
TRANSCRIPTIONAL REGULATION AND PROTECTIVE EFFICACY OF
192
TRANSCRIPTIONAL REGULATION AND PROTECTIVE EFFICACY OF BORDETELLA COLONIZATION FACTOR A (BcfA) IN BORDETELLA INFECTIONS BY NEELIMA SUKUMAR A Dissertation Submitted to the Graduate Faculty of WAKE FOREST UNIVESITY GRADUATE SCHOOL OF ARTS AND SCIENCES in Partial Fulfillment of the Requirements for the Degree of DOCTOR OF PHILOSOPHY Microbiology and Immunology May 2009 Winston-Salem, North Carolina Approved By: Rajendar Deora, Ph.D., Advisor ....................................................... Examining Committee: Mark O. Lively, Ph.D., Chairman ....................................................... Griffith Parks, Ph.D. ....................................................... Purnima Dubey, Ph.D. ....................................................... Sean Reid, Ph.D. .......................................................
Transcript of TRANSCRIPTIONAL REGULATION AND PROTECTIVE EFFICACY OF

TRANSCRIPTIONAL REGULATION AND PROTECTIVE EFFICACY OF
BORDETELLA COLONIZATION FACTOR A (BcfA) IN BORDETELLA
INFECTIONS
BY
NEELIMA SUKUMAR
A Dissertation Submitted to the Graduate Faculty of WAKE FOREST UNIVESITY GRADUATE SCHOOL OF ARTS AND SCIENCES
in Partial Fulfillment of the Requirements
for the Degree of
DOCTOR OF PHILOSOPHY
Microbiology and Immunology
May 2009
Winston-Salem, North Carolina
Approved By: Rajendar Deora, Ph.D., Advisor ....................................................... Examining Committee: Mark O. Lively, Ph.D., Chairman ....................................................... Griffith Parks, Ph.D. ....................................................... Purnima Dubey, Ph.D. ....................................................... Sean Reid, Ph.D. .......................................................

ACKNOWLEDGEMENTS
Over the past four years, I have realized that the adage “If you love
what you do, then you don’t have to work a single day in your life” is absolutely
true. I enjoyed every single moment of my graduate school. And this is not only
because I loved what I did, but also due to the great people I could share this
part of my life’s journey with.
First of all, I would like to express my sincere gratitude to my
mentor Dr. Rajendar Deora for the painstaking guidance, unconditional support
and advices which have helped me through out my research. Above all I am
thankful for the confidence he had in me. It is an honor to have worked with him.
I express my sincere acknowledgements to my committee members; Drs.
Griffith Parks, Dan Wozniak, Mark Lively, Sean Reid and Purnima Dubey.
Despite their busy schedules, they always extended whole hearted help and
provided insightful suggestions.
None of my research projects would have been complete without the help
of my Lab mates- both past and current; Dr. Meenu Mishra, Dr. Gina Parise
Sloan, Cheraton Love and Matt Conover. They are great friends and I enjoyed
working with them and will cherish all the moments I spent with them both inside
and outside of the lab.
Without the love and blessings of my parents; R. Sukumarakurup & Dr.
Sujatha and Mohanan Nair & Padmini, I wouldn’t even be in a graduate school.
They exemplify hard work, patience and trust; features that aid one in graduate
school. Also I would like to thank my brother Dr. Krishna Kumar, sister-in-law Dr.
ii

Jyotsana Menon and my twin sister Poornima Sukumar whose unfailing
encouragement and motivation have given me the strength to face the trials and
tribulations of the graduate school.
Although, only the last two years of my graduate school was shared with
my soul mate, my husband, Dr. Sunish Mohanan, he taught me some of the
greatest lessons- to believe in myself and always aim for the best. Since life
presented me with the best partner, I was positive that science would also gift
me. I would like to thank him for his patience and motivation.
Above all, I thank God for making me take the right decisions and blessing
me with a wonderful graduate school experience.
iii

TABLE OF CONTENTS
LIST OF FIGURES ..............................................................................................vi
LIST OF TABLES..............................................................................................viii
LIST OF ABBREVIATIONS ................................................................................ ix
ABSTRACT ........................................................................................................xii
CHAPTER I: INTRODUCTION.............................................................................1
The Bordetella Genus. ......................................................................................1
Bordetella as a Pathogen. .................................................................................2
The BvgAS Signal Transduction System...........................................................7
Virulence Factors. ...........................................................................................12
Adhesins. .....................................................................................................12
Toxins. .........................................................................................................16
Animal Models.................................................................................................19
Immune Responses to B. pertussis and B. bronchiseptica. ............................21
Innate Immune Responses. .........................................................................22
B cell responses. .........................................................................................24
T cell responses...........................................................................................26
Currently Available Vaccines against B. pertussis...........................................27
Vaccines against B. bronchiseptica................................................................29
References. .....................................................................................................33
CHAPTER II ( Published in Journal of Bacteriology 2007 May; 189(10): 3695-
704).....................................................................................................................48
iv

Introduction .....................................................................................................49
Materials and Methods ....................................................................................52
Results ............................................................................................................63
Discussion.......................................................................................................91
References......................................................................................................96
CHAPTER III (Published in Infection and Immunity 2009 Feb;77(2);885-95)
..........................................................................................................................100
Introduction ...................................................................................................101
Materials and Methods ..................................................................................105
Results ..........................................................................................................113
Discussion.....................................................................................................140
References....................................................................................................147
CHAPTER IV: DISCUSSION............................................................................152
Potential roles of BipA and BcfA in mediating respiratory tract colonization........
by Bordetella. ................................................................................................153
BvgAS-mediated regulation of bcfA expression. ...........................................158
Differential phase specific expression profile of BipA and BcfA. ...................162
BcfA as a vaccine candidate against B. bronchiseptica. ...............................163
BcfA as a Th1 response inducing adjuvant. ..................................................165
BcfA as a vaccine candidate against B. pertussis. ........................................167
References. ...................................................................................................171
Curriculum Vitae............................................................................................174
v

LIST OF FIGURES
CHAPTER I
Figure 1 Clinical manifestation of whooping cough and the corresponding B. pertussis load in infected individuals..................... 4 Figure 2 The BvgAS two component system of Bordetella spp..................... 10
CHAPTER II
Figure 1 Schematic representation of the similarity of BcfA with BipA............65 Figure 2 Quantification of bcfA transcriptional activity by β-galactosidase
assays in the wt, Bvg+, Bvgi and Bvg- phase locked strains of B. bronchiseptica...................................................................................69
Figure 3 Determination of the phase-dependent expression profiles of different Bvg-regulated genes in B. bronchiseptica by real time RT-PCR analysis...............................................................................71 Figure 4 Kinetics of transcriptional activation of different Bvg-activated genes and the Bvg-independent gene recA......................................75 Figure 5 The putative promoter region of bcfA................................................78 Figure 6 Electrophoretic Mobility Shift Assay...................................................81
Figure 7 BcfA is localized to the outer-membrane...........................................84
Figure 8 BcfA is expressed during infection.....................................................86
Figure 9 Colonization of rat respiratory tract by wt and isogenic mutant derivatives RB25 (ΔbipA), RKD110 (ΔbcfA) and MM101 (ΔbipAΔbcfA)......................................................................................90
vi

CHAPTER III
Figure 1 Immunization with BcfA protects mice against B. bronchiseptica challenge.........................................................................................115
Figure 2 Immunization with BcfA reduces lung pathology in mice challenged with RB50......................................................................118 Figure 3 Anti-BcfA antibody titers in immunized mice....................................122 Figure 4 Effect of adoptive transfer of BcfA-specific sera on respiratory tract colonization..............................................................................127 Figure 5 Opsonization with anti-BcfA serum enhances the phagocytosis of RB50 by J774 murine macrophages............................................130 Figure 6 Neutrophils are required for anti-BcfA antibody-mediated clearance of B. bronchiseptica.........................................................133 Figure 7 BcfA-induced production of IFN-γ and IL-4 in splenocytes..............137 Figure 8 Expression of BcfA among clinical isolates of B. bronchiseptica.....139 Figure 9 Model for BcfA-mediated protective immunity.................................145
CHAPTER IV
Figure 1 Potential roles of BipA and BcfA in B. bronchiseptica pathogenesis...................................................................................157
Figure 2 Model illustrating relative occupancy of BvgA-P to putative bcfA
promoter..........................................................................................161 Figure 3 Expression of BcfA by B. pertussis strains......................................170
vii

LIST OF TABLES
CHAPTER II Table 1 Strains and plasmids used in this study..............................................53 Table 2 Oligonucleotide primers used in this study.........................................56 CHAPTER III Table 1 Strains used in this study..................................................................106 Table 2 BcfA immunization reduces lung pathology......................................120
viii

LIST OF ABBREVIATIONS
ADP………………………………………………...Adenine Diphosphate
AIDS………………………………………………..Acquired Immune Deficiency
Syndrome
ATP…………………………………………………AdenineTriphosphate
BcfA....................................................................Bordetella Colonization Factor A
BipA....................................................................Bordetella Intermediate Phase
Protein A
Bvg………………………………………………….Bordetella virulence gene
BrkA....................................................................Bordetella Resistance to Killing A
cAMP……………………………………………….Cyclic Adenine Monophosphate
CDC………………………………………………...Centers for Disease Control
CFU………………………………………………....Colony Forming Units
CR…………………………………………………..Complement Receptor
CyaA………………………………………………. Adenylate Cyclase Toxin
D…………………………………………………….Aspartic Acid
DNA....................................................................Deoxyribo Nucleic Acid
DNT…………………………………………………Dermonecrotic Toxin
DTP…………………………………………………Diptheria-Tetnus-Pertussis
EMSA………………………………………………Electrophoretic Mobility Shift Assay
FHA…………………………………………………Filamentous Hemagglutinin
Fim………………………………………………….Fimbriae
G........................................................................Guanine nucleotide
ix

GTP…………………………………………….......Guanosine Triphosphate
H…………………………………………………….Histidine
HPt………………………………………………….Histidine Phosphotransferase
IAP………………………………………………….Intergrin Associated Protein
ICAM……………………………………………….Intercellular Adhesion Molecule
ID.......................................................................Infectious Dose
IFN-γ..................................................................Interferon-gamma
Ig……………………………………………………Immunoglobulin
IL……………………………………………………Interleukin
ITB......................................................................Infectious Tracheobronchitis
LPS.....................................................................Lipopolysaccharide
LRI......................................................................Leukocyte Response Intergrin
MHC...................................................................Major Histocompactability Complex
MgSO4................................................................Magnesium Sulphate
NO......................................................................Nitric Oxide
ORF………………………………………………...Open Reading Frame
PBP....................................................................Periplasmic Binding Protein
Pc........................................................................Pertussis Acellular vaccine
PCR………………………………………………...Polymerase Chain Reaction
PMN………………………………………………..Polymorphonuclear Leukocytes
PRDC.................................................................Porcine Reproductive and
Respiratory Disease Complex
Prn………………………………………………….Pertactin
x

PT…………………………………………………..Pertussis Toxin
Pw.......................................................................Pertussis Whole cell vaccine
RGD...................................................................Arginine-Glycine-Aspartic Acid
RT-PCR……………………………………………Reverse Transcriptase-Polymerase
Chain reaction
RTX………………………………………………...Repeats in Toxin
SCID..................................................................Severe Combined Immune
Deficiency
Th......................................................................T-Helper
TCT..…………………………………………........Tracheal Cytotoxin
TNF....................................................................Tumor Necrosis Factor
TLR………………………………………………...Toll Like Receptors
Wt…………………………………………………..Wild type
xi

ABSTRACT
Sukumar, Neelima
TRANSCRIPTIONAL REGULATION AND PROTECTIVE EFFICACY OF
BORDETELLA COLONIZATION FACTOR A (BcfA) IN BORDETELLA INFECTIONS
To successfully colonize their mammalian hosts many bacteria produce
multiple virulence factors that play essential roles in disease processes and pathogenesis. Some of these molecules are adhesins that allow efficient attachment to host cells, a prerequisite for successful host colonization. Bordetella spp. express a number of proteins which either play a direct role in attachment to the respiratory epithelia or exhibit similarity to previously known bacterial adhesins. One such recently identified protein is BipA. Despite similarity to intimins and invasins, its deletion from B. bronchiseptica did not result in any significant defect in respiratory tract colonization. We hypothesized the existence of a paralogous protein that could complement the function of BipA. In the studies described here, we report the identification and characterization of an ORF in B. bronchiseptica, designated as bcfA (Bordetella colonization factor A) that is similar to bipA. We show that in contrast to maximal expression of bipA in the Bvgi phase, bcfA is expressed at high levels in both the Bvg+ and the Bvgi phases. We have identified multiple sequence elements resembling the consensus BvgA binding site in the bcfA promoter region. Direct binding of purified BvgA to the bcfA promoter revealed differences in the DNA binding profiles of BvgA and BvgA-P to the promoter region. Utilizing an antibody raised against BcfA, we show that BcfA is localized in the outer membrane. Finally, we demonstrate that simultaneous deletion of both bipA and bcfA results in a defect in colonization of the rat trachea and that BcfA is expressed during Bordetella infectious cycle.
Based on our findings that BcfA is an outer membrane immunogenic protein
and is critical for murine respiratory tract colonization, we examined its utility in inducing a protective immune response against B. bronchiseptica in a mouse model of intranasal infection. Mice vaccinated with BcfA demonstrated reduced pathology in the lungs and harbored lower bacterial burdens in the respiratory tract. Immunization with BcfA led to the generation of BcfA-specific antibodies in both the serum and the lungs and passive immunization led to the reduction of B. bronchiseptica in the trachea and the lungs. These results suggest that protection after immunization with BcfA is mediated in part by antibodies against BcfA. To further investigate the mechanism of BcfA-induced immune clearance, we examined the role of neutrophils and macrophages. Our results demonstrate that neutrophils are critical for anti-BcfA antibody-mediated clearance and that opsonization with anti-BcfA serum enhances phagocytosis of B. bronchiseptica by murine macrophages. We show that immunization with BcfA results in the production of IFN-γ and subclasses of IgG antibodies that are consistent with the induction of a Th1 type immune response. In
xii

combination, our findings suggest that mechanism of BcfA-mediated immunity involves humoral and cellular responses. Expression of BcfA is conserved among multiple clinical isolates of B. bronchiseptica. Our results demonstrate the striking protective efficacy of BcfA-mediated immunization thereby highlighting its utility as a potential vaccine candidate. These results also provide a model for the development of cell-free vaccines against B. bronchiseptica.
xiii

CHAPTER I: INTRODUCTION
The Bordetella Genus.
Bordetellae are Gram- negative aerobic cocobaccilli that preferentially attach
to the ciliated respiratory epithelium of mammals (95). Currently there are ten known
Bordetella species. Among these, B. bronchiseptica (44), B. pertussis (12) and B.
parapertussis (40) are the most well-studied and are known as the classical species.
Other known species include B. avium, B. hinzii, B. holmesii, B. trematum, B. ansorpii
and B. petrii. B. bronchiseptica causes respiratory infections in a wide range of four
legged animals such dogs, cats, pigs, horses, non human primates (49) and
occasionally humans (151). Unlike B. bronchiseptica, B. pertussis has a restricted
host range and only infects humans (17). B. parapertussis exists in two host adapted
subspecies. B. parapertussishu is associated with respiratory illness in humans while
B. parapertussisov is ovine adapted and causes chronic respiratory infections in
sheep (60, 116). Based on a combination of different phylogenetic analysis tools such
as multilocus enzyme electrophoresis, insertion sequence polymorphisms and
genome sequencing, it is predicted that B. bronchiseptica is the ancestral organism
from which B. pertussis and B. parapertussis evolved independently into host
restricted species (35, 46, 114, 142). B. avium causes infections in poultry and is also
an opportunistic human pathogen (128, 134). Although majority of the Bordetella spp
are associated with respiratory tract infections, there are multiple species that have
variant pathogenic and survival characteristics. B. trematum has never been
associated with the respiratory tract, but has been isolated from ear and skin wound
1

infections (143). On the other hand, another unique species, B. petrii was isolated
from the environment and is considered to be the progenitor from which other
pathogenic Bordetella species evolved (144, 145). The range of differences in host
adaptation, virulence traits and survival niches highlight the versatility of the
Bordetella genus as a pathogen.
Bordetella as a Pathogen.
B. pertussis is the etiological agent of the highly communicable disease called
pertussis or whooping cough. This illness is most severe in children and can result in
mortality (52). Despite the availability and widespread use of vaccines, World Health
Organization estimates 48.5 million cases and 294,000 deaths worldwide resulting
from this disease in 2002. According to the Centers of Disease Control (CDC) in the
United States, more than 25,000 cases of pertussis were reported in the year 2004,
which is the highest incidence since 1959. In addition to its impact on infants and
young children, there is increasing prevalence of pertussis in adults and adolescents
(64). Pertussis is circulating and is highly ubiquitous in adults and adolescents of all
ages, even in previously infected or vaccinated populations. In fact, in the United
States, multiple reports estimate 1 million pertussis cases each year in this age group
(21). Moreover, case studies conducted in different countries including United States
reveal that more than 20% of adolescents and adults having prolonged cough illness
are colonized by B. pertussis (47, 109, 125). Although this disease is not as severe in
this age group, the significance of increasing incidence lies in the fact that
2

FIG. 1. Clinical manifestation of whooping cough and the corresponding B. pertussis
load in infected individuals. Following the 7-10 incubation period, the catarrhal phase
is manifested and is characterized by common cold-like symptoms. It is during this
stage that the bacterial numbers reach a peak and infected persons are at the
highest risk of transmission to uninfected individuals. The paroxysmal stage is
distinguished by the characteristic whooping cough and can last from 2 to 6 weeks.
The bacterial load begins to decline in this stage making the clinical diagnosis
difficult. The convalescent phase is the recovery stage. This phase is characterized
by the reduced incidence and severity of paroxysms and can last from 3 to 4 weeks
or upto several months.
3

Phases
Catarrhal
Paroxsymal
Convalescent
Bac
teria
l Num
bers
Incubation
3-4 weeks
or longer
7-10 days
1-2 weeks
2-4 weeks
Duration
4

adolescents and adults act as source for B. pertussis infections in partially immune
and nonimmune children especially in household settings.
The clinical manifestation of this disease occurs in three phases- catarrhal,
paroxysmal and convalescent (19). Following the 7-10 days incubation period, the
first stage of infection, the catarrhal phase, is manifested with runny nose, sneezing,
low grade fever and occasional mild cough (Fig. 1). These symptoms are similar to
the common cold or minor respiratory tract infections and thus pertussis is often
ignored during the initial diagnosis. This stage lasts for 1-2 weeks after which the
cough gradually attains severity and develops into the distinctive whooping cough in
the paroxysmal stage. The paroxysmal cough is characterized by bursts or
paroxysms of rapid, numerous (5-10), forceful coughs in a single expiration followed
by a massive inhalation, producing the characteristic whoop sound. Concurrent with
the paroxysms, cyanosis, eye bulging and posttussive vomiting can occur. The
paroxysmal stage can last 1-6 weeks and can even persist upto 10 weeks. It is during
the paroxysmal stage that pertussis is usually suspected; however the bacterial loads
begin to decline making the clinical diagnosis difficult. Recovery occurs during the
convalescent phase and is characterized by the gradual reduction in the incidence
and severity of paroxysms. The cough may completely disappear by 2-3 weeks or it
can take upto several months. Complications are common and include pneumonia,
otitis media, seizures and encephalopathy (95). In adults and adolescents and
partially immunized children, pertussis is manifested as a milder disease or may be
asymptomatic. B. pertussis infections in these persons may result in mild or
persistent, severe cough without the distinguishing whoop (37, 87, 88).
5

B. parapertussishu infections can result in mild pertussis like illness or severe
classic pertussis (10, 41). Coexistence and contributions of B. parapertussis in
pertussis infections may be underestimated due to difficulties in distinguishing
between the two human adapted species (68, 76).
B. bronchiseptica is the primary etiological cause and/or pre disposing factor
in a variety of veterinary infections such as Porcine Reproductive and Respiratory
Disease complex (PRDC), pneumonia and atrophic rhinitis in swine, Infectious
tracheobronchitis (ITB, Kennel Cough) in dogs and bronchopneumonia in guinea
pigs, rats, mice, rabbits, cats and non human primates (16, 95). Upper respiratory
illness by B. bronchiseptica in infant pigs is characterized by coughing and sneezing
followed by deformation of the nose bony structures leading to atrophy. In swine
populations, this disease is widely prevalent and results in loss of 17 million dollars
annually in the United States. Moreover, infection with B. bronchiseptica predisposes
pigs to infection by other bacterial and viral pathogens thus contributing to a multi-
factorial disease condition PRDC, which has an annual economic impact of 40 million
dollars in the United States. In dogs, the highly contagious ITB can manifest as mild
illness or severe cough with pneumonia. Although vaccination can considerably
reduce the severity of disease, B. bronchiseptica is frequently isolated from nasal
cavities of immunized animals, suggesting persistence of this pathogen in animals.
Although mainly implicated as an animal pathogen, there have been many recent
reports of B. bronchiseptica infections in humans. The majority of human infections
by B. bronchiseptica occur in immuno-compromised individuals such as AIDS and
cystic fibrosis patients (39, 71, 135). However, it has been also isolated from an
6

immunocompetent individual (118). There are also several reports of zoonotic
transmission of this organism from farm or pet animals (151).
The BvgAS Signal Transduction System.
Bordetellae control the expression of majority of their known virulence genes
and other factors through the BvgAS signal transduction system. In general, bacteria
utilize complex signaling mechanisms for eliciting adaptive responses to enhance
survival in constantly changing environment. These signaling systems are devised to
detect fluctuations in the environment and to trigger subsequent changes in gene
expression. The most common regulatory mechanism utilized by bacteria is the two
component signal transduction system consisting of a sensor kinase and a response
regulator. Typically, extracellular signals are processed by transfer of phosphate
group from histidine residue in the sensor to aspartate residue in the receiver (53).
The bvgAS locus encodes for a polydomain transmembrane sensor, BvgS and a
cytoplasmic response regulator, BvgA (28). This virulence regulatory system is 96 %
identical at the nucleotide level among B. pertussis, B. parapertussis and B.
bronchiseptica and is functionally interchangeable (94). However, BvgAS belongs to
a class of bacterial two component signal transduction systems, which deviates from
the biphasic paradigm in that communication between the transmembrane sensor
kinase (BvgS) and the cytoplasmic response regulator (BvgA) occurs via a
sophisticated four step His-Asp-His-Asp phosphorylation cascade (Fig. 2) (141).
Similar to other regulatory proteins, both BvgS and BvgA function as dimers. BvgS at
its N terminus contains two periplasmic binding protein domains (PBP) followed by
transmembrane linker region and cytoplasmically localized autokinase, receiver and
7

the C terminal histidine phosphotansferase (HPt) domains. In response to changes in
environmental signals, the autokinase domain catalyzes ATP hydrolysis and
phosphorylation of histidine (H) 729. The γ-phosphate moiety is subsequently
transferred to aspartic acid (D) 1023 on the receiver domain and then to H1172 of the
HPt domain. These intramolecular phosphotransfers eventually result in the
phosphorylation of BvgA (Fig. 2). BvgA is a typical response regulator composed of
N terminal receiver and C terminal DNA binding domains. Phosphorylation causes
dimerization of BvgA and enhances its capacity to bind Bvg-regulated promoters and
regulate transcription (9, 14, 127). This can either result in transcriptional activation or
repression of cognate genes based on the affinity and location of BvgA binding sites
with respect to the transcription initiation site (38).
A hallmark feature of the BvgAS signal transduction system is that it can cause
phenotypic transition of Bordetella among three known phases-Bvg+, Bvgi (Bvg-
intermediate) and Bvg- and potentially multiple unknown states (28). Bvg+ is the
virulent phase and is characterized by the expression of adhesins such as
filamentous hemagglutinin, fimbriae and pertactin and toxins such as adenylate
cyclase, dermonecrotic toxin and pertussis toxin in the case of B. pertussis, all
contributing to attachment and subsequent invasion. Bvg+ phase is sufficient for
establishing respiratory tract colonization (29). In the Bvg- phase, the avirulent stage,
BvgAS system is inactive and there is suppression of Bvg-activated loci and
expression of Bvg-repressed genes such as flagella (2, 3) and urease (99, 100) in B.
bronchiseptica. Bvgi phase is characterized by the expression of some of the Bvg+
phase factors and the maximal expression of a set of antigens of which only one-
8

FIG. 2. The BvgAS two component system of Bordetella spp. BvgS is the
transmembrane polydomain sensor kinase consisting of two periplasmic binding
protein domains, autokinase, receiver and histidine phosophotransferase domains.
When active, autokinase domain catalyzes the hydrolysis of ATP and initiates a
series of intramolecular phosphotransfers that eventually leads to the phosphorylation
of the cytoplasmic response regulator BvgA. Phosphorylated BvgA regulates
activation or repression of transcription of genes. When the BvgAS system is active,
Bordetella is in a Bvg+ phase and expresses factors with known or hypothesized role
in virulence such as adhesins (FHA, Fim, Prn) and toxins (CyaA, PT). Inactivation of
BvgAS system renders Bordetella to Bvg- phase. Under these conditions, Bvg-
activated factors are not expressed, but Bvg-repressed factors such as flagella and
urease are expressed. Bordetella also exsist in a Bvgi phase. Although the
phosphorylation status of BvgAS system is unknown in this phase, it is postulated
that BvgA-P is in an intermediate levels as compared to the Bvg+ and Bvg- phases.
The only known Bvgi phase- specific protein is BipA. In the laboratory, Bordetella can
be modulated between the different phases using nicotinic acid, MgSO4 or growth at
different temperature conditions.
9

P
BvgA
BvgS
Sulfate <30 C Nicotinic acid ·
Phosphotransfers
MotilityBvg
UreaseBipA
Bvgi
Bvg + Adenylate cyclase toxin
Pertactin Fimbriae
FHA
10

BipA has been identified at the molecular level (30, 138). In the Bvgi phase,
Bordetella exhibits decreased virulence and improved survival under nutrient limiting
conditions. Although the signals required for regulation of BvgAS in vivo have not yet
been identified, under laboratory conditions, modulation among these phases can be
achieved by varying temperature, sulphate anion or nicotinic acid concentrations (83)
or through specific mutations of the BvgAS system.
The third component of the bvg locus encodes for a regulatory protein called
BvgR. bvgR is located directly downstream of bvgAS and is transcribed convergently
(104). BvgR activity is induced by transcriptional activation of bvgR by activated BvgA
(103). BvgR is involved in the transcriptional repression of Bvg-repressed genes in
the Bvg+ phase. Deletion analysis revealed that the absence of BvgR-mediated
regulation of Bvg-repressed genes is detrimental for colonization by B. pertussis in a
mouse model of infection (105). This and other studies indicate that ectopic
expression of Bvg repressed genes can impede the development of infection (1,
105). Although the exact roles of Bvg repressed genes are unknown, it is
hypothesized that these factors are involved in survival outside the host and under
nutrient deprived conditions. Previous studies have demonstrated that Bvg-activated
factors are necessary and sufficient for colonization of the respiratory tract. On the
other hand, Bvgi is postulated to be important for transmission of bacteria, although
there is no experimental evidence for this hypothesis (138). These data therefore
suggests that a possible role for Bvg regulon is to sense whether the bacteria is
inside a host or outside and accordingly regulate expression of vital genes (28).
11

Virulence Factors. Bordetella spp produce an array of virulence determinants which
can be categorized into two broad groups- adhesins and toxins. Expression of
majority of these virulence associated factors is coordinately regulated by the BvgAS
system. Efficient colonization of the respiratory tract by Bordetellae requires the
interplay of multiple factors. The current literature provides evidence for unique
properties as well as synergistic and antagonistic roles for these factors in virulence
functions such as attachment, invasion, modulation and suppression of immune
responses. Furthermore, the presence of many of these antigens in currently
available vaccines, exemplify the importance of these factors in Bordetella
pathogenesis. Therefore, a better understanding the roles of these virulence
components in Bordetella-host interactions will contribute to the development of more
efficient therapeutic techniques as well as vaccines.
Adhesins. This section is a brief overview of the major adhesins expressed by
Bordetella spp along with their functions in pathogenesis.
Filamentous hemagglutinin (FHA).
One of the dominant adherence factors of Bordetellae is FHA; a large highly
immunogenic protein which is both surface associated and secreted in the
extracellular milieu (32, 33). FHA is a component of the currently available B.
pertussis acellular vaccines. In vitro studies utilizing different mammalian cell lines
indicate that FHA contains four binding domains that aid in attachment of Bordetella
to respiratory epithelium or host immune cells. The Arginine-Glycine-Aspartic Acid
12

(RGD) motif stimulates adherence to macrophages and leukocytes potentially
through the Complement Receptor Type 3 (CR3) and Leukocyte Response
Intergrin/Intergrin-Associated Protein (LRI/IAP) complex (74, 119). The RGD domain
has also been shown to facilitate attachment of Bordetella to bronchial epithelial cells
through interactions with very late antigen 5 (VLA-5) (75). This interaction results in
the upregulation of epithelial intercellular adhesion molecule 1 (ICAM-1) via the NFκB
signaling pathway. The carbohydrate recognition domain of FHA mediates adherence
to ciliated epithelial cells of the respiratory tract and macrophages (117). In addition,
FHA also has a heparin binding domain, which promotes binding to sulfated
carbohydrates (102). Finally FHA also possess a CR3 recognition motif, function for
which has not yet been identified. Furthermore, inhibition of CD4+ T cell proliferation
and induction of apoptosis by B. pertussis has been demonstrated to be FHA-
dependent (13). Studies using purified FHA revealed that this protein induces
immunosuppressive effects on macrophages and dendritic cells by downregulating
production of Interleukin (IL)-12 in an IL-10 dependent manner (97, 98).
In vivo studies utilizing mutant strains having in frame deletion of fha revealed
that FHA is essential for tracheal colonization by B. bronchiseptica (31). A role for
FHA in B. pertussis colonization has been more difficult to discern due to inconsistent
results. Utilizing mouse models, McGuirk et al. showed that in the absence of FHA, B.
pertussis showed defective colonization of lungs, while others found no difference
between the wild type strain (wt) and FHA mutant derivatives in their ability to
colonize the respiratory tract (50, 148). These conflicting results may be due to the
fact that mice are not natural hosts for B. pertussis and these models fail to represent
13

natural course of infection. Comparison of genome sequences from B. bronchiseptica
and B. pertussis indicates that these two closely related species encode FHA which
is similar but not identical. In fact, studies utilizing B. bronchiseptica strains
ectopically expressing B. pertussis FHA (FHA ) fail to colonize the rat trachea.
FHA could mediate attachment to epithelial cells in vitro but failed to protect B.
bronchiseptica from inflammation-mediated clearance (73). These studies highlight
the importance of FHA in modulating the immune responses and effecting successful
colonization and persistence of B. bronchiseptica.
Bp
Bp
Pertactin (Prn).
Prn is a Bvg-regulated surface associated protein belonging to the
autotransporter secretion system. Typically, these proteins direct their own export
across the outermembrane. The highly conserved C terminal β- barrel domain of
these proteins facilitates the transport of the N terminal passenger domain, which
confers the effector function (36). Prn contains the tripeptide RGD domain and is
proposed to function as an adhesin (42). However, in vitro studies comparing wt type
parent strain and mutants lacking Prn revealed no significant role for this protein in
adherence or invasion of HEp 2 cells (122). Additionaly prn- mutants of both B.
pertussis and B. bronchiseptica do not differ from their parental wt strains in the
ability to colonize and persist within the respiratory tracts in vivo. In contrast to the in
vitro and in vivo studies described above that fail to demonstrate a precise function
for this protein in Bordetella pathogenesis, data from several vaccine efficacy trials
conducted during the early 1990s demonstrate that antibodies against Prn are the
14

most vital to confer protection, suggesting a significant role in protective immunity
(20, 139). These vaccine efficacy evaluation studies have also suggested that
inclusion of Prn in acellular vaccines containing FHA and Pertussis toxin, augment its
efficacy in preventing B. pertussis infection (108). Moreover, a role for anti-Prn
antibodies in efficient phagocytosis of B. pertussis has been demonstrated (63).
Fimbriae (Fim).
Many Gram negative bacteria express filamentous, polymeric structures
localized to the cell surface called fimbriae (Fim). Bordetellae synthesize four fimbrial
serotypes including the predominantly expressed Fim2 and Fim3 and FimX and FimN
which are expressed only at low levels (78, 85, 111, 121). Several studies have
demonstrated a role for Fim in mediating adherence of Bordetella to the respiratory
epithelium and monocytes (58, 59). In vivo studies revealed that Fim is involved in
the efficient establishment of tracheal colonization and persistence (96). Fim is also
critical for production of appropriate serum antibody responses. Specifically, Fim aids
in eliciting an immunoglobulin (Ig) M response early during the infection as well as in
inducing the IgG2a component of host humoral immunity in a rat model of infection
(96). Furthermore, studies in mice showed that Fim is vital for inducing an anti-
inflammatory response and preventing killing of Bordetella by alveolar macrophages
(95). Fim is also a component of the accelluar vaccines and vaccine trials in children
suggested that antibody to Fim contributes to protective immunity. Moreover,
inclusion of Fim2/3 in vaccines has been shown to significantly enhance efficacy
(113).
15

