
Therapeutic efficacy of antiviral compounds in the ...
141
Graduate eses and Dissertations Iowa State University Capstones, eses and Dissertations 2019 erapeutic efficacy of antiviral compounds in the neonatal lamb model of respiratory syncytial virus infection of infants with pathogenesis of secondary Streptococcus pneumoniae infection Sarhad Alnajjar Iowa State University Follow this and additional works at: hps://lib.dr.iastate.edu/etd Part of the Pathology Commons , Pharmacology Commons , and the Virology Commons is Dissertation is brought to you for free and open access by the Iowa State University Capstones, eses and Dissertations at Iowa State University Digital Repository. It has been accepted for inclusion in Graduate eses and Dissertations by an authorized administrator of Iowa State University Digital Repository. For more information, please contact [email protected]. Recommended Citation Alnajjar, Sarhad, "erapeutic efficacy of antiviral compounds in the neonatal lamb model of respiratory syncytial virus infection of infants with pathogenesis of secondary Streptococcus pneumoniae infection" (2019). Graduate eses and Dissertations. 16959. hps://lib.dr.iastate.edu/etd/16959
Transcript of Therapeutic efficacy of antiviral compounds in the ...

Graduate Theses and Dissertations Iowa State University Capstones, Theses andDissertations
2019
Therapeutic efficacy of antiviral compounds in theneonatal lamb model of respiratory syncytial virusinfection of infants with pathogenesis of secondaryStreptococcus pneumoniae infectionSarhad AlnajjarIowa State University
Follow this and additional works at: https://lib.dr.iastate.edu/etd
Part of the Pathology Commons, Pharmacology Commons, and the Virology Commons
This Dissertation is brought to you for free and open access by the Iowa State University Capstones, Theses and Dissertations at Iowa State UniversityDigital Repository. It has been accepted for inclusion in Graduate Theses and Dissertations by an authorized administrator of Iowa State UniversityDigital Repository. For more information, please contact [email protected].
Recommended CitationAlnajjar, Sarhad, "Therapeutic efficacy of antiviral compounds in the neonatal lamb model of respiratory syncytial virus infection ofinfants with pathogenesis of secondary Streptococcus pneumoniae infection" (2019). Graduate Theses and Dissertations. 16959.https://lib.dr.iastate.edu/etd/16959

i
Therapeutic efficacy of antiviral compounds in the neonatal lamb model
of respiratory syncytial virus infection of infants with pathogenesis of
secondary Streptococcus pneumoniae infection
by
Sarhad S.A. Alnajjar
A dissertation submitted to the graduate faculty
in partial fulfillment of the requirements for the degree of
DOCTOR OF PHILOSOPHY
Major: Veterinary Pathology
Program of Study Committee:
Mark R. Ackermann, Co-major Professor
Jodi D. Smith, Co-major Professor
Jesse M Hostetter
Brett A Sponseller
Mitchell Van Palmer
The student author, whose presentation of the scholarship herein was approved by the
program of study committee, is solely responsible for the content of this dissertation. The
Graduate College will ensure this dissertation is globally accessible and will not permit
alterations after a degree is conferred.
Iowa State University
Ames, Iowa
2019
Copyright © Sarhad S.A. Alnajjar, 2019. All rights reserved.

ii
DEDICATION
This dissertation is dedicated to:
My wife
My family

iii
TABLE OF CONTENTS
LIST OF FIGURES ......................................................................................................... vii
LIST OF TABLES .......................................................................................................... viii
ACKNOWLEDGMENTS ................................................................................................ ix
ABSTRACT ...................................................................................................................... x
CHAPTER 1: GENERAL INTRODUCTION ...................................................................... 1
Statement of the Problem .................................................................................................. 1
Specific Aims .................................................................................................................... 2
Dissertation Organization .................................................................................................. 2
Literature Review .............................................................................................................. 3
Respiratory Syncytial Virus Structure and Strains ........................................................ 3
Disease Burden .............................................................................................................. 5
Animal Models of RSV Infection .................................................................................. 8
Immune Response to RSV Infection: .......................................................................... 11
Secondary Bacterial Infection ...................................................................................... 16
RSV Therapeutics ........................................................................................................ 18
References .................................................................................................................... 20
: STREPTOCOCCUS PNEUMONIAE INFECTION IN RESPIRATORY
SYNCYTIAL VIRUS INFECTED NEONATAL LAMBS ............................................... 35
Abstract ............................................................................................................................ 35
Introduction ..................................................................................................................... 36
Material and Methods ...................................................................................................... 38
Experimental Design .................................................................................................... 38
Infectious Agents ......................................................................................................... 39
Immunohistochemistry (IHC) ...................................................................................... 40
Quantitative Reverse Transcription Polymerase Chain Reaction (RT-qPCR) ............ 40
Hematoxylin-Eosin Staining and Histological Scoring of Lung Sections ................... 41
Statistical Analysis ....................................................................................................... 41
Results ............................................................................................................................. 42
Page

iv
Infected Lambs Showed Marked RSV and Spn Titer................................................... 42
RSV and Spn Induce a Well-recognized Macroscopic and Microscopic Lesion ......... 42
Discussion ........................................................................................................................ 44
References ........................................................................................................................ 48
Figures and Legends......................................................................................................... 52
: THERAPEUTIC EFFICACY OF JNJ-49214698, AN RSV FUSION
INHIBITOR, IN RSV-INFECTED NEONATAL LAMBS ................................................ 56
Authors Contributions .................................................................................................. 56
Acknowledgments ............................................................................................................ 57
Conflict of Interests ...................................................................................................... 57
Funding ......................................................................................................................... 57
Abstract ............................................................................................................................ 58
Key Words .................................................................................................................... 58
Introduction ...................................................................................................................... 58
Materials and Methods ..................................................................................................... 61
Compound and Dosing ................................................................................................. 61
Animals ......................................................................................................................... 61
Experimental Design .................................................................................................... 62
RSV Infection ............................................................................................................... 62
Assessed Parameters ..................................................................................................... 63
Statistical Analysis ....................................................................................................... 64
Results .............................................................................................................................. 64
JNJ-49214698 Efficiently Distributes to Different Lung Compartments of
Neonatal Lambs ............................................................................................................ 64
Treatment with JNJ-49214698 Reduces Incidence and Duration of
RSV-Associated Symptoms ......................................................................................... 65
Treatment with JNJ-49214698 Inhibits a Multicyclic RSV Infection in Neonatal
Lambs ........................................................................................................................... 66
Treatment with JNJ-49214698 Inhibits RSV-Induced Lung Pathology and Cellular
Immune Response of Neonatal Lambs ......................................................................... 67
Discussion ........................................................................................................................ 69
References ........................................................................................................................ 71
Figures and Legends......................................................................................................... 77
Page

v
Supplementary Data ........................................................................................................ 83
Materials and Methods ................................................................................................. 83
Compound and Dosing ................................................................................................ 83
Animals ........................................................................................................................ 83
Experimental Design .................................................................................................... 84
RSV Infection .............................................................................................................. 84
Animal Monitoring for Appearance of Clinical Signs and the Clinical Score ............ 85
Blood sampling for PK Analysis ................................................................................. 85
Lung Collection and Processing .................................................................................. 85
BALF Collection .......................................................................................................... 86
Quantification of JNJ-49214698 Exposure .................................................................. 87
Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR) ............ 88
Hematoxylin-Eosin Staining and Histological Scoring of Lung Sections ................... 89
Immunohistochemistry (IHC) of Lung Sections .......................................................... 90
RNAscope .................................................................................................................... 91
Infectious Focus-Forming Unit (IFFU) Assay ............................................................. 92
Statistical Analysis ....................................................................................................... 93
References ....................................................................................................................... 97
:THERAPEUTIC EFFICACY OF RESPIRATORY SYNCYTIAL
VIRUS NON-FUSION INHIBITOR (RSV-NFI) IN NEONATAL LAMBS
INFECTED WITH A HUMAN STRAIN OF RSV ............................................................ 99
Summary .......................................................................................................................... 99
Introduction ................................................................................................................... 100
Materials and Methods .................................................................................................. 101
Experimental Design .................................................................................................. 101
Tissue and Sample Collection .................................................................................... 102
Infectious Focus-Forming Unit (IFFU) Assay ........................................................... 102
Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR) .......... 103
Histologic Evaluation ................................................................................................. 103
Statistics ..................................................................................................................... 105
Results ........................................................................................................................... 105
Pharmacokinetics of Different Doses of RSV-NFI in the Neonatal Lambs .............. 105
Page

vi
Viral Load in BALF and Lavaged-Lung Samples of Neonatal Lambs ...................... 106
Treatment with RSV-NFI Significantly Reduced RSV-Induced Lung Lesion .......... 107
Treatment with RSV-NFI Significantly Decrease Inflammatory Chemokines .......... 108
Discussion ...................................................................................................................... 109
References ...................................................................................................................... 111
Figures and Legends....................................................................................................... 116
CHAPTER 5: GENERAL CONCLUSION ....................................................................... 125
Secondary Bacterial Co-Infection .................................................................................. 125
Efficacy of Anti-RSV Fusion Inhibitor .......................................................................... 125
Efficacy of Anti-RSV Non-Fusion Inhibitor .................................................................. 126
Future Directions ............................................................................................................ 127
References ...................................................................................................................... 128
Page

vii
LIST OF FIGURES
Figure 2-1:RSV and Spn titer in lung tissue and blood ....................................................... 52
Figure 2-2:Percent of lung tissue associated with RSV and/or Spn infection. ................... 53
Figure 2-3:Histologic lesions associated with RSV, Spn, and RSV-Spn
combined infection. ...................................................................................................... 54
Figure 2-4:Immunohistochemistry staining of RSV and Spn in FFPE lung tissue
sections. ........................................................................................................................ 55
Figure 3-1:Exposure of JNJ-49214698 in different body compartments of
neonatal lambs. ............................................................................................................ 77
Figure 3-2: Clinical signs score in different groups. ........................................................... 78
Figure 3-3:Effect of JNJ-49214698 on viral titers in BALF and lung tissue at
Day 6 p.i. ...................................................................................................................... 79
Figure 3-4: Effect of JNJ-49214698 on viral antigen and RNA in lung tissue at
Day 6 p.i. ...................................................................................................................... 80
Figure 3-5: Effect of JNJ-49214698 on development of gross lung lesions at
Day 6 p.i. ...................................................................................................................... 81
Figure 3-6:Effect of JNJ-49214698 on RSV-induced accumulated lung histopathology at
Day 6 p.i. ...................................................................................................................... 82
Figure 3-7: Effect of JNJ-49214698 on RSV-induced lung histopathology at
Day 6 p.i. ...................................................................................................................... 95
Figure 3-8:Effect of JNJ-49214698 on RSV-induced lung histopathology at
Day 6 p.i. ...................................................................................................................... 96
Figure 4-1:Experimental design. ....................................................................................... 116
Figure 4-2: Exposure of RSV-NFI in different body compartments. ............................... 117
Figure 4-3: Effect of RSV-NFI on viral titer in BALF and lung tissue
homogenates at day 6 p.i. ........................................................................................... 118
Figure 4-4: Effect of RSV-NFI on the development of RSV associated gross lesion. ..... 119
Figure 4-5: Effect of RSV-NFI on the development of RSV associated histologic
lesion. ......................................................................................................................... 120
Figure 4-6:Effect of RSV-NFI on RSV antigen. ............................................................... 121
Figure 4-7: Effect of RSV-NFI on RSV M37 RNA expression. ...................................... 122
Figure 4-8: Effect of RSV-NFI on the development of RSV associated histologic
lesion. ......................................................................................................................... 123
Figure 4-9: Effect of RSV-NFI inflammatory chemokine RNA expression in lung
tissue. ......................................................................................................................... 124
Page

viii
LIST OF TABLES
Table 1: Clinical signs observed in lambs in different groups throughout the study
period……………………………………………………………………………………94
Page

ix
ACKNOWLEDGMENTS
First of all I would like to thank God, the Most Gracious, the Most Merciful for helping
me throughout my life.
I would like to thank the Higher Committee for Education Development (HCED) in
Iraq for their support and funding of my study.
I would like to thank my major professor Dr. Mark Ackermann for his unwavering
guidance and support throughout my study program. Mark, your efforts are greatly
appreciated.
I would like to thank Dr. Jodi Smith, Dr. Jesse Hostetter, Dr. Brett Sponseller, and Dr.
Mitchell Palmer for serving in my program of study committee and for their constructive
comments and suggestions.
A special thank you to Jack Gallup. Thank you for sharing your expertise and help.
Thank you to Dr. Albert Van Geelen for your advice and mentoring in the laboratory. A
great thank you to our collaborators Drs. Dirk Roymans and Peter Rigaux for sharing their
scientific expertise and help in constructing, editing the scientific manuscripts. Great thanks
to Dr. David Verhoeven for his thoughts, ideas, and his constructive feedback.
A big thank you to my colleagues in the Veterinary Pathology, Immunobiology
programs for making my life enjoyable and for providing their help when asked especially
Dr. Panchan Sitthicharoenchai my friend, and lab mate, and also Dr. Saleh Albarrak thank
you for sharing your ideas, thoughts, and time.
A big thank you tothe Veterinary Pathology faculty and staff, who did not hesitate to
help and provide their expertise. Also, thank you for the laboratory animal veterinarian and
animal caretakers from Iowa State University Laboratory Animal Resources for their help
in ensuring the welfare of the animals and success of the research.
Dr. Mary Sauer, thank you for all your thoughts, time, and advice. My life would not
be this way without you. Debbie and Kurtis Younkin, what a great help you are. I cannot
find words that fulfill what you have done for my family and me during my study.
Finally, I would like to thank the wonderful Ames community, international family
(Stonebrook Church). Thank you for making us feel that we are home.

x
ABSTRACT
Respiratory Syncytial Virus (RSV)is one of the most common causes of acute lower
respiratory tract infection in humans that can cause severe infections in infants and the
elderly. It is estimated that annually 33 million lower respiratory tract infections in children
were associated with RSV worldwide. However, in the US alone there were 75000-125,000
hospitalizations due to RSV in infants per year. Although most RSV infections of the lower
respiratory tract are caused by RSV alone, occasionally, secondary bacterial infection further
increases lung damage and disease severity. Despite the widespread infections by RSV
throughout the world, till now there are no approved vaccines or therapeutic compounds to
treat RSV infection other than Ribavirin and prophylactic administration of Palivizumab.
There are several animal models of RSV infection, but neonatal lambs infected with RSV
have several advantages for modeling RSV infection in infants such as similarity in
pulmonary architecture, immune response, and respiratory tract size, viral replication, and
lesion development.
To further extend the neonatal lamb model to evaluate secondary bacterial infection
associated with RSV, lambs coinfected with RSV and Streptococcus pneumoniae were used
to determine feasibility, susceptibility and disease development. Lambs developed severe
disease when coinfected with both microorganisms with more severe suppurative bronchitis
and pneumonia. Streptococcus pneumonia infection enhanced the severity of RSV in lambs
when lambs were coinfected with both microorganisms.
To evaluate the efficacy of some antiviral compounds, two antiviral compounds
efficacy were tested in the neonatal lamb model of RSV infection. First, three regimens of

xi
JNJ-49214698, a small molecule RSV fusion protein inhibitor were tested (prophylactic,
early treatment, late treatment). JNJ-49214698 prevented RSV infection when given before
infection and reduced RSV induced lung lesion when used to treated established infection.
Additionally, late treatment at day 3 post RSV infection had a wide window for RSV
treatment. Secondly, 3 doses of RSV-NFI, an RSV non-fusion inhibitor were tested for
efficacy. RSV-NFI was well tolerated and reduced RSV induced lung lesions and viral titer.

