
Therapeutic potential of amniotic fluid-derived cells for treating the injured nervous system
49
Therapeutic potential of amniotic fluid-derived cells for treating the injured nervous system Kerry Rennie 1 , Julie Haukenfrers 1 , Maria Ribecco-Lutkiewicz 1 , Dao Ly 1 , Anna Jezierski 1,3 Brandon Smith 1 , Bogdan Zurakowski 1 , Marzia Martina 2,3 , Andrée Gruslin 3,4 , Mahmud Bani-Yaghoub 1,3 1 Neurogenesis and Brain Repair, National Research Council Canada, Bldg. M-54, Ottawa, ON, K1A 0R6, Canada 2 Synaptic Therapies and Devices, National Research Council Canada, Bldg. M-54, Ottawa, ON, K1A 0R6, Canada 3 Department of Cellular and Molecular Medicine, Faculty of Medicine, University of Ottawa, Ottawa, Canada 4 Department of Obstetrics and Gynecology, Faculty of Medicine, University of Ottawa, Ottawa, Canada Corresponding Author: Dr. Mahmud Bani Department of Translational Bioscience, National Research Council Canada, Bldg. M-54, Ottawa, ON, K1A 0R6, Canada Email: [email protected]) Phone: (613) 993-5723 Keywords: amniotic fluid, brain injury, cell-based therapy, gap junctions, micro-RNAs, stem cells Page 1 of 41 Page 1 of 49 Biochem. Cell Biol. Downloaded from www.nrcresearchpress.com by UNIVERSITY OF MICHIGAN on 06/13/13 For personal use only. This Just-IN manuscript is the accepted manuscript prior to copy editing and page composition. It may differ from the final official version of record.
Transcript of Therapeutic potential of amniotic fluid-derived cells for treating the injured nervous system

Therapeutic potential of amniotic fluid-derived cells for treating the injured nervous
system
Kerry Rennie1, Julie Haukenfrers
1, Maria Ribecco-Lutkiewicz
1, Dao Ly
1, Anna Jezierski
1,3
Brandon Smith1, Bogdan Zurakowski
1, Marzia Martina
2,3, Andrée Gruslin
3,4, Mahmud
Bani-Yaghoub1,3
1Neurogenesis and Brain Repair, National Research Council Canada, Bldg. M-54, Ottawa,
ON, K1A 0R6, Canada
2Synaptic Therapies and Devices, National Research Council Canada, Bldg. M-54, Ottawa,
ON, K1A 0R6, Canada
3Department of Cellular and Molecular Medicine, Faculty of Medicine, University of
Ottawa, Ottawa, Canada
4Department of Obstetrics and Gynecology, Faculty of Medicine, University of Ottawa,
Ottawa, Canada
Corresponding Author:
Dr. Mahmud Bani
Department of Translational Bioscience,
National Research Council Canada,
Bldg. M-54, Ottawa, ON, K1A 0R6,
Canada
Email: [email protected])
Phone: (613) 993-5723
Keywords: amniotic fluid, brain injury, cell-based therapy, gap junctions, micro-RNAs,
stem cells
Page 1 of 41Page 1 of 49B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIC
HIG
AN
on
06/1
3/13
For
pers
onal
use
onl
y. T
his
Just
-IN
man
uscr
ipt i
s th
e ac
cept
ed m
anus
crip
t pri
or to
cop
y ed
iting
and
pag
e co
mpo
sitio
n. I
t may
dif
fer
from
the
fina
l off
icia
l ver
sion
of
reco
rd.

Abstract
There is a need for improved therapy for acquired brain injury, which has proven resistant
to treatment by numerous drugs in clinical trials, and continues to represent one of the
leading causes of disability worldwide. Research into cell-based therapies for the treatment
of brain injury is growing rapidly, but the ideal cell source has yet to be determined.
Subpopulations of cells found in amniotic fluid, which is readily obtained during routine
amniocentesis, can be easily expanded in culture, have multipotent differentiation capacity,
are non-tumourigenic, and avoid the ethical complications associated with embryonic stem
cells, making them a promising cell source for therapeutic purposes. Beneficial effects of
amniotic fluid cell transplantation have been reported in various models of nervous system
injury. However, evidence that amniotic fluid cells can differentiate into mature, functional
neurons in vivo and incorporate into the existing circuitry to replace lost or damaged
neurons is lacking. The mechanisms by which amniotic fluid cells improve outcomes after
experimental nervous system injury remain unclear. However, studies reporting the
expression and release of neurotrophic, angiogenic and immunomodulatory factors by
amniotic fluid cells suggest they may provide neuroprotection and/or stimulate endogenous
repair and remodelling processes in the injured nervous system. In this paper, we address
recent research related to the neuronal differentiation of amniotic fluid-derived cells, the
therapeutic efficacy of these cells in animal models of nervous system injury, and the
possible mechanisms mediating the positive outcomes achieved by amniotic fluid cell
transplantation.
Page 2 of 41Page 2 of 49B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIC
HIG
AN
on
06/1
3/13
For
pers
onal
use
onl
y. T
his
Just
-IN
man
uscr
ipt i
s th
e ac
cept
ed m
anus
crip
t pri
or to
cop
y ed
iting
and
pag
e co
mpo
sitio
n. I
t may
dif
fer
from
the
fina
l off
icia
l ver
sion
of
reco
rd.

1. Introduction
Acquired brain injury, most commonly resulting from traumatic brain injury (TBI)
or stroke, is one of the leading causes of death and disability worldwide. Although the brain
is capable of a limited degree of endogenous repair and functional recovery, acquired brain
injury commonly leads to neurological deficits. Globally, approximately one-quarter to
one-half of brain injury patients exhibit substantial disability six months after moderate to
severe injury, and many are disabled even after mild brain injury (De Silva et al., 2009).
Unfortunately, the deficits are long-lasting for many brain injury patients. For instance, it is
estimated that 4% of Canadians are currently living with the long term cognitive, emotional
and/or sensory/motor sequelae of an acquired brain injury (The Ontario Brain Injury
Association 2011).
Despite decades of research and development leading to the testing of a multitude of
potentially neuroprotective compounds, translation to the clinic has been difficult. Although
neuroprotectants have been approved for clinical use in some countries (e.g. Japan), in
many countries there remains no approved clinically effective treatment for preventing
neuronal loss or repairing the brain after injury, beyond general strategies for promoting the
return of blood supply to the damaged area and reducing brain pressure in the immediate
post-injury phase (Auriel and Bornstein 2010; Gross et al. 2010; Labiche and Grotta 2004).
As a result, there is growing interest in novel therapeutic approaches, such as cell-based
therapy, to repair tissue damage and restore neurological function after acquired brain
injury (Bjorklund and Lindvall 2000; Schouten et al. 2004). In this paper, we will focus on
the growing body of research related to the potential use of human amniotic fluid (AF)-
derived cells as a therapy for nervous system injury.
Page 3 of 41Page 3 of 49B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIC
HIG
AN
on
06/1
3/13
For
pers
onal
use
onl
y. T
his
Just
-IN
man
uscr
ipt i
s th
e ac
cept
ed m
anus
crip
t pri
or to
cop
y ed
iting
and
pag
e co
mpo
sitio
n. I
t may
dif
fer
from
the
fina
l off
icia
l ver
sion
of
reco
rd.

2. Mechanisms of acquired brain injury
Two of the most frequent and heavily studied forms of acquired brain injury are TBI
and stroke. Although they have different etiologies, TBI and stroke share many common
mechanisms of injury (Bramlett and Dietrich 2004; Leker and Shohami 2002; Werner and
Engelhard 2007). Both are characterized by a primary phase, in which there is rapid
necrotic death of cells at the core of the injury (in TBI, where the mechanical impact is
centered, and in stroke, where cerebral perfusion falls below a critical threshold). Cells in
the area surrounding the injury core (the penumbra) may be dysfunctional, but remain
viable for a short period. However, the penumbra region is vulnerable to a secondary phase
of injury that evolves in response to the primary injury. Within a few hours, the primary
infarct can expand to encompass nearly the whole penumbra, and damage may continue to
progress for weeks or months after the initial insult (Hossmann 2006; Leker and Shohami
2002; Liu et al. 2010; Park et al. 2008).
Secondary injury occurs via multiple parallel and interacting pathways. The
excessive release and/or inadequate astrocytic uptake of glutamate following the primary
injury results in the overstimulation of glutamate receptors. In particular, activation of the
NMDA receptor allows excessive entry of calcium into the cell, which activates several
calcium-sensitive enzymes, including proteases and phospholipases, which directly damage
cellular proteins and lipids (Kalia et al. 2008; Lo et al. 2003; Waxman and Lynch 2005). In
addition, a prolonged rise in intracellular calcium levels stimulates the formation of reactive
oxygen species, which also damage DNA and impair the integrity of cell and organelle
membranes by initiating lipid peroxidation. Calcium overload, paired with the production
Page 4 of 41Page 4 of 49B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIC
HIG
AN
on
06/1
3/13
For
pers
onal
use
onl
y. T
his
Just
-IN
man
uscr
ipt i
s th
e ac
cept
ed m
anus
crip
t pri
or to
cop
y ed
iting
and
pag
e co
mpo
sitio
n. I
t may
dif
fer
from
the
fina
l off
icia
l ver
sion
of
reco
rd.

of free radicals, results in an increase in the permeability of the inner mitochondrial
membrane, impairing the production of energy substrates by oxidative phosphorylation. In
addition, the disruption of ion gradients across the mitochondrial membrane leads to
mitochondrial swelling, damage of the outer mitochondrial membrane, and the release of
cytochrome c into the cytoplasm. Cytochrome c initiates an apoptotic cell death program,
culminating in the activation of the apoptosis factor Caspase 3, which cleaves a number of
key cellular proteins, leading to the death of the cell ( reviewed in Bramlett and Dietrich
2004; Doyle et al. 2008; Hossmann 2006; Leker and Shohami 2002; Mergenthaler et al.
2004). In addition, elevated calcium concentration in the axon results in the destruction of
structural and transport proteins, leading to axonal swelling and synaptic disconnection.
Such white matter injury can be widespread throughout the brain even after a localized
ischemic or traumatic event, which may explain the broad nature of neurological symptoms
observed after focal injury (Bramlett and Dietrich 2004; Leker and Shohami 2002).
TBI and ischemia both stimulate an inflammatory response, with infiltration of
immune cells into the brain. Invading neutrophils release oxygen free radicals and
proteolytic enzymes, which damage resident cells. In addition, there is an upregulation in
the production of pro-inflammatory cytokines by activated microglia, resulting in further
damage (Bramlett and Dietrich 2004; Lakhan et al. 2009; Leker and Shohami 2002; Loane
and Byrnes 2010). Astrocytes proliferate, undergo morphological alterations, and exhibit
changes in the expression of multiple proteins including intermediate filament proteins such
as glial fibrillary acidic protein (GFAP), in a process known as reactive astrogliosis
(Sofroniew 2009). Over time, reactive astrocytes form a glial scar, isolating the injury site
from the surrounding healthy tissue to prevent the spread of damage, but also limiting the
Page 5 of 41Page 5 of 49B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIC
HIG
AN
on
06/1
3/13
For
pers
onal
use
onl
y. T
his
Just
-IN
man
uscr
ipt i
s th
e ac
cept
ed m
anus
crip
t pri
or to
cop
y ed
iting
and
pag
e co
mpo
sitio
n. I
t may
dif
fer
from
the
fina
l off
icia
l ver
sion
of
reco
rd.

capacity for axonal regeneration and migration of neuroblasts into the injury site (Fitch and
Silver 2008; Silver and Miller 2004). Figure 1 summarizes the evolution of
neuropathological changes occurring after brain injury.
The neuropathological features observed in TBI and stroke described above also
apply to other brain injury models. We (Bani-Yaghoub et al. 2008; Jezierski et al. 2012;
Tay et al. 2011) and others (Frontczak-Baniewicz et al. 2008; Jadhav et al. 2007) have used
surgically-induced brain injury models to mimic the incidental damage inflicted on the
brain during neurosurgical procedures. Such damage can occur, for instance, during the
excision of brain tumours or the removal of epileptic foci. As in TBI and stroke, the injury
site is characterized by the infiltration of macrophages (Frontczak-Baniewicz et al. 2008;
Frontczak-Baniewicz et al. 2011), invasion by activated microglia, the formation of a glial
scar (Frontczak-Baniewicz et al. 2011), the apoptotic death of neurons (Sulejczak et al.
2008), and an expanding zone of infarction over time (Frontczak-Baniewicz et al. 2011).
Figure 2 illustrates some of these changes after surgical lesion to the mouse motor cortex.
3. The challenge of reconstructing functional brain tissue
Both TBI and stroke can result in pan-necrosis (the loss of all cellular elements)
within the injury core, as well as the loss or damage of selectively vulnerable cell types in
surrounding areas (Katchanov et al. 2003; Kotapka et al. 1991; Lowenstein et al. 1992).
Amongst surviving neurons, many synaptic connections are lost after injury (Brown et al.
2008; Campbell et al. 2012), compromising the integrity of circuits which may support
sensory, motor, or cognitive functions. Repair of this type of damage is not a trivial feat,
owing to the diversity of cell types and the complexity of the circuitry involved (Chiu and
Page 6 of 41Page 6 of 49B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIC
HIG
AN
on
06/1
3/13
For
pers
onal
use
onl
y. T
his
Just
-IN
man
uscr
ipt i
s th
e ac
cept
ed m
anus
crip
t pri
or to
cop
y ed
iting
and
pag
e co
mpo
sitio
n. I
t may
dif
fer
from
the
fina
l off
icia
l ver
sion
of
reco
rd.

