
Tomato yellow leaf curl virus resistance gene ty-5tgc.ifas.ufl.edu/2013/2013/05 - Levin.pdfCloning...
49
Cloning and analysis of the Tomato yellow leaf curl virus resistance gene ty- 5
Transcript of Tomato yellow leaf curl virus resistance gene ty-5tgc.ifas.ufl.edu/2013/2013/05 - Levin.pdfCloning...

Cloning and analysis of the Tomato yellow leaf curl virus resistance
gene ty- 5

TYLCV - a monopartite begomovirus, posing increasing threat to tomato production
worldwide.

Management of TYLCV is difficult, expensive, limited, emphasizing
vector control - Multiple applications of insecticides. - Physical barriers (fine-mesh and/or UV-absorbing plastic screens).
Sus (control)
Sus (inoculated)

Breeding tomatoes that are resistant to the virus is considered highly effective in reducing yield
losses due to TYLCV.
מודבק מודבק
TY TY TY172
inoculated
Suceptible inoculated

All cultivated tomato accessions are susceptible to TYLCV - wild tomato
species were screened to identify and introgress resistance genes
.
מודבק מודבק
TY TY TY172
inoculated
S. chilense
S. pimpinellifolium
S. harbochaites
S. lycopersicum
Wild and domesticated Solanum species

- Ty- 1 and Ty- 3* from S. chilense on chromosome 6 Zamir et al. , 1 994; Ji and Scott, 2006; Ji et al. , 2007.
* Also effective against the bipartite ToMoV. * Ty- 1 and Ty- 3 are highly likely allelic (Verlaan et al. , 2012).
- Ty- 2 from S. habrochaites on Chromosome 11 (ToLCV?) Hanson et al. , 2000. - Ty- 4 from S. chilense on Chromosome 3 Ji et al. , 2008; 2009.
In addition several minor (relatively irreproducible) loci were identified on chromosomes 3 and 7 (Zamir et al. , 1994).
Tomato host loci identified and mapped

TY172 - Originated from 4 S. peruvianum accessions. - A tomato – BC3F6 with reduced “linkage drag”. - Usually symptomless upon inoculation with TYLCV. - Displays a recessive inheritance with 10-25% resistant plants in F2. - Friedmann et al. (1998) – TY172 resistance is controlled by 3 genes: 2 additive (1 partially dominant and 1 recessive); controlled epistatically by a third recessive gene.
M-82
TY172

TY172 - TY172 is resistant to Australian strain of ToLCV, Tomato yellow leaf curl Sardinia virus, Tomato yellow leaf curl Malaga virus, TYLCV-Almeria, and TYLCV-Mld (Lapidot M, unpublished). - Previous attempts to map its resistance genes failed due to lack of polymorphisms.

M 2528 P 284
ND 260 TOTAL 3072
Fingerprinting TY172 and M-82 with more than 3000 SNPs using illumina plex

Objective 1
To map genes controlling TYLCV resistance in TY172
Inoculated Non Inoculated

