
Transportation & Structure of Plants
47
TRANSPORT IN ANIMALS AND PLANTS
-
Upload
thivyaapriya -
Category
Science
-
view
293 -
download
2
Transcript of Transportation & Structure of Plants

TRANSPORT IN
ANIMALS AND PLANTS

THE PRESSURE–FLOW HYPOTHESIS
EXPLAINS
TRANSLOCATION IN PHLOEM
• Current experimental evidence supports the
translocation of dissolved sugar in phloem by the
pressure–flow hypothesis, which was first proposed
in 1926 by the German scientist Ernst Münch.

• The pressure–flow hypothesis states that solutes
(such as dissolved sugars) move in phloem by
means of a pressure gradient—that is, a difference
in pressure
• The pressure gradient exists between the source,
where the sugar is loaded into phloem, and the sink,
where the sugar is removed from phloem.
• According to the pressure flow hypothesis,
translocation which takes place in three stages
which involves a combination of active transport and
mass flow.

a) First stage occur at the leaves (source)
• Occurrence at leaves where photosynthesis is
carried out by mesophyll cells to produce organic
substances.
• Then, glucose is produced during photosynthesis
and condenses into sucrose which is a more
suitable soluble substance for transport.
• The leave ,which sucrose and other organic solutes
are actively loaded into sieve tubes.
• The loading is carried out by modified companion
cells called transfer cells.

• These modified companion cells contains numerous
mitochondria to provide Adenosine Triphosphate
(ATP) energy for the active loading and numerous
ingrowths (internal projections) of their cell wall to
increase surface area for more efficient loading.
• These transfer cells are used in active mechanism
to load into sieve tubes by cotransport
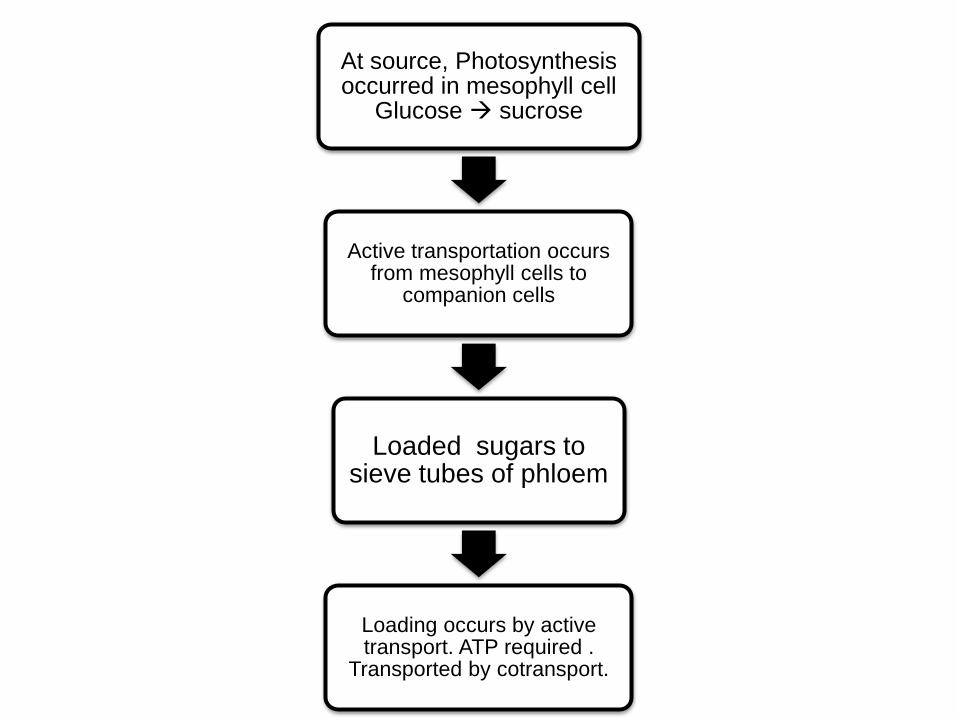
At source, Photosynthesis occurred in mesophyll cell
Glucose sucrose
Active transportation occurs from mesophyll cells to
companion cells
Loaded sugars to sieve tubes of phloem
Loading occurs by active transport. ATP required .
Transported by cotransport.
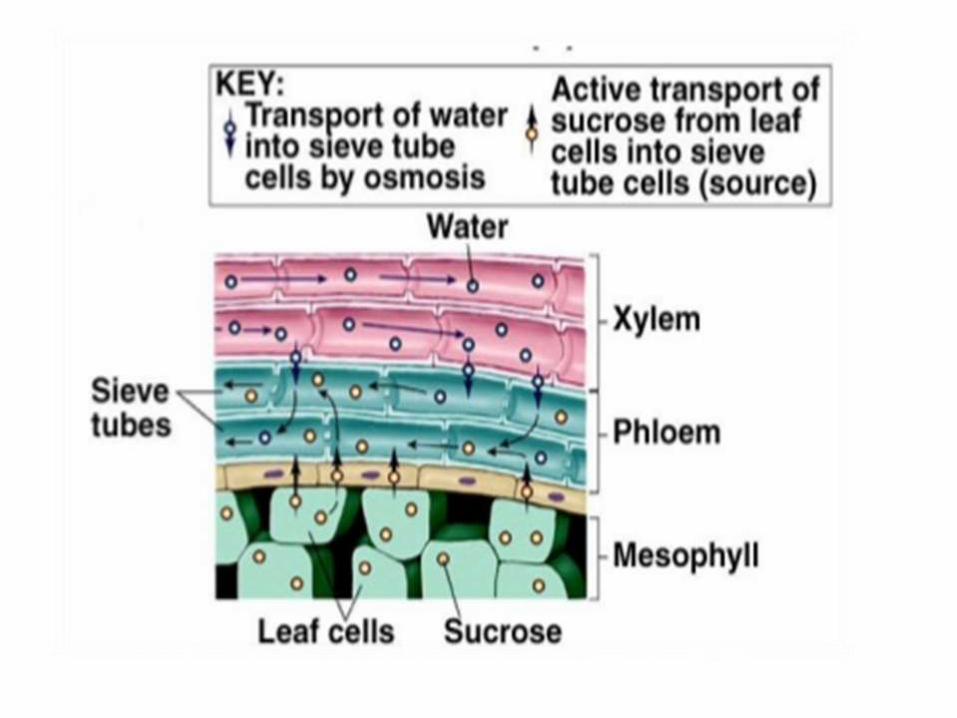