1
CHAPTER 1: GENERAL INTRODUCTION
Statement of the Problem
Respiratory Syncytial Virus (RSV) is one of the leading cause of acute lower respiratory tract
infection in infants. Each year there are about 33 million lower respiratory tract infection cases in
children under the age of five around the world and these often lead to significant hospital
admission and mortality[1]. Additionally, RSV is one of the leading cause of pneumonia in older
adults and high-risk (e.g., immunosuppressed) individuals [2,3]. RSV infections can occur year
round but peaks in the winter months [4,5]. RSV has similar hospitalization rate and disease
burden to influenza in elderly[3]. Secondary to RSV infection, about 40% of RSV infected children
develop co-infections with bacteria or at increased risk of secondary bacterial pneumonia [6].
Furthermore, RSV infection in the early childhood may have delayed sequelae such as airways
hyperreactivity, wheezing, and asthma in later childhood [7,8]. There is no vaccine or antiviral
agent available clinically against RSV except for prophylactic humanized monoclonal antibody
Palivizumab which has questionable efficacy and is expensive and Ribavirin [9,10].
There are many investigators developing new vaccines and therapeutic compounds for RSV
and many of these are in advanced human clinical trials. Some newly developed vaccines under
investigation provide an acceptable level of protection against RSV [11,12]. However, formalin-
inactivated vaccines developed in the 1960’s led to enhanced disease and even death upon RSV
infection in infants. Therefore, there are fears of adverse side effects of any new RSV vaccination
regimen [13]. Newly developed anti-RSV therapeutic compounds that have reached advanced
clinical trials include those targeting several critical viral components such as RSV fusion protein,
N protein, L protein, SH protein, and M2-1 protein, and are from four chemical classes including:

2
anti-RSV immunoglobulins, siRNA-interference, fusion inhibitors, and small molecule inhibitors
[14,15]. These therapeutic strategies require animal models to evaluate their efficacy.
Creating animal models for RSV investigations is challenging. There are several animal
models used to evaluate RSV infection, pathogenesis, and therapeutic efficacy and these include
but are not limited to: chimpanzees, baboons, sheep, cotton rats, mice, ferrets, and cattle [16]. Of
these, lambs have several features advantageous as a model of RSV infection of infants including
susceptibility to the human strains of RSV, lung development, structure and cellular morphology,
and lesion development and lesion composition [17,18].
Specific Aims
The goals of the studies done in this dissertation are to determine the extent to which RSV
infection can alter the susceptibility of the lung to Streptococcus pneumoniae (Spn), and test the
efficacy of novel anti-RSV compounds. The hypothesis is that RSV infection enhances Spn
infection and/or vice versa by lung damage induced by either pathogen and that anti-RSV (fusion
and non-fusion inhibitors) can reduce RSV infection and disease severity. This hypothesis was
tested by: 1) a study in which lambs were infected with Spn 3 day after initial RSV infection
(Chapter 2). 2) a study to test the efficacy of anti-RSV small molecule fusion protein inhibitor
(Chapter 3). 3) a study to test the efficacy of anti-RSV non-fusion inhibitor (Chapter 4).
Dissertation Organization
This dissertation describes the pathogenesis of secondary bacterial pneumonia in the neonatal
lamb model of RSV infection and two therapeutic approaches to RSV infection. The dissertation
composed of five chapters with the 1st one as general introduction and literature review. Chapters

3
2, 3, and 4 are composed of the three individual manuscript prepared for the peer review journal
submission, with the final chapter (Chapter 5) as the conclusion and future direction.
The first paper Spn infection in RSV infected neonatal lambs (Chapter 2) were submitted to
the journal Emerging Microbes and Infection. The second paper, Therapeutic efficacy of JNJ-
49214698, an RSV fusion inhibitor, in RSV-infected neonatal lambs, were partially published in
the journal Nature Communications, and the full manuscript was submitted to Frontiers in
Microbiology. The third manuscript, therapeutic efficacy of RSV-NFI in RSV infected lambs will
be submitted to a journal publishes work on antiviral compounds at a later time.
Literature Review
Respiratory Syncytial Virus Structure and Strains
RSV is an enveloped virus with non-segmented negative-sense single-stranded RNA within
the family of Pneumoviridae, genus Orthopneumovirus, [19,20]. The viral RNA is composed of
10 genes that encode 11 protein two of them non-structural protein. In general, Orthopneumovirus
have three externally protruded glycoproteins, the attachment (G), fusion (F), and small
hydrophobic (SH). These glycoproteins pin the outer lipid envelope obtained from the host cell
plasma membrane and attached to the underlining matrix (M) protein. The G protein has carboxy-
terminal located to the outside of the virion and is variable among RSV strains. The G protein
mediates viral attachment to cellular membrane. The F protein has amino-terminus oriented to the
outside of the viral envelope and is less variable between strains than the G protein and responsible
for the viral fusion and entry into the target cell. These two proteins are critical for the viral
replication and infection, although virions lacking G protein can penetrate and replicate in cell
culture, but less efficiently than virion having both G and F protein[21]. The G protein is heavily
glycosylated and attaches to glycosaminoglycans on the host cell surface[22]. Most neutralizing

4
antibodies are directed against G protein. Therefore, RSV has an immune evasion strategy by
which a secreted form of G protein (sG) is produced in order to avoid inhibition by neutralizing
antibody [23–25]. On the other hand, F protein is highly conserved, and upon attachment and
activation, F protein undergoes conformational changes leading to fusion of the virus to the target
cell membrane, which subsequently induce internalization of the virus into the cell cytoplasm [26–
28]. Interestingly, one study demonstrated that RSV internalized intact into the epithelial cell by
macropinocytosis initiated by the RSV attachment. This process leads to RSV virion to be present
within intracytoplasmic fluid-filled macropinocytosome, then followed by F protein cleavage and
viral fusion that leads to internalization of RSV to the cytoplasm and infection [29]. SH protein
function and location is not fully characterized [30,31]. However, SH protein was demonstrated
to have an antiapoptotic activity by inhibiting TNF-α signaling [32]. Another study demonstrated
that SH protein is a small hydrophobic protein (∼100 amino acid) called Viroporin, which enhance
infected cell permeability by forming a hydrophilic pore in the infected cell membrane [33]. The
non-segmented single-stranded RNA associated with five structural proteins, the first 3 are
nucleoprotein (N), large (L) protein, and phosphoprotein (P) to form the helical nucleocapsid. The
L protein considered the RNA dependent RNA polymerase used in viral genome replication.
Additionally, L protein responsible for the transcription of the positive sense viral mRNA, and
possess capping enzyme activity at the 5’end and polyadenylation at the 3’ end of the viral mRNA
[34]. N Protein forms a complex with the RSV RNA called ribonucleoprotein complex that acts
as a template for the L protein, while P protein serves as a link between L and N protein to facilitate
efficient and specific recognition of ribonucleoprotein complex by L protein [35]. The other two
proteins associated with the nucleocapsid are the M2-1 and M2-2 proteins which are transcriptional

5
enhancer proteins [36]. These proteins bind to the RNA and P protein to prevent premature
termination of transcription [37,38].
There are two RSV strains: an A and a B strain which differ substantially in the G protein
and noncoding portion of the genome but have less variability in the other structural
proteins[39,40]. These two RSV strains co-circulate clinically throughout the year with the
dominance of A strain. Furthermore, RSV group A causes more severe infection than group B
RSV [41–44].
Disease Burden
RSV is a common respiratory disease affecting all ages and especially children worldwide
[1,45,46]. In most infected individuals, RSV causes mild to moderate upper respiratory tract
infection characterized by fever, nasal congestion, cough and rhinorrhea that persists for several
days [47]. However, RSV can lead to severe acute lower respiratory tract infection (LRTI) in
infants, elderly and immunocompromised individuals [48–50].
All infants eventually become infected by RSV, and it is estimated that there were 33.8
million cases of RSV associated pneumonia worldwide in children under the age of five in 2005
[1]. Furthermore, about 3.4 million RSV associated pneumonia cases required hospital admission
with 66,000-199,000 resulted in mortality. Most cases of mortality occur in developing countries
[1]. Based on US National viral surveillance data from 1990-1999, RSV associated deaths in
infants was nine times the number of deaths related to influenza infection in the United States and
RSV was the second leading viral cause of death after influenza in children age 1-5 years and older
adults [51]. While the rate of RSV associated hospitalization is 30 per 1000 in the US, Japan
reported 60 hospital admissions per 1000 [47]. RSV seasonality is consistent throughout Europe
which accounts for 42-45% hospital admission due do LRTI [47]. In Belgium alone and during

6
2000 season, RSV was associated with 63% of acute LRTI in children under the age of 5 [52].
Another study in England and Wales concluded that RSV was associated with 60-80% mortality
more than that of influenza in a total of 15 winters from January 1975 to December 1990 [53]. In
Australia, a retrospective study of 3 RSV seasons in 1997-1999, concluded that RSV is a
significant cause of morbidity and low mortality with 11.4% of infants required admission to
intensive care unit [54]. In general, temperate reigns experience RSV year round with a peak in
winter months, while a specific RSV seasons were seen in tropical regions [55]. RSV considered
an important nosocomial agent since RSV aerosolized particles persist for an extended time in the
urgent care clinic and can be inhaled and infect other patients [56]. However, several risk factors
determine the severity of RSV infection in infants.
RSV infection has considerable morbidity and mortality in infants less than 3 months and
those having one or more risk factors [57]. There are several risk factors associated with the host
include not limited to premature birth, congenital heart diseases, and chronic lung diseases [58,59].
However, a high proportion of infants hospitalized due to RSV are healthy without any
involvement of any of these risk factors [60]. Other risk factors related to the host or the
environment are gender (males), young age (less than 6 months), number of siblings, daycare
attendance, and exposure to tobacco smoke[61–64]. Other risk factors that can contribute to the
severity of RSV infection are related to the virus itself. There is a significant association between
viral load the severity of infection [65–67]. In addition to the viral load, RSV type A is associated
with more severe respiratory illness in compare to type B RSV [41,68,69]. Considering all the
mentioned risk factors, RSV disease severity is an association of all these risk factors, and despite
the massive burden of RSV disease, there are no fully satisfactory treatment or vaccine strategies
available for RSV infection.

7
Only two therapeutic agents, Ribavirin and Palivizumab, were approved against RSV
infection but both have limitations in efficacy. Ribavirin, a nucleoside inhibitor, developed in 1972
as a virostatic agent for both RNA and DNA viruses[70]. Even though Ribavirin had a positive
impact in high-risk patients such as transplant and immune compromised patients [71,72], only
marginal clinical benefits were seen in RSV lower respiratory tract infection in general patients.
Also, Ribavirin has a degree of toxicity. Thus, Ribavirin is no longer recommended as anti-RSV
treatment[73–75]. Palivizumab is a humanized monoclonal antibody against RSV F protein and
thereby prevents/blocks virus fusion to the host cell as well as cell to cell fusion [76]. Palivizumab
use is restricted to premature infants 32-35 weeks gestation due to high cost and is used as
prophylactic treatment [77]. Adding to Palivizumab’s high cost, a Palivizumab-resistant variant
of RSV have been isolated from Palivizumab treated patients [78]. Vaccination and vaccine
development has been held back by the severe RSV infection which occurred in a 1960 vaccination
trial in which majority of vaccinated infants had a vaccine-enhanced disease that require
hospitalization with several fatalities [79]. Animal models for RSV infection is an essential step
in the investigation of RSV pathogenesis and the search for new therapeutics and vaccines.
RSV causes respiratory tract lesions that include severe bronchiolitis characterized by
epithelial cell degeneration and necrosis with areas of hyperplasia in response to the cell damage
along with syncytial cell formation. The bronchiolar lumen becomes partially occluded by
neutrophils, occasional macrophages, seroproteinaceous fluid, mucin, and cell debris.
Lymphocytes infiltrate the airway adventitia. The virus also infects ciliated epithelial cells of the
upper respiratory tract and bronchi. With time after infection, lesions can resolve. However, there
can be increases in Goblet cells and increased residual mast cells and eosinophils.

