
Trichostatin A (TSA) improves the development of rabbit-rabbit ...
9
RESEARCH ARTICLE Trichostatin A (TSA) Improves the Development of Rabbit-Rabbit Intraspecies Cloned Embryos, But Not Rabbit-Human Interspecies Cloned Embryos Li-Hong Shi, 1,2 Yi-Liang Miao, 1,2 Ying-Chun Ouyang, 1 Jun-Cheng Huang, 1,2 Zi-Li Lei, 1,2 Ji-Wen Yang, 1,2 Zhi-Ming Han, 1 Xiang-Fen Song, 1 Qing-Yuan Sun, 1 and Da-Yuan Chen 1 * The interspecies somatic cell nuclear transfer (iSCNT) technique for therapeutic cloning gives great promise for treatment of many human diseases. However, the incomplete nuclear reprogramming and the low blastocyst rate of iSCNT are still big problems. Herein, we observed the effect of TSA on the development of rabbit–rabbit intraspecies and rabbit– human interspecies cloned embryos. After treatment with TSA for 6 hr during activation, we found that the blastocyst rate of rabbit–rabbit cloned embryos was more than two times higher than that of untreated embryos; however, the blastocyst rate of TSA-treated rabbit– human interspecies cloned embryos decreased. We also found evident time-dependent histone deacetylation-reacetylation changes in rabbit–rabbit cloned embryos, but not in rabbit– human cloned embryos from fusion to 6 hr after activation. Our results suggest that TSA-treatment does not improve blastocyst development of rabbit– human iSCNT embryos and that abnormal histone deacetylation-reacetylation changes in iSCNT embryos may account for their poor blastocyst development. Developmental Dynamics 237:640 – 648, 2008. © 2008 Wiley-Liss, Inc. Key words: histone acetylation; embryos; intraspecies nuclear transfer; interspecies nuclear transfer; TSA Accepted 16 December 2007 INTRODUCTION The success of somatic cell cloning has shed light on species preservation, livestock propagation, and therapeu- tic cloning (Wakayama et al., 2001, 2005a,b; Tecirlioglu et al., 2006). Therapeutic cloning, which is used to generate patient-specific embryonic stem cells (ESCs) from nuclear trans- fer (NT) blastocysts, gives great prom- ise for regenerative medicine. How- ever, therapeutic cloning faces the ethical dispute and is limited by the availability of human oocytes in prac- tice. To secure human blastocysts and solve the limited resource of human oocytes, an interspecies somatic cell nuclear transfer (iSCNT) technique using human somatic cell and animal enucleated oocyte was suggested. The feasibility of interspecies cloning was demonstrated in previous studies, and blastocyst formation was possible af- ter transfer of mouse, bovine, porcine, rabbit, or human somatic cells into bo- vine and rabbit enucleated oocytes (Corley-Smith and Brandhorst, 1999; Dominko et al., 1999; Lanza et al., 2000; Kitiyanant et al., 2001; Chen et al., 2002; Saikhun et al., 2002). Al- though iSCNT offspring gaur (Lanza et al., 2000), mouflon (Loi et al., 2001), and African wild cats (Gomez et al., 2004) have been obtained from close- related species and iSCNT-ESCs were derived from un-related interspecies cloned blastocysts (Chen et al., 2003), the incomplete nuclear reprogram- ming and the low blastocyst rate of iSCNT are still big problems. Recently molecular analysis has elucidated that cloned embryos have 1 State Key Laboratory of Reproductive Biology, Institute of Zoology, Chinese Academy of Sciences, Beijing, China 2 Graduate School, Chinese Academy of Sciences, Beijing, China Grant sponsor: Special Funds for Major State Basic Research Projects; Grant number: 2001CB509905; Grant sponsor: Knowledge Innovation Project of Chinese Academy of Sciences; Grant number: KSCX2-YW-N-019. *Correspondence to: Da-Yuan Chen, State Key Laboratory of Reproductive Biology, Institute of Zoology, Chinese Academy of Sciences no. 5 Da-tun Road, Chaoyang, Beijing100101, PR China. E-mail: [email protected] DOI 10.1002/dvdy.21450 Published online 7 February 2008 in Wiley InterScience (www.interscience.wiley.com). DEVELOPMENTAL DYNAMICS 237:640 – 648, 2008 © 2008 Wiley-Liss, Inc.
Transcript of Trichostatin A (TSA) improves the development of rabbit-rabbit ...

RESEARCH ARTICLE
Trichostatin A (TSA) Improves theDevelopment of Rabbit-Rabbit IntraspeciesCloned Embryos, But Not Rabbit-HumanInterspecies Cloned EmbryosLi-Hong Shi,1,2 Yi-Liang Miao,1,2 Ying-Chun Ouyang,1 Jun-Cheng Huang,1,2 Zi-Li Lei,1,2
Ji-Wen Yang,1,2 Zhi-Ming Han,1 Xiang-Fen Song,1 Qing-Yuan Sun,1 and Da-Yuan Chen1*
The interspecies somatic cell nuclear transfer (iSCNT) technique for therapeutic cloning gives great promise fortreatment of many human diseases. However, the incomplete nuclear reprogramming and the low blastocystrate of iSCNT are still big problems. Herein, we observed the effect of TSA on the development of rabbit–rabbitintraspecies and rabbit–human interspecies cloned embryos. After treatment with TSA for 6 hr duringactivation, we found that the blastocyst rate of rabbit–rabbit cloned embryos was more than two times higherthan that of untreated embryos; however, the blastocyst rate of TSA-treated rabbit–human interspecies clonedembryos decreased. We also found evident time-dependent histone deacetylation-reacetylation changes inrabbit–rabbit cloned embryos, but not in rabbit–human cloned embryos from fusion to 6 hr after activation. Ourresults suggest that TSA-treatment does not improve blastocyst development of rabbit–human iSCNT embryosand that abnormal histone deacetylation-reacetylation changes in iSCNT embryos may account for their poorblastocyst development. Developmental Dynamics 237:640–648, 2008. © 2008 Wiley-Liss, Inc.
Key words: histone acetylation; embryos; intraspecies nuclear transfer; interspecies nuclear transfer; TSA
Accepted 16 December 2007
INTRODUCTION
The success of somatic cell cloning hasshed light on species preservation,livestock propagation, and therapeu-tic cloning (Wakayama et al., 2001,2005a,b; Tecirlioglu et al., 2006).Therapeutic cloning, which is used togenerate patient-specific embryonicstem cells (ESCs) from nuclear trans-fer (NT) blastocysts, gives great prom-ise for regenerative medicine. How-ever, therapeutic cloning faces theethical dispute and is limited by theavailability of human oocytes in prac-
tice. To secure human blastocysts andsolve the limited resource of humanoocytes, an interspecies somatic cellnuclear transfer (iSCNT) techniqueusing human somatic cell and animalenucleated oocyte was suggested. Thefeasibility of interspecies cloning wasdemonstrated in previous studies, andblastocyst formation was possible af-ter transfer of mouse, bovine, porcine,rabbit, or human somatic cells into bo-vine and rabbit enucleated oocytes(Corley-Smith and Brandhorst, 1999;Dominko et al., 1999; Lanza et al.,
2000; Kitiyanant et al., 2001; Chen etal., 2002; Saikhun et al., 2002). Al-though iSCNT offspring gaur (Lanzaet al., 2000), mouflon (Loi et al., 2001),and African wild cats (Gomez et al.,2004) have been obtained from close-related species and iSCNT-ESCs werederived from un-related interspeciescloned blastocysts (Chen et al., 2003),the incomplete nuclear reprogram-ming and the low blastocyst rate ofiSCNT are still big problems.