F1
Mapping populations
♀ TY172 X ♂ LA1589 (S. pimpinellifolium)
F2
F3 247 families 48 plants each (11,856)
I.
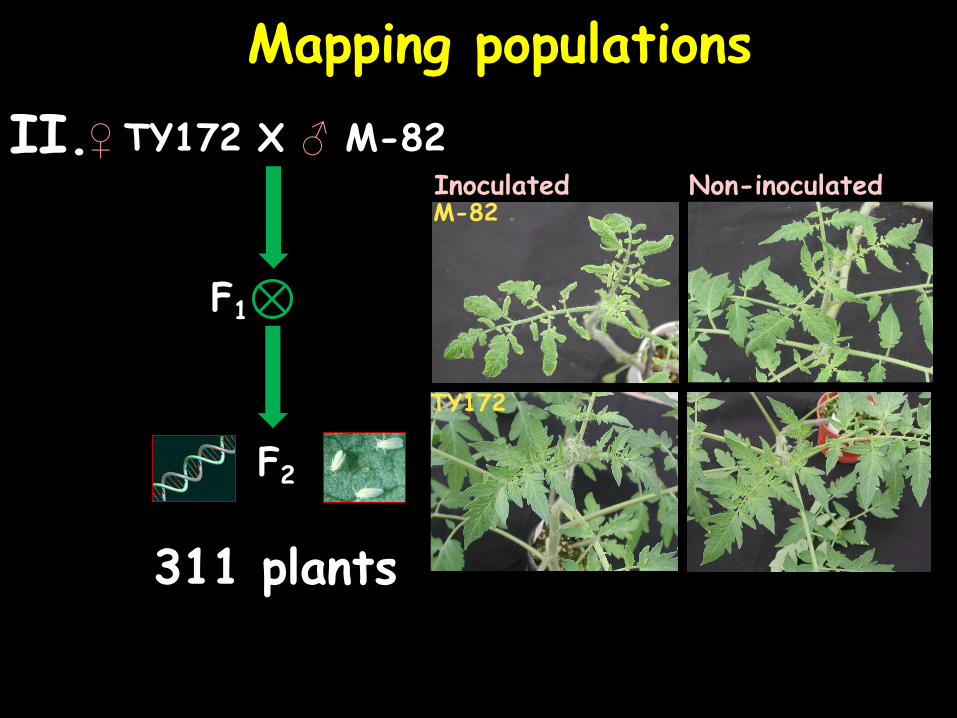
Mapping populations
♀ TY172 X ♂ M-82
F1
F2
II.
311 plants
Inoculated Non-inoculated M-82
TY172

Disease scale (DSI= Disease Severity Index)
Lapidot and Fridman, 2002
0 1
4 3
2

Major results
- 215 DNA markers (+ candidate genes). - 69 were polymorphic. - A major locus (termed ty- 5, Ch. 4 R2=39.7-46.6%, LOD= 33-35), and 4 minor loci (Ch. 1, 7, 9, 11, R2=12%, LOD~3) were found to control resistance in TY172.
Anbinder et al. (2009) Theor Appl Genet 119:519-530

Parental lines and their F1s
00.5
11.5
22.5
33.5
4
LA1589 TY172 X LA1589 TY172
עתוצ
ממ ה
יםומ
פטימ
הסת
מרחו
הדבקה ראשונההדבקה שניה
A A
BB
C C0
0.51
1.52
2.53
3.54
LA1589 TY172 X LA1589 TY172
עתוצ
ממ ה
יםומ
פטימ
הסת
מרחו
הדבקה ראשונההדבקה שניה
A A
BB
C C
00.5
11.5
22.5
33.5
4
M-82 TY172 X M-82 TY172
עתוצ
ממ ה
יםומ
פטימ
הסת
מרחו
C
B
A
00.5
11.5
22.5
33.5
4
M-82 TY172 X M-82 TY172
עתוצ
ממ ה
יםומ
פטימ
הסת
מרחו
C
B
A
Aver
age
DSI
Av
erag
e D
SI
1st inoculation 2nd inoculation
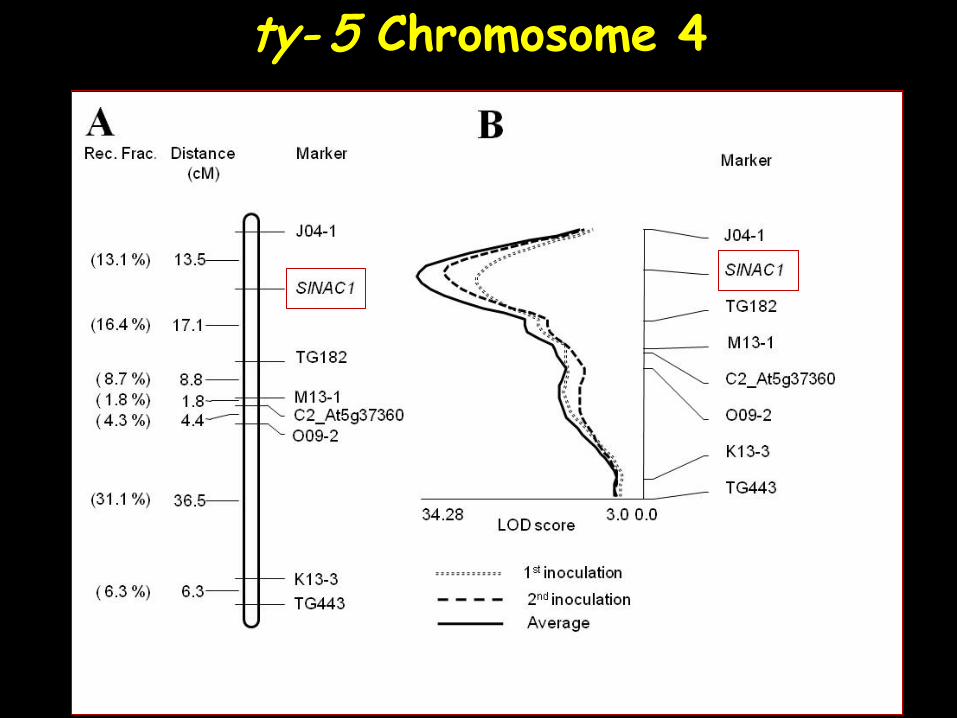
ty- 5 Chromosome 4