8
Animal Models of RSV Infection
Animal models are needed to study RSV pathogenesis and evaluate new therapeutics and
vaccines candidates. Animal models are considered the middle stage between tissue and cell
culture studies in vitro and human clinical trials. However, developing animal models for RSV
infections is very challenging due to the high degree of specificity of the hRSV to its natural host
and lack of virulence in other species. The specificity of hRSV is due to F protein, while lack of
virulence in other species is due to the inability to block interferon response [80,81]. Ideal RSV
models need to mimic several RSV disease aspects such as clinical signs and symptoms, viral
replication, upper and lower respiratory pathology, and immune response. Several animal models
have been developed to address some of these aspects if not all, but each model has its strengths
and limitations. In general, RSV animal models are either heterologous or cognate host-virus
models. hRSV can infect and replicate in heterologous host-virus models such as chimpanzees,
baboons sheep, Cotton rats, ferrets and mice, while related Orthopneumovirus specific to the
model was used in the cognate host-virus models such as murine pneumonia virus in mice model
and bovine RSV (bRSV) in calves [16]. In this section, several animal models of RSV infection
will be discussed concluding with the neonatal lamb model.
Chimpanzees seem ideal as RSV animal model due to anatomical similarity to human and
since RSV isolated originally from chimpanzees with respiratory tract infection in 1956 [82].
hRSV able to infect and replicate in the nasal sinuses and upper respiratory tract epithelium, and
induce disease symptoms similar to that found in human RSV associated upper respiratory tract
infection [83,84]. The drawback of this model is that chimpanzees rarely develop RSV LRTI.
There are also concerns with the substantial economic, ethical and emotional burden associated
with the use of chimpanzees. However, vaccine studies have benefited from chimpanzees since

9
chimpanzees tend to develop anti-RSV neutralizing antibody and their immune response similar
to that found in human [85,86]. Baboons have been used as well and are being bred in large
numbers at Oklahoma State University for use in RSV studies and other disease conditions [87,88].
Although rodents are an excellent animal model for experimental studies, they are considered
semi-permissive for hRSV replication, and need a large inoculum to induce mild to moderate RSV
disease[89,90]. Clinical signs are difficult to interpret in rodents, and RSV induces only mild to
moderate bronchiolitis and pneumonia[91,92]. Cotton rats are more permissive than mice for RSV
replication and considered the standard model for testing RSV therapeutics[93]. Mice, however,
have the advantage of wide variety of transgenetic mice and the availability of molecular markers.
Furthermore, mice can be used as a cognate host-virus model by usingmurine pneumonia virus,
which develop a disease in mice similar to RSV disease in human[94]. Murine pneumonia virus
targets bronchiolar epithelium and lead to severe disease with marked respiratory disease correlate
positively with the viral inoculum[94,95]. The drawback of murine pneumonia virus model of
hRSV is the far phylogenetic distance between the two viruses [96]. The critical disadvantage of
rodents as a model for RSV disease is the difference in lung anatomy, histology and immune
response between human and rodents that subsequently question the translations of studies done
in these models to human.
Another cognate host-virus model for hRSV is the bRSV in calves. bRSV induce a
respiratory disease in calves similar to what seen in human RSV. bRSV is more closely related to
hRSV than other non-RSV viruses and share about 38-41% homology on nucleotide level[97].
bRSV induces upper and lower respiratory tract infection in calves[98]. Furthermore, calves have
similar lung anatomy and histology, i.e., the presence of pharyngeal and nasopharyngeal tonsils,
the presence of ciliated pseudostratified epithelium and submucosal glands, and similar innate and

10
adaptive immune response to human [99,100]. The drawback of the calves bRSV model is the
large size of calves that need special housing and handling. Also, calves are not susceptible to
human strains of RSV. Thus, bRSV and hRSV are two distinct viruses that induce similar disease
process in their natural target host.
Neonatal lambs infected with RSV has several similarities to a human infant advantageous
for comparison. The most important criterion is that lambs are naturally susceptible to human,
ovine and bovine strains of RSV. hRSV replicates mostly in the lower part of the respiratory tract
of lamb lungs (bronchioles and bronchi) which models well bronchiolitis in infants and reduces
airflow to alveoli. hRSV replicates well in neonatal lamb respiratory tract airways with a peak of
viral replication at day 6 after intratracheal inoculation with A2 hRSV strain then declines with
time[17,18]. However, another study using hRSV M37 strain showed that peak viral replication
was at day 3 and persisted until day 6 post nebulization[101]. As in human infants, lambs have
variable clinical signs associated with hRSV infection. Clinical symptoms vary from mild such as
nausea, fever, reluctant to move, and reduce milk consumption to moderate and severe such as
cough, wheezing, expiratory efforts. Signs of infection appear as early as 2 days post infection
and progress till day 6 [17,101,102]. hRSV infected lambs develop lower respiratory tract
infection characterized by moderate to severe bronchiolitis and interstitial pneumonia. There is
modest thickening of alveolar septae due to edema, type II cell hyperplasia and, leukocytes
infiltration in the alveolar wall, and neutrophils and macrophages infiltration into the alveolar and
bronchiolar lumens. In addition, hRSV incites epithelial cell necrosis, hyperplasia of nearby
epithelial cells and syncytial cell formation in lambs [103] as in infants.
Additionally, immunological response to hRSV in neonatal lambs were characterized by the
Th1 proinflammatory response as it characterized by the elevated level of IFN-γ and TNFα

11
cytokine and decrease in the level of TGF-β and IL-10. Leukocyte recruitment to the lung were
associated with IL-8, MCP-1, and MIP-1α increase in the lung [18]. IL-10 increased to the highest
level in lambs lung at day 3 post-infection while other cytokine increase later in the course of the
disease [101,104]. Lambs have many similarities to human infant that make it ideal for studying
infant respiratory diseases. Lung development is similar between lambs and infants in terms of
alveolarization which occurs prenatally, similar airways size and branching (dichotomous
branching), presence of submucosal glands, and similar percent of club cells in the airways[105–
108]. Lambs can be born preterm similar to human. Since there is no transfer of immunoglobulin
from ewes to their lambs in utero, lambs can be colostrum deprived to avoid transmission of
maternal immunoglobulin to the lambs. Finally, the size of lambs allows for better evaluation of
the clinical signs, sampling, and lung evaluation.
Immune Response to RSV Infection:
Cell-mediated immune response guided by the proliferation of cytotoxic T cell is the
preferred response to clear many viral infections, although both cell-mediated and humoral
response needed for the ideal antiviral response. The innate immune response, however, has a huge
effect in limiting RSV replication and respiratory infection in infants.
The innate immune response is a nonspecific response aiming to prevent and reduce initial
infection to allow time for more specific acquired immune responses to develop. Innate barriers in
the respiratory tract such as mucociliary system act to prevent attachment of RSV to the airway
epithelial cell, which is the target cell for RSV. Additionally, in response to infection, infected or
neighboring cell upregulate genes, and produce cytokines and antimicrobial peptides and proteins,
which limit the microbial proliferation. There are several common cells in the bronchoalveolar
tree: ciliated bronchial epithelial cells, club cells, non-ciliated cells of the bronchiolar epithelium,

12
and pneumocytes (Type II and Type I). These cells sense the presence of RSV through pattern
recognition receptor most importantly TLR-2, TLR-4, TLR-6, TLR-7, TLR-8, Retinoic acid-
inducible gene I-liKe receptor, and MDA-5[109,110]. Activation of the TLRs eventually leads to
the production of cytokines that shape up the immune response and attract more immune cells to
the lung. Such cytokines include IL-8, IL-10, and IL-6 that induced by TLR4 activation, RANTES,
which produced in response to TLR3 activation, and IFN-α, IFN-β and IP10 production in response
to RIG-I activation[109,111–114]. IL-8, which is a neutrophils chemoattractant, is produced in
the respiratory tract during RSV infection and higher levels of IL-8 is associated with more severe
RSV bronchiolitis in human[115,116]. IL-10 and IL-6 are associated with the Th2 response.
RANTES is a chemoattractant for T cells and eosinophils and associated with severe RSV
bronchiolitis and induction of allergic cellular response [117–119]. IL-10 is an anti-inflammatory
cytokine produced by inflammatory cells mainly macrophages and elevated during acute RSV
infection[120,121]. IFN-α and IFN-β are type I interferons that act on reducing viral replication
and promote MHC type I upregulation and subsequently killing of viral infected cells. Although
IFN-α is elevated in RSV infected infants, its level is markedly lower than influenza-infected
patients[122–124]. That is may be due to the ability of RSV to resist Interferon through the
function of NS1 and NS2 proteins [125–127]. IFNγ, which is type II interferon, is associated with
Th1 response, and it is produced in part by natural killer and macrophages. Low level of IFNγ is
associated with severe RSV lower respiratory tract infection [128]. On the other hand, high level
of type III interferon, IFN-λ, is associated with increased RSV severity. Type III interferon has
similar activity to type I IFN[129].
Mucus and fluid covering the respiratory epithelium contain other innate antimicrobial
substances that act on deactivating microorganisms such as secretion of the submucosal glands

13
and antimicrobial peptides. Submucosal glands, which are present in both human and ruminant
lung airways, secrete lactoperoxidase, lactoferrin, and lysozyme into the mucosal surface.
Lactoperoxidase acts on thiocyanate and hydrogen peroxide present on the mucosal surface to
produce oxythiocyanate which has antimicrobial activity against bacteria and viruses[130,131].
Antimicrobial peptides present in the respiratory system include alpha and beta-defensins, and
cathelicidin. Significant antimicrobial proteins include surfactant protein A and D. Both SP- A
and D are produced by type II pneumocytes and club cells into the airways lumen. Surfactant
protein A and D are globular mannose-binding C-type lectins that bind to RSV F protein and
enhance clearance[132–134]. A higher level of SP-A and SP-D are associated with severe RSV
LRTI[135]. Beta-defensins are antimicrobial peptides secreted by respiratory epithelial cells either
constitutively such as HBD1 or inducible upon infection of the cell such as HBD2, 3, and 4.
Inducible HBD bind to RSV envelop preventing the virus from infecting the cell[136]. Sheep
produce Sheep beta defensins 1 and 2. In contrast, the antimicrobial peptide Cathelicidin is
secreted by leukocytes and stored in neutrophils and can be upregulated during viral
infection[137]. All these innate immune responses are directed towards carbohydrate or other
moieties rather than specific antigens and some are upregulated with RSV lower respiratory tract
infection. In addition to the ability to deactivate RSV, some of these molecules act as a
chemoattractant for other inflammatory and immune cells.
Several inflammatory cells are associated with severe RSV lower respiratory tract infection.
However, neutrophils are the predominant inflammatory cell that increased with the RSV both in
peripheral blood and in the respiratory system [138,139], and the increased number of neutrophils
in blood correlated with the severity of the disease and the peak of the viral load[140]. These
neutrophils are activated and producing neutrophil elastase[141], with neutrophils apoptosis and

14
NETosis to trap virions and prevent further spread of virions[142]. In contrast, eosinophils also
are activated during RSV infection, but they are associated with the healing process[143].
Eosinophils are increased following some RSV infections and contribute to asthma development
later in life and peripheral eosinophils are increased in children hospitalized for RSV LRTI along
with an increase in the Leukotriene C4, eosinophil-derived neurotoxin, and eosinophil cationic
protein in the respiratory tract [85,144,145]. Other inflammatory/ innate immune response
associated cells are macrophages, Natural killer cells, and Dendritic cells.
Although there is a predominance of neutrophil in the airway lavage fluid obtained from
severe RSV infected infant, changes in other cell type can be associated with the disease
outcomes[146,147]. Alveolar macrophages have an essential role in phagocytizing foreign bodies
and microorganisms leading to the subsequent microbial killing and antigen presentation. In
addition to the expression of RSV glycoproteins, alveolar macrophages associated with RSV
infection express immune modulatory molecules such as HLA-DR, interleukin-1β, and TNF as a
response to lung injury[148,149]. RSV immunoreactive staining were seen in alveolar
macrophages in lambs infected with hRSV[150]. Similar to macrophages, dendritic cells (DC)
internalize proteins, process and present antigen to other immune cells, and both conventional and
plasmacytoid DC are recruited to the respiratory system early in RSV infection. These DC are
activated and express a proinflammatory phenotype[151–153]. Most of these cells are nonspecific
and respond to RSV infection to either eliminate the virus or stimulate other type of immune cells
such as lymphocytes.
Lymphocytes are critical for RSV infection since they determine the magnitude and type of
the acquired immune response. However, T lymphocyte number in the blood is decreased in
patients with RSV associated LRTI, and the reduction in T lymphocytes number correlated with

15
the severity of RSV infection [138,154]. In contrast, there is an increase in the B lymphocytes
number in circulation during severe RSV bronchiolitis [155]. Both T and B lymphocytes are
needed to resolve RSV disease.
During RSV infection, there is a decrease in the number of all T lymphocyte population in
the blood, which is more pronounced in infants, and these peripheral lymphocytes are not activated
as is shown by the low level of expression of CD11a and CTL-4 markers[138,156,157]. Although
there is a predominance of CD4 T cell in the BALF obtained from early severe RSV infected
infants, there is a higher expansion of effector CD8 T cell with the course of infection[146,158].
Most research in RSV immunobiology suggests an imbalance in Th1/Th2 response, which
determines the severity of RSV infection. While Th1 responses associated with cytotoxic T
lymphocytes activation and IgG type immunoglobulin production result in resolution of RSV
infection, Th2 shifted responses are associated with increased mucus secretion, cellular infiltration
and atopic type reaction characterized by eosinophilia and eosinophil infiltration in the lung. Th1
response characterized by elevation of IFN-γ, IL-1, IL-2, IL-12, IL-18, and TNF-α, while TH2
response associated with the increase in IL-4, IL-5, IL-6, IL-9, IL-10, and IL-13[159]. Since TH1
and Th2 mutually inhibit each other by IFN-γ to suppress Th2 and IL-4 to inhibit Th1, INF-γ/IL-
4 ratio is used as an indicator for the Th2 bias[160]. There is lower IFN-γ and higher IL-4 leading
to higher IFN-γ/IL-4 ratio in RSV associated bronchiolitis, in addition to the higher IL-10/IL-12
ratio[161]. This high IFN-γ/IL-4 ratio indicates either poor Th1 response or enhanced Th2
response. The fact that IFNγ (Th1 cytokine) level in RSV infected infants is lower than what found
in Pneumovirus infected infants and that the IL-4 level (Th2 cytokine) is higher in RSV infected
infants, it suggests that theTh2 response is enhanced in RSV infected infants[162]. Another study