The Route of sugar
transport from
mesophyll cells to
sieve tube cells.
• At companion cells ,
sugars in cell walls are
actively loaded into
cytoplasm as known
Symplast route .
• The sugar then moves
into the adjoining sieve
tubes elements through
plasmodesmata.
• In some species, sugar
transport involves a
combination of symplast
and apoplast routes and
needs ATP energy to
actively load sugar into
sieve tube members.

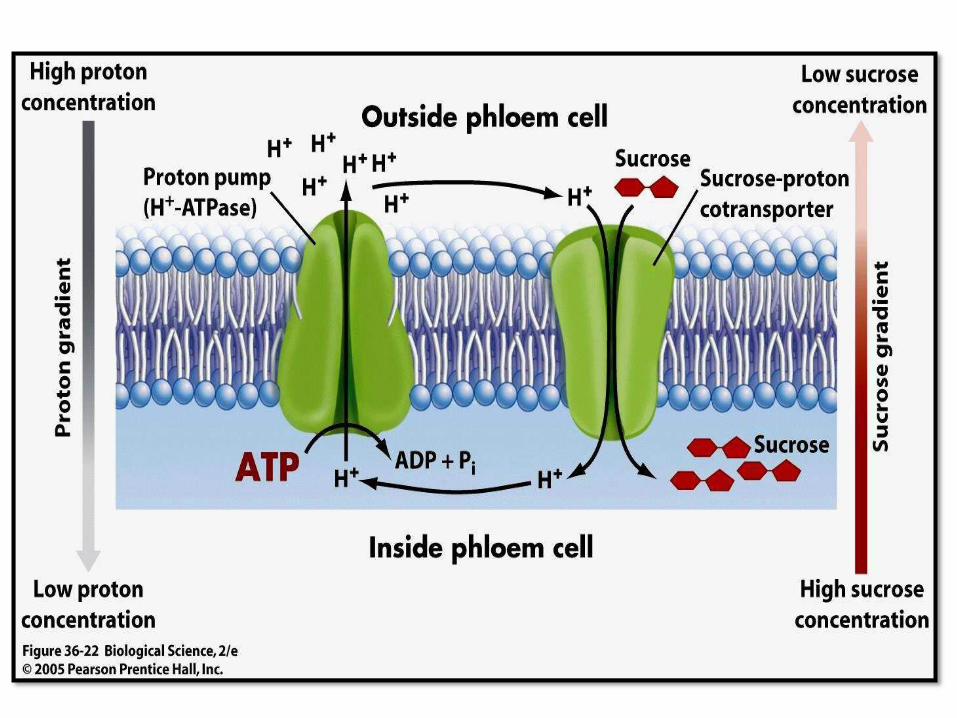
SUGAR LOADING
• Sugar loading involves a chemiosmostic
mechanism that uses a cotransport.
• Chemiosmostic mechanism is involved in the
transport of sucrose from transfer cells into sieve
tube.
• Proton pumps hydrogen ions (H+) out of the cell
membrane resulting in a proton gradient across the
membrane.
• A cotransport protein then carries the hydrogen ions
(H+) down its concentration gradient back into the
cell and sugar is transported into the cell as well.