Toxins. A different category of virulence determinants expressed by Bordetella spp
are toxins. The following section briefly discusses the toxins secreted by Bordetella
and their roles in bacterial invasion and host inflammatory responses.
Adenylate cyclase (CyaA).
CyaA is a bifunctional adenylate cyclase and hemolysin expressed by all three
classical species of Bordetella. The short 400 amino acid N terminal region confers
the catalytic activity of CyaA while the longer 1300 amino acid C terminal region
mediates the translocation of the catalytic domain into mammalian cell cytosol as well
as its function as a hemolysin (65). This calmodulin- sensitive toxin belongs to the
repeats in toxin (RTX) family of calcium dependent, pore forming cytotoxins (120,
124, 126). The receptor for CyaA has been identified as CD11b, a cell surface
glycoprotein, expressed by myeloid cells such as macrophages, dendritic cells,
neutrophils and natural killer cells. On entry into a mammalian cell, calmodulin
activates CyaA, which then catalyzes the enhanced conversion of cellular ATP to
cyclic AMP (cAMP). The supraphysiologic quantities of cAMP thus synthesized
interfere with intracellular signaling and result in altered cell physiology (23, 24, 150).
In many of the host immune effector cells, intoxication by cAMP results in inhibition of
bacteoricidal functions. Purified CyaA inhibits super oxide generation and chemotaxis
of peripheral blood monocytes and neutrophils. In addition, CyaA induces apoptosis
in macrophages and inhibits phagocytosis of B. pertussis by neutrophils (146, 147).
Furthermore studies using bone marrow derived dendritic cells revealed that CyaA
promoted upregulation of MHC class II and costimulatory molecule (CD80, CD83 and
16

CD86) expression and suppressed production of proinflammatory cytokines such as
IL-12 and Tumor Necrosis Factor (TNF)α (132). In accordance with its
immunomodulatory role in vitro, in vivo studies show that CyaA deficient mutants are
defective in causing lethal infection in infant mice (54, 148). These data thus suggest
an anti-inflammatory and anti-phagocytic role for CyaA in Bordetella infections.
Although not a component of acellular vaccines, studies has shown that
primary infection with B. pertussis induces anti-CyaA antibodies in children (22).
Moreover, studies utilizing convalescent serum samples revealed that antibodies
against CyaA promoted phagocytosis of B. pertussis by human neutrophils, thus
suggesting a role in infection-induced immunity (110).
Tracheal Cytotoxin (TCT).
TCT is a peptidoglycan derived disaccharide–tetrapeptide monomer
synthesized commonly by all gram-negative bacteria during growth and cell division.
Typically this monomer is recycled by recovering it back into the cell cytoplasm via a
membrane protein called AmpG (25, 26, 77). Bordetella spp. lack functional AmpG
and thus release this peptidoglycan fragment into the environment. The potency of
this toxin is reflected by the fact that TCT alone is sufficient and necessary to induce
specific cytopathology characteristic to B. pertussis infections in ciliated cells of
tracheal explants (48). TCT causes ciliostasis, cell blebbing and mitochondrial
damage. Additionally, TCT induces IL-1 α production and consequently nitric oxide
(NO) production in hamster tracheal epithelial cells (61, 62). TCT triggered NO
production is proposed to mediate annihilation of ciliated cells. In vivo, it is
17

hypothesized that TCT induces IL-1 production in non ciliated mucus secreting cells.
The resultant NO diffuses through the neighboring ciliated cells and causes severe
damage to the epithelium (45).
Dermonecrotic Toxin (DNT).
DNT is a heat labile A-B toxin consisting of an N terminal receptor-binding
domain and C terminal catalytic domain (34). The receptor for DNT has not yet been
identified. When intradermally injected, DNT induces necrotic lesions in laboratory
animals such as mice, guinea pigs and rabbits and is lethal for mice when delivered
intravenously (11, 72). In vitro studies utilizing purified DNT has shown that this toxin
stimulates modification and activation of the small GTP-binding protein Rho leading
to induction of DNA and protein synthesis, alterations to cell cytoskeleton and
inhibition of cell division (69, 70). DNT has also been associated with turbinate
atrophy and bronchopneumonia in B. bronchiseptica infected pigs (90, 123).
Pertussis Toxin (PT).
Unlike other toxins discussed above, PT is preferentially expressed only by B.
pertussis. Although B. bronchiseptica and B. parapertussis genomes contain genes
that can potentially express PT, mutations present in the promoters cause the genes
to be transcriptionally silent (4). PT is an ADP- ribosylating AB toxin composed of six
polypeptides designated as S1 to S5. The A subunit of the toxin comprises of S1
polypeptide and the B subunit is pentameric and is composed of S2, S3, S5 and two
S4 subunits (86, 112). S2 to S5 polypeptides form a ring like structure with S1 subunit
18

atop (79, 129). The B subunit mediates binding to eukaryotic cell membrane and
transports the enzymatically active S1 subunit into the cytoplasm, where it catalyzes
the transfer of ADP ribose to Guanine nucleotide (G) binding proteins. The ADP
ribosylation of different isoforms of G proteins leads to disruption of signal
transduction pathways within the cell (79). Studies have attributed both
immunomodulatory as well as an attachment function for PT (140). PT has been
suggested to mediate adherence of B. pertussis to ciliated epithelium of the
respiratory tract as well as immune cells such as macrophages. On the other hand,
PT has also been shown to have immunosuppressive effects. Studies revealed that
PT inhibits chemotaxis, oxidative responses and lysosome release in macrophages
and neutrophils (15, 101). In another study, a mutant strain lacking PT displayed
higher serum anti-Bordetella antibody responses as compared to the wt strain (18).
Nevertheless, PT is proposed to be the principal virulence factor responsible for all
the major pertussis-associated typical disease symptoms such as leukocytosis and
lymphocytosis and is a component of all the currently available acellular vaccines.
Animal Models.
Multiple animal models have been developed to gain insights into the function
of various virulence determinants and to study the progression of Bordetella
pathogenesis. For B. bronchiseptica, pathogen free rabbits, rats and mice are
commonly utilized. The infectious dose (ID)50 dose of this species for intranasal
inoculation is less than 200 Colony Forming Units (CFU) for rabbits, 25 CFU for rats
and 5 CFU for mice (95). Previous studies have shown that these model systems can
19

accurately simulate the characteristics of a natural infection by B. bronchiseptica. For
intranasal challenge either a high volume (25-50 μl) or low volume (5-10 μl) can be
used to deliver bacteria and both of these inoculation regimen lead to efficient
establishment and persistence in all animal models. In rats and mice, the nasal cavity
gets colonized persistently, while the trachea and larynx get cleared by 50-60 days
post inoculation with B. bronchiseptica (133). Apart from the upper respiratory tract,
the high volume treatment also consistently delivers bacteria to the lungs, which
eventually gets cleared by 50-70 days post inoculation (57).
Even though humans are the only known host for B. pertussis, a number of
animal species are used to study immune responses to this organism. In contrast to
B. bronchiseptica, for B. pertussis, a large infectious dose in a large volume is
commonly used to infect animals. The most prevalently used is the murine model of
intranasal or aerosol infection. Intranasal infection of mice with 5 x 105 CFU of
respective strains in 50 μl volume consistently and reproducibly delivers bacteria to
the nasal cavity, larynx, trachea and lungs (57). Unlike B. bronchiseptica, B. pertussis
does not persist for the life in mice and gets cleared from the respiratory tract by 20-
70 days post inoculation. Although mice do not display overt symptoms of the human
disease, the intranasal model replicates many of the attributes of pertussis: i) bacteria
rapidly multiply and the infection is limited to the respiratory tract, ii) young animals
display comparatively severe infections, iii) various systemic physiological and
neurological changes characteristic of human infection are observed in mice. One of
the apparent drawbacks of the murine model, for which it is often criticized, is its
inability to display the characteristic paroxysmal cough. Additionally, a rat model of B.
20

pertussis that can reproduce human pertussis illness with respect to course of
infection and cough production has been developed. This involves intrabronchially
injecting rats with agarose beads coated with challenge strain (55, 56).
Animal models are also critical for studying the protective efficacy of vaccines
and its pathophysiological responses. The gold standard to test the potency of
vaccines against B. pertussis is the lethal intracerebral challenge model or the
Kendrick test. In this model, vaccines are assessed based on its ability to protect
immunized mice against a lethal intracerebral challenge. Surprisingly, although the
previously available B. pertussis whole cell vaccines protect mice against the lethal
intracerebral challenge, the currently available new generation acellular vaccines fail
to pass the Kendrick test (27).
Knock out mice and immunodeficient mice such as Severe Combined
Immunodeficiency (SCID) mice have been frequently utilized for investigating the
interaction of Bordetella spp with the host immune system as well as to discern the
immunomodulatory role of specific virulence factors (73).
Immune Responses to B. pertussis and B. bronchiseptica. Bordetella spp exploit
both extracellular and intracellular host niches and employ different pathogenic
strategies to subvert immune responses. The majority of studies exploring immune
responses to Bordetella infection focus on development of new vaccine approaches,
its immunological properties and effector mechanisms that contribute to protection
against the human pathogen B. pertussis. Studies also provide evidence of
21

complimentary roles for both innate and adaptive immunity in bacterial clearance
subsequent to infections as well as vaccinations (57, 108).
Innate Immune Responses.
Neutrophils (PMN) and Macrophages.
PMNs are a major component of the initial immune responses vital for clearing
B. bronchiseptica infections. Infection with B. bronchiseptica results in rapid
recruitment of neutrophils to the lungs of infected mice. Consistent with this
observation, neutropenic mice succumb to B. bronchiseptica infections within 1-4
days post inoculation. Furthermore in vitro cytotoxicity assays using murine
macrophage cell line J774 and tunnel assay of B. bronchiseptica infected mice lungs
suggested that this species may induce apoptosis of alveolar macrophages. In
contrast, B. pertussis infection leads to cellular infiltration in lungs consisting
predominantly of macrophages and to a lesser extent neutrophils. In fact, neutropenic
mice survive B. pertussis infection. These studies thus demonstrate the differences in
immune responses elicited to the animal adapted and the human adapted Bordetella
spp and that alveolar macrophages are critical for limiting B. pertussis infection while
neutrophils are required for B. bronchiseptica clearance (57).
Toll like receptors (TLR).
Several studies have revealed a role for Toll like receptors (TLR) in innate
immune responses to Bordetella spp in mice. TLR4 is critical for LPS- induced
22

proinflammatory cytokine production in response to B. bronchiseptica infection.
Furthermore, TLR4 deficient mice show very drastic pathology in lungs and succumb
to B. bronchiseptica infection as early as 3 days post inoculation (93). Studies
investigating the mechanism of antibody mediated clearance of B. bronchiseptica
revealed that TLR4 is critical for recruitment of neutrophils and subsequent clearance
of antibody and complement opsonized bacteria (82). B. pertussis also causes a
more severe disease in TLR4 deficient mice. Studies have revealed that TLR 4-
signaling- induced IL-10 production inhibits inflammation and pathology in response
to B. pertussis infection (67). TLR4 is also vital for B. pertussis vaccine- mediated
protective immunity through the induction of Th1 and Th17 cells (66).
In vitro studies also demonstrate that B. bronchiseptica flagellin elicits
chemokine and cytokine production through TLR5 signaling (89). Despite these
convincing in vitro data, the in vivo role of TLR5 in B. bronchiseptica- host cell
interaction is unknown as flagellin expression is repressed on infection and ectopic
expression of flagellin has been shown to be inhibitory to Bordetella colonization (1).
Complement.
Complement system is an integral part of the host immune responses involved
in promoting clearance of pathogens like bacteria through opsonization –mediated
phagocytosis, killing, or augmentation of inflammation. Complement activation can
occur via three pathways: the antibody mediated classical pathway, antibody
independent alternate pathway and the mannose binding lectin pathway. Bordetella
pertussis expresses an autotransporter protein; BrkA (Bordetella resistance to killing)
23

that imparts resistance to the bacteoricidal activity of complement and provides
protection against antimicrobial peptides (43). Studies have demonstrated that the
disruption of brkA gene leads to enhanced deposition of C4 protein and subsequently
increasing susceptibility of B. pertussis to complement mediated killing. These
studies therefore, suggest that BrkA inhibits the classical pathway of complement
activation through preventing accumulation of C4 protein on bacterial surface (8). In
vitro serum killing assays suggest that B. pertussis is resistant to the alternate
pathway at lower naïve serum concentrations. In contrast, B. bronchiseptica is
resistant to killing by naïve serum, even at high concentrations, thereby suggesting
that this animal adapted species is resistant to the alternate pathway of complement
activation (57). C3 deficient mice are as efficient as wt mice in controlling Bordetella
infection as well as in the generation of infection-induced protective immunity.
However, studies have also demonstrated that C3, but not C5 and CR3 are vital for
vaccine induce immunity and antibody –mediated clearance of Bordetella from the
lower respiratory tract (82).
B cell responses.
There is definitive evidence primarily based on passive transfer experiments
that antibodies are critical for resolving Bordetella infections as well as for vaccine
mediated immune responses. Antibodies function through different mechanisms- 1)
Inhibition of adherence to prevent colonization, 2) neutralization and clearance of
toxins, 3) opsonization to enhance phagocytosis and 4) complement activation to
promote lysis (106). B cell deficient MuMT mice fail to clear Bordetella infection from
24

the trachea and lungs (81). Adoptive transfer of immune serum from mice infected
with B. bronchiseptica can rapidly clear infection of this organism from the lower
respiratory tract as early as 7 days post challenge. In contrast, B. pertussis can resist
clearance by passively transferred immune serum upto 7 days, after which it gets
cleared (80). Numerous studies investigating the role of individual virulence factors in
imparting protective immunity have revealed that passive immunization with
antibodies against FHA, Prn, Fim, PT and LPS can confer different levels of
protection (108). Moreover, studies have also shown a direct evidence for the critical
role of antibodies in humans by demonstrating that adoptive transfer of anti-B.
pertussis sera reduced the severity of disease in infected patients (51).
The efficiency of different antibodies directed against a range of virulence
factors to inhibit adherence has also been tested in vitro. Antibodies to FHA, Prn,
Fim, PT and LPS inhibit attachment of B. pertussis to human bronchial epithelial cells
(106). Also anti-FHA antibodies prevent attachment of B. pertussis to neutrophils,
while anti- CyaA sera promotes adherence and phagocytosis by neutrophils. Similarly
antibodies to Prn have also been shown to be crucial for phagocytosis of B. pertussis
by neutrophils. Natural infection as well as immunization with the whole cell vaccines
induces predominantly IgG2a antibodies, a subclass which is considered to be critical
for opsonization and complement fixation. IgG2a is also associated with greater
infection or vaccine induced protection. B. pertussis infection also induces the
mucosal, secretory isotype; IgA, both in mice and humans (106). IgA in convalescent
serum inhibits adherence of B. pertussis to ciliated epithelial cells in vitro and is
suggested to be critical for clearance of primary infection in vivo (106). Furthermore,
25

studies using IgA deficient mice also demonstrate that this isotype is important for
reducing B. bronchiseptica colonization from the upper respiratory tract (149).
T cell responses.
Direct evidence of the critical role for T cells in immunity is provided by the fact
that B. bronchiseptica and certain strains of B. pertussis cause lethal infection in
athymic as well as SCID mice (57). In sub-lethally irradiated mice, in which the T cells
are incapable of responding to antigens, B. pertussis causes a protracted infection.
Adoptive transfer of immune spleen cells from B. pertussis infected convalescent
mice imparted the recipient irradiated mice the ability to clear the infection within 2-3
weeks. Furthermore, studies investigating the contributions of individual
subpopulations of immune cells revealed that adoptive transfer of immune CD4+ T
cells, but not CD8+ T cells conferred the ability to clear B. pertussis infections in
irradiated mice (107). The vital role of CD4+ T cells in protective immunity is further
established by the study which demonstrated that intranasal immunization with
inactivated bacteria did not protect CD4 knock out mice against B. pertussis
challenge. In contrast, the same inactivated whole cell vaccine protected CD8 knock
out mice (84). Collectively these data suggest that CD4+ T cells, but not CD8+ T cells,
are important in both containing the primary infection as well as in vaccine mediated
protection.
Respiratory tract infection and vaccination with whole cell vaccines induce Th1
type responses with IFN-γ production. In contrast, the currently available acellular
vaccines generate Th2 cells that secrete IL-5, IL-4 and low levels of IFN-γ (66).
26

Addition of IL-12 to acellular vaccine led to priming of Th1 response and augmented
its protective efficacy (91). Consistent with these data, IFN-γ knockout and IL-12
deficient mice show greater bacterial load and delayed clearance while IL-10 knock-
out mice show faster clearance of infection (115). Apart from Th1 cells, recent in vitro
studies also demonstrated that B. bronchiseptica pulsed macrophages when
cocultured with CD4+ T cells, stimulate IL-17 production and preferentially induce
Th17 cells. Similarly immunization with B. pertussis whole cell vaccines, but not
acellular vaccines induced IL -17 producing T cells in addition to Th1 cells. Consistent
with these data, neutralization of IL-17 in vivo immediately prior to and after B.
pertussis challenge abrogated the protective efficacy of whole cell vaccines (66).
Currently Available Vaccines against B. pertussis.
Introduction of vaccines have decreased the incidence of whooping cough by
more than 80% as compared to the pre-vaccine era. The whole cell pertussis vaccine
(PW) containing killed whole B. pertussis organisms, was developed in the 1930s and
was combined with Diptheria (D), Tetanus (T) vaccines in 1940s and widely used in
clinical practice henceforth. PW is prepared by different methods, some of which
includes formalin fixing, heat killing, fractionation and extraction of B. pertussis (95).
Efficacy studies revealed that a primary series of four doses have 70-90%
effectiveness in preventing a serious pertussis illness. However, PW imparted
immunity has been suggested to wane over time with modest to no protection 5-10
years following the last dose (21). Furthermore, reactogenicity evaluations revealed
common local and systemic reactions in recipients of PW which was mainly attributed
27

to LPS. Studies have demonstrated that common local reactions are redness,
swelling and pain at the injection site and these occurred in 37-50% of DTP recipients
as compared to DT recipients. Systemic reactions to PW include drowsiness, fever,
vomiting and more serious events such as neurological diseases and death (5-7).
Although PW is still routinely used in many developing countries, at present this
vaccine is replaced by acellular pertussis vaccines in the United States due to the
adverse reactogenicity concerns.
The pediatric formulations of acellular vaccines (PC) were approved to be used
in United States in 1996. PC are subunit vaccines containing purified inactivated
components in varying concentrations. There are different combinations of PC
available that can range from monocomponent vaccines containing only PT to four
component vaccines consisting of PT, FHA, Fim and Prn. Although individual
antigens impart a certain level of protection, both murine model studies and clinical
trials demonstrate that multicomponent vaccines have higher estimated efficacy as
compared to monocomponent vaccines (108). PC in combination with Diphtheria and
Tetanus vaccines (DTaP) is administered in a series of four doses at 2 , 4 , 6 and 15-
18 months of age. A fifth dose is recommended for children who received the
primary four doses before the age of four, before joining school. Despite having high
effectiveness in clinical trials, this new generation PC does not pass the Kendrick test
and is considered to be less effective than PW. Due to increasing cases of pertussis
incidence in adults and adolescents, adult formulations of acellular vaccines were
approved by the CDC in 2005. Similar to PW, PC- imparted immunity is also
28

suggested to last for only 5-10 years following the last dose. Therefore, adult
formulations are theorized to booster immunity and lessen the incidence of pertussis.
Vaccines against B. bronchiseptica.
Currently available vaccines against B. bronchiseptica use live attenuated or
heat killed bacteria. Vaccines against kennel cough available in United States include
either individual heat killed or avirulent whole cells or in combination with other
vaccines such as live attenuated adenovirus type 2. Vaccines are administrated at 6-
8 and 10-12 weeks of age and subsequently annually. Similarly numerous vaccines
against atrophic rhinitis in pigs are available. These consist of inactivated or
nonpathogenic whole cells and are typically administered in combination with various
preparations of another important pathogen of swine, Pasterulla multocida. The
recommended schedule for vaccine administration in swine includes 1 dose prior to
delivery, shortly after birth and at 3-4 months of age (95). However, there are only
limited data on the safety and effectiveness of these vaccines. Although, heat killed
whole cell vaccines elicit high antibody responses, they do not induce as effective
protective immunity as infection induced immunity. Moreover, the basis for
attenuation of the commercially available live attenuated vaccines is unknown. As a
result, there is always the possibility of reversion of these strains back to the virulent
wild type form (136, 137). Therefore, current studies have been focusing on the use
of attenuated strains containing stable genetically defined mutations as vaccines.
These include strains with metabolic defects (aroA), as well as defects in production
29

of toxins such as DNT. A temperature sensitive urease strain defective in growth
above 340C has also been evaluated in a guinea pig model (130, 131). Furthermore,
the protective efficacy of a double mutant strain lacking adenylate cyclase toxin and
type III secretion system has been characterized using the mouse model system of
infection (92). However, the use of genetically defined attenuated strains still does not
resolve the possibility of reversion back into virulent form. The possibility of reversion
might be greater under competitive environments in the host such as co-infections
with other pathogenic organisms. Moreover, use of live attenuated strains for
vaccination purposes may increase the chances of zoonotic transmission especially
in immuno-compromised patients. Thus, development of acellular vaccines that can
provide protective immunity against B. bronchiseptica must be a priority.
A principal impediment towards development or improvement of vaccines for
B. pertussis and B. bronchiseptica is a gap in our understanding of the identity and
function of gene products that are critical for efficient respiratory tract infection. We
believe that efficient colonization and subsequent development of disease by
Bordetella require the interplay of multiple factors. The majority of current studies are
focused on the previously identified and characterized Bordetella factors which are
mentioned above and there is only limited focus on identification of novel antigens
that are vital for pathogenesis and with the potential to elicit protective immunity.
While several published data support the importance of the major virulence factors
namely, FHA, Fim, Prn CyaA, and PT in mediating attachment, colonization and
persistence, our unpublished data (Sukumar et al; in preparation) reveal that
simultaneous deletion of all these five factors does not abrogate upper respiratory
30

tract colonization by Bordetella. In fact, our observations provide evidence that
intranasal administration of this mutant strain results in protection from subsequent
challenge with wt B. bronchiseptica and B. pertussis in a mouse model of infection.
These studies highlight the contribution of hitherto unknown factors in Bordetella
infectious cycle. With the goal of identifying novel virulence factors and /or vaccine
candidates, we examined the genomic content of B. bronchiseptica. A computational
search was employed to identify unknown Open reading Frames (ORF) that have
potential BvgA binding motifs in the putative promoter regions. Our rationale for
searching ORFs which may be regulated by the BvgAS system was that the majority
of the previously characterized virulence factors are positively regulated by this
master regulatory system. Thus, our prediction was that, if BvgAS locus controls the
expression of a gene, then it may be important for Bordetella pathogenesis. Although
the search revealed multiple genes, we focused our study on an ORF that was
designated as a “putative adhesin” in the annotated B. bronchiseptica genome data
base and harbored homology to the previously characterized Bordetella protein BipA.
In Chapter II, we describe studies characterizing this ORF, which we designated as
bcfA. Using RT-PCR assays and EMSA, we confirm that the BvgAS system positively
regulates bcfA expression. Moreover, our studies indicate that, although homologous,
BipA and BcfA have differential phase specific expression patterns. We also
demonstrated that BcfA is localized to the outermembrane and that it is expressed in
vivo during Bordetella infection in rats. We also evaluated the role of these two
paralogous proteins in B. bronchiseptica colonization in the rat model of infection.
While individual deletions of BipA or BcfA did not abrogate B. bronchiseptica
31

colonization, a mutant strain deficient in both these proteins was defective in tracheal
colonization as compared to its wt parent strain. Based on this finding, we
hypothesize that BipA and BcfA have overlapping function in mediating tracheal
colonization by B. bronchiseptica.
In the subsequent Chapter III we explore the efficacy of BcfA as a vaccine
candidate in eliciting protective immune responses against B. bronchiseptica
infection. Our justifications for investigating the vaccine potential of BcfA were i) BcfA
is an outermembrane protein ii) BcfA-specific antibodies are produced during a wild
type B. bronchiseptica infection in rats. These results show that BcfA has the
potential to interact with the host immune system and induce at least the humoral
responses. Our studies reveal that both active and passive immunization with BcfA
provides protection against subsequent challenge with B. bronchiseptica in a mouse
model of infection. We also show that immunization with BcfA induces specific
antibodies in mice, with Ig2a being the predominant isotype. Data from this section
also demonstrate that neutrophils are critical for anti-BcfA antibody mediated
clearance of B. bronchiseptica from the lower respiratory tract. Finally we show that
immunization with BcfA induces a Th1 type response in splenocytes leading to IFN-γ
production. The utility of this antigen as a vaccine candidate is highlighted by the data
showing the conservation of expression of BcfA among multiple clinical isolates of B.
bronchiseptica.
32

REFERENCES 1. Akerley, B. J., P. A. Cotter, and J. F. Miller. 1995. Ectopic expression of
the flagellar regulon alters development of the Bordetella-host interaction. Cell 80:611-20.
2. Akerley, B. J., and J. F. Miller. 1993. Flagellin gene transcription in
Bordetella bronchiseptica is regulated by the BvgAS virulence control system. J Bacteriol 175:3468-79.
3. Akerley, B. J., D. M. Monack, S. Falkow, and J. F. Miller. 1992. The
bvgAS locus negatively controls motility and synthesis of flagella in Bordetella bronchiseptica. J Bacteriol 174:980-90.
4. Arico, B., and R. Rappuoli. 1987. Bordetella parapertussis and
Bordetella bronchiseptica contain transcriptionally silent pertussis toxin genes. J Bacteriol 169:2847-53.
5. Baraff, L. J., W. J. Ablon, and R. C. Weiss. 1983. Possible temporal
association between diphtheria-tetanus toxoid-pertussis vaccination and sudden infant death syndrome. Pediatr Infect Dis 2:7-11.
6. Baraff, L. J., C. R. Manclark, J. D. Cherry, P. Christenson, and S. M.
Marcy. 1989. Analyses of adverse reactions to diphtheria and tetanus toxoids and pertussis vaccine by vaccine lot, endotoxin content, pertussis vaccine potency and percentage of mouse weight gain. Pediatr Infect Dis J 8:502-7.
7. Baraff, L. J., W. D. Shields, L. Beckwith, G. Strome, S. M. Marcy, J. D.
Cherry, and C. R. Manclark. 1988. Infants and children with convulsions and hypotonic-hyporesponsive episodes following diphtheria-tetanus-pertussis immunization: follow-up evaluation. Pediatrics 81:789-94.
8. Barnes, M. G., and A. A. Weiss. 2001. BrkA protein of Bordetella
pertussis inhibits the classical pathway of complement after C1 deposition. Infect Immun 69:3067-72.
9. Beier, D., B. Schwarz, T. M. Fuchs, and R. Gross. 1995. In vivo
characterization of the unorthodox BvgS two-component sensor protein of Bordetella pertussis. J Mol Biol 248:596-610.
10. Bergfors, E., B. Trollfors, J. Taranger, T. Lagergard, V. Sundh, and G.
Zackrisson. 1999. Parapertussis and pertussis: differences and similarities in incidence, clinical course, and antibody responses. Int J Infect Dis 3:140-6.
33

11. Bordet, J. a. O. G. 1909. L'endotoxine coquelucheuse. Ann. Inst. Pasteur 23:415-419.
12. Bordet, J. a. O. G. 1906. Le microbe de la coqueluche. Ann. Inst. Pasteur
20:48-68. 13. Boschwitz, J. S., J. W. Batanghari, H. Kedem, and D. A. Relman. 1997.
Bordetella pertussis infection of human monocytes inhibits antigen-dependent CD4 T cell proliferation. J Infect Dis 176:678-86.
14. Boucher, P. E., and S. Stibitz. 1995. Synergistic binding of RNA
polymerase and BvgA phosphate to the pertussis toxin promoter of Bordetella pertussis. J Bacteriol 177:6486-91.
15. Bradford, P. G., and R. P. Rubin. 1985. Pertussis toxin inhibits
chemotactic factor-induced phospholipase C stimulation and lysosomal enzyme secretion in rabbit neutrophils. FEBS Lett 183:317-20.
16. Buonavoglia, C., and V. Martella. 2007. Canine respiratory viruses. Vet
Res 38:355-73. 17. Carbonetti, N. H. 2007. Immunomodulation in the pathogenesis of
Bordetella pertussis infection and disease. Curr Opin Pharmacol 7:272-8. 18. Carbonetti, N. H., G. V. Artamonova, C. Andreasen, E. Dudley, R. M.
Mays, and Z. E. Worthington. 2004. Suppression of serum antibody responses by pertussis toxin after respiratory tract colonization by Bordetella pertussis and identification of an immunodominant lipoprotein. Infect Immun 72:3350-8.
19. Cherry, J. D. 1999. Pertussis in the preantibiotic and prevaccine era, with
emphasis on adult pertussis. Clin Infect Dis 28 Suppl 2:S107-11. 20. Cherry, J. D., J. Gornbein, U. Heininger, and K. Stehr. 1998. A search
for serologic correlates of immunity to Bordetella pertussis cough illnesses. Vaccine 16:1901-6.
21. Cherry, J. D., and P. Olin. 1999. The science and fiction of pertussis
vaccines. Pediatrics 104:1381-3. 22. Cherry, J. D., D. X. Xing, P. Newland, K. Patel, U. Heininger, and M. J.
Corbel. 2004. Determination of serum antibody to Bordetella pertussis adenylate cyclase toxin in vaccinated and unvaccinated children and in children and adults with pertussis. Clin Infect Dis 38:502-7.
34

23. Confer, D. L., and J. W. Eaton. 1982. Phagocyte impotence caused by an invasive bacterial adenylate cyclase. Science 217:948-50.
24. Confer, D. L., A. S. Slungaard, E. Graf, S. S. Panter, and J. W. Eaton.
1984. Bordetella adenylate cyclase toxin: entry of bacterial adenylate cyclase into mammalian cells. Adv Cyclic Nucleotide Protein Phosphorylation Res 17:183-7.
25. Cookson, B. T., H. L. Cho, L. A. Herwaldt, and W. E. Goldman. 1989.
Biological activities and chemical composition of purified tracheal cytotoxin of Bordetella pertussis. Infect Immun 57:2223-9.
26. Cookson, B. T., A. N. Tyler, and W. E. Goldman. 1989. Primary
structure of the peptidoglycan-derived tracheal cytotoxin of Bordetella pertussis. Biochemistry 28:1744-9.
27. Corbel, M. J., and D. K. Xing. 1997. A consideration of control
requirements for acellular pertussis vaccines. Dev Biol Stand 89:343-7. 28. Cotter, P. A., and A. M. Jones. 2003. Phosphorelay control of virulence
gene expression in Bordetella. Trends Microbiol 11:367-73. 29. Cotter, P. A., and J. F. Miller. 1994. BvgAS-mediated signal transduction:
analysis of phase-locked regulatory mutants of Bordetella bronchiseptica in a rabbit model. Infect Immun 62:3381-90.
30. Cotter, P. A., and J. F. Miller. 1997. A mutation in the Bordetella
bronchiseptica bvgS gene results in reduced virulence and increased resistance to starvation, and identifies a new class of Bvg-regulated antigens. Mol Microbiol 24:671-85.
31. Cotter, P. A., M. H. Yuk, S. Mattoo, B. J. Akerley, J. Boschwitz, D. A.
Relman, and J. F. Miller. 1998. Filamentous hemagglutinin of Bordetella bronchiseptica is required for efficient establishment of tracheal colonization. Infect Immun 66:5921-9.
32. Coutte, L., R. Antoine, H. Drobecq, C. Locht, and F. Jacob-
Dubuisson. 2001. Subtilisin-like autotransporter serves as maturation protease in a bacterial secretion pathway. EMBO J 20:5040-8.
33. Coutte, L., E. Willery, R. Antoine, H. Drobecq, C. Locht, and F. Jacob-
Dubuisson. 2003. Surface anchoring of bacterial subtilisin important for maturation function. Mol Microbiol 49:529-39.
35