16
in RSV infected infant indicted the predominance of the Th2 cytokines in the nasopharyngeal fluid
[163]. Thus, Th2 enhanced response might be associated with the severe RSV bronchiolitis.
There is an increase in B-lymphocytes in the blood of RSV lower respiratory tract infected
infants. Humoral immune response to RSV infection includes IgM, IgA, and IgG production
within 5-10 days post infection. However, lower immunoglobulin responses are detected in
children under 6 months old. Both free and cell bounded anti-RSV IgA is present in the
nasopharyngeal secretion of RSV infected patients. Anti-RSV IgG is increased in RSV infected
patients in both IgG1 and IgG3 subclasses. Antibodies directed to F protein crossreact with
different RSV strains, while anti-G protein antibodies are strain specific [164,165]. Although both
anti-RSV IgG and IgA are associated with protection against RSV infection, nasal IgA is more
protective than serum neutralizing IgG antibody[166,167]. A similar finding was seen in children
where high IgA level seemed to be associated with recovery[168]. Another study indicated that
lack of IgA RSV-specific memory B cells in the blood of experimentally infected adults and this
may explain the susceptibility to recurrent infection with RSV [169].
Secondary Bacterial Infection
Secondary bacterial infection is a challenging potential sequel to viral pneumonia. There are
few reports of bacterial infections secondary to RSV infections due to the limitation in the
diagnosis of such infections i.e., the bacterial infection masked by the pathological changes
induced by the virus with difficulty in obtaining noninvasive specific diagnostic samples[170].
Thus, there are few reports on the magnitude/extent of bacterial infection secondary to RSV
infection. One study indicates approximately 40% of patients hospitalized due to viral respiratory
tract infection were associated with bacterial co-infection[171], while another study showed that
40% of children infected with RSV have bacterial coinfection or in high risk of bacterial

17
pneumonia [6]. The most common bacteria associated with secondary bacterial pneumonia are
Streptococcus pneumoniae, Staphylococcus aureus, Streptococcus pyogenes, and Haemophilus
influenza[172,6,173,174]. Because of a high percent of viral-bacterial coinfections, a synergistic
relationship between the two microbial agents that leads to enhanced host susceptibility to each
pathogen has been proposed.
Several in vitro and in vivo models investigated the relationship between viral and bacterial
respiratory infection and how these two microorganisms enhance virulence and persistence of both
pathogens. One study determined that IFN elevation induced by RSV infection enhanced
Pseudomonas aeruginosa biofilm formation in human bronchial epithelial cell culture along with
an increase in iron and Iron transport protein (Transferrin) secretion in the apical epithelium
promoting P. aeruginosa biofilm both in vitro and in vivo [175]. Another study with influenza
infected mice followed by Streptococcus pneumoniae (Spn) infected challenge revealed increases
in both viral and bacterial titers. It is thought that Spn leads to enhanced viral release from infected
cells, and alveolar macrophages impairment resulting in increased Spn [176]. An In vitro study
determined a 2-10 fold increase in the adherence of Spn to RSV infected respiratory cell line in
comparison to non-infected cells[177], while another study showed that 2-2.2 fold increase in
Haemophilus influenza and Streptococcus pneumoniae attachment to cells expressing RSV G
glycoprotein than cells infected with vector only. Furthermore, this bacterial adherence was
reduced to 78-84% when cells were incubated with anti-RSV G antibody [178]. In addition to
viral-specific characteristics that enhances bacterial coinfection, the viral infection itself lead to
alterations in host susceptibility to infection.
There are several potential mechanisms by which viruses predispose to secondary bacterial
infection. The most critical factor is epithelial necrosis and damage, which give access for

18
pathogenic bacterial to its receptors and to obtain iron and other micronutrients [179].
Additionally, viral infection leads to disabled innate barriers such as loss of mucociliary clearance
mechanisms leading to increased bacterial colonization and increased numbers of bacteria reaching
deeper location within the respiratory tree[180]. Viral infection creates a microenvironment
suitable for bacterial proliferation such as aggregation of fibrin, mucus, and necrotic cells, and the
occlusion of small airways leading to reduced O2 and CO2 concentration[181,182]. Furthermore,
bacterial coinfection increases viral titer and prolong the course of infection with some reports
demonstrating that bacterial infection increases susceptibility to viral infection [180,183].
RSV Therapeutics
RSV is a global burden and contributes significantly to increased hospital admission rates
and mortality due to severe bronchiolitis and pneumonia, and there is no effective specific
therapeutics for RSV infection. There are only two approved treatments, Ribavirin, and
palivizumab, although Ribavirin no longer being recommended for treatment and Palivizumab
recommended as a prophylactic treatment only in high-risk individuals. Additionally,
corticosteroids and bronchodilator have limited effects and benefits. The only therapeutic options
available in clinical settings are supportive cares such as oxygenation and intravenous fluid[184].
There is an increased interest from pharmaceutical companies to develop direct antiviral
compound suitable for treatment of RSV infection especially for infants and immunocompromised
individuals. Several compounds that differ in their characteristics and molecular targets have been
evaluated, and some have reach advanced clinical trials.
Four classes of anti-RSV therapeutics been developed and investigated: immunoglobulins,
siRNA-interference, fusion inhibitors, and small molecule inhibitors. Several types of
Immunoglobulins have been developed and most of them targeting RSV F protein. Both

19
polyclonal and monoclonal antibodies have been utilized as therapies with monoclonal showing
the higher neutralizing effect and lower adverse effects. siRNA is used to interfere with RSV-
directed protein synthesis. Anti-RSV therapeutic been developed target five different RSV
proteins[15].
Since RSV F protein is critical for RSV entry and spread to adjacent cells, most of the anti-
RSV therapeutics target F protein. Several F protein inhibitors are either neutralizing antibody
fragments or small molecules. Two compounds in this category (ALX-0171, REGN-2222) have
reached advanced clinical trials (phase 3 human clinical trials). ALX-0171 is trimeric Nanobody
(camelia antibody ) that binds the antigenic site II of RSV F protein [185], while REGN-2222 is a
fully human monoclonal antibody[186]. Both antibodies bind to prefusion F protein preventing
entry and multicyclic RSV infection. Other small molecules that target F protein have reached
advanced clinical trials. These anti F protein compounds bind to F protein preventing its functional
activity and transformation to post-fusion conformation. Since the transformation of the metastable
pre-fusion to the 6-helix bundle post-fusion conformation is critical for RSV entry to the cell, these
anti-F small molecule inhibitors deactivate RSV and limit the infection. Several molecules in this
category reached advanced clinical trails such as GS-5806, JNJ-53718678, and AK0529). One
downside for F protein inhibitor is the development of resistant viral mutation (escape mutations)
[187]. Another RSV target is the L-protein, which is the RNA-dependent- RNA polymerase.
Previously recommended anti-RSV compound Ribavirin is in this class. ALS-008176, a
nucleoside analog chain terminator, is a newly developed RSV polymerase inhibitor developed by
Alios Janssen and now in advanced clinical trials[188]. Other target proteins are N protein, SH
protein, and M2-1 protein. Several anti-RSV molecules developed targeting these proteins, and

20
they are progressing toward clinical trails [189–191]. Another possible anti-RSV infection
therapeutic is to target host immune response to infection.
The severe inflammatory and cellular infiltration associated with severe RSV infection lead
to the belief of bidirectional approach in developing anti-RSV lower respiratory tract infection
therapeutic, i.e., specific anti-RSV molecules and immunomodulatory therapeutic such as
chemokines and anti-leukotriene[192]. Blocking either of CCL3 (MIP-1α) or CCL5 (RANTES) in
RSV infected animal model showed a significant reduction in the recruitment of inflammatory
cells and increased survival[193,194]. Anti-leukotriene is also investigated since leukotriene is
prominent during RSV bronchiolitis and showed that anti-leukotriene administration reduces
inflammatory cellular infiltrate, reduced airway blockage, and reduced bronchiolitis in RSV
infected mice and infants [195,196]. Other immunomodulating agents such as surfactant protein,
vascular endothelial growth factor are also investigated and reduced RSV-associated pathology
[197].
References
1. Nair H, Nokes DJ, Gessner BD, et al. Global burden of acute lower respiratory infections due to
respiratory syncytial virus in young children: a systematic review and meta-analysis. The Lancet.
2010; 375(9725):1545–1555.
2. Falsey AR, Hennessey PA, Formica MA, Cox C, Walsh EE. Respiratory Syncytial Virus Infection
in Elderly and High-Risk Adults. N Engl J Med. 2005; 352(17):1749–1759.
3. Widmer K, Zhu Y, Williams JV, Griffin MR, Edwards KM, Talbot HK. Rates of Hospitalizations
for Respiratory Syncytial Virus, Human Metapneumovirus, and Influenza Virus in Older Adults. J
Infect Dis. 2012; :jis309.
4. Hon KL, Leung TF, Cheng WY, et al. Respiratory syncytial virus morbidity, premorbid factors,
seasonality, and implications for prophylaxis. J Crit Care. 2012; 27(5):464–468.
5. Mizuta K, Abiko C, Aoki Y, et al. Seasonal Patterns of Respiratory Syncytial Virus, Influenza A
Virus, Human Metapneumovirus, and Parainfluenza Virus Type 3 Infections on the Basis of Virus
Isolation Data between 2004 and 2011 in Yamagata, Japan. Jpn J Infect Dis. 2013; 66(2):140–145.

21
6. Thorburn K, Harigopal S, Reddy V, Taylor N, Saene HKF van. High incidence of pulmonary
bacterial co-infection in children with severe respiratory syncytial virus (RSV) bronchiolitis.
Thorax. 2006; 61(7):611–615.
7. Peebles RS. Viral infections, atopy, and asthma: Is there a causal relationship? J Allergy Clin
Immunol. 2004; 113(1, Supplement):S15–S18.
8. Wu P, Dupont WD, Griffin MR, et al. Evidence of a Causal Role of Winter Virus Infection during
Infancy in Early Childhood Asthma. Am J Respir Crit Care Med. 2008; 178(11):1123–1129.
9. Ramilo O, Lagos R, Sáez-Llorens X, et al. Motavizumab treatment of infants hospitalized with
respiratory syncytial virus infection does not decrease viral load or severity of illness. Pediatr
Infect Dis J. 2014; 33(7):703–709.
10. Lagos R, DeVincenzo JP, Muñoz A, et al. Safety and antiviral activity of motavizumab, a
respiratory syncytial virus (RSV)-specific humanized monoclonal antibody, when administered to
RSV-infected children. Pediatr Infect Dis J. 2009; 28(9):835–837.
11. Langley JM, Aggarwal N, Toma A, et al. A Randomized, Controlled, Observer-Blinded Phase 1
Study of the Safety and Immunogenicity of a Respiratory Syncytial Virus Vaccine With or
Without Alum Adjuvant. J Infect Dis. 2017; 215(1):24–33.
12. Teng MN, Tran KC. Enhancing immunogenicity of respiratory syncytial virus vaccine candidates
by altering NS1 function. J Allergy Clin Immunol. 2017; 139(2):AB270.
13. Blanco JCG, Pletneva LM, Otoa RO, Patel MC, Vogel SN, Boukhvalova MS. Preclinical
assessment of safety of maternal vaccination against respiratory syncytial virus (RSV) in cotton
rats. Vaccine. 2017; 35(32):3951–3958.
14. Jorquera PA, Tripp RA. Respiratory syncytial virus: prospects for new and emerging therapeutics.
Expert Rev Respir Med. 2017; 11(8):609–615.
15. Mazur NI, Martinón-Torres F, Baraldi E, et al. Lower respiratory tract infection caused by
respiratory syncytial virus: current management and new therapeutics. Lancet Respir Med. 2015;
3(11):888–900.
16. Bem RA, Domachowske JB, Rosenberg HF. Animal models of human respiratory syncytial virus
disease. Am J Physiol-Lung Cell Mol Physiol. 2011; 301(2):L148–L156.
17. Olivier A, Gallup J, De Macedo MMMA, Varga SM, Ackermann M. Human respiratory syncytial
virus A2 strain replicates and induces innate immune responses by respiratory epithelia of neonatal
lambs. Int J Exp Pathol. 2009; 90(4):431–438.
18. Sow FB, Gallup JM, Olivier A, et al. Respiratory syncytial virus is associated with an
inflammatory response in lungs and architectural remodeling of lung-draining lymph nodes of
newborn lambs. Am J Physiol-Lung Cell Mol Physiol. 2010; 300(1):L12–L24.
19. Rima B, Collins P, Easton A, et al. ICTV Virus Taxonomy Profile: Pneumoviridae. J Gen Virol.
2017; 98(12):2912–2913.

22
20. Human orthopneumovirus Subgroup A, complete cds. 2018 [cited 2019 Jan 24]; . Available from:
http://www.ncbi.nlm.nih.gov/nuccore/NC_038235.1
21. Techaarpornkul S, Barretto N, Peeples ME. Functional Analysis of Recombinant Respiratory
Syncytial Virus Deletion Mutants Lacking the Small Hydrophobic and/or Attachment
Glycoprotein Gene. J Virol. 2001; 75(15):6825–6834.
22. Hallak LK, Spillmann D, Collins PL, Peeples ME. Glycosaminoglycan Sulfation Requirements for
Respiratory Syncytial Virus Infection. J Virol. 2000; 74(22):10508–10513.
23. Palomo C, García-Barreno B, Peñas C, Melero JA. The G protein of human respiratory syncytial
virus: significance of carbohydrate side-chains and the C-terminal end to its antigenicity. J Gen
Virol. 1991; 72(3):669–675.
24. Bukreyev A, Yang L, Fricke J, et al. The Secreted Form of Respiratory Syncytial Virus G
Glycoprotein Helps the Virus Evade Antibody-Mediated Restriction of Replication by Acting as
an Antigen Decoy and through Effects on Fc Receptor-Bearing Leukocytes. J Virol. 2008;
82(24):12191–12204.
25. Bukreyev A, Yang L, Collins PL. The Secreted G Protein of Human Respiratory Syncytial Virus
Antagonizes Antibody-Mediated Restriction of Replication Involving Macrophages and
Complement. J Virol. 2012; 86(19):10880–10884.
26. Fleming EH, Kolokoltsov AA, Davey RA, Nichols JE, Roberts NJ. Respiratory Syncytial Virus F
Envelope Protein Associates with Lipid Rafts without a Requirement for Other Virus Proteins. J
Virol. 2006; 80(24):12160–12170.
27. Kumaria R, Iyer LR, Hibberd ML, Simões EA, Sugrue RJ. Whole genome characterization of non-
tissue culture adapted HRSV strains in severely infected children. Virol J. 2011; 8:372.
28. Lamb RA, Jardetzky TS. Structural basis of viral invasion: lessons from paramyxovirus F. Curr
Opin Struct Biol. 2007; 17(4):427–436.
29. Krzyzaniak MA, Zumstein MT, Gerez JA, Picotti P, Helenius A. Host Cell Entry of Respiratory
Syncytial Virus Involves Macropinocytosis Followed by Proteolytic Activation of the F Protein.
PLOS Pathog. 2013; 9(4):e1003309.
30. Huang YT, Collins PL, Wertz GW. Characterization of the 10 proteins of human respiratory
syncytial virus: Identification of a fourth envelope-associated protein. Virus Res. 1985; 2(2):157–
173.
31. Collins PL, Mottet G. Membrane orientation and oligomerization of the small hydrophobic protein
of human respiratory syncytial virus. J Gen Virol. 1993; 74(7):1445–1450.
32. Fuentes S, Tran KC, Luthra P, Teng MN, He B. Function of the Respiratory Syncytial Virus Small
Hydrophobic Protein. J Virol. 2007; 81(15):8361–8366.
33. Gan S-W, Tan E, Lin X, et al. The Small Hydrophobic Protein of the Human Respiratory
Syncytial Virus Forms Pentameric Ion Channels. J Biol Chem. 2012; 287(29):24671–24689.