Rao 2011). The morphological complexity of the tissue lost or damaged after traumatic or
ischemic injury to the cortex can be appreciated by Golgi-Cox staining (see Figure 3) which
emphasizes the enormity of the task of reconstructing functional brain tissue.
Excitatory pyramidal neurons, which extend long axonal projections to connect
with cells in distant brain regions, make up the majority (70-80%) of neocortical neurons
(Markram et al. 2004). While pyramidal neurons (Figure 3; arrows) have common
structural and physiological characteristics, they differ in terms of their location within the
cortical laminae and their patterns of connectivity (Markram et al. 2004; Molyneaux et al.
2007). For instance, pyramidal projection neurons can be classified as associative,
commisural, or corticofugal, based on their predominant output to other neocortical areas,
the contralateral hemisphere, or subcortical targets, respectively (Molyneaux et al. 2007).
Each pyramidal neuron also receives input from distinct cortical and subcortical regions
(Aronoff et al. 2010; Miyashita et al. 1994; Porter and White 1983), and each input arrives
at a layer-specific area of the cell's dendritic arbour (Spruston 2008; Zilles 1990). In
addition to pyramidal neurons, the cortex contains local-circuit interneurons (Figure 3;
arrowheads), which make up 20-30% of the neuron population. This is a diverse group of
cells which differ in terms of morphology, physiology, connectivity, neurochemistry,
molecular characteristics and location within the cortical laminae (Wonders and Anderson
2006).
Pyramidal cells and interneurons form an intricate circuitry within the cortex.
While pyramidal neurons provide the major output from the cortex, interneurons serve to
modulate cortical excitability, provide lateral inhibition, and affect the timing and
Page 7 of 41Page 7 of 49B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIC
HIG
AN
on
06/1
3/13
For
pers
onal
use
onl
y. T
his
Just
-IN
man
uscr
ipt i
s th
e ac
cept
ed m
anus
crip
t pri
or to
cop
y ed
iting
and
pag
e co
mpo
sitio
n. I
t may
dif
fer
from
the
fina
l off
icia
l ver
sion
of
reco
rd.

synchronization of neuronal activity across clusters of pyramidal neurons (Markram et al.
2004). It is estimated that each cortical pyramidal neuron makes roughly 1 x 104 synapses
(Huttenlocher 1990), and that there are approximately 1-2 x109
synapses per mm3 of
cortical neuropil (DeFelipe et al. 1999). This sophisticated circuitry arises over a long
period of time during development (months to years in humans) and involves the
elaboration of dendritic trees and axonal arbourizations, and the formation of synapses
(Huttenlocher 1990). Refinement of the circuitry by pruning of overabundant synapses
reportedly continues over decades in some cortical regions (Petanjek et al. 2011).Thus, a
wide variety of cells, which normally exist as part of a complex functional circuit formed
over a prolonged period of development, are lost after injury.
4. Limitations of endogenous brain repair after injury
In the aftermath of traumatic or ischemic brain injury, a number of processes are
initiated that might represent the brain’s attempt to mount an endogenous repair response
(Wieloch and Nikolich 2006). For instance, an increased proliferation of neural stem and
progenitor cells residing in the subventricular zone has been reported in several models of
injury and ischemia (Arvidsson et al. 2002; Chen et al. 2003; Chirumamilla et al. 2002; Jin
et al. 2001; Parent et al. 2002; Ramaswamy et al. 2005; Zhang et al. 2001). Although a
small proportion of these cells may successfully migrate to the damaged cortex or striatum,
the rate of survival of newly born cells is quite low, likely due to the cytotoxic environment
surrounding the injury. Furthermore, the majority of surviving cells differentiate into glia,
with only a small number of cells becoming neurons (Arvidsson et al. 2002; Kernie and
Page 8 of 41Page 8 of 49B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIC
HIG
AN
on
06/1
3/13
For
pers
onal
use
onl
y. T
his
Just
-IN
man
uscr
ipt i
s th
e ac
cept
ed m
anus
crip
t pri
or to
cop
y ed
iting
and
pag
e co
mpo
sitio
n. I
t may
dif
fer
from
the
fina
l off
icia
l ver
sion
of
reco
rd.

Parent 2010; Parent et al. 2002). Thus, while the adult brain is capable of supporting the
integration of newly born neurons into pre-existing circuitry, the generation of new neurons
to replace dead or damaged ones is not a widespread phenomenon (Chiu and Rao 2011),
and endogenous neurogenesis does not appear to occur on a scale that is sufficient to
replace a substantial portion of cells lost due to injury (Kernie and Parent 2010).
One reason for the lack of endogenous cell replacement after injury may be that the
specification of neuronal subtypes is a highly complex process that is not normally
recapitulated in the adult brain. During development, different cell types establish the
telencephalon in a spatiotemporal manner, encounter different extracellular signals, and
express different transcriptional programs, all of which influence their eventual neuronal
phenotype (Molyneaux et al. 2007; Sansom and Livesey 2009; Wonders and Anderson
2006). The signals that promote differentiation into particular neuronal subtypes, as well as
axonal extension towards specific targets, may not be present in the adult brain. The lack of
appropriate signals for neuronal specification and axonal targeting in the adult likely
hampers the ability of endogenous cells to replace the full complement of neuronal
subtypes lost after injury and to re-establish functional circuits.
Structural plasticity after brain injury also occurs in the form of axonal sprouting
and synaptogenesis (Carmichael 2003; Fitch and Silver 2008). Both TBI and ischemia
stimulate the expression of growth-promoting genes which mediate signaling within growth
cone membranes, reorganization of cytoskeletal elements, and axonal sprouting
(Carmichael et al. 2005; Stroemer et al. 1995). Subsequently, immunoreactivity for
synaptophysin, a protein found in synaptic vesicles, increases, suggesting that synapse
Page 9 of 41Page 9 of 49B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIC
HIG
AN
on
06/1
3/13
For
pers
onal
use
onl
y. T
his
Just
-IN
man
uscr
ipt i
s th
e ac
cept
ed m
anus
crip
t pri
or to
cop
y ed
iting
and
pag
e co
mpo
sitio
n. I
t may
dif
fer
from
the
fina
l off
icia
l ver
sion
of
reco
rd.

formation occurs in the penumbra (Stroemer et al. 1995). However, the inhibitory
environment surrounding an injury may be non-permissive for efficient sprouting and
synaptogenesis. Within the glial scar, growth-inhibitory molecules such as tenascin,
semaphorins, ephrins, and chondroitin sulfate proteoglycans are produced by reactive
astrocytes (Fitch and Silver 2008; Galtrey and Fawcett 2007; Gervasi et al. 2008). As
regenerating axons approach the glial scar, dystrophic endings form on the axon terminal,
and growth is halted (Silver and Miller 2004). Structural plasticity in the post-injury brain
is therefore limited.
5. Amniotic fluid as a source of cells for cell-based therapy
The notion that newly born cells may replace some of those lost or damaged by TBI
or stroke has stimulated an interest in developing cell-based therapies aimed at enhancing
the repair of damaged tissue after nervous system injury. Stem cells from various sources,
including embryonic stem (ES) cells, embryonic and adult neural stem (NS) cells and stem
cells derived from other adult sources such as adipose tissue and bone marrow have been
investigated as a potential therapy for experimental brain injury (reviewed in Boncoraglio
et al. 2010; Burns et al. 2009; Harting et al. 2008; Hess and Borlongan 2008; Honmou et
al. 2012; Jain 2009; Miljan and Sinden 2009; Stabenfeldt et al. 2011; Yu and Morshead
2011). Ultimately, the choice of the most appropriate cell source will be governed by
several factors, including accessibility, ability to generate sufficient quantities of cells,
safety, ethical restrictions and of course, efficacy. Each of the cell types that have
commonly been employed to date in preclinical testing has advantages and drawbacks. ES
cells, although possessing a high capacity for differentiation, have the potential to induce
Page 10 of 41Page 10 of 49B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIC
HIG
AN
on
06/1
3/13
For
pers
onal
use
onl
y. T
his
Just
-IN
man
uscr
ipt i
s th
e ac
cept
ed m
anus
crip
t pri
or to
cop
y ed
iting
and
pag
e co
mpo
sitio
n. I
t may
dif
fer
from
the
fina
l off
icia
l ver
sion
of
reco
rd.

tumour formation in the brain (Molcanyi et al. 2009; Riess et al. 2007). In addition, the
ethical debate over the use of cells from discarded embryos has not been resolved and poses
a barrier to the development of clinically applicable therapy. In contrast, while adult stem
cells pose a reduced risk of tumorigenicity and fewer ethical restrictions, difficulties
expanding adult stem cells from some sources in culture have limited their use (Wright et
al. 2006), although advances in the understanding of culture conditions that promote
expansion may overcome this limitation (eg. Ribeiro et al. 2012). Because each of these
cell types is associated with shortcomings that may impede the development of clinically
useful cell-based therapies, there is a demand for further research into alternative sources of
cells for use in regenerative medicine.
Cells derived from amniotic fluid (AF) have a number of characteristics that make
them attractive candidates as a potential source of cells for tissue engineering and cell
replacement strategies (Kaviani et al. 2001; Wilson et al. 2012). As a result, AF cells have
been extensively studied in a variety of experimental models of injury and disease over the
past decade (reviewed elsewhere; Joo et al. 2012; Klemmt et al. 2011; Parolini et al. 2009).
AF, which is routinely obtained by amniocentesis for prenatal screening and diagnosis,
contains a heterogeneous population of cells derived from fetal skin, gastrointestinal,
respiratory and urinary tracts (Fauza 2004). The suggestion that AF might contain
multipotent fetal-derived cells (Prusa and Hengstschlager 2002) led to the identification of
a small subpopulation of actively dividing AF cells which express OCT4, a marker of
pluripotent stem cells (Prusa et al. 2003). Subsequently, AF-derived cells selected based
on the expression of stem cell factor (C-KIT, CD117) were found to express OCT4 along
with a number of markers of somatic stem cells such as CD29, CD44, CD73, CD90 and
Page 11 of 41Page 11 of 49B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIC
HIG
AN
on
06/1
3/13
For
pers
onal
use
onl
y. T
his
Just
-IN
man
uscr
ipt i
s th
e ac
cept
ed m
anus
crip
t pri
or to
cop
y ed
iting
and
pag
e co
mpo
sitio
n. I
t may
dif
fer
from
the
fina
l off
icia
l ver
sion
of
reco
rd.

CD105 (De Coppi et al. 2007). The multi-lineage potential of AF-derived cells was
reported by De Coppi et al (2007), who demonstrated that clonal cell lines derived from C-
KIT+ AF cells were capable of differentiating into cell types representing all three germ
layers. A recent genome-wide analysis of AF cells has shown that these cells have a distinct
transcriptional signature, compared with other stem cells (Maguire et al. 2013). Unlike stem
cells derived from adult sources, AF cells are easily cultured and can be used to generate a
large number of cells. For instance, we have shown that a minimum of 2-4x108
cells can be
produced from a single cell clone after only 9 passages (Figure 4A) while maintaining their
karyotypic stability (Figure 4B). An important feature of these cells is that they express
markers of the neural lineage, such as Nestin, Brn-2 (pou3f2) and Neurofilament (see
Figure 5A-D), suggesting that distinct populations of AF cells may have a higher potential
to differentiate along the neural lineage. The use of more specific markers (e.g. for neural
progenitors, neurons and glia) will provide a better understanding of the differentiation
capacity of these cells.
In general, transplanted AF cells may contribute to nervous system repair in two
different ways. Exogenous cells may serve as direct replacements for lost or damaged cells,
which requires that the cells differentiate into the appropriate neuronal subtypes, acquire
functional properties of the desired cell types, and integrate into the existing circuitry.
Another possibility is that transplanted cells may provide support for surviving host cells,
offer protection from the toxic environment surrounding the injury, and/or stimulate
endogenous repair mechanisms (Boucherie and Hermans 2009; De Feo et al. 2012; Einstein
and Ben-Hur 2008). Of course, these possibilities are not mutually exclusive. Here, we
Page 12 of 41Page 12 of 49B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIC
HIG
AN
on
06/1
3/13
For
pers
onal
use
onl
y. T
his
Just
-IN
man
uscr
ipt i
s th
e ac
cept
ed m
anus
crip
t pri
or to
cop
y ed
iting
and
pag
e co
mpo
sitio
n. I
t may
dif
fer
from
the
fina
l off
icia
l ver
sion
of
reco
rd.

evaluate the potential for using human AF cells for both cell replacement, and for support
functions in nervous system injury.
6. The application of AF cells as a neuron replacement strategy
Criteria for classifying a cell as a neuron have been previously described (Song et
al. 2002; Yang et al. 2011) and can be used to evaluate the neuronal differentiation capacity
of AF-derived cells. To be regarded as neurons, AF-derived cells should exhibit typical
neuronal morphology, including cellular polarization and the extension of neurites from the
cell body. This should be accompanied by the expression of neuron-specific gene products,
such as neurofilaments, microtubules, microtubule-associated proteins (MAP), and synaptic
proteins. In addition, putative neurons must display electrophysiological and neurochemical
characteristics that would enable intra- and intercellular communication. Thus, they should
be capable of maintaining a stable resting membrane potential and generating action
potentials with characteristic changes in membrane voltage when stimulated. Furthermore,
differentiated neurons should show evidence of synapse formation, by electrophysiological
recording or electron microscopic examination of pre- and post-synaptic specializations
(Song et al. 2002; Yang et al. 2011).
AF-derived cells have repeatedly been reported to acquire neuronal morphology and
express neuronal markers, including beta III tubulin, NeuN, MAP2, neurofilament, and
neuron-specific enolase (NSE) under conditions of neural induction in vitro (Bossolasco et
al. 2006; De Coppi et al. 2007; Jezierski et al. 2010; Kim et al. 2007; Mareschi et al. 2009;
Prusa et al. 2004; Tsai et al. 2006; Tsai et al. 2004). However, in the majority of reports,
neuronal differentiation has been assessed only by changes in morphology and expression
Page 13 of 41Page 13 of 49B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIC
HIG
AN
on
06/1
3/13
For
pers
onal
use
onl
y. T
his
Just
-IN
man
uscr
ipt i
s th
e ac
cept
ed m
anus
crip
t pri
or to
cop
y ed
iting
and
pag
e co
mpo
sitio
n. I
t may
dif
fer
from
the
fina
l off
icia
l ver
sion
of
reco
rd.

