
Transcriptional profiles of response to terbinafine in Trichophyton rubrum
8
GENOMICS AND PROTEOMICS Transcriptional profiles of response to terbinafine in Trichophyton rubrum Wenliang Zhang & Lu Yu & Jian Yang & Lingling Wang & Junping Peng & Qi Jin Received: 6 November 2008 / Revised: 15 January 2009 / Accepted: 2 February 2009 / Published online: 24 February 2009 # Springer-Verlag 2009 Abstract Trichophyton rubrum is a good model for the study of human pathogenic filamentous fungi. The antifun- gal agent terbinafine (TRB) shows specific activity against T. rubrum. To identify the transcriptional profiles of response to TRB in T. rubrum, a cDNA microarray was constructed from the expressed sequence tags of different phase cDNA libraries and transcriptional profiles of the response to TRB were determined. Exposure to TRB decreased the transcription of some genes involved in the ergosterol biosynthesis pathway, including ERG2, ERG4, ERG24, and ERG25, and induced the expression of some genes involved in lipid metabolism such as ERG10, ERG13, and INO1. TRB affected transcription of multidrug-resistance genes and some genes encoding ribosomal proteins. Keywords Transcriptional profiles . Trichophyton rubrum . Terbinafine . Microarray . Antifungal Introduction Dermatophytosis is a common disease that can affect a large proportion of the human population (Vander Straten et al. 2003). The main causative agent of dermatophytosis is Trichophyton rubrum (Elewski et al. 2002), a widespread filamentous fungus that can infect keratinized tissue such as skin, nails and, rarely, hair (Costa et al. 2002; Jennings et al. 2002; Monod et al. 2002). T. rubrum is known to account for up to 69.5% of all dermatophyte infections (Chen and Friedlander 2001; Coloe and Baird 1999), which are often intractable, and relapse occurs frequently after cessation of antifungal therapy (Mukherjee et al. 2003). The prevalence of T. rubrum infections and its anthrophilic nature make it a good model for the study of human pathogenic filamentous fungi. Among the antifungal agents, allylamines such as terbinafine (TRB) are very widely used, both orally and topically, in the therapy of superficial mycoses, including dermatophyte onychomycosis, dermatomycoses, tinea, and piedra (Ryder 1992; Jessup et al. 2000; Aly 2001). Common therapeutic strategies based on the use of TRB are generally considered effective (Degreef and DeDoncker 1994; Crawford et al. 2002), but resistance against the antifungal agents has been reported (Mukherjee et al. 2003). The allylamines are potent inhibitors of the fungal squalene epoxidase enzyme (Ryder 1992), which is involved in the conversion of squalene to squalene epoxide, a precursor in the biosynthesis of ergosterol (Favre and Ryder 1996). As a consequence of this inhibition, fungal cells accumulate squalene and become depleted of ergos- terol (Paltauf et al. 1982; Leber et al. 1995). The cDNA microarray is a good tool for drug target identification in a survey of the global effects mediated by antifungal agents. Although the mechanisms of action of Appl Microbiol Biotechnol (2009) 82:1123–1130 DOI 10.1007/s00253-009-1908-9 Electronic supplementary material The online version of this article (doi:10.1007/s00253-009-1908-9) contains supplementary material, which is available to authorized users. W. Zhang : J. Yang : L. Wang : J. Peng : Q. Jin (*) State Key Laboratory for Molecular Virology and Genetic Engineering, Beijing 100176, China e-mail: [email protected] Q. Jin Institute of Pathogen Biology, Chinese Academy of Medical Sciences, Beijing 100730, China L. Yu Key Laboratory of Zoonosis, Ministry of Education, Institute of Zoonosis, Jilin University, Changchun 130062, China
-
Upload
wenliang-zhang -
Category
Documents
-
view
218 -
download
0
Transcript of Transcriptional profiles of response to terbinafine in Trichophyton rubrum

GENOMICS AND PROTEOMICS
Transcriptional profiles of response to terbinafinein Trichophyton rubrum
Wenliang Zhang & Lu Yu & Jian Yang & Lingling Wang &
Junping Peng & Qi Jin
Received: 6 November 2008 /Revised: 15 January 2009 /Accepted: 2 February 2009 /Published online: 24 February 2009# Springer-Verlag 2009
Abstract Trichophyton rubrum is a good model for thestudy of human pathogenic filamentous fungi. The antifun-gal agent terbinafine (TRB) shows specific activity againstT. rubrum. To identify the transcriptional profiles ofresponse to TRB in T. rubrum, a cDNA microarray wasconstructed from the expressed sequence tags of differentphase cDNA libraries and transcriptional profiles of theresponse to TRB were determined. Exposure to TRBdecreased the transcription of some genes involved in theergosterol biosynthesis pathway, including ERG2, ERG4,ERG24, and ERG25, and induced the expression of somegenes involved in lipid metabolism such as ERG10,ERG13, and INO1. TRB affected transcription ofmultidrug-resistance genes and some genes encodingribosomal proteins.
Keywords Transcriptional profiles . Trichophyton rubrum .
Terbinafine .Microarray . Antifungal
Introduction
Dermatophytosis is a common disease that can affect alarge proportion of the human population (Vander Straten etal. 2003). The main causative agent of dermatophytosis isTrichophyton rubrum (Elewski et al. 2002), a widespreadfilamentous fungus that can infect keratinized tissue such asskin, nails and, rarely, hair (Costa et al. 2002; Jennings etal. 2002; Monod et al. 2002). T. rubrum is known toaccount for up to 69.5% of all dermatophyte infections(Chen and Friedlander 2001; Coloe and Baird 1999), whichare often intractable, and relapse occurs frequently aftercessation of antifungal therapy (Mukherjee et al. 2003). Theprevalence of T. rubrum infections and its anthrophilicnature make it a good model for the study of humanpathogenic filamentous fungi.