23
34. Das K, Arnold E. Negative-Strand RNA Virus L Proteins: One Machine, Many Activities. Cell.
2015; 162(2):239–241.
35. Yabukarski F, Lawrence P, Tarbouriech N, et al. Structure of Nipah virus unassembled
nucleoprotein in complex with its viral chaperone. Nat Struct Mol Biol. 2014; 21(9):754–759.
36. Berthiaume L, Joncas J, Pavilanis V. Comparative structure, morphogenesis and biological
characteristics of the respiratory syncytial (RS) virus and the pneumonia virus of mice (PVM).
Arch Für Gesamte Virusforsch. 1974; 45(1–2):39–51.
37. Fearns R, Collins PL. Role of the M2-1 Transcription Antitermination Protein of Respiratory
Syncytial Virus in Sequential Transcription. J Virol. 1999; 73(7):5852–5864.
38. Tran T-L, Castagné N, Dubosclard V, et al. The Respiratory Syncytial Virus M2-1 Protein Forms
Tetramers and Interacts with RNA and P in a Competitive Manner. J Virol. 2009; 83(13):6363–
6374.
39. Wertz GW, Moudy RM. Antigenic and genetic variation in human respiratory syncytial virus.
Pediatr Infect Dis J. 2004; 23(1):S19.
40. Mufson MA, Örvell C, Rafnar B, Norrby E. Two Distinct Subtypes of Human Respiratory
Syncytial Virus. J Gen Virol. 1985; 66(10):2111–2124.
41. Walsh EE, McConnochie KM, Long CE, Hall CB. Severity of Respiratory Syncytial Virus
Infection Is Related to Virus Strain. J Infect Dis. 1997; 175(4):814–820.
42. Zhang Z, Du L, Chen X, et al. Genetic Variability of Respiratory Syncytial Viruses (RSV)
Prevalent in Southwestern China from 2006 to 2009: Emergence of Subgroup B and A RSV as
Dominant Strains. J Clin Microbiol. 2010; 48(4):1201–1207.
43. Arbiza J, Delfraro A, Frabasile S. Molecular epidemiology of human respiratory syncytial virus in
Uruguay: 1985-2001 - A review. Mem Inst Oswaldo Cruz. 2005; 100(3):221–230.
44. Panayiotou C, Richter J, Koliou M, Kalogirou N, Georgiou E, Christodoulou C. Epidemiology of
respiratory syncytial virus in children in Cyprus during three consecutive winter seasons (2010–
2013): age distribution, seasonality and association between prevalent genotypes and disease
severity. Epidemiol Amp Infect. 2014; 142(11):2406–2411.
45. Do AHL, Doorn HR van, Nghiem MN, et al. Viral Etiologies of Acute Respiratory Infections
among Hospitalized Vietnamese Children in Ho Chi Minh City, 2004–2008. PLOS ONE. 2011;
6(3):e18176.
46. Paes BA, Mitchell I, Banerji A, Lanctôt KL, Langley JM. A Decade of Respiratory Syncytial
Virus Epidemiology and Prophylaxis: Translating Evidence into Everyday Clinical Practice
[Internet]. Can. Respir. J. 2011 [cited 2018 Jan 10]. Available from:
https://www.hindawi.com/journals/crj/2011/493056/abs/
47. Simoes EA, Carbonell-Estrany X. Impact of severe disease caused by respiratory syncytial virus in
children living in developed countries. Pediatr Infect Dis J. 2003; 22(2 Suppl):S13-18; discussion
S18-20.

24
48. Falsey AR, McElhaney JE, Beran J, et al. Respiratory Syncytial Virus and Other Respiratory Viral
Infections in Older Adults With Moderate to Severe Influenza-like Illness. J Infect Dis. 2014;
209(12):1873–1881.
49. Whimbey E, Ghosh S. Respiratory syncytial virus infections in immunocompromised adults. Curr
Clin Top Infect Dis. 2000; 20:232–255.
50. Hynicka LM, Ensor CR. Prophylaxis and Treatment of Respiratory Syncytial Virus in Adult
Immunocompromised Patients. Ann Pharmacother. 2012; 46(4):558–566.
51. Thompson WW, Shay DK, Weintraub E, et al. Mortality Associated With Influenza and
Respiratory Syncytial Virus in the United States. JAMA. 2003; 289(2):179–186.
52. Ducoffre G, Cauchi P, Hendrickx E. Respiratory syncytialvirus epidemiology in Belgium in 1998,
1999, and 2000 (Poster). 2001. 2001; 29:352.
53. Nicholson KG. Impact of influenza and respiratory syncytial virus on mortality in England and
Wales from January 1975 to December 1990. Epidemiol Amp Infect. 1996; 116(1):51–63.
54. Numa A. Outcome of respiratory syncytial virus infection and a cost–benefit analysis of
prophylaxis. J Paediatr Child Health. 2000; 36(5):422–427.
55. Bloom-Feshbach K, Alonso WJ, Charu V, et al. Latitudinal Variations in Seasonal Activity of
Influenza and Respiratory Syncytial Virus (RSV): A Global Comparative Review. PLOS ONE.
2013; 8(2):e54445.
56. Lindsley WG, Blachere FM, Davis KA, et al. Distribution of Airborne Influenza Virus and
Respiratory Syncytial Virus in an Urgent Care Medical Clinic. Clin Infect Dis. 2010; 50(5):693–
698.
57. Law BJ, Carbonell-estrany X, Simoes EAF. An update on respiratory syncytial virus
epidemiology: a developed country perspective. Respir Med. 2002; 96:S1–S7.
58. Kristensen K, Hjuler T, Ravn H, Simões EAF, Stensballe LG. Chronic Diseases, Chromosomal
Abnormalities, and Congenital Malformations as Risk Factors for Respiratory Syncytial Virus
Hospitalization: A Population-Based Cohort Study. Clin Infect Dis. 2012; 54(6):810–817.
59. GOUYON J-B, ROZÉ J-C, GUILLERMET-FROMENTIN C, et al. Hospitalizations for
respiratory syncytial virus bronchiolitis in preterm infants at <33 weeks gestation without
bronchopulmonary dysplasia: the CASTOR study. Epidemiol Infect. 2013; 141(4):816–826.
60. Hall CB, Weinberg GA, Iwane MK, et al. The Burden of Respiratory Syncytial Virus Infection in
Young Children. N Engl J Med. 2009; 360(6):588–598.
61. Law BJ, Langley JM, Allen U, et al. The Pediatric Investigators Collaborative Network on
Infections in Canada Study of Predictors of Hospitalization for Respiratory Syncytial Virus
Infection for Infants Born at 33 Through 35 Completed Weeks of Gestation. Pediatr Infect Dis J.
2004; 23(9):806.
62. Boyce TG, Mellen BG, Mitchel EF, Wright PF, Griffin MR. Rates of hospitalization for
respiratory syncytial virus infection among children in Medicaid. J Pediatr. 2000; 137(6):865–870.

25
63. Figueras-Aloy J, Carbonell-Estrany X, Quero-Jiménez J, et al. FLIP-2 Study: Risk Factors Linked
to Respiratory Syncytial Virus Infection Requiring Hospitalization in Premature Infants Born in
Spain at a Gestational Age of 32 to 35 Weeks. Pediatr Infect Dis J. 2008; 27(9):788.
64. Bradley JP, Bacharier LB, Bonfiglio J, et al. Severity of Respiratory Syncytial Virus Bronchiolitis
Is Affected by Cigarette Smoke Exposure and Atopy. Pediatrics. 2005; 115(1):e7–e14.
65. Saleeby E, M C, Bush AJ, Harrison LM, Aitken JA, DeVincenzo JP. Respiratory Syncytial Virus
Load, Viral Dynamics, and Disease Severity in Previously Healthy Naturally Infected Children. J
Infect Dis. 2011; 204(7):996–1002.
66. Buckingham SC, Bush AJ, Devincenzo JP. Nasal quantity of respiratory syncytical virus correlates
with disease severity in hospitalized infants. Pediatr Infect Dis J. 2000; 19(2):113–117.
67. Fodha I, Vabret A, Ghedira L, et al. Respiratory syncytial virus infections in hospitalized infants:
Association between viral load, virus subgroup, and disease severity. J Med Virol. 2007;
79(12):1951–1958.
68. Gilca R, De Serres G, Tremblay M, et al. Distribution and Clinical Impact of Human Respiratory
Syncytial Virus Genotypes in Hospitalized Children over 2 Winter Seasons. J Infect Dis. 2006;
193(1):54–58.
69. Imaz MS, Sequeira MD, Videla C, et al. Clinical and epidemiologic characteristics of respiratory
syncytial virus subgroups A and B infections in Santa Fe, Argentina. J Med Virol. 2000; 61(1):76–
80.
70. Snell NJC. Ribavirin - current status of a broad spectrum antiviral agent. Expert Opin
Pharmacother. 2001; 2(8):1317–1324.
71. Shah DP, Ghantoji SS, Shah JN, et al. Impact of aerosolized ribavirin on mortality in 280
allogeneic haematopoietic stem cell transplant recipients with respiratory syncytial virus
infections. J Antimicrob Chemother. 2013; 68(8):1872–1880.
72. McColl MD, Corser RB, Bremner J, Chopra R. Respiratory syncytial virus infection in adult BMT
recipients: effective therapy with short duration nebulised ribavirin. Bone Marrow Transplant.
1998; 21(4):423.
73. Moler FW, Steinhart CM, Ohmit SE, Stidham GL. Effectiveness of ribavirin in otherwise well
infants with respiratory syncytial virus-associated respiratory failure. J Pediatr. 1996; 128(3):422–
428.
74. Meert KL, Sarnaik AP, Gelmini MJ, Lieh-Lai MW. Aerosolized ribavirin in mechanically
ventilated children with respiratory syncytial virus lower respiratory tract disease: A prospective,
double-blind, randomized trial. Crit Care Med. 1994; 22(4):566.
75. Krilov LR. Respiratory syncytial virus disease: update on treatment and prevention. Expert Rev
Anti Infect Ther. 2011; 9(1):27–32.
76. Huang K, Incognito L, Cheng X, Ulbrandt ND, Wu H. Respiratory Syncytial Virus-Neutralizing
Monoclonal Antibodies Motavizumab and Palivizumab Inhibit Fusion. J Virol. 2010;
84(16):8132–8140.

26
77. Wang D, Bayliss S, Meads C. Palivizumab for immunoprophylaxis of respiratory syncytial virus
(RSV) bronchiolitis in high-risk infants and young children: a systematic review and additional
economic modelling of subgroup analyses. Health Technol Assess Winch Engl. 2011; 15(5):iii–
124.
78. Adams O, Bonzel L, Kovacevic A, Mayatepek E, Hoehn T, Vogel M. Palivizumab-Resistant
Human Respiratory Syncytial Virus Infection in Infancy. Clin Infect Dis. 2010; 51(2):185–188.
79. Kim HW, Canchola JG, Brandt CD, et al. RESPIRATORY SYNCYTIAL VIRUS DISEASE IN
INFANTS DESPITE PRIOR ADMINISTRATION OF ANTIGENIC INACTIVATED
VACCINE. Am J Epidemiol. 1969; 89(4):422–434.
80. Schlender J, Zimmer G, Herrler G, Conzelmann K-K. Respiratory Syncytial Virus (RSV) Fusion
Protein Subunit F2, Not Attachment Protein G, Determines the Specificity of RSV Infection. J
Virol. 2003; 77(8):4609–4616.
81. Bossert B, Conzelmann K-K. Respiratory Syncytial Virus (RSV) Nonstructural (NS) Proteins as
Host Range Determinants: a Chimeric Bovine RSV with NS Genes from Human RSV Is
Attenuated in Interferon-Competent Bovine Cells. J Virol. 2002; 76(9):4287–4293.
82. Morris JA, R. E. Blount J, Savage RE. Recovery of Cytopathogenic Agent from Chimpanzees with
Goryza. Proc Soc Exp Biol Med. 1956; 92(3):544–549.
83. Belshe RB, Richardson LS, London WT, et al. Experimental respiratory syncytial virus infection
of four species of primates. J Med Virol. 1977; 1(3):157–162.
84. Whitehead SS, Bukreyev A, Teng MN, et al. Recombinant Respiratory Syncytial Virus Bearing a
Deletion of either the NS2 or SH Gene Is Attenuated in Chimpanzees. J Virol. 1999; 73(4):3438–
3442.
85. Hancock GE, Smith JD, Heers KM. Serum Neutralizing Antibody Titers of Seropositive
Chimpanzees Immunized with Vaccines Coformulated with Natural Fusion and Attachment
Proteins of Respiratory Syncytial Virus. J Infect Dis. 2000; 181(5):1768–1771.
86. McLellan JS, Chen M, Joyce MG, et al. Structure-Based Design of a Fusion Glycoprotein Vaccine
for Respiratory Syncytial Virus. Science. 2013; 342(6158):592–598.
87. Murthy KK, Salas MT, Carey KD, Patterson JL. Baboon as a nonhuman primate model for
vaccine studies. Vaccine. 2006; 24(21):4622–4624.
88. Papin JF, Wolf RF, Kosanke SD, et al. Infant baboons infected with respiratory syncytial virus
develop clinical and pathological changes that parallel those of human infants. Am J Physiol-Lung
Cell Mol Physiol. 2013; 304(8):L530–L539.
89. Prince GA, Jenson AB, Horswood RL, Camargo E, Chanock RM. The pathogenesis of respiratory
syncytial virus infection in cotton rats. Am J Pathol. 1978; 93(3):771–791.
90. Prince GA, Horswood RL, Berndt J, Suffin SC, Chanock RM. Respiratory syncytial virus
infection in inbred mice. Infect Immun. 1979; 26(2):764–766.