SlNAC1 (Selth et al. , 2005) - Encodes a member of the NAC-domain protein family. - Implicated in the replication of ToLCV, by interacting with the its replication enhancer protein (REn). - ToLCV and TYLCSV induce SlNAC1 expression specifically in infected susceptible cells, and this up-regulation requires REn. - Over-expression of SlNAC1 results in a substantial increase in viral DNA accumulation.
i.e. NAC1 may play an important role in the process by which REn enhances replication of ToLCV and possibly other
begomoviruses.

300 bp
400 bp
500 bp
LA1589 PKILPTQLAPMPPRPRSTPANDYFYFESSESMTRMHTTNSSSGSEHVLSPC 240 M82 PKILPTQLAPMPPRPRSTPANDYFYFESSESMTRMHTTNSSSGSEHVLSPC 240 TY172 PKILPTQLAPMPPRPRSTPANDCFYFESSESMTRMHTTNSSSGSEHVLSPC 240
Tyrosine212-to-Cysteine
ty- 5 (NAC1 ?)

500 bp 400 bp 300 bp
ty- 5 (NAC1 ?) could be traced back to the S. peruvianum progenitors of
TY172

NAC1 seemed preliminarily to be transcriptionally functional
NAC
1 ex
pres
sion

C
B
A
C
A A
Susceptible tail Resistant tail
ty- 5 (NAC1 ?)

0
0.5
1
1.5
2
2.5
SlNAC1 C2_At4g22260 SlSUMO
????
???
????
????
?? ?
????
SS
SR
RR C
B A
B B B B A A
0
0.5
1
1.5
2
2.5
NAC1 C2_At4g22260 SlSUMO
DSI
SS
SR
RR C
B A
B B B B A A
Minor resistance QTLs ♀ TY172 X ♂ LA1589

Genotype (SlNAC1 )
Fruit number
Fruit weight (kg)
Plant weight (kg)
Average fruit weight (gr)
Harvest index
Total mass (kg)
SS 55B 2.2B 1.09B 38.0A 2.1A 3.3B SR 62B 2.2B 1.17B 36.0A 2.0A 3.3B RR 84A 3.1A 2.09A 37.7A 1.7A 5.2A
♀ TY172 X ♂ M-82 F2

Summer 2010
Analysis of large F1, F2 and BC1F2 populations originating from crosses carried out, in reciprocal manner, between TY172 and the susceptible processing line M-82: (1) the minor QTLs we have previously identified are in effect not reproducible, and (2) ty- 5 alone can yield highly resistant plants with practically no extra-chromosomal effects.
VE ~ 0.1 VP ~ 0.5 VA ~ 0.5
h2 BS = (VP-VE)/VP = (0.5-0.1)/0.5 = 0.8 h2 NS = VA/VP = 0.5/0.5 = 1

Conclusions - ty- 5 is highly likely the sole major resistance locus in TY172 - increasing the likelihood of its successful cloning.
- The association between NAC1 and symptom severity at the ty- 5 locus and its role in enhancing the replication of begomoviruses merits its further study.