Among the antifungal agents, allylamines such asterbinafine (TRB) are very widely used, both orally andtopically, in the therapy of superficial mycoses, includingdermatophyte onychomycosis, dermatomycoses, tinea, andpiedra (Ryder 1992; Jessup et al. 2000; Aly 2001).Common therapeutic strategies based on the use of TRBare generally considered effective (Degreef and DeDoncker1994; Crawford et al. 2002), but resistance against theantifungal agents has been reported (Mukherjee et al.2003). The allylamines are potent inhibitors of the fungalsqualene epoxidase enzyme (Ryder 1992), which isinvolved in the conversion of squalene to squalene epoxide,a precursor in the biosynthesis of ergosterol (Favre andRyder 1996). As a consequence of this inhibition, fungalcells accumulate squalene and become depleted of ergos-terol (Paltauf et al. 1982; Leber et al. 1995).
The cDNA microarray is a good tool for drug targetidentification in a survey of the global effects mediated byantifungal agents. Although the mechanisms of action of
Appl Microbiol Biotechnol (2009) 82:1123–1130DOI 10.1007/s00253-009-1908-9
Electronic supplementary material The online version of this article(doi:10.1007/s00253-009-1908-9) contains supplementary material,which is available to authorized users.
W. Zhang : J. Yang : L. Wang : J. Peng :Q. Jin (*)State Key Laboratory for Molecular Virologyand Genetic Engineering,Beijing 100176, Chinae-mail: [email protected]
Q. JinInstitute of Pathogen Biology,Chinese Academy of Medical Sciences,Beijing 100730, China
L. YuKey Laboratory of Zoonosis, Ministry of Education,Institute of Zoonosis, Jilin University,Changchun 130062, China

antifungal agents against some model fungi such asSaccharomyces cerevisiae and Candida albicans have beenstudied by cDNA microarray (Bammert and Fostel 2000;De Backer et al. 2001; Zhang et al. 2002), the inhibitorymechanism of TRB against T. rubrum is poorly understood.
In our experiment, a cDNA microarray was constructedfrom the expressed sequence tags (ESTs) of the cDNA libraryof T. rubrum. To identify class-specific changes in geneexpression, we used a cDNA microarray to examine changesin transcriptional profiles of T. rubrum in response to TRB.
Materials and methods
Fungus and material
The T. rubrum clinical isolate BMU 01672 used in thisstudy was provided by Professor Ruoyu Li (ResearchCenter for Medical Mycology, Peking University). Theisolate was confirmed as T. rubrum by morphologicidentification of both microscopic and macroscopic charac-teristics (De Hoog et al. 2000; Larone 2002) as well as byPCR amplification and sequencing of the 18S ribosomalDNA and internal transcribed spacer (ITS) regions (Jacksonet al. 1999). Potato dextrose agar (PDA), yeast extract,peptone, and D-glucose used for the strain cultures werefrom Difco. TRB was obtained from Sigma (St. Louis, MO,USA). Stock solutions of various concentrations were madein dimethyl sulfoxide (DMSO; Sigma).
Preparation of cell cultures, construction of cDNA libraries,sequencing, and annotation
Potato dextrose agar (39 g/l) was inoculated with a fewhyphae of T. rubrum and incubated at 28°C for 2 to3 weeks until good conidiation was produced. The mixtureof conidia and hyphal fragments was collected in distilledwater. Construction of cDNA libraries, cDNA sequencing,Phred quality assessment, and computational analysis weredone as described (Wang et al. 2006; Yu et al. 2007).
cDNA microarray
T7 and SP6 universal primers were used for amplificationof PCR fragments used for printing the microarray chip.Purified PCR products were resuspended in 12 Cl ofspotting solution containing 50% dimethyl sulfoxide toproduce the microarrays. A set of microarrays containing atotal of 11,232 spots (10,250 clones in the form of PCRproducts and 982 controls, including blank, negative, andpositive controls) were spotted in duplicate on b aminopropylsilan-coated GAP II slides (Corning) with a Cartesianarrayer. Slides were subsequently placed in a blocking
solution containing 0.2 M succinic anhydride and 0.05 Msodium borate prepared in 1-methyl-2-pyrrolidinone for20 min, washed for 2 min in 95°C water, and rinsed fivetimes in 95% ethanol. Slides were spin dried and stored forfuture hybridizations (Diehl et al. 2001; Peng et al. 2006).
MIC determinations
Test concentrations of TRB ranged from 0.001 to 0.5 µg/ml. MIC is defined as the lowest concentration at which thegrowth of the organism was inhibited 100% compared withgrowth in the control. All tests were done in duplicate andthe results were read visually. MIC results were recorded inunits of micrograms per milliliter. The antifungal suscepti-bility tests were done as described (Zhang et al. 2007).
Cell culture and drug exposure for microarray experiments
For a single microarray experiment, six 100-ml cultureswere prepared (three independent 100-ml cultures weregrown for TRB). TRB was added to three cultures at aconcentration equivalent to 0.5 MIC (0.01 µg/ml) andincubated at 30°C. Three control cultures were treated withDMSO (0.01 µg/ml). Six hours after the addition of TRB,three control cultures and three drug-treated cultures wereharvested and frozen in liquid nitrogen. The culture of T.rubrum clinical isolate BMU 01672 and the experimentalprotocol were as described (Yu et al. 2007).