27
91. Prince GA, Prieels JP, Slaoui M, Porter DD. Pulmonary lesions in primary respiratory syncytial
virus infection, reinfection, and vaccine-enhanced disease in the cotton rat (Sigmodon hispidus).
Lab Investig J Tech Methods Pathol. 1999; 79(11):1385–1392.
92. Stokes KL, Chi MH, Sakamoto K, et al. Differential Pathogenesis of Respiratory Syncytial Virus
Clinical Isolates in BALB/c Mice. J Virol. 2011; 85(12):5782–5793.
93. Boukhvalova MS, Blanco JCG. The Cotton Rat Sigmodon Hispidus Model of Respiratory
Syncytial Virus Infection. Chall Oppor Respir Syncytial Virus Vaccines [Internet]. Springer,
Berlin, Heidelberg; 2013 [cited 2018 Jan 17]. p. 347–358. Available from:
https://link.springer.com/chapter/10.1007/978-3-642-38919-1_17
94. Bonville CA, Bennett NJ, Koehnlein M, et al. Respiratory dysfunction and proinflammatory
chemokines in the pneumonia virus of mice (PVM) model of viral bronchiolitis. Virology. 2006;
349(1):87–95.
95. Rosenberg HF, Domachowske JB. Pneumonia virus of mice: severe respiratory infection in a
natural host. Immunol Lett. 2008; 118(1):6–12.
96. Sacco RE, Durbin RK, Durbin JE. Animal models of respiratory syncytial virus infection and
disease. Curr Opin Virol. 2015; 13:117–122.
97. Elvander M, Vilcek S, Baule C, Uttenthal A, Ballagi-Pord√°ny A, Bel√°k S. Genetic and antigenic
analysis of the G attachment protein of bovine respiratory syncytial virus strains. J Gen Virol.
1998; 79(12):2939–2946.
98. Valarcher J-F, Taylor G. Bovine respiratory syncytial virus infection. Vet Res. 2007; 38(2):153–
180.
99. Choi HK, Finkbeiner WE, Widdicombe JH. A comparative study of mammalian tracheal mucous
glands. J Anat. 2000; 197(3):361–372.
100. Casteleyn C, Breugelmans S, Simoens P, Van den Broeck W. The tonsils revisited: review of the
anatomical localization and histological characteristics of the tonsils of domestic and laboratory
animals. Clin Dev Immunol. 2011; 2011.
101. Larios Mora A, Detalle L, Van Geelen A, et al. Kinetics of Respiratory Syncytial Virus (RSV)
Memphis Strain 37 (M37) Infection in the Respiratory Tract of Newborn Lambs as an RSV
Infection Model for Human Infants. PLoS ONE [Internet]. 2015 [cited 2016 Jan 7]; 10(12).
Available from: http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4671688/
102. Derscheid RJ, Geelen A van, McGill JL, et al. Human Respiratory Syncytial Virus Memphis 37
Grown in HEp-2 Cells Causes more Severe Disease in Lambs than Virus Grown in Vero Cells.
Viruses. 2013; 5(11):2881–2897.
103. Grosz DD, Geelen A van, Gallup JM, Hostetter SJ, Derscheid RJ, Ackermann MR. Sucrose
stabilization of Respiratory Syncytial Virus (RSV) during nebulization and experimental infection.
BMC Res Notes. 2014; 7:158.

28
104. Derscheid RJ, Geelen A van, Gallup JM, et al. Human Respiratory Syncytial Virus Memphis 37
Causes Acute Respiratory Disease in Perinatal Lamb Lung. BioResearch Open Access. 2014;
3(2):60–69.
105. Alcorn DG, Adamson TM, Maloney JE, Robinson PM. A morphologic and morphometric analysis
of fetal lung development in the sheep. Anat Rec. 1981; 201(4):655–667.
106. Plopper CG, Mariassy AT, Lollini LO. Structure as Revealed by Airway Dissection. Am Rev
Respir Dis. 1983; 128(2P2):S4–S7.
107. Zosky GR, Sly PD. Animal models of asthma. Clin Exp Allergy. 2007; 37(7):973–988.
108. Scheerlinck J-PY, Snibson KJ, Bowles VM, Sutton P. Biomedical applications of sheep models:
from asthma to vaccines. Trends Biotechnol. 2008; 26(5):259–266.
109. Liu P, Jamaluddin M, Li K, Garofalo RP, Casola A, Brasier AR. Retinoic Acid-Inducible Gene I
Mediates Early Antiviral Response and Toll-Like Receptor 3 Expression in Respiratory Syncytial
Virus-Infected Airway Epithelial Cells. J Virol. 2007; 81(3):1401–1411.
110. Scagnolari C, Midulla F, Pierangeli A, et al. Gene Expression of Nucleic Acid-Sensing Pattern
Recognition Receptors in Children Hospitalized for Respiratory Syncytial Virus-Associated Acute
Bronchiolitis. Clin Vaccine Immunol. 2009; 16(6):816–823.
111. Rudd BD, Burstein E, Duckett CS, Li X, Lukacs NW. Differential Role for TLR3 in Respiratory
Syncytial Virus-Induced Chemokine Expression. J Virol. 2005; 79(6):3350–3357.
112. Murawski MR, Bowen GN, Cerny AM, et al. Respiratory Syncytial Virus Activates Innate
Immunity through Toll-Like Receptor 2. J Virol. 2009; 83(3):1492–1500.
113. Kurt-Jones EA, Popova L, Kwinn L, et al. Pattern recognition receptors TLR4 and CD14 mediate
response to respiratory syncytial virus. Nat Immunol. 2000; 1(5):398.
114. Haynes LM, Moore DD, Kurt-Jones EA, Finberg RW, Anderson LJ, Tripp RA. Involvement of
Toll-Like Receptor 4 in Innate Immunity to Respiratory Syncytial Virus. J Virol. 2001;
75(22):10730–10737.
115. Smyth RL, Mobbs KJ, O’Hea U, Ashby D, Hart CA. Respiratory syncytial virus bronchiolitis:
Disease severity, interleukin-8, and virus genotype*. Pediatr Pulmonol. 2002; 33(5):339–346.
116. Adams O, Weis J, Jasinska K, Vogel M, Tenenbaum T. Comparison of human metapneumovirus,
respiratory syncytial virus and Rhinovirus respiratory tract infections in young children admitted
to hospital. J Med Virol. 2015; 87(2):275–280.
117. Noah TL, Becker S. Chemokines in Nasal Secretions of Normal Adults Experimentally Infected
with Respiratory Syncytial Virus. Clin Immunol. 2000; 97(1):43–49.
118. John AE, Berlin AA, Lukacs NW. Respiratory syncytial virus-induced CCL5/RANTES
contributes to exacerbation of allergic airway inflammation. Eur J Immunol. 2003; 33(6):1677–
1685.

29
119. Murai H, Terada A, Mizuno M, et al. IL-10 and RANTES are Elevated in Nasopharyngeal
Secretions of Children with Respiratory Syncytial Virus Infection. Allergol Int. 2007; 56(2):157–
163.
120. McNamara PS, Flanagan BF, Hart CA, Smyth RL. Production of Chemokines in the Lungs of
Infants with Severe Respiratory Syncytial Virus Bronchiolitis. J Infect Dis. 2005; 191(8):1225–
1232.
121. Roe MFE, Bloxham DM, Cowburn AS, O’Donnell DR. Changes in helper lymphocyte chemokine
receptor expression and elevation of IP-10 during acute respiratory syncytial virus infection in
infants. Pediatr Allergy Immunol. 2011; 22(2):229–234.
122. Nakayama T, Sonoda S, Urano T, Sasaki K, Maehara N, Makino S. Detection of alpha-interferon
in nasopharyngeal secretions and sera in children infected with respiratory syncytial virus. Pediatr
Infect Dis J. 1993; 12(11):925.
123. Spann KM, Tran K-C, Chi B, Rabin RL, Collins PL. Suppression of the Induction of Alpha, Beta,
and Gamma Interferons by the NS1 and NS2 Proteins of Human Respiratory Syncytial Virus in
Human Epithelial Cells and Macrophages. J Virol. 2004; 78(8):4363–4369.
124. Barik S. Respiratory Syncytial Virus Mechanisms to Interfere with Type 1 Interferons. Chall
Oppor Respir Syncytial Virus Vaccines [Internet]. Springer, Berlin, Heidelberg; 2013 [cited 2018
Jan 22]. p. 173–191. Available from: https://link.springer.com/chapter/10.1007/978-3-642-38919-
1_9
125. Spann KM, Tran KC, Collins PL. Effects of Nonstructural Proteins NS1 and NS2 of Human
Respiratory Syncytial Virus on Interferon Regulatory Factor 3, NF-κB, and Proinflammatory
Cytokines. J Virol. 2005; 79(9):5353–5362.
126. Lo MS, Brazas RM, Holtzman MJ. Respiratory Syncytial Virus Nonstructural Proteins NS1 and
NS2 Mediate Inhibition of Stat2 Expression and Alpha/Beta Interferon Responsiveness. J Virol.
2005; 79(14):9315–9319.
127. Ling Z, Tran KC, Teng MN. Human Respiratory Syncytial Virus Nonstructural Protein NS2
Antagonizes the Activation of Beta Interferon Transcription by Interacting with RIG-I. J Virol.
2009; 83(8):3734–3742.
128. Eichinger KM, Egaña L, Orend JG, et al. Alveolar macrophages support interferon gamma-
mediated viral clearance in RSV-infected neonatal mice. Respir Res. 2015; 16:122.
129. Selvaggi C, Pierangeli A, Fabiani M, et al. Interferon lambda 1–3 expression in infants
hospitalized for RSV or HRV associated bronchiolitis. J Infect. 2014; 68(5):467–477.
130. Conner GE, Salathe M, Forteza R. Lactoperoxidase and Hydrogen Peroxide Metabolism in the
Airway. Am J Respir Crit Care Med. 2002; 166(supplement_1):S57–S61.
131. Wijkstrom-Frei C, El-Chemaly S, Ali-Rachedi R, et al. Lactoperoxidase and Human Airway Host
Defense. Am J Respir Cell Mol Biol. 2003; 29(2):206–212.
132. LeVine AM, Gwozdz J, Stark J, Bruno M, Whitsett J, Korfhagen T. Surfactant protein-A enhances
respiratory syncytial virus clearance in vivo. J Clin Invest. 1999; 103(7):1015–1021.

30
133. Sano H, Nagai K, Tsutsumi H, Kuroki Y. Lactoferrin and surfactant protein A exhibit distinct
binding specificity to F protein and differently modulate respiratory syncytial virus infection. Eur J
Immunol. 2003; 33(10):2894–2902.
134. LeVine AM, Elliott J, Whitsett JA, et al. Surfactant Protein-D Enhances Phagocytosis and
Pulmonary Clearance of Respiratory Syncytial Virus. Am J Respir Cell Mol Biol. 2004;
31(2):193–199.
135. Kerr MH, Paton JY. Surfactant Protein Levels in Severe Respiratory Syncytial Virus Infection.
Am J Respir Crit Care Med. 1999; 159(4):1115–1118.
136. Kota S, Sabbah A, Chang TH, et al. Role of Human β-Defensin-2 during Tumor Necrosis Factor-
α/NF-κB-mediated Innate Antiviral Response against Human Respiratory Syncytial Virus. J Biol
Chem. 2008; 283(33):22417–22429.
137. Barlow PG, Beaumont PE, Cosseau C, et al. The Human Cathelicidin LL-37 Preferentially
Promotes Apoptosis of Infected Airway Epithelium. Am J Respir Cell Mol Biol. 2010; 43(6):692–
702.
138. O’Donnell DR, Carrington D. Peripheral blood lymphopenia and neutrophilia in children with
severe respiratory syncytial virus disease. Pediatr Pulmonol. 2002; 34(2):128–130.
139. Smith P, Wang S-Z, Dowling K, Forsyth K. Leucocyte populations in respiratory syncytial virus-
induced bronchiolitis. J Paediatr Child Health. 2001; 37(2):146–151.
140. Lukens MV, Pol AC van de, Coenjaerts FEJ, et al. A Systemic Neutrophil Response Precedes
Robust CD8+ T-Cell Activation during Natural Respiratory Syncytial Virus Infection in Infants. J
Virol. 2010; 84(5):2374–2383.
141. Emboriadou M, Hatzistilianou M, Magnisali C, et al. Human Neutrophil Elastase in RSV
Bronchiolitis. Ann Clin Lab Sci. 2007; 37(1):79–84.
142. Cortjens B, De Boer OJ, De Jong R, et al. Neutrophil extracellular traps cause airway obstruction
during respiratory syncytial virus disease. J Pathol. 2016; 238(3):401–411.
143. Xu H, Schultze-Mosgau A, Agic A, Diedrich K, Taylor RN, Hornung D. Regulated upon
activation, normal T cell expressed and secreted (RANTES) and monocyte chemotactic protein 1
in follicular fluid accumulate differentially in patients with and without endometriosis undergoing
in vitro fertilization. Fertil Steril. 2006; 86(6):1616–1620.
144. Dimova-Yaneva D, Russell D, Main M, Brooker RJ, Helms PJ. Eosinophil activation and
cysteinyl leukotriene production in infants with respiratory syncytial virus bronchiolitis. Clin Exp
Allergy. 2004; 34(4):555–558.
145. Okamoto N, Ikeda M, Okuda M, et al. Increased Eosinophilic Cationic Protein in Nasal Fluid in
Hospitalized Wheezy Infants with RSV Infection. Allergol Int. 2011; 60(4):467–472.
146. Everard ML, Swarbrick A, Wrightham M, et al. Analysis of cells obtained by bronchial lavage of
infants with respiratory syncytial virus infection. Arch Dis Child. 1994; 71(5):428–432.