Objective 2
To identify and analyze the Tomato yellow leaf curl virus resistance gene ty- 5
Inoculated Non Inoculated

Populations
♀ TY172 X ♂ M-82
F1
F2
5662 segregating F2, F3, F4, BC1F1, BC1F2, BC1F3, BC1F4, BC1F5, BC2F2, BC2F3 and BC2F4 seedlings originating from TY172 X M-82 were inoculated with TYLCV and genotyped with 27 polymorphic markers spanning NAC1 .

Twenty seven markers Marker ~Distance from NAC1
(Kb) NAC1 0
31 6 22 6.7 41 7.6 34 8.7 25 18.5 1.8 33.2 RP5 184.3 RP3 186.5 7.4 218.7 DJ 220.7
RING 231.7 4.4 249.9
PELOTA 268.2 4.8 270.7 9.4 271.6 5.4 273.9 3.1 291.6 9.8 301.3 0.1 316.2 0.3 333.3 0.4 340.8 0.5 348.9 5.8 351 3.8 579.3 4.9 693
C2_At5g37360 43,552
- A 4394 bp genomic region spanning the NAC1 gene sequence in M-82 was blasted against the tomato WGS chromosomes (version SL2.40) at the Solanaceae Genomics Network (http://solgenomics.net/). - First, PCR primers were designed for sequences flanking the NAC1 gene region approximately every 50 Kbp, then every 10 Kbp and finally every 3 Kbp. Altogether, 257 800-900 bp fragments were sequenced.

Results
No contiguous introgression was observed spanning the ty- 5 locus but rather scattered islands of SNPs.
Genomic DNA sequence comparisons between TY172 and its susceptible counterparts did not yield significant insertions
or deletions usually characterizing DNA sequences of species distantly related to the cultivated tomato, including
S. peruvianum thought to be the origin of ty- 5.
S. peruvianum may not be the origin of ty- 5 or that TY172 already went through
considerable BC-breeding