of neuronal markers, and these changes do not prove the neuronal functionality of the cells.
It has been reported that certain conditions used to induce neuronal differentiation, such as
serum withdrawal and addition of DMSO, can induce morphological changes in cells due to
disruption of the actin cytoskeleton, causing them to resemble neurons and express
neuronal antigens possibly as a stress response (Croft and Przyborski 2006; Lu et al. 2004;
Neuhuber et al. 2004). These changes do not appear to represent true neuronal
differentiation, as the neuronal characteristics may be unstable (Croft and Przyborski 2006).
For instance, when C-KIT+ AF cells were cultured in neural induction media (N2 media
plus 3-isobutyl-1-methylxanthine or 2’-O-dibutyryl-cAMP) a small population of cells
developed neuronal morphology and expression of BIII-tubulin (Prasongchean et al. 2012).
However, these changes were reversible upon removal of the induction media, indicating
that the cells had not undergone true neuronal differentiation (Prasongchean et al. 2012).
Although most studies use growth factors to induce neuronal differentiation (e.g. De Coppi
et al. 2007), rather than serum withdrawal, DMSO treatment or the neural induction media
used by Prasongchean et al (2012), these results highlight the need for functional studies to
validate the neuronal differentiation capacity of AF cells.
A limited number of studies have evaluated the functional properties of AF cells
after culture in neuronal differentiation media. Tsai et al. (2006) reported that after
dopaminergic induction, AF cells produced dopamine in response to depolarization induced
by high K+ stimulation, and work by De Coppi et al. (2007) suggested that neuronally
differentiated AF cells were capable of secreting l-glutamate. However, as pointed out by
Toselli et al (2008), this characteristic is not exclusive to neurons. Electrophysiological
studies suggest that after neuronal induction, AF cells express a barium-sensitive K+
Page 14 of 41Page 14 of 49B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIC
HIG
AN
on
06/1
3/13
For
pers
onal
use
onl
y. T
his
Just
-IN
man
uscr
ipt i
s th
e ac
cept
ed m
anus
crip
t pri
or to
cop
y ed
iting
and
pag
e co
mpo
sitio
n. I
t may
dif
fer
from
the
fina
l off
icia
l ver
sion
of
reco
rd.

channel (De Coppi et al. 2007) and a tetrodotoxin-sensitive voltage-gated sodium channel
(Mareschi et al. 2009), both of which are characteristic of neurons. However, as of yet, the
generation of action potentials by neuronally differentiated AF cells has not been
demonstrated, and evidence that AF cells can form functional synapses after neural
induction is completely lacking. Therefore, the ability of AF cells to differentiate into
mature functional neurons requires further investigation.
In a handful of in vivo studies, AF cells have been transplanted into the brain or
peripheral nerves after various types of injury (Bigini et al. 2012; Cipriani et al. 2007; De
Coppi et al. 2007; Donaldson et al. 2009; Pan et al. 2007; Pan et al. 2006; Prasongchean et
al. 2012; Rehni et al. 2007). In some cases, the phenotypes of transplanted cells were not
examined, so it is unclear whether the cells underwent neuronal differentiation. In other
studies, histological examination has generally failed to provide evidence that AF cells
differentiated into mature neurons. For example, the majority of human cells detected at 10,
30 and 90 days after transplantation of human AF cells into the striatum of rats subjected to
focal ischemia expressed the astrocytic marker GFAP (Cipriani et al. 2007). Although a
few human cells expressing doublecortin (a marker of immature neuroblasts) were found 10
days after transplantation, no cells expressing the neuronal marker beta III tubulin were
found at any time point (Cipriani et al. 2007). In another study, AF cells transplanted into
the striatum of rats treated with the dopaminergic toxin 6-OHDA failed to acquire a
dopaminergic phenotype, and survived for only a short period (less than three weeks) in the
brain (Donaldson et al. 2009). The ability of AF cells to integrate into host circuitry, and
express functional characteristics of neurons in vivo, has not been verified. Thus, there is so
Page 15 of 41Page 15 of 49B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIC
HIG
AN
on
06/1
3/13
For
pers
onal
use
onl
y. T
his
Just
-IN
man
uscr
ipt i
s th
e ac
cept
ed m
anus
crip
t pri
or to
cop
y ed
iting
and
pag
e co
mpo
sitio
n. I
t may
dif
fer
from
the
fina
l off
icia
l ver
sion
of
reco
rd.

far little evidence that AF-derived cells can be used to directly replace lost or damaged
neurons in the injured nervous system.
7. The application of AF cells for neuroprotection and repair
Even for cell types which have more convincingly been demonstrated to
differentiate into neurons in vitro and in vivo, it is not clear that replacement of lost cells is
the underlying mechanism for the improvements observed after their transplantation into
brain-injured animals (Boucherie and Hermans 2009; van Velthoven et al. 2009). In the
field of regenerative medicine, attention has shifted somewhat to the ability of transplanted
cells to provide neuroprotection and to stimulate repair of damaged nervous tissue
(Boucherie and Hermans 2009; Carletti et al. 2011; Chen et al. 2007; De Feo et al. 2012;
Farin et al. 2009; van Velthoven et al. 2009).
In fact, despite the limited evidence for neuronal differentiation of AF cells in vivo,
a number of studies have demonstrated beneficial effects of AF cell transplantation in
animal models of nervous system injury (summarized in Table 1). For instance, the
administration of AF cells of human or rat origin into a surgically-induced sciatic nerve gap
or nerve crush injury in rats has repeatedly been reported to result in significant functional
improvement, as well as enhanced recovery of compound action potential amplitude and
conduction latency (Cheng et al. 2010; Pan et al. 2009a; Pan et al. 2007; Pan et al. 2009b;
Pan et al. 2006; Pan et al. 2009c). Transplanted AF cells were not observed in the injury
site a month after surgery, and did not penetrate into the regenerating nerve, suggesting that
the observed improvements could not be accounted for by cell replacement (Pan et al.
2007). In a model of embryonic spinal cord thoracic crush injury, Prasongchean et al.
Page 16 of 41Page 16 of 49B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIC
HIG
AN
on
06/1
3/13
For
pers
onal
use
onl
y. T
his
Just
-IN
man
uscr
ipt i
s th
e ac
cept
ed m
anus
crip
t pri
or to
cop
y ed
iting
and
pag
e co
mpo
sitio
n. I
t may
dif
fer
from
the
fina
l off
icia
l ver
sion
of
reco
rd.

(2012) demonstrated that transplantation of C-KIT+ AF cells into the spinal cord at the
time of injury improved survival, even though grafted cells were negative for the neural
markers DCX and GFAP. In vitro co-culture of spinal cord slices with AF cells in a
transwell system prevented cell death and axonal degeneration, further supporting the
notion that the survival-promoting effect of AF cells was not due to differentiation and
integration into the tissue, but rather, due to secreted factors released by AF cells
(Prasongchean et al. 2012). AF cells injected into the lateral ventricles of mice three days
after focal ischemia induced by middle cerebral artery occlusion were shown to improve
somatosensory and cognitive function four days later (Rehni et al. 2007). Although the
authors did not examine the phenotype of injected cells, the fact that improvements were
observed after only a few days suggests that the beneficial effects of AF cells were likely
not due to cell replacement, since the differentiation and integration of transplanted cells
would presumably require a longer time.
7.1. Paracrine effects of AF cells
If AF cells do not replace lost or damaged cells in the injured nervous system, what
accounts for the beneficial effects of AF cell transplantation observed in the above
experiments? Studies using other stem cell types have suggested that transplanted cells may
enhance the survival of host neurons through the release of trophic factors, stimulate
endogenous repair through the recruitment of progenitor cells and promotion of neurite
outgrowth, and render the peri-lesion environment less toxic and more favourable to
regeneration by modulating the immune response and scar formation (Chiu and Rao 2011).
To date, the mechanisms by which damaged neural tissue may benefit from AF cells have
Page 17 of 41Page 17 of 49B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIC
HIG
AN
on
06/1
3/13
For
pers
onal
use
onl
y. T
his
Just
-IN
man
uscr
ipt i
s th
e ac
cept
ed m
anus
crip
t pri
or to
cop
y ed
iting
and
pag
e co
mpo
sitio
n. I
t may
dif
fer
from
the
fina
l off
icia
l ver
sion
of
reco
rd.

yet to be clearly identified. However, AF cells have been shown to express and secrete a
number of factors that could potentially support neuroprotective and/or reparative
functions. For instance, AF cells secrete vascular endothelial growth factor (VEGF),
stromal cell derived factor- 1 (CXCL12) and IL-8, all of which regulate angiogenesis
(Mirabella et al. 2011; Yoon et al. 2010), and in fact, the pro-angiogenic capacity of AF
cells has been demonstrated in mouse hindlimb ischemia (Mirabella et al. 2011) and
ischemic skin flap (Mirabella et al. 2012) models. Enhanced angiogenesis is also beneficial
in animal models of stroke (Fan and Yang 2007; Gertz et al. 2012; Ma et al. 2012),
traumatic brain injury (Li et al. 2012; Xiong et al. 2010) and nerve injury (Pereira Lopes et
al. 2011). Thus, the secretion of pro-angiogenic factors by AF cells might contribute to
improved function in models of nervous system injury. In addition, RT-PCR analysis has
indicated that some AF cells express a number of neurotrophic factors, such as BDNF,
GDNF, CNTF, NGF and NT-3 (Pan et al. 2007). Since neurotrophic factors have
frequently been shown to be neuroprotective in ischemic stroke models (Beck et al. 1994;
Duarte et al. 2012; Ferrer et al. 1998; Kiprianova et al. 1999; Kitagawa et al. 1998a;
Kitagawa et al. 1998b; Miyazaki et al. 1999; Schabitz et al. 1997; Wang et al. 1997;
Yamashita et al. 1997), traumatic brain injury (Minnich et al. 2010), and peripheral nerve
injury (Fine et al. 2002; Kokai et al. 2011; Wood et al. 2009), it is possible that some of the
beneficial effects of AF cells may be explained by the release of trophic factors. AF cells
have also been shown to secrete a number of immune-modulating cytokines such as Il-6,
and growth related oncogene (GRO) and monocyte chemotactic protein (MCP) family
members (Moorefield et al. 2011) which might serve to limit the damage after nervous
system injury. Whether secretion of angiogenic, trophic, or immune-modulating factors
Page 18 of 41Page 18 of 49B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIC
HIG
AN
on
06/1
3/13
For
pers
onal
use
onl
y. T
his
Just
-IN
man
uscr
ipt i
s th
e ac
cept
ed m
anus
crip
t pri
or to
cop
y ed
iting
and
pag
e co
mpo
sitio
n. I
t may
dif
fer
from
the
fina
l off
icia
l ver
sion
of
reco
rd.

actually does account for the positive effects of AF cell transplantation in neural injury
models remains to be determined. This issue warrants further investigation, in order to
capitalize on the beneficial properties of AF cells.
Beyond their natural capacity for providing protection and promoting repair of
neural tissue, AF cells can also be engineered to deliver specific factors to sites of nervous
system damage (see Figure 6). For instance, Cheng et al. (2010) transplanted AF cells
expressing human GDNF into the site of a sciatic nerve crush injury. Transplantation of
GDNF-transduced AF cells resulted in improved electrophysiological and functional
recovery, and prompted an upregulation of GDNF receptor expression at the distal end of
the crushed nerve (Cheng et al. 2010). We are currently evaluating the possibility that
GDNF-expressing AF cells might also be useful for the treatment of surgically-induced
brain injury.
7.2. Transfer of small molecules from AF cells to host tissue
An additional potential mechanism by which AF cells might modulate damage or
mediate repair in the injured nervous system is through the direct transfer of small species
to at-risk cells via gap junction channels. Gap junction channels are permeable to many
small molecules, including metabolites, amino acids, nucleotides and second messengers
(Harris 2007). One type of molecule of particular interest, owing to their capacity to
regulate multiple targets, are microRNAs (miRs). miRs are short (21-23 nucleotide) non-
coding RNA molecules that interact mainly with the 3’UTR of mRNA transcripts, resulting
in translational repression or degradation of the target mRNA (Bartel 2004; Nelson et al.
2003). A number of miRs, including miR-223 (Harraz et al. 2012), miR-181c (Zhang et al.
Page 19 of 41Page 19 of 49B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIC
HIG
AN
on
06/1
3/13
For
pers
onal
use
onl
y. T
his
Just
-IN
man
uscr
ipt i
s th
e ac
cept
ed m
anus
crip
t pri
or to
cop
y ed
iting
and
pag
e co
mpo
sitio
n. I
t may
dif
fer
from
the
fina
l off
icia
l ver
sion
of
reco
rd.