Recently molecular analysis haselucidated that cloned embryos have
1State Key Laboratory of Reproductive Biology, Institute of Zoology, Chinese Academy of Sciences, Beijing, China2Graduate School, Chinese Academy of Sciences, Beijing, ChinaGrant sponsor: Special Funds for Major State Basic Research Projects; Grant number: 2001CB509905; Grant sponsor: Knowledge InnovationProject of Chinese Academy of Sciences; Grant number: KSCX2-YW-N-019.*Correspondence to: Da-Yuan Chen, State Key Laboratory of Reproductive Biology, Institute of Zoology, Chinese Academy ofSciences no. 5 Da-tun Road, Chaoyang, Beijing100101, PR China. E-mail: [email protected]
DOI 10.1002/dvdy.21450Published online 7 February 2008 in Wiley InterScience (www.interscience.wiley.com).
DEVELOPMENTAL DYNAMICS 237:640–648, 2008
© 2008 Wiley-Liss, Inc.

abnormal epigenetic modifications in-cluding DNA methylation and histonemodifications (Dean et al., 2001; Kanget al., 2001; Santos et al., 2003;Ohgane et al., 2004). Histone modifi-cations including acetylation, phos-phorylation, methylation, and ubiq-uitination played a key role in post-translational modifications (Fischle etal., 2003). The global and loci patternsof histone acetylation contributed sig-nificantly to nuclear reprogramming(Turner, 2000). Histone acetylationhas the greatest potential for unfold-ing chromatin to recruit differenttranscriptional factors, and thus acet-ylation is related to the active genes,and removal of acetylated groups byhistone deacetylases (HDACs) is gen-erally associated with gene silencing(Gao et al., 2002; Adcock and Lee,2006). Therefore, improper patterns ofhistone acetylation might severely af-fect the development of cloned em-bryos.
There have been several reports onchanges of histone acetylation in ei-ther donor somatic cells or NT em-bryos after treatment with TSA,which is a reversible and specific his-tone deacetylase inhibitor. Treatmentof donor cells before NT with TSA orsodium butyrate (NaBu), anotherHDAC inhibitor, improved the effi-ciency of the cloning (Enright et al.,2003; Yang et al., 2007). Additionally,TSA-treated cloned embryos followingactivation developed to blastocyststage at a higher rate, and the blasto-cyst quality and the subsequent full-term development were greatly im-proved, too (Rybouchkin et al., 2006).Furthermore, the establishment ratesof NT-ESCs from TSA-treated clonedblastocysts following activation werethree times higher than those of un-
treated groups (Kishigami et al.,2006). The global loci patterns of his-tone acetylation were involved in thenuclear reprogramming.
Here, we aimed to clarify the effectof TSA on development of rabbit–rab-bit intraspecies and rabbit–human in-terspecies cloned embryos. We foundthat TSA had a dramatic effect on rab-bit–rabbit cloned embryo develop-ment, while this drug had little effecton rabbit–human cloned embryo de-velopment. In addition, we observedthe dynamic changes of histone acety-lation in rabbit–rabbit cloned embryosand rabbit–human interspecies clonedembryos.
RESULTS
Effect of TSA on theDevelopment ofRabbit–Rabbit SCNTEmbryos to Blastocyst Stage
To evaluate the effect of TSA on rab-bit–rabbit and rabbit–human clonedembryos, we treated the activated em-bryos for 6 hr with 100 nM TSA. Al-though TSA-treated rabbit–rabbitcloned embryos developed to 2- and4-cell stage at a similar rate as un-treated groups, the development tomorula and blastocyst stage increasedfrom 26.42 and 22.64% to 47.06 and45.01%, respectively (Table 1).
Effect of TSA on theDevelopment ofRabbit–Human iSCNTEmbryos to Blastocyst Stage
In rabbit–human interspecies clonedembryos, however, the developmentalrate at various stages in the TSA-
treated group was not statistically dif-ferent from that of the untreatedgroup, but even a bit lower (P � 0.05)(Table 2). To rule out the toxicity ofthe long-term TSA-treatment, weshortened the TSA-treated time to 3hr following activation. TSA-treatedembryos increased a little in morulaand blastocyst rate, but there were nosignificant differences between TSA-treated and control groups (Table 3).
Dynamic Changes of HistoneAcetylation in Rabbit–RabbitIntraspecies SCNT EmbryosFrom Fusion to 6 hr AfterActivation
Previous studies have shown that thehistone acetylation level decreasedmarkedly in the somatic nuclei whenthey were transferred into the enucle-ated oocytes (Kim et al., 2003; Ry-bouchkin et al., 2006; Wang et al.,2007). To address this view in SCNTand iSCNT embryos, we observed thedynamic changes of histone acetyla-tion in SCNT and iSCNT embryosfrom fusion to 6 hr after activation.
As shown in Figure 1, slightdeacetylation of AcH4/K12 occurred inrabbit donor nuclei from 0.5 to 1.5 hrafter fusion (8 of 10 and 7 of 12, P �0.05), whereas reacetylation tookplace abruptly 3 hr after fusion, withrapid changes in nuclear size (10 of12) (Fig. 1A and B, P � 0.05). Duringactivation, with the pseudo-pro-nuclear formation, the staining ofAcH4/K12 kept increasing from 3 hr to6 hr without TSA-treatment (6 of 8and 7 of 10, respectively) (Fig. 1A andB). At 3 and 6 hr after activation, com-pared with the untreated intraspeciescloned embryos, the staining intensity
TABLE 1. Development of Rabbit–Rabbit Intraspecies Cloned Embryos After Treatment With TSA for 6 hra
TreatmentNo. ofoocytes
No. ofreconstructedoocytes
No. ofembryosfused (%)
No. of 2-cellembryos (%)
No. of 4-cellembryos (%)
No. ofmorula(%)
No. ofblastocysts(%)
Non-treated 176 173 106(61.27)a
77(72.64)a
56(52.83)a
28(26.42)a
24(22.64)b
TSA6hr (100 nM) 168 166 102(61.45)a
63(61.76)a
59(57.84)a
48(47.06)b
46(45.01)b
aValues with different superscripts within the same column differ significantly (P � 0.05). Note: Fused (%), fusedoocytes/reconstructed oocytes; development rate of embryos at different stages, number of embryos/number of fused embryos.