• The ATP supplies energy to pump protons out of the
sieve tube elements, producing a proton gradient that
drives the uptake of sugar through specific channels by
the cotransport of protons back into the sieve tube
elements.
• The sugar therefore accumulates in the sieve tube
element.
• The increase in dissolved sugars in the sieve tube
element at the source-a concentration that is 2 to 3
times as great as in surrounding cells-decreases
(makes more negative) the water potential of that cell.
• As a result, water moves by osmosis from the xylem
cells into the sieve tubes, increasing the turgor pressure
(hydrostatic pressure) inside them.
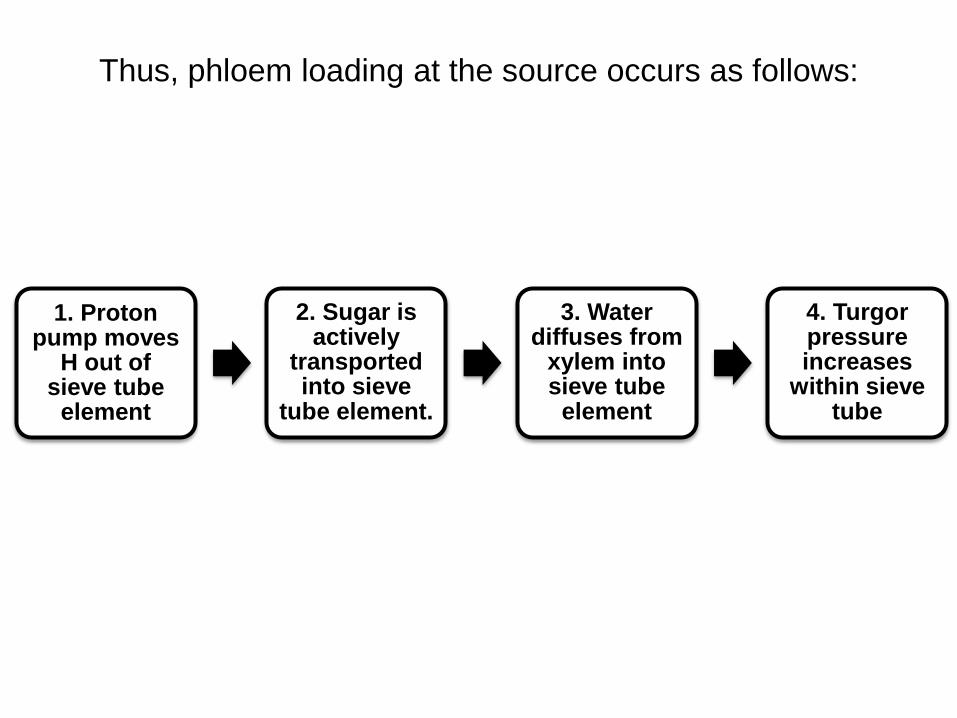
Thus, phloem loading at the source occurs as follows:
1. Proton pump moves
H out of sieve tube
element
2. Sugar is actively
transported into sieve
tube element.
3. Water diffuses from
xylem into sieve tube
element
4. Turgor pressure increases
within sieve tube