2012a), and miR-21 (Buller et al. 2010; Zhang et al. 2012b), have been shown to confer
neuroprotection in in vitro or in vivo models of neural injury, and thus the delivery of
specific miRs to the injured nervous system may represent a valid therapeutic strategy.
Previous work has shown that miRs can pass between pairs of glioma cells (Katakowski et
al. 2010), cardiac myocytes (Kizana et al. 2009) and from bone marrow stromal cells to
breast cancer cells (Lim et al. 2011) likely through gap junctions composed of connexin 43
(Cx43), to affect target genes in the receiving cell. We have hypothesized that AF cells
might also be capable of transferring miRs to host cells in the injured brain via gap
junctions to promote cell survival or repair.
AF cells form gap junction channels made of CX43 (Figure 5D and Jezierski et al.
2012). Dye transfer experiments in our laboratory indicated that AF cells can form
functional gap junctions with other AF cells and with cortical cells in vitro (Jezierski et al.
2012). Interestingly, the expression of Cx43 is upregulated after brain injury (Haupt et al.
2007; Ohsumi et al. 2010; Rouach et al. 2002), possibly increasing the likelihood that
transplanted cells might be able to establish gap junctional communication with vulnerable
host tissue.
To test the capacity of AF cells to transfer miRs through gap junctions, we injected
a fluorescently labelled morpholino mimic of miR-21 into individual AF cells, and
observed the successful transfer of fluorescent signal to neighbouring AF cells (Figure 7A-
D) and cortical astrocytes (Figure 7E-G) in vitro. The transfer of miRs from AF cells to
host cells therefore seems plausible. Whether AF cells can in fact transfer endogenous miRs
to injured neural tissue, which miRs are transferred, and whether this occurs on a scale
sufficient to impact the expression of miR target genes in the receiving cell remains to be
Page 20 of 41Page 20 of 49B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIC
HIG
AN
on
06/1
3/13
For
pers
onal
use
onl
y. T
his
Just
-IN
man
uscr
ipt i
s th
e ac
cept
ed m
anus
crip
t pri
or to
cop
y ed
iting
and
pag
e co
mpo
sitio
n. I
t may
dif
fer
from
the
fina
l off
icia
l ver
sion
of
reco
rd.

determined. An alternative approach might be to harness the ability of AF cells to transfer
miRs by engineering HAF cells to produce specific miRs that are expected to have
protective or reparative functions.
8. Future Directions
The reported neural differentiation potential of AF–derived cells has stimulated an
interest in developing AF cell-based therapies for treating damaged or diseased neural
tissue. Research on the ability of AF cells to replace lost or damaged neurons in vivo is still
in its infancy, and major questions remain to be answered. One issue which needs further
exploration is whether certain subpopulations of AF cells might be more suitable for use in
neural applications than others. For instance, Arnhold et al (2011) reported enhanced
neuronal differentiation capacity of C-KIT- AF cells relative to C-KIT
+ cells, although
these differences have not yet been examined in vivo. Furthermore, the question of whether
AF cells should be pre-differentiated towards a neuronal fate or towards specific neuronal
subtypes in vitro prior to transplantation should be addressed. In order to do so, a better
understanding of the culture conditions needed to direct AF cells towards specific neuronal
subtypes will be necessary.
On the other hand, it is possible that AF cells may provide benefit independent of
any neuronal differentiation or integration into the host circuitry, and neuronal
differentiation of AF cells may not be required in order to achieve positive outcomes. In
fact, research thus far supports this idea, as transplantation of AF cells resulted in improved
outcomes after peripheral nerve injury and brain ischemia, without directly replacing lost
cells (Pan et al. 2007; Pan et al. 2006; Rehni et al. 2007). However, the mechanisms by
Page 21 of 41Page 21 of 49B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIC
HIG
AN
on
06/1
3/13
For
pers
onal
use
onl
y. T
his
Just
-IN
man
uscr
ipt i
s th
e ac
cept
ed m
anus
crip
t pri
or to
cop
y ed
iting
and
pag
e co
mpo
sitio
n. I
t may
dif
fer
from
the
fina
l off
icia
l ver
sion
of
reco
rd.

which AF cells provide neuroprotection and/or stimulate repair require further research.
Factors secreted by AF cells, and the conditions under which this occurs, should be
explored in greater detail.
References
Arnhold S, Gluer S, Hartmann K, Raabe O, Addicks K, Wenisch S and Hoopmann M.
2011, Amniotic-fluid stem cells: growth dynamics and differentiation potential after
a CD-117-based selection procedure, Stem Cells Int 2011:715341
Aronoff R, Matyas F, Mateo C, Ciron C, Schneider B and Petersen CCH. 2010, Long-range
connectivity of mouse primary somatosensory barrel cortex, EurJ Neurosci
31(12):2221-2233
Arvidsson A, Collin T, Kirik D, Kokaia Z and Lindvall O. 2002, Neuronal replacement
from endogenous precursors in the adult brain after stroke, Nat Med 8(9):963-970
Auriel E and Bornstein NM. 2010, Neuroprotection in acute ischemic stroke--current status,
J Cell Mol Med 14(9):2200-2202
Bani-Yaghoub M, Tremblay RG, Ajji A, Nzau M, Gangaraju S, Chitty D, Zurakowski B
and Sikorska M. 2008, Neuroregenerative strategies in the brain: emerging
significance of bone morphogenetic protein 7 (BMP7), Biochem Cell Biol
86(5):361-369
Bartel DP. 2004, MicroRNAs: genomics, biogenesis, mechanism, and function, Cell
116(2):281-297
Beck T, Lindholm D, Castren E and Wree A. 1994, Brain-derived neurotrophic factor
protects against ischemic cell damage in rat hippocampus, J Cereb Blood Flow
Metab 14(4):689-692
Bigini P, Diana V, Barbera S, Fumagalli E, Micotti E, Sitia L, Paladini A, Bisighini C, De
Grada L, Coloca L, Colombo L, Manca P, Bossolasco P, Malvestiti F, Fiordaliso F,
Forloni G, Morbidelli M, Salmona M, Giardino D, Mennini T, Moscatelli D, Silani
V and Cova L. 2012, Longitudinal tracking of human fetal cells labeled with super
paramagnetic iron oxide nanoparticles in the brain of mice with motor neuron
disease, PLoS One 7(2):e32326
Bjorklund A and Lindvall O. 2000, Cell replacement therapies for central nervous system
disorders, Nat Neurosci 3(6):537-544
Boncoraglio GB, Bersano A, Candelise L, Reynolds BA and Parati EA. 2010, Stem cell
transplantation for ischemic stroke, Cochrane Database Syst Rev(9):CD007231
Bossolasco P, Montemurro T, Cova L, Zangrossi S, Calzarossa C, Buiatiotis S, Soligo D,
Bosari S, Silani V, Deliliers GL, Rebulla P and Lazzari L. 2006, Molecular and
phenotypic characterization of human amniotic fluid cells and their differentiation
potential, Cell Res 16(4):329-336
Boucherie C and Hermans E. 2009, Adult stem cell therapies for neurological disorders:
benefits beyond neuronal replacement? J Neurosci Res 87(7):1509-1521
Bramlett HM and Dietrich WD. 2004, Pathophysiology of cerebral ischemia and brain
trauma: similarities and differences, J Cereb Blood Flow Metab 24(2):133-150
Page 22 of 41Page 22 of 49B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIC
HIG
AN
on
06/1
3/13
For
pers
onal
use
onl
y. T
his
Just
-IN
man
uscr
ipt i
s th
e ac
cept
ed m
anus
crip
t pri
or to
cop
y ed
iting
and
pag
e co
mpo
sitio
n. I
t may
dif
fer
from
the
fina
l off
icia
l ver
sion
of
reco
rd.

Brown CE, Wong C and Murphy TH. 2008, Rapid morphologic plasticity of peri-infarct
dendritic spines after focal ischemic stroke, Stroke 39(4):1286-1291
Buller B, Liu X, Wang X, Zhang RL, Zhang L, Hozeska-Solgot A, Chopp M and Zhang
ZG. 2010, MicroRNA-21 protects neurons from ischemic death, FEBS J
277(20):4299-4307
Burns TC, Verfaillie CM and Low WC. 2009, Stem cells for ischemic brain injury: a
critical review, J Comp Neurol 515(1):125-144
Campbell JN, Low B, Kurz JE, Patel SS, Young MT and Churn SB. 2012, Mechanisms of
dendritic spine remodeling in a rat model of traumatic brain injury, J Neurotrauma
29(2):218-234
Carletti B, Piemonte F and Rossi F. 2011, Neuroprotection: the emerging concept of
restorative neural stem cell biology for the treatment of neurodegenerative diseases,
Curr Neuropharmacol 9(2):313-317
Carmichael ST. 2003, Plasticity of cortical projections after stroke, Neuroscientist 9(1):64-
75
Carmichael ST, Archibeque I, Luke L, Nolan T, Momiy J and Li S. 2005, Growth-
associated gene expression after stroke: evidence for a growth-promoting region in
peri-infarct cortex, Exp Neurol 193(2):291-311
Centers for Disease Control and Prevention NCfIPaC. 2012 Injury Prevention &
Control:Traumatic Brain Injury Atlanta, GA.
Chen HI, Bakshi A, Royo NC, Magge SN and Watson DJ. 2007, Neural stem cells as
biological minipumps: a faster route to cell therapy for the CNS? Curr Stem Cell
Res Ther 2(1):13-22
Chen XH, Iwata A, Nonaka M, Browne KD and Smith DH. 2003, Neurogenesis and glial
proliferation persist for at least one year in the subventricular zone following brain
trauma in rats, J Neurotrauma 20(7):623-631
Cheng FC, Tai MH, Sheu ML, Chen CJ, Yang DY, Su HL, Ho SP, Lai SZ and Pan HC.
2010, Enhancement of regeneration with glia cell line-derived neurotrophic factor-
transduced human amniotic fluid mesenchymal stem cells after sciatic nerve crush
injury, J Neurosurg 112(4):868-879
Chirumamilla S, Sun D, Bullock MR and Colello RJ. 2002, Traumatic brain injury induced
cell proliferation in the adult mammalian central nervous system, J Neurotrauma
19(6):693-703
Chiu AY and Rao MS. 2011, Cell-based therapy for neural disorders--anticipating
challenges, Neurotherapeutics 8(4):744-752
Cipriani S, Bonini D, Marchina E, Balgkouranidou I, Caimi L, Grassi Zucconi G and
Barlati S. 2007, Mesenchymal cells from human amniotic fluid survive and migrate
after transplantation into adult rat brain, Cell Biol Int 31(8):845-850
Croft AP and Przyborski SA. 2006, Formation of neurons by non-neural adult stem cells:
potential mechanism implicates an artifact of growth in culture, Stem Cells
24(8):1841-1851
De Coppi P, Bartsch G, Siddiqui MM, Xu T, Santos CC, Perin L, Mostoslavsky G, Serre
AC, Snyder EY, Yoo JJ, Furth ME, Soker S and Atala A. 2007, Isolation of
amniotic stem cell lines with potential for therapy, Nat Biotech 25(1):100-106
Page 23 of 41Page 23 of 49B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIC
HIG
AN
on
06/1
3/13
For
pers
onal
use
onl
y. T
his
Just
-IN
man
uscr
ipt i
s th
e ac
cept
ed m
anus
crip
t pri
or to
cop
y ed
iting
and
pag
e co
mpo
sitio
n. I
t may
dif
fer
from
the
fina
l off
icia
l ver
sion
of
reco
rd.

De Feo D, Merlini A, Laterza C and Martino G. 2012, Neural stem cell transplantation in
central nervous system disorders: from cell replacement to neuroprotection, Curr
Opin Neurol 25(3):322-333
DeFelipe J, Marco P, Busturia I and Merchan-Perez A. 1999, Estimation of the number of
synapses in the cerebral cortex: methodological considerations, Cereb Cortex
9(7):722-732
De Silva MJ, Roberts I, Perel P, Edwards P, Kenward MG, Fernandes J, Shakur H and
Patel V. 2009, Patient outcome after traumatic brain injury in high-, middle- and
low-income countries: analysis of data on 8927 patients in 46 countries, Int J
Epidemiol 38(2):452-458
Donaldson AE, Cai J, Yang M and Iacovitti L. 2009, Human amniotic fluid stem cells do
not differentiate into dopamine neurons in vitro or after transplantation in vivo,
Stem Cells Dev 18(7):1003-1012
Doyle KP, Simon RP and Stenzel-Poore MP. 2008, Mechanisms of ischemic brain damage,
Neuropharmacology 55(3):310-318
Duarte EP, Curcio M, Canzoniero LM and Duarte CB. 2012, Neuroprotection by GDNF in
the ischemic brain, Growth Factors 30(4):242-257
Einstein O and Ben-Hur T. 2008, The changing face of neural stem cell therapy in
neurologic diseases, Arch Neurol 65(4):452-456
Fan Y and Yang GY. 2007, Therapeutic angiogenesis for brain ischemia: a brief review, J
Neuroimmune Pharmacol 2(3):284-289
Farin A, Liu CY, Langmoen IA and Apuzzo ML. 2009, Biological restoration of central
nervous system architecture and function: part 3-stem cell- and cell-based
applications and realities in the biological management of central nervous system
disorders: traumatic, vascular, and epilepsy disorders, Neurosurgery 65(5):831-859;
discussion 859
Fauza D. 2004, Amniotic fluid and placental stem cells, Best Pract Res Clin Obstet
Gynaecol 18(6):877-891
Ferrer I, Ballabriga J, Marti E, Perez E, Alberch J and Arenas E. 1998, BDNF up-regulates
TrkB protein and prevents the death of CA1 neurons following transient forebrain
ischemia, Brain Pathol 8(2):253-261
Fine EG, Decosterd I, Papaloizos M, Zurn AD and Aebischer P. 2002, GDNF and NGF
released by synthetic guidance channels support sciatic nerve regeneration across a
long gap, Eur J Neurosci 15(4):589-601
Fitch MT and Silver J. 2008, CNS injury, glial scars, and inflammation: Inhibitory
extracellular matrices and regeneration failure, Exp Neurol 209(2):294-301
Frechette M, Rennie K and Pappas BA. 2009, Developmental forebrain cholinergic lesion
and environmental enrichment: behaviour, CA1 cytoarchitecture and neurogenesis,
Brain Res 1252:172-182
Frontczak-Baniewicz M, Chrapusta SJ and Sulejczak D. 2008, Long-term consequences of
surgical brain injury - characteristics of the neurovascular unit and formation and
demise of the glial scar in a rat model, Folia Neuropathol 49(3):204-218
Galtrey CM and Fawcett JW. 2007, The role of chondroitin sulfate proteoglycans in
regeneration and plasticity in the central nervous system, Brain Res Rev 54(1):1-18
Page 24 of 41Page 24 of 49B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIC
HIG
AN
on
06/1
3/13
For
pers
onal
use
onl
y. T
his
Just
-IN
man
uscr
ipt i
s th
e ac
cept
ed m
anus
crip
t pri
or to
cop
y ed
iting
and
pag
e co
mpo
sitio
n. I
t may
dif
fer
from
the
fina
l off
icia
l ver
sion
of
reco
rd.