34. Cowell, J. L., E. L. Hewlett, and C. R. Manclark. 1979. Intracellular localization of the dermonecrotic toxin of Bordetella pertussis. Infect Immun 25:896-901.
35. Cummings, C. A., M. M. Brinig, P. W. Lepp, S. van de Pas, and D. A.
Relman. 2004. Bordetella species are distinguished by patterns of substantial gene loss and host adaptation. J Bacteriol 186:1484-92.
36. Dautin, N., and H. D. Bernstein. 2007. Protein secretion in gram-
negative bacteria via the autotransporter pathway. Annu Rev Microbiol 61:89-112.
37. De Serres, G., R. Shadmani, B. Duval, N. Boulianne, P. Dery, M.
Douville Fradet, L. Rochette, and S. A. Halperin. 2000. Morbidity of pertussis in adolescents and adults. J Infect Dis 182:174-9.
38. Deora, R., H. J. Bootsma, J. F. Miller, and P. A. Cotter. 2001. Diversity
in the Bordetella virulence regulon: transcriptional control of a Bvg-intermediate phase gene. Mol Microbiol 40:669-83.
39. Dworkin, M. S., P. S. Sullivan, S. E. Buskin, R. D. Harrington, J.
Olliffe, R. D. MacArthur, and C. E. Lopez. 1999. Bordetella bronchiseptica infection in human immunodeficiency virus-infected patients. Clin Infect Dis 28:1095-9.
40. Eldering, G., and P. Kendrick. 1938. Bacillus Para-Pertussis: A Species
Resembling Both Bacillus Pertussis and Bacillus Bronchisepticus but Identical with Neither. J Bacteriol 35:561-72.
41. Eldering, G., and P. L. Kendrick. 1952. Incidence of parapertussis in the
Grand Rapids area as indicated by 16 years' experience with diagnostic cultures. Am J Public Health Nations Health 42:27-31.
42. Emsley, P., G. McDermott, I. G. Charles, N. F. Fairweather, and N. W.
Isaacs. 1994. Crystallographic characterization of pertactin, a membrane-associated protein from Bordetella pertussis. J Mol Biol 235:772-3.
43. Fernandez, R. C., and A. A. Weiss. 1994. Cloning and sequencing of a
Bordetella pertussis serum resistance locus. Infect Immun 62:4727-38. 44. Ferry, N. S., and H. C. Klix. 1918. Studies Relative to the Apparent Close
Relationship between Bact. pertussis and B. bronchisepticus: II. Complement Fixation Tests. J Bacteriol 3:309-12.
45. Flak, T. A., and W. E. Goldman. 1999. Signalling and cellular specificity
of airway nitric oxide production in pertussis. Cell Microbiol 1:51-60.
36

46. Gerlach, G., F. von Wintzingerode, B. Middendorf, and R. Gross. 2001. Evolutionary trends in the genus Bordetella. Microbes Infect 3:61-72.
47. Gilberg, S., E. Njamkepo, I. P. Du Chatelet, H. Partouche, P. Gueirard,
C. Ghasarossian, M. Schlumberger, and N. Guiso. 2002. Evidence of Bordetella pertussis infection in adults presenting with persistent cough in a french area with very high whole-cell vaccine coverage. J Infect Dis 186:415-8.
48. Goldman, W. E., D. G. Klapper, and J. B. Baseman. 1982. Detection,
isolation, and analysis of a released Bordetella pertussis product toxic to cultured tracheal cells. Infect Immun 36:782-94.
49. Goodnow, R. A. 1980. Biology of Bordetella bronchiseptica. Microbiol
Rev 44:722-38. 50. Goodwin, M. S., and A. A. Weiss. 1990. Adenylate cyclase toxin is
critical for colonization and pertussis toxin is critical for lethal infection by Bordetella pertussis in infant mice. Infect Immun 58:3445-7.
51. Granstrom, M., A. M. Olinder-Nielsen, P. Holmblad, A. Mark, and K.
Hanngren. 1991. Specific immunoglobulin for treatment of whooping cough. Lancet 338:1230-3.
52. Greenberg, D. P., C. H. von Konig, and U. Heininger. 2005. Health
burden of pertussis in infants and children. Pediatr Infect Dis J 24:S39-43. 53. Gross, R., B. Arico, and R. Rappuoli. 1989. Families of bacterial signal-
transducing proteins. Mol Microbiol 3:1661-7. 54. Gueirard, P., and N. Guiso. 1993. Virulence of Bordetella bronchiseptica:
role of adenylate cyclase-hemolysin. Infect Immun 61:4072-8. 55. Hall, E., R. Parton, and A. C. Wardlaw. 1998. Responses to acellular
pertussis vaccines and component antigens in a coughing-rat model of pertussis. Vaccine 16:1595-603.
56. Hall, E., R. Parton, and A. C. Wardlaw. 1999. Time-course of infection
and responses in a coughing rat model of pertussis. J Med Microbiol 48:95-8.
57. Harvill, E. T., P. A. Cotter, and J. F. Miller. 1999. Pregenomic
comparative analysis between bordetella bronchiseptica RB50 and Bordetella pertussis tohama I in murine models of respiratory tract infection. Infect Immun 67:6109-18.
37

58. Hazenbos, W. L., C. A. Geuijen, B. M. van den Berg, F. R. Mooi, and R. van Furth. 1995. Bordetella pertussis fimbriae bind to human monocytes via the minor fimbrial subunit FimD. J Infect Dis 171:924-9.
59. Hazenbos, W. L., B. M. van den Berg, C. W. Geuijen, F. R. Mooi, and
R. van Furth. 1995. Binding of FimD on Bordetella pertussis to very late antigen-5 on monocytes activates complement receptor type 3 via protein tyrosine kinases. J Immunol 155:3972-8.
60. Heininger, U., K. Stehr, S. Schmitt-Grohe, C. Lorenz, R. Rost, P. D.
Christenson, M. Uberall, and J. D. Cherry. 1994. Clinical characteristics of illness caused by Bordetella parapertussis compared with illness caused by Bordetella pertussis. Pediatr Infect Dis J 13:306-9.
61. Heiss, L. N., T. A. Flak, J. R. Lancaster, Jr., M. L. McDaniel, and W. E.
Goldman. 1993. Nitric oxide mediates Bordetella pertussis tracheal cytotoxin damage to the respiratory epithelium. Infect Agents Dis 2:173-7.
62. Heiss, L. N., S. A. Moser, E. R. Unanue, and W. E. Goldman. 1993.
Interleukin-1 is linked to the respiratory epithelial cytopathology of pertussis. Infect Immun 61:3123-8.
63. Hellwig, S. M., M. E. Rodriguez, G. A. Berbers, J. G. van de Winkel,
and F. R. Mooi. 2003. Crucial role of antibodies to pertactin in Bordetella pertussis immunity. J Infect Dis 188:738-42.
64. Hewlett, E. L., and K. M. Edwards. 2005. Clinical practice. Pertussis--not
just for kids. N Engl J Med 352:1215-22. 65. Hewlett, E. L., V. M. Gordon, J. D. McCaffery, W. M. Sutherland, and
M. C. Gray. 1989. Adenylate cyclase toxin from Bordetella pertussis. Identification and purification of the holotoxin molecule. J Biol Chem 264:19379-84.
66. Higgins, S. C., A. G. Jarnicki, E. C. Lavelle, and K. H. Mills. 2006.
TLR4 mediates vaccine-induced protective cellular immunity to Bordetella pertussis: role of IL-17-producing T cells. J Immunol 177:7980-9.
67. Higgins, S. C., E. C. Lavelle, C. McCann, B. Keogh, E. McNeela, P.
Byrne, B. O'Gorman, A. Jarnicki, P. McGuirk, and K. H. Mills. 2003. Toll-like receptor 4-mediated innate IL-10 activates antigen-specific regulatory T cells and confers resistance to Bordetella pertussis by inhibiting inflammatory pathology. J Immunol 171:3119-27.
68. Hoppe, J. E. 1999. Update on respiratory infection caused by Bordetella parapertussis. Pediatr Infect Dis J 18:375-81.
38

69. Horiguchi, Y., T. Nakai, and K. Kume. 1991. Effects of Bordetella bronchiseptica dermonecrotic toxin on the structure and function of osteoblastic clone MC3T3-e1 cells. Infect Immun 59:1112-6.
70. Horiguchi, Y., N. Sugimoto, and M. Matsuda. 1993. Stimulation of DNA
synthesis in osteoblast-like MC3T3-E1 cells by Bordetella bronchiseptica dermonecrotic toxin. Infect Immun 61:3611-5.
71. Huebner, E. S., B. Christman, S. Dummer, Y. W. Tang, and S.
Goodman. 2006. Hospital-acquired Bordetella bronchiseptica infection following hematopoietic stem cell transplantation. J Clin Microbiol 44:2581-3.
72. Iida, T., and T. Okonogi. 1971. Lienotoxicity of Bordetella pertussis in
mice. J Med Microbiol 4:51-61. 73. Inatsuka, C. S., S. M. Julio, and P. A. Cotter. 2005. Bordetella
filamentous hemagglutinin plays a critical role in immunomodulation, suggesting a mechanism for host specificity. Proc Natl Acad Sci U S A 102:18578-83.
74. Ishibashi, Y., S. Claus, and D. A. Relman. 1994. Bordetella pertussis
filamentous hemagglutinin interacts with a leukocyte signal transduction complex and stimulates bacterial adherence to monocyte CR3 (CD11b/CD18). J Exp Med 180:1225-33.
75. Ishibashi, Y., D. A. Relman, and A. Nishikawa. 2001. Invasion of human
respiratory epithelial cells by Bordetella pertussis: possible role for a filamentous hemagglutinin Arg-Gly-Asp sequence and alpha5beta1 integrin. Microb Pathog 30:279-88.
76. Iwata, S., T. Aoyama, A. Goto, H. Iwai, Y. Sato, H. Akita, Y. Murase, T.
Oikawa, T. Iwata, S. Kusano, and et al. 1991. Mixed outbreak of Bordetella pertussis and Bordetella parapertussis in an apartment house. Dev Biol Stand 73:333-41.
77. Jacobs, C., B. Joris, M. Jamin, K. Klarsov, J. Van Beeumen, D.
Mengin-Lecreulx, J. van Heijenoort, J. T. Park, S. Normark, and J. M. Frere. 1995. AmpD, essential for both beta-lactamase regulation and cell wall recycling, is a novel cytosolic N-acetylmuramyl-L-alanine amidase. Mol Microbiol 15:553-9.
78. Kania, S. A., S. Rajeev, E. H. Burns, Jr., T. F. Odom, S. M. Holloway,
and D. A. Bemis. 2000. Characterization of fimN, a new Bordetella bronchiseptica major fimbrial subunit gene. Gene 256:149-55.
39

79. Katada, T., M. Tamura, and M. Ui. 1983. The A protomer of islet-activating protein, pertussis toxin, as an active peptide catalyzing ADP-ribosylation of a membrane protein. Arch Biochem Biophys 224:290-8.
80. Kirimanjeswara, G. S., L. M. Agosto, M. J. Kennett, O. N. Bjornstad,
and E. T. Harvill. 2005. Pertussis toxin inhibits neutrophil recruitment to delay antibody-mediated clearance of Bordetella pertussis. J Clin Invest 115:3594-601.
81. Kirimanjeswara, G. S., P. B. Mann, and E. T. Harvill. 2003. Role of
antibodies in immunity to Bordetella infections. Infect Immun 71:1719-24. 82. Kirimanjeswara, G. S., P. B. Mann, M. Pilione, M. J. Kennett, and E. T.
Harvill. 2005. The complex mechanism of antibody-mediated clearance of Bordetella from the lungs requires TLR4. J Immunol 175:7504-11.
83. Lacey, B. W. 1960. Antigenic modulation of Bordetella pertussis. J Hyg
(Lond) 58:57-93. 84. Leef, M., K. L. Elkins, J. Barbic, and R. D. Shahin. 2000. Protective
immunity to Bordetella pertussis requires both B cells and CD4(+) T cells for key functions other than specific antibody production. J Exp Med 191:1841-52.
85. Livey, I., C. J. Duggleby, and A. Robinson. 1987. Cloning and
nucleotide sequence analysis of the serotype 2 fimbrial subunit gene of Bordetella pertussis. Mol Microbiol 1:203-9.
86. Locht, C., and J. M. Keith. 1986. Pertussis toxin gene: nucleotide
sequence and genetic organization. Science 232:1258-64. 87. Long, S. S., H. W. Lischner, A. Deforest, and J. L. Clark. 1990.
Serologic evidence of subclinical pertussis in immunized children. Pediatr Infect Dis J 9:700-5.
88. Long, S. S., C. J. Welkon, and J. L. Clark. 1990. Widespread silent
transmission of pertussis in families: antibody correlates of infection and symptomatology. J Infect Dis 161:480-6.
89. Lopez-Boado, Y. S., L. M. Cobb, and R. Deora. 2005. Bordetella
bronchiseptica flagellin is a proinflammatory determinant for airway epithelial cells. Infect Immun 73:7525-34.
90. Magyar, T., N. Chanter, A. J. Lax, J. M. Rutter, and G. A. Hall. 1988.
The pathogenesis of turbinate atrophy in pigs caused by Bordetella bronchiseptica. Vet Microbiol 18:135-46.
40

91. Mahon, B. P., M. S. Ryan, F. Griffin, and K. H. Mills. 1996. Interleukin-12 is produced by macrophages in response to live or killed Bordetella pertussis and enhances the efficacy of an acellular pertussis vaccine by promoting induction of Th1 cells. Infect Immun 64:5295-301.
92. Mann, P., E. Goebel, J. Barbarich, M. Pilione, M. Kennett, and E.
Harvill. 2007. Use of a genetically defined double mutant strain of Bordetella bronchiseptica lacking adenylate cyclase and type III secretion as a live vaccine. Infect Immun 75:3665-72.
93. Mann, P. B., K. D. Elder, M. J. Kennett, and E. T. Harvill. 2004. Toll-like
receptor 4-dependent early elicited tumor necrosis factor alpha expression is critical for innate host defense against Bordetella bronchiseptica. Infect Immun 72:6650-8.
94. Martinez de Tejada, G., J. F. Miller, and P. A. Cotter. 1996.
Comparative analysis of the virulence control systems of Bordetella pertussis and Bordetella bronchiseptica. Mol Microbiol 22:895-908.
95. Mattoo, S., and J. D. Cherry. 2005. Molecular pathogenesis,
epidemiology, and clinical manifestations of respiratory infections due to Bordetella pertussis and other Bordetella subspecies. Clin Microbiol Rev 18:326-82.
96. Mattoo, S., J. F. Miller, and P. A. Cotter. 2000. Role of Bordetella
bronchiseptica fimbriae in tracheal colonization and development of a humoral immune response. Infect Immun 68:2024-33.
97. McGuirk, P., P. A. Johnson, E. J. Ryan, and K. H. Mills. 2000.
Filamentous hemagglutinin and pertussis toxin from Bordetella pertussis modulate immune responses to unrelated antigens. J Infect Dis 182:1286-9.
98. McGuirk, P., and K. H. Mills. 2000. Direct anti-inflammatory effect of a
bacterial virulence factor: IL-10-dependent suppression of IL-12 production by filamentous hemagglutinin from Bordetella pertussis. Eur J Immunol 30:415-22.
99. McMillan, D. J., E. Medina, C. A. Guzman, and M. J. Walker. 1999.
Expression of urease does not affect the ability of Bordetella bronchiseptica to colonise and persist in the murine respiratory tract. FEMS Microbiol Lett 178:7-11.
100. McMillan, D. J., M. Shojaei, G. S. Chhatwal, C. A. Guzman, and M. J.
Walker. 1996. Molecular analysis of the bvg-repressed urease of Bordetella bronchiseptica. Microb Pathog 21:379-94.
41

101. Meade, B. D., P. D. Kind, J. B. Ewell, P. P. McGrath, and C. R. Manclark. 1984. In vitro inhibition of murine macrophage migration by Bordetella pertussis lymphocytosis-promoting factor. Infect Immun 45:718-25.
102. Menozzi, F. D., C. Gantiez, and C. Locht. 1991. Interaction of the
Bordetella pertussis filamentous hemagglutinin with heparin. FEMS Microbiol Lett 62:59-64.
103. Merkel, T. J., P. E. Boucher, S. Stibitz, and V. K. Grippe. 2003. Analysis
of bvgR expression in Bordetella pertussis. J Bacteriol 185:6902-12. 104. Merkel, T. J., and S. Stibitz. 1995. Identification of a locus required for
the regulation of bvg-repressed genes in Bordetella pertussis. J Bacteriol 177:2727-36.
105. Merkel, T. J., S. Stibitz, J. M. Keith, M. Leef, and R. Shahin. 1998.
Contribution of regulation by the bvg locus to respiratory infection of mice by Bordetella pertussis. Infect Immun 66:4367-73.
106. Mills, K. H. 2001. Immunity to Bordetella pertussis. Microbes Infect 3:655-
77. 107. Mills, K. H., A. Barnard, J. Watkins, and K. Redhead. 1993. Cell-
mediated immunity to Bordetella pertussis: role of Th1 cells in bacterial clearance in a murine respiratory infection model. Infect Immun 61:399-410.
108. Mills, K. H., M. Ryan, E. Ryan, and B. P. Mahon. 1998. A murine model
in which protection correlates with pertussis vaccine efficacy in children reveals complementary roles for humoral and cell-mediated immunity in protection against Bordetella pertussis. Infect Immun 66:594-602.
109. Mink, C. M., J. D. Cherry, P. Christenson, K. Lewis, E. Pineda, D.
Shlian, J. A. Dawson, and D. A. Blumberg. 1992. A search for Bordetella pertussis infection in university students. Clin Infect Dis 14:464-71.
110. Mobberley-Schuman, P. S., B. Connelly, and A. A. Weiss. 2003.
Phagocytosis of Bordetella pertussis incubated with convalescent serum. J Infect Dis 187:1646-53.
111. Mooi, F. R., H. G. van der Heide, A. R. ter Avest, K. G. Welinder, I.
Livey, B. A. van der Zeijst, and W. Gaastra. 1987. Characterization of fimbrial subunits from Bordetella species. Microb Pathog 2:473-84.
42

112. Nicosia, A., M. Perugini, C. Franzini, M. C. Casagli, M. G. Borri, G. Antoni, M. Almoni, P. Neri, G. Ratti, and R. Rappuoli. 1986. Cloning and sequencing of the pertussis toxin genes: operon structure and gene duplication. Proc Natl Acad Sci U S A 83:4631-5.
113. Olin, P., F. Rasmussen, L. Gustafsson, H. O. Hallander, and H.
Heijbel. 1997. Randomised controlled trial of two-component, three-component, and five-component acellular pertussis vaccines compared with whole-cell pertussis vaccine. Ad Hoc Group for the Study of Pertussis Vaccines. Lancet 350:1569-77.
114. Parkhill, J., M. Sebaihia, A. Preston, L. D. Murphy, N. Thomson, D. E.
Harris, M. T. Holden, C. M. Churcher, S. D. Bentley, K. L. Mungall, A. M. Cerdeno-Tarraga, L. Temple, K. James, B. Harris, M. A. Quail, M. Achtman, R. Atkin, S. Baker, D. Basham, N. Bason, I. Cherevach, T. Chillingworth, M. Collins, A. Cronin, P. Davis, J. Doggett, T. Feltwell, A. Goble, N. Hamlin, H. Hauser, S. Holroyd, K. Jagels, S. Leather, S. Moule, H. Norberczak, S. O'Neil, D. Ormond, C. Price, E. Rabbinowitsch, S. Rutter, M. Sanders, D. Saunders, K. Seeger, S. Sharp, M. Simmonds, J. Skelton, R. Squares, S. Squares, K. Stevens, L. Unwin, S. Whitehead, B. G. Barrell, and D. J. Maskell. 2003. Comparative analysis of the genome sequences of Bordetella pertussis, Bordetella parapertussis and Bordetella bronchiseptica. Nat Genet 35:32-40.
115. Pilione, M. R., and E. T. Harvill. 2006. The Bordetella bronchiseptica
type III secretion system inhibits gamma interferon production that is required for efficient antibody-mediated bacterial clearance. Infect Immun 74:1043-9.
116. Porter, J. F., K. Connor, and W. Donachie. 1994. Isolation and
characterization of Bordetella parapertussis-like bacteria from ovine lungs. Microbiology 140 ( Pt 2):255-61.
117. Prasad, S. M., Y. Yin, E. Rodzinski, E. I. Tuomanen, and H. R. Masure.
1993. Identification of a carbohydrate recognition domain in filamentous hemagglutinin from Bordetella pertussis. Infect Immun 61:2780-5.
118. Rath, B. A., K. B. Register, J. Wall, D. M. Sokol, and R. B. Van Dyke.
2008. Persistent Bordetella bronchiseptica pneumonia in an immunocompetent infant and genetic comparison of clinical isolates with kennel cough vaccine strains. Clin Infect Dis 46:905-8.
119. Relman, D., E. Tuomanen, S. Falkow, D. T. Golenbock, K. Saukkonen,
and S. D. Wright. 1990. Recognition of a bacterial adhesion by an
43

integrin: macrophage CR3 (alpha M beta 2, CD11b/CD18) binds filamentous hemagglutinin of Bordetella pertussis. Cell 61:1375-82.
120. Rhodes, C. R., M. C. Gray, J. M. Watson, T. L. Muratore, S. B. Kim, E.
L. Hewlett, and C. M. Grisham. 2001. Structural consequences of divalent metal binding by the adenylyl cyclase toxin of Bordetella pertussis. Arch Biochem Biophys 395:169-76.
121. Riboli, B., P. Pedroni, A. Cuzzoni, G. Grandi, and F. de Ferra. 1991.
Expression of Bordetella pertussis fimbrial (fim) genes in Bordetella bronchiseptica: fimX is expressed at a low level and vir-regulated. Microb Pathog 10:393-403.
122. Roberts, M., N. F. Fairweather, E. Leininger, D. Pickard, E. L. Hewlett,
A. Robinson, C. Hayward, G. Dougan, and I. G. Charles. 1991. Construction and characterization of Bordetella pertussis mutants lacking the vir-regulated P.69 outer membrane protein. Mol Microbiol 5:1393-404.
123. Roop, R. M., 2nd, H. P. Veit, R. J. Sinsky, S. P. Veit, E. L. Hewlett, and
E. T. Kornegay. 1987. Virulence factors of Bordetella bronchiseptica associated with the production of infectious atrophic rhinitis and pneumonia in experimentally infected neonatal swine. Infect Immun 55:217-22.
124. Rose, T., P. Sebo, J. Bellalou, and D. Ladant. 1995. Interaction of calcium with Bordetella pertussis adenylate cyclase toxin. Characterization of multiple calcium-binding sites and calcium-induced conformational changes. J Biol Chem 270:26370-6.
125. Rosenthal, S., P. Strebel, P. Cassiday, G. Sanden, K. Brusuelas, and
M. Wharton. 1995. Pertussis infection among adults during the 1993 outbreak in Chicago. J Infect Dis 171:1650-2.
126. Rossi, C., J. Homand, C. Bauche, H. Hamdi, D. Ladant, and J.
Chopineau. 2003. Differential mechanisms for calcium-dependent protein/membrane association as evidenced from SPR-binding studies on supported biomimetic membranes. Biochemistry 42:15273-83.
127. Scarlato, V., A. Prugnola, B. Arico, and R. Rappuoli. 1990. Positive
transcriptional feedback at the bvg locus controls expression of virulence factors in Bordetella pertussis. Proc Natl Acad Sci U S A 87:10067.
128. Sebaihia, M., A. Preston, D. J. Maskell, H. Kuzmiak, T. D. Connell, N.
D. King, P. E. Orndorff, D. M. Miyamoto, N. R. Thomson, D. Harris, A. Goble, A. Lord, L. Murphy, M. A. Quail, S. Rutter, R. Squares, S. Squares, J. Woodward, J. Parkhill, and L. M. Temple. 2006. Comparison of the genome sequence of the poultry pathogen Bordetella
44

avium with those of B. bronchiseptica, B. pertussis, and B. parapertussis reveals extensive diversity in surface structures associated with host interaction. J Bacteriol 188:6002-15.
129. Sekura, R. D., F. Fish, C. R. Manclark, B. Meade, and Y. L. Zhang.
1983. Pertussis toxin. Affinity purification of a new ADP-ribosyltransferase. J Biol Chem 258:14647-51.
130. Shimizu, T. 1978. Prophylaxis of Bordetella bronchiseptica infection in
guinea pigs by intranasal vaccination with live strain ts-S34. Infect Immun 22:318-21.
131. Shimizu, T., and H. Ishikawa. 1982. Some characteristics of a urease-
negative, temperature- sensitive strain of Bordetella bronchiseptica as a live, attenuated vaccine. Infect Immun 36:198-201.
132. Skinner, J. A., A. Reissinger, H. Shen, and M. H. Yuk. 2004. Bordetella
type III secretion and adenylate cyclase toxin synergize to drive dendritic cells into a semimature state. J Immunol 173:1934-40.
133. Sloan, G. P., C. F. Love, N. Sukumar, M. Mishra, and R. Deora. 2007.
The Bordetella Bps polysaccharide is critical for biofilm development in the mouse respiratory tract. J Bacteriol 189:8270-6.
134. Spears, P. A., L. M. Temple, D. M. Miyamoto, D. J. Maskell, and P. E. Orndorff. 2003. Unexpected similarities between Bordetella avium and other pathogenic Bordetellae. Infect Immun 71:2591-7.
135. Spilker, T., A. A. Liwienski, and J. J. LiPuma. 2008. Identification of
Bordetella spp. in respiratory specimens from individuals with cystic fibrosis. Clin Microbiol Infect 14:504-6.
136. Stevenson, A., and M. Roberts. 2002. Use of a rationally attenuated
Bordetella bronchiseptica as a live mucosal vaccine and vector for heterologous antigens. Vaccine 20:2325-35.
137. Stevenson, A., and M. Roberts. 2003. Use of Bordetella bronchiseptica
and Bordetella pertussis as live vaccines and vectors for heterologous antigens. FEMS Immunol Med Microbiol 37:121-8.
138. Stockbauer, K. E., B. Fuchslocher, J. F. Miller, and P. A. Cotter. 2001.
Identification and characterization of BipA, a Bordetella Bvg-intermediate phase protein. Mol Microbiol 39:65-78.
139. Storsaeter, J., H. O. Hallander, L. Gustafsson, and P. Olin. 1998.
Levels of anti-pertussis antibodies related to protection after household exposure to Bordetella pertussis. Vaccine 16:1907-16.
45

140. Tuomanen, E., and A. Weiss. 1985. Characterization of two adhesins of Bordetella pertussis for human ciliated respiratory-epithelial cells. J Infect Dis 152:118-25.
141. Uhl, M. A., and J. F. Miller. 1994. Autophosphorylation and
phosphotransfer in the Bordetella pertussis BvgAS signal transduction cascade. Proc Natl Acad Sci U S A 91:1163-7.
142. van der Zee, A., F. Mooi, J. Van Embden, and J. Musser. 1997.
Molecular evolution and host adaptation of Bordetella spp.: phylogenetic analysis using multilocus enzyme electrophoresis and typing with three insertion sequences. J Bacteriol 179:6609-17.
143. Vandamme, P., M. Heyndrickx, M. Vancanneyt, B. Hoste, P. De Vos,
E. Falsen, K. Kersters, and K. H. Hinz. 1996. Bordetella trematum sp. nov., isolated from wounds and ear infections in humans, and reassessment of Alcaligenes denitrificans Ruger and Tan 1983. Int J Syst Bacteriol 46:849-58.
144. von Wintzingerode, F., G. Gerlach, B. Schneider, and R. Gross. 2002.
Phylogenetic relationships and virulence evolution in the genus Bordetella. Curr Top Microbiol Immunol 264:177-99.
145. von Wintzingerode, F., A. Schattke, R. A. Siddiqui, U. Rosick, U. B.
Gobel, and R. Gross. 2001. Bordetella petrii sp. nov., isolated from an anaerobic bioreactor, and emended description of the genus Bordetella. Int J Syst Evol Microbiol 51:1257-65.
146. Weingart, C. L., P. S. Mobberley-Schuman, E. L. Hewlett, M. C. Gray,
and A. A. Weiss. 2000. Neutralizing antibodies to adenylate cyclase toxin promote phagocytosis of Bordetella pertussis by human neutrophils. Infect Immun 68:7152-5.
147. Weingart, C. L., and A. A. Weiss. 2000. Bordetella pertussis virulence
factors affect phagocytosis by human neutrophils. Infect Immun 68:1735-9.
148. Weiss, A. A., and M. S. Goodwin. 1989. Lethal infection by Bordetella
pertussis mutants in the infant mouse model. Infect Immun 57:3757-64. 149. Wolfe, D. N., G. S. Kirimanjeswara, E. M. Goebel, and E. T. Harvill.
2007. Comparative role of immunoglobulin A in protective immunity against the Bordetellae. Infect Immun 75:4416-22.
46

150. Wolff, J., G. H. Cook, A. R. Goldhammer, and S. A. Berkowitz. 1980. Calmodulin activates prokaryotic adenylate cyclase. Proc Natl Acad Sci U S A 77:3841-4.
151. Woolfrey, B. F., and J. A. Moody. 1991. Human infections associated
with Bordetella bronchiseptica. Clin Microbiol Rev 4:243-55.
47

Chapter II
Differential Bvg-Phase Dependent Regulation and Role in
Pathogenesis of Two Bordetella Paralogs
Neelima Sukumar, Meenu Mishra, Gina Parise, Tomoo Ogi and Rajendar Deora
This chapter has been published in Journal of Bacteriology 2007 May;189(10):3695-
704 and is reprinted with permission. Experiments for Figure 9 were done by Dr.
Meenu Mishra. All animal work were performed with the help of Drs. Meenu Mishra
and Gina Parise-Sloan.
48

