
Thermal denaturation of recombinant human lysozyme from rice: effect of pH and comparison with human...
7
ORIGINAL PAPER Thermal denaturation of recombinant human lysozyme from rice: effect of pH and comparison with human milk lysozyme Eduardo Castillo • Indira Franco • Marı ´a D. Pe ´rez • Miguel Calvo • Lourdes Sa ´nchez Received: 28 June 2011 / Revised: 21 September 2011 / Accepted: 12 October 2011 / Published online: 29 October 2011 Ó Springer-Verlag 2011 Abstract Thermal denaturation of recombinant human lysozyme from rice has been studied by differential scan- ning calorimetry at acidic (4.2), neutral (7.2) and basic (9.2) pH levels at various heating rates, and it has been compared with thermal denaturation of human lysozyme isolated from milk at the same pH levels at a heating rate of 10 °C/min. Data obtained from heat-induced unfolding and subsequent refolding after heating indicate that thermal denaturation of both lysozymes undergoes a two-state process. The maximum temperature of the endothermic peaks and the enthalpy change of denaturation indicate that recombinant and milk lysozymes possess similar thermo- stability, which is higher at acidic than at neutral pH. On the other hand, both proteins are more thermolabile at basic pH. Lysozyme from human milk shows a higher tendency to aggregate than recombinant human lysozyme from rice during the thermal denaturation process. Keywords Human recombinant lysozyme Human milk lysozyme Differential scanning calorimetry Thermodynamical parameters Introduction Lysozymes are hydrolytic enzymes that cleave the b-(1,4) glycosidic bond between N-acetylmuramic acid and N-acetylglucosamine in peptidoglycan, the major bacterial cell wall polymer. Lysozymes are divided into six families according to genetical, immunological and structural sim- ilarities [1, 2]. Most lysozymes are implicated in defensive bactericidal systems in a wide range of taxonomically diverse organisms including, among others, plants, inver- tebrate and vertebrate animals [2]. One of these lysozyme families is the c-type group, to which hen egg-white lysozyme (HEWL) and human lysozyme (hLz) belong. HEWL makes up to 3.4% of the total egg-white protein content, whereas hLz is present in different human secre- tions, such as tears, saliva and milk. Particularly, human milk is rich in lysozyme, although this concentration has great individual and temporal variations. The concentration of lysozyme in colostrum is about 100 mg/L, then decreases to 20–40 mg/L and in the second half of the first year is increased, especially in the weaning stage, when it reaches concentrations of 800–1,000 mg/L [3]. This con- centration is about 3,000 times higher than that of cow’s milk, which ranges between 0.05 and 0.21 mg/mL [4, 5]. In addition, the enzymatic activity of hLz is 10 times greater than that of bovine lysozyme [6], and it has been reported that both proteins are antigenically different [7], which suggests the existence of structural differences between them. Lysozyme plays a role in the development of the gas- trointestinal microbiota in the newborn. It has been found that the predominant anaerobic flora in infants fed breast milk is composed of Bifidobacterium bifidum, contrary to that observed to those who are fed formula, in which the major microbiota is composed of coliform bacteria [8]. One of the reasons for which the intestinal flora of infants fed breast milk is rich in Bifidobacterium, bacteria essential for health and well-being of the child, may be due in part to the high content of hLz. Lysozyme plays a dual role: by hydrolyzing the peptidoglycan, it kills the harmful bacteria E. Castillo I. Franco M. D. Pe ´rez M. Calvo L. Sa ´nchez (&) Tecnologı ´a de los Alimentos, Facultad de Veterinaria, Universidad de Zaragoza, Miguel Servet 177, 50013 Zaragoza, Spain e-mail: [email protected] 123 Eur Food Res Technol (2011) 233:1067–1073 DOI 10.1007/s00217-011-1612-8
-
Upload
eduardo-castillo -
Category
Documents
-
view
212 -
download
0
Transcript of Thermal denaturation of recombinant human lysozyme from rice: effect of pH and comparison with human...

ORIGINAL PAPER
Thermal denaturation of recombinant human lysozymefrom rice: effect of pH and comparison with human milk lysozyme
Eduardo Castillo • Indira Franco • Marıa D. Perez •
Miguel Calvo • Lourdes Sanchez
Received: 28 June 2011 / Revised: 21 September 2011 / Accepted: 12 October 2011 / Published online: 29 October 2011
� Springer-Verlag 2011
Abstract Thermal denaturation of recombinant human
lysozyme from rice has been studied by differential scan-
ning calorimetry at acidic (4.2), neutral (7.2) and basic
(9.2) pH levels at various heating rates, and it has been
compared with thermal denaturation of human lysozyme
isolated from milk at the same pH levels at a heating rate of
10 �C/min. Data obtained from heat-induced unfolding and
subsequent refolding after heating indicate that thermal
denaturation of both lysozymes undergoes a two-state
process. The maximum temperature of the endothermic
peaks and the enthalpy change of denaturation indicate that
recombinant and milk lysozymes possess similar thermo-
stability, which is higher at acidic than at neutral pH. On
the other hand, both proteins are more thermolabile at basic
pH. Lysozyme from human milk shows a higher tendency
to aggregate than recombinant human lysozyme from rice
during the thermal denaturation process.