Gertz K, Kronenberg G, Kalin RE, Baldinger T, Werner C, Balkaya M, Eom GD,
Hellmann-Regen J, Krober J, Miller KR, Lindauer U, Laufs U, Dirnagl U, Heppner
FL and Endres M. 2012, Essential role of interleukin-6 in post-stroke angiogenesis,
Brain 135(Pt 6):1964-1980
Gervasi NM, Kwok JC and Fawcett JW. 2008, Role of extracellular factors in axon
regeneration in the CNS: implications for therapy, Regen Med 3(6):907-923
Ginsberg M, James D, Ding BS, Nolan D, Geng F, Butler JM, Schachterle W, Pulijaal VR,
Mathew S, Chasen ST, Xiang J, Rosenwaks Z, Shido K, Elemento O, Rabbany SY,
Rafii S. 2012, Efficient direct reprogramming of mature amniotic cells into
endothelial cells by ETS factors and TGFβ suppression. Cell 151(3):559-75
Glaser EM and Van der Loos H. 1981. Analysis of thick brain sections by obverse-reverse
computer microscopy: application of a new, high clarity Golgi-Nissl stain, J
Neurosci Methods 4(2):117-125
Gross AK, Norman J and Cook AM. 2010, Contemporary pharmacologic issues in the
management of traumatic brain injury, J Pharm Pract 23(5):425-440
Harraz MM, Eacker SM, Wang X, Dawson TM and Dawson VL. 2012, MicroRNA-223 is
neuroprotective by targeting glutamate receptors, Proc Natl Acad Sci U S A
109(46):18962-18967
Harris AL. 2007, Connexin channel permeability to cytoplasmic molecules, Prog Biophys
Mol Bio 94:120-143
Harting MT, Baumgartner JE, Worth LL, Ewing-Cobbs L, Gee AP, Day MC and Cox CS,
Jr. 2008, Cell therapies for traumatic brain injury, Neurosurg Focus 24(3-4):E18
Haupt C, Witte OW and Frahm C. 2007, Up-regulation of Connexin43 in the glial scar
following photothrombotic ischemic injury, Mol Cell Neurosci 35(1):89-99
Hess DC and Borlongan CV. 2008, Cell-based therapy in ischemic stroke, Expert Rev
Neurother 8(8):1193-1201
Honmou O, Onodera R, Sasaki M, Waxman SG and Kocsis JD. 2012, Mesenchymal stem
cells: therapeutic outlook for stroke, Trends Mol Med 18(5):292-297
Hossmann KA. 2006, Pathophysiology and therapy of experimental stroke, Cell Mol
Neurobiol 26(7-8):1057-1083
Huttenlocher PR. 1990, Morphometric study of human cerebral cortex development,
Neuropsychologia 28(6):517-527
Jadhav V, Solaroglu I, Obenaus A and Zhang JH. 2007, Neuroprotection against surgically
induced brain injury, Surg Neurol 67(1):15-20
Jain KK. 2009, Cell therapy for CNS trauma, Mol Biotechnol 42(3):367-376
Jezierski A, Gruslin A, Tremblay R, Ly D, Smith C, Turksen K, Sikorska M and Bani-
Yaghoub M. 2010, Probing stemness and neural commitment in human amniotic
fluid cells, Stem Cell Rev 6(2):199-214
Jezierski A, Rennie K, Tremblay R, Zurakowski B, Gruslin A, Sikorska M and Bani-
Yaghoub M. 2012, Human amniotic fluid cells form functional gap junctions with
cortical cells, Stem Cells Int 2012:607161
Jin K, Minami M, Lan JQ, Mao XO, Batteur S, Simon RP and Greenberg DA. 2001,
Neurogenesis in dentate subgranular zone and rostral subventricular zone after focal
cerebral ischemia in the rat, Proc Natl Acad Sci U S A 98(8):4710-4715
Page 25 of 41Page 25 of 49B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIC
HIG
AN
on
06/1
3/13
For
pers
onal
use
onl
y. T
his
Just
-IN
man
uscr
ipt i
s th
e ac
cept
ed m
anus
crip
t pri
or to
cop
y ed
iting
and
pag
e co
mpo
sitio
n. I
t may
dif
fer
from
the
fina
l off
icia
l ver
sion
of
reco
rd.

Joo S, Ko IK, Atala A, Yoo JJ and Lee SJ. 2012, Amniotic fluid-derived stem cells in
regenerative medicine research, Arch Pharm Res 35(2):271-280
Kalia LV, Kalia SK and Salter MW. 2008, NMDA receptors in clinical neurology:
excitatory times ahead, Lancet Neurol 7(8):742-755
Katakowski M, Buller B, Wang X, Rogers T and Chopp M. 2010, Functional microRNA is
transferred between glioma cells, Cancer Res 70(21):8259-8263
Katchanov J, Waeber C, Gertz K, Gietz A, Winter B, Bruck W, Dirnagl U, Veh RW and
Endres M. 2003, Selective neuronal vulnerability following mild focal brain
ischemia in the mouse, Brain Pathol 13(4):452-464
Kaviani A, Perry TE, Dzakovic A, Jennings RW, Ziegler MM, Fauza DO. 2001, The
amniotic fluid as a source of cells for fetal tissue engineering. J Pediatr Surg
36(11):1662-5.
Kernie SG and Parent JM. 2010, Forebrain neurogenesis after focal ischemic and traumatic
brain injury, Neurobiol Dis 37(2):267-274
Kim J, Lee Y, Kim H, Hwang KJ, Kwon HC, Kim SK, Cho DJ, Kang SG and You J. 2007,
Human amniotic fluid-derived stem cells have characteristics of multipotent stem
cells, Cell Prolif 40(1):75-90
Kiprianova I, Freiman TM, Desiderato S, Schwab S, Galmbacher R, Gillardon F and
Spranger M. 1999, Brain-derived neurotrophic factor prevents neuronal death and
glial activation after global ischemia in the rat, J Neurosci Res 56(1):21-27
Kitagawa H, Abe K, Hayashi T, Mitsumoto Y, Koga N and Itoyama Y. 1998a,
Ameliorative effect of glial cell line-derived neurotrophic factor on brain edema
formation after permanent middle cerebral artery occlusion in rats, Neurol Res
20(4):333-336
Kitagawa H, Hayashi T, Mitsumoto Y, Koga N, Itoyama Y and Abe K. 1998b, Reduction
of ischemic brain injury by topical application of glial cell line-derived neurotrophic
factor after permanent middle cerebral artery occlusion in rats, Stroke 29(7):1417-
1422
Kizana E, Cingolani E and Marban E. 2009, Non-cell-autonomous effects of vector-
expressed regulatory RNAs in mammalian heart cells, Gene Ther 16(9):1163-1168
Klemmt PA, Vafaizadeh V and Groner B. 2011, The potential of amniotic fluid stem cells
for cellular therapy and tissue engineering, Expert Opin Biol Ther 11(10):1297-
1314
Kokai LE, Bourbeau D, Weber D, McAtee J and Marra KG. 2011, Sustained growth factor
delivery promotes axonal regeneration in long gap peripheral nerve repair, Tissue
Eng Part A 17(9-10):1263-1275
Kotapka MJ, Gennarelli TA, Graham DI, Adams JH, Thibault LE, Ross DT and Ford I.
1991, Selective vulnerability of hippocampal neurons in acceleration-induced
experimental head injury, J Neurotrauma 8(4):247-258
Labiche LA and Grotta JC. 2004, Clinical trials for cytoprotection in stroke, NeuroRx
1(1):46-70
Lakhan SE, Kirchgessner A and Hofer M. 2009, Inflammatory mechanisms in ischemic
stroke: therapeutic approaches, J Transl Med 7:97
Page 26 of 41Page 26 of 49B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIC
HIG
AN
on
06/1
3/13
For
pers
onal
use
onl
y. T
his
Just
-IN
man
uscr
ipt i
s th
e ac
cept
ed m
anus
crip
t pri
or to
cop
y ed
iting
and
pag
e co
mpo
sitio
n. I
t may
dif
fer
from
the
fina
l off
icia
l ver
sion
of
reco
rd.

Leker RR and Shohami E. 2002, Cerebral ischemia and trauma-different etiologies yet
similar mechanisms: neuroprotective opportunities, Brain Res Brain Res Rev
39(1):55-73
Li S, Wei M, Zhou Z, Wang B, Zhao X and Zhang J. 2012, SDF-1alpha induces
angiogenesis after traumatic brain injury, Brain Res 1444:76-86
Lim PK, Bliss SA, Patel SA, Taborga M, Dave MA, Gregory LA, Greco SJ, Bryan M,
Patel PS and Rameshwar P. 2011, Gap junction-mediated import of microRNA
from bone marrow stromal cells can elicit cell cycle quiescence in breast cancer
cells, Cancer Res 71(5):1550-1560
Liu S, Levine SR and Winn HR. 2010, Targeting ischemic penumbra: part I - from
pathophysiology to therapeutic strategy, J Exp Stroke Transl Med 3(1):47-55
Liu T, Guo L, Liu Z, Huang Y and Cheng W. 2011, Induction of dopaminergic neuronal-
like cells from CD44+ human amniotic fluids that are ameliorative to behavioural
recovery in a Parkinson's disease rat model, Int J Mol Med 28(5): 745-752
Lo EH, Dalkara T and Moskowitz MA. 2003, Mechanisms, challenges and opportunities in
stroke, Nat Rev Neurosci 4(5):399-414
Loane DJ and Byrnes KR. 2010, Role of microglia in neurotrauma, Neurotherapeutics
7(4):366-377
Lowenstein DH, Thomas MJ, Smith DH and McIntosh TK. 1992, Selective vulnerability of
dentate hilar neurons following traumatic brain injury: a potential mechanistic link
between head trauma and disorders of the hippocampus, J Neurosci 12(12):4846-
4853
Lu P, Blesch A and Tuszynski MH. 2004, Induction of bone marrow stromal cells to
neurons: differentiation, transdifferentiation, or artifact?, J Neurosci Res 77(2):174-
191
Ma Y, Zechariah A, Qu Y and Hermann DM. 2012, Effects of vascular endothelial growth
factor in ischemic stroke, J Neurosci Res 90(10):1873-1882
Maguire CT, Demarest BL, Hill JT, Palmer JD, Brothman AR, Yost HJ, Condic ML. 2013,
Genome-wide analysis reveals the unique stem cell identity of human amniocytes.
PLoS One 8(1):e53372.
Mareschi K, Rustichelli D, Comunanza V, De Fazio R, Cravero C, Morterra G, Martinoglio
B, Medico E, Carbone E, Benedetto C and Fagioli F. 2009, Multipotent
mesenchymal stem cells from amniotic fluid originate neural precursors with
functional voltage-gated sodium channels, Cytotherapy 11(5):534-547
Markram H, Toledo-Rodriguez M, Wang Y, Gupta A, Silberberg G and Wu C. 2004,
Interneurons of the neocortical inhibitory system, Nat Rev Neurosci 5(10):793-807
Mergenthaler P, Dirnagl U and Meisel A. 2004, Pathophysiology of stroke: lessons from
animal models, Metab Brain Dis 19(3-4):151-167
Miljan EA and Sinden JD. 2009, Stem cell treatment of ischemic brain injury, Curr Opin
Mol Ther 11(4):394-403
Minnich JE, Mann SL, Stock M, Stolzenbach KA, Mortell BM, Soderstrom KE, Bohn MC
and Kozlowski DA. 2010, Glial cell line-derived neurotrophic factor (GDNF) gene
delivery protects cortical neurons from dying following a traumatic brain injury,
Restor Neurol Neurosci 28(3):293-309
Page 27 of 41Page 27 of 49B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIC
HIG
AN
on
06/1
3/13
For
pers
onal
use
onl
y. T
his
Just
-IN
man
uscr
ipt i
s th
e ac
cept
ed m
anus
crip
t pri
or to
cop
y ed
iting
and
pag
e co
mpo
sitio
n. I
t may
dif
fer
from
the
fina
l off
icia
l ver
sion
of
reco
rd.