• At its destination (the sink), sugar is unloaded by various
mechanisms, both active and passive, from the sieve
tube elements.
• With the loss of sugar, the water potential in the sieve
tube elements at the sink increases (becomes less
negative).
• Therefore, water moves out of the sieve tubes by
osmosis and into surrounding cells where the water
potential is more negative.
• Most of this water diffuses back to the xylem to be
transported upward.
• This water movement decreases the turgor pressure
inside the sieve tubes at the sink.

• Thus, phloem unloading at the sink proceeds as follows:
Sugar is transported out
of sieve tube element
Water diffuses out of sieve tube element and into
xylem
Turgor pressure decreases
within sieve tube

The Pressure Flow-
Hypothesis
• Sugar is actively loaded into
the sieve tube element at the
source.
• As a result, water diffuses from
the xylem into the sieve tube
element.
• At the sink, the sugar is
actively or passively unloaded,
and water diffuses from the
sieve tube element into the
xylem.
• The pressure gradient within
the sieve tube, from source to
sink, causes translocation from
the area of higher turgor
pressure (the source) to the
area of lower turgor pressure
(the sink).
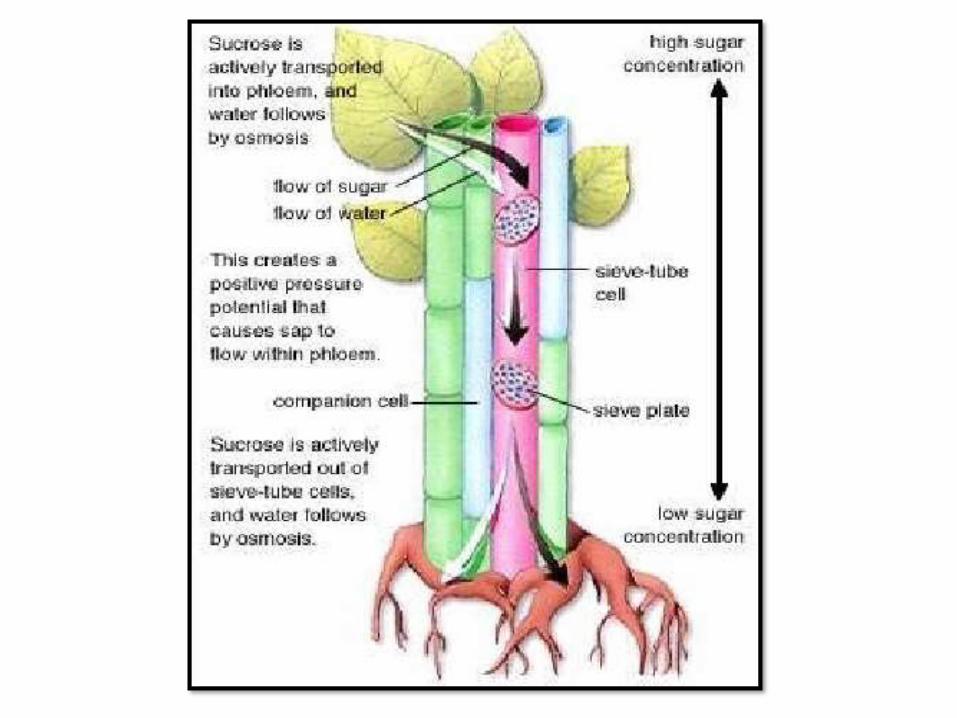


Sucrose
1.When high concentration of
sugar at sieve tubes
2.Water potential decreases
3.Water diffuse in from xylem to
sieve tube
4.IncreaseTurgor Pressure in Sieve
tube.
1.When low concentration of
sugar in sieve tubes.
2.Water potential increases
3.Water diffuse out of the sieve tube
to xylem
4.DecreaseTurgor pressure in sieve
tube.