RNA preparation
Total RNA was isolated using the QIAGEN RNeasy plantmini kit (QIAGEN, Inc., Valencia, CA, USA) according tothe manufacturer’s instructions. Three independent sets ofRNA from control and three independent sets of RNA fromdrug-treated cells were used to prepare six independentcDNA sets. An aliquot of poly(A)+ mRNA was isolatedwith the Oligotex mRNA mini kit (QIAGEN) following themanufacturer’s instructions.
Microarray hybridization
First-strand cDNA was synthesized using Superscript II RT(Life Technologies/Invitrogen, Carlsbad, CA, USA).Second-strand cDNA was synthesized as follows: 5×second-strand buffer; 0.2 mM deoxynucleoside triphos-phate mix; Escherichia coli DNA ligase (10U); E. coliDNA polymerase I (40 U); and E. coli RNase H (2 U) wereadded to a first-strand reaction tube and incubated for 2 h at16°C. Double-stranded cDNA (dscDNA) was purifiedusing QIAquick columns (QIAGEN) by following themanufacturer’s instructions. dscDNA was then fluores-cently labeled using BioPrime DNA labeling system (Life
1124 Appl Microbiol Biotechnol (2009) 82:1123–1130

Technologies/Invitrogen). Those representing RNA fromdrug-treated cells were labeled using Cy5, and thoserepresenting RNA from control cells were labeled usingCy3. Labeled cDNA was purified using QIAquick columnsand mixed with 2 Cg poly(A), 5× Denhardt’s, 3× SSC,24 Cg yeast tRNA, 25 mM HEPES, pH 7.0, and 0.25%SDS. The mixture was heated at 100°C for 2 min, cooled toroom temperature, and applied to the array slides underglass coverslips. Hybridization was performed at 65°Covernight in a Micro hybridization incubator (RobbinsScientific, Sunnyvale, CA). Slides were washed in 2× SSC(0.1% SDS), 1× SSC, and 0.2× SSC sequentially and thenwere scanned at 5 Cm on a GenePix 4000B scanner (AxonInstruments, Inc.). The Cy5- and Cy3-labeled DNAsamples were scanned at 635 and 532 nm, respectively.
Data analysis
GenePix 6.0 software (Axon Instruments) was used forimage analysis and data visualization. Before data analysis,signals were normalized using a locally weighted scatter-plot smoothing regression algorithm in the MIDAS soft-ware package (Cleveland 1979; Saeed et al. 2003), with thesmoothing parameter set to 0.33. Genes were considered tobe expressed differentially if: (1) average expressionchanged by at least 1.5-fold in three independent experi-ments with triplicate RNA samples; and (2) the change ingene expression was in the same direction (increased ordecreased) in three experiments.
Quantitative real-time RT-PCR
Quantitative real-time reverse transcription (RT)-PCR wasused to verify the microarray results. Gene-specific primerswere designed for the genes of interest and are shown inTable 1. All the RT-PCR reactions were performed asdescribed (Yu et al. 2007).
Results
MIC determination
Assays were first done in microplates as described in‘Materials and methods’. The inhibition assays wererepeated in large-scale experiments because the microarrayexperiments use 100-ml cultures and the MIC values maybe different in small-scale and large-scale experiments. TheMIC value was determined as 0.02 µg/ml for TRB, and themicroplate and large-scale assay results were in agreement.These results showed that the T. rubrum clinical isolateBMU 01672 used here was susceptible to TRB.
Global analysis of the transcriptional response to TRB
A large number of genes were affected, both positively andnegatively, by exposure to TRB. In total, 337 genes wereexpressed differentially upon exposure to TRB; 193showed increases in expression and 144 showed decreases
Target Sequence (5′–3′) (directiona) Amplicon size (bp)
18S CGCTGGCTTCTTAGAGGGACTAT (F) 51TGCCTCAAACTTCCATCGACTT (R)
ERG10 TTTGGGTGTCTTGCGTGAG (F) 78CACCCAAAAGAGTAGTCAGGAT (R)
INO1 TCTCAAGGCTATCAAGGAAGG (F) 100GAAGGTTGTCAGCGGTATCAT (R)
ERG4 CACCGTACTCAGAGGGACATT (F) 95GGGACATAGTTCGGCTTGC (R)
ERG24 GTGAGTAGACCCCAGGTGAAG (F) 88CCTTCCCTTGGTTTTACTTCG (R)
DW687448 AATCACGGAGGATGTGCTTT (F) 88ACCCGTGCTGTCGTAAAAG (R)
DW707836 GCTGAATCTGACAGGGAACG (F) 90TGTCGGAACCCATCAAGTC (R)
EB801711 ACGAGGGTTCGGCAATAG (F) 96TTTCGCAGAGCGGGAGA (R)
DW698412 TCCAGCCCACTGCTACAAG (F) 94CGTAAAGAAGCCACCCAATG (R)
MDR1 AGTTTTCCCAACCCTACCC (F) 100GGAGTTCGTGATGAGTGCC (R)
DW708349 TCAAGGGATACGAGCAGATG (F) 79AGCCGTCCCTTTCTTCTGC (R)
Table 1 Gene-specific primersused for real-time RT-PCRassays
aF forward, R reverse
Appl Microbiol Biotechnol (2009) 82:1123–1130 1125
in expression. (The data sets’ GEO number is GSE6951.)The distribution of TRB-responsive genes and theirbiological roles are shown in Table S1 of the Electronicsupplementary material.
