
TOLERANCE MECHANISMS IN CADMIUM-EXPOSED Eichhornia ... · land, which are now described as denuded,...
10
W1-06 1 TOLERANCE MECHANISMS IN CADMIUM-EXPOSED Eichhornia crassipes Mart. Solms, A PHYTOREMEDIATOR G.C. Rivero 1,2,3* , J.M. Puzon 1,2 , and H.P. Alcantara 1,2 1 Natural Sciences Research Institute and 2 Institute of Biology, College of Science, University of the Philippines, Diliman, Quezon City 1105, PHIL 3 DBSES, College of Science and Mathematics, University of the Philippines, Mindanao 8022, Philippines *Presenting author & author for correspondence: [email protected]; telefax (+632) 9205471 Abstract: Eichhornia crassipes (Mart.) Solms. plants were grown in nutrient solutions with 0.05, 0.5 and 5.0 mg/L CdCl 2 for 7 d. Chlorophyll content was significantly affected by leaf age and cadmium treatment levels. ICP-AES analyses of Cd and S contents revealed the localization of Cd in all subcellular components of the cells from young and mature laminae of Cd-exposed plants. The ultimate sequestration of Cd 2+ was in the vacuoles. The presence of S in the vacuoles indicated its role in cadmium chelation and sequestration. The sulfhydryl-containing peptides detected through DTNB assay exhibited peaks in the RP-HPLC chromatographs with retention times falling within the range of the GSH standard. The GSH-like peptide levels and Cd +2 concentrations showed a direct relationship and were found to be highest in Cd-treated mature laminae. Further characterization through CD and FTIR showed the GSH nature of the Cd-binding biomolecule. These observations revealed that the GSH-like peptide is the major Cd-chelating biomolecule in E. crassipes laminae. The enhanced synthesis of this peptide, possibly another form of GSH, as well as the ultimate retention of Cd in the vacuoles of the laminae, constitute the cellular mechanisms for Cd tolerance and homeostasis in E. crassipes leaves. The DTNB-reactive substances were then detected from the isolated protoplasts of young and mature laminae of Cd-exposed E. crassipes. ICP-AES analyses revealed that protoplasts from young and mature laminae of Cd-exposed plants contained significantly higher Cd 2+ ions than those from the control plants. The Cd 2+ that accumulated in the protoplasts from both young and mature laminae of the Cd-treated plants influenced the enhanced production of SH-containing compounds. Moreover, the young protoplasts from both the control and treated plants contained significantly higher levels of Cd 2+ , S 2 , and SH-containing compounds, indicating that young protoplasts were more capable of responding against Cd toxicity. The results of our studies provided an understanding of the efficacy by which E. crassipes plants accumulate high Cd levels. Our results provide information that explains why E. crassipes would be effective for Cd phytoremediation. Keywords: NIAES, cadmium, cadmium-binding biomolecules, Eichhornia crassipes, phytoremediation, vacuolar sequestration Introduction Cadmium is naturally present in soils at very low concentrations, i.e., slightly more than 1.0 mg kg -1 [17]. It is released into the environment by urban trafficking, “metal-working” industries, heating systems, power stations, cement factories, waste incinerators, and phosphate fertilizers. In the areas that have been subjected to mining, the concentration can be high, varying from 100-600 mg kg -1 dry weight. In particular, mining activities in the Philippines have resulted in massive environmental problems. The United Nations Environmental Programme (UNEP) records reveal that the Philippines is among one of the countries with insufficient mine tailings dam leading to the surface impoundments of the toxic waste from the mining process [28]. Sadly, disastrous consequences on the environment through time are borne by the local people. Since ours is a mineral- rich archipelago, the Philippine Government in 1992 has been pursuing plans to revitalize the mining industry [1]. The current regulation stipulates that mining be carried out in compliance with international standards and that environmental and social problems be addressed effectively. Rehabilitation efforts of polluted areas however, have been lagging behind extraction activities, leading to continuous dumping of toxic effluents into the surrounding ecosystems. One particular example is the Lafayette mining operations in Rapu-Rapu Island in the province of Albay in the Bicol Region, southern Luzon, which started mining gold, silver, copper, and zinc in April 2005 [2]. The poor implementation of environmental safeguards resulted in spills into the sea and around the island of cyanide and other contaminants from the mine tailings, resulting in massive fish kills after heavy rains. In 2006, a fishkill occurred in Mirikpitik Creek, one of the creeks leading out of the mine grounds [2]. The lower stretch of the creek was found to be affected by acid mine drainage. The waters in this section of the creek was found acidic, with the characteristic yellow solid precipitate ochre, composed of iron oxides and hydroxides. The creek is significantly impacted and has resulted in very high levels of heavy metals, particularly Cd, Cu, and Zn [2]. These metals were present in dissolved forms at levels which were several hundred times higher than typical background levels for these metals in the river water. The mine-impacted towns in Rapu-Rapu Island are mainly agricultural land, which are now described as denuded, barren, acidic, and unproductive due to open-pit mining, tunneling, and associated activities and by-products. Moreover, the marine life of the island has consequently been affected by varying amounts of these pollutants since aquatic systems inevitably serve as the dumping ground for mine wastes. It has been our interest for over a decade to evaluate how plant species thrive in polluted and stressful conditions. Reports are abundantly available on these plant species which may directly be integrated into the ecosystem or may serve as immobilizers of pollutants. E. crassipes, one of these plant species, may be a good phytoremediator due to its heavy metal- hyperaccumulating property, large biomass, and rapid proliferation. Yet, the mechanisms involving metal uptake by
Transcript of TOLERANCE MECHANISMS IN CADMIUM-EXPOSED Eichhornia ... · land, which are now described as denuded,...