50 plants displaying recombination events among the polymorphic markers developed were identified among the 5,662 segregating plants. These plants were allowed to self-pollinate in order to produce either segregating or non-segregating populations
that were again genotyped and inoculated.
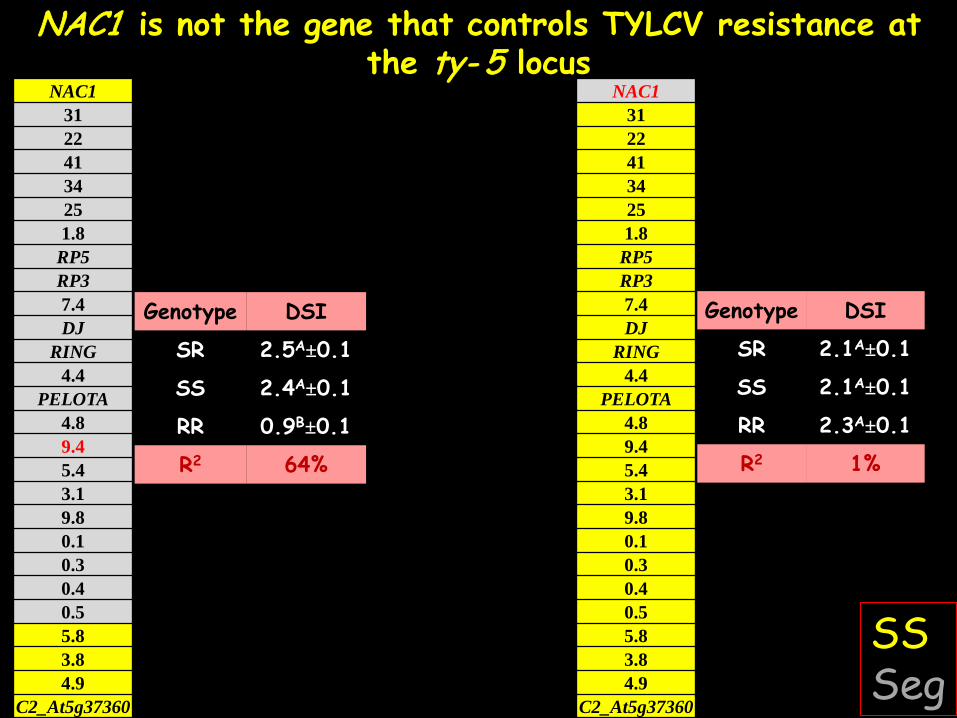
NAC1 is not the gene that controls TYLCV resistance at the ty- 5 locus
DSI Genotype
2.5A±0.1 SR
2.4A±0.1 SS
0.9B±0.1 RR
64% R2
NAC1 31 22 41 34 25 1.8
RP5 RP3 7.4 DJ
RING 4.4
PELOTA 4.8 9.4 5.4 3.1 9.8 0.1 0.3 0.4 0.5 5.8 3.8 4.9
C2_At5g37360
DSI Genotype
2.1A±0.1 SR
2.1A±0.1 SS
2.3A±0.1 RR
1% R2
NAC1 31 22 41 34 25 1.8
RP5 RP3 7.4 DJ
RING 4.4
PELOTA 4.8 9.4 5.4 3.1 9.8 0.1 0.3 0.4 0.5 5.8 3.8 4.9
C2_At5g37360
SS Seg