INTRODUCTION
Bordetellae are small aerobic, gram-negative coccobacilli that colonize the
respiratory tracts of humans and animals (26). Of the three classical species, B.
pertussis infects only humans and causes the acute respiratory disease known as
whooping cough (38). B. parapertussis strains can be divided into two genetically
distinct types, those which infect humans causing a pertussis-like illness, and those
which cause respiratory infections in sheep (26, 36). In contrast to the former two
species, B. bronchiseptica has a broad host range infecting a variety of nonhuman
animals (17, 26). It typically establishes asymptomatic infections but can cause
atrophic rhinitis in pigs, kennel cough in dogs, snuffles in rabbits and
bronchopneumonia in guinea pigs (17).
Efficient and productive colonization of the respiratory tract by Bordetella
requires interplay of multiple factors that allow bacterial adherence to the respiratory
epithelium leading to the eventual development of disease. The majority of these
virulence determinants are regulated by a two component regulatory system known
as BvgAS. BvgA is a DNA-binding response regulator and BvgS is a transmembrane
sensor protein kinase (26, 37). On perception of a signal, BvgS gets
autophosphorylated at a histidine residue (34). Phosphorylation of BvgA at a
conserved aspartate residue then occurs through a series of phosphotransfer
reactions from BvgS (5, 35). Phosphorylated BvgA (BvgA-P) can bind to the cognate
promoter regions of the Bvg-regulated genes and mediates activation and/or
repression of transcription (4, 29). A striking feature of the BvgAS regulatory circuitry
is its ability to control the transition among multiple phenotypic states of which at least
49

three phenotypic phases, the Bvg+, Bvg- and Bvgi (Bvg-intermediate) are known.
Each of these phases is characterized by the differential expression of known Bvg-
regulated gene products (9, 12). For example, during the Bvg+ phase a variety of
Bvg-activated factors including adhesins and toxins are maximally expressed and the
Bvg-repressed genes are minimally expressed (9, 12). For both B. pertussis and B.
bronchiseptica, it has been demonstrated that the Bvg+ phase is necessary and
sufficient for respiratory tract colonization (1, 10)
Transition to the Bvg- phase occurs as a result of either mutational inactivation
of BvgAS or growth in the presence of modulating signals (sulfate anion, nicotinic
acid or growth at low temperature). This phase is characterized by expression of the
Bvg-repressed factors (e. g. flagella in B. bronchiseptica and outermembrane
proteins of unknown function in B. pertussis) and the repression of Bvg-activated
genes (26). It has been suggested that this phase may be responsible for survival of
B. bronchiseptica in the environment (9). The Bvgi phase is expressed either as a
result of specific genetic mutations in BvgS or by growth of wild type (wt) Bordetella
strains in the presence of semi-modulating concentrations of chemical signals (9).
The Bvgi phase is principally distinguished by the maximal expression of a set of
antigens of which BipA (Bordetella intermediate phase protein A) is the first to be
identified at the molecular level (12, 33). The role of the Bvgi phase in Bordetella
infectious cycle is presently unclear.
BipA shares at its amino terminus similarity with intimin proteins of
enteropathogenic (EPEC) and enterohemorrhagic (EHEC) E. coli and with invasins of
Yersinia species, thereby leading to suggestions that BipA plays a role in the
50

Bordetella infectious cycle (33). To date a significant demonstrable role for BipA in
Bordetella pathogenesis has been elusive.
In this study, we report the identification of a B. bronchiseptica ORF termed
bcfA, which is a paralog of bipA. We demonstrate that the Bvg-phase dependent
expression profile of bcfA is strikingly different from that of bipA. In contrast to
maximal expression of bipA in the Bvgi phase, bcfA is expressed at high levels both
in the Bvg+ and the Bvgi phases. We have identified DNA sequences similar to the
consensus BvgA binding site in the region upstream of the bcfA ORF. Utilizing
electrophoretic mobility shift assay (EMSA), we observed differences in the nature of
DNA binding between BvgA and BvgA-P to the bcfA promoter region. Higher order
BvgA-DNA complexes were observed in the presence of acetyl phosphate. Our
results also document that BcfA is an outer membrane protein and that it is
expressed during Bordetella infection of rats. By comparing strains, harboring single
and double deletion mutations in bipA and bcfA, in intranasally infected rats, we show
that BipA and BcfA have an overlapping function in mediating efficient colonization of
trachea.
51

MATERIALS AND METHODS
Bacterial strains, plasmids, media and growth conditions. The bacterial strains
and plasmids used in this study are listed in Table 1. Bordetella strains were
maintained on Bordet Gengou (BG) agar (Becton Dickinson Microbiology Systems,
MD, USA) supplemented with 7.5% defibrinated sheep blood. For RNA extraction
and β-galactosidase assays, cells were grown in Stainer Scholte (SS) broth at 37°C
with shaking (32). E. coli strains were grown in Luria-Bertani medium at 37°C with
shaking. The different growth media were supplemented with appropriate antibiotics
(μg/ml) as needed, ampicillin, 100; chloramphenicol, 50; gentamycin sulfate 25;
streptomycin, 50; kanamycin, 25.
LacZ transcriptional fusions and β-galactosidase assays. For constructing lacZ
transcriptional fusion, a 640 bp DNA fragment consisting of 445 bp upstream and 195
bp downstream of the bcfA translational site was amplified from B. bronchiseptica
using BcfA5 and BcfA6 primers. The PCR fragment was cloned as a blunt ended
fragment into the EcoRV site of linearized pSTBlue-1 vector (Novagen, USA)
resulting in plasmid pRKD22. This plasmid was subsequently digested with EcoR1
and the resultant fragment was then cloned into EcoR1 site of the suicide plasmid
pEGZ (13) leading to the creation of the fusion plasmid pRKD23. This places the bcfA
promoter fragment upstream of a promoterless lacZ gene. pRKD23 was then
integrated into the genome of the different Bordetella strains by a single crossover at
the bcfA locus as described previously (12). β-galactosidase assays were performed
as previously described (11, 12). For detecting the β-galactosidase units under
52

TABLE 1. Strains and plasmids used in this study Strain/Plasmid Description ReferencesRB50 RB53 RB53i RB54 SM10λpir pSTBlue-1 pRKD22 pEGZ pRKD23 pRE112 pRKD40 pET24(a) pNS101 BL21(DE3)pLysE RKD101(ΔbcfA) RB25 (ΔbipA) MM101 (ΔbipAΔbcfA)
Wt B. bronchiseptica strain Bvg+ phase locked derivative Bvgi phase locked derivative Bvg- phase locked, ΔbvgS th-1 thr leu tonA lacY supE, recA::RP4–2-Tc::Mu Km
R (λ pir) Blunt end cloning vector; Novagen pSTBlue-1 derivative containing the 640 bp promoter region of bcfA cloned into the EcoRV site of pSTBlue1 lacZ transcriptional fusion vector pEGZ derivative; EcoRI fragment from pRKD22 cloned into EcoRI site of pEGZ upstream of promoter less lacZ Allelic exchange vector; Cmr
pRE112 derivative, bcfA deletion plasmid T7 based expression plasmid, Novagen pET24(a) derivative, BcfA overexpression plasmid Competent cells, RB50 derivative with in frame chromosomal deletion of bcfA RB50 derivative with in frame chromosomal deletion of bipA ΔbipA derivative with in frame chromosomal deletion of bcfA
(9) (9) (9) (9) (31)
This study (13) This study (14) This study This study This study (33) This study
53

conditions where the BvgAS system is modulated, the wt strain was grown in the
presence of 40 mM MgSO4.
RNA isolation and Real Time RT-PCR. Total RNA was isolated using the RNeasy
kit from Qiagen (MD, USA), treated with RNase free DNase-1 (Invitrogen, CA, USA)
to degrade contaminating DNA. 2-3 μg of RNA was primed with random hexamers
(Invitrogen, CA, USA) and cDNA was prepared utilizing the reverse transcriptase
enzyme (RT) Superscript III (Invitrogen, CA, USA) following manufacturer’s protocol.
Specific primers (Table 2) for various genes were designed using the ABI PRISM
Primer Express software (PE Applied Biosystems) in order to obtain similar-sized
amplicons. Diluted (1:25) reverse-transcription products obtained as above and a
blank control without cDNA were used as template and amplified using the TaqMan
Universal PCR Master mix (Applied Biosystems). The Master mix contains dNTPs
with dUTP, AmpliTaq Gold DNA polymerase, Amperase UNG, optimized buffer and a
passive reference dye. For each PCR, a mixture (20 μl) containing template cDNA, 1x
Master Mix, 250 nM of each sense and antisense primer and 500 nM of the TaqMan
probe was placed in 96-well optically clear PCR plates (Greiner Bio-one, USA).
Amplification and detection of the specific products were performed on an ABI PRISM
7000 Sequence Detection System (PE Biosystems) with the following cycle protocol:
one cycle at 50°C for 2 min and one cycle at 95°C for 10 min followed by 40 cycles at
95°C for 15 s and 60°C for 1 min. The threshold value was set manually and kept
constant for all comparison groups. To control for variation in RNA quantity and
quality, the constitutively expressed Bordetella gene recA was used as the
54

endogenous reference control. The relative quantification of gene expression was
performed using the comparative CT (threshold cycle) method according to the
manufacturer's instructions (User Bulletin # 2: ABI PRISM Sequence Detection
System). The critical threshold cycle (CT) was defined as the cycle at which the
fluorescence became detectable above the background fluorescence, and was
inversely proportional to the logarithm of the initial number of template molecules. The
ΔCT value was determined by subtracting recA CT value from the gene-specific CT
values. The ΔΔ CT was calculated by subtracting the obtained ΔCT value with the
ΔCT-calibrator value. For measurement of the relative levels of Bvg-activated genes
bcfA, fhaB (encodes filamentous hemagglutinin), bipA and cyaA (encodes adenylate
cyclase toxin), the ΔCT values obtained from the Bvg- phase locked cells was
designated as the calibrator, since in this strain there is lowest expression of these
target genes. Similarly, for measurement of the relative levels of the Bvg-repressed
gene flaA (codes for flagellin), the ΔCT values obtained from the Bvg+ phase locked
cells was designated as the calibrator, since there is lowest expression of flaA in this
strain. We also performed a validation experiment to ensure that the efficiencies of
the target amplification and that of the reference amplification were similar by
determining the ΔCT values with template dilution. Data from at least three
measurements each carried out on at least two different batches of RNA were
plotted. Error bars represent standard deviation from independent values.
Time point analysis. To modulate bvg-activity, the wt B. bronchiseptica strain RB50
was grown in 50 ml of SS medium in the presence of 40 mM MgSO4 for 18 h. At time
55

TABLE 2. Oligonucleotide primers used in this study a,b
Primer Sequence/Reference BcfA1 BcfA2 BcfA3 BcfA4 BcfA5 BcfA6 BcfA7 BcfA8 BcfA9 BcfA10 BcfA11 BcfA12 BcfA13 FhaF FhaR BipAF BipAR PrnF PrnR RecF RecR RecA9 RecA10 RecA11 Fha9 Fha10 Fha11 Fla9 Fla10 Fla11 BipA109 BipA110 BipA111
5’-CTAGTCTAGACCTACATATCCGTAGGATTG -3’ 5’-TTGGCGCGCCGTCTGCGCGACCCGCAGCAT -3’ 5’-TTGGCGCGCCTGATCGGTGGCGAGGGCGGC-3’ 5’-CGGGGTACCCTTGAACAGCGGCAGCACGTCG -3’ 5’-CGCATGGGATTCTCCCGGGTA-3’ 5’-GCTGTCGGCGGCATCCTGGCGATC -3’ 5’-GGCGAGCGCGACCGCGTCTTGCT-3’ 5’-GGCTGAGATTGACGCCAAGCTGCA-3’ 5’-CGCCATGCCTTGAA-3’ 5’-[DFAM]-GGGCGCCAACGCATT-[DTAM]-3’ 5’-TGGTTCTTGCCGAA-3’ 5’-CGCGGATCCGTGAAGCAAGCCATCCACG-3’ 5’-CCCAAGCTTCCCAGCAGGCCGCCCTC-3 5’-ATCCGACCTACACCGAATGG-3’5’-GAGTGTGCGCCGATTTTCAG-3’5’-GGGTCTGCCCTTTCTGCGCAATCTG-3’5’-GCCGATCACCTTGGTCTGCTCCAG-3’5’-GCCGCTGCAGCCGGAAGACCTTC-3’ 5’-GCAGCTCCGTCGCGACGATGTCG-3’ (13) (13) 5’-ACGTGCAATACGCCTCCAA-3’ 5’- [DFAM]-TGGGCGTCAACCTGACCGACCT-[DTAM]-3’ 5’-TGTCCGGCTGGGAGATCA-3’ 5’-TGTCCGCCATGGAGTATTTCA-3’ 5’-[DFAM]-CCCGGTGAGCCTGACAGCCCT-[DTAM]-3’ 5’-CCAGCAGATAATCCAGGAGTTCAT-3’ 5’-GATCCAGCAGGAAGTCAACCA-3’ 5’- [DFAM]-AAATCAACCGCATCGCCGAGCA-[DTAM]-3’ 5’-GACCTGATGCCGTTGAAGTC-3’ 5’-GGCCCAGGTCAATGATGTCTT-3’ 5’-[DFAM]-AACCTGGCTCGGGAATCGGGTC-[DTAM]-3’ 5’-CCTTGCAGATTGCGCAGA-3’
a Sequences in bold represent restriction enzyme sites b [DFAM]- 6 carboxyfluorescein: Reporter fluorochrome. b [DTAM]- 6-carboxy-tetramethyl-rhodamine: Quencher fluorochrome.
56

zero, the culture was spun down and re-suspended in 100 ml of SS media lacking
MgSO4. At times 5 min, 30 min, 1 h and 4 h, 5 ml of the bacterial culture was utilized
for total RNA preparation as described above.
After reverse transcription of the RNA, an aliquot of cDNA (5%) was used as
template in subsequent RT-PCR. In order to eliminate possible interference by
genomic DNA, mock reactions without RT were also performed. RT-PCR was carried
out as previously described (11, 12) with gene-specific primers. Primer pairs used are
listed in Table 2. Genomic DNA, prepared from the wt strain RB50, was used as the
positive control for PCR. Aliquots of amplified products obtained, were
electrophoresed on 1% agarose gels. Images of the ethidium-bromide-stained gels
were captured by Alpha Innotech Gel Doc System (Alpha Innotech Corporation).
Electrophoretic mobility shift assays. The purified DNA fragment was end labeled
by T4 polynucleotide kinase (New England Biolabs, MA) with [γ-32P]ATP (Amersham
Biosciences, NJ). Unincorporated radioactivity was removed by passage through G-
50 quick spin columns (Amersham Biosciences, NJ). Each reaction (20 μl), contained
indicated amounts of purified BvgA or BvgA-P and the radiolabeled promoter DNA in
1x binding buffer (10 mM Tris-HCl (pH 7.8), 2 mM MgCl2 50 mM NaCl, 1 mM DTT,
0.5 μg of poly (dI-dC), 0.01% NP-40, 100 ng of BSA and 10% glycerol).
Phosphorylation of BvgA was carried out as described earlier by incubation of the
protein at room temperature for 15 min in 1x binding buffer containing 20 mM acetyl
phosphate (11, 29). The reaction mixtures were incubated at 37oC for 15 min to allow
57

binding of BvgA/BvgA-P to radiolabeled promoter. The samples were
electrophoresed and visualized by autoradiography as previously described (11, 29).
For non-specific competition, a DNA fragment corresponding to the internal
region (+453 to +978) of bcfA ORF was amplified using primers BcfA7 and BcfA8.
Indicated concentrations of this fragment were included in the binding reactions. For
specific competition, bcfA promoter fragment was amplified using primers BcfA5 and
BcfA6 and indicated concentrations of the unlabeled DNA fragment was included in
the reaction.
Overexpression, purification of BcfA and antibody production. The entire bcfA
ORF excluding the stop codon was cloned as a BamHI-HindIII fragment in similarly
digested pET24(a) plasmid (Novagen), thereby resulting in the BcfA overexpression
plasmid pNS101. Amplification was carried out using recombinant Pfu DNA
polymerase (Stratagene, CA). E. coli BL21(DE3)pLysE cells containing pNS101
were grown in 2XTY (16 g of Bacto tryptone, 10 g of yeast extract, and 5 g/liter NaCl
plus 0.4% glucose) in the presence of kanamycin and chloramphenicol at 37 °C. The
cells were grown to an O.D.600 of 0.8-1.0 and induced by IPTG for 2 h. The cells
were harvested by centrifugation, resuspended in 30 ml of TGED buffer (10 mM
Tris-HCl (pH 7.9), 0.1 mM EDTA (pH 8.0), 0.2 mM DTT, and 0.05% sodium
deoxycholate, 5 % glycerol and 2 mM PMSF), incubated on ice for 20 min, and lysed
by passing through a French pressure cell three times at 14,000-16,000 p.s.i. The
lysate was centrifuged and the soluble BcfA was purified from the clarified lysate
58

using the T7. Tag affinity purification kit (Novagen) according to the manufacturer’s
protocol.
For antibody production, the cells after induction, were resuspended in the
TGED buffer, lysed by passing through French pressure cell and centrifuged. The
pellet was washed once with TGED buffer containing 0.5M NaCl, 10 ml of 1x
Bugbuster protein extraction reagent (Novagen), 10 μl of Benzonase nuclease and 1
mg/ml of lysozyme. After centrifugation, the cell pellet was resuspended in 5 ml of
TGED buffer containing 6M guanidine hydrochloride and incubated on ice for 10
min. Another 5 ml of cold TGED buffer was added and the suspension was
incubated on ice for an additional 10 min. The lysate was spun down and the
supernatant was dialyzed in 2 l of cold TGED buffer at 4oC for 18 h. The dialyzate
was centrifuged and the pellet containing overproduced BcfA was resuspended in
protein loading buffer (50 mM Tris-HCl (pH 6.8), 2 % SDS, 10 % glycerol and 5 mM
DTT) and was electrophoresed on a standard SDS polyacrylamide protein gel. The
band corresponding to the BcfA protein was excised and utilized for production of
anti-BcfA antibodies in rats. Antibody production was carried out on a fee for service
basis by Covance.
Preparation of Cellular Fractions. Stationary phase cultures of the different strains
were centrifuged at 17,000 rpm, and the cell pellets were resuspended in the cell
disruption buffer (10mM Tris-HCl (pH 8.0), 20% sucrose, 1 mM EDTA and 0.1 mg/mL
lysozyme). After incubation on ice for 10 min, the samples were frozen in dry ice
followed by thawing in cold water. The bacterial cells were sonicated on ice,
59

centrifuged initially at 3,700 rpm for 10 min to pellet unlysed cells. The clarified
suspension obtained was spun at 17,000 rpm for 1 h and the pellet resuspended in
an appropriate volume of 1x PBS. For separation of inner and outer membrane
proteins, the membrane fractions were incubated with 2% Triton X-100 for 30 min on
ice followed by centrifugation at 17,000 rpm for 1 h. The pellet (Triton X-100-
insoluble) consisting of the outer membrane proteins was utilized for SDS-PAGE
analysis.
SDS-PAGE and immunoblot analysis. Proteins from outer membrane fractions or
purified recombinant BcfA were separated on SDS-polyacrylamide gels, transferred
to nitrocellulose membranes and probed with anti-BcfA (1:5,000) or anti-BipA
antibody (1:5,000 dilution) (33) or rat serum (1:1,000 dilution) harvested from rats
infected with the indicated strains 30 days post-inoculation. The secondary antibody
(1:2,000) used, was either the goat anti-rat IgG or the goat anti rabbit IgG conjugated
to horseradish peroxidase. For detection of proteins, the Amersham ECL Western
blotting system was utilized.
Construction of deletion strains. Based on the pre-annotated sequence
information of the wt B. bronchiseptica strain RB50 present in the Sanger Center
database, we had designed primers to delete a region encompassing amino-acids
10-882 of the then 903 amino acid long bcfA ORF. Subsequent to the construction of
the deletion strains and the testing of these strains in animal models, we found that in
the updated database, the bcfA ORF was extended to include an upstream in frame
60

stretch of nucleotides encoding 66 amino acids. Thus, based on the current
annotation, the in-frame deletion of bcfA encompasses a region corresponding to
amino-acids 76-958 of the 969 amino acid long ORF.
A XbaI-AscI fragment (407 bp) containing sequences corresponding to the 5’
end including the first 75 amino acids of the bcfA ORF was amplified from the
chromosome of RB50 using primers BcfA1 and BcfA2. A second AscI-KpnI fragment
(525 bp) containing sequences corresponding to the 3’ end of bcfA including the last
11 amino-acids of the bcfA ORF was also amplified using primers BcfA3 and BcfA4.
These fragments were digested with respective restriction enzymes and were used
for three way ligation with Xba1 and Kpn1 digested suicide vector pRE112 (Cmr) (14)
resulting in plasmid pRKD40. This plasmid was transformed into SM10λpir and
mobilized from this strain into RB50. After conjugation, cointegrants were selected on
BG agar containing chloramphenicol and streptomycin. Colonies arising from second
recombination events were selected on LB-agar containing 7.5% sucrose as
described previously (11, 14). The genotype of the deletion strain RKD110 (ΔbcfA)
was confirmed by PCR and subsequent DNA sequencing.
MM101, the ΔbipAΔbcfA strain was constructed essentially as described
above for the ΔbcfA strain except that the parental strain used was the previously
described ΔbipA strain, RB25 (33).
Rat colonization Experiments. Four-five week old female Wistar rats (Charles
River Laboratories) were lightly anesthetized with halothane and intranasally
inoculated with 5 μl of sterile PBS alone or 40-100 CFUs of either the wt or the
61

various mutant strains. The number of delivered CFUs was confirmed by plating on
BG agar containing streptomycin (50μg/ml). Colonization levels in the respiratory
tract were determined by sacrificing rats 12 or 30 days post inoculation and
removing the entire nasal septum and 1cm of trachea. These were then
homogenized in 200 μL of sterile PBS and various dilutions of the homogenate were
plated on BG agar containing streptomycin to determine the number of CFUs.
62

RESULTS
Identification of BcfA, a parolog of BipA in B. bronchiseptica. It was previously
reported that a BipA-deficient strain of B. bronchiseptica colonized the rabbit
respiratory tract as efficiently as the wt strain, suggesting that this protein is not
essential for colonization (33). Since Bordetellae express multiple proteins that have
either been demonstrated or been predicted to be involved in attachment to the
respiratory epithelium (26), we hypothesized that the absence of BipA in B.
bronchiseptica could be compensated by either a known or yet to be identified
alternative factor, thereby leading to no apparent effect on colonization. In particular,
we explored the possibility of a paralogous protein in B. bronchiseptica. We searched
the recently sequenced B. bronchiseptica genome database for ORFs homologous to
BipA. BLASTP (2) searches revealed the presence of an ORF (BB0110) that
displayed 49% identity in N-terminal 809 amino acids to the BipA protein. In addition,
these searches also revealed similarity of BB0110 to intimins from EPEC and EHEC
(8, 15), invasins from Yersinia species (21) and the putative E. coli adhesin EaeH
(accession number AAZ57201). In the annotated B. bronchiseptica genome
database, the protein encoded by BB0110 has been designated as a putative
adhesin and based on our finding that it plays a role in respiratory tract colonization
(see below), we have designated this ORF as bcfA, Bordetella colonization factor A.
The B. bronchiseptica BcfA protein. The bcfA ORF has the potential to code for a
969 amino acid protein with a predicted molecular weight of 102 kDa (Fig. 1). The
63

FIG. 1. Schematic representation of the similarity of BcfA with BipA. Alignment of
amino acid sequences of BipA and BcfA was performed using the Needleman-
Wunsch global alignment algorithm by utilizing EMBOSS Needle available at
European Bioinformatics institute website. The regions of homology between these
two proteins are indicated by the dashed regions. The putative signal sequences are
represented by the shaded region. The black rectangle represents the region of BipA
and BcfA that displays similarity to intimins and invasins.
64

969
1578
BcfA
1
461
491 671 1158
BipA
A B
1
65

BipA protein of B. bronchiseptica is comparatively a larger protein having 1578 amino
acids and a molecular weight of 164.5 kDa (Fig. 1). Analysis of the predicted BcfA
protein sequence by SignalP 3.0 method (3) revealed that it contains an unusually
long signal sequence of 44 amino acids (signal peptide probability of 0.942) (Fig. 1),
including the characteristic positively charged N-region, the hydrophobic core H-
region and the C-region with a consensus cleavage site of a sec-dependent leader
peptide. The predicted cleavage site of the signal sequence is located between Ala44
and Gln45 residues (cleavage site probability of 0.928). The presence of a canonical
signal sequence in BcfA suggests that this protein can traverse the inner membrane.
In contrast to BcfA, the BipA protein lacks a canonical signal sequence.
Alignment of the amino acid sequences of BcfA and BipA revealed two main
regions of homology (Fig. 1). The N-terminal region of BcfA (amino acid residues 11-
461) is similar to N-terminal region of BipA (residues 29-491). The N-terminal regions
of BcfA and BipA are also similar to intimins from enteropathogenic and
enterohemorrhagic E. coli and invasins from Yersinia spp (Fig. 1). This region is
predicted to anchor these proteins in the outer membrane (25). The second region of
significant similarity between BipA and BcfA extends from amino acid residues 473-
969 of BcfA and 671-1158 of BipA (Fig. 1). The BipA protein possesses a 291 amino
acid long C-terminal surface-exposed domain and it appears that this domain is
lacking in BcfA (33).
Expression of bcfA is regulated by BvgAS in B. bronchiseptica. The BvgAS
locus regulates the expression of the majority of virulence gene expression in
66

Bordetella (26). The homology of bcfA with bipA, which is transcriptionally activated
by BvgAS (12, 33), prompted us to determine whether BvgAS controlled bcfA
expression. We cloned a DNA fragment, encompassing regions both upstream and
downstream of the predicted bcfA start codon, upstream of the promoter-less lacZ
gene contained in the suicide plasmid pEGZ (13). The resultant bcfA-lacZ fusion was
integrated into different strains of B. bronchiseptica by homologous recombination.
Measurement of the β-galactosidase activities from log-phase cultures of these
strains indicated that while bcfA is expressed at high levels in the wt strain, it is
expressed at low levels in the Bvg- phase locked strain (Fig. 2). Growth of the wt
strain in the presence of MgSO4, a known modulator of BvgAS activity, resulted in
very low levels of β-galactosidase activity (Fig. 2). Taken together these results
suggest that expression of bcfA is positively regulated by BvgAS.
bcfA and bipA demonstrate distinct Bvg-phase-dependent expression profile.
Previously, we and others have shown that the various Bvg-regulated genes exhibit
differences in their phase-dependent expression profiles (9, 12). Specifically, we have
demonstrated that in contrast to other known Bvg-regulated genes, bipA is expressed
maximally in the Bvgi phase and at low levels in the Bvg+ phase (12). To compare the
phase-dependent expression profile of bcfA directly with that of bipA and other Bvg-
regulated genes, we performed real time RT-PCR assays. We isolated total RNA
from Bvg+, Bvgi and Bvg- phase locked strains and generated cDNA through reverse
transcription. Primers and probes were designed to anneal to internal regions of
67

FIG. 2. Quantification of bcfA transcriptional activity by β-galactosidase assays in the
wt, Bvg+, Bvgi and Bvg- phase locked strains of B. bronchiseptica. The bcfA-lacZ
fusion was integrated in the various strains and the β-galactosidase activity was
determined after growth to mid-log phase in SS broth at 37°C. For modulating the
BvgAS activity, wt strain carrying bcfA-lacZ fusion was grown in the presence of 40
mM MgSO4. Error bars represent standard deviation.
68

0
200
600
1000
1400
wt Bvg+ Bvg
iBvg
-+MgSO4
β-ga
lact
osid
ase
units
Strains
69

FIG. 3. Determination of the phase-dependent expression profiles of different Bvg-
regulated genes in B. bronchiseptica by real time RT-PCR analysis. bcfA (A); bipA
(B); fhaB (C); cyaA (D) and flaA (E). cDNA prepared from various strains was used
as template for real time RT-PCR as described in the Materials and Methods. The
ΔCT value was determined by subtracting recA CT value from the gene-specific CT
values. ΔΔCT values for each gene was obtained by subtracting the ΔCT value of
either the Bvg- phase locked strain (for bcfA, bipA, fhaB, cyaA) or the Bvg+ phase
locked strain (for flaA) from the gene-specific ΔCT value. The relative expression
levels of respective genes (y-axes) were then calculated by the formula 2-ΔΔCT. Thus,
for bcfA, bipA, fhaB and cyaA the relative level of expression in the Bvg- phase locked
strain is 20=1. Similarly, the expression of flaA in Bvg+ phase locked strain is also 1.
Bars indicate standard deviation of three independent values obtained from two
different RNA batches. Y axes represent the relative expression levels of the
indicated gene.
70

cya
02468
10 12 14 16 18
0123
4567
bipA
B
0102030405060708090
bcfA
A
fha
02040
6080
100120
140C
D
Bvg+
Bvgi
Bvg-
Rel
ativ
e ex
pres
sion
Rel
ativ
e ex
pres
sion
Rel
ativ
e ex
pres
sion
0
2
4
6
8
10
12 fla
ER
elat
ive
expr
essi
on
si
onve
es
expr
Rel
ati
71

respective genes (Materials and Methods and Table 2). Expression levels of recA, a
Bvg-independent gene, were used as normalization controls. As shown in Fig. 3A,
bcfA was expressed at high levels in both the Bvg+ and the Bvgi phases. bcfA specific
RNA was barely detectable from Bvg- phase locked samples thereby confirming the
results of the β-galactosidase assays above. Strikingly, these results demonstrate
that the two paralogs bcfA and bipA have differential Bvg-dependent expression
profiles, since bipA is maximally expressed in the Bvgi phase and at a low level in the
Bvg+ phase (Fig. 3B and (11, 12)). Our data further suggest that the expression
profile of bcfA resembles that of the previously known Bvg-activated gene fhaB (6, 9).
As shown in Fig. 3C and consistent with previously published data, fhaB is expressed
at a high level in both the Bvg+ and Bvgi phases (9). As expected from previous
results (9, 12), cyaA and flaA were expressed maximally only in the Bvg+ and the
Bvg- phases (Figs. 3D and Fig. 3E), respectively, thereby further corroborating the
validity of the real time RT-PCR assays in accurately measuring the expression
patterns of Bvg-regulated genes.
We independently confirmed the phase-dependent expression pattern of bcfA
by measuring the β-galactosidase activities from log-phase cultures of the Bvg+ and
the Bvgi phase locked strains. Our results (Fig. 2) indicate that bcfA is expressed at
high levels in both the Bvg+ and Bvgi phases.
Kinetics of transcriptional activation of bipA and bcfA. Previous studies have
revealed the differential kinetics of expression of bvg-regulated genes upon induction
of the BvgAS activity either due to a temperature-shift (25°C to 37°C) or by first
72

growing in the presence of chemical modulators followed by growth in their absence
(22, 24). We compared the transcriptional activation of bcfA in B. bronchiseptica over
time following the induction of the BvgAS system. RB50 cells were modulated first by
growing them in the presence of 40 mM MgSO4 for 18h. At time zero, cells were spun
down and re-suspended in media without MgSO4 for the induction of the BvgAS
system. After 0, 5, 30, 60 and 240 min. of induction, total RNA was isolated from cells
and kinetics of promoter activation was determined by RT-PCR using primers specific
for fha, prn, bipA and bcfA. We also utilized primers specific for a bvg-independent
gene recA, as a normalization standard. RT-PCR assays revealed that of all the bvg-
regulated genes tested, only fhaB was transcribed at very low levels, suggesting very
efficient modulation of the BvgAS activity by overnight growth in presence of MgS04.
It was also not possible previously, using a slightly different protocol to completely
inhibit the expression of fhaB. As shown in Fig. 4, fhaB and bipA were reproducibly
activated as early as 5 min after induction and prn was transcribed after 30 min of
induction (Fig. 4). In accordance with our results, it has been shown previously that
the expression of fhaB and bipA was activated almost immediately after the switch to
inducing conditions, whereas there was a delay in the expression of prn (22, 24). Our
results clearly demonstrate that transcriptional activation of bcfA was reproducibly
observed within 5 min of growth under inducing conditions, suggesting that its
activation kinetics parallels to that of fhaB and bipA (Fig. 4). Control reactions were
performed to confirm the specificity and the reliability of the RT-PCR assay. No
detectable PCR products were obtained in the absence of RT indicating that
contaminating DNA was absent from the RNA preparation (data not shown).
73

FIG.4. Kinetics of transcriptional activation of different Bvg-activated genes and the
Bvg-independent gene recA. Wt strain was grown in liquid cultures under modulating
conditions (in the presence of 40 mM MgSO4) for 18h, spun and re-suspended in
media without MgSO4. Total RNA was isolated at indicated times and cDNA was
prepared as described in the Material and Methods. RT-PCR was used to detect the
transcript levels of indicated genes.
74

fha
5 30 60 240
+MgS
O4
bcfA
bipA
recAg0
Time (min)
prn
fha
5 30 60 240
+MgS
O4
bcfAbcfA
bipA
recArecAg0
Time (min)
prn
75