Keywords Human recombinant lysozyme � Human milk
lysozyme � Differential scanning calorimetry �Thermodynamical parameters
Introduction
Lysozymes are hydrolytic enzymes that cleave the b-(1,4)
glycosidic bond between N-acetylmuramic acid and
N-acetylglucosamine in peptidoglycan, the major bacterial
cell wall polymer. Lysozymes are divided into six families
according to genetical, immunological and structural sim-
ilarities [1, 2]. Most lysozymes are implicated in defensive
bactericidal systems in a wide range of taxonomically
diverse organisms including, among others, plants, inver-
tebrate and vertebrate animals [2]. One of these lysozyme
families is the c-type group, to which hen egg-white
lysozyme (HEWL) and human lysozyme (hLz) belong.
HEWL makes up to 3.4% of the total egg-white protein
content, whereas hLz is present in different human secre-
tions, such as tears, saliva and milk. Particularly, human
milk is rich in lysozyme, although this concentration has
great individual and temporal variations. The concentration
of lysozyme in colostrum is about 100 mg/L, then
decreases to 20–40 mg/L and in the second half of the first
year is increased, especially in the weaning stage, when it
reaches concentrations of 800–1,000 mg/L [3]. This con-
centration is about 3,000 times higher than that of cow’s
milk, which ranges between 0.05 and 0.21 mg/mL [4, 5]. In
addition, the enzymatic activity of hLz is 10 times greater
than that of bovine lysozyme [6], and it has been reported
that both proteins are antigenically different [7], which
suggests the existence of structural differences between
them.
Lysozyme plays a role in the development of the gas-
trointestinal microbiota in the newborn. It has been found
that the predominant anaerobic flora in infants fed breast
milk is composed of Bifidobacterium bifidum, contrary to
that observed to those who are fed formula, in which the
major microbiota is composed of coliform bacteria [8]. One
of the reasons for which the intestinal flora of infants fed
breast milk is rich in Bifidobacterium, bacteria essential for
health and well-being of the child, may be due in part to the
high content of hLz. Lysozyme plays a dual role: by
hydrolyzing the peptidoglycan, it kills the harmful bacteria
E. Castillo � I. Franco � M. D. Perez � M. Calvo �L. Sanchez (&)
Tecnologıa de los Alimentos, Facultad de Veterinaria,
Universidad de Zaragoza, Miguel Servet 177,
50013 Zaragoza, Spain
e-mail: [email protected]
123
Eur Food Res Technol (2011) 233:1067–1073
DOI 10.1007/s00217-011-1612-8

and, in addition, it induces the liberation of N-acetylglu-
cosamine, which is an important bifidogenic factor. Thus,
despite the high content of lysozyme in breast milk, the
number of bifidobacteria is increased in the intestinal tract,
suggesting that these bacteria are resistant to the antibac-
terial activity of this enzyme [9]. It has also been docu-
mented that breastfed babies suffer fewer infections, and
these are shorter than those of infants fed with infant for-
mula [10]. In Japan, studies were conducted supplementing
infant formula with HEWL, and it was found that the
intestinal microbiota of infants fed with that milk was
richer in bifidobacteria than that of those fed with com-
mercially available infant formulae [11]. These results
suggest that supplementation of infant formula with lyso-
zyme would be very convenient. However, the addition of
egg protein that is the main source of lysozyme for com-
mercial use in infant food may cause allergic reactions, so
it would be desirable to supplement infant formula with
hLz [12]. Furthermore, the structure of hen egg lysozyme,
as well as its enzymatic and antibacterial activities, is
different to those of human milk.
Human lysozyme has been isolated from milk, pancreatic
juice [13], neutrophils [14] and urine of hemodialysis
patients [15]. Since human milk is the main source of this
protein, it is obviously impossible to obtain enough hLz to be
added to commercial products. However, hLz has been
expressed successfully in transgenic cattle [16] and in Pichia
pastoris [17]. The expression of recombinant proteins in rice
is an alternative to produce large quantities of proteins with
low cost. The use of plants for large-scale expression of
human proteins offers some advantages over microbial or
mammalian cell culture systems, because transgenic plants
do not carry or propagate human diseases or mammalian
viral vectors [18]. In addition, the expression of recombinant
proteins in edible plant tissues could reduce protein purifi-
cation requirements for some applications [19]. Rice is a
particular well-suited crop for this purpose, since their seeds
allow for long-term storage and hold GRAS status. There-
fore, rice-based foods are regarded as hypoallergenic, and
rhLz obtained from transgenic rice could be used in a par-
tially purified form as a baby formula ingredient [20].