DSI Genotype
3.5A±0.1 SR
3.6A±0.1 SS
1.5B±0.1 RR
80% R2
NAC1 31 22 41 34 25 1.8
RP5 RP3 7.4 DJ
RING 4.4
PELOTA 4.8 9.4 5.4 3.1 9.8 0.1 0.3 0.4 0.5 5.8 3.8 4.9
C2_At5g37360
DSI Genotype
3.4A±0.1 SR
3.6A±0.1 SS
1.2B±0.2 RR
60% R2
NAC1 31 22 41 34 25 1.8
RP5 RP3 7.4 DJ
RING 4.4
PELOTA 4.8 9.4 5.4 3.1 9.8 0.1 0.3 0.4 0.5 5.8 3.8 4.9
C2_At5g37360
The ty- 5 gene was further narrowed down into a 23,250 bp fragment, including markers 4.4 and 4.8, containing two genes: a CALCIUM DEPENDENT PROTEIN
KINASE (SlCDPK2; Kamiyoshihara et al. 2010) and a PELOTA gene homolog (Becker et al. 2012)
SS RR Seg
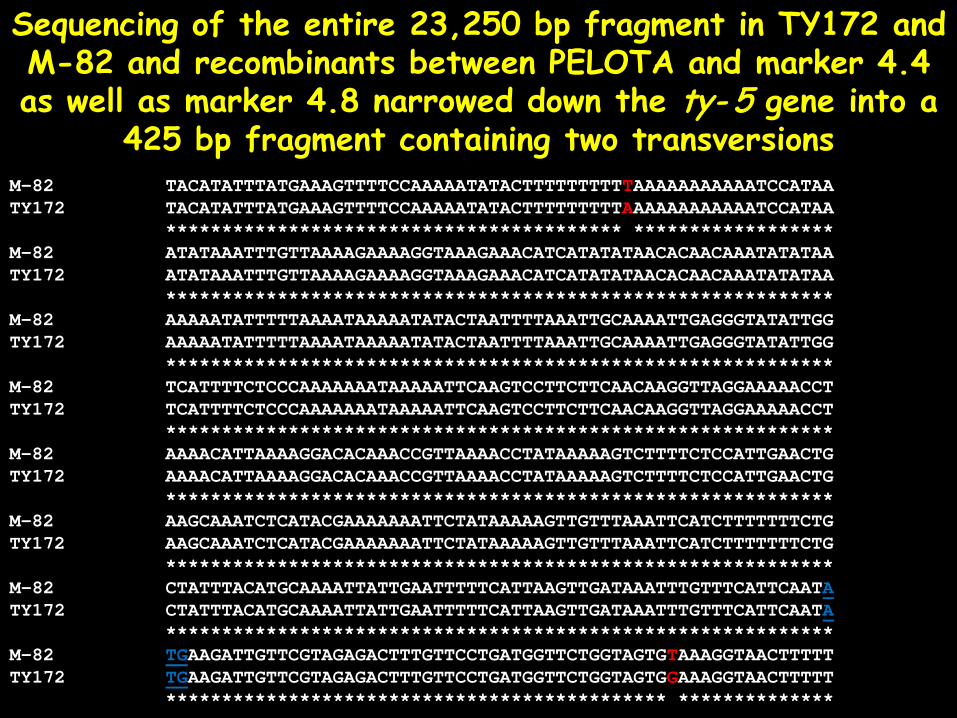
Sequencing of the entire 23,250 bp fragment in TY172 and M-82 and recombinants between PELOTA and marker 4.4 as well as marker 4.8 narrowed down the ty- 5 gene into a
425 bp fragment containing two transversions M-82 TACATATTTATGAAAGTTTTCCAAAAATATACTTTTTTTTTTAAAAAAAAAAATCCATAA TY172 TACATATTTATGAAAGTTTTCCAAAAATATACTTTTTTTTTAAAAAAAAAAAATCCATAA ***************************************** ****************** M-82 ATATAAATTTGTTAAAAGAAAAGGTAAAGAAACATCATATATAACACAACAAATATATAA TY172 ATATAAATTTGTTAAAAGAAAAGGTAAAGAAACATCATATATAACACAACAAATATATAA ************************************************************ M-82 AAAAATATTTTTAAAATAAAAATATACTAATTTTAAATTGCAAAATTGAGGGTATATTGG TY172 AAAAATATTTTTAAAATAAAAATATACTAATTTTAAATTGCAAAATTGAGGGTATATTGG ************************************************************ M-82 TCATTTTCTCCCAAAAAAATAAAAATTCAAGTCCTTCTTCAACAAGGTTAGGAAAAACCT TY172 TCATTTTCTCCCAAAAAAATAAAAATTCAAGTCCTTCTTCAACAAGGTTAGGAAAAACCT ************************************************************ M-82 AAAACATTAAAAGGACACAAACCGTTAAAACCTATAAAAAGTCTTTTCTCCATTGAACTG TY172 AAAACATTAAAAGGACACAAACCGTTAAAACCTATAAAAAGTCTTTTCTCCATTGAACTG ************************************************************ M-82 AAGCAAATCTCATACGAAAAAAATTCTATAAAAAGTTGTTTAAATTCATCTTTTTTTCTG TY172 AAGCAAATCTCATACGAAAAAAATTCTATAAAAAGTTGTTTAAATTCATCTTTTTTTCTG ************************************************************ M-82 CTATTTACATGCAAAATTATTGAATTTTTCATTAAGTTGATAAATTTGTTTCATTCAATA TY172 CTATTTACATGCAAAATTATTGAATTTTTCATTAAGTTGATAAATTTGTTTCATTCAATA ************************************************************ M-82 TGAAGATTGTTCGTAGAGACTTTGTTCCTGATGGTTCTGGTAGTGTAAAGGTAACTTTTT TY172 TGAAGATTGTTCGTAGAGACTTTGTTCCTGATGGTTCTGGTAGTGGAAAGGTAACTTTTT ********************************************* **************