EFFECT OF TSA ON INTERSPECIES CLONED EMBRYOS 641
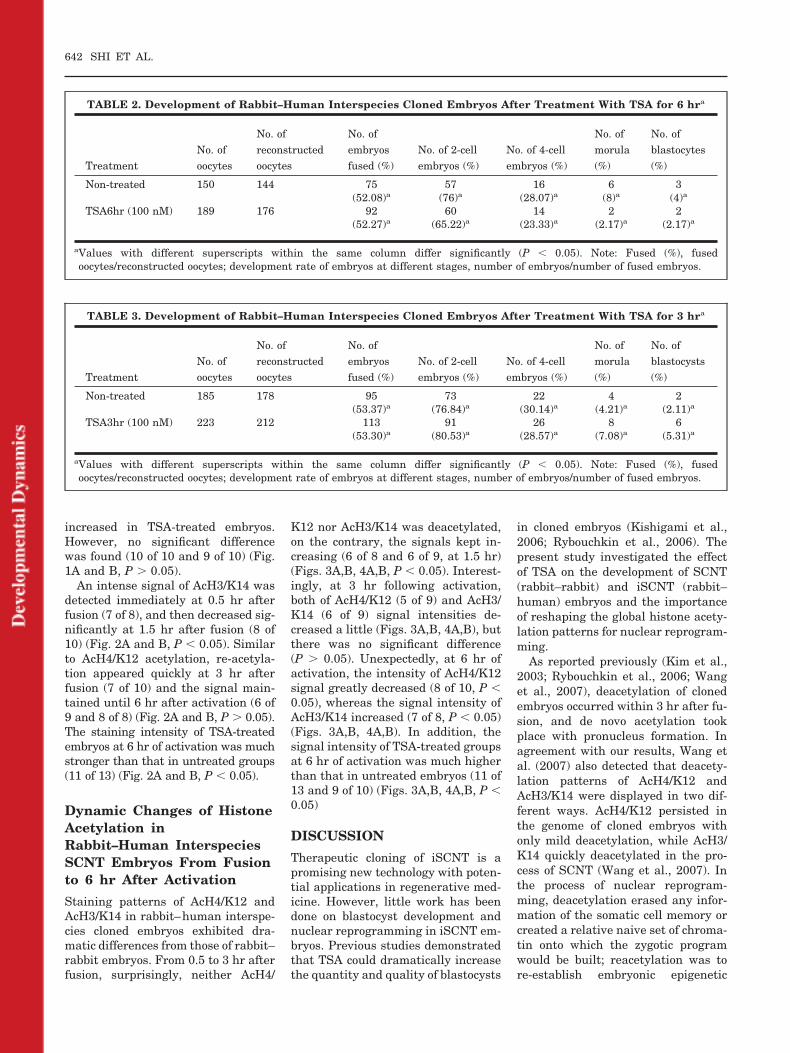
increased in TSA-treated embryos.However, no significant differencewas found (10 of 10 and 9 of 10) (Fig.1A and B, P � 0.05).
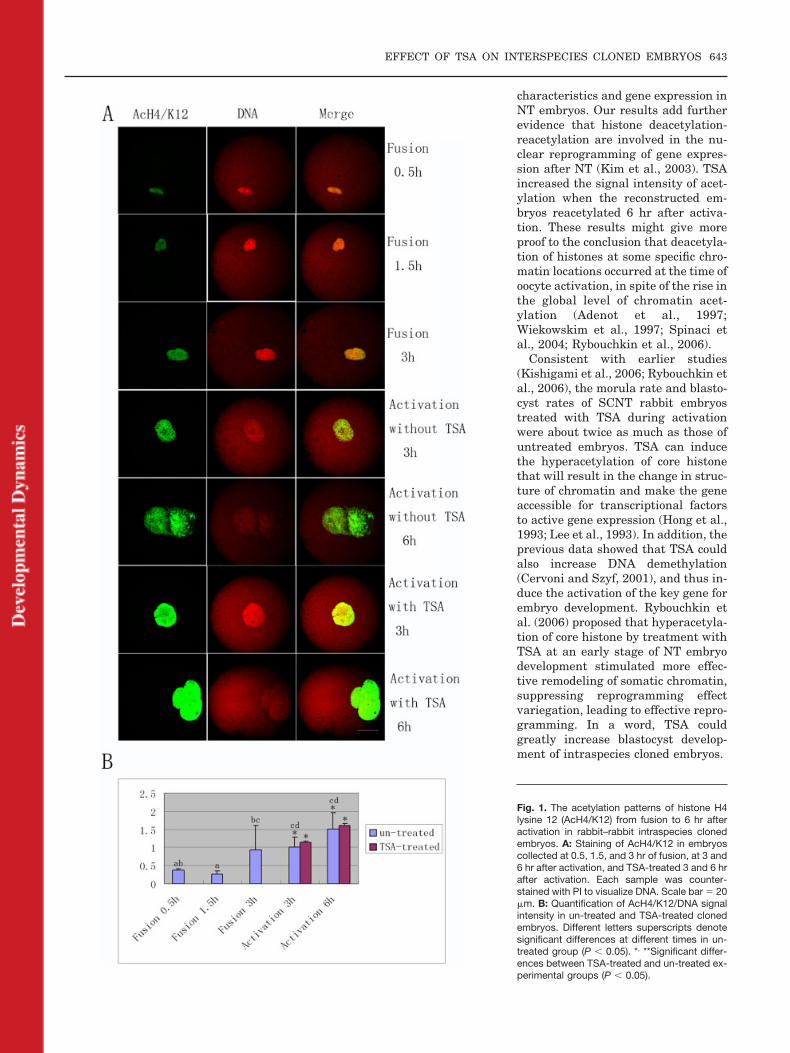
An intense signal of AcH3/K14 wasdetected immediately at 0.5 hr afterfusion (7 of 8), and then decreased sig-nificantly at 1.5 hr after fusion (8 of10) (Fig. 2A and B, P � 0.05). Similarto AcH4/K12 acetylation, re-acetyla-tion appeared quickly at 3 hr afterfusion (7 of 10) and the signal main-tained until 6 hr after activation (6 of9 and 8 of 8) (Fig. 2A and B, P � 0.05).The staining intensity of TSA-treatedembryos at 6 hr of activation was muchstronger than that in untreated groups(11 of 13) (Fig. 2A and B, P � 0.05).