The use of rhLz as a food additive or ingredient requires
the knowledge of its thermal stability; therefore, the pur-
pose of this study was the determination of the thermal
stability of rhLz by differential scanning calorimetry
(DSC), a technique that provides useful information con-
cerning factors that affect protein stability [21]. The ther-
moresistance of rhLz was determined at three different pH
levels: acidic, neutral and basic, since no thermal stability
studies of rhLz has been carried out at pH above the acidic
range. In addition, the thermoresistance of rhLz has been
compared with that of lysozyme from human milk by the
same technique.
Materials and methods
Proteins and human milk
Recombinant hLz isolated from rice was kindly provided
by Ventria Bioscience (Sacramento, CA) as a white powder
of a high purity above 98% as determined by native PAGE.
Human milk samples were generously donated by
healthy mothers by mediation of Dr. Luis Ros from the
Hospital Miguel Servet of Zaragoza (Spain).
Isolation of lysozyme from human milk
Milk was skimmed by centrifugation at 2,5009g at 4 �C for
30 min. Caseins were separated by acid precipitation adding
0.1 M HCl to reach a pH of 4.5 and subsequent centrifuga-
tion at 2,5009g at room temperature for 30 min. The clear
whey obtained was dialyzed against 50 mM sodium acetate
buffer, pH 4.5 overnight. Afterward, to avoid undesired
interactions between hLz and other whey proteins, 100 lL
of Triton X-100 was added to 250 mL of the dialyzed whey,
and then, it was applied on a SP-Sepharose column
(/1.5 cm 9 8 cm) previously equilibrated with 50 mM
sodium acetate buffer, pH 4.5 containing 50 mM NaCl. The
column was washed with the same buffer, and the bound
proteins were sequentially eluted with 50 mM sodium ace-
tate buffer, pH 4.5 and a NaCl stepwise gradient (0.2, 0.5 and
1 M). Fractions were collected and analyzed by SDS–
PAGE, and those with the highest content of hLz were
concentrated by ultrafiltration and applied to a Sephadex
G-50 column (/1 cm 9 80 cm), which was eluted using
1 M NaCl and 10 mM potassium phosphate buffer, pH 7.4.
The fractions collected were analyzed by SDS–PAGE, and
those containing pure hLz were dialyzed against distilled
water and lyophilized.
Enzymatic activity assay
Lysozyme activity was determined by a turbidimetric
technique, measuring the decrease in absorbance at 450 nm
versus time of a Micrococcus lysodeikticus suspension
(Sigma Aldrich, Steinheim, Germany). A fresh suspension
of M. lysodeikticus (0.15 mg/mL) in 100 mM phosphate
buffer, pH 6.2, was used as substrate. For each sample,
950 lL of substrate was placed in a cuvette to which 50 lL
of sample was added. Absorbance decrease was recorded
versus time and the activity of each sample calculated from
the slope of the curve.
Differential scanning calorimetry (DSC)
Solutions of rhLz and of hLz at a concentration of 100 mg/
mL were prepared in three different buffers: 100 mM
1068 Eur Food Res Technol (2011) 233:1067–1073
123

sodium acetate buffer, pH 4.2; 15 mM potassium phosphate,
pH 7.4; and 100 mM sodium carbonate buffer, pH 9.2. For
the DSC study of hLz, 100 mM sodium carbonate buffer, pH
8.8, and 100 mM sodium phosphate buffer, pH 8.0, were also
employed. Samples and references (10 lL) were introduced
into aluminum pans (TA Instruments, New Castle, USA) and
sealed for analysis. The references consisted of pans con-
taining the corresponding buffer. DSC analysis of both
lysozymes was performed in a DuPont Thermal Analyser
(model DSC 10, Nemours, Germany), using a Thermal
Analyst 2000 System. DSC scans were programmed in the
temperature range of 35–110 �C at different heating rates: 2,
3, 4, 5, 7, 10 and 20 �C/min for rhLz and at 10 �C/min for
hLz. Lysozymes were analyzed using 3–5 replicates.
Pans containing samples of rhLz from rice treated at pH
7.2 and at a heating rate of 10 �C/min were left to cool at
4 �C during 24 h and then rescanned in the same condi-
tions of the first measurement in order to evaluate a pos-
sible renaturation of the protein after heating.