Validation of microarray data by real-time RT-PCR
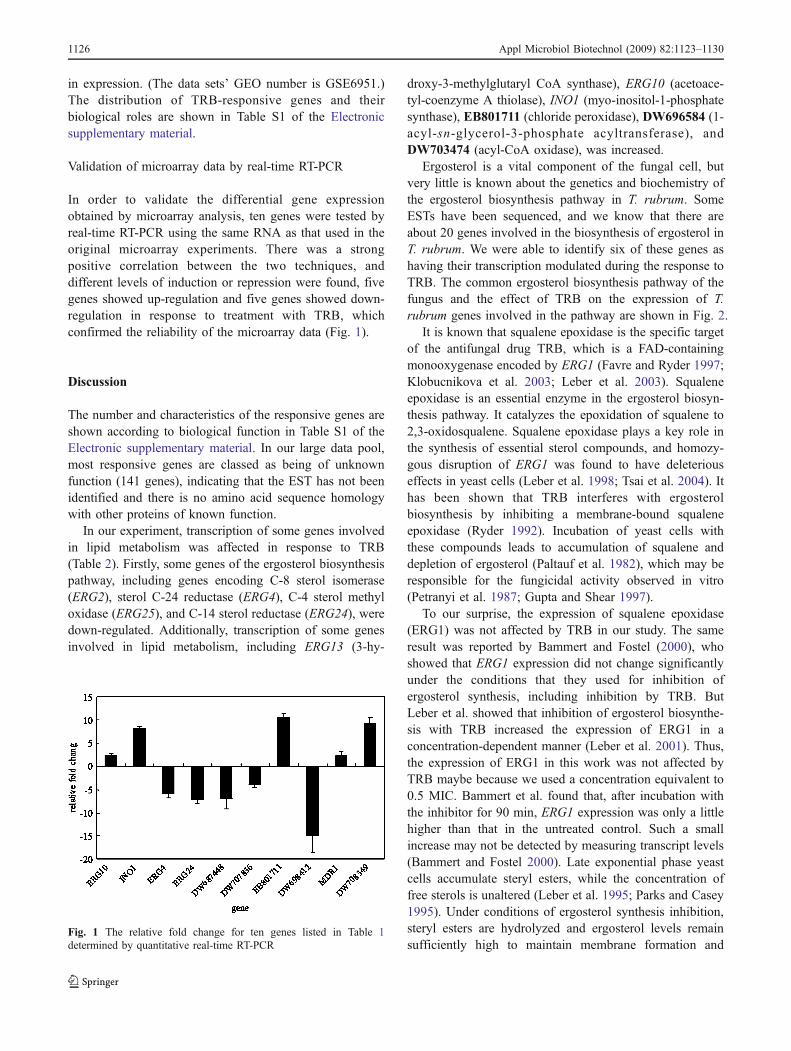
In order to validate the differential gene expressionobtained by microarray analysis, ten genes were tested byreal-time RT-PCR using the same RNA as that used in theoriginal microarray experiments. There was a strongpositive correlation between the two techniques, anddifferent levels of induction or repression were found, fivegenes showed up-regulation and five genes showed down-regulation in response to treatment with TRB, whichconfirmed the reliability of the microarray data (Fig. 1).
Discussion
The number and characteristics of the responsive genes areshown according to biological function in Table S1 of theElectronic supplementary material. In our large data pool,most responsive genes are classed as being of unknownfunction (141 genes), indicating that the EST has not beenidentified and there is no amino acid sequence homologywith other proteins of known function.
In our experiment, transcription of some genes involvedin lipid metabolism was affected in response to TRB(Table 2). Firstly, some genes of the ergosterol biosynthesispathway, including genes encoding C-8 sterol isomerase(ERG2), sterol C-24 reductase (ERG4), C-4 sterol methyloxidase (ERG25), and C-14 sterol reductase (ERG24), weredown-regulated. Additionally, transcription of some genesinvolved in lipid metabolism, including ERG13 (3-hy-
droxy-3-methylglutaryl CoA synthase), ERG10 (acetoace-tyl-coenzyme A thiolase), INO1 (myo-inositol-1-phosphatesynthase), EB801711 (chloride peroxidase), DW696584 (1-acyl-sn-glycerol-3-phosphate acyltransferase), andDW703474 (acyl-CoA oxidase), was increased.
Ergosterol is a vital component of the fungal cell, butvery little is known about the genetics and biochemistry ofthe ergosterol biosynthesis pathway in T. rubrum. SomeESTs have been sequenced, and we know that there areabout 20 genes involved in the biosynthesis of ergosterol inT. rubrum. We were able to identify six of these genes ashaving their transcription modulated during the response toTRB. The common ergosterol biosynthesis pathway of thefungus and the effect of TRB on the expression of T.rubrum genes involved in the pathway are shown in Fig. 2.
It is known that squalene epoxidase is the specific targetof the antifungal drug TRB, which is a FAD-containingmonooxygenase encoded by ERG1 (Favre and Ryder 1997;Klobucnikova et al. 2003; Leber et al. 2003). Squaleneepoxidase is an essential enzyme in the ergosterol biosyn-thesis pathway. It catalyzes the epoxidation of squalene to2,3-oxidosqualene. Squalene epoxidase plays a key role inthe synthesis of essential sterol compounds, and homozy-gous disruption of ERG1 was found to have deleteriouseffects in yeast cells (Leber et al. 1998; Tsai et al. 2004). Ithas been shown that TRB interferes with ergosterolbiosynthesis by inhibiting a membrane-bound squaleneepoxidase (Ryder 1992). Incubation of yeast cells withthese compounds leads to accumulation of squalene anddepletion of ergosterol (Paltauf et al. 1982), which may beresponsible for the fungicidal activity observed in vitro(Petranyi et al. 1987; Gupta and Shear 1997).