W1-06
1
TOLERANCE MECHANISMS IN CADMIUM-EXPOSED Eichhornia crassipes Mart. Solms, A PHYTOREMEDIATOR
G.C. Rivero1,2,3*, J.M. Puzon1,2
, and H.P. Alcantara1,2
1Natural Sciences Research Institute and 2Institute of Biology, College of Science, University of the Philippines, Diliman, Quezon City 1105, PHIL
3DBSES, College of Science and Mathematics, University of the Philippines, Mindanao 8022, Philippines *Presenting author & author for correspondence: [email protected]; telefax (+632) 9205471
Abstract: Eichhornia crassipes (Mart.) Solms. plants were grown in nutrient solutions with 0.05, 0.5 and 5.0 mg/L CdCl2 for 7 d. Chlorophyll content was significantly affected by leaf age and cadmium treatment levels. ICP-AES analyses of Cd and S contents revealed the localization of Cd in all subcellular components of the cells from young and mature laminae of Cd-exposed plants. The ultimate sequestration of Cd2+ was in the vacuoles. The presence of S in the vacuoles indicated its role in cadmium chelation and sequestration. The sulfhydryl-containing peptides detected through DTNB assay exhibited peaks in the RP-HPLC chromatographs with retention times falling within the range of the GSH standard. The GSH-like peptide levels and Cd+2 concentrations showed a direct relationship and were found to be highest in Cd-treated mature laminae. Further characterization through CD and FTIR showed the GSH nature of the Cd-binding biomolecule. These observations revealed that the GSH-like peptide is the major Cd-chelating biomolecule in E. crassipes laminae. The enhanced synthesis of this peptide, possibly another form of GSH, as well as the ultimate retention of Cd in the vacuoles of the laminae, constitute the cellular mechanisms for Cd tolerance and homeostasis in E. crassipes leaves. The DTNB-reactive substances were then detected from the isolated protoplasts of young and mature laminae of Cd-exposed E. crassipes. ICP-AES analyses revealed that protoplasts from young and mature laminae of Cd-exposed plants contained significantly higher Cd2+ ions than those from the control plants. The Cd2+ that accumulated in the protoplasts from both young and mature laminae of the Cd-treated plants influenced the enhanced production of SH-containing compounds. Moreover, the young protoplasts from both the control and treated plants contained significantly higher levels of Cd2+, S2, and SH-containing compounds, indicating that young protoplasts were more capable of responding against Cd toxicity. The results of our studies provided an understanding of the efficacy by which E. crassipes plants accumulate high Cd levels. Our results provide information that explains why E. crassipes would be effective for Cd phytoremediation. Keywords: NIAES, cadmium, cadmium-binding biomolecules, Eichhornia crassipes, phytoremediation, vacuolar sequestration Introduction Cadmium is naturally present in soils at very low concentrations, i.e., slightly more than 1.0 mg kg-1 [17]. It is released into the environment by urban trafficking, “metal-working” industries, heating systems, power stations, cement factories, waste incinerators, and phosphate fertilizers. In the areas that have been subjected to mining, the concentration can be high, varying from 100-600 mg kg-1 dry weight. In particular, mining activities in the Philippines have resulted in massive environmental problems. The United Nations Environmental Programme (UNEP) records reveal that the Philippines is among one of the countries with insufficient mine tailings dam leading to the surface impoundments of the toxic waste from the mining process [28]. Sadly, disastrous consequences on the environment through time are borne by the local people. Since ours is a mineral-rich archipelago, the Philippine Government in 1992 has been pursuing plans to revitalize the mining industry [1]. The current regulation stipulates that mining be carried out in compliance with international standards and that environmental and social problems be addressed effectively. Rehabilitation efforts of polluted areas however, have been lagging behind extraction activities, leading to continuous dumping of toxic effluents into the surrounding ecosystems. One particular example is the Lafayette mining operations in Rapu-Rapu Island in the province of Albay in the Bicol Region, southern Luzon, which started mining gold, silver, copper, and zinc in April 2005 [2]. The poor implementation of environmental safeguards resulted in spills into the sea and around the island of cyanide and other contaminants from the mine tailings, resulting in massive fish kills after heavy rains. In 2006, a fishkill occurred in Mirikpitik Creek, one of the creeks leading out of the mine grounds [2]. The lower stretch of the creek was found to be affected by acid mine drainage. The waters in this section of the creek was found acidic, with the characteristic yellow solid precipitate ochre, composed of iron oxides and hydroxides. The creek is significantly impacted and has resulted in very high levels of heavy metals, particularly Cd, Cu, and Zn [2]. These metals were present in dissolved forms at levels which were several hundred times higher than typical background levels for these metals in the river water. The mine-impacted towns in Rapu-Rapu Island are mainly agricultural land, which are now described as denuded, barren, acidic, and unproductive due to open-pit mining, tunneling, and associated activities and by-products. Moreover, the marine life of the island has consequently been affected by varying amounts of these pollutants since aquatic systems inevitably serve as the dumping ground for mine wastes. It has been our interest for over a decade to evaluate how plant species thrive in polluted and stressful conditions. Reports are abundantly available on these plant species which may directly be integrated into the ecosystem or may serve as immobilizers of pollutants. E. crassipes, one of these plant species, may be a good phytoremediator due to its heavy metal-hyperaccumulating property, large biomass, and rapid proliferation. Yet, the mechanisms involving metal uptake by

W1-06
2
hyperaccumulating plant species, such as E. crassipes, still remain poorly understood. The efficiency of a plant species involved in the remediation of polluted environment is dependent on the understanding of the physiological basis of the metabolic action or processes responsible for the uptake and subsequently, tolerance of the heavy metals. To date, studies still remain inadequate on the specific biochemical mechanisms underlying the morphological and physiological responses which enable E. crassipes to accumulate and tolerate the toxic metal, Cd. Thus, our studies have been to primarily address these aspects, important in assessing the efficacy of E. crassipes as a Cd phytoremediator. Our work on Cd and E. crassipes involved the following: (1) assessment of the morphological effects of Cd uptake; (2) determination of the subcellular localization of accumulated Cd in the leaves of the plant; (3) detection of Cd-binding biomolecules in the leaf tissues; (4) characterization of Cd-binding biomolecules in the leaf tissues; and, (5) detection of DTNB-reactive biomolecules from isolated protoplasts of the leaf. The characterization of the Cd-binding biomolecules are still partial. We are pursuing the chemical characterization of these biomolecules and the evaluation of crucial enzymes in their metabolism. Materials and Methods A. Plant Collection, Cultivation, Treatment, and Harvest E. crassipes plants were maintained at the Botanical Gardens II, Institute of Biology, College of Science, U.P. Diliman. Healthy, vigorous plantlets from common stolons served as the source plants. From these, 48 plants of relatively the same age were transferred to the Institute of Biology Plant Physiology Laboratory. A half-strength Hoagland [9] solution was modified with CdCl2 (Baker cadmium chloride 2.5 hydrate crystals; 80% A.C.S. specifications), to prepare CdCl2 concentrations. The unmodified half-strength Hoagland solution served as the control. Twelve plastic basins (38cm x 27cm x 35cm) were filled with 6 L of the different solutions. Each basin contained 4 plants. Three replicates for each treatment were maintained under continuous illumination (40-W fluorescent tubes, 30 µEm-2s-1) with constant aeration to ensure healthy plants [11, 12]. After 7 days of treatment, the plants were harvested; the laminae were classified as to young and mature on the basis of their position in the plant [19]. B. Evaluation of the Subcellular Localization of Cd2+ in the Leaves Differential Centrifugation The method of Hernandez et al. [8] for the differential centrifugation of subcellular fractions was adopted. The young and mature laminae from E. crassipes plants treated with 5.0 ppm CdCl2 have been reported to accumulate Cd as determined by flame atomic absorption spectophotometry (FAAS) [19]. These were selected and harvested for the subcellular localization of Cd. Isolation and Purification of the Protoplasts, Vacuoles, and Vacuolar Components The method of Vogeli-Lange and Wagner [26] was used to isolate and purify the protoplasts and vacuoles from tissues of young and mature E. crassipes leaves. The vacuoles were subsequently isolated using the modified method of Matoh et al. [15]. Aliquots of these protoplasts were used for inductively coupled plasma - atomic emission spectrometry (ICP-AES) Cd analysis and subsequent isolation of vacuoles, samples of which were subjected to phosphatase assay. Vacuoles were disrupted by sonication in a bath sonifier (Branson Sonifier 450) and were subsequently frozen. After thawing, tonoplast membranes were sedimented by centrifugation at for 1 h at 40oC (100,000 x g Beckman Coulter Optima TM U-90k ultracentrifuge). The supernatant and the pellets were immediately assayed for Cd content by ICP-AES. C. Detection of the Cadmium-Binding Biomolecules through DTNB Assay The method of Grill et al. [7] for gel filtration chromatography as modified by Kubota et al. [14] and Josue et al. [12] was adopted. Samples derived thereafter were subjected separately to ICP-AES analysis, 5,5’-dithiobis(2-nitro-benzoic acid) (DTNB) assay, and further spectral elucidation of Cd-binding peptides. D. Isolation and Characterization of the SH-containing, Cadmium-Binding Biomolecules The lyophilized gel filtration fractions that showed the highest detected SH-containing biomolecules and highest absorbance at 254 nm were used to isolate the Cd-binding peptides and were subsequently subjected to Reversed Phase-High Pressure Liquid Chromatography (RP-HPLC), Circular Dichroism (CD) Spectrophotometry, and Fourier Transformed- Infrared (FT-IR) Spectrometry.