Mirabella T, Cilli M, Carlone S, Cancedda R and Gentili C. 2011, Amniotic liquid derived
stem cells as reservoir of secreted angiogenic factors capable of stimulating neo-
arteriogenesis in an ischemic model, Biomaterials 32(15):3689-3699
Mirabella T, Hartinger J, Lorandi C, Gentili C, van Griensven M and Cancedda R. 2012,
Proangiogenic soluble factors from amniotic fluid stem cells mediate the
recruitment of endothelial progenitors in a model of ischemic fasciocutaneous flap,
Stem Cells Dev 21(12):2179-2188
Miyashita E, Keller A and Asanuma H. 1994, Input-output organization of the rat vibrissal
motor cortex, Exp Brain Res 99(2):223-232
Miyazaki H, Okuma Y, Fujii Y, Nagashima K and Nomura Y. 1999, Glial cell line-derived
neurotrophic factor protects against delayed neuronal death after transient forebrain
ischemia in rats, Neuroscience 89(3):643-647
Molcanyi M, Riess P, Haj-Yasein NN, Bentz K, Loehr M, Kuchta J, Zivcak J, Stenzel W,
Miletic H, Hescheler J, Neugebauer E, Hampl JA, Ernestus RI and Schafer U. 2009,
Developmental potential of the murine embryonic stem cells transplanted into the
healthy rat brain--novel insights into tumorigenesis, Cell Physiol Biochem 24(1-
2):87-94
Molyneaux BJ, Arlotta P, Menezes JR and Macklis JD. 2007, Neuronal subtype
specification in the cerebral cortex, Nat Rev Neurosci 8(6):427-437
Moorefield EC, McKee EE, Solchaga L, Orlando G, Yoo JJ, Walker S, Furth ME and
Bishop CE. 2011, Cloned, CD117 selected human amniotic fluid stem cells are
capable of modulating the immune response, PLoS One 6(10):e26535
Nelson P, Kiriakidou M, Sharma A, Maniataki E and Mourelatos Z. 2003, The microRNA
world: small is mighty, Trends Biochem Sci 28(10):534-540
Neuhuber B, Gallo G, Howard L, Kostura L, Mackay A and Fischer I. 2004, Reevaluation
of in vitro differentiation protocols for bone marrow stromal cells: disruption of
actin cytoskeleton induces rapid morphological changes and mimics neuronal
phenotype, J Neurosci Res 77(2):192-204
Ohsumi A, Nawashiro H, Otani N, Ooigawa H, Toyooka T and Shima K. 2010, Temporal
and spatial profile of phosphorylated connexin43 after traumatic brain injury in rats,
J Neurotrauma 27(7):1255-1263
Pan HC, Chen CJ, Cheng FC, Ho SP, Liu MJ, Hwang SM, Chang MH and Wang YC.
2009a, Combination of G-CSF administration and human amniotic fluid
mesenchymal stem cell transplantation promotes peripheral nerve regeneration,
Neurochem Res 34(3):518-527
Pan HC, Cheng FC, Chen CJ, Lai SZ, Lee CW, Yang DY, Chang MH and Ho SP. 2007,
Post-injury regeneration in rat sciatic nerve facilitated by neurotrophic factors
secreted by amniotic fluid mesenchymal stem cells, J Clin Neurosci 14(11):1089-
1098
Pan HC, Chin CS, Yang DY, Ho SP, Chen CJ, Hwang SM, Chang MH and Cheng FC.
2009b, Human amniotic fluid mesenchymal stem cells in combination with
hyperbaric oxygen augment peripheral nerve regeneration, Neurochem Res
34(7):1304-1316
Page 28 of 41Page 28 of 49B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIC
HIG
AN
on
06/1
3/13
For
pers
onal
use
onl
y. T
his
Just
-IN
man
uscr
ipt i
s th
e ac
cept
ed m
anus
crip
t pri
or to
cop
y ed
iting
and
pag
e co
mpo
sitio
n. I
t may
dif
fer
from
the
fina
l off
icia
l ver
sion
of
reco
rd.

Pan HC, Yang DY, Chiu YT, Lai SZ, Wang YC, Chang MH and Cheng FC. 2006,
Enhanced regeneration in injured sciatic nerve by human amniotic mesenchymal
stem cell, J Clin Neurosci 13(5):570-575
Pan HC, Yang DY, Ho SP, Sheu ML, Chen CJ, Hwang SM, Chang MH and Cheng FC.
2009c, Escalated regeneration in sciatic nerve crush injury by the combined therapy
of human amniotic fluid mesenchymal stem cells and fermented soybean extracts,
Natto, J Biomed Sci 16:75
Parent JM, Vexler ZS, Gong C, Derugin N and Ferriero DM. 2002, Rat forebrain
neurogenesis and striatal neuron replacement after focal stroke, Ann Neurol
52(6):802-813
Park E, Bell JD and Baker AJ. 2008, Traumatic brain injury: can the consequences be
stopped?, CMAJ 178(9):1163-1170
Parolini O, Soncini M, Evangelista M and Schmidt D. 2009, Amniotic membrane and
amniotic fluid-derived cells: potential tools for regenerative medicine? Regen Med
4(2):275-291
Pereira Lopes FR, Lisboa BC, Frattini F, Almeida FM, Tomaz MA, Matsumoto PK,
Langone F, Lora S, Melo PA, Borojevic R, Han SW and Martinez AM. 2011,
Enhancement of sciatic nerve regeneration after vascular endothelial growth factor
(VEGF) gene therapy, Neuropathol Appl Neurobiol 37(6):600-612
Petanjek Z, Judas M, Simic G, Rasin MR, Uylings HB, Rakic P and Kostovic I. 2011,
Extraordinary neoteny of synaptic spines in the human prefrontal cortex, Proc Natl
Acad Sci U S A 108(32):13281-13286
Porter LL and White EL. 1983, Afferent and efferent pathways of the vibrissal region of
primary motor cortex in the mouse, J Comp Neurol 214(3):279-289
Prasongchean W, Bagni M, Calzarossa C, De Coppi P and Ferretti P. 2012, Amniotic fluid
stem cells increase embryo survival following injury, Stem Cells Dev 21(5):675-688
Prusa AR and Hengstschlager M. 2002, Amniotic fluid cells and human stem cell research:
a new connection, Med Sci Monit 8(11):RA253-257
Prusa AR, Marton E, Rosner M, Bernaschek G and Hengstschlager M. 2003, Oct-4-
expressing cells in human amniotic fluid: a new source for stem cell research?, Hum
Reprod 18(7):1489-1493
Prusa AR, Marton E, Rosner M, Bettelheim D, Lubec G, Pollack A, Bernaschek G and
Hengstschlager M. 2004, Neurogenic cells in human amniotic fluid, Am J Obstet
Gynecol 191(1):309-314
Ramaswamy S, Goings GE, Soderstrom KE, Szele FG and Kozlowski DA. 2005, Cellular
proliferation and migration following a controlled cortical impact in the mouse,
Brain Res 1053(1-2):38-53
Rehni AK, Singh N, Jaggi AS and Singh M. 2007, Amniotic fluid derived stem cells
ameliorate focal cerebral ischaemia-reperfusion injury induced behavioural deficits
in mice, Behav Brain Res 183(1):95-100
Ribeiro, D, Laguna Goya R, Ravindran G, Vuono R, Parish CL, Foldi C, Piroth T, Yang S,
Parmar M, Nikkhah G, Hjerling-Leffler J, Lindvall O, Barker RA and Arenas E.
2012. Efficient expansion and dopaminergic differentiation of human fetal ventral
midbrain neural stem cells by midbrain morphogens, Neurobiol Dis 49C:118-127
Page 29 of 41Page 29 of 49B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIC
HIG
AN
on
06/1
3/13
For
pers
onal
use
onl
y. T
his
Just
-IN
man
uscr
ipt i
s th
e ac
cept
ed m
anus
crip
t pri
or to
cop
y ed
iting
and
pag
e co
mpo
sitio
n. I
t may
dif
fer
from
the
fina
l off
icia
l ver
sion
of
reco
rd.

Riess P, Molcanyi M, Bentz K, Maegele M, Simanski C, Carlitscheck C, Schneider A,
Hescheler J, Bouillon B, Schafer U and Neugebauer E. 2007, Embryonic stem cell
transplantation after experimental traumatic brain injury dramatically improves
neurological outcome, but may cause tumors, J Neurotrauma 24(1):216-225
Rouach N, Avignone E, Meme W, Koulakoff A, Venance L, Blomstrand F and Giaume C.
2002, Gap junctions and connexin expression in the normal and pathological central
nervous system, Biol Cell 94(7-8):457-475
Sansom SN and Livesey FJ. 2009, Gradients in the brain: the control of the development of
form and function in the cerebral cortex, Cold Spring Harb Perspect Biol
1(2):a002519
Schabitz WR, Schwab S, Spranger M and Hacke W. 1997, Intraventricular brain-derived
neurotrophic factor reduces infarct size after focal cerebral ischemia in rats, J Cereb
Blood Flow Metab 17(5):500-506
Schouten JW, Fulp CT, Royo NC, Saatman KE, Watson DJ, Snyder EY, Trojanowski JQ,
Prockop DJ, Maas AI and McIntosh TK. 2004, A review and rationale for the use of
cellular transplantation as a therapeutic strategy for traumatic brain injury, J
Neurotrauma 21(11):1501-1538
Silver J and Miller JH. 2004, Regeneration beyond the glial scar, Nat Rev Neurosci
5(2):146-156
Sofroniew MV. 2009, Molecular dissection of reactive astrogliosis and glial scar formation,
Trends Neurosci 32(12):638-647
Soler R, Füllhase C, Hanson A, Campeau L, Santos C and Andersson KE.2012, Stem cell
therapy ameliorates bladder dysfunction in an animal model of Parkinson disease, J
Urol 187(4): 1491-1497
Song HJ, Stevens CF and Gage FH. 2002, Neural stem cells from adult hippocampus
develop essential properties of functional CNS neurons, Nat Neurosci 5(5):438-445
Spruston N. 2008, Pyramidal neurons: dendritic structure and synaptic integration, Nat Rev
Neurosci 9(3):206-221
Stabenfeldt SE, Irons HR and Laplaca MC. 2011, Stem cells and bioactive scaffolds as a
treatment for traumatic brain injury, Curr Stem Cell Res Ther 6(3):208-220
Stroemer RP, Kent TA and Hulsebosch CE. 1995, Neocortical neural sprouting,
synaptogenesis, and behavioral recovery after neocortical infarction in rats, Stroke
26(11):2135-2144
Sulejczak D, Grieb P, Walski M and Frontczak-Baniewicz M. 2008, Apoptotic death of
cortical neurons following surgical brain injury, Folia Neuropathol 46(3):213-219
Tajiri N, Acosta S, Glover LE, Bickford PC, Jacotte Simancas A, Yasuhara T, Date I,
Solomita MA, Antonucci I, Stuppia L, Kaneko Y and Borlongan CV. 2012,
Intravenous grafts of amniotic fluid-derived stem cells induce endogenous cell
proliferation and attenuate behavioral deficits in ischemic stroke rats, PLOS One
7(8): e43779
Tay LL, Tremblay RG, Hulse J, Zurakowski B, Thompson M and Bani-Yaghoub M. 2011,
Detection of acute brain injury by Raman spectral signature, Analyst 136(8):1620-
1626
The Ontario Brain Injury Association. 2011 Ontario Brain Injury Association, Ontario
Brain Injury Association, St. Catharines, ON.
Page 30 of 41Page 30 of 49B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIC
HIG
AN
on
06/1
3/13
For
pers
onal
use
onl
y. T
his
Just
-IN
man
uscr
ipt i
s th
e ac
cept
ed m
anus
crip
t pri
or to
cop
y ed
iting
and
pag
e co
mpo
sitio
n. I
t may
dif
fer
from
the
fina
l off
icia
l ver
sion
of
reco
rd.

Toselli M, Cerbai E, Rossi F and Cattaneo E. 2008, Do amniotic fluid-derived stem cells
differentiate into neurons in vitro?, Nat Biotechnol 26(3):269-270
Tsai MS, Hwang SM, Tsai YL, Cheng FC, Lee JL and Chang YJ. 2006, Clonal amniotic
fluid-derived stem cells express characteristics of both mesenchymal and neural
stem cells, Biol Reprod 74(3):545-551
Tsai MS, Lee JL, Chang YJ and Hwang SM. 2004, Isolation of human multipotent
mesenchymal stem cells from second-trimester amniotic fluid using a novel two-
stage culture protocol, Hum Reprod 19(6):1450-1456
van Velthoven CT, Kavelaars A, van Bel F and Heijnen CJ. 2009, Regeneration of the
ischemic brain by engineered stem cells: fuelling endogenous repair processes,
Brain Res Rev 61(1):1-13
Wang Y, Lin SZ, Chiou AL, Williams LR and Hoffer BJ. 1997, Glial cell line-derived
neurotrophic factor protects against ischemia-induced injury in the cerebral cortex, J
Neurosci 17(11):4341-4348
Waxman EA and Lynch DR. 2005, N-methyl-D-aspartate receptor subtypes: multiple roles
in excitotoxicity and neurological disease, Neuroscientist 11(1):37-49
Werner C and Engelhard K. 2007, Pathophysiology of traumatic brain injury, Brit J
Anaesth 99(1):4-9
Wieloch T and Nikolich K. 2006, Mechanisms of neural plasticity following brain injury,
Curr Opin Neurobiol 16(3):258-264
Wilson PG, Devkota L, Payne T, Crisp L, Winter A, Wang Z. 2012, Clonal populations of
amniotic cells by dilution and direct plating: evidence for hidden diversity. Stem
Cells Int. 2012
Wonders CP and Anderson SA. 2006, The origin and specification of cortical interneurons,
Nat Rev Neurosci 7(9):687-696
Wood MD, Moore AM, Hunter DA, Tuffaha S, Borschel GH, Mackinnon SE and
Sakiyama-Elbert SE. 2009, Affinity-based release of glial-derived neurotrophic
factor from fibrin matrices enhances sciatic nerve regeneration, Acta Biomater
5(4):959-968
Wright LS, Prowse KR, Wallace K, Linskens MH and Svendsen CN. 2006, Human
progenitor cells isolated from the developing cortex undergo decreased
neurogenesis and eventual senescence following expansion in vitro, Exp Cell Res
312(11):2107-2120
Xiong Y, Mahmood A and Chopp M. 2010, Angiogenesis, neurogenesis and brain recovery
of function following injury, Curr Opin Investig Drugs 11(3):298-308
Yamashita K, Wiessner C, Lindholm D, Thoenen H and Hossmann KA. 1997, Post-
occlusion treatment with BDNF reduces infarct size in a model of permanent
occlusion of the middle cerebral artery in rat, Metab Brain Dis 12(4):271-280
Yang N, Ng YH, Pang ZP, Sudhof TC and Wernig M. 2011, Induced neuronal cells: how to
make and define a neuron, Cell Stem Cell 9(6):517-525
Yoon BS, Moon JH, Jun EK, Kim J, Maeng I, Kim JS, Lee JH, Baik CS, Kim A, Cho KS,
Lee HH, Whang KY and You S. 2010, Secretory profiles and wound healing effects
of human amniotic fluid-derived mesenchymal stem cells, Stem Cells Dev
19(6):887-902
Page 31 of 41Page 31 of 49B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIC
HIG
AN
on
06/1
3/13
For
pers
onal
use
onl
y. T
his
Just
-IN
man
uscr
ipt i
s th
e ac
cept
ed m
anus
crip
t pri
or to
cop
y ed
iting
and
pag
e co
mpo
sitio
n. I
t may
dif
fer
from
the
fina
l off
icia
l ver
sion
of
reco
rd.

