 47, 1–10.
Grant, D.F., Hammock, B.D., 1992. Genetic and molecular evidence for a trans-actingregulatory locus controlling glutathione S-transferase-2 expression in Aedesaegypti. Molecular and Biochemical Parasitology 234, 169–176.
Hayes, J.D., Pulford, D.J., 1995. The glutathione S-transferase supergene family:regulation of GST and the contribution of the isoenzymes to cancer chemo-protection and drug resistance. Critical Reviews in Biochemistry and MolecularBiology 30, 445–600.
Huang, X., Maddan, A., 1999. CAP3: a DNA sequence assembly program. GenomeResearch 9, 868–877.
Huang, H.S., Hu, N.T., Yao, Y.E., Wu, C.Y., Chiang, S.W., 1998. Molecular cloning andheterologous expression of a glutathione S-transferase involved in insecticideresistance from the diamondback moth, Plutella xylostella. Insect Biochemistryand Molecular Biology 28, 651–658.
Lehrach, H., Diamond, D., Wozney, J.M., Boedtker, H., 1977. RNA molecular weightdetermination by gel electrophoresis under denaturing conditions, a criticalreexamination. Biochemistry 16, 4743–4751.
Lewis, J.B., Sawicki, R.M., 1971. Characterization of the resistant mechanism todiazinon, parathion and diazoxon in the organophosphorous resistant SKAstrain of house flies (Musca domestica L.). Pesticide Biochemistry and Physiology1, 275–285.
Li, X., Schuler, M.A., Berenbaum, M.R., 2007. Molecular mechanisms of metabolicresistance to synthetic and natural xenobiotics. Annual Review of Entomology52, 231–253.
Ortelli, F., Rossiter, L.C., Vontas, J., Ranson, H., Hemingway, J., 2003. Heterologousexpression of four glutathione transferase genes genetically linked to a majorinsecticide-resistance locus from the malaria vector Anopheles gambiae. Bio-chemical Journal 373, 957–963.
Prapanthadara, L., Ranson, H., Somboon, P., Hermingway, J., 1998. Cloning, expres-sion and characterization of an insect class I glutathione S-transferase fromAnopheles gambiae. Journal of Biological Chemistry 272, 5464–5468.
Ranson, H., Hemingway, J., 2005. Mosquito glutathione S-transferases. Methods inEnzymology 401, 226–241.
Ranson, H., Rossiter, L., Ortelli, F., Jesen, B., Wang, X., Roth, C.W., Collins, F.H.,Hemingway, J., 2001. Identification of a novel class of insect glutathione S-transferases involved in resistance to DDT in the malaria vector Anophelesgambiae. Biochemical Journal 359, 295–304.
Sambrook, J., Fritsch, E.F., Maniatis, T., 1989. Molecular Cloning—A LaboratoryManual, 2nd ed. Cold Spring Harbor Laboratory Press, New York.
Sawicki, R., Singh, S.P., Mondal, A.K., Benes, H., Zimniak, P., 2003. Cloning, expressionand biochemical characterization of one epsilon class (GST-3) and ten delta-class (GST-1) glutathione S-transferase from Drosophlia melanogaster, andidentification of additional nine members of the epsilon class. BiochemicalJournal 370, 661–669.
Sheehan, D., Meade, G., Foley, V.M., Dowd, C.A., 2001. Structure, function andevolution of glutathione transferases: implications for classification non-mam-malian members of an ancient enzyme superfamily. Biochemical Journal 360,1–16.
Snyder, M.J., Walding, J.K., Feyerreisen, R., 1995. Glutathione S-transferases fromlarval Manduca sexta midgut: sequence of two cDNAs and enzyme induction.Insect Biochemistry and Molecular Biology 25, 455–465.
Sonoda, S., Tsumuki, H., 2005. Studies on glutathione S-transferase gene involvedin chlorfluazuron resistance of the diamondback moth, Plutella xylostella L.(Lepidoptera: Yplutellisae). Pesticide Biochemistry and Physiology 82,94–101.
Tang, A.H., Tu, C.P., 1994. Biochemical characterization of Drosophila glutathione S-transferases D1 and D21. Journal of Biological Chemistry 269, 27876–27884.
Vontas, J.G., Small, G.J., Hemingway, J., 2001. Glutathione S-transferases as anti-oxidant defence agents confer pyrethroid resistance in Nilaparvata lugens.Biochemical Journal 357, 65–72.
Vontas, J.G., Hejazi, M.J., Hawkes, N.J., Cosmidis, N., Loukas, M., Janes, R.W., Hemi-ngway, J., 2002. Resistance-associated point mutations of organophosphateinsensitive acetylcholinesterase, in the olive fruit fly Bactrocera oleae. InsectMolecular Biology 11, 329–336.
Wang, Y., Qiu, L., Ranson, H., Lumjuan, N., Hemingway, J., Setzer, W.N., Meehan,E.J., Chen, L., 2008. Structure of an insect epsilon class glutathione S-transfer-ase from the malaria vector Anopheles. Journal of Structural Biology 164,228–235.
Wu, M.C., Lu, K.H., 2008. Juvenile hormone induction of glutathione s-transferasesactivity in the larval fat body of the common cutworm, Spodoptera litura(Lepidoptera:Noctuidae). Archives of Insect Biochemistry and Physiology 68,232–240.
Yamamoto, K., Nagaoka, S., Banno, Y., Aso, Y., 2008. Biochemical properties of anomega-class glutathione S-transferase of the silkmoth, Bombyx mori. Compara-tive Biochemistry and Physiology C: Pharmacology Toxicology and Endocrinol-ogy 149, 461–467.
Yu, Q.Y., Lu, C., Li, B., Fang, S., Zou, W.D., Dai, F.Y., Zhang, Z., Xiang, Z.H., 2008.Identification, genomic organization and expression pattern of glutathione S-transferase in the silkworm, Bombyx mori. Insect Biochemistry and MolecularBiology 38, 1158–1164.
Zheng, S.C., Deng, H.M., Ladd, T., Tomkins, B.L., Krell, P.J., Feng, Q.L., 2007. Cloningand characterization of two glutathione S-transferase cDNAs in the sprucebudworm, Choristoneura fumiferana. Archives of Insect Biochemistry and Phy-siology 66, 146–157.


