Dynamic Changes of HistoneAcetylation inRabbit–Human InterspeciesSCNT Embryos From Fusionto 6 hr After Activation
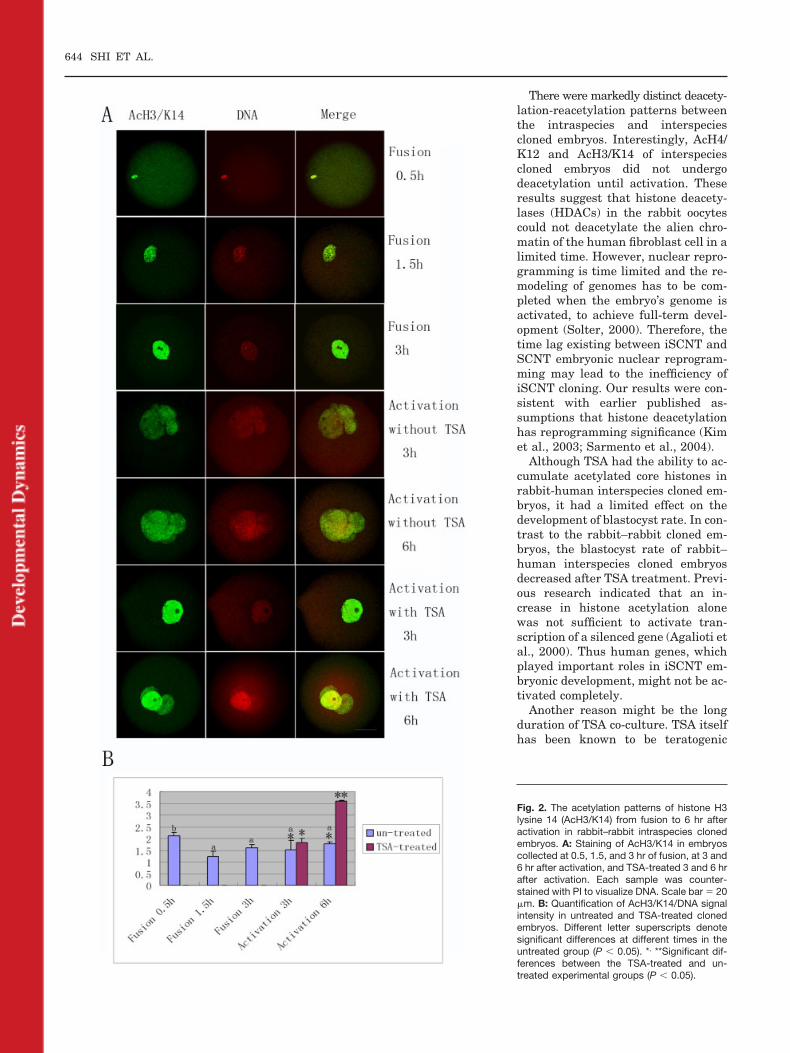
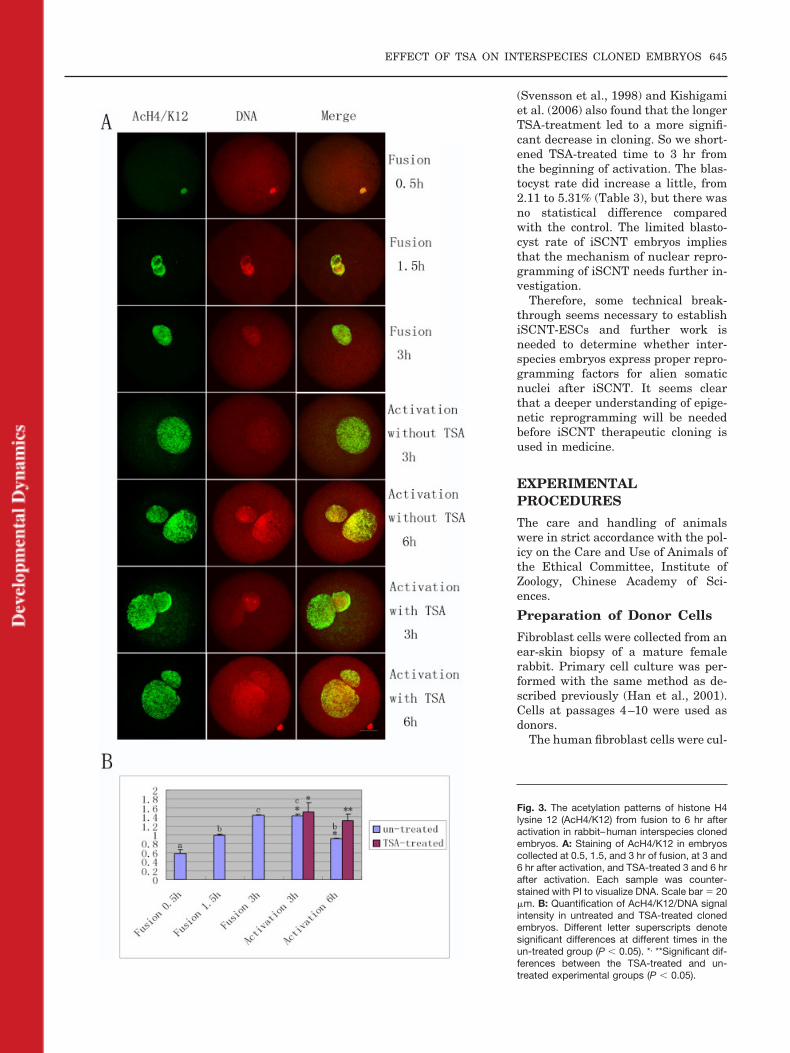
Staining patterns of AcH4/K12 andAcH3/K14 in rabbit–human interspe-cies cloned embryos exhibited dra-matic differences from those of rabbit–rabbit embryos. From 0.5 to 3 hr afterfusion, surprisingly, neither AcH4/
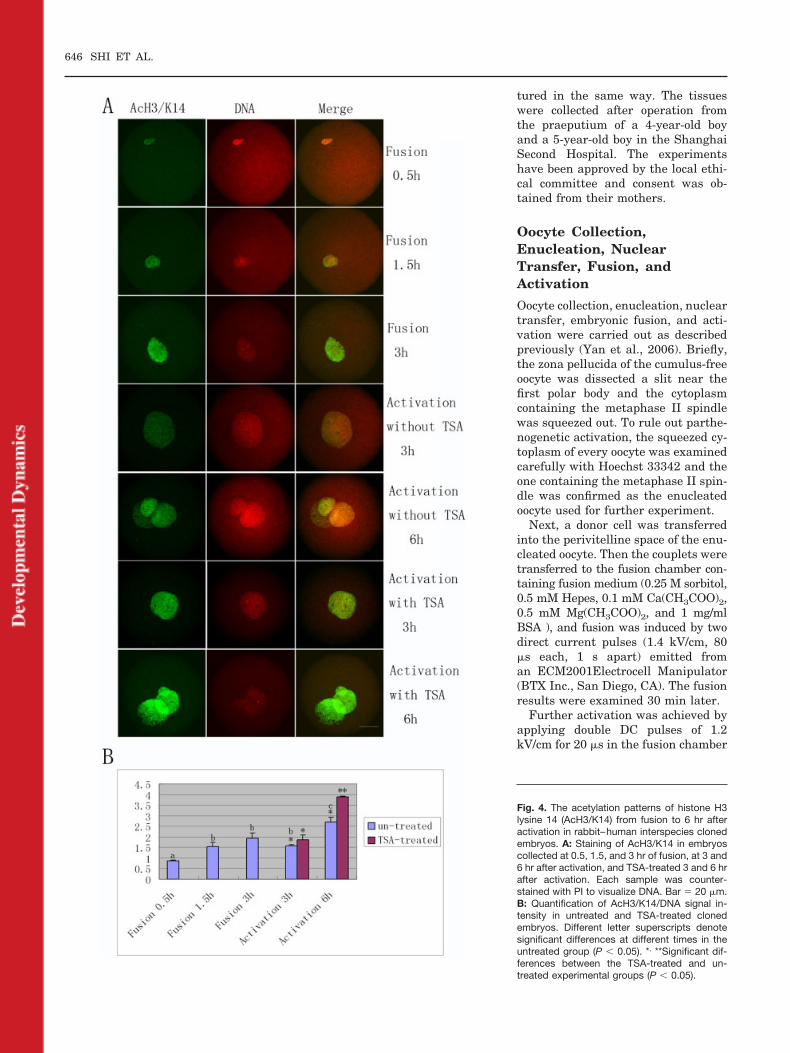
K12 nor AcH3/K14 was deacetylated,on the contrary, the signals kept in-creasing (6 of 8 and 6 of 9, at 1.5 hr)(Figs. 3A,B, 4A,B, P � 0.05). Interest-ingly, at 3 hr following activation,both of AcH4/K12 (5 of 9) and AcH3/K14 (6 of 9) signal intensities de-creased a little (Figs. 3A,B, 4A,B), butthere was no significant difference(P � 0.05). Unexpectedly, at 6 hr ofactivation, the intensity of AcH4/K12signal greatly decreased (8 of 10, P �0.05), whereas the signal intensity ofAcH3/K14 increased (7 of 8, P � 0.05)(Figs. 3A,B, 4A,B). In addition, thesignal intensity of TSA-treated groupsat 6 hr of activation was much higherthan that in untreated embryos (11 of13 and 9 of 10) (Figs. 3A,B, 4A,B, P �0.05)
DISCUSSION
Therapeutic cloning of iSCNT is apromising new technology with poten-tial applications in regenerative med-icine. However, little work has beendone on blastocyst development andnuclear reprogramming in iSCNT em-bryos. Previous studies demonstratedthat TSA could dramatically increasethe quantity and quality of blastocysts
in cloned embryos (Kishigami et al.,2006; Rybouchkin et al., 2006). Thepresent study investigated the effectof TSA on the development of SCNT(rabbit–rabbit) and iSCNT (rabbit–human) embryos and the importanceof reshaping the global histone acety-lation patterns for nuclear reprogram-ming.
As reported previously (Kim et al.,2003; Rybouchkin et al., 2006; Wanget al., 2007), deacetylation of clonedembryos occurred within 3 hr after fu-sion, and de novo acetylation tookplace with pronucleus formation. Inagreement with our results, Wang etal. (2007) also detected that deacety-lation patterns of AcH4/K12 andAcH3/K14 were displayed in two dif-ferent ways. AcH4/K12 persisted inthe genome of cloned embryos withonly mild deacetylation, while AcH3/K14 quickly deacetylated in the pro-cess of SCNT (Wang et al., 2007). Inthe process of nuclear reprogram-ming, deacetylation erased any infor-mation of the somatic cell memory orcreated a relative naive set of chroma-tin onto which the zygotic programwould be built; reacetylation was tore-establish embryonic epigenetic
TABLE 2. Development of Rabbit–Human Interspecies Cloned Embryos After Treatment With TSA for 6 hra
TreatmentNo. ofoocytes
No. ofreconstructedoocytes
No. ofembryosfused (%)
No. of 2-cellembryos (%)