Yu F and Morshead CM. 2011, Adult stem cells and bioengineering strategies for the
treatment of cerebral ischemic stroke, Curr Stem Cell Res Ther 6(3):190-207
Zhang L, Dong LY, Li YJ, Hong Z and Wei WS. 2012a, The microRNA miR-181c
controls microglia-mediated neuronal apoptosis by suppressing tumor necrosis
factor, J Neuroinflamm 9:211
Zhang L, Dong LY, Li YJ, Hong Z and Wei WS. 2012b, miR-21 represses FasL in
microglia and protects against microglia-mediated neuronal cell death following
hypoxia/ischemia, Glia 60(12):1888-1895
Zhang RL, Zhang ZG, Zhang L and Chopp M. 2001, Proliferation and differentiation of
progenitor cells in the cortex and the subventricular zone in the adult rat after focal
cerebral ischemia, Neuroscience 105(1):33-41
Zilles K. 1990 Anatomy of the neocortex: cytoarchitecture and myeloarchitecture., in The
Cerebral Cortex of the Rat (Kolb BT, R.C. ed) pp 77-113, MIT, Cambridge, MA
Page 32 of 41Page 32 of 49B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIC
HIG
AN
on
06/1
3/13
For
pers
onal
use
onl
y. T
his
Just
-IN
man
uscr
ipt i
s th
e ac
cept
ed m
anus
crip
t pri
or to
cop
y ed
iting
and
pag
e co
mpo
sitio
n. I
t may
dif
fer
from
the
fina
l off
icia
l ver
sion
of
reco
rd.

Acknowledgement:
The authors would like to thank Dr. Marianna Sikorska, Dr. Robert Monette and Tanya
Comas for discussions and technical assistance.
Page 33 of 41Page 33 of 49B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIC
HIG
AN
on
06/1
3/13
For
pers
onal
use
onl
y. T
his
Just
-IN
man
uscr
ipt i
s th
e ac
cept
ed m
anus
crip
t pri
or to
cop
y ed
iting
and
pag
e co
mpo
sitio
n. I
t may
dif
fer
from
the
fina
l off
icia
l ver
sion
of
reco
rd.

Table 1. Applications of AF cells from different sources in animal models of nervous
system injury or disease. The effects of AF cells administered via various routes on the
behavioural and histological outcomes associated with each model are described. Where
possible, an indication of whether the cells survived in vitro and expressed neuronal
markers is included.
Model Source of AF cells
Route of administration
Outcomes (behavioural and histological) AF cell survival and phenotype Reference
MCAO (rat) Human
Intra-striatal injection one week post MCAO
Not examined
Few GFAP+ AF cells in striatum 30 and 90 days post-injection. Very few DCX+ AF cells 10 days post-injection. No βIII-tubulin+ AF cells at any time point.
Cipriani et al. 2007
MCAO (mouse)
Mouse ICV three days post MCAO
Improved neurological severity score, cognitive score, and somatosensory score 4 days after cell transplantation. No assessment of extent of brain damage.
Not examined Rehni et al. 2007
MCAO (rat) Rat IV, 35 days after MCAO
Reduced infarct volume and improved cognitive and motor functions one month after transplantation.
Not examined Tajiri et al. 2012
Thoracic crush injury (E2.5 chick embryo)
Rat
Implanted in the spinal cord at the time of injury
Reduced hemorrhage and increased survival of chick embryos after injury.
Grafted cells were negative for neural markers DCX, GFAP and Tuj1.
Prasongchean et al. 2012
Sciatic nerve resection (rat)
Human
AF cells in Surgicel and fibrin glue implanted in the surgical site
Improved ankle angle when walking, improved compound muscle action potential and nerve conduction velocities 8 weeks after transplantation.
Not examined Pan et al. 2006
Sciatic nerve crush (rat)
Rat
AF cells in Surgicel and fibrin glue implanted in the surgical site
Improved sciatic function index, ankle angle, compound muscle action potential and nerve conduction latencies 1, 2 and 4 weeks after transplantation
AF cells observed around the crush site, but not penetrating into the nerve. Cells did not survive for more than a month.
Pan et al. 2007
Sciatic nerve crush (rat)
Human
AF cells in Matrigel or in Surgicel/fibrin glue implanted in the surgical site
Effects of AF cells on motor and electrophysiological recovery were augmented using GDNF-transduced AF cells, or co-treatment with hyperbaric oxygen, granulocyte-colony stimulating factor or fermented soybean extract
Cell phenotype not examined
Cheng et al. 2010; Pan et al. 2009a; 2009b; 2009c
6-OHDA Parkinson's disease model (rat)
Human Intrastriatal injection
Not examined
AF cells were only detected in the brain of 1 rat (out of 12). AF cells observed at 3 days, but not at 3 weeks after transplant, and expressed markers of neural progenitors and neurons, but not dopaminergic neurons.
Donaldson et al. 2009
Page 34 of 41Page 34 of 49B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIC
HIG
AN
on
06/1
3/13
For
pers
onal
use
onl
y. T
his
Just
-IN
man
uscr
ipt i
s th
e ac
cept
ed m
anus
crip
t pri
or to
cop
y ed
iting
and
pag
e co
mpo
sitio
n. I
t may
dif
fer
from
the
fina
l off
icia
l ver
sion
of
reco
rd.

Abbreviations: AF: amniotic fluid; ALS: amyotrophic lateral sclerosis; DA: dopamine; DCX: doublecortin; E:
embryonic day; GDNF: glial cell-derived neurotrophic factor; GFAP: glial fibrillary acidic protein; ICV:
intracerebroventricular; IV; intravenous; MCAO: middle cerebral artery occlusion; OB: olfactory bulb; 6-OHDA:
6-hydroxydopamine; TH: tyrosine hydroxylase.
6-OHDA Parkinson's disease model (rat)
Human
Injection into the medial forebrain bundle and ventral tegmental area
CD44+ AF cells reduced apomorphine-induced rotational behaviour
CD44+ AF cells survived for at least 3 weeks, and expressed TH
Liu et al., 2011
6-OHDA Parkinson's disease model (rat)
Human Instrastriatal injection
Improved bladder function and survival of host DA neurons 14 but not 28 days after AF cell injection.
AF cells survived for less than 28 days. No assessment of cell phenotype.
Soler, 2012
Genetic twitcher model of leukodystrophy (mouse)
Human ICV Not examined
Survival of grafted cells for at least 2 months, found throughout the brain, including hippocampus, OB and periventricular areas in both wt and twitcher mice. Cell phenotypes not examined.
De Coppi et al., 2007
Genetic wobbler model of ALS (mouse)
Human ICV No effect on mouse survival. AF cells remained in the ventricular system and did not migrate into parenchyma.
Bigini et al., 2012
Page 35 of 41Page 35 of 49B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIC
HIG
AN
on
06/1
3/13
For
pers
onal
use
onl
y. T
his
Just
-IN
man
uscr
ipt i
s th
e ac
cept
ed m
anus
crip
t pri
or to
cop
y ed
iting
and
pag
e co
mpo
sitio
n. I
t may
dif
fer
from
the
fina
l off
icia
l ver
sion
of
reco
rd.

Figure Legends
Figure 1: Time course of events occurring after brain injury. The approximate temporal
sequence of various neuropathological and reparative processes triggered by brain injury or stroke
is depicted on a timeline spanning from hours to weeks. Shading of the bars indicates the relative
intensity of the process over time, with the darkest colouring indicating the peak intensity of each
process. Necrotic cell death begins within minutes of the insult and is followed by the initiation of
apoptosis (programmed cell death), which can continue for weeks, expanding the zone of infarction.
This is accompanied by morphological and gene expression changes in both microglia (microglial
activation) and astrocytes (astrogliosis). Growth inhibitory genes are expressed in the tissue
surrounding the injury site, preventing axonal extension and neuroblast migration within the
damaged area. In response to the injury, the brain launches endogenous repair processes, including
an increase in the expression of growth-promoting genes, the formation of synapses
(synaptogenesis), and the production of new neurons (neurogenesis).
Figure 2: Mouse model of surgically induced cortical injury. To produce the cortical injury, the
part of the skull overlying the left motor cortex was removed. Injury to the motor cortex was
performed with a sterile needle to remove neural tissue to a depth of 1mm, using the
following coordinates: from AP -0.25mm to -1.0mm, Lat +0.7mm to AP +1.25mm to
+3.0mm, Lat +2.4mm relative to Bregma. Full details of the surgical procedure were
previously described (Jezierski et al., 2012). This method produces a consistent injury with
only slight variability in lesion placement across animals. The changes shown here have
been consistently observed across multiple studies. A: A 3D reconstruction of a lesioned mouse
Page 36 of 41Page 36 of 49B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIC
HIG
AN
on
06/1
3/13
For
pers
onal
use
onl
y. T
his
Just
-IN
man
uscr
ipt i
s th
e ac
cept
ed m
anus
crip
t pri
or to
cop
y ed
iting
and
pag
e co
mpo
sitio
n. I
t may
dif
fer
from
the
fina
l off
icia
l ver
sion
of
reco
rd.

brain showing the area of damage in the left primary motor cortex (orange). B: Cresyl violet
staining of a typical coronal section from a lesioned mouse brain showing the extent of damage in
primary motor cortex. C-D: Immunostaining for neurons (MAP2; red) and astrocytes (GFAP;
green) in sham (C) and lesioned (D) mouse motor cortex one week after surgery. Loss of neurons
and neurites as well as an intense astrocytic reaction in the peri-lesion cortex are evident in the
lesioned mouse, compared to sham. E-F: Immunostaining for microglia/macrophages (Iba-1; green)
in sham (E) and lesioned (F) cortex one week after surgery demonstrates the strong inflammatory
response that occurs after surgical lesion. G-H: TUNEL (green) staining at the border of the injury
site indicates that while very low levels of cell death (almost none) are seen in sham brains (G) cell
death continues to occur even 4 weeks after lesion (H). Blue staining in C-H is Hoechst. Scale bar:
2mm (B); 100µm (C-F); 50µm (G-H).
Figure 3: Complexity and diversity of cellular morphology and connectivity in mouse motor
cortex. Golgi-Cox staining of mouse motor cortex reveals the complexity and diversity of cellular
morphology and connectivity within the cortex. Saline-perfused mouse brains (n=4) were fixed in
Golgi-Cox solution (Glaser and van der Loos 1981) for 14 days, rinsed several times in distilled
water, and transferred to 30% sucrose. 200µm sections were cut on a vibratome and mounted on
gelatinized slides. The sections were developed by incubating in ammonium hydroxide followed by
Kodak film fix, and then were dehydrated, cleared and coverslipped. Complete details of the
procedure were previously described (Frechette et al. 2009). The images shown here are
representative images of a non-injured hemisphere taken from a single animal. A. A low
magnification view of motor cortex, showing diverse cell types throughout the cortex. Several
pyramidal neurons with long apical processes and extensively branched basal trees can be seen
(indicated with arrows), as well as structurally varied interneurons (arrowheads) .B. A pyramidal
neuron, demonstrating the highly branched morphology of the basal dendrites and long apical
Page 37 of 41Page 37 of 49B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIC
HIG
AN
on
06/1
3/13
For
pers
onal
use
onl
y. T
his
Just
-IN
man
uscr
ipt i
s th
e ac
cept
ed m
anus
crip
t pri
or to
cop
y ed
iting
and
pag
e co
mpo
sitio
n. I
t may
dif
fer
from
the
fina
l off
icia
l ver
sion
of
reco
rd.

process. C. A high magnification image of spines on the basal dendrites of a pyramidal neuron.
Scale bar: 200µm (A); 50µm (B); 3µm (C).
Figure 4. Proliferative potential and karyotypic stability of AF cells
A. Samples of amniotic fluid were obtained from the Ottawa Hospital (Ottawa, ON, Canada)
following routine amniocentesis carried out on pregnant women at 15 to 35 weeks gestation with
written consent and performed under the guidelines established by the Ottawa Hospital and
National Research Council Canada - Research Ethics Boards. In brief, AF was diluted with PBS
and spun at 250 x g for 5 mins; pellets were re-suspended in expansion media composed of DMEM
supplemented with 20% FBS. The cells were subsequently plated in a 10 cm dish, passaged by
trypsinization and serially expanded with a split ratio of 1:3 at 70% confluence (Jezierski et al.
2010). For the generation of single cell clones, a single cell suspension was prepared by gentle
trypsinization and individual AF cells were deposited, by Fluorescence Activated Cell Sorter
(FACS), with a cell density of one cell per well of a 96-well plate in 100ul expansion media.
Resulting clones (e.g. F5) were then sub-cultured first into 24-well plates and thereafter expanded
serially to 10cm plates and subsequently used for a number of cellular and molecular purposes. B.
Single clone (F5) cells were shown to preserve their karyotypic stability after multiple passages in
culture. Single clone F5 cells were grown in DME + 20% FBS until they reached about 70%
confluence. They were then incubated for 4h in medium containing 0.1µg/ml Karyomax Colcemid
solution (Invitrogen), harvested by trypsinization, and spun at 175xg for 5 minutes at room
temperature. Cells were then incubated for 20 minutes at room temperature in 10ml of hypotonic
solution (0.075M KCl) and fixed with fresh methanol/acetic acid fixative (3:1 vol/vol). The fixed
cell suspension was dropped onto a clean glass slide. Slides were incubated with Hoechst (1 µg/ml)
Page 38 of 41Page 38 of 49B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIC
HIG
AN
on
06/1
3/13
For
pers
onal
use
onl
y. T
his
Just
-IN
man
uscr
ipt i
s th
e ac
cept
ed m
anus
crip
t pri
or to
cop
y ed
iting
and
pag
e co
mpo
sitio
n. I
t may
dif
fer
from
the
fina
l off
icia
l ver
sion
of
reco
rd.