b) Second stage (Translocated in the stem
from source to the sink by Mass Flow)
• Sink- is a area where organic substances translocate
from the source are used or stored.E.g.: stem tubes,
tap roots, fruits and seeds.
• The concentration of sucrose in the sink is lower than
that found in the source and sieve tubes as sugar is
continuously being consumed or converted into
starch to be stored .
• This allows sucrose to diffuse out of sieve tubes
down the concentration gradient and its continuous
flow from the source to the sink.
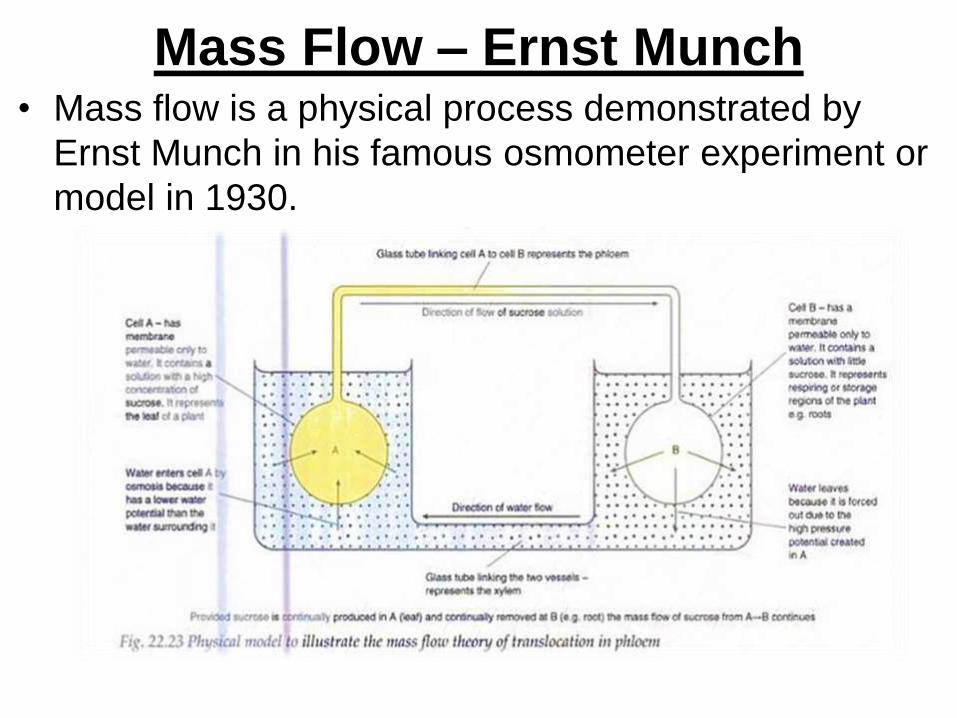
Mass Flow – Ernst Munch• Mass flow is a physical process demonstrated by
Ernst Munch in his famous osmometer experiment or
model in 1930.

• In mass flow, Munch’s model demonstrates that fluid flows
from region of high hydrostatic pressure to region of low
hydrostatic pressure.
• As fluid flow, it carried the whole mass of different substance.
• In osmometer A, concentrated sucrose solution (leaf) has
lower water potential. Water flows into it from a high water
potential region (xylem vessel) to a low water potential region
(leaf cells) by osmosis.
• This create high hydrostatic pressure in A and forces sucrose
solution to enter into the connecting tube (sieve tube) and
pass to B (root cell)
• As the flow of mass from osmometer A to osmometer B
continues, the sucrose solution is pushed along and finally
appears in B.

• In B, contain water / dilute sugar solution, water moves out
from a higher water potential region by the hydrostatic
pressure gradient produced and redistributed through
connecting tube (xylem vessels) between the two container.
• Mass flow continues until the concentration of sugar solution in
A and B are equal (balanced).
• In nature, equilibrium is not reached because solutes are
constantly synthesized at source A and utilized at the sink B.





C) The third stage of Pressure Flow.
• The sucrose and other organic substances are
actively unloaded at the sink (root cells) involving
companion cells and energy.
• Here, sucrose converted into insoluble starch, used
for cellular respiration and synthesis the cellulose of
cell wall.
• Now, the water potential in the cell sap of the root
cells is reduced. Water follows the organic solutes
from the sieve tube into the root cells by osmosis.
• This reduces the hydrostatic pressure at the sink.