From the transition peaks obtained by DSC, several
thermodynamic parameters were obtained, such as the
temperature of maximum heat absorption (Tp), the onset
temperature (Ti) and the enthalpy change (DHap) for the
denaturation process of proteins. The last parameter (DHap)
was calculated by integrating the peak area using a straight
baseline drawn from the onset to the end of thermal tran-
sition. The value of van’t Hoff enthalpy was calculated
according to the following equation:
DHVH ¼4RT2
p
DT1=2
where Tp is the temperature of maximum heat absorption,
DT1/2 is the endothermic peak width at its half-height and R
is the universal gas constant.The activation energy (Ea) for
the denaturation process of rhLz from rice was calculated
according to Kissinger’s method [22]. Thus, the activation
energy of a denaturation process can be calculated from the
heating rate (b) and the temperature of maximum heat
absorption (Tp) using the Arrhenius equation:
LnbT2
p
!¼ �Ea
R� 1
T
where R is the universal gas constant and the temperatures
are given in Kelvin degrees.
Results
Isolation of lysozyme from human milk
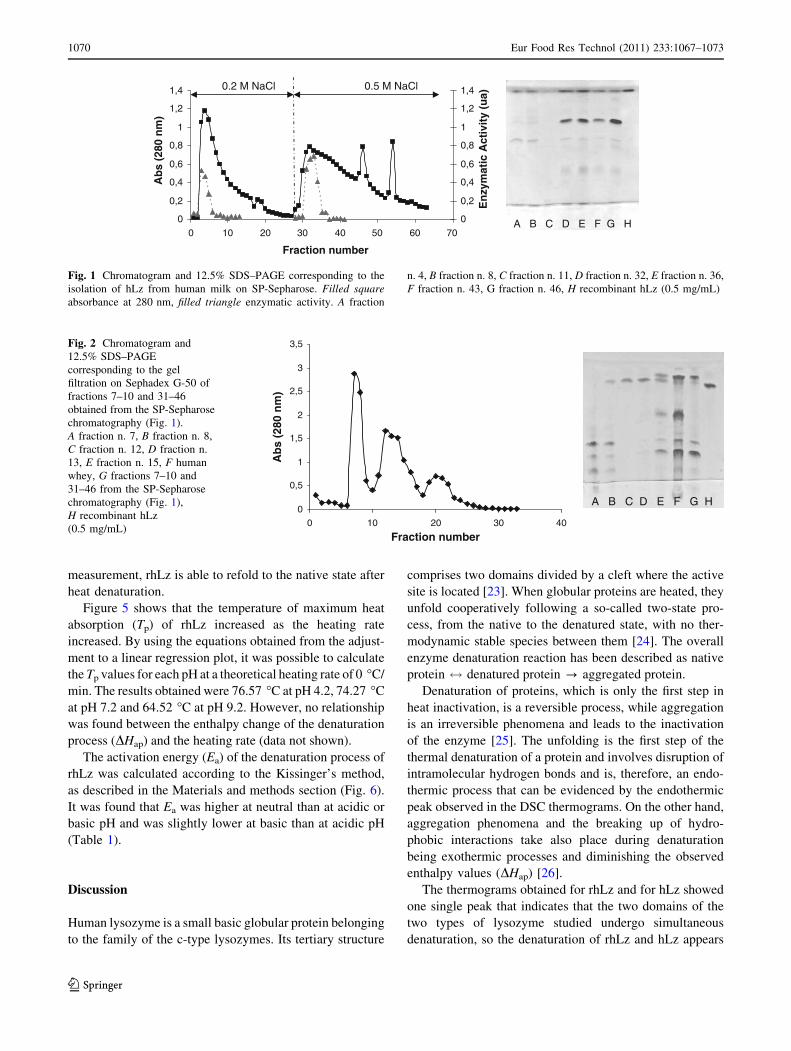
Figure 1 shows the chromatogram and the enzymatic
activity as well as the corresponding SDS–PAGE of frac-
tions eluted with 50 mM sodium acetate buffer, pH 4.5
containing 0.2 and 0.5 M NaCl. The elution with 1 M NaCl
did not result in any measurable peak. As it is shown by
SDS–PAGE, besides hLz other whey proteins were present
in the two peaks obtained. The fractions with the highest
hLz content and enzymatic activity (fractions 7–10 and
31–43) were concentrated and subjected to gel filtration.
After the analysis of fractions by SDS–PAGE, it was found
that only fraction C contained pure lysozyme (Fig. 2),
because this protein interacts with other whey proteins and
cannot be easily separated from them. This fraction was
concentrated by ultrafiltration, dialyzed against distilled
water and lyophilized.
DSC of rhLz from rice to hLz from milk
Thermal stability of rhLz was studied at three different pH
levels, 4.2, 7.2 and 9.2, and at different heating rates by
means of DSC. Thermal stability of hLz was also studied at
the same pH levels as rhLz at a heating rate of 10 �C/min.