To our surprise, the expression of squalene epoxidase(ERG1) was not affected by TRB in our study. The sameresult was reported by Bammert and Fostel (2000), whoshowed that ERG1 expression did not change significantlyunder the conditions that they used for inhibition ofergosterol synthesis, including inhibition by TRB. ButLeber et al. showed that inhibition of ergosterol biosynthe-sis with TRB increased the expression of ERG1 in aconcentration-dependent manner (Leber et al. 2001). Thus,the expression of ERG1 in this work was not affected byTRB maybe because we used a concentration equivalent to0.5 MIC. Bammert et al. found that, after incubation withthe inhibitor for 90 min, ERG1 expression was only a littlehigher than that in the untreated control. Such a smallincrease may not be detected by measuring transcript levels(Bammert and Fostel 2000). Late exponential phase yeastcells accumulate steryl esters, while the concentration offree sterols is unaltered (Leber et al. 1995; Parks and Casey1995). Under conditions of ergosterol synthesis inhibition,steryl esters are hydrolyzed and ergosterol levels remainsufficiently high to maintain membrane formation and
Fig. 1 The relative fold change for ten genes listed in Table 1determined by quantitative real-time RT-PCR
1126 Appl Microbiol Biotechnol (2009) 82:1123–1130
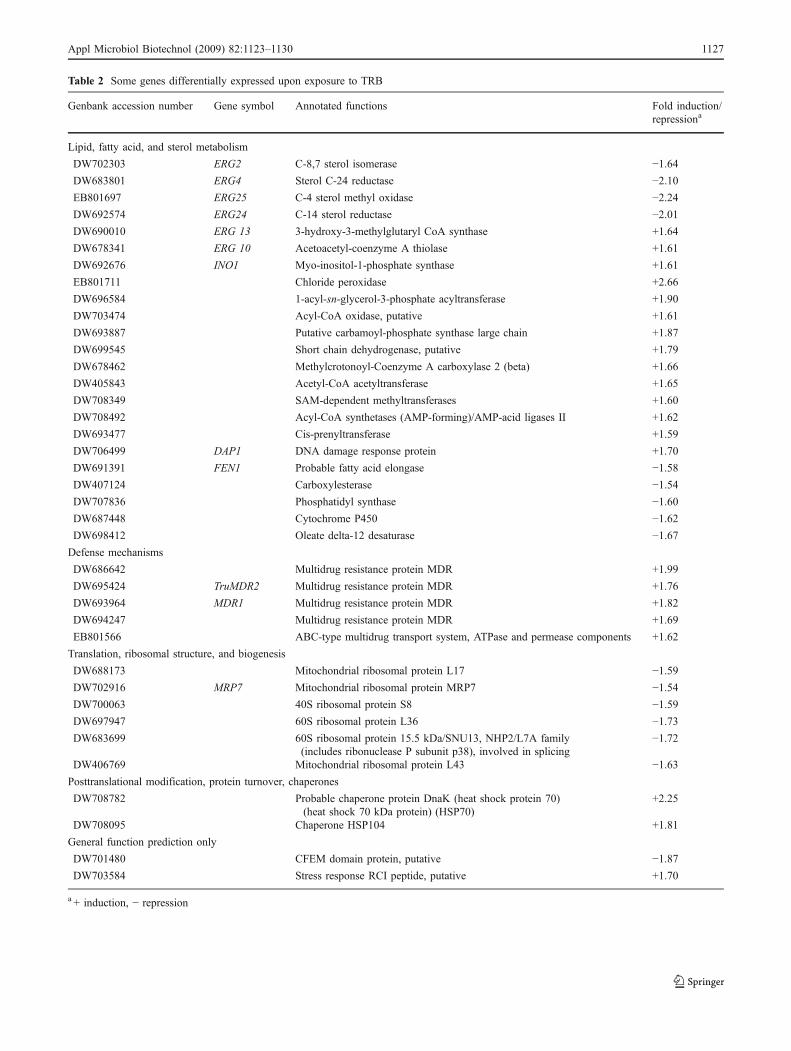
Table 2 Some genes differentially expressed upon exposure to TRB
Genbank accession number Gene symbol Annotated functions Fold induction/repressiona
Lipid, fatty acid, and sterol metabolism
DW702303 ERG2 C-8,7 sterol isomerase −1.64DW683801 ERG4 Sterol C-24 reductase −2.10EB801697 ERG25 C-4 sterol methyl oxidase −2.24DW692574 ERG24 C-14 sterol reductase −2.01DW690010 ERG 13 3-hydroxy-3-methylglutaryl CoA synthase +1.64
DW678341 ERG 10 Acetoacetyl-coenzyme A thiolase +1.61
DW692676 INO1 Myo-inositol-1-phosphate synthase +1.61
EB801711 Chloride peroxidase +2.66
DW696584 1-acyl-sn-glycerol-3-phosphate acyltransferase +1.90
DW703474 Acyl-CoA oxidase, putative +1.61
DW693887 Putative carbamoyl-phosphate synthase large chain +1.87
DW699545 Short chain dehydrogenase, putative +1.79
DW678462 Methylcrotonoyl-Coenzyme A carboxylase 2 (beta) +1.66
DW405843 Acetyl-CoA acetyltransferase +1.65
DW708349 SAM-dependent methyltransferases +1.60
DW708492 Acyl-CoA synthetases (AMP-forming)/AMP-acid ligases II +1.62
DW693477 Cis-prenyltransferase +1.59
DW706499 DAP1 DNA damage response protein +1.70
DW691391 FEN1 Probable fatty acid elongase −1.58DW407124 Carboxylesterase −1.54DW707836 Phosphatidyl synthase −1.60DW687448 Cytochrome P450 −1.62DW698412 Oleate delta-12 desaturase −1.67Defense mechanisms
DW686642 Multidrug resistance protein MDR +1.99
DW695424 TruMDR2 Multidrug resistance protein MDR +1.76
DW693964 MDR1 Multidrug resistance protein MDR +1.82
DW694247 Multidrug resistance protein MDR +1.69
EB801566 ABC-type multidrug transport system, ATPase and permease components +1.62
Translation, ribosomal structure, and biogenesis
DW688173 Mitochondrial ribosomal protein L17 −1.59DW702916 MRP7 Mitochondrial ribosomal protein MRP7 −1.54DW700063 40S ribosomal protein S8 −1.59DW697947 60S ribosomal protein L36 −1.73DW683699 60S ribosomal protein 15.5 kDa/SNU13, NHP2/L7A family
(includes ribonuclease P subunit p38), involved in splicing−1.72
DW406769 Mitochondrial ribosomal protein L43 −1.63Posttranslational modification, protein turnover, chaperones
DW708782 Probable chaperone protein DnaK (heat shock protein 70)(heat shock 70 kDa protein) (HSP70)
+2.25
DW708095 Chaperone HSP104 +1.81
General function prediction only
DW701480 CFEM domain protein, putative −1.87DW703584 Stress response RCI peptide, putative +1.70
a + induction, − repression