Identification of BvgA box like sequences upstream of the bcfA ORFs. Based on
our results demonstrating BvgAS-mediated activation of bcfA expression (Figs. 2 and
3), we hypothesized that regions upstream of bcfA ORF will harbor BvgA binding
sites. Scanning of the sequences upstream of the predicted translational start site of
the bcfA ORF revealed the presence of two directly joined inverted repeats (IR2 and
IR3) that were similar to previously identified BvgA binding sites (Fig. 5) and (6, 12).
In addition to the conjoined inverted repeat, we also detected additional overlapping
inverted repeat sequences (IR1) where the two halves were separated by multiple
nucleotides (Fig. 5). For some Bvg-regulated promoters it has been shown that BvgA
binding repeats are not conjoined but are separated by multiple nucleotides-2 for
cyaA, 10 for ptx (encodes for pertussis toxin) and 27 and 37 nucleotides respectively
for bipA (7, 11, 12, 23). The presence of BvgA binding sites upstream of the ORFs
combined with our finding that bcfA is positively regulated by BvgAS suggests that
BvgAS control of bcfA expression is direct.
BvgA and BvgA-P bind to promoter region of bcfA. The presence of putative
BvgA binding elements upstream of bcfA ORF led us to speculate that BvgA will bind
the bcfA promoter. To demonstrate BvgA occupancy to the bcfA promoter, we
conducted EMSA by utilizing a PCR fragment that was essentially similar to the
fragment used for β-galactosidase assays (see above and Materials and Methods).
Purified recombinant BvgA and BvgA phosphorylated in vitro with acetyl phosphate
(BvgA-P) were used for this analysis. These reactions also contained poly(dI-dC) as
a nonspecific competitor DNA. The results of EMSA are shown in Fig. 6A. Both BvgA
76

FIG. 5. The putative promoter region of bcfA. Arrows on top of the individual DNA
sequences designate sequence elements (bold) that are similar to the consensus BvgA
binding site. The predicted translational start codon is italicized.
77

IR1
IR2
IR3
ACAGAAACCTGAATTTAATGGAGTTCCTGTCATCACGAAAGGTTCCATTTTTTTGTGCACTGGCGCTTTTGGTCTGGCTGCCTA
TCAAGGAAATATCCTACATATCCGTAGGATTGGGCTGGCATCAGGACCGCACGGATCCAAATAATCTTCCTACATCGATTCTCC
GATATGTCTGCATAGCTCACGGGTTGGCAGGTGTTGTCGGTCCAACCAAGGGCCGTATCCGCCGGAAGTCGAAATTTCCTCTCT
GCCATTTCCTCCGATCGCCACGCCGGCGACCGGCGAGTCTTCTTGCGCCCTGAATGGCGCATTCCAAGGGCCACGGCCCGCGGG
CATGGCGGATCGATCGTGAACTTGGGAAAGGAGTAATCCGTGAAGCAAGCCATCCACGCCGTTGCGTTCCGCCATGATGCGCTC
HS1
IR1
IR2
IR3
ACAGAAACCTGAATTTAATGGAGTTCCTGTCATCACGAAAGGTTCCATTTTTTTGTGCACTGGCGCTTTTGGTCTGGCTGCCTA
TCAAGGAAATATCCTACATATCCGTAGGATTGGGCTGGCATCAGGACCGCACGGATCCAAATAATCTTCCTACATCGATTCTCC
GATATGTCTGCATAGCTCACGGGTTGGCAGGTGTTGTCGGTCCAACCAAGGGCCGTATCCGCCGGAAGTCGAAATTTCCTCTCT
GCCATTTCCTCCGATCGCCACGCCGGCGACCGGCGAGTCTTCTTGCGCCCTGAATGGCGCATTCCAAGGGCCACGGCCCGCGGG
CATGGCGGATCGATCGTGAACTTGGGAAAGGAGTAATCCGTGAAGCAAGCCATCCACGCCGTTGCGTTCCGCCATGATGCGCTC
HS1
78

and BvgA-P retarded the mobility of the radiolabeled DNA fragment. As a result of
phosphorylation, occupancy of BvgA to the radiolabeled DNA fragment gave rise to
higher order protein-DNA complexes suggestive of multiple binding sites or
multimerization of BvgA as a result of phosphorylation (Fig. 6A, compare lanes 2-6
with lanes 8-12, respectively). In order to confirm the specificity of BvgA binding, we
also performed competition EMSAs with increasing concentrations of specific (Fig.
6B) and non specific competitors (Fig. 6C). For specific competition, we used
unlabeled bcfA promoter fragment and as a non specific competitor the DNA
fragment corresponding to +453 to +978 of the bcfA coding region was utilized. There
was a gradual loss of BvgA binding activity with increasing concentrations of the
specific competitor (Fig. 6B). In contrast, in the presence of the non-specific
competitor, there was no significant loss in the binding affinity of BvgA-P. Therefore,
these results suggest that interaction of BvgA-P to bcfA promoter is DNA sequence
specific.
BcfA is an outer membrane protein. Due to the homology of BcfA to BipA,
particularly in the N-terminal region (Fig. 1), we hypothesized that BcfA will also be
localized to the outer membrane. We constructed two isogenic strains, RKD110
(ΔbcfA), and a double deletion strain, MM101 (ΔbipAΔbcfA). These strains are in-
frame deletions of the respective genes and are non-polar. Notably, these strains are
comparable to the wt strain with respect to growth in laboratory cultures (data not
shown). We purified the outer membranes from the wt, Bvg- phase locked strain,
RKD110 (ΔbcfA), RB25 (ΔbipA) and MM101 (ΔbipAΔbcfA), as the Triton X-100
79

FIG. 6. Electrophoretic Mobility Shift Assay.
A. 32P-end labeled bcfA promoter fragment was incubated with varying
concentrations of either BvgA (lanes 2-6) or BvgA-P (lanes 8-12). The DNA-protein
complexes were separated by electrophoresis on 4% non-denaturing polyacrylamide
gel and visualized by autoradiography. Lane 1,7 DNA alone. Lanes 2-6, 0.12, 0.6,
1.2 1.8 and 2.4 μg of BvgA, respectively. Lanes 8-12, same as lanes 2-5 except that
20 mM acetyl phosphate was added to the reactions.
B. Specific competition (lanes 3-5) was carried out using 10, 50 and 100 fold excess
of unlabeled bcfA promoter fragment. Lane1, DNA alone. 2 – 7, BvgA-P- 1.8μg. The
reactions were carried out in the presence of 20 mM acetyl phosphate.
C. Non-specific competition (lanes 3-7) was performed with 10,100,150 and 450 fold
excess of a 525 bp fragment encompassing a region corresponding to the bcfA ORF.
Lane1, DNA alone. 2 – 7, BvgA-P- 1.8μg. The reactions were carried out in the
presence of 20 mM acetyl phosphate.
80

1
0 0BvgA
BvgA-P
2 3 4 5 6 7 8 9 10 11 12
B C
A
1 2 3 4 5 6
Sp. competitor
BvgA-P0
1 2 3 4 5 6 7
Non sp. competitor
BvgA-P0
7
1
0 0BvgA
BvgA-P0 0
BvgABvgA-P
2 3 4 5 6 7 8 9 10 11 12
B C
A
1 2 3 4 5 6
Sp. competitor
BvgA-P0
1 2 3 4 5 6 7
Non sp. competitor
BvgA-P0
7
81

insoluble fractions (see Material and Methods). To facilitate the detection of BcfA,
polyclonal sera were raised against a BcfA-T7-tagged fusion protein purified from E.
coli (see Materials and Methods). Western-blotting with anti-BcfA antibody detected a
polypeptide that migrated at a mobility corresponding to ≈ 100 kDa in the outer
membrane fractions isolated from the wt and the RB25 (ΔbipA) strains but not from
the Bvg- phase locked strain, the RKD110 (ΔbcfA) and the MM101 (ΔbipAΔbcfA)
strains. The absence of a cross-reactive polypeptide band in the Bvg- phase locked
strain, RKD110 (ΔbcfA) and MM101 (ΔbipAΔbcfA) fractions confirms that the
polyclonal serum is specifically recognizing BcfA. As a positive control for a known
outer membrane protein, we probed for BipA. A previously described anti-BipA
antibody (33) raised against the C-terminus of BipA recognized a polypeptide present
in the outer membrane fractions prepared from wt and RKD110 (ΔbcfA) strains but
not from that prepared from the Bvg- phase locked, the RB25 (ΔbipA) and MM101
(ΔbipAΔbcfA) strains (Fig. 7).
BcfA is expressed during the Bordetella infectious cycle. To evaluate whether
BcfA is expressed during infection, we used sera from rats infected for 30 days with
wt, RKD110 (ΔbcfA), RB25 (ΔbipA) and MM101 (ΔbipAΔbcfA) strains as a probe to
detect the purified BcfA protein in western blotting assays. While the serum from the
wt and the RB25 (ΔbipA)-infected rats recognized the purified BcfA protein, sera from
RKD110 (ΔbcfA) and MM101 (ΔbipAΔbcfA) infected rats (Fig. 8) showed no
significant reactivity. Western blot analysis consistently revealed two bands, one
around 100 kDa corresponding to approximately the predicted size of the BcfA
82

FIG.7. BcfA is localized to the outer-membrane. Outer membrane proteins were
prepared by cellular fractionation of the indicated strains and were subjected to
western blot analysis using polyclonal antibodies raised against purified B.
bronchiseptica BcfA and BipA. The molecular weight markers used are indicated.
83

wt
Δbc
fA
Δbi
pA
Δbi
pAΔ
bcfA
Anti-BipA antibody180 115
B
180115
82w
t
Δbc
fA
Δbi
pA
Δbi
pAΔ
bcfA
Rec
. Bcf
A
A
Anti-BcfA antibodyw
t
Δbc
fA
Δbi
pA
Δbi
pAΔ
bcfA
Anti-BipA antibody180 115
B
180115
82w
t
Δbc
fA
Δbi
pA
Δbi
pAΔ
bcfA
Rec
. Bcf
A
A
Anti-BcfA antibody
84

FIG. 8. BcfA is expressed during infection. The purified recombinant BcfA protein
was subjected to SDS-PAGE and western blot analysis using serum collected 30
days post inoculation from rats infected with the indicated strains.
85
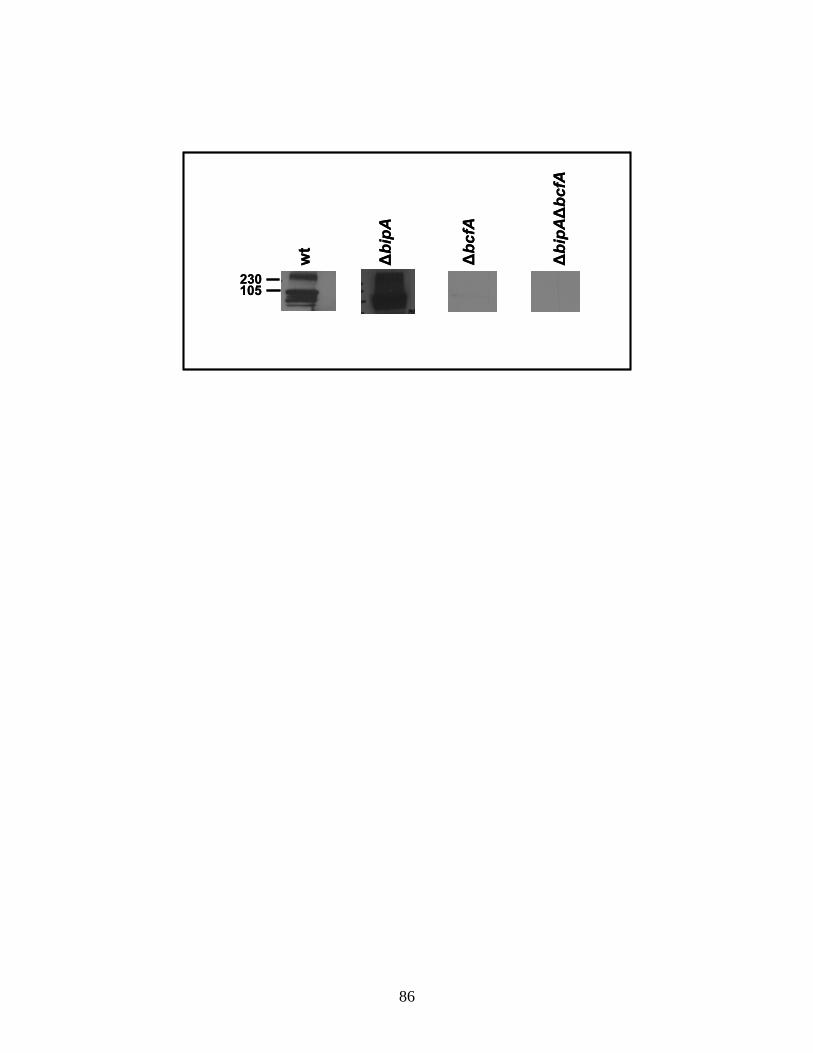
wt
Δbc
fA
Δbi
pAΔ
bcfA
Δbi
pA
105230
wt
Δbc
fA
Δbi
pAΔ
bcfA
Δbi
pA
105230
86

protein and another higher molecular weight polypeptide which migrated at greater
than 230 kDa. This higher molecular weight peptide was not recognized by sera from
RKD110 (ΔbcfA) and MM110 (ΔbipAΔbcfA) infected rats, thereby suggesting that this
polypeptide might be a higher molecular weight (possibly a dimer) form of BcfA (Fig.
8). However, this higher molecular weight band was not seen with outer membrane
fractions (Fig. 7), suggesting that it might also be an artifact of the purification
procedure. In addition to sera from rats infected with RB50 for 30 days, sera collected
from rats infected with the wt strain for 12 days also recognized purified BcfA (data
not shown).
Overlapping function of BipA and BcfA in colonization of the lower respiratory
tract. The similarity of BcfA to BipA and the failure to detect a function for bipA in
colonization of the upper respiratory tract led us to hypothesize that BcfA either alone
or in combination with BipA will play a role in respiratory tract colonization. We tested
the effect of these deletions on respiratory tract colonization. Groups of six female
Wistar rats were inoculated intranasally with the wt, RB25 (ΔbipA), RKD110 (ΔbcfA)
and MM110 (ΔbipAΔbcfA) strains. For intranasal inoculation, we utilized the well
studied and frequently utilized low volume-low-inoculum protocol (1, 18). Twelve days
and thirty days post-inoculation, animals were sacrificed and colonization levels were
determined by removing the entire nasal septum and 1 cm of the trachea. Each
tissue was homogenized in 200 μl of PBS, aliquots were diluted and viable colonies
were enumerated by plating on BG agar containing blood.
87

Consistent with previously published results (1, 18), high numbers of bacteria
ranging from 104 to 106 CFUs were recovered from the nasal septums and tracheas
of the wt-inoculated animals at both time points (Fig. 9). Although some animals (one
each for nasal septum at 12 days and 30 days and one for trachea at 12 days) did
not get colonized by the RB25 (ΔbipA) strain, there was no statistically significant
difference in the mean colonization levels as a result of the deletion of bipA. Note that
the failure of the RB25 (ΔbipA) strain to elicit a significant colonization defect is
consistent with previously reported results (33). Similarly, the difference in mean
colonization levels for the wt strain and the RKD110 (ΔbcfA) strain was not
statistically significant at any of the time points tested. In contrast to the individual
deletion of bipA and bcfA, a combined deletion of these two genes resulted in a
drastic defect in the colonization of the trachea at both the early and late time points
(Fig. 9). Notably, for five rats at 12 days and for four rats at 30 days, the number of
CFUs recovered from the trachea was either at or below the lower limit of detection
(Fig. 9). At 12 days, three animals did not get colonized in the nasal septum with the
double deletion strain, but this defect in nasal colonization was not apparent at thirty
days when all of the six animals were colonized and the mean colonization level at
this time point was not significantly different when compared to the wt strain. These
results thus suggest that BipA and BcfA have overlapping roles in colonization of the
trachea.
88

FIG. 9. Colonization of rat respiratory tract by wt and isogenic mutant derivatives
RB25 (ΔbipA), RKD110 (ΔbcfA) and MM101 (ΔbipAΔbcfA). (A) 12 days and (B) 30
days post inoculation. 4-5 weeks old female Wistar rats were inoculated with the
respective strains. The entire nasal septum and 1cm of trachea were harvested at
indicated times, homogenized and aliquots were plated on BG agar plates containing
7.5 % defibrinated blood. The plates also contain 50 μg/ml of streptomycin. The
resultant CFUs were enumerated. Horizontal bars indicate mean of each group. The
dashed lines represent the lower limit of detection.
89

wt
ΔbipAΔbcfA
∆bipA∆bcfA
Nasal Septum Trachea
1.0E+00
1.0E+01
1.0E+02
1.0E+03
1.0E+04
1.0E+05
1.0E+06
1.0E+07
1.0E+00
1.0E+01
1.0E+02
1.0E+03
1.0E+04
1.0E+05
1.0E+06
1.0E+00
1.0E+01
1.0E+02
1.0E+03
1.0E+04
1.0E+05
1.0E+06
1.0E+00
1.0E+01
1.0E+02
1.0E+03
1.0E+04
1.0E+05
1.0E+06
A
B Nasal Septum Trachea
wt
ΔbipAΔbcfA
∆bipA∆bcfA
Nasal Septum Trachea
1.0E+00
1.0E+01
1.0E+02
1.0E+03
1.0E+04
1.0E+05
1.0E+06
1.0E+07
1.0E+00
1.0E+01
1.0E+02
1.0E+03
1.0E+04
1.0E+05
1.0E+06
1.0E+00
1.0E+01
1.0E+02
1.0E+03
1.0E+04
1.0E+05
1.0E+06
1.0E+00
1.0E+01
1.0E+02
1.0E+03
1.0E+04
1.0E+05
1.0E+06
1.0E+00
1.0E+01
1.0E+02
1.0E+03
1.0E+04
1.0E+05
1.0E+06
1.0E+00
1.0E+01
1.0E+02
1.0E+03
1.0E+04
1.0E+05
1.0E+06
A
B Nasal Septum Trachea
90

DISCUSSION
Efficient colonization of the mammalian respiratory tract by Bordetella occurs by the
production of a collection of virulence factors that include a wide variety of adhesins
and several toxins. Because of the presence of multiple potential adhesin molecules,
it is easy to envision the existence of redundancy in the adherence mechanisms of
Bordetella to cultured epithelial cells in vitro or to the respiratory tract. One or more
adhesin molecules can mediate their action by acting either in a concerted manner or
synergistically. In this study, we report the identification of a Bordetella gene bcfA, a
paralog of the previously identified Bordetella gene bipA (39). We were encouraged
to search for a gene homologous to bipA because a previous report (39) failed to
identify a role for BipA in respiratory tract colonization. With the aid of BLAST
searches, we have detected an ORF in B. bronchiseptica that harbors significant
similarity to BipA. Comparison of BcfA amino acid sequence revealed most significant
sequence identity of the N-terminal region with the corresponding regions of BipA and
intimins of EPEC and EHEC and invasins of Yersinia species. The N-terminal region
in intimins and invasins is required for outer membrane localization and for the export
of the carboxyl termini of these proteins (25, 39). Western Blot analysis of the outer
membrane fractions demonstrated that BcfA, like BipA is localized in the outer
membrane.
The BvgAS signal transduction system is the principal regulator of virulence
gene expression in all the three Bordetella species. We investigated the Bvg-
mediated control of bcfA expression in B. bronchiseptica. Our data show that BvgAS
positively regulates the expression of bcfA. We and others have previously reported
91

that bipA is expressed maximally in the Bvg-intermediate phase (12, 16, 33). Using
real time RT-PCR and β-galctosidase assays to quantitate the expression of bcfA
under different phenotypic phases, we show that in contrast to bipA, bcfA is
expressed maximally both in the Bvg+ and Bvgi phases. In this respect, the
expression profile of bcfA resembles that of another well-studied Bordetella adhesin,
FHA (Fig. 3) and (9, 11). Our kinetic transcriptional activation results indicated that
bipA and bcfA are activated concurrently after the initiation of an inducing signal.
Although results from these two studies seem contradictory at first, it is to be noted
that these represent two different ways by which the transition between the different
activation states of BvgA can occur. In the phase-locked mutant strains, the BvgAS
system is locked in its different activation states (ranging from maximal activation
(Bvg+-phase), sub-maximal (Bvgi-phase) to a minimal (Bvg--phase). In contrast, when
Bordetella are grown overnight in the presence of chemical modulators followed by
subsequent growth in their absence, the activity of BvgA transitions from that of
minimal activation state at early time-points to a maximal state at late time-points.
Thus, it can be hypothesized that genes that have a requirement for a low
concentration of BvgA-P to be activated, will be turned on first. While there are
obvious differences in the arrangement and the affinity of various BvgA binding sites
of fhaB, bipA and bcfA, a common feature of the promoters of all these genes is the
presence of a high affinity BvgA binding site in the form of inverted heptanucleotide
repeats (centered at -88.5 for fha (6), IR1 for bipA (11, 12) and IR2 for bcfA, our
unpublished results). Indeed we and others have demonstrated that under in vitro
conditions using purified BvgA and RNA polymerase, activation of fhaB and bipA
92

occurs at a relatively low concentration of BvgA-P (22, 29). We hypothesize that
similar to fhaB and bipA, bcfA will require a low concentration of BvgA-P to be
activated in vitro and are currently in the process of testing this hypothesis.
Encouraged by our finding of sequence identity of BcfA with BipA and other
bacterial adhesins, we investigated whether BcfA singly or in unison with BipA will
play a role in respiratory tract colonization. We utilized both single as well as double
deletion mutants. Our results utilizing an intranasal rat model of colonization show
that there was no significant demonstrable phenotype in respiratory tract colonization
as a result of individually deleting either bipA or bcfA. Both BipA and BcfA deficient
strains are similar to their isogenic wild type parent in their ability to colonize the nose
and the trachea. However, a combined deficiency of BipA and BcfA practically
resulted in an absence of tracheal colonization, showing that the two proteins indeed
have a role in Bordetella pathogenesis.
These findings raise an important question concerning the function of these
two proteins in Bordetella infections. One of our first hypotheses was that the function
of BipA and BcfA will be different. Based on our inability to detect a defect as a result
of individual deletion of either BipA or BcfA, this scenario seems highly unlikely.
Rather our data indicate that BipA and BcfA have either collective and/or overlapping
functions in Bordetella lifecycle in the host. We believe that this hypothesis is more
likely since a defect in respiratory tract colonization is apparent only after the
disruption of the function of both genes.
What role do these two proteins precisely play in colonization of the respiratory
tract? One simplest explanation is that these proteins are acting as adhesins, i.e.
93

they are directly involved in attachment to the respiratory epithelium. By utilizing
epithelial cell lines and tracheal explants, we are currently testing whether BcfA in
directly involved in attachment. It is also possible that the defect in colonization
observed as a result of gene deletions is not dependent directly on adherence to
respiratory epithelia and that BipA and BcfA might be involved in modulating the
components of the innate and/or adaptive immune systems. In this context, it is
important to note that the major Bordetella adhesin FHA plays an accessory role in
down-regulating the innate immune responses resulting in increased bacterial
persistence (20, 27, 28).
In conclusion, continued studies directed towards deciphering the mechanistic
basis of the role of BipA and BcfA in Bordetella life cycle within its hosts will provide
important insights towards understanding the multipartite nature of Bordetella-host
interactions. Additionally, studies targeted towards dissecting the biochemical basis
of the BvgAS–mediated control of these two genes will allow a clear and detailed
understanding of how a single regulatory locus directs the observed variations in
signal-dependent gene expression patterns.
94

ACKNOWLEDGMENTS
We thank Dan Wozniak for critical reading of the manuscript. Research in the
laboratory of RD is supported by funds from Wake Forest University Health Sciences,
National Research Initiative of the USDA Cooperative State Research, Education and
Extension Service, grant number # 35604-16874." and by NIH grant # R21
AI071054.
95

REFERENCES
1. Akerley, B. J., P. A. Cotter, and J. F. Miller. 1995. Ectopic Expression of the Flagellar Regulon Alters Development of the Bordetella Host Interaction. Cell 80:611-620.
2. Altschul, S. F., W. Gish, W. Miller, E. W. Myers, and D. J. Lipman. 1990. Basic Local Alignment Search Tool3. Journal of Molecular Biology 215:403-410. 3. Bendtsen, J. D., H. Nielsen, G. von Heijne, and S. Brunak. 2004. Improved prediction of signal peptides: SignalP 3.04. Journal of Molecular Biology 340:783-795.
4. Boucher, P. E., A. E. Maris, M. S. Yang, and S. Stibitz. 2003. The response regulator BvgA and RNA polymerase a subunit C-terminal domain bind simultaneously to different faces of the same segment of promoter DNA. Molecular Cell 11:163-173.
5. Boucher, P. E., F. D. Menozzi, and C. Locht. 1994. The Modular Architecture of Bacterial Response Regulators - Insights Into the Activation Mechanism of the Bvga Transactivator of Bordetella-Pertussis . Journal of Molecular Biology 241:363-377.
6. Boucher, P. E., K. Murakami, A. Ishihama, and S. Stibitz. 1997. Nature of DNA binding and RNA polymerase interaction of the Bordetella pertussis BvgA transcriptional activator at the fha promoter. Journal of Bacteriology 179:1755-1763.
7. Boucher, P. E. and S. Stibitz. 1995. Synergistic Binding of Rna-Polymerase and Bvga Phosphate to the Pertussis Toxin Promoter of Bordetella-Pertussis. Journal of Bacteriology 177:6486-6491.
8. Celli, J., W. Y. Deng, and B. B. Finlay. 2000. Enteropathogenic Escherichia coli (EPEC) attachment to epithelial cells: exploiting the host cell cytoskeleton from the outside. Cellular Microbiology 2:1-9.
9. Cotter, P. A. and J. F. Miller. 1997. A mutation in the Bordetella bronchiseptica bvgS gene results in reduced virulence and increased resistance to starvation, and identifies a new class of Bvg-regulated antigens. Molecular Microbiology 24:671-685.
10. de Tejada, G. M., P. A. Cotter, U. Heininger, A. Camilli, B. J. Akerley, J. J. Mekalanos, and J. F. Miller. 1998. Neither the Bvg(-) phase nor the vrg6 locus of Bordetella pertussis is required for respiratory infection in mice. Infection and Immunity 66:2762-2768.
96

11. Deora, R. 2002. Differential regulation of the Bordetella bipA gene: Distinct roles for different BvgA binding sites. Journal of Bacteriology 184:6942-6951.
12. Deora, R., H. J. Bootsma, J. F. Miller, and P. A. Cotter. 2001. Diversity in the Bordetella virulence regulon: transcriptional control of a Bvg-intermediate phase gene. Molecular Microbiology 40:669-683.
13. deTejada, G. M., J. F. Miller, and P. A. Cotter. 1996. Comparative analysis of the virulence control systems of Bordetella pertussis and Bordetella bronchiseptica. Molecular Microbiology 22:895-908.
14. Edwards, R. A., L. H. Keller, and D. M. Schifferli. 1998. Improved allelic exchange vectors and their use to analyze 987P fimbria gene expression 2. Gene 207:149-157.
15. Frankel, G., A. D. Phillips, L. R. Trabulsi, S. Knutton, G. Dougan, and S. Matthews. 2001. Intimin and the host cell - is it bound to end in Tir(s)?. Trends in Microbiology 9:214-218. 16. Fuchslocher, B., L. L. Millar, and P. A. Cotter. 2003. Comparison of bipA Alleles within and across Bordetella species 1. Infection and Immunity 71:3043-3052.
17. Goodnow, R. A. 1980. Biology of Bordetella bronchiseptica. Microbiol.Rev. 44:722-738.
18. Harvill, E. T., P. A. Cotter, and J. F. Miller. 1999. Pregenomic comparative analysis between Bordetella bronchiseptica RB50 and Bordetella pertussis Tohama I in murine models of respiratory tract infection. Infection and Immunity 67:6109-6118.
19. Heininger, U., P. A. Cotter, H. W. Fescemyer, G. M. de Tejada, M. H. Yuk, J. F. Miller, and E. T. Harvill. 2002. Comparative phenotypic analysis of the Bordetella parapertussis isolate chosen for genomic sequencing. Infection and Immunity 70:3777-3784.
20. Inatsuka, C. S., S. M. Julio, and P. A. Cotter. 2005. Bordetella filamentous hemagglutinin plays a critical role in immunomodulation, suggesting a mechanism for host specificity. Proceedings of the National Academy of Sciences of the United States of America 102:18578-18583. 21. Isberg, R. R. and P. Barnes. 2001. Subversion of integrins by enteropathogenic Yersinia. Journal of Cell Science 114:21-28. 22. Jones, A. M., P. E. Boucher, C. L. Williams, S. Stibitz, and P. A. Cotter. 2005. Role of BvgA phosphorylation and DNA binding affinity in control of Bvg-mediated phenotypic phase transition in Bordetella pertussis. Molecular Microbiology 58:700-713.
97

23. Karimova, G. and A. Ullmann. 1997. Characterization of DNA binding sites for the BvgA protein of Bordetella pertussis. Journal of Bacteriology 179:3790-3792. 24. Kinnear, S. M., P. E. Boucher, S. Stibitz, and N. H. Carbonetti. 1999. Analysis of BvgA activation of the pertactin gene promoter in Bordetella pertussis. Journal of Bacteriology 181:5234-5241.
25. Leong, J. M., R. S. Fournier, and R. R. Isberg. 1990. Identification of the Integrin Binding Domain of the Yersinia-Pseudotuberculosis Invasin Protein. Embo Journal 9:1979-1989. 26. Mattoo, S. and J. D. Cherry. 2005. Molecular pathogenesis, epidemiology, and clinical manifestations of respiratory infections due to Bordetella pertussis and other Bordetella subspecies. Clinical Microbiology Reviews 18:326-+. 27. McGuirk, P., C. McCann, and K. H. G. Mills. 2002. Pathogen-specific T regulatory 1 cells induced in the respiratory tract by a bacterial molecule that stimulates interleukin 10 production by dendritic cells: A novel strategy for evasion of protective T helper type 1 responses by Bordetella pertussis. Journal of Experimental Medicine 195:221-231. 28. McGuirk, P. and K. H. G. Mills. 2000. Direct anti-inflammatory effect of a bacterial virulence factor: IL-10-dependent suppression of IL-12 production by filamentous hemagglutinin from Bordetella pertussis. European Journal of Immunology 30:415-422. 29. Mishra, M. and R. Deora. 2005. Mode of action of the Bordetella BvgA protein: Transcriptional activation and repression of the Bordetella bronchiseptica bipA promoter. Journal of Bacteriology 187:6290-6299. 30. Relman, D., E. Tuomanen, S. Falkow, D. T. Golenbock, K. Saukkonen, and S. D. Wright. 1990. Recognition of A Bacterial Adhesin by An Integrin - Macrophage Cr3 (Alpha-M-Beta-2, Cd11B Cd18) Binds Filamentous Hemagglutinin of Bordetella-Pertussis. Cell 61:1375-1382. 31. Simon, R., U. Priefer, and A. Puhler. 1983. A Broad Host Range Mobilization System for Invivo Genetic-Engineering - Transposon Mutagenesis in Gram-Negative Bacteria. Bio-Technology 1:784-791.
32. Stainer, D. a. S. M. 1971. A simple chemically defined medium for the production of phase I Bordetella pertussis. J.Gen.Microbiology 211-220.
33. Stockbauer, K. E., B. Fuchslocher, J. F. Miller, and P. A. Cotter. 2001. Identification and characterization of BipA, a Bordetella Bvg-intermediate phase protein. Molecular Microbiology 39:65-78.
98

34. Uhl, M. A. and J. F. Miller. 1996. Central role of the BvgS receiver as a phosphorylated intermediate in a complex two-component phosphorelay. Journal of Biological Chemistry 271:33176-33180.
35. Uhl, M. A. and J. F. Miller. 1996. Integration of multiple domains in a two-component sensor protein: The Bordetella pertussis BVgAS phosphorelay. Embo Journal 15:1028-1036.
36. Watanabe M and Nagai, M. Whooping cough due to Bordetella parapertussis: an unresolved problem. Expert Rev Anti Infect Ther. 2[3], 447-454. 2004. 37. West, A. H. and A. M. Stock. 2001. Histidine kinases and response regulator proteins in two-component signaling systems. Trends in Biochemical Sciences 26:369-376.
38. Yeh, S. H. 2003. Pertussis: persistent pathogen, imperfect vaccines. Expert.Rev.Vaccines. 2:113-127.
39. Yu, L., E. A. Frey, R. A. Pfuetzner, A. L. Creagh, D. G. Knoechel, C. A. Haynes, B. B. Finlay, and N. C. J. Strynadka. 2000. Crystal structure of enteropathogenic Escherichia coli intimin-receptor complex. Nature 405:1073-1077.
99