No. of 4-cellembryos (%)
No. ofmorula(%)
No. ofblastocytes(%)
Non-treated 150 144 75(52.08)a
57(76)a
16(28.07)a
6(8)a
3(4)a
TSA6hr (100 nM) 189 176 92(52.27)a
60(65.22)a
14(23.33)a
2(2.17)a
2(2.17)a
aValues with different superscripts within the same column differ significantly (P � 0.05). Note: Fused (%), fusedoocytes/reconstructed oocytes; development rate of embryos at different stages, number of embryos/number of fused embryos.
TABLE 3. Development of Rabbit–Human Interspecies Cloned Embryos After Treatment With TSA for 3 hra
TreatmentNo. ofoocytes
No. ofreconstructedoocytes
No. ofembryosfused (%)
No. of 2-cellembryos (%)
No. of 4-cellembryos (%)
No. ofmorula(%)
No. ofblastocysts(%)
Non-treated 185 178 95(53.37)a
73(76.84)a
22(30.14)a
4(4.21)a
2(2.11)a
TSA3hr (100 nM) 223 212 113(53.30)a
91(80.53)a
26(28.57)a
8(7.08)a
6(5.31)a
aValues with different superscripts within the same column differ significantly (P � 0.05). Note: Fused (%), fusedoocytes/reconstructed oocytes; development rate of embryos at different stages, number of embryos/number of fused embryos.
642 SHI ET AL.
characteristics and gene expression inNT embryos. Our results add furtherevidence that histone deacetylation-reacetylation are involved in the nu-clear reprogramming of gene expres-sion after NT (Kim et al., 2003). TSAincreased the signal intensity of acet-ylation when the reconstructed em-bryos reacetylated 6 hr after activa-tion. These results might give moreproof to the conclusion that deacetyla-tion of histones at some specific chro-matin locations occurred at the time ofoocyte activation, in spite of the rise inthe global level of chromatin acet-ylation (Adenot et al., 1997;Wiekowskim et al., 1997; Spinaci etal., 2004; Rybouchkin et al., 2006).
Consistent with earlier studies(Kishigami et al., 2006; Rybouchkin etal., 2006), the morula rate and blasto-cyst rates of SCNT rabbit embryostreated with TSA during activationwere about twice as much as those ofuntreated embryos. TSA can inducethe hyperacetylation of core histonethat will result in the change in struc-ture of chromatin and make the geneaccessible for transcriptional factorsto active gene expression (Hong et al.,1993; Lee et al., 1993). In addition, theprevious data showed that TSA couldalso increase DNA demethylation(Cervoni and Szyf, 2001), and thus in-duce the activation of the key gene forembryo development. Rybouchkin etal. (2006) proposed that hyperacetyla-tion of core histone by treatment withTSA at an early stage of NT embryodevelopment stimulated more effec-tive remodeling of somatic chromatin,suppressing reprogramming effectvariegation, leading to effective repro-gramming. In a word, TSA couldgreatly increase blastocyst develop-ment of intraspecies cloned embryos.