31
147. McNamara PS, Ritson P, Selby A, Hart CA, Smyth RL. Bronchoalveolar lavage cellularity in
infants with severe respiratory syncytial virus bronchiolitis. Arch Dis Child. 2003; 88(10):922–
926.
148. Cirino NM, Panuska JR, Villani A, et al. Restricted replication of respiratory syncytial virus in
human alveolar macrophages. J Gen Virol. 1993; 74(8):1527–1537.
149. Midulla F, Villani A, Panuska JR, et al. Respiratory Syncytial Virus Lung Infection in Infants:
Immunoregulatory Role of Infected Alveolar Macrophages. J Infect Dis. 1993; 168(6):1515–1519.
150. Meyerholz DK, Grubor B, Fach SJ, et al. Reduced clearance of respiratory syncytial virus
infection in a preterm lamb model. Microbes Infect. 2004; 6(14):1312–1319.
151. Gill MA, Long K, Kwon T, et al. Differential Recruitment of Dendritic Cells and Monocytes to
Respiratory Mucosal Sites in Children with Influenza Virus or Respiratory Syncytial Virus
Infection. J Infect Dis. 2008; 198(11):1667–1676.
152. Gill MA, Palucka AK, Barton T, et al. Mobilization of Plasmacytoid and Myeloid Dendritic Cells
to Mucosal Sites in Children with Respiratory Syncytial Virus and Other Viral Respiratory
Infections. J Infect Dis. 2005; 191(7):1105–1115.
153. Kerrin A, Fitch P, Errington C, et al. Differential lower airway dendritic cell patterns may reveal
distinct endotypes of RSV bronchiolitis. Thorax. 2017; 72(7):620–627.
154. Welliver TP, Garofalo RP, Hosakote Y, et al. Severe Human Lower Respiratory Tract Illness
Caused by Respiratory Syncytial Virus and Influenza Virus Is Characterized by the Absence of
Pulmonary Cytotoxic Lymphocyte Responses. J Infect Dis. 2007; 195(8):1126–1136.
155. Raes M, Peeters V, Alliet P, et al. Peripheral blood T and B lymphocyte subpopulations in infants
with acute respiratory syncytial virus brochiolitis. Pediatr Allergy Immunol. 1997; 8(2):97–102.
156. Ayukawa H, Matsubara T, Kaneko M, Hasegawa M, Ichiyama T, Furukawa S. Expression of
CTLA-4 (CD152) in peripheral blood T cells of children with influenza virus infection including
encephalopathy in comparison with respiratory syncytial virus infection. Clin Exp Immunol. 2004;
137(1):151–155.
157. Mayumi Koga SF Takashi Matsuoka, Tomoyo Matsubara, Kumiko Katayama. Different
Expression of ICAM-1 and LFA-1 Alpha by Peripheral Leukocytes During Respiratory Syncytial
Virus and Influenza Virus Infection in Young Children. Scand J Infect Dis. 2000; 32(1):7–11.
158. Heidema J, Lukens MV, Maren WWC van, et al. CD8+ T Cell Responses in Bronchoalveolar
Lavage Fluid and Peripheral Blood Mononuclear Cells of Infants with Severe Primary Respiratory
Syncytial Virus Infections. J Immunol. 2007; 179(12):8410–8417.
159. Berger A. Th1 and Th2 responses: what are they? BMJ. 2000; 321(7258):424.
160. Openshaw PJM, Tregoning JS. Immune Responses and Disease Enhancement during Respiratory
Syncytial Virus Infection. Clin Microbiol Rev. 2005; 18(3):541–555.

32
161. Legg JP, Hussain IR, Warner JA, Johnston SL, Warner JO. Type 1 and Type 2 Cytokine
Imbalance in Acute Respiratory Syncytial Virus Bronchiolitis. Am J Respir Crit Care Med. 2003;
168(6):633–639.
162. Román M, Calhoun WJ, Hinton KL, et al. Respiratory Syncytial Virus Infection in Infants Is
Associated with Predominant Th-2-like Response. Am J Respir Crit Care Med. 1997; 156(1):190–
195.
163. Bermejo-Martin JF, Garcia-Arevalo MC, Lejarazu ROD, et al. Predominance of Th2 cytokines,
CXC chemokines and innate immunity mediators at the mucosal level during severe respiratory
syncytial virus infection in children. Eur Cytokine Netw. 2007; 18(3):45–50.
164. Welliver RC, Kaul TN, Putnam TI, Sun M, Riddlesberger K, Ogra PL. The antibody response to
primary and secondary infection with respiratory syncytial virus: Kinetics of class-specific
responses. J Pediatr. 1980; 96(5):808–813.
165. McGill A, Greensill J, Marsh R, Craft A w., Toms G l. Detection of human respiratory syncytial
virus genotype specific antibody responses in infants. J Med Virol. 2004; 74(3):492–498.
166. Walsh E, Falsey AR. Humoral and Mucosal Immunity in Protection from Natural Respiratory
Syncytial Virus Infection in Adults. J Infect Dis. 2004; 190(2):373–378.
167. Gonzalez ML, Jozwik AA, Habibi MS, Openshaw PJM, Chiu C. quality of antigen-specific B-cell
responses as a correlate of protection against Rsv: 569. Immunology [Internet]. 2014 [cited 2018
Feb 16]; 143. Available from: https://insights.ovid.com/immunology/immu/2014/12/002/quality-
antigen-specific-cell-responses-correlate/193/00004227
168. Tsutsumi H, Matsuda K, Yamazaki H, Ogra PL, Chiba S. Different kinetics of antibody responses
between IgA and IgG classes in nasopharyngeal secretion in infants and children during primary
respiratory syncytial virus infection. Pediatr Int. 1995; 37(4):464–468.
169. Habibi MS, Jozwik A, Makris S, et al. Impaired Antibody-mediated Protection and Defective IgA
B-Cell Memory in Experimental Infection of Adults with Respiratory Syncytial Virus. Am J
Respir Crit Care Med. 2015; 191(9):1040–1049.
170. Bartlett JG. Diagnostic Tests for Agents of Community-Acquired Pneumonia. Clin Infect Dis.
2011; 52(suppl_4):S296–S304.
171. Falsey AR, Becker KL, Swinburne AJ, et al. Bacterial Complications of Respiratory Tract Viral
Illness: A Comprehensive Evaluation. J Infect Dis. 2013; 208(3):432–441.
172. Korppi M, Leinonen M, Koskela M, Mäkelä PH, Launiala K. Bacterial coinfection in children
hospitalized with respiratory syncytial virus infections. Pediatr Infect Dis J. 1989; 8(10):687–692.
173. Madhi SA, Klugman KP. A role for Streptococcus pneumoniae in virus-associated pneumonia. Nat
Med. 2004; 10(8):811–813.
174. DeLeo FR, Musser JM. Axis of Coinfection Evil. J Infect Dis. 2010; 201(4):488–490.

33
175. Hendricks MR, Lashua LP, Fischer DK, et al. Respiratory syncytial virus infection enhances
Pseudomonas aeruginosa biofilm growth through dysregulation of nutritional immunity. Proc Natl
Acad Sci. 2016; 113(6):1642–1647.
176. Smith AM, Adler FR, Ribeiro RM, et al. Kinetics of Coinfection with Influenza A Virus and
Streptococcus pneumoniae. PLOS Pathog. 2013; 9(3):e1003238.
177. Hament J-M, Aerts PC, Fleer A, et al. Enhanced Adherence of Streptococcus pneumoniae to
Human Epithelial Cells Infected with Respiratory Syncytial Virus. Pediatr Res. 2004; 55(6):972–
978.
178. Avadhanula V, Wang Y, Portner A, Adderson E. Nontypeable Haemophilus influenzae and
Streptococcus pneumoniae bind respiratory syncytial virus glycoprotein. J Med Microbiol. 2007;
56(9):1133–1137.
179. McCullers JA. Insights into the Interaction between Influenza Virus and Pneumococcus. Clin
Microbiol Rev. 2006; 19(3):571–582.
180. McCullers JA. The co-pathogenesis of influenza viruses with bacteria in the lung. Nat Rev
Microbiol. 2014; 12(4):252–262.
181. Loosli CG, Stinson SF, Ryan DP, Hertweck MS, Hardy JD, Serebrin R. The destruction of type 2
pneumocytes by airborne influenza PR8-A virus; its effect on surfactant and lecithin content of the
pneumonic lesions of mice. Chest. 1975; 67(2 Suppl):7S-14S.
182. Levandowski RA, Gerrity TR, Garrard CS. Modifications of lung clearance mechanisms by acute
influenza A infection. Transl Res. 1985; 106(4):424–427.
183. Nguyen DT, Louwen R, Elberse K, et al. Streptococcus pneumoniae Enhances Human Respiratory
Syncytial Virus Infection In Vitro and In Vivo. PLOS ONE. 2015; 10(5):e0127098.
184. Ralston SL, Lieberthal AS, Meissner HC, et al. Clinical Practice Guideline: The Diagnosis,
Management, and Prevention of Bronchiolitis. Pediatrics. 2014; 134(5):e1474–e1502.
185. Detalle L, Stohr T, Palomo C, et al. Generation and Characterization of ALX-0171, a Potent Novel
Therapeutic Nanobody for the Treatment of Respiratory Syncytial Virus Infection. Antimicrob
Agents Chemother. 2016; 60(1):6–13.
186. Sivapalasingam S, Caballero-Perez D, Houghton M, et al. Phase 1 Study Evaluating Safety,
Tolerability, Pharmacokinetics and Immunogenicity of REGN2222 in Healthy Adults: A New
Human Monoclonal RSV-F Antibody for RSV Prevention. Open Forum Infect Dis [Internet]. 2015
[cited 2018 Feb 13]; 2(suppl_1). Available from:
https://academic.oup.com/ofid/article/2/suppl_1/912/2635143
187. Yan D, Lee S, Thakkar VD, Luo M, Moore ML, Plemper RK. Cross-resistance mechanism of
respiratory syncytial virus against structurally diverse entry inhibitors. Proc Natl Acad Sci. 2014;
111(33):E3441–E3449.
188. DeVincenzo JP, McClure MW, Symons JA, et al. Activity of Oral ALS-008176 in a Respiratory
Syncytial Virus Challenge Study. N Engl J Med. 2015; 373(21):2048–2058.

34
189. Chapman J, Abbott E, Alber DG, et al. RSV604, a Novel Inhibitor of Respiratory Syncytial Virus
Replication. Antimicrob Agents Chemother. 2007; 51(9):3346–3353.
190. Bailly B, Richard C-A, Sharma G, et al. Targeting human respiratory syncytial virus transcription
anti-termination factor M2-1 to inhibit in vivo viral replication. Sci Rep. 2016; 6:25806.
191. Li Y, To J, Verdià-Baguena C, et al. Inhibition of the Human Respiratory Syncytial Virus Small
Hydrophobic Protein and Structural Variations in a Bicelle Environment. J Virol. 2014;
88(20):11899–11914.
192. F. Rosenberg H, B. Domachowske J. Inflammatory Responses to Respiratory Syncytial Virus
(RSV) Infection and the Development of Immunomodulatory Pharmacotherapeutics [Internet].
2012 [cited 2018 Feb 13]. Available from:
http://orst.library.ingentaconnect.com/content/ben/cmc/2012/00000019/00000010/art00002
193. Bonville CA, Easton AJ, Rosenberg HF, Domachowske JB. Altered Pathogenesis of Severe
Pneumovirus Infection in Response to Combined Antiviral and Specific Immunomodulatory
Agents. J Virol. 2003; 77(2):1237–1244.
194. Miller AL, Gerard C, Schaller M, Gruber AD, Humbles AA, Lukacs NW. Deletion of CCR1
Attenuates Pathophysiologic Responses during Respiratory Syncytial Virus Infection. J Immunol.
2006; 176(4):2562–2567.
195. Welliver RC, Hintz KH, Glori M, Welliver RC. Zileuton Reduces Respiratory Illness and Lung
Inflammation, during Respiratory Syncytial Virus Infection, in Mice. J Infect Dis. 2003;
187(11):1773–1779.
196. Bisgaard H. A Randomized Trial of Montelukast in Respiratory Syncytial Virus Postbronchiolitis.
Am J Respir Crit Care Med. 2003; 167(3):379–383.
197. Meyerholz DK, Gallup JM, Lazic T, De Macedo MMA, Lehmkuhl HD, Ackermann MR.
Pretreatment with Recombinant Human Vascular Endothelial Growth Factor Reduces Virus
Replication and Inflammation in a Perinatal Lamb Model of Respiratory Syncytial Virus Infection.
Viral Immunol. 2007; 20(1):188–196.

35
: STREPTOCOCCUS PNEUMONIAE INFECTION IN
RESPIRATORY SYNCYTIAL VIRUS INFECTED NEONATAL LAMBS
A paper submitted to Emerging Microbes and Infections
Sarhad Alnajjar1,2,3, Panchan Sitthicharoenchai1, Jack Gallup1, Mark Ackermann3,5, David
Verhoeven4*
1Departments of Veterinary Pathology and 4Veterinary Microbiology and Preventative
Medicine, College of Veterinary Medicine, Iowa State University, Ames, Iowa
2Department of Veterinary Pathology, College of Veterinary Medicine, Baghdad
University, Baghdad, Iraq
3Department of Biomedical Sciences, Carlson College of Veterinary Medicine, Oregon
State University
5LambCure, LLC, Corvallis, Oregon
Abstract
Background: Respiratory syncytial virus (RSV) is the primary cause of viral bronchiolitis
resulting in hospitalization and the most frequent cause of secondary respiratory bacterial infection
especially by Streptococcus pneumoniae (Spn) in infants. While murine studies have demonstrated
enhanced morbidity during a viral/bacterial co-infection, human meta-studies have conflicting
results. Moreover, less is known about the pathogenesis of Spn serotype 22F and especially the
co-pathologies between RSV and Spn dual infections.