Chapter III
Active and Passive Immunization with Bordetella Colonization
Factor A (BcfA) Protects Mice against Respiratory Challenge with
Bordetella bronchiseptica.
Neelima Sukumar, Cheraton F. Love, Matt S. Conover, Nancy D. Kock,
Purnima Dubey and Rajendar Deora
This chapter has been published in Infection and Immunity, 2009 Feb;77(2):885-95
and is reprinted with permission. Neelima Sukumar performed all the experiments
described in this chapter. All animal work were done in collaboration with Cheraton
Love and Matt Conover. Pathology evaluations were carried out by Dr. Nancy Kock.
100

INTRODUCTION
Respiratory pathogens are a major cause of morbidity and mortality in
humans and animals, making the development of efficacious vaccines that protect
against these infections a top priority. Bordetellae are small aerobic, Gram-negative
coccobacilli that colonize the respiratory tracts of humans and animals (31).
Bordetella pertussis infects only humans and causes the acute respiratory disease
whooping cough (6). B. parapertussis strains can be divided into two genetically
distinct types, those which infect humans causing a pertussis-like illness, and those
which cause respiratory infections in sheep (22, 38). B. avium mainly infects
commercially grown turkeys and wild and domesticated birds (43, 45). In contrast,
B. bronchiseptica has a broader host range and is considered a co-contributor to a
number of respiratory syndromes in agriculturally important food producing animals,
pets and nonhuman primates (17). B. bronchiseptica is also a primary etiological
agent and/or a pre disposing factor that results in Porcine Reproductive and
Respiratory Disease complex (PRDC), pneumonia and atrophic rhinitis (AR) in
swine, infectious tracheobronchitis ((ITB) or kennel cough) in dogs and
bronchopneumonia in sheep, guinea pigs, rats, mice, rabbits, cats and non human
primates (5, 31). According to the 2000 National Animal Health Monitoring System
(NAHMS) survey, respiratory disease was the greatest cause of mortality in swine,
accounting for 28.9 percent of nursery deaths and 39.1 percent of deaths in
grower/finisher pigs. The annual economic impact of AR and PRDC in USA alone is
estimated to be about $17 and $40 million, respectively. B. bronchiseptica is also
capable of infecting humans, mostly immunocompromized individuals with AIDS or
101

cystic fibrosis (14, 26, 46, 52), although it was recently isolated from an
immunocompetent individual (39).
Currently available and proposed vaccines against this pathogen include
live, attenuated, heat killed or genetically modified bacteria (2, 30, 32, 48, 49).
Problems associated with these various whole-cell vaccination approaches include:
persistence of the vaccine strain in animals, poor induction of an antibody response
and/or protective immunity and retention of some of the virulence characteristics by
the vaccine strains (2, 30, 32, 48, 49). The genetic mutations that result in the
attenuation of many of the commercially available live attenuated vaccines is
unknown, making it likely that these strains may revert to virulent forms because of
survival pressures in the host, such as co-infections with other pathogenic
organisms. B. bronchiseptica can predispose animals to other infectious agents or
exacerbate the disease symptoms. For example, B. bronchiseptica colonization
leads to increased severity of Canine parainfluenza virus-2 infections and
predisposition of pigs and rabbits to subsequent Pasturella multocida colonization
(8, 12, 15). Infection of porcine tracheal rings with B. bronchiseptica has also been
shown to enhance the adherence of P. multocida (13).
Despite vaccination, animals continue to be carriers resulting in outbreak
among herds. For laboratory animals like rats, mice and rabbits, experimental
infection with B. bronchiseptica results in a chronic and asymptomatic colonization
of the upper respiratory tract. We have been able to isolate B. bronchiseptica from
the rat nasopharynx even 85 days after inoculation (our unpublished results) and
this bacterium has previously been reported to exist in this site for the life of the
102

infected animals (30). Theoretically, persistent colonization of the upper respiratory
tract of the animals vaccinated with live or attenuated strains can create a reservoir
of infectious bacteria from which animal-animal and zoonotic transmission can
occur. Although transmission of a vaccine strain to humans has not been
experimentally proven, a number of such human cases have occurred in individuals
exposed to either infected, sick or recently immunized farm and companion animals
(20).
We propose that an effective acellular vaccination regimen capable of
providing long lasting protective immunity will limit the spread of B. bronchiseptica
not only among animals in a herd but also from animals to humans. For B.
pertussis, there has been a shift to acellular vaccines because of high frequency of
side effects and multiple adverse reactions associated with the whole cell vaccines
(34). Similarly, development of acellular vaccines capable of protecting against B.
bronchiseptica should be given a priority.
BcfA (Bordetella colonization factor) is an outer membrane protein which is
positively regulated by the BvgAS signal transduction system (50). We were
encouraged to examine the role of BcfA in protective immunity because our
previously published research revealed its role in the colonization of the rat trachea.
In addition, sera from rats infected with B. bronchiseptica specifically recognized
BcfA (50). In the current report, we have evaluated the immunogenicity and
protective efficacy of BcfA in an intranasal mouse model of respiratory infection.
Both active and passive immunization with BcfA provided protection against
subsequent intranasal challenge with B. bronchiseptica, which significantly
103

correlated with the production of subclasses of IgG antibodies with high opsonic
activity. Our results also suggested a role for a Th1 type cellular response in BcfA-
mediated protection. Finally, we demonstrated that BcfA is expressed by multiple
clinical isolates of B. bronchiseptica. Data presented in the current study
underscore the potential utility of an acellular vaccine approach for B.
bronchiseptica and highlight the importance of BcfA as a critical protective antigen
against B. bronchiseptica infections.
104

MATERIALS AND METHODS
Bacterial strains, media and growth conditions. The bacterial strains used in
this study are listed in Table 1. B. bronchiseptica wild type (wt) strain RB50 and the
isogenic mutant strain RKD110 (ΔbcfA) has been described previously (50). The
clinical strains of B. bronchiseptica were a kind gift from Dr. T.L. Nicholson and Dr.
K. Register at USDA-ARS. All the strains were maintained on Bordet-Gengou agar
(Becton Dickinson Microbiology Systems) containing 7.5% defibrinated sheep blood
(BG-agar). For RB50 and RKD110 (ΔbcfA), BG-agar was supplemented with
50μg/ml of streptomycin (SM) (Research Products International Corp). For animal
inoculations, single colonies of RB50 were inoculated into Stainer-Scholte broth
(47), cultured overnight, followed by sub-culture to OD600 of ≈ 1 at 37oC.
Over-expression and purification of BcfA. For over-expression of BcfA, E. coli
BL21(DE3)/pLysE cells containing the previously described bcfA over-expression
plasmid, pNS101 were used (50). Bacterial growth conditions, protein induction and
cell extract preparation were carried out as described previously (50). BcfA was
purified using a T7.Tag affinity purification kit (Novagen) as described (50).
Mouse immunizations. Five-six week old female C57/BL6 mice were obtained
from Charles River Laboratories. All experimental procedures were performed in
compliance with institutional regulations and were approved by the animal care and
use committee of WFUHS .
105

TABLE 1. Strains used in this study.
B. bronchiseptica strain isolated from a dog in United States
MBORD685
RB50 derivative having in-frame chromosomal deletion of bcfA (50) RKD110
B. bronchiseptica strain isolated from a sea otter in California (1999) SO3287-99
B. bronchiseptica strain isolated from a pig in Hungary (1993) (3) KM22
B. bronchiseptica strain isolated from a seal in Scottish coast (1999) M584/99/1
B. bronchiseptica strain isolated from a Koala in Australia MBORD698
B. bronchiseptica strain isolated from a cat in United States MBORD631
B. bronchiseptica strain isolated from a horse in United States MBORD628
Wild type B. bronchiseptica strain (9) RB50
Description/ Reference Strain
106

Active immunizations. Groups of four C57/BL6 mice were intraperitoneally
injected with 10 or 30μg of purified BcfA adsorbed to 50μg of alum (Sigma). The
control group of mice received 50μg of alum only. Three weeks later, all the mice
received another dose of the respective immunogen. A week after the second
immunization, mice were sedated with isoflurane (Abbott Laboratories) and
challenged intranasally with 5 x 105 CFU of RB50 in 25μl of sterile PBS. The
number of CFU delivered was confirmed by plating an aliquot of the inoculum on
BG-agar containing SM. One and 6 days post-challenge, mice were sacrificed and
nasal septum, trachea and left lung were harvested. Colonization levels were
determined by homogenizing these tissues in sterile PBS and plating various
dilutions of the homogenates on BG-agar containing SM. Prior to the second
immunization (post-first dose) and at the time of challenge (post-second dose),
blood was drawn from the tail to collect serum for ELISAs.
Passive immunizations. BcfA hyperimmune serum was generated in rats by
Covance as described previously (50). Excised pieces of standard SDS-
polyacrylamide protein gels containing the band corresponding to the BcfA protein
were utilized to immunize rats for the generation of anti-BcfA antibodies. Pre-
immune serum was collected from these rats prior to immunization with BcfA. Since
we were not able to obtain enough pre-immune serum, naïve rat serum (Invitrogen)
was utilized for some of these experiments. To obtain anti-B. bronchiseptica sera,
rats were intranasally infected with RB50 and convalescent phase serum was
collected 30 days post-inoculation.
107

Groups of five C57/BL6 mice were intraperitoneally injected with 200μl of
either anti-BcfA sera, anti-B. bronchiseptica sera, pre-immune serum, or sterile
PBS. Three to four hours later, they were intranasally challenged with 5 x 105 CFU
of RB50 in 25μl of sterile PBS. Mice were sacrificed 3 and 7 days post-challenge
and colonization levels were determined as described above.
Lung histopathology. The right lung from all mice were immersed in 10% neutral
buffered formalin (EMD Chemicals Inc.) just after sacrifice for at least 24 h,
trimmed, embedded in paraffin, processed routinely for histology, cut at 4-6μm,
stained with haematoxylin and eosin (H & E), and examined by light microscopy.
The sections were scored qualitatively for inflammation and injury, degree of overall
cellularity, thickness of alveolar walls, bronchiolar and vascular degeneration,
edema, hemorrhage, and degree and type of inflammatory cellular infiltration by
NK, a board certified veterinary pathologist, in a blind manner.
Neutrophil depletion. RB6-8C5 hybridoma cells were a kind gift from Dr. G.
Huffnagle (University of Michigan) (23, 29). Growth of these and purification of RB6-
8C5 monoclonal antibodies were performed as described previously (23, 29). For in
vivo neutrophil depletion, groups of five C57/BL6 mice were intraperitoneally
injected with 1mg of RB6-8C5 monoclonal antibody. Previous studies have shown
that this treatment regimen is able to deplete neutrophils for 1-2 weeks in mice (28).
A separate group of mice was injected with 1ml of sterile PBS. To determine the
efficiency of neutrophil depletion, blood collected from the tails was analyzed by
108

IDEXX laboratories for complete blood cell counts. The treatment was found to be
98% effective in depleting neutrophils (data not shown). One day post-
administration of RB6-8C5 antibodies, mice were intraperitoneally injected with
200μl of either anti-BcfA sera or naïve rat serum. Three to four h afterwards, they
were intranasally challenged with 5 x 105 CFU of RB50 in 25μl of sterile PBS. Two
days post-challenge, they were sacrificed and colonization levels were determined
as described above. We were unable to extend the infection of neutropenic mice
with B. bronchiseptica beyond two days, because 4 of 5 mice succumbed to
infection by 3 days.
ELISAs. Serum and lung homogenate antibody responses to BcfA were quantified
by coating 96 well flat-bottomed immuno plates (Nalge Nunc International) with
purified BcfA protein. The plates were incubated at 4oC overnight in a humidified
chamber. The wells were then washed 3 times with phosphate buffered saline
containing 0.05% Tween 20 (PBST) (EMD Chemicals Inc.). Blocking for non-
specific interactions was carried out by the addition of 200μl of 5% milk in PBST per
well and plates were incubated at 37oC for 1 h. Lung homogenates or serum
(100μl) from immunized mice or BcfA hyperimmune serum at various dilutions were
added and plates were incubated at 37oC for 2 h. Wells were washed 3 times with
PBST and bound antibodies were detected using HRP-conjugated goat anti-mouse
(Bio-Rad Laboratories) or goat anti-rat IgG (Rockland Inc.) (1:2000) antibodies.
Plates were washed 5 times with PBST and 3,3’, 5,5’-Tetramethyl benzidine
(Sigma) was used as the substrate. Absorbance at OD450 was determined using the
109

Labsystems Multiskan Plus plate reader. Absorbance was plotted against dilution
and the end point titer was determined as the inverse of the highest dilution giving
an OD450 reading 4-5 times above the background. Negative titers were plotted as
zero.
Specific class and isotypes of antibodies present in BcfA hyperimmune
serum and serum from BcfA-immunized mice were determined using rat and mouse
Immunoglobulin Isotyping ELISA Kit (BD Pharmingen), respectively according to
manufacturer’s instructions. Titers of the respective isotypes in the pre-immune
serum from rats or serum from alum-immunized mice were assayed by ELISA and
the OD450 values from these were subtracted from values obtained for BcfA
hyperimmune serum and serum from BcfA-immunized mice, respectively. The
resulting values were then plotted against the dilutions and the end point titer was
determined as the inverse of the highest dilution that gave an OD450 reading 4-5
times above the background. Negative titers were plotted as zero.
Opsonophagocytosis assay. The murine macrophage cell line J774A.1 and
murine monocyte/macrophage cell line RAW 264.7 were cultured in Dulbecco’s
Modified Eagle’s Medium (DMEM) (Invitrogen) supplemented with 10% fetal bovine
serum (FBS) (HyClone) and 4mM L- glutamine (Invitrogen). Approximately 2 x 105
cells were seeded into wells of a 24 well cell culture plate (Corning Incorporated)
and incubated overnight with 5% CO2. For opsonophagocytosis assay, BcfA
hyperimmune serum and naïve rat serum were heat inactivated by incubation at
55oC for 15 min. RB50 or RKD110 (ΔbcfA) was grown to an OD600 of 1 and ≈ 2 x
110

106 cells were incubated with either heat inactivated BcfA hyperimmune serum (at
1% or 10%) or naïve rat serum (10%) or PBS in 100μl at 37oC for 30 min. The
assay was performed by incubating macrophage cells with the above mentioned
mixture of serum or PBS containing bacteria, for 1 h. This was followed by
gentamycin (100μg/ml) (Invitrogen) treatment for 1 h to kill the extra-cellular
bacteria, washed twice with PBS to remove adherent bacteria, lysed with water and
different dilutions were plated to enumerate the number of phagocytosed bacteria.
The fold CFU for each treatment was calculated by dividing the intracellular CFU
obtained from different serum groups by the CFU obtained from the PBS control.
The assay was performed in triplicates and repeated 2-3 times.
T cell cytokine assays. Groups of five C57/BL6 mice were immunized at 0 and 3
weeks with 2 doses of either 30μg of BcfA adsorbed to alum or 50μg of alum only,
as described above. Two and four weeks after administrations of the second dose,
mice were sacrificed and spleens were harvested. Spleens were homogenized in
RPMI (Invitrogen) and red blood cells were lysed using ACK lysing buffer (Lonza).
Splenocytes were counted and approximately 2 x 106 cells were added to each well
of a 96 well plate in RPMI media supplemented with gentamycin. The splenocytes
were re-stimulated with 10μg of purified BcfA protein or medium alone.
Supernatants were harvested after 72 h of incubation and analyzed for IFN-γ, IL-4,
IL-10, IL-12 and TNF-α production using respective ELISA kits (BD OptEIA)
according to manufacturer’s instructions. Using the standards provided with the kit,
111

a standard curve was plotted for each cytokine and the respective cytokine
concentrations in the samples were derived from the standard curve.
SDS-PAGE and immunoblot analyses. Membrane fractions were prepared from
different B. bronchiseptica strains as described previously (50), resolved using
SDS-PAGE, transferred to nitrocellulose membranes (Osmonics Inc.) and probed
with enriched anti-BcfA antibody (1:5000). Anti-BcfA serum was enriched by
repeated incubation with overnight cultures of RKD110 (ΔbcfA) for 3-4 h as
described previously (50). Goat anti-rat IgG (Rockland Inc.) conjugated to HRP
(1:2000) was used as the secondary antibody and proteins were detected using the
Amersham ECL system.
112

RESULTS
Active immunization with BcfA induces protective immunity against B.
bronchiseptica infections. We have previously demonstrated that the outer-
membrane protein BcfA promotes colonization of the rat trachea (50). Our results
also suggested that BcfA-specific antibodies were produced during infection of
animals with B. bronchiseptica (50). Therefore, we hypothesized that the immune
response elicited against BcfA will provide protective immunity against a
subsequent B. bronchiseptica infection. To examine this hypothesis, mice were
intraperitoneally injected with two doses of either 10 or 30μg of purified BcfA
adsorbed to alum. One week after the second immunization, mice were challenged
with RB50 utilizing an intranasal inoculation regimen that seeds and leads to
colonization of the entire respiratory tract (30, 44). In control mice receiving only
alum, high numbers of bacteria were recovered from the nose, the trachea and the
lungs at 1 and 6 days following challenge (Figs. 1A and B, respectively). Three out
of four control mice showed signs of Bordetellosis and one mouse succumbed to
the infection at 6 days post-challenge. In contrast, mice immunized with either
doses of BcfA did not display any signs of Bordetellosis and had remarkably lower
bacterial burdens at both time-points post-challenge (Fig. 1). The reduction of
colonization due to immunization with BcfA was most dramatic in the lungs and the
trachea and at six days post-inoculation. Half of the mice immunized with the lower
dose and all the mice immunized with the higher dose of BcfA had no detectable
bacteria in the lungs (Fig. 1B). Both the immunizing doses resulted in lowering of
the bacterial burdens in tracheas of all of the vaccinated mice (Fig. 1B) and two of
113

FIG. 1. Immunization with BcfA protects mice against B. bronchiseptica challenge.
Mice were immunized intraperitoneally at 0 and 3 weeks with either 10 or 30μg of
BcfA adsorbed to alum or alum only. One week after the second immunization,
mice were intranasally challenged with 5 x 105 CFU of RB50 in a 25μl volume. Mice
were sacrificed at 1 day (A) and 6 days (B) post-challenge and the number of CFU
was determined in the nasal septum, trachea and lungs. Individual symbols
represent a single mouse. The dashed line represents the lower limits of CFU
detection. Black bars represent mean colonization of respective groups. A statistical
analysis was carried out using an unpaired two-tailed Student t test to compare the
CFU obtained from the respective groups of BcfA-immunized mice to that of mice
receiving alum. The asterisks indicate the range of the different P values (one
asterisk, ≤0.05; two asterisks, ≤0.005 and three asterisks, ≤0.0005).
114

0
1
2
3
4
5
6
Log 1
0 CFU
BcfA 10μg
BcfA 30μg
Alum
Trachea Nasal Septum Lungs
0
1
2
3
4
5
6
7
Log 1
0 CFU
Trachea Nasal Septum Lungs
A
B
*
*
* *
*** *
***
**
**
*
1 day
6 days
115

the four mice were cleared of bacteria from this site. Although immunization with
BcfA conferred protection against nasal challenge, compared to the other
respiratory organs, there was only a modest reduction in the bacterial burden. This
observation is consistent with the fact that B. bronchiseptica is extremely difficult to
clear from the nose (18). Our results thus demonstrate that immunization with BcfA
elicits immunity that is protective against B. bronchiseptica infections.
Immunization with BcfA reduces lung pathology in B. bronchiseptica infected
mice. Infection of mice with B. bronchiseptica results in significant lung pathology
and inflammation (21). We next sought to determine whether the reduced morbidity
observed as a result of B. bronchiseptica infection in BcfA-vaccinated mice,
compared to that observed in the control mice, correlated with reduced pulmonary
injury. Lungs from both groups of mice were excised at six days following challenge
with RB50 and examined microscopically. Control mice had pneumonia,
characterized by extensive neutrophilic infiltration of the parenchyma (Fig. 2A) while
BcfA-immunized mice had only modest cellular infiltration (Figs. 2B and 2C). Injury
scores from both groups are given in Table 2. Those of the control mice averaged
10.0, while those immunized with 10 or 30μg of BcfA were markedly lower at 3.0
and 2.7, respectively. This attests that immunization with BcfA considerably
diminishes the pulmonary injury in mice infected with B. bronchiseptica.
Immunization with BcfA induces high antibody responses. Our next objective
was to evaluate whether immunization with BcfA was able to induce the production
116

FIG. 2. Immunization with BcfA reduces lung pathology in mice challenged with
RB50. Representative H & E stained lung sections harvested six days post-
challenge with RB50 from mice immunized with alum only (A) or with 10μg (B) and
30μg (C) of BcfA adsorbed to alum. The sections were examined and evaluated in
a blinded manner. Magnification x 10.
117

C
A
B
118

TABLE 2. BcfA immunization reduces lung pathology. Mice were immunized with
10 or 30μg of BcfA adsorbed to alum or alum only and challenged with RB50 as
described in the legend to Fig. 1. Six days post-challenge, mice were sacrificed and
the right lung were harvested and processed for H & E staining. The sections were
examined by NK blinded to the treatment groups. The total score for each group
and the average score for indicated parameters are represented with ± standard
deviations. Unpaired two-tailed Student t test was used to compare pathology
scores between BcfA immunized and alum immunized mice. The asterisks indicate
the range of the different P values (one asterisk, ≤0.05; two asterisks, ≤0.005).
119

0.4 ± 0.6 0.09 ± 0.3 0 ± 0 Perivascular/Peribronchiolar Lymphocytes
10 ± 5.4 2.7 ± 2 3 ± 1.2 Average Total Score
0.4 ± 0.6 0.2 ± 0.4 0 ± 0 Alveolar Macrophages
1 ± 0.7 0.09 ± 0.3 0 ± 0 Intrabronchial PMNs
2 ± 1.0 0.3 ± 0.5 0.7 ± 0.5 Alveolar/ Interstitial PMNs
0.6 ± 0.9 0.09 ± 0.3 0.1 ± 0.4 Hemorrhage
1.2 ± 0.8 0 ± 0 0.4 ± 0.5 Edema
1 ± 0.7 0.18 ± 0.4 0 ±0 Degeneration
1.8 ± 0.5 1.5 ±0.5 1.7 ± 0.5 Alveolar Walls
1.6 ± 1.1 0.27 ± 0.5 0 ± 0 Consolidation
Alum BcfA 30μg BcfA 10μg Pathology parameters
* **
120

FIG. 3. Anti-BcfA antibody titers in immunized mice.
Anti-BcfA antibody titers in sera (A and B) and lung homogenates (C) collected from
mice immunized with 10μg or 30μg of BcfA adsorbed to alum or alum only.
A. Mouse serum was collected three weeks subsequent to delivery of the first dose
(post-first dose) and immediately before challenge with RB50 (post-second dose).
Total anti-BcfA IgG titers were determined using BcfA as an antigen by ELISA.
Values represented are mean titers from 3-5 mice and bars represent ± standard
deviation. A statistical analysis was carried out using an unpaired two-tailed Student
t test. The asterisks indicate the range of the different P values (one asterisk,
≤0.05; two asterisks, ≤0.005).
B. Mouse serum was collected immediately before challenge with RB50. Titers of
anti-BcfA-specific IgA, IgM and the different IgG isotypes were measured utilizing
specific ELISA kits as described in the Materials and Methods. Each symbol
represents an individual mouse. Black bars represent mean titer.
C. Total anti-BcfA IgG titers was determined from the lung homogenates of
individual mice immunized with either 10μg or 30μg of BcfA adsorbed to alum, alum
only or PBS. Lung homogenates were prepared six days post-challenge with RB50
as described in the Materials and Methods. Values represented are mean titers
from 3-4 mice and bars represent ± standard deviation. A statistical analysis was
carried out using an unpaired two-tailed Student t test. The asterisks indicate the
range of the different P values (one asterisk, ≤0.05; three asterisks, ≤0.0005).
121

0
500
BcfA 10μg BcfA 30μgAlum
700
10700
20700
30700
40700
50700
60700
Post- First Dose Post- Second Dose
**
* *
*
A.
122

0
10
20
30
40
50
60
70
80
90
IgG1 IgG2a IgG2b IgG3 IgA IgM
Tite
r ( X
10,
000)
B.
C.
0
10
20
30
40
50
BcfA 10μg BcfA 30μg Alum
Tota
l IgG
Tite
r
*** *
PBS
123

of specific antibodies in mice. A single dose of either 10 or 30μg of BcfA elicited
high levels of anti-BcfA IgG in the sera (Fig. 3A). After administration of the second
dose of BcfA, IgG levels were much higher, indicating a booster effect. In control
mice, the anti-BcfA IgG titers were at undetectable or background levels (Fig.
3A).While immunization with BcfA resulted in the induction of specific IgM
antibodies in three of the immunized mice, IgA antibodies were not detected (Fig.
3B). We also determined the levels of the different subclasses of anti-BcfA IgG
antibodies. While IgG1, IgG2b and IgG3 anti-BcfA antibodies were detected in the
sera of some of the BcfA-immunized mice, IgG2a was detected in all of the
immunized mice.
We also assayed lung homogenates from immunized mice 6 days post-
challenge with RB50 for antibody production. Whereas BcfA-immunized mice
generated a dose dependent antibody response in the lungs after infection with
RB50, mice which received only alum or PBS had considerably lower levels of anti-
BcfA antibodies (Fig. 3C). Therefore, these results are consistent with the
conclusion that clearance of B. bronchiseptica from the lower respiratory tract
correlates with the presence of high levels of specific antibodies in both the sera
and the lung homogenate.
Passive transfer of anti-BcfA antibodies provides protection against B.
bronchiseptica challenge. A critical role for anti-Bordetella antibodies in both
vaccine and infection-induced immunity against B. bronchiseptica has been
demonstrated (27) (18). We hypothesized that passive immunization with anti-BcfA
124

antibodies would protect mice against B. bronchiseptica challenge. For production
of hyperimmune serum, rats were immunized with excised polyacrylamide gel
fragments containing the recombinant BcfA protein and pre-immune and immune
rat sera were generated as described in the Materials and Methods. We have
previously shown that this polyclonal serum specifically recognizes BcfA (50).
ELISA showed that while in immune rats antibody titers specific for BcfA were very
high (≈1:70,000), in pre-immune rats, the levels of these specific antibodies were
undetectable.
For passive immunization experiments, groups of 5 mice were
intraperitoneally injected with either the BcfA hyperimmune serum, pre-immune rat
serum, the convalescent serum (from rats infected with B. bronchiseptica for 30
days) or sterile PBS, 3-4 hours prior to challenge with 5 x 105 CFU of B.
bronchiseptica. Three and 7 days post-challenge, mice were sacrificed and
bacterial burdens in the different respiratory organs were enumerated as described
in the Materials and Methods. Mice that received the BcfA-specific serum, harbored
lower bacterial burdens at 3 days post-challenge in both the trachea and the lungs,
compared to mice that received only PBS (Fig. 4A). At 7 days post-challenge, the
numbers of bacteria recovered from these sites was further reduced and in some
animals bacteria were not detected (Fig. 4B). Consistent with previous reports (27),
convalescent phase serum from B. bronchiseptica infected rats also resulted in the
lowering of bacterial numbers in the lungs at three days post-challenge and the
infection being cleared from both the trachea and the lungs of some mice at 7 days
post-challenge (Figs. 4A and B). Control mice which received pre-immune rat
125

FIG. 4. Effect of adoptive transfer of BcfA-specific sera on respiratory tract
colonization. Mice were intraperitoneally injected with anti-BcfA hyperimmune
serum, convalescent phase anti-RB50 serum, pre-immune serum or sterile PBS.
Three-four hours later, mice were intranasally challenged with 5 x 105 CFU of RB50
in 25μl volume. Three (A) and seven (B) days post-challenge, mice were sacrificed
and bacterial colonization in the nasal septum, trachea and lungs was determined.
Dashed line represents lower limits of CFU detection. Individual symbols represent
a single mouse. Black bars represent mean colonization of respective group.
Unpaired two-tailed Student t test was used to determine statistical significance.
The groups of mice receiving anti-BcfA hyperimmune serum and convalescent
phase anti-RB50 serum were compared to that of mice receiving sterile PBS. The
asterisks indicate the range of the different P values (one asterisk, ≤0.05; two
asterisks, ≤0.005; three asterisks, ≤0.0005). Differences between the CFU obtained
from pre-immune treated mice and PBS treated mice were not statistically
significant.
126
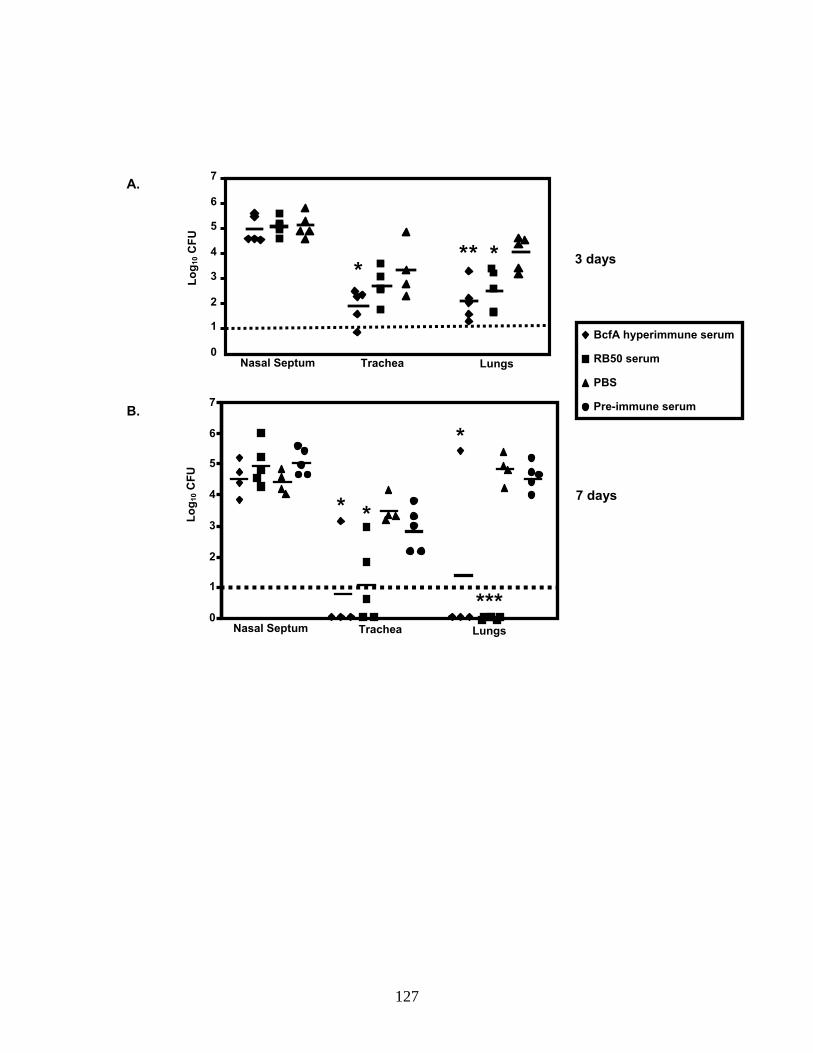
0
1
2
3
4
5
6
7
0 2
Log 1
0 CFU
BcfA hyperimmune serum
RB50 serum
PBS
Pre-immune serum
Trachea Nasal Septum Lungs
Trachea Nasal Septum Lungs 0
1
2
3
4
5
6
7 Lo
g 10 C
FU
***
*
*
A.
B.
3 days
7 days
*
*
***
127

serum or PBS prior to challenge showed high bacterial burdens in the lower
respiratory tract. In contrast to that observed in the trachea and the lungs, adoptive
transfer of either the BcfA hyperimmune serum or anti-B. bronchiseptica serum had
no significant effect on nasal septum colonization. These results thus suggest that
the protection from B. bronchiseptica infections observed in the lungs and the
trachea was mediated at least partially by antibodies generated in response to
immunization with BcfA. Further, these results also demonstrate that passively
transferred BcfA antibodies are efficient in protecting mice against B. bronchiseptica
challenge similar to those generated during an infection with the wt bacteria.
Anti-BcfA sera are opsonic. To better understand the mechanism of passive
protection, we next determined the levels of antibodies other than IgG and the
various IgG isotypes present in the rat sera utilized for passive protection. While
significant levels of IgM were detected in the anti-BcfA rat sera, no IgA was
detected. Isotyping analysis also revealed that whereas IgG1 was the predominant
isotype, IgG2a was present at lower levels (Fig. 5A).
The presence of IgG1, IgG2a and IgM in the immune serum has been
correlated with high opsonic activity (42). We examined the efficiency of BcfA
hyperimmune rat serum to promote opsonization and phagocytosis of B.
bronchiseptica by J774 murine macrophage cells (Fig. 5B) and RAW 264.7, the
murine monocyte/macrophage cells. These cells are frequently utilized to study
Bordetella pathogenesis and for phagocytic assays (24, 25, 41, 51). Naïve rat
serum or PBS was used as negative control. Opsonization with BcfA-specific sera
128