Fig. 1. The acetylation patterns of histone H4lysine 12 (AcH4/K12) from fusion to 6 hr afteractivation in rabbit–rabbit intraspecies clonedembryos. A: Staining of AcH4/K12 in embryoscollected at 0.5, 1.5, and 3 hr of fusion, at 3 and6 hr after activation, and TSA-treated 3 and 6 hrafter activation. Each sample was counter-stained with PI to visualize DNA. Scale bar � 20�m. B: Quantification of AcH4/K12/DNA signalintensity in un-treated and TSA-treated clonedembryos. Different letters superscripts denotesignificant differences at different times in un-treated group (P � 0.05). *, **Significant differ-ences between TSA-treated and un-treated ex-perimental groups (P � 0.05).
EFFECT OF TSA ON INTERSPECIES CLONED EMBRYOS 643
There were markedly distinct deacety-lation-reacetylation patterns betweenthe intraspecies and interspeciescloned embryos. Interestingly, AcH4/K12 and AcH3/K14 of interspeciescloned embryos did not undergodeacetylation until activation. Theseresults suggest that histone deacety-lases (HDACs) in the rabbit oocytescould not deacetylate the alien chro-matin of the human fibroblast cell in alimited time. However, nuclear repro-gramming is time limited and the re-modeling of genomes has to be com-pleted when the embryo’s genome isactivated, to achieve full-term devel-opment (Solter, 2000). Therefore, thetime lag existing between iSCNT andSCNT embryonic nuclear reprogram-ming may lead to the inefficiency ofiSCNT cloning. Our results were con-sistent with earlier published as-sumptions that histone deacetylationhas reprogramming significance (Kimet al., 2003; Sarmento et al., 2004).
Although TSA had the ability to ac-cumulate acetylated core histones inrabbit-human interspecies cloned em-bryos, it had a limited effect on thedevelopment of blastocyst rate. In con-trast to the rabbit–rabbit cloned em-bryos, the blastocyst rate of rabbit–human interspecies cloned embryosdecreased after TSA treatment. Previ-ous research indicated that an in-crease in histone acetylation alonewas not sufficient to activate tran-scription of a silenced gene (Agalioti etal., 2000). Thus human genes, whichplayed important roles in iSCNT em-bryonic development, might not be ac-tivated completely.
Another reason might be the longduration of TSA co-culture. TSA itselfhas been known to be teratogenic
Fig. 2. The acetylation patterns of histone H3lysine 14 (AcH3/K14) from fusion to 6 hr afteractivation in rabbit–rabbit intraspecies clonedembryos. A: Staining of AcH3/K14 in embryoscollected at 0.5, 1.5, and 3 hr of fusion, at 3 and6 hr after activation, and TSA-treated 3 and 6 hrafter activation. Each sample was counter-stained with PI to visualize DNA. Scale bar � 20�m. B: Quantification of AcH3/K14/DNA signalintensity in untreated and TSA-treated clonedembryos. Different letter superscripts denotesignificant differences at different times in theuntreated group (P � 0.05). *, **Significant dif-ferences between the TSA-treated and un-treated experimental groups (P � 0.05).
644 SHI ET AL.
(Svensson et al., 1998) and Kishigamiet al. (2006) also found that the longerTSA-treatment led to a more signifi-cant decrease in cloning. So we short-ened TSA-treated time to 3 hr fromthe beginning of activation. The blas-tocyst rate did increase a little, from2.11 to 5.31% (Table 3), but there wasno statistical difference comparedwith the control. The limited blasto-cyst rate of iSCNT embryos impliesthat the mechanism of nuclear repro-gramming of iSCNT needs further in-vestigation.
Therefore, some technical break-through seems necessary to establishiSCNT-ESCs and further work isneeded to determine whether inter-species embryos express proper repro-gramming factors for alien somaticnuclei after iSCNT. It seems clearthat a deeper understanding of epige-netic reprogramming will be neededbefore iSCNT therapeutic cloning isused in medicine.
EXPERIMENTALPROCEDURES
The care and handling of animalswere in strict accordance with the pol-icy on the Care and Use of Animals ofthe Ethical Committee, Institute ofZoology, Chinese Academy of Sci-ences.
Preparation of Donor Cells
Fibroblast cells were collected from anear-skin biopsy of a mature femalerabbit. Primary cell culture was per-formed with the same method as de-scribed previously (Han et al., 2001).Cells at passages 4–10 were used asdonors.
The human fibroblast cells were cul-
Fig. 3. The acetylation patterns of histone H4lysine 12 (AcH4/K12) from fusion to 6 hr afteractivation in rabbit–human interspecies clonedembryos. A: Staining of AcH4/K12 in embryoscollected at 0.5, 1.5, and 3 hr of fusion, at 3 and6 hr after activation, and TSA-treated 3 and 6 hrafter activation. Each sample was counter-stained with PI to visualize DNA. Scale bar � 20�m. B: Quantification of AcH4/K12/DNA signalintensity in untreated and TSA-treated clonedembryos. Different letter superscripts denotesignificant differences at different times in theun-treated group (P � 0.05). *, **Significant dif-ferences between the TSA-treated and un-treated experimental groups (P � 0.05).
EFFECT OF TSA ON INTERSPECIES CLONED EMBRYOS 645
tured in the same way. The tissueswere collected after operation fromthe praeputium of a 4-year-old boyand a 5-year-old boy in the ShanghaiSecond Hospital. The experimentshave been approved by the local ethi-cal committee and consent was ob-tained from their mothers.
Oocyte Collection,Enucleation, NuclearTransfer, Fusion, andActivation
Oocyte collection, enucleation, nucleartransfer, embryonic fusion, and acti-vation were carried out as describedpreviously (Yan et al., 2006). Briefly,the zona pellucida of the cumulus-freeoocyte was dissected a slit near thefirst polar body and the cytoplasmcontaining the metaphase II spindlewas squeezed out. To rule out parthe-nogenetic activation, the squeezed cy-toplasm of every oocyte was examinedcarefully with Hoechst 33342 and theone containing the metaphase II spin-dle was confirmed as the enucleatedoocyte used for further experiment.
Next, a donor cell was transferredinto the perivitelline space of the enu-cleated oocyte. Then the couplets weretransferred to the fusion chamber con-taining fusion medium (0.25 M sorbitol,0.5 mM Hepes, 0.1 mM Ca(CH3COO)2,0.5 mM Mg(CH3COO)2, and 1 mg/mlBSA ), and fusion was induced by twodirect current pulses (1.4 kV/cm, 80�s each, 1 s apart) emitted froman ECM2001Electrocell Manipulator(BTX Inc., San Diego, CA). The fusionresults were examined 30 min later.
Further activation was achieved byapplying double DC pulses of 1.2kV/cm for 20 �s in the fusion chamber
Fig. 4. The acetylation patterns of histone H3lysine 14 (AcH3/K14) from fusion to 6 hr afteractivation in rabbit–human interspecies clonedembryos. A: Staining of AcH3/K14 in embryoscollected at 0.5, 1.5, and 3 hr of fusion, at 3 and6 hr after activation, and TSA-treated 3 and 6 hrafter activation. Each sample was counter-stained with PI to visualize DNA. Bar � 20 �m.B: Quantification of AcH3/K14/DNA signal in-tensity in untreated and TSA-treated clonedembryos. Different letter superscripts denotesignificant differences at different times in theuntreated group (P � 0.05). *, **Significant dif-ferences between the TSA-treated and un-treated experimental groups (P � 0.05).