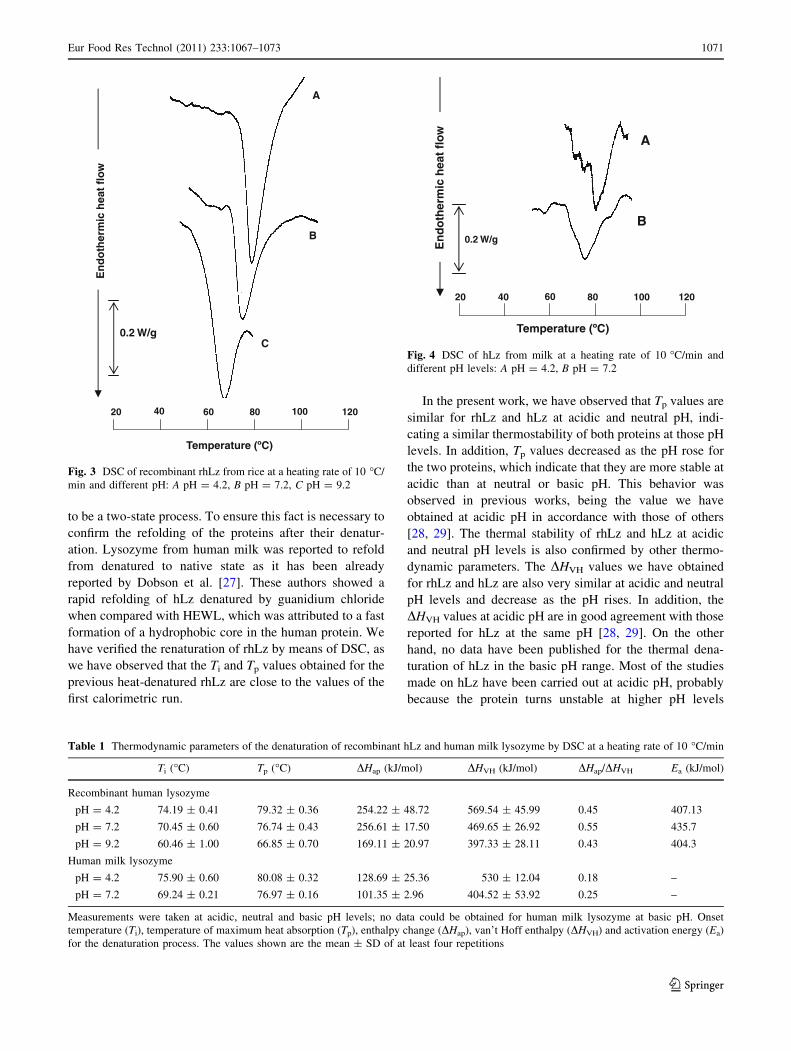
Figure 3 shows the thermograms obtained for rhLz
treated at the three different pH levels at a heating rate of
10 �C/min. All measurements resulted in thermograms
showing a major endothermic peak at all pH levels and
heating rates studied, with the exception of samples treated
at pH 9.2 at heating rates of 2 and 3 �C/min, which did not
give any endothermic peak. This fact indicates that rhLz is
less stable at basic than at acidic or neutral pH.
The thermograms obtained for hLz are shown in Fig. 4.
They present also one single denaturation peak at acidic and
neutral pH levels, but it was not possible to obtain any
thermogram at pH 9.2. In addition, the study was also carried
out at pH values of 8.8 and 8.0, but in any case, a thermogram
could be obtained at a heating rate of 10 �C/min.
The thermodynamic parameters obtained at a heating
rate of 10 �C/min for the denaturation of rhLz and hLz at
the three pH levels are shown in Table 1. For both lyso-
zymes, the temperature of maximum heat absorption (Tp)
decreases as the pH rises, which points out to a greater
stability of both proteins at acidic pH. In addition, Tp
values at acidic and neutral pH levels are the same for both
lysozymes, which indicate that the thermal stability of rhLz
is comparable with that of hLz at those pH levels.
The thermogram obtained in the renaturation study of
rhLz carried out at pH 7.2 and at a heating rate of 10 �C/
min also showed one single peak, with an onset tempera-
ture (Ti) of 69.95 �C and a temperature of maximum heat
absorption of 76.63 �C, both values close to that of the first
measurement, 70.45 and 76.74 �C, respectively. However,
the enthalpy change of the second measurement was of
75.79 kJ/mL, about 70% lower than that of the value
obtained in the first run. All these data indicate that despite
aggregation that is evidenced by the DHap/DHVH ratio and
by the drop in the enthalpy change in the second
Eur Food Res Technol (2011) 233:1067–1073 1069
123
measurement, rhLz is able to refold to the native state after
heat denaturation.
Figure 5 shows that the temperature of maximum heat
absorption (Tp) of rhLz increased as the heating rate
increased. By using the equations obtained from the adjust-
ment to a linear regression plot, it was possible to calculate
the Tp values for each pH at a theoretical heating rate of 0 �C/
min. The results obtained were 76.57 �C at pH 4.2, 74.27 �C
at pH 7.2 and 64.52 �C at pH 9.2. However, no relationship
was found between the enthalpy change of the denaturation
process (DHap) and the heating rate (data not shown).