Appl Microbiol Biotechnol (2009) 82:1123–1130 1127

growth for some time (Leber et al. 1995), and, thus, do notlead to induced ERG1 expression (Leber et al. 2001).Ergosterol has been reported to regulate its own synthesisby negative feedback mechanisms (Pinto et al. 1985;Servouse and Karst 1986; Casey et al. 1992; Smith et al.1996).
In this experiment, we observed that genes ERG10 andERG13 were up-regulated by TRB, which indicates that lowlevels of ergosterol increase the enzymatic activities ofacetoacetyl-CoA thiolase and HMG-CoA synthase, twoearly enzymes in the ergosterol biosynthesis pathway(Servouse and Karst 1986), and this result is consistentwith a previous report (Pasrija et al. 2005). The expressionof ERG24, ERG4, and ERG25 was down-regulated, perhapsbecause they are functionally downstream of ERG1, andtheir reduction is in response to ergosterol depletion.
In this study, TRB induced the transcription ofmultidrug-resistance (MDR) genes, including DW686642,TruMDR2, and MDR1. The reduced intracellular accumu-lation has been correlated with over-expression of MDRefflux transporter genes of the ATP-binding cassette (ABC),and the major facilitator superfamily (MFS) classes (Lupetti
et al. 2002, Fachin et al. 2006). Multidrug- and drug-specific efflux systems are responsible for clinicallysignificant resistance to chemotherapeutic agents in thetreatment of pathogenic bacteria, fungi, parasites, andcancer cells (Paulsen et al. 1996). It has long beenspeculated that fungi have efflux pumps that allow themto avoid the effects of metabolic poisons present in theenvironment. Urban et al. proposed that the fungal trans-porters initiate associations between host and fungal cells(Urban et al. 1999).
We found that transcription of some genes encodingribosomal proteins, including DW688173 (mitochondrialribosomal protein L17), MRP7 (mitochondrial ribosomalprotein MRP7), and DW700063 (40S ribosomal proteinS8), was down-regulated significantly (see Table S1 of theElectronic supplementary material). Small reductions in theexpression of ribosomal protein genes allow energy to beredistributed, which allows increased expression of genesinvolved in protective responses, while maintaining a basallevel of protein synthesis (Vanden Bossche et al. 1987).
It was found that TRB induced some genes that encodestress response-related proteins in T. rubrum, such as
Fig. 2 Effect of TRB on theexpression of T. rubrum genesinvolved in the ergosterol bio-synthetic pathway. Boldfacesindicate genes with increasedtranscript levels and shadedareas indicate genes withdecreased transcript levels. Thesterol biosynthesis inhibitor(TRB) examined in this studywas underlined and shown tothe right of the gene encodingthe targeted enzyme. CoAcoenzyme A, HMG-CoAsynthase 3-hydroxy-3-methylglutaryl CoA synthase,FPP synthase farnesylpyrophosphate synthetase
1128 Appl Microbiol Biotechnol (2009) 82:1123–1130

DW708782 (HSP70) and DW708095 (HSP104), andoxidative-stress proteins such as DW703584 (stress re-sponse RCI peptide).
In addition, expression of the gene DW701480 encodingthe CFEM domain protein was inhibited. CFEM mightserve as a characteristic signature for a subset of proteinsthat function in the extracellular environment. SomeCFEM-containing proteins are proposed to have importantroles in fungal pathogenesis (Kulkarni et al. 2003), whichmay be of benefit in studying the interaction betweenpathogen and host.
In conclusion, the T. rubrum microarray studies de-scribed here revealed the changes of the transcriptionallevel of genes exposed to the allylamine antifungal agentTRB and laid the groundwork for an antifungal drugdevelopment program employing cDNA microarrays toidentify gene expression profiles.