W1-06
3
Results and Discussion
Morpho-physiological effects of Cd An initial morpho-physiological study was conducted to assess the uptake and accumulation of Cd2+ in all young and mature organs of E. crassipes, i.e., roots, stems, petioles, and laminae [19]. AAS analyses revealed that significantly high Cd levels were taken up by various vegetative organs of E. crassipes. Based on linear regression analysis, Cd levels increased with an increase in concentration, as well as, prolonged Cd exposure. Generally, mature vegetative organs took up greater amounts of cadmium compared with the younger organs. The chlorophyll contents of young and mature laminae significantly changed with varying cadmium concentrations and age (Table 1). The total chlorophyll contents of young laminae significantly increased on the 7th day of cadmium exposure at all levels (Table 1). The total chlorophyll contents of mature laminae significantly decreased in both cadmium-treated and control plants (Table 1). The overall total chlorophyll content levels of the mature laminae had relatively lower contents when compared to those of young laminae. This was attributed to the near senescent state of the mature laminae [4, 13]; whereas, the young laminae tissues were more photosynthetically active. The effect then of cadmium toxicity on the total chlorophyll contents was more pronounced in the mature than in the young laminae, the reduction in the mature laminae being by 20-94%. Exposure of plants to cadmium may have possibly altered certain enzymes required for chlorophyll production. Thus, the amount of total chlorophyll in the laminae of cadmium-treated plants significantly decreased. The chl a, chl b, and total chlorophyll contents of the young laminae however, generally increased after 7 days. This is a compensating mechanism for tolerance. B. Subcellular Localization of Cadmium The results of the ICP-AES analyses of cadmium contents (Table 2) demonstrated the mobility and localization of Cd2+ in all subcellular fractions, i.e., Fraction I containing the cell wall materials, Fraction II containing the nuclei, mitochondria, chloroplasts, Fraction III containing the microsomes, and Fraction IV containing the cytosol. The results also revealed that more Cd2+ ions were accumulated in subcellular fractions and components of the mature laminae, compared with those derived from the young laminae. In 5.0 ppm Cd-exposed plants, the Cd2+ level in the cell wall-containing fraction of the young laminae was only 5.9% relative to the Cd2+ level in the corresponding fraction from the mature laminae. The fraction containing nuclei, mitochondria, and chloroplasts exhibited 13.5%, compared to the Cd2+ level in young laminae of the corresponding fraction in the mature laminae. The Cd2+ accumulated in the cytosol-containing fraction of young laminae was only 7.3%, compared to the Cd2+ level accumulated in the corresponding fraction of mature laminae. The combined Cd contents of the protoplasmic components, i.e., Fractions II, III, and IV had values of 0.073 and 0.625 mg/kg in young and mature laminae, respectively. These values represented 57.5% and 40.6% of the combined Cd contents of all subcellular fractions in young and mature laminae, respectively. These results indicate that more than 50% of the Cd contents were localized in the cell walls of mature laminae; while more than 50% Cd were localized in the protoplasmic fractions of young laminae. Mature leaves possess thickened, specialized cell walls due to the large deposition of lignin in the secondary walls of xylem cells. These lignified cell walls, as well as, the pectin in the middle lamella [25] adsorbed more Cd ions. On the other hand, young leaves serve as “sinks” to the various metabolic activities, including the nondiscriminatory uptake of non-essential ions in the cytosol, like the toxic heavy metals [12]. Higher levels of Cd were localized in all subcellular fractions and components of mature laminae, compared to the young counterpart. The combined Cd contents of all subcellular fractions, i.e., Fraction I, II, III, and IV (Table 2) had values of 0.127 and 1.539 mg/kg in young and mature laminae, respectively. Moreover, vacuoles and their vacuolar components isolated from mature laminae exhibited higher Cd2+ levels than those from young laminae (Table 3). No Cd2+ ions were detected in the tonoplasts of vacuoles from Cd-exposed young laminae. This could be explained by the presence of larger, more developed, specialized vacuoles in mature laminae tissues. In young tissues, vacuoles are found to be less prominent, usually present as small provacuoles, with less developed tonoplasts [25]. Higher Cd2+ levels were localized in the vacuoles isolated from mature laminae since in mature tissues, a single, central vacuole occupies up to 95% of a mature plant cell volume. Further, specialized protein vacuoles in mature tisues, called protein bodies, serve as storage of proteins, hydrolytic enzymes, peptides, and amino acids, possibly including Cd-binding biomolecules in E. crassipes laminae. The results of ICP-AES analyses of Cd and S contents (Table 4) generally demonstrate the localization of Cd in all subcellular components of the cells from young and mature laminae of 5.0 ppm Cd-exposed E. crassipes plants, and the ultimate sequestration of Cd ions in the vacuoles, together with S-containing compounds. The presence of S in the vacuoles indicates its role, as well as those of other S-containing compounds, in the chelation and sequestration of Cd into the vacuoles [3]. The S-containing tripeptide GSH has been reported to be transported into the vacuoles [21] possibly with Cd in the case of E. crassipes.