646 SHI ET AL.

containing fusion medium 3 hr afterfusion. Then the activated embryoswere washed three times with M199supplemented with 10% FBS (culturemedium) and used for further experi-ment.
Reconstructed EmbryosCollection and TSA-Treatment
TSA solution prepared in dimethyl-sulfoxide (DMSO) was diluted in cul-ture medium to yield a final concen-tration of 100 nM, which was based ona previous study (Rybouchkin et al.,2006). When the embryos weretreated with TSA, correspondingly,0.1% DMSO was used as a control.
Five randomly selected cloned em-bryos were fixed at fusion for 0.5, 1.5,and 3 hr. During activation, the recon-structed embryos were cultured inM199 supplemented with 10% FBS(culture medium) with or without 100nM TSA for 6 hr. Five of the clonedembryos were collected after activa-tion of 3 and 6 hr including untreatedand TSA-treated embryos, respec-tively.
Immunofluorescent ConfocalMicroscopy
After removing the zona pellucida inacidic M2 medium (pH 2.5), embryoswere fixed with 4% paraformaldehydein PBS (pH 7.4) for 1 hr and perme-abilized with 1% Triton X-100 over-night at 4°C. Then the embryos wereblocked in 1% BSA-supplementedPBS for 1 hr at room temperature andincubated overnight at 4°C with rab-bit polyclonal anti-acetyl lysine 12 inhistone H4 (AcH4/K12) antibody andanti-acetyl lysine14 in histone H3(AcH3/K14) antibody, which were pur-chased from Upstate Biotechnology(Lake Placid, NY), diluted 1:300 inblocking solution. After three washesin PBS containing 0.1% Tween-20 and0.01% Triton X-100 (washing solution)for 5 min each, the embryos were la-beled with secondary FITC-conju-gated antibody diluted 1:100 for 1 hrat room temperature. After extensivewashing, DNA of samples was coun-terstained with propidium iodide (PI).Finally, samples were mounted be-tween a cover slip and glass slide sup-ported by four columns of a mixture of
petroleum jelly and paraffin (9:1).Each experiment was repeated atleast three times, and 5 randomly se-lected reconstructed embryos were ex-amined each time. Slides werescanned using a confocal laser-scan-ning microscope (Zeiss LSM 510METE, Germany) with an argon/kryp-ton laser at 488 and 563 nm and two-channel scanning for detection offluorescein isothiocyanate and PI, re-spectively. Exposure times from fluo-rescent light were kept constant forthe respective channel. Intensity ofimages was determined using an im-age analyzer system; the intensity ofacetylated histone H4-lysine 12(AcH4/K12) or acetylated histone H3-lysine 14 (AcH3/K14) was compared tothat of PI signal (Wee et al., 2006).The fluorescent intensity was ana-lyzed by a Paired-Samples t-test withSPSS software and P � 0.05 was con-sidered to be significant. The imageswere merged using Adobe Photoshop7.0 software.
Statistical Analysis
The data were pooled from at leastthree replicates and analyzed by aChi-square test. Differences at P �0.05 were considered to be significant.
ACKNOWLEDGMENTSThe authors thank Hui-Zhen Sheng inthe Second Medicine Hospital ofShanghai for providing the human fi-broblast cell. We also thank Shi-WenLi for sample scanning, and QiangWang, Shen Yin, Jun-Shu Ai, Li-YingYan, and Man-Xi Jiang for their tech-nical help and comments on themanuscript.
REFERENCES
Adcock IM, Lee KY. 2006. Abnormal his-tone acetylase and deacetylase expres-sion and function in lung inflammation.Inflamm Res 55:311–321.
Adenot PG, Mercier Y, Renard JP, Thomp-son EM. 1997. Differential H4 acetyla-tion of paternal and maternal chromatinprecedes DNA replication and differen-tial transcriptional activity in pronucleiof 1-cell mouse embryos. Development124:4615–4625.
Agalioti T, Lomvardas S, Parekh B, Yie J,Maniatis T, Thanos D. 2000. Ordered re-cruitment of chromatin modifying andgeneral transcription factors to the IFN-beta promoter. Cell 103:667–678.
Cervoni N, Szyf M. 2001. Demethylase ac-tivity is directed by histone acetylation.J Biol Chem 276:40778–40787.
Chen DY, Wen DC, Zhang YP, Sun QY,Han ZM, Liu ZH, Shi P, Li JS, XiangyuJG, Lian L, Kou ZH, Wu YQ, Chen YC,Wang PY, Zhang HM. 2002. Interspeciesimplantation and mitochondria fate ofpanda-rabbit cloned embryos. Biol Re-prod 67:637–642.
Chen Y, He ZX, Liu A, Wang K, Mao WW,Chu JX, Lu Y, Fang ZF, Shi YT, YangQZ, Chen da Y, Wang MK, Li JS, HuangSL, Kong XY, Shi YZ, Wang ZQ, Xia JH,Long ZG, Xue ZG, Ding WX, Sheng HZ.2003. Embryonic stem cells generated bynuclear transfer of human somatic nu-clei into rabbit oocytes. Cell Res 13:251–263.
Corley-Smith GE, Brandhorst BP. 1999.Preservation of endangered species andpopulations: a role for genome banking,somatic cell cloning, and androgenesis?Mol Reprod Dev 53:363–367.
Dean W, Santos F, Stojkovic M, Zakhartch-enko V, Walter J, Wolf E, Reik W. 2001.Conservation of methylation reprogram-ming in mammalian development: aber-rant reprogramming in cloned embryos.Proc Natl Acad Sci USA 98:13734–13738.
Dominko T, Mitalipova M, Haley B, Bey-han Z, Memili E, McKusick B, First NL.1999. Bovine oocyte cytoplasm supportsdevelopment of embryos produced by nu-clear transfer of somatic cell nuclei fromvarious mammalian species. Biol Reprod60:1496–1502.
Enright BP, Kubota C, Yang X, Tian XC.2003. Epigenetic characteristics and de-velopment of embryos cloned from donorcells treated by trichostatin A or 5-aza-2’-deoxycytidine. Biol Reprod 69:896–901.
Fischle W, Wang Y, Allis CD. 2003. His-tone and chromatin cross-talk. CurrOpin Cell Biol 15:172–183.
Gao L, Cueto MA, Asselbergs F, Atadja P.2002. Cloning and functional character-ization of HDAC11, a novel member ofthe human histone deacetylase family.J Biol Chem 277:25748–25755.
Gomez MC, Pope CE, Giraldo A, Lyons LA,Harris RF, King AL, Cole A, Godke RA,Dresser BL. 2004. Birth of African Wild-cat cloned kittens born from domesticcats. Cloning Stem Cells 6:247–258.