stain for ten minutes and mounted with Vectashield (Vector). Pictures of the metaphase spreads
were taken using a 40x objective (Axiovert Zeiss).
Figure 5. Expression of neural markers by AF cells
A. RT-PCR analysis of the mRNA harvested from AF single cell clone F5 cells shows the
expression of Nestin (Nes), Brn2 (pou3f2) and Neurofilament Light Chain (NFL). β-ACTIN (ACTB)
and undifferentiated human nTera2 (NT2) cells were used as internal and positive controls,
respectively. NTC, No Template Control. B. Western blotting confirms the expression of NESTIN
protein isoforms in F5 clone and the parent population, AF sample 1 (AF*) compared to another AF
Sample (AF**), suggesting that some AF samples harbour the potential to differentiate along the
neural lineage. β-ACTIN (ACTB) and undifferentiated human NT2 cells were used as internal and
positive controls, respectively, whereas HEK293 cells were used as a negative control. C-D.
Immunocytochemistry for NES expression in F5 cells. Hoechst was used as a counter stain. Scale
bar: 25 µm
Figure 6. GDNF-secreting AF cells
A. AF cells were infected with Tet07-CMV-GDNF-DsRed vector to constitutively secrete GDNF.
These GDNF-secreting cells can be seeded on biodegradable, biocompatible polymer scaffolds
(Bani-Yaghoub et al. 2008) or injected directly into tissue of interest to deliver GDNF. B. AF cells
infected with GDNF-DsRed and DsRed backbone control in vitro. Scale bar: 50µm.
Figure 7. Transfer of morpholino mimic of miR-21 from AF cells to neighbouring AF and
cortical astrocytes in vitro
Cell-to-cell transfer of a fluorescently labeled morpholino mimic of miR 21 (GeneTools) between
adjacent AF cells was assessed using a whole-cell patch-clamp technique. Cells were cultured on
Page 39 of 41Page 39 of 49B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIC
HIG
AN
on
06/1
3/13
For
pers
onal
use
onl
y. T
his
Just
-IN
man
uscr
ipt i
s th
e ac
cept
ed m
anus
crip
t pri
or to
cop
y ed
iting
and
pag
e co
mpo
sitio
n. I
t may
dif
fer
from
the
fina
l off
icia
l ver
sion
of
reco
rd.

coverslips and bathed in a solution containing 135mM NaCl, 5mM KCl, 2mM MgCl2, 2mM CaCl2,
10mM HEPES and 10mM glucose (pH 7.4). Whole-cell patch-clamp experiments were performed
at room temperature using borosilicate pipettes (resistance 3-5MΩ) filled with a solution containing
130mM K+-gluconate, 10mM KCl, 2mM MgCl2, 10mM HEPES, 0.5mM EGTA, 4mM ATP,
0.2mM GTP (pH7.4). Morpholino oligos were added to the patch pipette solution to concentrations
of 0.25-0.5mM. Whole-cell patch-clamp recordings were monitored using an Axopatch 1D
amplifier (Axon Instrument Inc) under visual control using differential interference contrast and
infrared video microscopy (IR-DIC; Olympus), Whole-cell access resistance ranged from 5 to 15
MΩ and was monitored throughout the experiments. The cell-to-cell transfer of morpholino oligos
was recorded using a Fluo-View FV1000 Olympus confocal microscope (Olympus). Images were
acquired every minute using a 40X water immersion objective (NA 0.8), laser excitation
wavelength 559nm and BP wavelength 570-640nm. A. The morpholino was injected into a single
AF cell (asterisk; image acquired 1 minute post-injection) and B. The cell-to-cell morpholino
transfer was recorded in real-time (Supplementary Video 1) between adjacent AF cells (arrows;
image acquired 40 minutes post-injection). C. Phase contrast showing patch pipette microinjection
into a single AF cell. D. CX43 expression (arrowhead) at the cell-cell boundary between the patch-
clamped AF cell and adjacent morpholino-receiving cells. CX43 expression was determined by
immunocytochemical staining of the cells, which were fixed after completion of the morpholino
transfer experiment. E-G. Similarly, miR-21 morpholino transfer from patch-clamped AF cell
(asterisk) to mouse cortical astrocytes (arrows) was recorded between minute 1 (E) and 40 (F) post-
injection. As seen in the video (supplementary material), transfer of the morpholino was observed in
the receiving cell as early as 5 minutes post-injection. Scale bar: 25 µm
Page 40 of 41Page 40 of 49B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIC
HIG
AN
on
06/1
3/13
For
pers
onal
use
onl
y. T
his
Just
-IN
man
uscr
ipt i
s th
e ac
cept
ed m
anus
crip
t pri
or to
cop
y ed
iting
and
pag
e co
mpo
sitio
n. I
t may
dif
fer
from
the
fina
l off
icia
l ver
sion
of
reco
rd.

Supplementary Figure 1. miR expressing AF cells were engineered by retroviral infection. EGFP
was used as reporter gene in the same vector [miR-132F1R1/pMX clone 22; original vector pMIR-
REPORT Luciferase (Applied Biosystems)]. A. miR-132 EGFP expression, B. phase, C. Merged,
Scale bar: 150 µm
Page 41 of 41Page 41 of 49B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIC
HIG
AN
on
06/1
3/13
For
pers
onal
use
onl
y. T
his
Just
-IN
man
uscr
ipt i
s th
e ac
cept
ed m
anus
crip
t pri
or to
cop
y ed
iting
and
pag
e co
mpo
sitio
n. I
t may
dif
fer
from
the
fina
l off
icia
l ver
sion
of
reco
rd.

Fig
ure
1
Page 42 of 49B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIC
HIG
AN
on
06/1
3/13
For
pers
onal
use
onl
y. T
his
Just
-IN
man
uscr
ipt i
s th
e ac
cept
ed m
anus
crip
t pri
or to
cop
y ed
iting
and
pag
e co
mpo
sitio
n. I
t may
dif
fer
from
the
fina
l off
icia
l ver
sion
of
reco
rd.

Fig
ure
2
MA
P2
//
GFA
PH
oe
ch
st
Iba
-1/H
oe
ch
st
TU
NE
L/H
oe
ch
st
A B
C D
E F
G H
Page 43 of 49B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIC
HIG
AN
on
06/1
3/13
For
pers
onal
use
onl
y. T
his
Just
-IN
man
uscr
ipt i
s th
e ac
cept
ed m
anus
crip
t pri
or to
cop
y ed
iting
and
pag
e co
mpo
sitio
n. I
t may
dif
fer
from
the
fina
l off
icia
l ver
sion
of
reco
rd.

Fig
ure
3
A
B C
Page 44 of 49B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIC
HIG
AN
on
06/1
3/13
For
pers
onal
use
onl
y. T
his
Just
-IN
man
uscr
ipt i
s th
e ac
cept
ed m
anus
crip
t pri
or to
cop
y ed
iting
and
pag
e co
mpo
sitio
n. I
t may
dif
fer
from
the
fina
l off
icia
l ver
sion
of
reco
rd.

A
B
C
D
E
F
G
H
21 3 4 5 6 7 8 9 10 11 12
96 Well Plate
24 Well Plate
12 Well Plate
6 Well Plate
10 mm Plate
Amniocentesis
10 mm Plate
Passage 1
Single CellClone AF1*(F5)
Passage 6
37-10x10
45x10
51x10
52x10
61.75x10
OutputApproximate CellNumber at 70%
Confluency
FreezeCells
SampleCollection
andAnalysis
At Least82-4x10
at Passage 9
MorePassages
A
B
iPSCells
Delivery ofBeneficialFactors
Fig
ure
4Page 45 of 49
Bio
chem
. Cel
l Bio
l. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y U
NIV
ER
SIT
Y O
F M
ICH
IGA
N o
n 06
/13/
13Fo
r pe
rson
al u
se o
nly.
Thi
s Ju
st-I
N m
anus
crip
t is
the
acce
pted
man
uscr
ipt p
rior
to c
opy
editi
ng a
nd p
age
com
posi
tion.
It m
ay d
iffe
r fr
om th
e fi
nal o
ffic
ial v
ersi
on o
f re
cord
.
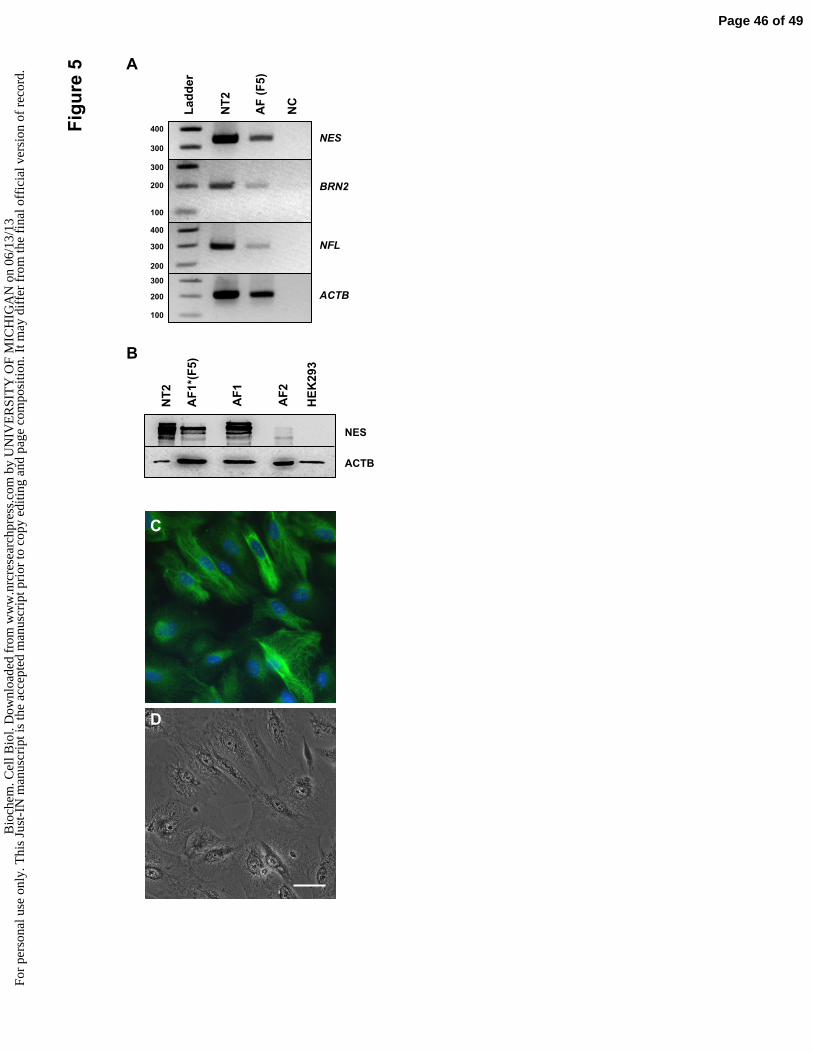
NES
BRN2
NFL
ACTB
300
400
200
300
100
200
400
300
200
300
100
La
dd
er
NT
2
AF
(F
5)
NC
A
C
D
NES
ACTB
NT
2
AF
1*(
F5
)
1
HE
K29
3
B
AF
2A
F
Fig
ure
5
Page 46 of 49B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIC
HIG
AN
on
06/1
3/13
For
pers
onal
use
onl
y. T
his
Just
-IN
man
uscr
ipt i
s th
e ac
cept
ed m
anus
crip
t pri
or to
cop
y ed
iting
and
pag
e co
mpo
sitio
n. I
t may
dif
fer
from
the
fina
l off
icia
l ver
sion
of
reco
rd.

AF Cells
GDNF LentivectorGDNF-secreting AF Cells
Transcription
CMV GDNF DsRed
AG
DN
F-D
sR
ed
DsR
ed
Scaffolds
Injection
Fig
ure
6
B DC
E GF
Page 47 of 49B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIC
HIG
AN
on
06/1
3/13
For
pers
onal
use
onl
y. T
his
Just
-IN
man
uscr
ipt i
s th
e ac
cept
ed m
anus
crip
t pri
or to
cop
y ed
iting
and
pag
e co
mpo
sitio
n. I
t may
dif
fer
from
the
fina
l off
icia
l ver
sion
of
reco
rd.

A B
C D
**
E
F
*
G
*
*
Fig
ure
7
Page 48 of 49B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIC
HIG
AN
on
06/1
3/13
For
pers
onal
use
onl
y. T
his
Just
-IN
man
uscr
ipt i
s th
e ac
cept
ed m
anus
crip
t pri
or to
cop
y ed
iting
and
pag
e co
mpo
sitio
n. I
t may
dif
fer
from
the
fina
l off
icia
l ver
sion
of
reco
rd.

Su
pp
lem
en
tary
Fig
ure
1 A
C
B
Page 49 of 49B
ioch
em. C
ell B
iol.
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
UN
IVE
RSI
TY
OF
MIC
HIG
AN
on
06/1
3/13
For
pers
onal
use
onl
y. T
his
Just
-IN
man
uscr
ipt i
s th
e ac
cept
ed m
anus
crip
t pri
or to
cop
y ed
iting
and
pag
e co
mpo
sitio
n. I
t may
dif
fer
from
the
fina
l off
icia
l ver
sion
of
reco
rd.


