W1-06
4
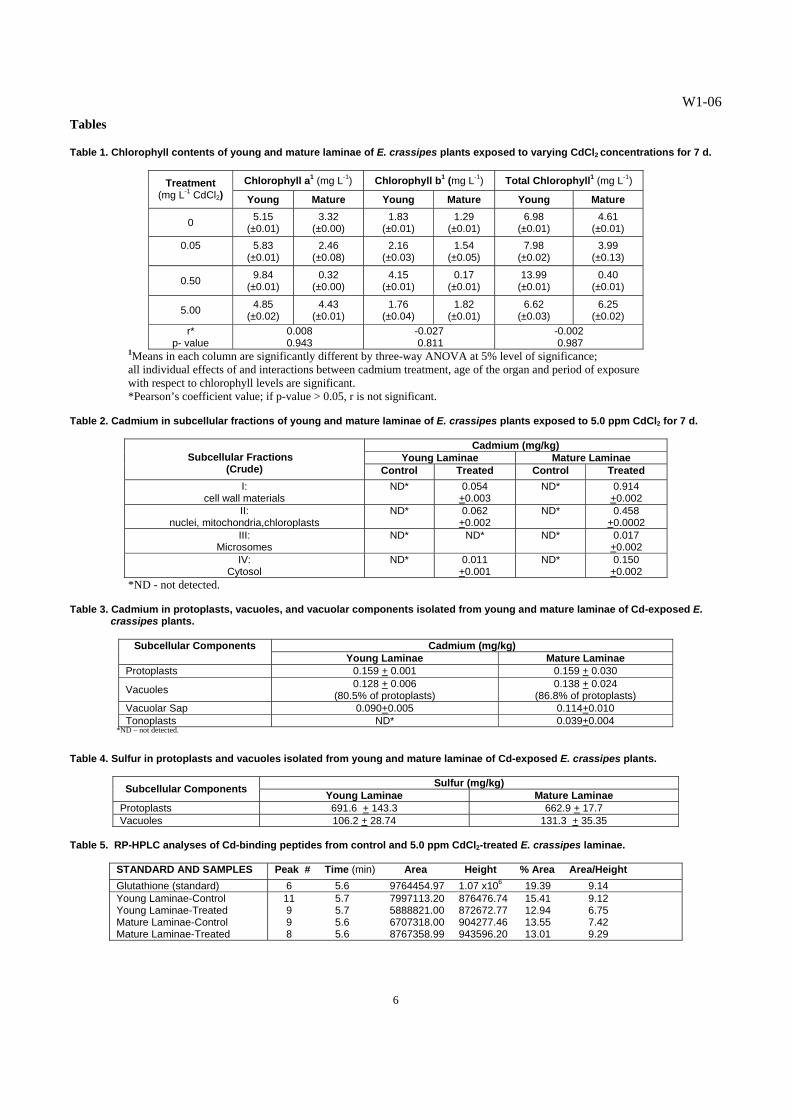
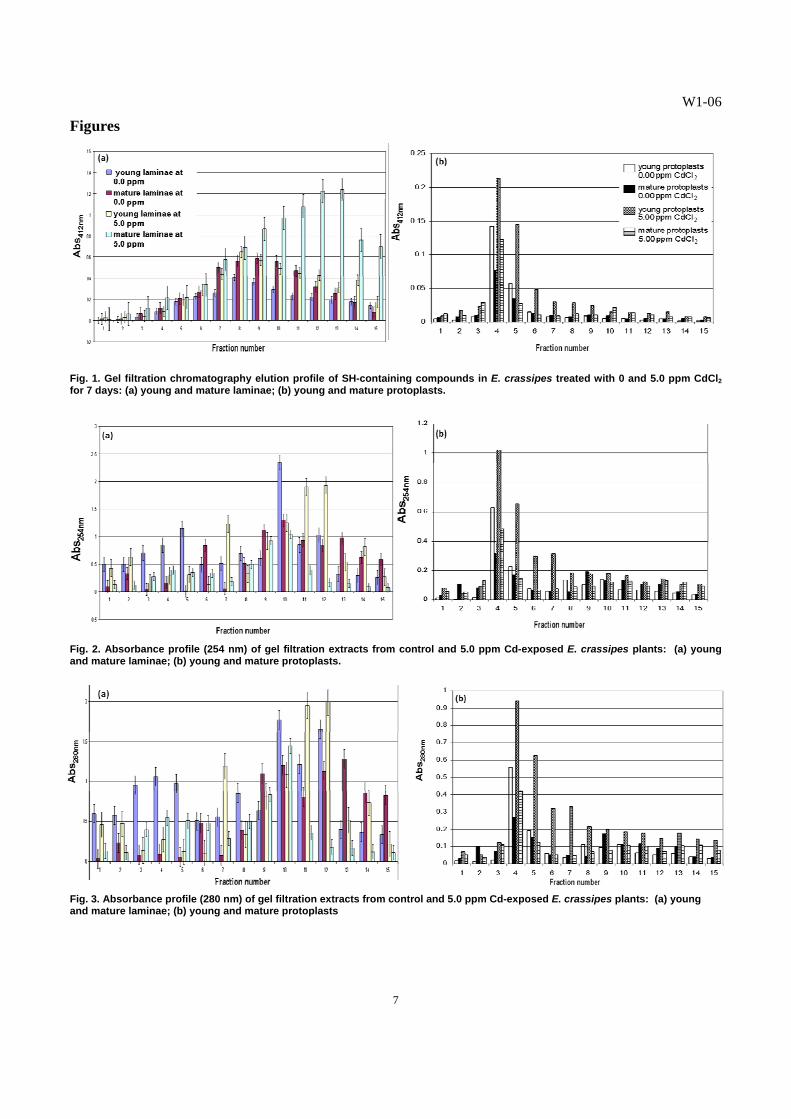
C. Detection of Cadmium-Binding Biomolecules1 Higher amounts of SH-containing biomolecules were detected in the young and mature laminae of treated plants, compared with those in the young and mature laminae of the control (Fig. 1a). The isolated young protoplasts from the laminae of the Cd-exposed E. crassipes plants displayed significantly higher amounts of SH-containing compounds, as indicated by higher elution profiles for all the fractions tested, peaking at fractions 4 and 5 (Fig. 1b). The mature protoplasts from both the Cd-treated and control laminae contained the least amounts of SH-containing compounds. The Cd2+ that accumulated in the vegetative organs influenced the production of SH-containing compounds in E. crassipes. In Cd-treated plants, young organs were found to accumulate lower amounts of Cd2+ than mature organs. The SH-containing compounds, like glutathione and phytochelatins, protect plant cells from the interference of the toxic heavy metals that may have gained entry into the protoplasm [11, 12]. The results indicate that more SH-containing substances were synthesized when the plants were exposed to the toxic heavy metal. It has been established in many plant species that soon as Cd enters the roots and is translocated in the leaves, Cd stimulated the synthesis of specialized SH-containing biomolecules involved in Cd chelation and sequestration into the vacuoles [18]. In this study, the enhanced synthesis of SH-containing biomolecules seemed to be one of the major defense strategies of E. crassipes in coping with Cd toxicity. Exposure to Cd would induce an “additional sink” increasing the need for thiol compounds by cells [16]. The results on the absorbance readings of the gel filtration fractions at 254 nm (Fig. 2a) suggest that young leaves from control and treated plants contain more S-metal complexes, compared to the fractions from the mature leaves. The fractions obtained from the gel filtration chromatography of the isolated young and mature protoplasts from the laminae of E. crassipes plants were also examined for S-metal complexes at 254 nm (Fig. 2b). The young protoplasts from the Cd-treated E. crassipes plants exhibited the highest absorbance profile at fractions 4 and 5 indicating the highest amounts of S-metal complexes. This was followed by the young protoplasts from the control E. crassipes plants, with the peak at fraction 4 (Fig. 2b). Least amounts of S-metal complexes were noted in all mature protoplasts of E. crassipes. The presence of more sulfur-metal complexes in young leaves is possibly linked to higher rates of photosynthesis, as reflected by higher chlorophyll content in young leaves, probably resulting from higher metabolic activity of the ferredoxin-thioredoxin system, which are Fe-S proteins exhibiting the S-S and SH groups, respectively [25]. Moreover, the S-containing, Cd-binding peptides, like GSH, MTs and PCs, contain Cys sulfhydryl residues which bind and sequester available Cd2+ into stable complexes, resulting to high absorbance at 254 nm [4]. The