• There exists a hydrostatic pressure gradient in the
sieve tube which causes the passive mass flow of
water and dissolved solutes from source to the sink
region due to the differences in water potential
between the leaves and roots.
• If sugar continues to be produces in the source and
converted to starch or to be oxidized at the sink , the
gradient will be maintained and the mass flow
continues.
• The return of excess water from the sink (root) to the
leaves (source) through xylem vessels is brought by
transpiration pull.

Supporting The Pressure Flow
Hypothesis
There are different pieces of evidences that support the
hypothesis.
• Firstly, there is an exudation of solution from the phloem
when the stem is cut or punctured by the mouthparts of an
aphid - A classical experiment demonstrating the
translocation function of phloem, indicating that the phloem
sap is under pressure
Phloem translocation is difficult to study in plants. Because
phloem cells are under pressure, cutting into phloem to
observe it releases the pressure and causes the contents of the
sieve tube elements (the phloem sap) to exude and mix with
the contents of other severed cells that are also unavoidably cut.

• In the 1950s, scientists developed a unique
research tool to avoid contaminating the phloem
sap: aphids, which are small insects that insert
their mouthparts into phloem sieve tubes for
feeding
• The pressure in the punctured phloem drives the
sugar solution through the aphid’s mouthpart into
its digestive system.
• When the aphid’s mouthpart is severed from its
body by a laser beam, the sugar solution
continues to flow through the mouthpart at a rate
proportional to the pressure in phloem.
• This rate can be measured, and the effects on
phloem transport.


• Secondly, concentration gradients of organic solutes are
proved to be present between the sink and the source.
• Thirdly, when viruses or growth chemicals are applied to a
well-illuminated (actively photosynthesizing) leaf, they are
translocated downwards to the roots. Yet, when applied to
shaded leaves, such downward translocation of chemicals
does not occur, hence showing that diffusion is not a possible
process involved in translocation.

Against The Pressure Flow Hypothesis
• Some argue that mass flow is a passive process while sieve
tube vessels are supported by companion cells. Hence, the
hypothesis neglects the living nature of phloem.
• It is difficult to make measurements of transporting in the
phloem due to disruptions caused to the phloem.
• In this hypothesis, substances cannot flow opposite
directions in the same sieve tubes.
• In actual fact, phloem contains many sieve tubes and
different solutes could travel in opposite directions at the
same time in different sieve tube with different sources and
sinks.

The Electro-osmosis Hypothesis
• This mechanism is proposed by Spanner
• Electroosmosis is the movement of ions in an electrical field
through a fixed porous which is electrically charged by
carrying water and any dissolved solutes.

• The sieve plates and phloem protein are normally negatively
charged, thus forming a fixed porous surface with an electrical
charge
• As mass flow occur through the negatively charged sieve
plates, anions will be repelled but cations will be able to pass
through.
• When mass flow occur downwards through the phloem, the
repulsed anions will accumulates above the sieve plates so
that the cell above the sieve plate will become negative.
• The sieve plate will now be a fixed porous surface within a
electrical field, such as is needed for electroosmosis to occur.
• When a critical potential difference across the sieve plate is
reached , protons (H+ ions) surge from the wall of upper cell
into its cytoplasm by lowering its pH and making the
cytoplasm above the sieve tube to positively charged.