The activation energy (Ea) of the denaturation process of
rhLz was calculated according to the Kissinger’s method,
as described in the Materials and methods section (Fig. 6).
It was found that Ea was higher at neutral than at acidic or
basic pH and was slightly lower at basic than at acidic pH
(Table 1).
Discussion
Human lysozyme is a small basic globular protein belonging
to the family of the c-type lysozymes. Its tertiary structure
comprises two domains divided by a cleft where the active
site is located [23]. When globular proteins are heated, they
unfold cooperatively following a so-called two-state pro-
cess, from the native to the denatured state, with no ther-
modynamic stable species between them [24]. The overall
enzyme denaturation reaction has been described as native
protein $ denatured protein ? aggregated protein.
Denaturation of proteins, which is only the first step in
heat inactivation, is a reversible process, while aggregation
is an irreversible phenomena and leads to the inactivation
of the enzyme [25]. The unfolding is the first step of the
thermal denaturation of a protein and involves disruption of
intramolecular hydrogen bonds and is, therefore, an endo-
thermic process that can be evidenced by the endothermic
peak observed in the DSC thermograms. On the other hand,
aggregation phenomena and the breaking up of hydro-
phobic interactions take also place during denaturation
being exothermic processes and diminishing the observed
enthalpy values (DHap) [26].
The thermograms obtained for rhLz and for hLz showed
one single peak that indicates that the two domains of the
two types of lysozyme studied undergo simultaneous
denaturation, so the denaturation of rhLz and hLz appears
0 10 20 30 40 50 60 70A B C D E F G H 0
0,2
0,4
0,6
0,8
1
1,2
1,4
Fraction number
Ab
s (2
80 n
m)
0
0,2
0,4
0,6
0,8
1
1,2
1,4
En
zym
atic
Act
ivit
y (u
a)
0.2 M NaCl 0.5 M NaCl
Fig. 1 Chromatogram and 12.5% SDS–PAGE corresponding to the
isolation of hLz from human milk on SP-Sepharose. Filled squareabsorbance at 280 nm, filled triangle enzymatic activity. A fraction
n. 4, B fraction n. 8, C fraction n. 11, D fraction n. 32, E fraction n. 36,
F fraction n. 43, G fraction n. 46, H recombinant hLz (0.5 mg/mL)
A B C D E F G H 0
0,5
1
1,5
2
2,5
3
3,5
0 10 20 30 40
Fraction number
Ab
s (2
80 n
m)
Fig. 2 Chromatogram and
12.5% SDS–PAGE
corresponding to the gel
filtration on Sephadex G-50 of
fractions 7–10 and 31–46
obtained from the SP-Sepharose
chromatography (Fig. 1).
A fraction n. 7, B fraction n. 8,
C fraction n. 12, D fraction n.
13, E fraction n. 15, F human
whey, G fractions 7–10 and
31–46 from the SP-Sepharose
chromatography (Fig. 1),
H recombinant hLz
(0.5 mg/mL)
1070 Eur Food Res Technol (2011) 233:1067–1073
123
to be a two-state process. To ensure this fact is necessary to
confirm the refolding of the proteins after their denatur-
ation. Lysozyme from human milk was reported to refold
from denatured to native state as it has been already
reported by Dobson et al. [27]. These authors showed a
rapid refolding of hLz denatured by guanidium chloride
when compared with HEWL, which was attributed to a fast
formation of a hydrophobic core in the human protein. We
have verified the renaturation of rhLz by means of DSC, as
we have observed that the Ti and Tp values obtained for the
previous heat-denatured rhLz are close to the values of the
first calorimetric run.