Acknowledgments We are grateful to Ruoyu Li (Research Centerfor Medical Mycology, Peking University) for providing strain T.rubrum BMU 01672 and for helpful discussions. Financial supportsfor this work came from the National High Technology Research andDevelopment Program of China (accession number 2006AA020504)and National Key Technologies R&D Programme (accession number2002BA711A14).
References
Aly R, Forney R, Bayles C (2001) Treatments for common superficialfungal infections. Dermatol Nurs 13:91–101
Bammert GF, Fostel JM (2000) Genome-wide expression patterns inSaccharomyces cerevisiae: comparison of drug treatments andgenetic alterations affecting biosynthesis of ergosterol. Antimi-crob Agents Chemother 44:1255–1265
Casey WM, Keesler GA, Parks LW (1992) Regulation of partitionedsterol biosynthesis in Saccharomyces cerevisiae. J Bacteriol174:7283–7288
Chen BK, Friedlander SF (2001) Tinea capitis update: a continuingconflict with an old adversary. Curr Opin Pediatr 13:331–335
Cleveland WS (1979) Robust locally weighted regression andsmoothing scatterplots. J Am Stat Assoc 74:829–836
Coloe SV, Baird RW (1999) Dermatophyte infections in Melbourne:trends from 1961/64 to 1995/96. Pathology 31:395–397
Costa M, Passos XS, Souza LK, Miranda AT, Lemos JA, Oliveira JG,Silva MR (2002) Epidemiology and etiology of dermatophytosisin Goiania, GO. Brazil Rev Soc Bras Med Trop 35:19–22
Crawford F, Young P, Godfrey C, Bell-Syer SE, Hart R, Brunt E,Russell I (2002) Oral treatments for toenail onychomycosis: asystematic review. Arc Dermatol 138:811–816
De Backer MD, Ilyina T, Ma XJ, Vandoninck S, Luyten WH, VandenBossche H (2001) Genomic profiling of the response of Candidaalbicans to itraconazole treatment using a DNA microarray.Antimicrob Agents Chemother 45:1660–1670
De Hoog GS, Guarro J, Gene J, Figueras MJ (2000) Atlas of ClinicalFungi, 2nd Ed. Centraalbureau voor Schimmelcultures, Utrecht,The Netherlands and University Rovirai Virgili, Reus, Spain
Degreef HJ, DeDoncker PR (1994) Current therapy of dermatophy-tosis. J Am Acad Dermatol 31:S25–S30
Diehl F, Grahlmann S, Beier M, Hoheisel JD (2001) ManufacturingDNA-microarrays of high spot homogeneity and reducedbackground signal. Nucleic Acids Res 29:e38
Elewski BE, Leyden J, Rinaldi MG, Atillasoy E (2002) Officepractice-based confirmation of onychomycosis: a U.S. nation-wide prospective survey. Arch Intern Med 162:2133–2138
Fachin AL, Ferreira-Nozawa MS, Maccheroni W, Martinez-Rossi NM(2006) Role of the ABC transporter TruMDR2 in terbinafine, 4-nitroquinoline N-oxide and ethidium bromide susceptibility inTrichophyton rubrum. J Med Microbiol 55:1093–1099
Favre B, Ryder NS (1996) Characterization of squalene epoxidaseactivity from the dermatophyte Trichophyton rubrum and itsinhibition by terbinafine and other antimycotic agents. Anti-microb Agents Chemother 40:443–447
Favre B, Ryder NS (1997) Cloning and expression of squaleneepoxidase from pathogenic yeast Candida albicans. Gene189:119–126
Gupta AK, Shear NH (1997) Terbinafine: an update. J Am AcadDermatol 37:979–988
Jackson CJ, Barton RC, Evans EGV (1999) Species identification andstrain differentiation of dermatophyte fungi by analysis ofribosomal-DNA intergenic spacer regions. J Clin Microbiol37:931–936
Jennings MB, Weinberg JM, Koestenblatt EK, Lesczczynski C (2002)Study of clinically suspected onychomycosis in a podiatricpopulation. J Am Podiat Med Assoc 92:327–330
Jessup CJ, Ryder NS, Ghannoum MA (2000) An evaluation of the invitro activity of terbinafine. Med Mycol 38:155–159
Klobucnikova V, Kohut P, Leber R, Fuchsbichler S, Schweighofer N,Turnowsky F, Hapala I (2003) Terbinafine resistance in apleiotropic yeast mutant is caused by a single point mutation inthe ERG1 gene. Biochem Biophys Res Commun 309:666–671
Kulkarni RD, Kelkar HS, Dean RA (2003) An eight-cysteine-containing CFEM domain unique to a group of fungal membraneproteins. Trends Biochem Sci 28:118–121
Larone DH (2002) Medically important fungi: a guide to identification,4th ed. American Society for Microbiology, Washington, DC
Leber R, Fuchsbichler S, Klobucnikova V, Schweighofer N, Pitters E,Wohlfarter K, Lederer M, Landl K, Ruckenstuhl C, Hapala I,Turnowsky F (2003) Molecular mechanism of terbinafineresistance in S. cerevisiae. Antimicrob Agents Chemother47:3890–3900
Leber R, Landl K, Zinser E, Ahorn H, Spök A, Kohlwein SD,Turnowsky F, Daum G (1998) Dual localization of squaleneepoxidase, Erg1p, in yeast reflects a relationship between theendoplasmic reticulum and lipid particles. Mol Biol Cell 9:375–386