fractions obtained from the laminae (Fig. 3a) and isolated protoplasts (Fig. 3b) of treated plants which exhibited high absorbance readings at 280 nm may contain biomolecules or peptides, which may have been synthesized by E. crassipes plants in response to cadmium stress, thus adding support to the observed increase in SH-containing peptides in the Cd-treated plants. D. Partial Characterization of Cadmium-Binding Biomolecules1 The RP-HPLC chromatographs of glutathione (GSH) standard and gel filtration fractions from young and mature laminae of control and Cd-exposed plants are presented in Figs. 4a-e, respectively. The GSH standard used in the RP-HPLC runs had a retention time of 5:56 min (Fig. 4a). The profiles of extracts from laminae of control and 5.0 ppm Cd-exposed plants reflected a peak at the range of 5:58 to 5:72 min retention time, indicating the presence of GSH-like peptides. The GSH-like substances were concentrated in the laminar part of E. crassipes leaves as evidenced by the display of peaks in these RP-HPLC chromatographs (Figs. 4b-e). The GSH-like substances and other Cd-binding substances may be involved in the translocation of Cd2+ from the roots up to the shoot of the plant, rather than existing as a relatively static means to sequester Cd2+ [6, 24]. Glutathione-like substances were also produced mainly in the leaves of Ipomoea aquatica exposed to mercury and were reported to mediate Hg-binding and detoxification in this plant [11, 12]. The differential responses to Cd treatment, with respect to the area/height values, obtained from the fractions of the laminae are age-dependent. The high area/height value of the major peak of fractions (Table 5) obtained from mature laminae of Cd-treated plants suggests a Cd-enhanced synthesis of a GSH-like substance. In contrast, there was no apparent increase in the rate of synthesis of GSH-like substance in the young laminae of Cd-exposed plants, as compared to that of the control. These results demonstrate the concomitant increase in the rate of GSH synthesis, in response to an increase in Cd accumulation in E. crassipes leaves, as evidenced by the higher Cd level in the mature laminae, as compared to that of young laminae. Moreover, the higher concentration of GSH-like substance in the young laminae of the control, relative to those of Cd-treated plants, suggests the involvement of GSH in the regulation of many enzymes associated with photosynthetic metabolism, specifically related to the ferredoxin-thioredoxin system [18]. The general activation of sulfur metabolism induced by Cd2+ is a likely consequence of the increase in demand for SH-containing peptides [16, 27, 29], like the change in GSH levels in Cd-exposed E. crassipes. The CD spectra of the SH-containing extracts confirm the GSH nature of the cadmium-binding biomolecule, reflecting a strong absorbance in the far-UV to UV region, [10, 20], i.e., 200-240 nm in E. crassipes. The bands in the spectra of the extracts from the laminae of E. crassipes resembled the bands present in the spectrum of the GSH standard (Fig. 5a), more
1Submitted for publication, The Philippine Agricultural Scientist