In the present work, we have observed that Tp values are
similar for rhLz and hLz at acidic and neutral pH, indi-
cating a similar thermostability of both proteins at those pH
levels. In addition, Tp values decreased as the pH rose for
the two proteins, which indicate that they are more stable at
acidic than at neutral or basic pH. This behavior was
observed in previous works, being the value we have
obtained at acidic pH in accordance with those of others
[28, 29]. The thermal stability of rhLz and hLz at acidic
and neutral pH levels is also confirmed by other thermo-
dynamic parameters. The DHVH values we have obtained
for rhLz and hLz are also very similar at acidic and neutral
pH levels and decrease as the pH rises. In addition, the
DHVH values at acidic pH are in good agreement with those
reported for hLz at the same pH [28, 29]. On the other
hand, no data have been published for the thermal dena-
turation of hLz in the basic pH range. Most of the studies
made on hLz have been carried out at acidic pH, probably
because the protein turns unstable at higher pH levels
40 60 100
Temperature (ºC)
En
do
ther
mic
hea
t fl
ow
0.2 W/g
A
B
C
20 80 120
Fig. 3 DSC of recombinant rhLz from rice at a heating rate of 10 �C/
min and different pH: A pH = 4.2, B pH = 7.2, C pH = 9.2
60 100 120
Temperature (ºC)
En
do
ther
mic
hea
t fl
ow
0.2 W/g
A
B
20 40 80
Fig. 4 DSC of hLz from milk at a heating rate of 10 �C/min and
different pH levels: A pH = 4.2, B pH = 7.2
Table 1 Thermodynamic parameters of the denaturation of recombinant hLz and human milk lysozyme by DSC at a heating rate of 10 �C/min
Ti (�C) Tp (�C) DHap (kJ/mol) DHVH (kJ/mol) DHap/DHVH Ea (kJ/mol)
Recombinant human lysozyme
pH = 4.2 74.19 ± 0.41 79.32 ± 0.36 254.22 ± 48.72 569.54 ± 45.99 0.45 407.13
pH = 7.2 70.45 ± 0.60 76.74 ± 0.43 256.61 ± 17.50 469.65 ± 26.92 0.55 435.7
pH = 9.2 60.46 ± 1.00 66.85 ± 0.70 169.11 ± 20.97 397.33 ± 28.11 0.43 404.3
Human milk lysozyme
pH = 4.2 75.90 ± 0.60 80.08 ± 0.32 128.69 ± 25.36 530 ± 12.04 0.18 –
pH = 7.2 69.24 ± 0.21 76.97 ± 0.16 101.35 ± 2.96 404.52 ± 53.92 0.25 –
Measurements were taken at acidic, neutral and basic pH levels; no data could be obtained for human milk lysozyme at basic pH. Onset
temperature (Ti), temperature of maximum heat absorption (Tp), enthalpy change (DHap), van’t Hoff enthalpy (DHVH) and activation energy (Ea)
for the denaturation process. The values shown are the mean ± SD of at least four repetitions
Eur Food Res Technol (2011) 233:1067–1073 1071
123

[30–32]. In fact, in our study, it was not possible to obtain
any thermogram for hLz at the three basic pH levels
studied: 9.2, 9.0 and 8.8. In addition, it was found that Tp
and DHap values were lower at basic pH for rhLz, which
confirms the thermal sensitivity of lysozyme at basic pH.
The lower DHap values obtained for hLz may be due to a
higher tendency of hLz to aggregate, since the DHVH val-
ues are very similar. These slight differences observed in
the thermodynamic values obtained for denaturation of
rhLz compared to those of hLz at basic pH may be due to
differences in the folding patterns or mechanisms at that
pH, since the behavior and the values obtained at acidic
and neutral pH levels are very similar for both proteins.
The production of recombinant proteins in bioreactors is
a rapid and often inexpensive process; however, this
technique implies a high rate of misfolded or aggregated
pH 4.2
0 5 10 15 20 2576
78
80
82
84
Heating rate (ºC/min)
Tp
(ºC
)
pH 7.2
0 5 10 15 20 2574
76
78
80
Heating rate (ºC/min)
Tp
(ºC
)
pH 9.2
0 5 10 15 20 2564
66
68
70
Heating rate (ºC/min)
Tp
(ºC
)
Fig. 5 Effect of the heating rate
on the temperature of maximum
heat absorption (Tp) for
recombinant rhLz from rice
pH 4.2
2.81 2.82 2.83 2.84 2.85 2.86-12
-11
-10
-9
-8
1/Tp (x 103)(K-1)
Ln
(β
/Tp
2 )
pH 7.2
2.83 2.84 2.85 2.86 2.87 2.88 2.89-12
-11
-10
-9
-8
1/Tp (x 103)(K-1)
Ln
(β
/Tp
2 )
pH 9.2
2.92 2.93 2.94 2.95 2.96-11
-10
-9
-8
1/Tp (x 103)(K-1)
Ln
(β
/Tp
2 )
Fig. 6 Kissinger plot for heat
denaturation of rhLz from rice.
Tp is the temperature of
maximum heat absorption, and
b is the heating rate
1072 Eur Food Res Technol (2011) 233:1067–1073
123

proteins [33]. Therefore, the expression of proteins whose
tridimensional structure is indispensable for its biological
functions, as is the case of human lysozyme, should be
conducted by other approaches. Plants are suitable for this
purpose, since their endoplasmic reticulum enables proper
folding of proteins [34]. The thermodynamic parameters
we have obtained for the thermal denaturation of rhLz from
rice evidence that the thermal stability of this protein is
similar to that of hLz from milk at both acidic and neutral
pH levels, which also points out to a correspondence in
their folding structure. On the other hand, both proteins are
more thermolabile at basic pH, though rhLz is more labile.