• The increased positive charged generated by the H+ surge
pushes other cations mainly ( K+) by electrical repulsion,
through the sieve plate from the upper to the lower cell and
therefore, electroosmosis occurs.
• The Potassium ions are then secreted on the other side of the
sieve plates.
• The presence of Positive K+ ions on the other side of the sieve
plate induces negative charges on the other side of the sieve
creating a potential different across the sieve plates.
• This surge the hydrated Potassium ions carries water
molecules and dissolved solutes like sucrose across the sieve
plates.
• With this energy causes an electro-osmosis flow of polar water
molecules and dissolved solutes through the sieve pores to
the adjacent sieve tube element.
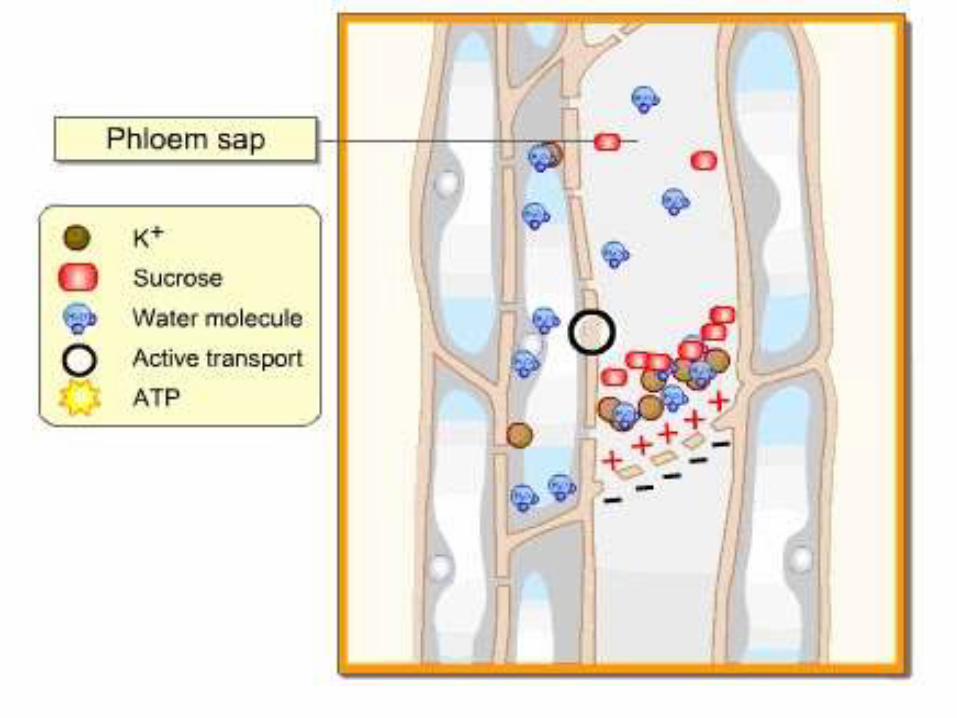

Supporting Electro-Osmosis
• The energy derived from potential difference
shows that translocation is an active
transport.
• The presence of porous sieve plates in all
sieve tubes of plants.
• High concentration of K+ ions have been
found in phloem sap.

Against Electro-Osmosis
• There is a lack of evidence and no
detailed mechanism of how substance or
solutes move between the sieve plates.

The Cytoplasmic Streaming
Hypothesis
• Was proposed by Thaine in 1962.
• The cytoplasm of plant cells is often observed to move
around within the cell, a process called Streaming
• It has been proposed that solutes might be carried from one
end of a sieve tube element to the other by streaming and
then transferred across sieve plates by active transport.
• Both streaming and transfer through the sieve plates would
be energy-dependent, explaining high turnover of ATP in
phloem cells
• Different solutes move through the sieve pore at different
rates due to their different molecular characteristics and level
of impermeability of sieve plates to different solutes.



Supporting C.S.H
• Upwards and downwards movement of
solutes that occur within the confines of the
sieve tube explains the two way flow
substance.
• Low temperature and metabolic poison affect
cytoplasmic streaming indicating an active
process is involved.

Against C.S.H
• Cytoplasmic streaming is not enough to
account for the rate of translocation
observed in phloem.
• Cytoplasmic streaming has been observed
in immature sieve tube only.

Peristalsis Waves• Transcellular strands contain contractile protein which are
present in some phloem sieve tubes.
• sieve tube is filled with fine cytoplasmic filaments
• continuous from sieve tube to the next
• thru pores of sieve plate.
• contain phloem sap tube constrict + relax alternately
• pushing sap from one sieve tube to the next.
• constriction + relaxation/peristaltic movement form a pattern of wave = peristaltic wave
• can be at diff speed + in opposite direction (in sieve tube)
• depends on metabolic energy/ATP.• Their rhythmic contraction produces peristalsis waves that
facilitate the long-distance transport of solutes in phloem.



