W1-06
5
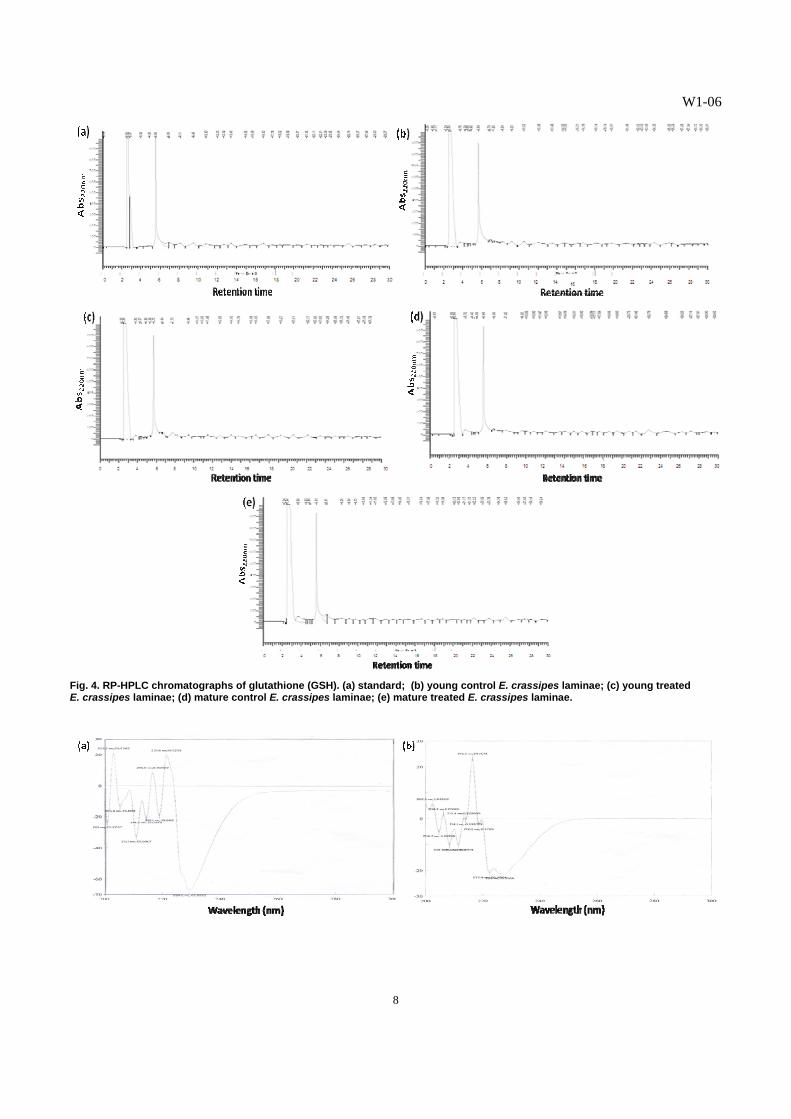
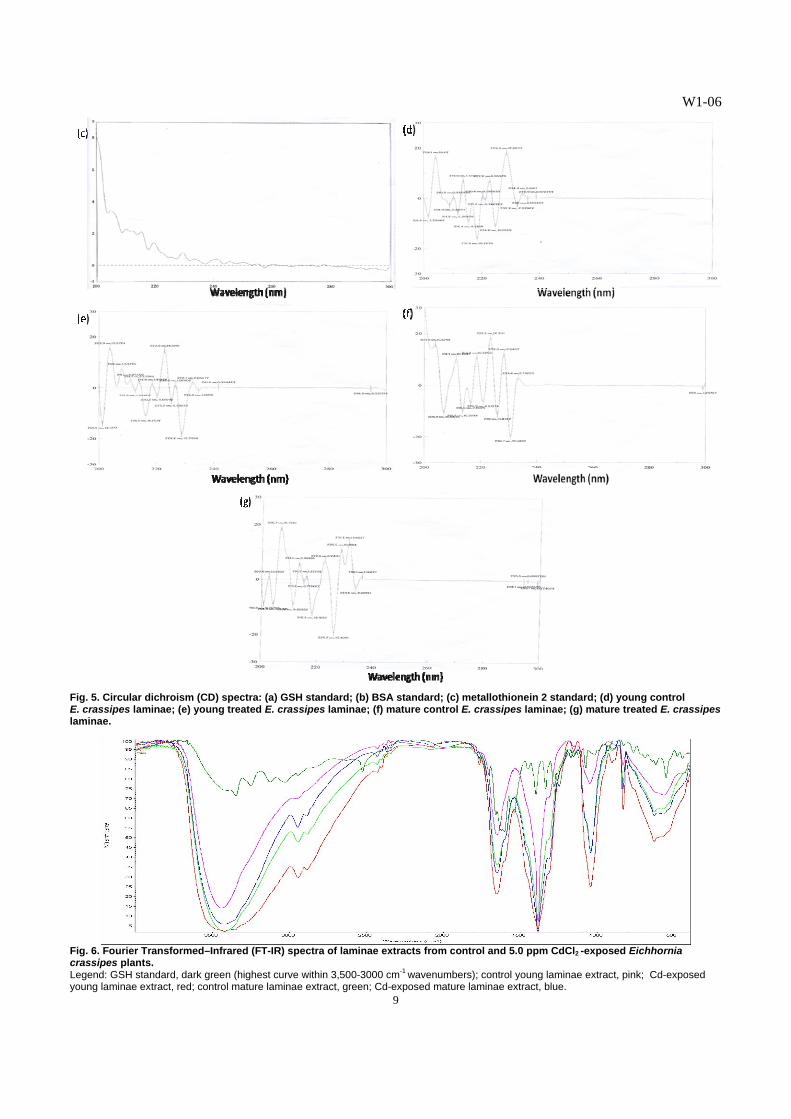
than those of the BSA (Fig. 5b) and metallothionein 2 (Fig. 5c) standards. The spectrum of the GSH standard exhibited corresponding peaks and troughs found in spectra of the various extracts from the laminae (Figs. 5d-e), indicating that the Cd-binding substance isolated from E. crassipes leaves exhibited GSH-like characteristics. However, some peaks and troughs in the spectra of the extracts were not found in the GSH spectrum. We may be looking at a different form of GSH. The presence of minor bands in the vicinity of the 300 nm region and within the 235-240 nm region of the spectra derived from the extracts of the young and mature laminae of Cd-treated plants suggest the presence of Cd-induced, sulfur-containing substances in E. crassipes, possibly GSSG and other substances with S=S bonds. The disulfide chromophores absorb maximally within these regions [14]. The results also indicate that the CD spectrum of Cd-binding peptide in E. crassipes in the far-UV at wavelengths, ca. 210 nm, revealed metal binding, as determined by different spectroscopic studies [20]. The FT-IR spectra of the purified GSH-like substances isolated from the laminae demonstrated patterns of transmittance similar to the GSH spectrum (Fig. 6), although that of the GSH standard had more number of peaks and more peaks at higher % transmittance in the regions of 500-3,000 cm-1 wave numbers. The bonds present in the peptide, viz., C-N, C=O, and N-H, reflect absorptions in these regions [22, 23]. The transmittance patterns displayed by the four spectra of GSH-like substances from the laminae revealed close resemblance to each other in the regions from 2,500 to 3,400 cm-1. The bonds present in the peptide, specifically N-H, absorb maximally within this region of wave numbers [22, 23]. The results obtained from the FT-IR spectra of GSH-like peptides from the laminae of E. crassipes (Fig. 6) corresponded with the data on GSH concentration obtained from the RP-HPLC (Table 5). In the FT-IR spectra of the different extracts from the laminae, the GSH-like substances from the young laminae of control plants and that from the mature laminae of Cd-treated plants had the highest transmittance values, followed by that of the GSH-like substance from the mature laminae of control plants. The lowest transmittance values were displayed by the spectrum of GSH-like substance from the young laminae of Cd-treated plants. Likewise, the RP-HPLC data revealed the highest concentration of GSH-like substance in the young laminae of control plants and in the mature laminae of Cd-exposed plants; followed by that of the mature laminae from control plants; and, the least amount of GSH-like substance in the young laminae of Cd-treated plants. These results revealed that the SH-containing Cd-binding peptide in the laminae of E. crassipes plants resembled the spectral characteristics of GSH. This GSH-like peptide is the major chelating biomolecule for cadmium, and may be involved in the sequestration of this heavy metal into the vacuoles of E. crassipes leaves.