FIG. 5. Opsonization with anti-BcfA serum enhances the phagocytosis of RB50 by
J774 murine macrophages.
(A) Characterization of anti-BcfA antibodies in BcfA-specific hyperimmune serum.
Titers of different antibody classes and IgG isotypes was measured using specific
ELISA kit as described in the Materials and Methods. The titers obtained were
normalized to the titers of the pre-immune serum. ND = not detectable.
(B) Approximately, 2 x 106 CFU of RB50 or RKD110 (ΔbcfA) was incubated with
either 1 or 10% heat inactivated anti-BcfA hyperimmune serum, 10% naïve rat
serum or sterile PBS at 37o C for 30 min followed by incubation with 2 x 105 J774
cells for 1 h. Extracellular bacteria were killed by treatment with 100μg/ml of
gentamycin for 1 h followed by washing twice with sterile PBS to remove adherent
bacteria. The cells were lysed and the CFU of phagocytosed bacteria were
determined by plating on BG-agar containing SM. Results are expressed as fold
CFU of intracellular bacteria over the PBS treatment and are representative of three
independent experiments performed in triplicates. Bars represent ± standard
deviation. Statistical analysis was carried out using an unpaired two-tailed Student t
test. Asterisk indicates the P value of ≤0.05.
129

010
30
50
70
90
IgG1 IgG2a IgG2b IgG2c IgA IgM
Tite
rs (x
100
)
ND ND ND
A.
0
1
2
3
4
5
6
7
RB50
10% 1% 10% 1% 10% 10%
ΔbcfA
BcfA hyperimmune serum Naïve rat serum
PBS
Fold
intr
acel
lula
r CFU
*
B.
130

increased the efficiency of uptake of RB50 by J774 macrophages when compared
to incubation with naïve rat serum or PBS (Fig. 5B). In contrast, there were no
significant differences in the uptake of the RKD 110 (ΔbcfA) strain on opsonization
with either the BcfA-serum or naïve rat serum and PBS, thereby confirming the
specificity of BcfA-antibody mediated opsonization. Similar results were obtained
with the RAW 264.7 cells (data not shown). These results therefore suggest that
one of the mechanisms for the observed BcfA-mediated protection is increased
opsonization of B. bronchiseptica for phagocytosis.
Neutrophils are critical for anti-BcfA antibody-mediated bacterial clearance.
Neutrophils are a vital component of the immune responses responsible for clearing
B. bronchiseptica infections, since neutropenic mice succumb to B. bronchiseptica
infections within 1-4 days post-inoculation (21, 28). We hypothesized that
neutrophils will be a critical component of the BcfA-induced protective immunity. To
evaluate this possibility, we rendered mice neutropenic by injection of the anti-Gr1
antibody (RB6-8C5) which specifically depletes neutrophils without affecting
resident and circulating macrophages and lymphocytes (11, 28). Groups of 4-5
mice were administered either PBS or the RB6-8C5 antibody, followed by adoptive
transfer of either naïve rat sera or the anti-BcfA sera from rats. Subsequently, these
mice were inoculated with RB50. We examined only the lungs of these mice since
the effect of passive transfer of serum on bacterial clearance is greatest in lower
respiratory tract (Fig. 4 and (27)). Consistent with results presented in Fig. 4A, PBS-
injected mice which received BcfA-specific sera had significantly lower numbers of
131

FIG. 6. Neutrophils are required for anti-BcfA antibody-mediated clearance of B.
bronchiseptica. Mice were intraperitoneally injected with 1mg of RB6-8C5 antibody
or PBS. One day later mice were intraperitoneally injected with 200μl of anti-BcfA
serum or naïve rat serum followed by intranasal challenge with 5 x 105 CFU of
RB50 in a 25μl volume. Mice were sacrificed 2 days post-challenge and bacterial
colonization in the lungs were determined. Dashed line represents lower limits of
CFU detection. Black bars represent mean colonization of respective group.
Unpaired two-tailed Student t test was used to determine statistical significance.
Asterisks indicate the P value of ≤0.05.
132

0
1
2
3
4
5
6
7
8
9Lo
g 10 C
FU
RB
6-8C
5
PBS Treatment
* BcfA hyperimmune serum
Naïve rat serum
133

bacteria in the lungs as compared to those that received the naïve rat serum (Fig.
6). In contrast, neutropenic mice (RB6-8C5-injected) that received BcfA-specific
sera harbored comparatively greater numbers of bacteria in the lungs. These mice
harbored greater than 1000-fold more CFU in the lungs than those observed in
PBS-treated control mice. As expected from previous results (Figs. 4A and B and
(28), neutropenic mice, which received naïve rat serum also harbored high bacterial
burdens in the lungs (Fig. 6). The failure of adoptively transferred BcfA-specific sera
to provide protection against B. bronchiseptica challenge in neutropenic mice
suggests that neutrophils are required for anti-BcfA antibody-mediated clearance of
B. bronchiseptica from the lower respiratory tract.
BcfA induces the production of high levels of IFN-γ in ex-vivo stimulated
splenocytes. In order to characterize the cellular responses stimulated by BcfA
vaccination, we examined BcfA-induced cytokine production by splenocytes ex
vivo. Groups of 5 mice received two successive (at three weeks interval) doses of
either 30μg of BcfA adsorbed to alum or alum alone. Two and 4 weeks following the
administration of the second dose, spleens were excised, processed as described
in the Materials and Methods followed by stimulation with 10μg of BcfA. The culture
supernatant was collected and analyzed for the production of IFN-γ (a prototype
Th1 cytokine), IL-4 (a prototype Th2 cytokine) IL-10, IL-12 and TNF-α. BcfA-
stimulated splenocytes, collected two weeks after immunization with BcfA, secreted
high amounts of IFN-γ and low levels of IL-4 (Fig. 7, left panel). Four weeks post-
immunization with BcfA, there was ≈ 3-fold increase in the production of IFN-γ by
134

splenocytes exposed to BcfA (Fig. 7B). However, IL-4 was not detected. Very little
IL-10, IL-12 or TNF-α were detected in the cultures of splenocytes from BcfA-
immunized mice incubated with BcfA. Re-stimulation of spleen cells from mice
infected with alum alone produced either very low or undetectable levels of all the
cytokines tested (Fig. 7). These data are consistent with the conclusion that
immunization with BcfA elicits a Th1-biased immune response with high levels of
IFN-γ production, which has been previously reported to contribute to efficient
clearance of B. bronchiseptica infection (19, 37).
BcfA expression is prevalent in clinical isolates of B. bronchiseptica.
The sequenced laboratory strains and clinical isolates of the three classical
Bordetella spp. vary in the expression of different virulence factors (4, 16). In order
to strengthen the utility of BcfA-based therapeutics for treatment of B.
bronchiseptica infections, it is critical to determine if BcfA is expressed by
circulating clinical isolates of B. bronchiseptica. We performed SDS-PAGE and
immunoblot analyses on the membrane fractions of a number of B. bronchiseptica
strains isolated from a variety of animal species. Using the affinity-enriched anti-
BcfA serum, we detected the presence of a protein band in all the B. bronchiseptica
strains tested here, which corresponded to the BcfA protein from RB50 (Fig. 8).
135

FIG. 7. BcfA-induced production of IFN-γ and IL-4 in splenocytes. Mice were
immunized at 0 and 3 weeks with either 30μg of BcfA adsorbed to alum or 50μg of
alum only. Two and four weeks post-immunization, splenocytes were harvested and
re-stimulated with 10μg of purified BcfA protein. Supernatants were harvested 3
days post-stimulation and analyzed for IFN-γ and IL-4 production using respective
ELISA kits. Re-stimulations were carried out in triplicates for each individual mouse
and mean of the value from individual mice is represented with ± standard
deviations. Unpaired two-tailed Student t test was performed to determine statistical
significance. Asterisks indicate the P value of ≤0.0005.
136

0
4000
8000
12000
16000
20000
Tota
l pg BcfA
Alum
IFN-γ IL-4 IFN-γ IL-4 2 weeks 4 weeks
***
***
137

FIG. 8. Expression of BcfA among clinical isolates of B. bronchiseptica. Membrane
fractions of respective strains were prepared as described in the Materials and
Methods, subjected to SDS-PAGE followed by immunoblot analyses utilizing anti-
BcfA serum. For MBORD628, the amount of protein loaded corresponds to 0.03X of
the amount loaded for RB50.
138

139

DISCUSSION
There is an urgent need to search for new less-virulent or less-aggressive
vaccines for B. bronchiseptica. Similar to the switch from whole cell vaccines to
cell-free vaccines for B. pertussis, there exists a strong rationale for the use of
acellular vaccines containing defined antigens for B. bronchiseptica. A prerequisite
towards development of these vaccines is a complete understanding of the
antigens which elicit a protective immune response. In this report, we have
examined the protective efficacy of the protein antigen, BcfA in a murine respiratory
challenge model of B. bronchiseptica. Immunization of mice with purified BcfA
resulted in lowering of bacterial burdens as early as 1 day post-challenge and
complete clearance of B. bronchiseptica infection from the lower respiratory tract 7
days subsequent to challenge.
The observed protection due to immunization with BcfA appeared to be
mediated by antibodies specific to BcfA. Active immunization with BcfA resulted in
the production of high titers of IgG and IgM anti-BcfA antibodies in the sera of mice,
suggesting the induction of a strong systemic response. In addition, IgG to BcfA
were also detected in the lungs after challenge with RB50. Although it is possible
that the lung IgG may be derived from systemic IgG reaching the lungs by passive
diffusion, the presence of BcfA-specific antibodies in the lungs is important, since it
shows the generation of a localized antibody response in the actual site of B.
bronchiseptica colonization. Moreover, the BcfA immune serum but not the pre-
immune serum protected mice in the passive immunization and challenge
experiments. While adoptive transfer of immune serum from mice infected or
140

vaccinated with B. bronchiseptica can clear bacteria from the lower respiratory tract
(18), there is limited data available on the efficacy of antibodies against individual
protein components in protection against B. bronchiseptica infections. Strikingly, we
observed that the hyperimmune serum generated against purified BcfA was as
efficient as anti-Bordetella antibodies in mediating this protection.
We found that anti-BcfA antibodies of IgG2a and IgG1 subclasses were
either produced in mice as a result of vaccination with BcfA or were the major
antibody isotypes present in the hyperimmune serum. Although the exact
mechanism by which different IgG subclasses protect against B. bronchiseptica is
not clear, in general these antibodies induce efficient opsonization and
phagocytosis of the bacteria. Indeed, we found that anti-BcfA serum led to greater
internalization of B. bronchiseptica cells by macrophages. These data thus suggest
that one of the mechanisms of BcfA-mediated protection is by induction of large
amounts of specific antibodies with high opsonic activity which leads to enhanced
phagocytosis.
In addition to the role of antibodies, we also investigated the contribution of
cell-mediated immunity in BcfA-induced protection. Immunization with BcfA
preferably induced a Th1 response with high IFN-γ production. Production of this
cytokine has previously been shown to be important for clearance of B.
bronchiseptica (19, 37). The dominant Th1 pattern observed due to vaccination with
BcfA was also consistent with the titers of major BcfA-specific IgG isotypes. In all of
the vaccinated mice, IgG2a was detected, whereas IgG1 was the major isotype in
the hyperimmune BcfA-specific rat serum. Although the correlation between
141

induction of different IgG subclasses and Th1/Th2 activity has not been extensively
characterized, IgG2a in the mouse and IgG1 in the rat are Th1 cytokine driven
antibody subclass (7, 10, 36). Very little is known about specific cellular immune
responses generated as a result of vaccination with B. bronchiseptica proteins.
However, our results demonstrating the induction of a Th1 response by B.
bronchiseptica BcfA is different from the Th2 response induced by component
vaccines against B. pertussis. In contrast to acellular vaccines, infection with B.
bronchiseptica and B. pertussis and immunization with the respective whole cell
vaccines induces a Th1 type response (1, 33, 35, 40). CD4+Th1 cells have been
demonstrated to result in more efficient immunity to B. pertussis compared to CD4+
Th2 cells (1). Induction of a Th1 type response and IFN-γ production as a
consequence of BcfA immunization might explain the superior protective efficacy of
this protein even when administered as a single antigen.
An important prerequisite towards the development of an individual protein-
based therapeutics for treatment of bacterial infections is the ubiquitous
demonstration of its expression among circulating isolates. We have demonstrated
that BcfA is expressed by a wide variety of B. bronchiseptica strains isolated from
multiple animal species. These data further strengthen the utility of BcfA as a
potential vaccine candidate. Previous studies demonstrated that individual
immunization with B. pertussis antigens such as Fha, Pertactin and Pertussis toxin
imparts reasonable protective efficacy. However, vaccines containing multiple
components lead to enhanced protection compared to monocomponent vaccines
(35). Similarly, we believe that despite the remarkable efficacy of BcfA, a
142

multivalent vaccine containing other known Bordetella antigens such as Fha,
Pertactin or Fimbriae will be more efficacious. Future studies will have to address
whether immunization with BcfA alone or with composite vaccines will protect
against large animals like pigs and dogs, in which B. bronchiseptica causes
disease. Finally, the broader potential utility of BcfA as a vaccine candidate is
highlighted by our preliminary results which demonstrate the presence of
polypeptides that cross-react with anti-BcfA serum in the closely related human
pathogens B. pertussis and B. parapertussis (our unpublished results). We are
currently investigating whether these bands correspond to orthologs of BcfA and
whether BcfA can confer protection against these human adapted species. A
positive outcome will be highly significant and will promote the inclusion of BcfA in
the current acellular vaccines for B. pertussis.
143

FIG. 9. Model for BcfA-mediated protective immunity.
Based on the in vivo and in vitro analyses described here, we propose a model for
BcfA-induced immune clearance of B. bronchiseptica. We believe that immunization
with BcfA elicits specific antibodies, which have the ability to enhance opsonization
and phagocytosis of B. bronchiseptica. Findings with neutropenic mice and
opsonophagocytosis assays using J774 macrophage cells reveal a role for
neutrophils and macrophages in the subsequent phagocytosis and clearance of
Bordetella. Also our data demonstrate the induction of IFN-γ and Th1 type responses
in BcfA primed splenocytes. Therefore, we propose that apart from antibodies, BcfA-
immunization also results in the induction of CD 4+ Th1 cells, which is vital for
protective immunity to Bordetella.
144

Challenge with Bordetella
Immunization with BcfA
T
IFNγ
B
Antibodies Phagocytosis
Ciliated Epithelial cells of Respiratory tract
B T
145

ACKNOWLEDGMENTS
We thank Drs. Dan Wozniak and Steven Mizel for critical reading of this manuscript.
We thank Drs. T. Nicholson and K. Register for the different clinical isolates and Dr.
G. Huffnagle for a gift of RB6-8C5 hybridoma cells. Research in the laboratory of
R.D. is supported by funds from National Research Initiative of the USDA
Cooperative State Research, Education and Extension Service (grant 35604-16874)
and the NIH (grant R21AI071054 and 1R01AI075081).
146

REFERENCES
1. Barnard, A., B. P. Mahon, J. Watkins, K. Redhead, and K. H. Mills. 1996. Th1/Th2 cell dichotomy in acquired immunity to Bordetella pertussis: variables in the in vivo priming and in vitro cytokine detection techniques affect the classification of T-cell subsets as Th1, Th2 or Th0. Immunology 87:372-80. 2. Bey, R. F., F. J. Shade, R. A. Goodnow, and R. C. Johnson. 1981. Intranasal vaccination of dogs with liver avirulent Bordetella bronchiseptica: correlation of serum agglutination titer and the formation of secretory IgA with protection against experimentally induced infectious tracheobronchitis. Am. J. Vet. Res. 42:1130-2. 3. Brockmeier, S. L., and K. B. Register. 2007. Expression of the dermonecrotic toxin by Bordetella bronchiseptica is not necessary for predisposing to infection with toxigenic Pasteurella multocida. Vet. Microbiol.125: 284-9 4. Buboltz, A. M., T. L. Nicholson, M. R. Parette, S. E. Hester, J. Parkhill, and E. T. Harvill. 2008. Replacement of adenylate cyclase toxin in a lineage of Bordetella bronchiseptica. J. Bacteriol. 190:5502-11. 5. Buonavoglia, C., and V. Martella. 2007. Canine respiratory viruses. Vet. Res. 38:355-73. 6. Carbonetti, N. H. 2007. Immunomodulation in the pathogenesis of Bordetella pertussis infection and disease. Curr. Opin. Pharmacol. 7:272-8. 7. Cazorla, S. I., P. D. Becker, F. M. Frank, T. Ebensen, M. J. Sartori, R. S. Corral, E. L. Malchiodi, and C. A. Guzman. 2008. Oral vaccination with Salmonella enterica as a cruzipain-DNA delivery system confers protective immunity against Trypanosoma cruzi. Infect. Immun. 76:324-33. 8. Chanter, N., T. Magyar, and J. M. Rutter. 1989. Interactions between Bordetella bronchiseptica and toxigenic Pasteurella multocida in atrophic rhinitis of pigs. Res. Vet. Sci. 47:48-53. 9. Cotter, P. A., and J. F. Miller. 1997. A mutation in the Bordetella bronchiseptica bvgS gene results in reduced virulence and increased resistance to starvation, and identifies a new class of Bvg-regulated antigens. Mol. Microbiol. 24:671-85. 10. Cuturi, M. C., R. Josien, D. Cantarovich, L. Bugeon, I. Anegon, S. Menoret, H. Smit, P. Douillard, and J. P. Soulillou. 1994. Decreased anti-donor major histocompatibility complex class I and increased class II alloantibody response in allograft tolerance in adult rats. Eur. J. Immunol. 24:1627-31.
147

11. Czuprynski, C. J., J. F. Brown, N. Maroushek, R. D. Wagner, and H. Steinberg. 1994. Administration of anti-granulocyte mAb RB6-8C5 impairs the resistance of mice to Listeria monocytogenes infection. J. Immunol. 152:1836-46. 12. Deeb, B. J., R. F. DiGiacomo, B. L. Bernard, and S. M. Silbernagel. 1990. Pasteurella multocida and Bordetella bronchiseptica infections in rabbits. J. Clin. Microbiol. 28:70-5. 13. Dugal, F., M. Belanger, and M. Jacques. 1992. Enhanced adherence of Pasteurella multocida to porcine tracheal rings preinfected with Bordetella bronchiseptica. Can. J. Vet.Res. 56:260-4. 14. Dworkin, M. S., P. S. Sullivan, S. E. Buskin, R. D. Harrington, J. Olliffe, R. D. MacArthur, and C. E. Lopez. 1999. Bordetella bronchiseptica infection in human immunodeficiency virus-infected patients. Clin. Infect. Dis. 28:1095-9. 15. Elias, B., M. Albert, S. Tuboly, and P. Rafai. 1992. Interaction between Bordetella bronchiseptica and toxigenic Pasteurella multocida on the nasal mucosa of SPF piglets. J. Vet. Med. Sci. 54:1105-10. 16. Fennelly, N. K., F. Sisti, S. C. Higgins, P. J. Ross, H. van der Heide, F. R. Mooi, A. Boyd, and K. H. Mills. 2008. Bordetella pertussis expresses a functional type III secretion system that subverts protective innate and adaptive immune responses. Infect. Immun. 76:1257-66. 17. Goodnow, R. A. 1980. Biology of Bordetella bronchiseptica. Microbiol. Rev. 44:722-38. 18. Gopinathan, L., G. S. Kirimanjeswara, D. N. Wolfe, M. L. Kelley, and E. T. Harvill. 2007. Different mechanisms of vaccine-induced and infection-induced immunity to Bordetella bronchiseptica. Microbes Infect. 9:442-8. 19. Gueirard, P., P. Minoprio, and N. Guiso. 1996. Intranasal inoculation of Bordetella bronchiseptica in mice induces long-lasting antibody and T-cell mediated immune responses. Scand. J. Immunol. 43:181-92. 20. Gueirard, P., C. Weber, A. Le Coustumier, and N. Guiso. 1995. Human Bordetella bronchiseptica infection related to contact with infected animals: persistence of bacteria in host. J. Clin. Microbiol. 33:2002-6. 21. Harvill, E. T., P. A. Cotter, and J. F. Miller. 1999. Pregenomic comparative analysis between Bordetella bronchiseptica RB50 and Bordetella pertussis tohama I in murine models of respiratory tract infection. Infect. Immun. 67:6109-18. 22. Heininger, U., K. Stehr, S. Schmitt-Grohe, C. Lorenz, R. Rost, P. D. Christenson, M. Uberall, and J. D. Cherry. 1994. Clinical characteristics of illness
148

caused by Bordetella parapertussis compared with illness caused by Bordetella pertussis. Pediatr. Infect. Dis. J. 13:306-9. 23. Herring, A. C., N. R. Falkowski, G. H. Chen, R. A. McDonald, G. B. Toews, and G. B. Huffnagle. 2005. Transient neutralization of tumor necrosis factor alpha can produce a chronic fungal infection in an immunocompetent host: potential role of immature dendritic cells. Infect. Immun. 73:39-49. 24. Hewlett, E. L., G. M. Donato, and M. C. Gray. 2006. Macrophage cytotoxicity produced by adenylate cyclase toxin from Bordetella pertussis: more than just making cyclic AMP! Mol. Microbiol. 59:447-59. 25. Huang, X. Z., and L. E. Lindler. 2004. The pH 6 antigen is an antiphagocytic factor produced by Yersinia pestis independent of Yersinia outer proteins and capsule antigen. Infect. Immun. 72:7212-9. 26. Huebner, E. S., B. Christman, S. Dummer, Y. W. Tang, and S. Goodman. 2006. Hospital-acquired Bordetella bronchiseptica infection following hematopoietic stem cell transplantation. J. Clin. Microbiol. 44:2581-3. 27. Kirimanjeswara, G. S., P. B. Mann, and E. T. Harvill. 2003. Role of antibodies in immunity to Bordetella infections. Infect. Immun. 71:1719-24. 28. Kirimanjeswara, G. S., P. B. Mann, M. Pilione, M. J. Kennett, and E. T. Harvill. 2005. The complex mechanism of antibody-mediated clearance of Bordetella from the lungs requires TLR4. J. Immunol. 175:7504-11. 29. Lopez, S., A. J. Marco, N. Prats, and C. J. Czuprynski. 2000. Critical role of neutrophils in eliminating Listeria monocytogenes from the central nervous system during experimental murine listeriosis. Infect. Immun. 68:4789-91. 30. Mann, P., E. Goebel, J. Barbarich, M. Pilione, M. Kennett, and E. Harvill. 2007. Use of a genetically defined double mutant strain of Bordetella bronchiseptica lacking adenylate cyclase and type III secretion as a live vaccine. Infect. Immun. 75:3665-72. 31. Mattoo, S., and J. D. Cherry. 2005. Molecular pathogenesis, epidemiology, and clinical manifestations of respiratory infections due to Bordetella pertussis and other Bordetella subspecies. Clin. Microbiol. Rev. 18:326-82. 32. McArthur, J. D., N. P. West, J. N. Cole, H. Jungnitz, C. A. Guzman, J. Chin, P. R. Lehrbach, S. P. Djordjevic, and M. J. Walker. 2003. An aromatic amino acid auxotrophic mutant of Bordetella bronchiseptica is attenuated and immunogenic in a mouse model of infection. FEMS Microbiol. Lett. 221:7-16.
149

33. McGuirk, P., and K. H. Mills. 2002. Pathogen-specific regulatory T cells provoke a shift in the Th1/Th2 paradigm in immunity to infectious diseases. Trends Immunol. 23:450-5. 34. Miller, D. L., E. M. Ross, R. Alderslade, M. H. Bellman, and N. S. Rawson. 1981. Pertussis immunisation and serious acute neurological illness in children. Br. Med. J. (Clin. Res. Ed.) 282:1595-9. 35. Mills, K. H., M. Ryan, E. Ryan, and B. P. Mahon. 1998. A murine model in which protection correlates with pertussis vaccine efficacy in children reveals complementary roles for humoral and cell-mediated immunity in protection against Bordetella pertussis. Infect. Immun. 66:594-602. 36. Philips, J. R., W. Brouwer, M. Edwards, S. Mahler, J. Ruhno, and A. M. Collins. 1999. The effectiveness of different rat IgG subclasses as IgE-blocking antibodies in the rat basophil leukaemia cell model. Immunol. Cell. Biol. 77:121-6. 37. Pilione, M. R., and E. T. Harvill. 2006. The Bordetella bronchiseptica type III secretion system inhibits gamma interferon production that is required for efficient antibody-mediated bacterial clearance. Infect. Immun. 74:1043-9. 38. Porter, J. F., K. Connor, and W. Donachie. 1994. Isolation and characterization of Bordetella parapertussis-like bacteria from ovine lungs. Microbiology 140 ( Pt 2):255-61. 39. Rath, B. A., K. B. Register, J. Wall, D. M. Sokol, and R. B. Van Dyke. 2008. Persistent Bordetella bronchiseptica pneumonia in an immunocompetent infant and genetic comparison of clinical isolates with kennel cough vaccine strains. Clin. Infect. Dis. 46:905-8. 40. Redhead, K., J. Watkins, A. Barnard, and K. H. Mills. 1993. Effective immunization against Bordetella pertussis respiratory infection in mice is dependent on induction of cell-mediated immunity. Infect. Immun. 61:3190-8. 41. Ross, P. J., E. C. Lavelle, K. H. Mills, and A. P. Boyd. 2004. Adenylate cyclase toxin from Bordetella pertussis synergizes with lipopolysaccharide to promote innate interleukin-10 production and enhances the induction of Th2 and regulatory T cells. Infect. Immun. 72:1568-79. 42. Schlageter, A. M., and T. R. Kozel. 1990. Opsonization of Cryptococcus neoformans by a family of isotype-switch variant antibodies specific for the capsular polysaccharide. Infect. Immun. 58:1914-8. 43. Sebaihia, M., A. Preston, D. J. Maskell, H. Kuzmiak, T. D. Connell, N. D. King, P. E. Orndorff, D. M. Miyamoto, N. R. Thomson, D. Harris, A. Goble, A. Lord, L. Murphy, M. A. Quail, S. Rutter, R. Squares, S. Squares, J. Woodward,
150

J. Parkhill, and L. M. Temple. 2006. Comparison of the genome sequence of the poultry pathogen Bordetella avium with those of B. bronchiseptica, B. pertussis, and B. parapertussis reveals extensive diversity in surface structures associated with host interaction. J. Bacteriol. 188:6002-15. 44. Sloan, G. P., C. F. Love, N. Sukumar, M. Mishra, and R. Deora. 2007. The Bordetella Bps Polysaccharide is Critical for Biofilm Development in the Mouse Respiratory Tract. J. Bacteriol.189: 750-760. 45. Spears, P. A., L. M. Temple, D. M. Miyamoto, D. J. Maskell, and P. E. Orndorff. 2003. Unexpected similarities between Bordetella avium and other pathogenic Bordetellae. Infect. Immun. 71:2591-7. 46. Spilker, T., A. A. Liwienski, and J. J. LiPuma. 2008. Identification of Bordetella spp. in respiratory specimens from individuals with cystic fibrosis. Clin. Microbiol. Infect. 14:504-6. 47. Stainer, D. W., and M. J. Scholte. 1970. A simple chemically defined medium for the production of phase I Bordetella pertussis. J.Gen. Microbiol. 63:211-20. 48. Stevenson, A., and M. Roberts. 2002. Use of a rationally attenuated Bordetella bronchiseptica as a live mucosal vaccine and vector for heterologous antigens. Vaccine 20:2325-35. 49. Stevenson, A., and M. Roberts. 2003. Use of Bordetella bronchiseptica and Bordetella pertussis as live vaccines and vectors for heterologous antigens. FEMS Immunol. Med. Microbiol. 37:121-8. 50. Sukumar, N., M. Mishra, G. P. Sloan, T. Ogi, and R. Deora. 2007. Differential Bvg phase-dependent regulation and combinatorial role in pathogenesis of two Bordetella paralogs, BipA and BcfA. J. Bacteriol. 189:3695-704. 51. West, N. P., H. Jungnitz, J. T. Fitter, J. D. McArthur, C. A. Guzman, and M. J. Walker. 2000. Role of phosphoglucomutase of Bordetella bronchiseptica in lipopolysaccharide biosynthesis and virulence. Infect. Immun. 68:4673-80. 52. Woolfrey, B. F., and J. A. Moody. 1991. Human infections associated with Bordetella bronchiseptica. Clin. Microbiol. Rev. 4:243-55.
151

CHAPTER IV: DISCUSSION
The mammalian respiratory tract is equipped with specialized mutltitiered
defense mechanisms to prevent the entry and infection by pathogens. The common
basic defenses at this site include mechanical barriers such as the mucociliatory
escalator consisting of the ciliated epithelium and innate factors such as defensins,
lysozyme and phagocytic cells. If these are compromised, specific adaptive immunity
becomes operational. Despite the presence of sophisticated pulmonary host defense
systems, the respiratory tract acts as the main port of entry for many infectious
agents. These include several pathogenic bacteria that have evolved mechanisms to
overcome the protective host responses present in the respiratory tract and
successfully colonize and exert pathological infections. Understanding the nature of
bacterial – host interactions in the respiratory tract is of interest not only to study the
mechanisms used by pathogens to evade host immune responses, but also for
designing therapeutic and prophylactic strategies. This is highly relevant because
pathogens of the respiratory tract are the foremost cause of mortality and morbidity in
humans and animals. One of the major respiratory pathogens in agriculturally
important, food producing and pet animals is the Gram-negative bacterium Bordetella
bronchiseptica. In the studies described here, we have identified and characterized a
novel virulence factor of Bordetella spp, Bordetella colonization factor A (BcfA). We
show that BcfA is a Bvg-regulated outer membrane protein and that a strain lacking
bcfA, along with its paralog bipA is defective in colonization of the rat trachea.
Additionally, we show that BcfA-specific antibodies are produced during a wild type
152