Han ZM, Chen DY, Li JS, Sun QY, WangPY, Huang Y, Du J. 2001. The culture offibroblasts from diaphragm of giantpanda. In Vitro Cell Dev Biol Anim 37:644–645.
Hong L, Schroth GP, Matthews HR, Yau P,Bradbury EM. 1993. Studies of the DNAbinding properties of histone H4 aminoterminus. Thermal denaturation studiesreveal that acetylation markedly reducesthe binding constant of the H4 “tail” toDNA. J Biol Chem 268:305–314.
Kang YK, Koo DB, Park JS, Choi YH,Chung AS, Lee KK, Han YM. 2001. Ab-errant methylation of donor genome incloned bovine embryos. Nat Genet 28:173–177.
EFFECT OF TSA ON INTERSPECIES CLONED EMBRYOS 647

Kim JM, Liu H, Tazaki M, Nagata M, AokiF. 2003. Changes in histone acetylationduring mouse oocyte meiosis. J Cell Biol162:37–46.
Kishigami S, Mizutani E, Ohta H, HikichiT, Thuan NV, Wakayama S, Bui HT,Wakayama T. 2006. Significant improve-ment of mouse cloning technique bytreatment with trichostatin A after so-matic nuclear transfer. Biochem BiophysRes Commun 340:183–189.
Kitiyanant Y, Saikhun J, Chaisalee B,White KL, Pavasuthipaisit K. 2001. So-matic cell cloning in Buffalo (Bubalusbubalis): effects of interspecies cytoplas-mic recipients and activation proce-dures. Cloning Stem Cells 3:97–104.
Lanza RP, Cibelli JB, Diaz F, Moraes CT,Farin PW, Farin CE, Hammer CJ, WestMD, Damiani P. 2000. Cloning of an en-dangered species (Bos gaurus) using in-terspecies nuclear transfer. Cloning 2:79–90.
Lee DY, Hayes JJ, Pruss D, Wolffe AP.1993. A positive role for histone acetyla-tion in transcription factor access to nu-cleosomal DNA. Cell 72:73–84.
Loi P, Ptak G, Barboni B, Fulka J Jr, Cap-pai P, Clinton M. 2001. Genetic rescue ofan endangered mammal by cross-speciesnuclear transfer using post-mortem so-matic cells. Nat Biotechnol 19:962–964.
Ohgane J, Wakayama T, Senda S,Yamazaki Y, Inoue K, Ogura A, Marh J,Tanaka S, Yanagimachi R, Shiota K.2004. The Sall3 locus is an epigenetichotspot of aberrant DNA methylation as-sociated with placentomegaly of clonedmice. Genes Cells 9:253–260.
Rybouchkin A, Kato Y, Tsunoda Y. 2006.Role of histone acetylation in reprogram-ming of somatic nuclei following nucleartransfer. Biol Reprod 74:1083–1089.
Saikhun J, Pavasuthipaisit K, Jaruansu-wan M, Kitiyanant Y. 2002. Xenonuclear
transplantation of buffalo (Bubalusbubalis) fetal and adult somatic cell nu-clei into bovine (Bos indicus) oocyte cy-toplasm and their subsequent develop-ment. Theriogenology 57:1829–1837.
Santos F, Zakhartchenko V, Stojkovic M,Peters A, Jenuwein T, Wolf E, Reik W,Dean W. 2003. Epigenetic marking cor-relates with developmental potential incloned bovine preimplantation embryos.Curr Biol 13:1116–1121.
Sarmento OF, Digilio LC, Wang Y, PerlinJ, Herr JC, Allis CD, Coonrod SA. 2004.Dynamic alterations of specific histonemodifications during early murine devel-opment. J Cell Sci 117:4449–4459.
Solter D. 2000. Mammalian cloning: ad-vances and limitations. Nat Rev Genet1:199–207.
Spinaci M, Seren E, Mattioli M. 2004. Ma-ternal chromatin remodeling duringmaturation and after fertilization inmouse oocytes. Mol Reprod Dev 69:215–221.
Svensson K, Mattsson R, James TC, Went-zel P, Pilartz M, MacLaughlin J, MillerSJ, Olsson T, Eriksson UJ, Ohlsson R.1998. The paternal allele of the H19 geneis progressively silenced during earlymouse development: the acetylation sta-tus of histones may be involved in thegeneration of variegated expression pat-terns. Development 125:61–69.
Tecirlioglu RT, Guo J, Trounson AO. 2006.Interspecies somatic cell nuclear trans-fer and preliminary data for horse-cow/mouse iSCNT. Stem Cell Rev 2:277–287.
Turner BM. 2000. Histone acetylation andan epigenetic code. Bioessays 22:836–845.
Wakayama S, Kishigami S, Van Thuan N,Ohta H, Hikichi T, Mizutani E, Yanagi-machi R, Wakayama T. 2005a. Propaga-tion of an infertile hermaphrodite mouselacking germ cells by using nuclear
transfer and embryonic stem cell tech-nology. Proc Natl Acad Sci USA 102:29–33.
Wakayama S, Mizutani E, Kishigami S,Thuan NV, Ohta H, Hikichi T, Bui HT,Miyake M, Wakayama T. 2005b. Micecloned by nuclear transfer from somaticand ntES cells derived from the sameindividuals. J Reprod Dev 51:765–772.
Wakayama T, Tabar V, Rodriguez I, PerryAC, Studer L, Mombaerts P. 2001. Dif-ferentiation of embryonic stem cell linesgenerated from adult somatic cells bynuclear transfer. Science 292:740–743.
Wang F, Kou Z, Zhang Y, Gao S. 2007.Dynamic reprogramming of histone acet-ylation and methylation in the first cellcycle of cloned mouse embryos. Biol Re-prod 77:1007–1016.
Wee G, Koo DB, Song BS, Kim JS, KangMJ, Moon SJ, Kang YK, Lee KK, HanYM. 2006. Inheritable histone H4 acety-lation of somatic chromatins in clonedembryos. J Biol Chem 281:6048–6057.
Wiekowskim., Miranda M, Nothias JY, De-pamphils ML. 1997. Change in histonesynthesis and modification at the begin-ning of mouse development correlatewith the establishment of chromatin me-diated repression of transcription. J CellSci 110:1147–1158
Yan LY, Huang JC, Zhu ZY, Lei ZL, ShiLH, Nan CL, Zhao ZJ, Ouyang YC, SongXF, Sun QY, Chen DY. 2006. NuMA dis-tribution and microtubule configurationin rabbit oocytes and cloned embryos.Reproduction 132:869–876.
Yang F, Hao R, Kessler B, Brem G, Wolf E,Zakhartchenko V. 2007. Rabbit somaticcell cloning: effects of donor cell type,histone acetylation status and chimericembryo complementation. Reproduction133:219–230.
648 SHI ET AL.


