Summary and Conclusions
Eichhornia crassipes accumulated the heavy metal Cd in all subcellular fractions of the laminae, but higher levels were noted in mature laminae than in young laminae. The results also demonstrated the ultimate sequestration of Cd2+ in the vacuoles of the laminae, with higher levels present in mature laminae than in young laminae. The presence of S in the vacuoles indicated the probable role of S-containing substances in the chelation and sequestration of Cd into the vacuoles. The SH-containing substances were concentrated in the laminae of E. crassipes leaves. The levels of GSH-like biomolecule showed a direct relationship with the levels of Cd in the vacuoles, as well as Cd contents of the SH-containing gel filtration fractions. The amplified production of DTNB-reactive substances, i.e., SH-containing compounds in the protoplasts of the Cd-exposed E. crassipes plants and the subsequent binding of Cd2+ ions to these compounds, is responsible for the Cd tolerance exhibited by E. crassipes. The sulfhydryl-containing biomolecule detected through DTNB assay exhibited RP-HPLC chromatographs with retention times falling within the range of that reflected by the GSH standard. Further characterization through CD spectrophotometry and FT-IR spectrometry showed the GSH nature of the SH-containing Cd-binding peptide. The CD spectra of the extracts from the laminae of E. crassipes resembled more the spectrum of the GSH standard, than those of the metallothionein 2 and BSA standards. The CD spectrum of the GSH standard exhibited corresponding peaks and troughs found in spectra of the various extracts from the laminae, indicating the GSH-like nature of the Cd-binding substance isolated from E. crassipes leaves. The FT-IR spectra of the GSH-like substances isolated from the laminae demonstrated patterns of transmittance similar to the GSH spectrum, although that of the GSH standard had more number of peaks and more peaks at higher % transmittance in the regions of 3,000-500 cm-1 wave numbers. These observations revealed a GSH-like biomolecule as the major chelating agent for Cd in E. crassipes. The results herein provide a biological baseline and initial biochemical analyses which lends understanding to the efficacy by which E. crassipes accumulate high levels of Cd without manifesting impaired morpho-physiological symptoms, making it thus an effective phytoremediator. The elucidation and complete characterization of these Cd-biomolecules and the evaluation of enzyme synthesis in the Cd-accumulating E. crassipes are the subjects of the on-going research activities by the authors.
W1-06
6
Tables Table 1. Chlorophyll contents of young and mature laminae of E. crassipes plants exposed to varying CdCl2 concentrations for 7 d.
Chlorophyll a1 (mg L-1) Chlorophyll b1 (mg L-1) Total Chlorophyll1 (mg L-1) Treatment (mg L-1 CdCl2) Young Mature Young Mature Young Mature
0 5.15 (±0.01)
3.32 (±0.00)
1.83 (±0.01)
1.29 (±0.01)
6.98 (±0.01)
4.61 (±0.01)
0.05
5.83 (±0.01)
2.46 (±0.08)
2.16 (±0.03)
1.54 (±0.05)
7.98 (±0.02)
3.99 (±0.13)
0.50 9.84 (±0.01)
0.32 (±0.00)
4.15 (±0.01)
0.17 (±0.01)
13.99 (±0.01)
0.40 (±0.01)
5.00 4.85 (±0.02)
4.43 (±0.01)
1.76 (±0.04)
1.82 (±0.01)
6.62 (±0.03)
6.25 (±0.02)
r* p- value
0.008 0.943
-0.027 0.811
-0.002 0.987
1Means in each column are significantly different by three-way ANOVA at 5% level of significance; all individual effects of and interactions between cadmium treatment, age of the organ and period of exposure with respect to chlorophyll levels are significant. *Pearson’s coefficient value; if p-value > 0.05, r is not significant.
Table 2. Cadmium in subcellular fractions of young and mature laminae of E. crassipes plants exposed to 5.0 ppm CdCl2 for 7 d.
Cadmium (mg/kg) Young Laminae Mature Laminae
Subcellular Fractions
(Crude) Control Treated Control Treated I:
cell wall materials ND* 0.054
+0.003 ND* 0.914
+0.002 II:
nuclei, mitochondria,chloroplasts ND* 0.062
+0.002 ND* 0.458
+0.0002 III:
Microsomes ND* ND* ND* 0.017
+0.002 IV:
Cytosol ND* 0.011
+0.001 ND* 0.150
+0.002 *ND - not detected.
Table 3. Cadmium in protoplasts, vacuoles, and vacuolar components isolated from young and mature laminae of Cd-exposed E.
crassipes plants.
Cadmium (mg/kg) Subcellular Components Young Laminae Mature Laminae
Protoplasts 0.159 + 0.001 0.159 + 0.030
Vacuoles 0.128 + 0.006 (80.5% of protoplasts)
0.138 + 0.024 (86.8% of protoplasts)
Vacuolar Sap 0.090+0.005 0.114+0.010 Tonoplasts ND* 0.039+0.004
*ND – not detected.
Table 4. Sulfur in protoplasts and vacuoles isolated from young and mature laminae of Cd-exposed E. crassipes plants.
Sulfur (mg/kg) Subcellular Components Young Laminae Mature Laminae Protoplasts 691.6 + 143.3 662.9 + 17.7 Vacuoles 106.2 + 28.74 131.3 + 35.35
Table 5. RP-HPLC analyses of Cd-binding peptides from control and 5.0 ppm CdCl2-treated E. crassipes laminae.
STANDARD AND SAMPLES Peak # Time (min) Area Height % Area Area/Height Glutathione (standard) 6 5.6 9764454.97 1.07 x106 19.39 9.14 Young Laminae-Control Young Laminae-Treated Mature Laminae-Control Mature Laminae-Treated
11 5.7 7997113.20 876476.74 15.41 9.12 9 5.7 5888821.00 872672.77 12.94 6.75 9 5.6 6707318.00 904277.46 13.55 7.42 8 5.6 8767358.99 943596.20 13.01 9.29
W1-06
7
Figures
Fig. 1. Gel filtration chromatography elution profile of SH-containing compounds in E. crassipes treated with 0 and 5.0 ppm CdCl2 for 7 days: (a) young and mature laminae; (b) young and mature protoplasts.
Fig. 2. Absorbance profile (254 nm) of gel filtration extracts from control and 5.0 ppm Cd-exposed E. crassipes plants: (a) young and mature laminae; (b) young and mature protoplasts.
Fig. 3. Absorbance profile (280 nm) of gel filtration extracts from control and 5.0 ppm Cd-exposed E. crassipes plants: (a) young and mature laminae; (b) young and mature protoplasts
W1-06
8
Fig. 4. RP-HPLC chromatographs of glutathione (GSH). (a) standard; (b) young control E. crassipes laminae; (c) young treated E. crassipes laminae; (d) mature control E. crassipes laminae; (e) mature treated E. crassipes laminae.
W1-06
9
Fig. 5. Circular dichroism (CD) spectra: (a) GSH standard; (b) BSA standard; (c) metallothionein 2 standard; (d) young control E. crassipes laminae; (e) young treated E. crassipes laminae; (f) mature control E. crassipes laminae; (g) mature treated E. crassipes laminae.