This finding could indicate a slight difference in the tridi-
mensional structure of the recombinant protein compared
to that of human milk that makes rhLz being less stable
upon heating at basic pH. This fact was not previously
described, since most published studies have been carried
out at acidic pH.
Acknowledgments This research has been carried out supported by
the European Social Fund and by a research fellow grant from the
Ministerio de Educacion y Ciencia of Spanish Government.
References
1. Olsen OM, Nilsen IW, Sletten K, Myrnes B (2003) Comp Bio-
chem Phys B 136:107–115
2. Ibrahim HR, Higashiguchi S, Juneja LR, Kim M, Yamamoto T
(1996) J Agric Food Chem 44:1416–1423
3. Braun OH, Sandkuhler H (1985) J Pediatr Gastr Nutr 4:583–586
4. Parry RM, Chandan RC, Shahani KM (1968) J Dairy Sci
51:606–607
5. Piccini R, Binda E, Belotti M, Casirani G, Zecconi A (2005) Vet
Res 36:747–757
6. Li N, Yu Z, Meng Q, Yu H, Fan B, Yu S, Fei J, Wang L, Dai Y
(2006) J Dairy Sci 89:2911–2918
7. Eitenmiller RR, Friend BA, Shahani KM, Ball EM (1974) J Food
Sci 39:930–933
8. Hanson LA, Winberg J (1972) Arch Dis Child 47:845–848
9. Modler HW (1994) Int Dairy J 4:383–407
10. Dewey KG, Heinig MJ, Nommsen-Rivers LA (1995) J Pediatr
126:696–702
11. Proctor VA, Cunningham FE (1988) Crit Rev Food Sci
26:359–395
12. Carlsson B, Cruz JR, Garcıa B, Hanson LA, Urrutia JJ (1979)
Immune factors in human milk. In: Visser HKA (ed) Nutrition
and metabolismus of the fetus and infants. The Hague, The
Netherlands, pp 263–271
13. Wang CS, Kloer HU (1984) Anal Biochem 139:224–227
14. Lollike K, Kjeldsen L, Sengeløv H, Borregaard N (1995)
Leukemia 9:206–209
15. Takai I, Oda O, Shinzato T, Ohbayashi K, Yamanaka N, Maeda
K (1996) J Chromatogr B Biomed Appl 685:21–25
16. Yang B, Wang J, Tang B, Liu Y, Guo C, Yang P, Yu T, Li R,
Zhao J, Zhang L, Dai Y, Li N (2011) PLoS One 16:e17593
17. Xiong R, Chen J (2008) Biotechnol Appl Biochem 51:129–134
18. Goldstein DA, Thomas JA (2004) Q J Med 97:705–716
19. Twyman RM, Stoger E, Schillberg S, Christou P, Fischer R
(2003) Trends Biotechnol 21:570–578
20. Gastanaduy A, Cordano A, Graham GG (1990) J Pediatr Gas-
troenterol Nutr 11:240–246
21. Llabres M, Cueto M, Dorta MJ, Munguıa O (2003) Int J Pharm
252:159–166
22. Kissinger HE (1957) Anal Chem 29:1702–1706
23. Jolles P, Jolles J (1984) Mol Cell Biochem 63:165–189
24. Privalov PL, Griko YV, Freire E, Privalov G, van Dael H (1995) J
Mol Biol 252:447–459
25. Makki F, Durance TD (1996) Food Res Int 29:635–645
26. Harwalkar VR, Ma CY, Maurice TJ (1990) Instrumentation and
techniques of thermal analysis in food research. In: Harwalkar
VR, Ma CY (eds) Thermal analysis of foods. Elsevier, Essex,
pp 1–15
27. Dobson CM, Hooke SD, Radford SE (1994) Biochemistry
33:5867–5876
28. Koshiba T, Tsumoto K, Masaki K, Kawano K, Nitta K, Kumagai
I (1998) Protein Eng 11:683–690
29. Kuroki R, Kawakita S, Nakamura H, Yutani K (1992) Proc Natl
Acad Sci USA 89:6803–6807
30. Imoto T, Hashimoto Y, Munemura O, Masumoto K, Ueda T
(2001) Biol Pharm Bull 24:1102–1107
31. Yutani K, Takano K, Ogasahara K, Kaneda H, Yagamata Y, Fujii
S, Kanaya E, Kikuchi M, Oobatake M (1995) J Mol Biol
254:62–76
32. Yutani K, Takano K, Tsuchimori K, Yagamata Y (1999) Eur J
Biochem 266:675–682
33. Clark EDB (1998) Curr Opin Biotechnol 9:157–163
34. Boothe J, Nykiforuk C, Shen Y, Zaplachinski S, Szarka S,
Kuhlman P, Murray E, Morck D, Moloney MM (2010) Plant
Biotechnol J 8:588–606
Eur Food Res Technol (2011) 233:1067–1073 1073
123


