Amino acid sequence of the PELOTA gene in the resistant TY172 line in comparison to its
susceptible counterparts (the Valine16-to-Glycine substitution in TY172 is in red)
M82 MKIVRRDFVPDGSGSVKIIPEEADDLWVAYNLIAEGDTVLAVTVRKVLREAASGGRDAER 60 TY-172 MKIVRRDFVPDGSGSGKIIPEEADDLWVAYNLIAEGDTVLAVTVRKVLREAASGGRDAER 60 *************** ******************************************** M82 VKLKLEIKVENVEYDKEGSALRIRGKNILENEHVKIGAFHTLEIEQHRPFVLRKVVWDSL 120 TY-172 VKLKLEIKVENVEYDKEGSALRIRGKNILENEHVKIGAFHTLEIEQHRPFVLRKVVWDSL 120 ************************************************************ M82 AREVLRQASDPSASADLAVVLMQEGLAHILLIGKSVTITRSRIESSIPRKHGPAIAGYDK 180 TY-172 AREVLRQASDPSASADLAVVLMQEGLAHILLIGKSVTITRSRIESSIPRKHGPAIAGYDK 180 ********************************************:*************** M82 ALNKFFDNVLQAFVKHVDFKVVRCAVIASPGFTKDQFHRHLLLEAERKQLRPIIENKSRI 240 TY-172 ALNKFFDNVLQAFVKHVDFKVVRCAVIASPGFTKDQFHRHLLLEAERKQLRPIIENKSRI 240 ************************************************************ M82 ILVHTTSGYKHSLKEVMDAPNVMTMIKDTKAAKEVQALKDFFNMLSNDPDRACYGPKHVE 300 TY-172 ILVHTTSGYKHSLKEVMDAPNVMTMIKDTKAAKEVQALKDFFNMLSNDPDRACYGPKHVE 300 ************************************************************ M82 VAHERLAIQTLLITDELFRSSDVETRKKYANLVDSVKDSGGTALIFSSMHVSGEQLNQLT 360 TY-172 VAHERLAIQTLLITDELFRSSDVETRKKYANLVDSVKDSGGTALIFSSMHVSGEQLNQLT 360 ************************************************************ M82 GIAAILRFPLPELEDIEM 378 TY-172 GIAAILRFPLPELEDIEM 378 ******************

Average relative transcript levels of PELOTA do not differ between resistant and susceptible plants
either inoculated or not
Days post inoculation Line Treatment n 7 14 21 28 TY172 Inoculated 10 2.2A±0.1 4.4A±0.9 1.1A±0.2 1.9A±0.2
Non-inoculated 10 2.1A±0.2 2.9A±0.4 1.1A±0.1 2.0A±0.3 R13 Inoculated 10 1.3A±0.4 2.3A±0.3 1.1A±0.2 1.4A±0.3
Non-inoculated 10 2.6A±0.5 2.6A±0.3 1.0A±0.2 1.6A±0.2
i.e. the SNP observed at the promoter region of PELOTA is not controlling resistance

Over-expression of PELOTA- M- 82 in TY172 renders this resistant line
susceptible Average DSI ± SE Average virus copy number ± SE
Line Transgenic Non-transgenic Transgenic Non-transgenic TYT-6 1.4A ± 0.2 0B ± 0 53,129A±4,714 15,113B±5,471 TYT-10 1.8A ± 0.1 0B ± 0 58,803A±6,400 16,612B±2331 TY172 - 0 ± 0 - 17,275±3728 R13 - 4 ± 0 - 82,408±7325
But its symptom buildup is slower

47 cm
Top view Side view
Non transgenic
25 cm
Transgenic

Over-expression of PELOTA-TY1 72 in R13 does not affect this
highly susceptible line Average DSI±SE
Line Season Transgenic Non-transgenic TYT-2 Spring 2.8A±0.2 2.2A±0.3 TYT-11 Summer 4.0A±0.0 4.0A±0.0 TY172 Spring - 0.1±0.1 R13 Spring - 3.4±0.1 TY172 Summer - 0.0±0.0 R13 Summer - 4.0±0.0
i.e. pelota- TY1 72 is a-loss-of-function allele

What is PELOTA
Ribosome-driven protein biosynthesis is comprised of four phases: initiation, elongation, termination and recycling and
PELOTA is implicated in the later phase as part of ribosome recycling complexes containing also an ABC-type
ATPase - ABCE1.
Interestingly, recessive resistance genes against plant viruses have been linked to components of the eukaryotic translation initiation complex (eIF4e and eIF4g protein
families).