Fig. 6. Fourier Transformed–Infrared (FT-IR) spectra of laminae extracts from control and 5.0 ppm CdCl2 -exposed Eichhornia crassipes plants. Legend: GSH standard, dark green (highest curve within 3,500-3000 cm-1 wavenumbers); control young laminae extract, pink; Cd-exposed young laminae extract, red; control mature laminae extract, green; Cd-exposed mature laminae extract, blue.

W1-06
10
Acknowledgments We are grateful to the Natural Sciences Research Institute and the Institute of Biology for the research grant and support. References
[1] URL: An Overview of Minerals Potential and Opportunity in the Philippines. Mines and GeoSciences Bureau. Available at: http://clients.westminster-digital.co.uk/minesite/microsite/events/philippines/index.aspx.
[2] Brigden, K. and J. Cotter. 2006. Pollution from the Lafayette mine, Rapu Rapu (Philippines) during 30-day trial run, Greenpeace Research Laboratories Technical Note.
[3] Clemens, S. 2001. Molecular mechanisms of plant tolerance and homeostasis, Planta, 212: 475-486. [4] Drazkiewicz, M., A. Tukendorf, and T. Baszynskit. 2003. Age-dependent response of maize leaf segments to cadmium treatment:
effect on chlorophyll fluorescence and phytochelatin accumulation, J Plant Physiol, 160: 247-254. [5] Fujita, M. and T. Kawanishi. 1987. Cd-Binding Complexes from the Root Tissues of Various Higher Plants Cultivated in Cd2+-
Containing Medium, Plant and Cell Physiology, 28(2): 379-382. [6] Gong, J.M., D.A. Lee, and J. I. Schroader. 2003. Long-distance root-to-shoot transport of phytochelatins and cadmium in
Arabidopsis, Proc. Natl Acad. Sci. USA, 100(17): 10118-10123. [7] Grill, E., E. Winnacker, and M. Zenk. 1987. Phytochelatins, a class of heavy-metal-binding peptides from plants, are functionally
analogous to metallothioneins, Proc. Natl. Acad. Sci. USA, 84: 439-443. [8] Hernandez, L.E., E. Lozano-Rodriguez, A. Garate, and R. Carpena-Ruiz. 1998. Influence of cadmium on the uptake, tissue
accumulation and subcellular distribution of manganese in pea seedlings, Plant Science, 132: 139-151. [9] Hoagland, D. and D. I. Arnon. 1938. The water culture method for growing plants without soil, Calif. Agric. Exp. Sta. Circ, 347: 1-
39. [10] Jasco Corporation. 1995. CD/ORD Measurement Manual, Jasco Corp.,Tokyo, Japan, 1-69. [11] Josue, R.D.R., J.J.M. Puzon, J.A. Villanueva, and G.C. Rivero. 2006. Glutathione mediates Hg-binding and detoxification in
Ipomoea aquatica Forsk., Poster Presentation. 8th International Conference on Mercury as Global Pollutant. Madison, Wisconsin, U.S.A. 6-12 August 2006.
[12] Josue, R.D.R., J.A. Villanueva, and G.C. Rivero. 2004. Mercury uptake and phytochelatin production in Ipomoea aquatica Forsk, Philipp. Agr. Sci., 87(3): 312-321.
[13] Krupa, Z. and M. Moniak. 1998. The stage of leaf maturity implicates the response of the photosynthetic apparatus to cadmium toxicity, Plant Sci, 138: 149-156.
[14] Kubota, H., K. Sato, T. Yamada, and T. Maitani. 2000. Phytochelatin homologs induced in hairy roots of horseradish, Phytochem, 53: 239-245.
[15] Matoh, T., J. Watanabe, and E. Takahashi. 1987. Sodium, potassium, chloride, and betaine concentrations in isolated vacuoles from salt-grown Atriplex gmelini leaves, Plant Physiology, 84: 173-177.
[16] Nocito, F., L. Pirovano, M. Cocucci, and G. Sacchi. 2002. Cadmium-Induced Sulfate Uptake in Maize Roots, Plant Physiology, 129: 1-8.
[17] Peterson, P.J. and B.J. Alloway. 1979. Cadmium in soils and vegetation, In: Webb, M. (ed.) The chemistry, biochemistry and biology of cadmium. Elsevier: North Holland Biomedical Press, 93-132.
[18] Pietrini, F., M. Iannelli, S. Pasqualini, and A. Massacci. 2003. Interaction of Cadmium with Glutathione and Photosynthesis in Developing Leaves and Chloroplasts of Phragmites australis (Cav.) Trin. ex Steudel, Plant Physiology, 133: 829–83
[19] Puzon, J.M., G.C. Rivero, and M.E. Montaño. 2008. Vacuolar sequestration as a coping mechanism in cadmium-treated Eichhornia crassipes (Mart.) Solms, The Philippine Agricultural Scientist, 91: 123-133.
[20] Schaffer, A. 1991. Absorption, Circular Dichroism, and Magnetic Circular Dichroism Spectroscopy of Metallothionein, In: Methods in Enzymology, vol. 205, Part B: Metallothionein and Related Molecules. (J.F. Riordan and B.J. Vallee (eds.) Academic Press, Inc. U.S.A, 529-540.
[21] Smirnoff, N. 1995. Antioxidant systems and plant response to the environment, In: Environment and Plant Metabolism, N. Smirnoff (ed.). BIOS Scientific Publishers Limited. Oxford. U. K. pp. 222-224.
[22] Solomons, T.W.G. 1996. Spectroscopic methods of structure determination, In: Organic Chemistry. 6th ed. University of South Florida. John Wiley and Sons, Inc. Canada.
[23] Streitwieser, A. and C. Heathcock. 1985. Introduction to Organic Chemistry. 3rd ed. Macmillan Publishing Company. [24] Steffens, J.C. 1990. The heavy-metal-binding peptides of plants, Annu. Rev. Plant Physiol. Plant Mol. Biol., 41: 553-575 [25] Taiz, L. and E. Zeiger. 1998. Plant Physiology. 2nd ed. Sinauer Associates, Inc.,U.S.A. 792 pp. [26] Vogeli-Lange, R. and G. J. Wagner. 1990. Subcellular localization of cadmium and cadmium-binding peptides in tobacco leaves,
Plant Physiology, 92: 1086- 1093. [27] Wawrzynski, A., E. Kopera, A. Wawrzynski, J. Kanisha, W. Bal, and A. Sirko. 2006. Effects of simultaneous expression of
heterologous genes involved in PC biosynthesis on thiol content and Cd accumulation in tobacco plants, J. Exp. Bot., 57 (10): 2173-2182.
[28] URL: www.unep.org [29] Yamaguchi, Y., T. Nakamura, Y. Tatsumi, K. Nakamura, and H. Sano. 2006. Efficient assimilation of sulfide by transgenic rice
plants over-expressing a rice cysteine synthase, Plant Biotech, 23: 117-122.


