Leber R, Zenz R, Schröttner K, Fuchsbichler S, Pühringer B,Turnowsky F (2001) A novel sequence element is involved inthe transcriptional regulation of expression of the ERG1(squalene epoxidase) gene in Saccharomyces cerevisiae. Eur JBiochem 268:914–924
Leber R, Zinser E, Hrastnik C, Paltauf F, Daum G (1995) Export ofsteryl esters from lipid particles and release of free sterols in theyeast, Saccharomyces cerevisiae. Biochim Biophys Acta1234:119–126
Lupetti A, Danesi R, Campa M, Del Tacca M, Kelly S (2002)Molecular basis of resistance to azole antifungals. Trends MolMed 8:76–81
Monod M, Jaccoud S, Zaugg C, Lechenn B, Baudraz F, Panizzon R(2002) Survey of dermatophyte infections in the Lausanne area(Switzerland). Dermatology 205:201–203
Mukherjee PK, Leidich SD, Isham N, Leitner I, Ryder NS, GhannoumMZ (2003) Clinical Trichophyton rubrum strain exhibitingprimary resistance to terbinafine. Antimicrob Agents Chemother47:82–86
Appl Microbiol Biotechnol (2009) 82:1123–1130 1129

Paltauf F, Daum G, Zuder G, Högenauer G, Schulz G, Seidl G (1982)Squalene and ergosterol biosynthesis in fungi treated withnaftifine, a new antimycotic agent. Biochim Biophys Acta712:268–273
Parks LW, Casey WM (1995) Physiological implications of sterolbiosynthesis in yeast. Annu Rev Microbiol 49:95–116
Pasrija R, Krishnamurthy S, Prasad T, Ernst JF, Prasad R (2005)Squalene epoxidase encoded by ERG1 affects morphogenesisand drug susceptibilities of Candida albicans. J AntimicrobChemother 55:905–913
Paulsen IT, Brown MH, Skurray RA (1996) Proton-dependentmultidrug efflux systems. Microbiol Rev 60:575–608
Peng J, Zhang X, Yang J, Wang J, Yang E, Bin W, Wei C, Sun M, JinQ (2006) The use of comparative genomic hybridization tocharacterize genome dynamics and diversity among the serotypesof Shigella. BMC genomics 7:218
Petranyi G, Meingassner JG, Mieth H (1987) Activity of terbinafine inexperimental fungal infections of laboratory animals. AntimicrobAgents Chemother 31:1558–1561
Pinto WJ, Lozano R, Nes WR (1985) Inhibition of sterol biosynthesisby ergosterol and cholesterol in Saccharomyces cerevisiae.Biochim Biophys Acta 836:89–95
Ryder NS (1992) Terbinafine: mode of action and properties of thesqualene epoxidase inhibition. Br J Dermatol 126(Suppl. 39):2–7
Saeed AI, Sharov V, White J, Li J, Liang W, Bhagabati N, Braisted J,Klapa M, Currier T, Thiagarajan M, Sturn A, Snuffin M,Rezantsev A, Popov D, Ryltsov A, Kostukovich E, BorisovskyI, Liu Z, Vinsavich A, Trush V, Quackenbush J (2003) TM4: afree, open-source system for microarray data management andanalysis. Biotechniques 34:374–378
Servouse M, Karst F (1986) Regulation of early enzymes of ergosterolbiosynthesis in Saccharomyces cerevisiae. Biochem J 240:541–547
Smith SJ, Crowley JH, Park SLW (1996) Transcriptional regulation byergosterol in the yeast Saccharomyces cerevisiae. Mol Cell Biol16:5427–5432
Tsai HF, Bard M, Izumikawa K, Krol AA, Sturm AM, Culbertson NT,Pierson CA, Bennett JE (2004) Candida glabrata mutant withincreased sensitivity to azoles and to low oxygen tension.Antimicrob Agents Chemother 48:2483–2489
Urban M, Bhargava T, Hamer JE (1999) An ATP-driven efflux pumpis a novel pathogenicity factor in rice blast disease. EMBO J18:512–521
Vanden Bossche H, Willemsens G, Marichal P (1987) Anti-Candidadrugs—the biochemical basis for their activity. Clin MicrobiolRev 15:57–72
Vander Straten MR, Hossain MA, Ghannoum MA (2003) Cutaneousinfections dermatophytosis, onychomycosis, and tinea versicolor.Infect Dis Clin North Am 17:87–112
Wang L, Ma L, Leng W, Liu T, Yu L, Yang J, Yang L, Zhang W, ZhangQ, Dong J, Xue Y, Zhu Y, Xu X, Wan Z, Ding G, Yu F, Tu K, Li Y,Li R, Shen Y, Jin Q (2006) Analysis of the dermatophyte Tricho-phyton rubrum expressed sequence tags. BMC Genomics 7:255
Yu L, Zhang W, Wang L, Yang J, Liu T, Peng J, Leng W, Chen L, LiR, Jin Q (2007) Transcriptional profiles of the response toketoconazole and amphotericin B in Trichophyton rubrum.Antimicrob Agents Chemother 51:144–153
Zhang L, Zhang Y, Zhou Y, An S, Zhou Y, Cheng J (2002) Responseof gene expression in Saccharomyces cerevisiae to amphotericinB and nystatin measured by microarrays. J Antimicrob Chemo-ther 49:905–915
Zhang W, Yu L, Leng W, Wang X, Wang L, Deng X, Yang J, Liu T,Peng J, Wang J, Li S, Jin Q (2007) cDNA microarray analysis ofthe expression profiles of Trichophyton rubrum in response tonovel synthetic fatty acid synthase inhibitor PHS11A. FungalGenet Biol 4:1252–1261
1130 Appl Microbiol Biotechnol (2009) 82:1123–1130


