What is PELOTA Recycling of ribosomes for a new round of translation initiation is essential to protein synthesis. ABCE1 can
dissociate ribosomes into subunits either after canonical termination by release factors (RF1) or after recognition
of stalled ribosomes by messenger RNA surveillance factors PELOTA, an eukaryotic RF1 (eRF1) paralog

Does the resistant PELOTA allele induce a yield penalty?
Yield components, per plant, of segregating non-inoculated BC3F2 plants in comparison to their recurrent parent M-82
Genotype Total fruit weight (Kg)
Fruit number (n)
Plant weight (gr)
M-82 3.1A±0.2 52A±3 511A±44 SS 2.7A±0.1 41B±2 510A±31 SR 1.9B±0.1 34BC±2 355B±31 RR 1.7B±0.1 33C±2 413AB±31

Conclusions - PELOTA is the gene that encodes TYLCV-resistance at the ty- 5 locus.
- PELOTA- TY1 72 is a loss-of-function allele (mutant) that induce yield penalty in non-infected plants.

Ty-1/3 in TY172
ty- 5 (R2=39%)
Ty- 1 /3 (UF_Ty- 3- P1 9) (R2=7%)
N DSI
SS SS 27 3.2A±0.2 SR SS 104 2.7AB±0.1 SS SR 70 2.4BC±0.1 SR SR 187 2.1CD±0.1 SS RR 43 2.0CD±0.2 SR RR 82 1.8D±0.2 RR SS 43 0.7E±0.1 RR SR 103 0.4E±0.0 RR RR 59 0.2E±0.0

Ty-1/3 in TY172
ty- 5 TY- 1 /3 (UF_Ty- 3- P1 9) N DSI R13 = Susceptible line 24 3.2A±0.2
SS SS 27 3.2A±0.2 SR SS 104 2.7AB±0.1
R13 X TY172 9 2.6AB±0.3 SS SR 70 2.4BC±0.1 SR SR 187 2.1CD±0.1 SS RR 43 2.0CD±0.2 SR RR 82 1.8D±0.2 RR SS 43 0.7E±0.1 RR SR 103 0.4E±0.0 RR RR 59 0.2E±0.0
TY172 12 0.0E±0.0
(+parental lines and their F1)

Acknowledgments
Funded by a Queensland (Australia) – Israel and by US – Israel Binational Agricultural Research and Development Fund (No. QB-9302-03 and IS-4409-11) to IL and ML.
- Ilana Anbinder - Uri Karniel - Doron Fogel - Dr. Moshe Lapidot - Dr. Moshe Reuveni - Dr. Dana Gelbart - Dr. Dalia Evenor - The late Dr. Shlomo Cohen - Sahadia Nahon - Haviva Shlomo - Lea Chen - Zion Machbash

The ribosome-bound PELOTA-ABCE1 complex
Becker et al. Nature 482: 501-506 2012

Scheme of archaeal and eukaryotic ribosome recycling bridging termination with initiation

Model of ribosome recycling in Archaea. After termination, ABCE1 is recruited to the 70S ribosome by binding to aRF1. ATP and ribosome binding induces a conformational switch into a closed state of ABCE1,
which powers ribosome dissociation and release of aRF1. Finally, ATP hydrolysis triggers the dissociation of ABCE1 from the 30S subunit. In addition, the conformational changes that drive 70S ribosome dissociation
(ATP-switch) and putative interaction sites with aRF1, 30S and RNA are illustrated.

Today - it is apparent that the name TYLCV has been given to a heterogeneous complex
of begomoviruses
Tomato yellow leaf curl disease (TYLCD) is induced by over 10 different viruses, mostly
monopartite


