Bordetella bronchiseptica infection. Subsequently, we describe studies demonstrating
the efficacy of BcfA as a protective antigen against B. bronchiseptica infections. Our
results strongly suggest that both active and passive immunization with BcfA provides
protection against subsequent challenge with B. bronchiseptica in a mouse model of
infection. Also, through ELISAs, we show that BcfA specific antibodies are produced
after immunization, with Ig2a being the predominant isotype. Furthermore,
investigation of the mechanistic basis of BcfA-induced immunity revealed a role for
neutrophils and macrophages in phagocytosis. Finally we show that immunization
with BcfA induces a Th1 type response in splenocytes leading to IFN-γ production,
which has been previously shown to aid clearance of Bordetella infections.
Potential roles of BipA and BcfA in mediating respiratory tract colonization
by Bordetella.
Our results demonstrate that individual deletion of either bipA or bcfA does not
have any significant effect on respiratory tract colonization by Bordetella. Strikingly,
we have found that concomitant deletion of bipA and bcfA results in a defect in
colonization of the rat trachea, thereby suggesting a potential overlapping role for
these two proteins in Bordetella pathogenesis. Although our data demonstrate a
prerequisite for BipA and BcfA in B. bronchiseptica colonization, the exact function of
these two paralogs is unknown. A crucial and initial step in colonization is the
attachment of bacteria to host cells. Therefore, one possibility is that these proteins
function as adhesins and mediate attachment of Bordetella to epithelial cells of
respiratory tract (Fig. 1A). Future studies evaluating the attachment efficiency of the
153

individual deletion strains, the double deletion ΔbipA-ΔbcfA strain and the wt strain to
tissue culture cell lines and cells directly acquired from respiratory tissues of animals
will facilitate addressing this hypothesis.
The respiratory tract is equipped with a multitude of innate and adaptive
immune mechanisms to efficiently eliminate pathogens. The mucociliatory escalator
is an effective process to clear pathogenic organisms from the respiratory tract and
this is complemented by other innate defenses such as neutrophils and macrophages
and adaptive immune responses. Therefore, another significant prerequisite for
successful colonization by Bordetella, subsequent to adherence, is to evade these
host defenses. Numerous pathogens including Bordetella spp. induce anti-
inflammatory immune responses to enhance their persistence (25). Since the ΔbipA-
ΔbcfA strain demonstrated a defective colonization of the rat trachea, another
persuasive hypothesis is that BipA and BcfA are involved in host immune
suppression and thus deletion of these proteins might make the ΔbipA-ΔbcfA strain
susceptible to enhanced immunological clearance. We speculate that BipA and BcfA
may diminish clearance of B. bronchiseptica from the trachea by modulating either
innate or adaptive immune responses including adherence to and phagocytosis by
immune cells such as neutrophils and macrophages, inhibition of the complement
system, altered cytokine and chemokine secretion or antibody production (Fig. 1B).
The role of BipA and BcfA in altering cytokine production as well as adherence and
phagocytosis can be evaluated in vitro by utilizing the epithelial cell lines (L2, Hep2)
or immune cell-derived lines (J774, RAW or isolated macrophages, neutrophils)
infected with either wt or single or double mutant ΔbipA-ΔbcfA strains. To address the
154

roles of these proteins in affecting components of immunity in vivo, the well
characterized mouse model of infection as well as mice defective in specific immune
responses can be utilized (17). Differences between the mutant and the wt strains in
the degree of inflammation in the lungs, recruitment of inflammatory cells, cytokine
production profile and eliciting serum antibody responses can be evaluated using
these models.
In the low volume-low inoculum rat colonization model, the defect in
colonization of the ΔbipA-ΔbcfA strain was surprisingly restricted to only the lower
respiratory tract. The failure to colonize the trachea and not the nasal cavity is a
phenotype common to several Bordetella mutants including strains deficient in FHA,
Cya and Fim (11, 22). While these data may provide evidence for stricter
requirements to colonize trachea as compared to the nasal cavity, it is important to
note that the model of infection that we adopted, utilized low volume (5 μl) to deliver
the inoculum dosage. It has been previously demonstrated that in this model,
Bordetella initially colonizes the nasal cavity from which the bacteria seeds and
colonizes the lower respiratory tract (1). Therefore, the lack of tracheal colonization
in the absence of BipA and BcfA could also be due to failure of Bordetella to traffic
from the nasal cavity to the trachea (Fig. 1C). If this is true, then the wt and the
ΔbipA-ΔbcfA strains will be indistinguishable in their ability to colonize the trachea in
a high volume model of intranasal infection (50 μl) or when delivered through the
intratracheal inoculation route, both of which directly deposit bacteria into trachea.
155

FIG. 1. Potential roles of BipA and BcfA in B. bronchiseptica pathogenesis.
A. BipA and BcfA as adhesins mediating attachment of Bordetella to the respiratory
epithelium.
B. BipA and BcfA as immunomodulators causing the suppression of the host immune
responses that are responsible for the clearance of Bordetella.
C. The lack of colonization of the trachea and not the nasal cavity by the ΔbipA-ΔbcfA
strain in a low volume-low inoculum dosage model of respiratory colonization might
also be due to the inefficient seeding of this strain from the nasal cavity to the
trachea. Therefore, another possibility is that BipA and BcfA function as mediators of
trafficking Bordetella from the upper respiratory tract to the lower respiratory tract.
156

A B
Attachment
Ciliated Epithelial cells of trachea
Immunomodulation Trafficking
T B
T
B
Ciliated Epithelial cells of
Nasal Septum
Ciliated Epithelial cells of trachea
trachea
NeutrophilBordetella BipA B B-cell T
C
BcfA T cell
Macrophage
157

Previous studies have shown the differential expression of bacterial virulence
factors
Evidently, all the proposed functions of BipA and BcfA are not mutually
exclus
BvgAS-mediated regulation of bcfA expression.
signal transduction system that
in a host tissue specific and time dependent manner with the progression of
the infectious cycle. Therefore, we cannot exclude the possibility that BipA and BcfA
are expressed only in the lower respiratory tract in a tissue specific manner at the
time points that we investigated. Also B. bronchiseptica causes prolonged
colonization of the nasal cavity despite clearance from the lower respiratory tract. We
and others have observed that B. bronchiseptica can be isolated from the nasal
cavities of infected animals for at least 270 days (15). Our studies were restricted to
12 and 30 days post inoculation, consequently, the possibility of contribution of BipA
and BcfA to nasal cavity colonization at further stages of infection cannot be ruled
out.
ive; in fact it is highly likely that these proteins may have multiple effector
mechanisms. This hypothesis is in agreement with several other studies investigating
the role of Bordetella factors such as FHA, PT and CyaA. It has been demonstrated
that all these factors have multiple functions in vivo including adherence and host
immunomodulation.
BvgAS is the master regulatory two component
controls the expression of the majority of the virulence factors of Bordetella spp (8).
Given the importance of the BvgAS system, our finding that expression of bcfA is
regulated by this system correlates with its role in Bordetella pathogenicity. The
158

BvgAS – mediated regulation of bcfA is evident by RT-PCR assays as well as by the
presence of multiple sequence elements resembling the consensus BvgA binding site
in the regions upstream of the bcfA ORF. Furthermore, EMSA demonstrates the
direct binding of purified BvgA to the putative bcfA promoter. Although EMSAs clearly
demonstrate the specific binding of BvgA to the bcfA promoter region, they do not
reveal the relative occupancy of individual binding sites. Thus, we can postulate at
least two models to explain the formation of higher order complexes with increasing
concentration of BvgA-P - i) With increasing concentration there is more binding of
BvgA-P dimers to a single binding site or ii) At higher concentrations multiple binding
sites are occupied by BvgA-P (Fig. 2). One of the striking features common to all of
the characterized BvgA-activated promoters is the presence of multiple BvgA binding
sites (2, 13, 26). Hence, this fact along with analysis of the expression profile of bcfA
and identification of several inverted heptads, we speculate that the bcfA promoter
will harbor multiple BvgA binding sites. We are currently conducting DNase I foot
printing assays to further delineate the above mentioned scenarios. Knowledge of
the nature and organization of BvgA binding sites would also enable us to predict the
specific mechanism of Bvg-mediated regulation of phase- specific expression of bcfA
(12). We believe that our studies along with others will provide new insights into how
a single virulence regulatory system controls the expression of a multitude of
virulence factors. Furthermore, these studies will provide the foundation to devise
strategies geared towards disrupting this regulatory circuitry as a preventive measure
against Bordetella pathogenesis. We believe that the complexity in the gene
expression states exhibited by the Bvg regulon may not be
159

FIG. 2. Model illustrating relative occupancy of phosphorylated BvgA (BvgA-P) to the
putative bcfA promoter.
Higher order complex formation seen in Electrophoretic Mobility Shift Assays with
increasing concentrations of BvgA-P may be a resultant of multiple BvgA-P dimers
binding to a single binding site (A) or though occupation of multiple binding sites (B).
160

+1
BvgA-P
BvgA-P
+1
0 0BvgA BvgA-P
BvgA-P
A B
161

unique and other pathogenic bacteria will show similar patterns of gene regulation.
Therefore, results from our studies will serve as a model system for understanding
the dynamics of virulence gene expression in other pathogens.
Differential phase specific expression profile of BipA and BcfA.
Previous studies have shown that Bvg+ phase is sufficient and necessary for
establishing respiratory tract infections and that the Bvgi phase displays decreased
ability to colonize the host (9, 10). Both real-time RT-PCR and β-galactosidase
assays indicate that despite their homology, surprisingly BipA and BcfA have distinct
phase specific expression profiles. Our studies demonstrate that BipA is expressed
only in the Bvgi phase while BcfA is expressed both in the Bvg+ and Bvgi phases.
However, our rat colonization model provides evidence for overlapping functions for
these proteins, since only the ΔbipA-ΔbcfA strain is defective in colonization of the
trachea. These seemingly contrasting observations may be explained on the basis
that our phase specific expression data are restricted to the transcript levels under in
vitro conditions and do not define protein levels generated during an infection. It is
also possible that low level of expression of BipA in the Bvg+ phase is sufficient for
Bordetella colonization. Nevertheless, future studies involving promoter exchange
between BipA and BcfA would address the in vivo significance of the differential
expression pattern of these paralogs.
162

BcfA as a vaccine candidate against B. bronchiseptica.
Despite the availability of a natural mouse infection model as well as other well
characterized animal models, there is only limited focus on the development as well
as efficacy evaluations of vaccines against B. bronchiseptica. An effective vaccination
regimen which provides long lasting protective immunity can easily limit the spread of
this pathogen both among animals in a herd as well as from animals to humans (29).
Whole cell vaccines consisting of inactivated or attenuated strains are currently being
administered to animals, although there is limited information on the safety, efficacy
and stability of these strains. Genetic attenuation of some of the vaccine strains is
still unknown, and there is increasing focus on the use of genetically defined mutants
as live vaccines (28, 29). While these studies reveal a great potential for defined
attenuated vaccine strains in clearing and protecting animals against a lethal B.
bronchiseptica infection, they fail to report the reversion rate of these mutants.
Moreover, in the murine model of infection, in which these strains are tested, bacteria
colonize the upper respiratory tract for the life of the animals (21, 27). Persistent
colonization of the upper respiratory tract of the vaccinated animals by B.
bronchiseptica might create a reservoir of infectious bacteria from which other
animals and even zoonotic infections can occur. Lack of clearance from the
respiratory tract combined with the possibility of reversion of these strains to a more
virulent form emphasizes the need for the development of superior vaccines. In the
context of these observations, our studies demonstrating the remarkable
immunogenicity of a single protein, BcfA, raise the novel concept of employing
acellular vaccines containing defined antigens against B. bronchiseptica. We propose
163

that BcfA in combination with other protective antigens will be a safer and more
efficient vaccine.
Although active immunization with BcfA had striking effects on colonization of
B. bronchiseptica in the lower respiratory tract, it only slightly lowered the bacterial
numbers in the upper respiratory tract. Likewise, passive immunization also had
minimal effects on nasal cavity colonization. As discussed above, this may be due to
the differences in organ specific requirements for colonization and the immune
responses generated in the lower and upper respiratory tract (18). The majority of the
studies in the Bordetella field are restricted to the mechanism of pathogenesis and
bacterial elimination from the lower respiratory tract. Consequentially, the
characteristics of Bordetella or the host immune system that enable this bacterium to
persist in the upper respiratory tract are still unclear. However, it is it important to note
that in our studies we used the intraperitoneal route to administer the antigen, which
is widely accepted to be a poor inducer of mucosal responses. The failure to induce
mucosal responses is evident in the isotyping data which demonstrates a lack of IgA
production both in immunized mice and rats. Previous reports have demonstrated
that IgA is critical for decreasing B. bronchiseptica infection in the upper respiratory
tract (30). Therefore we believe that, by adopting a distinct route of BcfA
administration, namely intranasal immunization, it may be possible to elicit a local
secretory response in the mucosa resulting in either more drastic reduction or
clearance of Bordetella from the upper respiratory tract.
Our studies evaluating the immunogenicity of BcfA are restricted to only its
vaccine potential. Although our data confirm that immunization with BcfA confer
164

relatively high levels of protection against B. bronchiseptica, it does not address the
therapeutic potential of this protein in clearing an established infection. Future
studies can be devised to verify whether administration of BcfA antigen or anti-BcfA
antibodies 7-10 days post inoculation of B. bronchiseptica can accelerate clearance
in a mouse model of infection.
BcfA as a Th1 response inducing adjuvant.
Another significant new finding of our studies is the induction of Th1 response
by BcfA antigen immunization. Ex vivo restimulation of splenocytes harvested from
immunized mice with BcfA resulted in the induction of IFN-γ and Th1 type responses.
In agreement with these results, we also found that immunization with BcfA induces
predominantly IgG1 in rats and IgG2a in mice, both of which are IFN-γ driven
isotypes. Obviously, these findings are surprising considering other studies which
indicate that B. pertussis protein-based vaccines induce a more adjuvant mediated
Th2 response (16, 24). On the contrary, either infection or immunization with B.
pertussis whole cell vaccines induce Th1 responses (24). Consistent with the well
established role of Th1 cells in protective immunity against Bordetella spp, both
natural infection as well as whole cell vaccine induced immunity is considered to be
superior to acellular vaccine induced immunity (23). To further enhance the efficacy
of acellular vaccines, many studies have proposed the identification and use of
effective adjuvants, which can preferentially induce the more appropriate Th1 cell
subtype. Also of note are studies demonstrating that addition of IL-12 to acellular
vaccine augmented its protective efficacy mainly through priming a Th1 biased
165

response (20). At present we do not have an explanation for why BcfA induces a Th1
response. Further investigations into this property of BcfA will involve testing whether
immunization with BcfA alone in the absence of any adjuvant or in the presence of a
different adjuvant other than alum results in Th1 response and IFN-γ production will
substantiate our observations. We are currently testing a BcfA-flagellin fusion protein
and evaluating whether in combination with a different adjuvant, flagellin, BcfA can
still induce a Th1 response. Positive outcome from this experiment will confirm our
results with BcfA immunizations supplemented with alum as an adjuvant. If the ability
to preferentially induce Th1 responses is inherent to the BcfA antigen and is
independent of the adjuvant used, we propose the use of BcfA as a Th1 promoting
adjuvant for antigen vaccines. However, further investigation into this possibility is
essential.
Our assumption that BcfA induces preferentially a Th1 response is based on
differential cytokine secretion. Detection of high amounts of IFN-γ in ex vivo
restimulated splenocytes from BcfA-immunized mice as compared to alum-
immunized mice, is the basis of our theory. Since we used whole splenocyte
populations in these investigations, we cannot rule out the possible contributions of
other immune cells such as natural killer (NK) cells in IFN-γ production. Future
experiments utilizing purified CD4+ T cells will provide further evidence for induction
of Th1 phenotype on BcfA immunization.
166

BcfA as a vaccine candidate against B. pertussis.
There are a plethora of studies describing the immunogenicity of inactivated
whole cells, individual antigens or antigen combinations against B. pertussis
infections (5, 19, 24). A common premise for many of these studies is the fact that
although there are multiple side effects associated with the whole cells vaccines, they
provide superior protection against a B. pertussis infection as compared to the
currently available acellular vaccines (14). The greater efficacy of whole cell vaccines
may be attributed to the presence of additional antigens and factors that contribute to
adjuvanticity such as LPS, which at high levels has been previously shown to
enhance IL-12 production and elicit Th1 responses. In addition several studies have
demonstrated that five, four, three or even two component vaccines have higher
immunogenicity than monocomponent vaccines (24). Consistent with the lower
efficacy of acellular vaccines, in the past two decades, there has been an increase in
the number of whooping cough cases reported in many of countries including USA
where immunization with pertussis vaccine is routinely carried out (6, 7). Gene
profiling studies have revealed that the currently circulating isolates are constantly
mutating antigens such as Prn and PT which are major components of the acellular
vaccines (3, 4). Consequently, antigens included in the acellular vaccines may fail to
impart protection. These observations signify the requirement to identify new vaccine
candidates. Therefore our studies identifying a novel vaccine candidate is highly
encouraging. Currently we are examining the expression of BcfA in B. pertussis.
Although the open reading frame of bcfA gene is disrupted by an insertion sequence,
our current results from SDS-PAGE and western blot analyses utilizing the BcfA
167

polyclonal serum on whole cell lysates of B. pertussis reveal cross reactive bands
(Fig. 3). Our preliminary data also reveal the expression of BcfA in recent clinical
isolates of B. pertussis. This observation is significant as it shows the conservation of
BcfA among currently circulating host adapted strains. The relevance of this result is
further justified in the light of pathogenomic studies demonstrating that the current
circulating isolates of B. pertussis are losing genetic material continuously and the
lost genetic material appears to be unimportant for virulence and pathogenesis (3).
Further studies are required to investigate whether these bands correspond to
orthologs of BcfA and whether it can confer protection against this human adapted
species. We speculate that inclusion of new antigens such as BcfA may improve the
immunogenic activity of the currently available vaccines.
In conclusion, our studies describe a novel antigen of Bordetella spp, BcfA, its
role in B. bronchiseptica colonization and its utility as immunogen to elicit protective
immune responses. We believe that our studies will be a stimulus for identifying new
vaccine targets and for developing novel therapeutic interventions against Bordetella
infections.
168

FIG. 3. Expression of BcfA by B. pertussis strains. Whole cell lysates of B. pertussis
strains Bp536 (laboratory strain), M3984, S49560 (clinical strains) and B.
bronchiseptica strains RB50 and ΔbcfA were separated by SDS-PAGE and analyzed
by Western blotting with anti-BcfA hyperimmune serum.
169

RB
50
Δbc
fA
S495
60
M39
84
Bp5
36
115 kDa
Clinical Strains B. bronchiseptica
B. pertussis
170

REFERENCES 1. Akerley, B. J., P. A. Cotter, and J. F. Miller. 1995. Ectopic expression of
the flagellar regulon alters development of the Bordetella-host interaction. Cell 80:611-20.
2. Boucher, P. E., and S. Stibitz. 1995. Synergistic binding of RNA
polymerase and BvgA phosphate to the pertussis toxin promoter of Bordetella pertussis. J Bacteriol 177:6486-91.
3. Bouchez, V., V. Caro, E. Levillain, G. Guigon, and N. Guiso. 2008.
Genomic content of Bordetella pertussis clinical isolates circulating in areas of intensive children vaccination. PLoS ONE 3:e2437.
4. Buboltz, A. M., T. L. Nicholson, M. R. Parette, S. E. Hester, J. Parkhill,
and E. T. Harvill. 2008. Replacement of adenylate cyclase toxin in a lineage of Bordetella bronchiseptica. J Bacteriol 190:5502-11.
5. Cherry, J. D. 2007. Immunity to pertussis. Clin Infect Dis 44:1278-9. 6. Cherry, J. D., S. J. Chang, D. Klein, M. Lee, S. Barenkamp, D.
Bernstein, R. Edelman, M. D. Decker, D. P. Greenberg, W. Keitel, J. Treanor, and J. I. Ward. 2004. Prevalence of antibody to Bordetella pertussis antigens in serum specimens obtained from 1793 adolescents and adults. Clin Infect Dis 39:1715-8.
7. Cherry, J. D., and P. Olin. 1999. The science and fiction of pertussis
vaccines. Pediatrics 104:1381-3. 8. Cotter, P. A., and A. M. Jones. 2003. Phosphorelay control of virulence
gene expression in Bordetella. Trends Microbiol 11:367-73. 9. Cotter, P. A., and J. F. Miller. 1994. BvgAS-mediated signal transduction:
analysis of phase-locked regulatory mutants of Bordetella bronchiseptica in a rabbit model. Infect Immun 62:3381-90.
10. Cotter, P. A., and J. F. Miller. 1997. A mutation in the Bordetella
bronchiseptica bvgS gene results in reduced virulence and increased resistance to starvation, and identifies a new class of Bvg-regulated antigens. Mol Microbiol 24:671-85.
11. Cotter, P. A., M. H. Yuk, S. Mattoo, B. J. Akerley, J. Boschwitz, D. A.
Relman, and J. F. Miller. 1998. Filamentous hemagglutinin of Bordetella bronchiseptica is required for efficient establishment of tracheal colonization. Infect Immun 66:5921-9.
171

12. Deora, R. 2002. Differential regulation of the Bordetella bipA gene: distinct roles for different BvgA binding sites. J Bacteriol 184:6942-51.
13. Deora, R. 2004. Multiple mechanisms of bipA gene regulation by the
Bordetella BvgAS phosphorelay system. Trends Microbiol 12:63-5. 14. Gopinathan, L., G. S. Kirimanjeswara, D. N. Wolfe, M. L. Kelley, and E.
T. Harvill. 2007. Different mechanisms of vaccine-induced and infection-induced immunity to Bordetella bronchiseptica. Microbes Infect 9:442-8.
15. Harvill, E. T., P. A. Cotter, and J. F. Miller. 1999. Pregenomic
comparative analysis between bordetella bronchiseptica RB50 and Bordetella pertussis tohama I in murine models of respiratory tract infection. Infect Immun 67:6109-18.
16. Higgins, S. C., A. G. Jarnicki, E. C. Lavelle, and K. H. Mills. 2006.
TLR4 mediates vaccine-induced protective cellular immunity to Bordetella pertussis: role of IL-17-producing T cells. J Immunol 177:7980-9.
17. Inatsuka, C. S., S. M. Julio, and P. A. Cotter. 2005. Bordetella
filamentous hemagglutinin plays a critical role in immunomodulation, suggesting a mechanism for host specificity. Proc Natl Acad Sci U S A 102:18578-83.
18. Kirimanjeswara, G. S., P. B. Mann, and E. T. Harvill. 2003. Role of
antibodies in immunity to Bordetella infections. Infect Immun 71:1719-24. 19. Leef, M., K. L. Elkins, J. Barbic, and R. D. Shahin. 2000. Protective
immunity to Bordetella pertussis requires both B cells and CD4(+) T cells for key functions other than specific antibody production. J Exp Med 191:1841-52.
20. Mahon, B. P., M. S. Ryan, F. Griffin, and K. H. Mills. 1996. Interleukin-
12 is produced by macrophages in response to live or killed Bordetella pertussis and enhances the efficacy of an acellular pertussis vaccine by promoting induction of Th1 cells. Infect Immun 64:5295-301.
21. Mann, P., E. Goebel, J. Barbarich, M. Pilione, M. Kennett, and E.
Harvill. 2007. Use of a genetically defined double mutant strain of Bordetella bronchiseptica lacking adenylate cyclase and type III secretion as a live vaccine. Infect Immun 75:3665-72.
22. Mattoo, S., J. F. Miller, and P. A. Cotter. 2000. Role of Bordetella
bronchiseptica fimbriae in tracheal colonization and development of a humoral immune response. Infect Immun 68:2024-33.
172

23. Mills, K. H. 2001. Immunity to Bordetella pertussis. Microbes Infect 3:655-77.
24. Mills, K. H., M. Ryan, E. Ryan, and B. P. Mahon. 1998. A murine model
in which protection correlates with pertussis vaccine efficacy in children reveals complementary roles for humoral and cell-mediated immunity in protection against Bordetella pertussis. Infect Immun 66:594-602.
25. Pilione, M. R., and E. T. Harvill. 2006. The Bordetella bronchiseptica
type III secretion system inhibits gamma interferon production that is required for efficient antibody-mediated bacterial clearance. Infect Immun 74:1043-9.
26. Roy, C. R., J. F. Miller, and S. Falkow. 1990. Autogenous regulation of
the Bordetella pertussis bvgABC operon. Proc Natl Acad Sci U S A 87:3763-7.
27. Shimizu, T., and H. Ishikawa. 1982. Some characteristics of a urease-
negative, temperature- sensitive strain of Bordetella bronchiseptica as a live, attenuated vaccine. Infect Immun 36:198-201.
28. Stevenson, A., and M. Roberts. 2002. Use of a rationally attenuated
Bordetella bronchiseptica as a live mucosal vaccine and vector for heterologous antigens. Vaccine 20:2325-35.
29. Stevenson, A., and M. Roberts. 2003. Use of Bordetella bronchiseptica
and Bordetella pertussis as live vaccines and vectors for heterologous antigens. FEMS Immunol Med Microbiol 37:121-8.
30. Wolfe, D. N., G. S. Kirimanjeswara, E. M. Goebel, and E. T. Harvill.
2007. Comparative role of immunoglobulin A in protective immunity against the Bordetellae. Infect Immun 75:4416-22.
173

CURRICULUM VITAE
NAME: Neelima Sukumar ADDRESS:
Residence: 390 Sailway Road Winston-Salem, NC 27127 Phone: (336) 602-2541; Cell: (336) 406-6633 Business: Department of Microbiology & Immunology Wake Forest University School of Medicine Medical Center Boulevard Winston-Salem, North Carolina 27157 Phone: (336) 716-1211 FAX: (336) 716-9928 E-mail: [email protected]
EDUCATION
2004-Present Wake Forest University School of Medicine Winston-Salem, NC Advisor: Dr. Rajendar Deora Ph.D.: Projected May 2009 Major: Microbiology & Immunology
Dissertation Title: Bvg-dependent Regulation and Protective Efficacy of BcfA GPA: 4.0
1999-2004 Kerala Agricultural University Vellayani, Kerala, India
Bachelor of Science in Agriculture GPA: 9.0/10 (Ranked 1st in the University) FELLOWSHIPS 2004 Dean’s Fellowship, Wake Forest University Graduate School of Arts & Sciences
1999-2003 Indian Council of Agricultural Research Merit Scholarship
174

1999-2003 Kerala Agricultural University Merit
Scholarship 1999-2003 Cochin Refineries School Higher Education Scholarship
PATENTS:
Deora R, Mishra M, Sukumar N. Role of a Bordetella outer-membrane protein BcfA in protective immunity and as a vaccine candidate. Applied, 2008
PUBLICATIONS
JOURNAL ARTICLES:
Sukumar N, Love CF, Conover MS, Kock ND, Dubey P, Deora R. Active and Passive Immunization with Bordetella Colonization Factor A (BcfA) Protects Mice against respiratory challenge with Bordetella bronchiseptica. Infect Immun. 2009 Feb;77(2):885-95. Featured in the Spot light section Sukumar N, Mishra M, Sloan GP, Ogi T, Deora R. Differential Bvg phase-dependent regulation and combinatorial role in pathogenesis of two Bordetella paralogs, BipA and BcfA. J Bacteriol. 2007 May;189(10):3695-704. Sloan GP, Love CF, Sukumar N, Mishra M, Deora R. The Bordetella Bps polysaccharide is critical for biofilm development in the mouse respiratory tract. J Bacteriol. 2007 Nov;189(22):8270-6.
ABSTRACTS: Sukumar N, Love CF, Conover MS, Kock ND, Dubey P, Deora R. BcfA as a vaccine candidate against Bordetella infections in humans and animals. Charlotte Biotechnology Conference, Barnhardt Student Activities Center, UNC Charlotte, October 28, 2008. Sukumar N, Love CF, Conover MS, Kock ND, Dubey P, Deora R. Immunization with BcfA induces protective immunity against B. bronchiseptica infections. Graduate Student Festival, National Institutes of Health, Bethesda, Maryland, September 11-12, 2008. Sukumar N, Mishra M, Sloan GP, Ogi T, Deora R. Differential Bvg phase-dependent regulation and combinatorial role in pathogenesis
175

of two Bordetella paralogs, BipA and BcfA. American Society for Microbiology 107th General meeting, Metro Convention Center, Toronto, Canada, May 21-25, 2007. Sloan GP, Love CF, Sukumar N, Mishra M, Deora R. Biofilm Formation of Bordetella in the Murine Respiratory Tract. American Society for Microbiology 107th General meeting, Metro Convention Center, Toronto, Canada, May 21-25, 2007. Sloan GP, Love CF, Sukumar N, Mishra M, Deora R. Role of the Bordetella BPS Exopolysaccaride in Promoting Biofilm Development and Persistent Colonization of the Mammalian Respiratory Tract. American Society for Microbiology Conference on Biofilm, Quebec City, Canada, March 25-29 2007. Sukumar N, Mishra M, Sloan GP, Ogi T, Deora R. Differential Bvg phase-dependent Regulation and Combinatorial Role in Pathogenesis of two Bordetella Paralogs, BipA and BcfA. Mid-Atlantic Microbial Pathogenesis Meeting, Wintergreen Resort, Wintergreen, Virginia, February 11-13, 2007. Sukumar N, Mishra M, Sloan GP, Ogi T, Deora R. Synergistic Role of Two Homologous Proteins BipA and BcfA in Respiratory Tract Colonization by Bordetella. 2005 NC ASM Annual meeting The Jane S. McKimmon Conference Center 1101 Gorman Street, NC State University, Raleigh, NC,October 7, 2005
ORAL PRESENTATIONS:
Sukumar N, Love CF, Conover MS, Kock ND, Dubey P, Deora R. Immunization with Bordetella Colonization Factor A (BcfA) Protects Mice against respiratory challenge with Bordetella bronchiseptica. 2008 NC ASM Annual meeting, Frank Family Science Center Guilford College, Greensboro, NC, October 10, 2008 Sukumar N, Mishra M, Sloan GP, Ogi T, Deora R. Differential Bvg phase-dependent Regulation and Combinatorial Role in Pathogenesis of two Bordetella Paralogs, BipA and BcfA. Mid-Atlantic Microbial Pathogenesis Meeting, Wintergreen Resort, Wintergreen, Virginia, February 11-13, 2007. Sukumar N, Mishra M, Sloan GP, Ogi T, Deora R. Synergistic Role of Two Homologous Proteins BipA and BcfA in Respiratory Tract Colonization by Bordetella. 2005 NC ASM Annual meeting The Jane S. McKimmon Conference Center 1101 Gorman Street, NC State University, Raleigh, NC, October 7, 2005
176

Sukumar N, Deora R. Role of two paralogous proteins BipA and BcfA in Bordetella pathogenesis. Fall Symposium, Department of Microbiology and Immunology, Wake Forest University Baptist Medical Center, Winston-Salem, NC, August, 2006. Sukumar N, Deora R. Role of Bvg regulated genes bcfA and bipA in respiratory tract colonization. Fall Symposium, Department of Microbiology and Immunology, Commons Conference Room, Wake Forest University Baptist Medical Center, Winston-Salem, NC, August 23, 2005.
AWARDS: Recipient of Second Prize, Annual Graduate Student Poster Competition, Charlotte Biotechnology Conference, Barnhardt Student Activities Center, UNC Charlotte, October 28, 2008. Selected to Participate in the Annual Graduate Student Poster Competition, Charlotte Biotechnology Conference, Barnhardt Student Activities Center, UNC Charlotte, October 28, 2008. Selected to Participate in the National Graduate Student Research Festival (Poster Presentation) – NIH, Bethesda, Maryland, September 11-12, 2008. Mid-Atlantic Microbial Pathogenesis Meeting Student Travel Award: Mid-Atlantic Microbial Pathogenesis Meeting, Wintergreen Resort, Wintergreen, Virginia, February 11-13, 2007.
Gold Medal from Kerala Agricultural University, Vellayani, India: For the student with highest GPA, 2003.
Bronze Medal, Duke of Edinburg: For Extracurricular activities, Kochi Refineries School, India, 1999.
RESEARCH SKILLS: Molecular Techniques: DNase-1 Footprinting, EMSA, Primer Extension, RACE-PCR, RNA and DNA isolation, Protein over-expression and purification, Gel electrophoresis, Western Blotting, RT-PCR, Reporter Assays, Cloning and Mutagenesis.
Immunological Assays: ELISA, Cytokine Assays, Splenocyte Re-stimulation Assays, Opsonophagocytosis Assays.
177

Animal Models: Worked with mice and rats, transgenic mice, Immunizations, Harvesting different organs. Adhoc review of manuscripts.
PROFESSIONAL APPOINTMENTS & ACTIVITIES
Graduate School Activities Member of Wake Forest University Graduate Student Association, 2004- Present
PROFESSIONAL MEMBERSHIPS
Member of American Society for Microbiology. Member of American Association for the Advancement of Science.
178

179


