36
Objective: The objective of this study was to determine interactions and mechanisms that
contribute to co-pathogen-induced morbidity using a neonatal lamb model naturally permissive to
infection by both pathogens.
Methods: Colostrum deprived lambs (aged 3-5 days) were randomly divided into four
groups. Two of the groups were nebulized with RSV M37 (1.27 x 107 IFFU/mL), and the other
two groups nebulized with cell–conditioned mock media. At day 3 post-infection, one RSV group
(RSV/Spn) and one mock-nebulized group (Spn only) were infected with (2x106 cfu of Spn)
intratracheally. At day 6 post-infection all lambs were humanely euthanized, and bacterial/viral
pathogeneses were assessed by culture, viral infectious focus forming unit assay, RT-qPCR,
immunohistochemistry, and histopathology.
Results: Lambs dually infected with RSV and Spn had higher RSV titers, but lower Spn
than the other comparable groups. Additionally, lung lesions were observed to be more intense in
the RSV/Spn group characterized by increased interalveolar wall thickness accompanied by
neutrophil and lymphocyte infiltration.
Conclusions: Despite lower Spn in lungs, lambs co-infected with RSV demonstrated greater
morbidity and tissue histopathology. Thus, the perception of enhanced disease severity may be
due to observed lesion development rather than elevated bacterial pathogenesis.
Introduction
Respiratory Syncytial Virus is one of the leading cause of severe lower respiratory infection
in infants under the age of five leading to 600,000 deaths worldwide [1]. RSV is a member of the
pneumoviridae family that infects all infants by the age of two years [2]. Although mild to

37
moderate upper respiratory tract infection is the most common form of infection, severe lower
respiratory tract can develop leading to severe pneumonia and/or bronchiolitis that leads to
hospitalization and sometimes death [3–5]. Lower respiratory tract infection increases the
susceptibility to secondary bacterial infection(s) leading to severe and life-threatening pneumonia
[6]. Streptococcus pneumoniae (Spn) is one of the most common bacterial infections that occurs
concurrently with respiratory viruses such as influenza and RSV [7].
Spn is a Gram-positive facultative anaerobic bacterial pathogen that causes invasive disease
including sepsis, meningitis, and pneumonia. Similar to RSV, Spn cause severe illness and presents
with higher incidence in both children and the elderly worldwide [8]. Pneumococcal pneumonia
is one of the leading causes of bacterial pneumonia in children worldwide, responsible for about
11% of all deaths in children under the age of five (700,000-1 million every year). Most of these
deaths occur in developing countries [9]. Spn vaccines are effective in reducing the incidence of
pneumonia caused by the serotypes contained in the vaccine [10]. However, the emergence of
non-vaccine serotypes and persistence of antibiotic-resistant Spn such as serotype 19A highlights
the importance of more investigation into Spn pathogenesis and therapy. Since Spn has an essential
role in secondary bacterial infections following viral pneumonia or viral-bacterial co-infection
[7,11], animal modeling for the purpose of understanding viral-bacterial co-infections are crucial
to investigating therapeutics that combat both. Moreover, most studies have concentrated on
influenza and Spn co-infections but mainly in murine models with few mechanistic studies done
in humans other than calculation of frequencies of co-infections with these two pathogens. Despite
the importance of RSV/Spn co-infections, far fewer studies have been done in this area as
compared to influenza/Spn. Furthermore, less is known about emergent serotype 22F pathogenesis
[12,13]. We have extensively used a neonatal lamb model to mimic RSV lower respiratory tract

38
infection in infants as a preclinical model to evaluate the efficacy of new therapeutics [14,15] and
to understand RSV pathogenesis [16–18]. Sheep are also permissive to Spn infection and have
served as a model of Spn sepsis that appears to manifest clinical signs similar to human infection
[19,20]. Thus, in the present study, we investigated a co-infection of neonatal lambs using
RSV/Spn as an alternative large animal model to better understand viral/bacterial co-infections in
the young with the objective of gaining more insight into serotype 22F pathogenesis in the lung.
Material and Methods
Experimental Design
Animals: A total of 20, 2-3 day-old, colostrum-deprived lambs, were randomly divided into
four groups with 5 animals per group: RSV only, RSV-Spn co-infection, Spn only, and uninfected
control. Animal use was approved by the Institutional Animal Care and Use Committee of Iowa
State University. Two groups were nebulized with RSV M37 (1.27x107 IFFU/mL) on day 0. One
of the RSV infected groups was injected intratracheally with 2 ml normal saline as a mock Spn
infection (RSV group), while the second RSV-infected group was injected intratracheally with 2
ml solution containing Spn serotype 22F (2x106 CFU/ml) 3 days post-RSV nebulization (RSV-
Spn group). The other two groups were nebulized with cell-conditioned mock media containing
20% sucrose at day 0 and injected intratracheally with either normal saline (control group) or
solution containing Spn (2x106 CFU/ml) at day 3 post nebulization (Spn group). At day 6 post-
RSV infection, all lambs were humanely euthanized. Autopsy was performed to evaluate the
macroscopic lung lesions. Lung samples were collected including sterile lung tissue for bacterial
isolation, frozen lung sample for RT-qPCR, bronchioalveolar lavage fluid (BALF) from right
caudal lung lobe for RSV infectious focus forming unit (IFFU) assay and RT-qPCR, and lung

39
pieces from different lobes were fixed in 10% neutral buffered formalin for histological
assessment.
Infectious Agents
Lambs were infected with RSV strain M37, purchased from Meridian BioSciences
(Memphis, TN, USA). This strain is a wild type A RSV isolated from the respiratory secretions of
an infant hospitalized for bronchiolitis [21,22]. M37 was grown in HELA cells and stored at -80°C
in media containing 20% sucrose [23]. 6 mL of 1.27 x 107 IFFU/mL in media containing 20%
sucrose or cell-conditioned mock media (also containing 20% sucrose) was nebulized using PARI
LC Sprint™ nebulizers to each lamb over the course of 25-30 minutes resulting in the total
inhalation of about 3 mL by each lamb [23]. Spn serotype 22F was grown overnight at 37°C in
Todd Hewitt media containing 2% yeast extract, 50 g/ml of gentamicin, and 10% bovine serum.
Colony forming units (CFUs) were calculated by OD600 with confirmation by dilution plating on
Tryptic Soy Agar (TSA) plates with 5% sheep blood containing gentamicin.
Lung RSV viral and Spn bacterial titers
BALF collected from right caudal lobe at necropsy was used to evaluate RSV IFFU. BALF
were spun for 5 minutes at 3,000g to pellet large debris. Supernatants were then spun through 0.45
am Costar SPIN-X filters (microcentrifuge 15,600g) for 5 minutes. The resulting BALF samples
were applied to HELA cells grown to 70% confluence in 12-well culture plates (Fisher Scientific,
Hanover Park, IL) at full strength, and three serial dilutions (1:10, 1:100, and 1:1000); all samples
were tested in triplicate to determine the viral titer. Lung tissue samples were used to determine
Spn titer. Lung tissue samples were placed in 500 μl of sterile PBS and were mechanically
homogenized. Lung homogenates were pelleted at 100x g, for 5 minutes. Supernatants were
serially diluted and applied to 5% sheep blood TSA plates containing gentamycin.

40
Immunohistochemistry (IHC)
Formalin-fixed paraffin-embedded tissue sections were used for IHC which was performed
according to a previously published protocol in our laboratory [18,24]. Briefly, after
deparaffinization and rehydration, antigen retrieval was performed in 10mM TRIZMA base (pH
9.0), 1mM EDTA buffer, and 0.05% Tween 20 with boiling under pressure for up to 15 minutes.
Polyclonal goat anti-RSV antibody (Millipore/Chemicon, Temecula, CA; Cat. No. AB1128) was
used as the primary antibody after two blocking steps. The first blocking was with 3% bovine
serum albumin in Tris-buffered saline+0.05% Tween 20 (TBS-T) and the second was 20% normal
swine serum in TBS-T for 15 minutes each. Primary antibody was followed by application of
biotinylated rabbit anti-goat secondary antibody (KP&L; Cat. No. 16-13-06). Signal development
was accomplished using a 1:200 dilution of streptavidin-horseradish peroxidase (Invitrogen; Cat.
No. 43-4323) for 30 minutes followed by incubation with Nova Red chromagen solution (Vector;
Cat. No. SK-4800). Positive signal was quantified in both bronchioles and alveoli for each tissue
section, and a score of 0-4 was assigned according to an integer-based scale of: 0=no positive
alveoli/bronchioles, 1=1-10 positive alveoli/bronchioles, 2=11-39 positive alveoli/bronchioles,
3=40-99 positive alveoli/bronchioles, 4=>100 positive alveoli/bronchioles. IHC for Spn was
performed using rabbit anti-Streptococcus pneumoniae polyclonal antibody (Thermo Fisher
scientific cat. # PA-7259) followed by biotin-labeled goat anti-rabbit IgG antibody (Thermo Fisher
Scientific Cat.#: 65-6140). Five random images were taken for each tissue section that were then
analyzed by the quantitative Halo program.
Quantitative Reverse Transcription Polymerase Chain Reaction (RT-qPCR)
BALF and lung tissue homogenates in Trizol were used to assess RSV mRNA expression by RT-
qPCR. The assay was performed as published previously in our laboratory [18,24,25]. Briefly,

41
RNA isolation from lung tissue and BALF was performed using the TRIzol method followed by
standard DNase treatment. RT-qPCR was carried out using One-Step Fast qRT-PCR Kit master
mix (Quanta, BioScience, Gaithersburg, MD) in a StepOnePlus™ qPCR machine (Applied
Biosystems, Carlsbad, CA) in conjunction with PREXCEL-Q assay-optimizing calculations.
Primers and probe for RSV M37 nucleoprotein were designed based on RSV accession number
M74568. Forward primer: 5′-GCTCTTAGCAAAGTCAAGTTGAACGA; reverse primer: 5′-
TGCTCCGTTGGATGGTGTATT; hydrolysis probe: 5′-6FAM-
ACACTCAACAAAGATCAACTTCTGTCATCCAGC-TAMRA.
Hematoxylin-Eosin Staining and Histological Scoring of Lung Sections
hematoxylin-eosin stained sections were examined via light microscope. An integer-based score
of 0-4 was assigned for each parameter (bronchiolitis, syncytial cells, epithelial necrosis, epithelial
hyperplasia, alveolar septal thickening, neutrophils in bronchial lumen, neutrophils in alveolar
lumen, alveolar macrophages, peribronchial lymphocytic infiltration, perivascular lymphocytic
infiltration, lymphocytes in alveolar septa, fibrosis), with 4 as the highest score. A final score was
calculated by adding up all measured scores to form a 0-48 score, with 48 as the highest, which is
called the accumulative histopathological lesion score.
Statistical Analysis
Statistical analysis used the Wilcoxon signed-rank test for non-parametric parameters such as
accumulative microscopic lesion scoring, followed by nonparametric comparisons for each pair
also using the Wilcoxon method. One-way ANOVA was followed by all pairs comparison by the
Tukey-Kramer HSD method for gross lesion scores and viral titer analyses by RT-qPCR and IFFU
assays.

42
Results
Infected Lambs Showed Marked RSV and Spn Titer
RSV titers and Spn colony-forming units were measured in this study to evaluate the degree
of infection by each pathogen and to investigate the possible effect(s) of co-infection in the
combined RSV-Spn group on the replication of each infectious agent. As measured by IFFU,
infectious RSV was detected in both RSV and RSV-Spn groups (P<0.0001) (Figure 1a). RSV titer
trended (not significant) 0.15 fold higher in the RSV-Spn group than RSV alone. A similar trend
was observed when assessing RSV mRNA detected in BALF by RT-qPCR. RSV mRNA was
elevated in both RSV and RSV-Spn groups (7.28 and 7.31 virion/ml) (Figure 1b). Furthermore,
similar to the viable virus titer increase, RSV virions measured by RT-qPCR in the lung of the
RSV-Spn group were 0.6 fold higher than the RSV-only group (Figure 1c). In contrast, Spn was
isolated in the lung tissue of both the Spn only and the RSV-Spn groups (56902.75 and 5456
CFU/ g, respectively) (p<0.0001) (Figure 1d). In contrast to the RSV/Spn group, the bacterial
titer was 9.4 fold higher in the Spn only infected group (p<0.05) (Figure 1d). Interestingly, Spn
titers in lambs that died before the end of the study were the highest of their groups. Lamb number
22 that was found dead 36hr after Spn infection in the RSV-Spn group had 59,302 CFU/ g, while
lamb number 9 that was euthanized 48 hr after Spn infection in the Spn group had a titer of
9,302,325 CFU/ g (Figure 1d). Spn was detected in the blood in both Spn infected groups -
indicating bacteremia/sepsis (Figure 1e).
RSV and Spn Induce a Well-recognized Macroscopic and Microscopic Lesion
Percent of lung tissue with gross lesions related to either infectious agent was determined at
necropsy coupled with post-necropsy retrospective qualitative analyses. Both RSV and Spn-related
lesions were found scattered across the lung surface in all lung lobes. Pinpoint dark red areas of

43
lung consolidation characterized RSV lesions. These areas were obvious in RSV and RSV-Spn
groups (P<0.001, and p<0.005, respectively). There were no significant differences in the
percentage of lung RSV macroscopic lesions detected between RSV and RSV-Spn groups (Figure
2a). Spn gross lesions are characterized by larger sizes of lung consolidation with bright red color
- which was seen to a lesser extent when compared to RSV lesions (Figure 2b). When combining
lesions observed associated with both RSV and Spn, there was a significant increase in observed
lesions within both RSV-infected groups in comparison to the Spn only group (p<0.001) (Figure
2c).
Microscopic lesions observed within the lung tissue reflected the infectious agent used and
contradicted our initial expectations (i.e. microscopic lesions caused by RSV infection were
multifocal areas of interstitial pneumonia and bronchiolitis scattered randomly and
homogeneously throughout the lung tissue). However, Spn induced diffuse homogenous and subtle
pathological changes in the lung tissue. Infection with either Spn, RSV, or both, markedly increase
microscopic lesions (accumulative microscopic lesion score) associated with the disease in
comparison to the control group (p<0.05) (Figure 3a-f). Additionally, the combined RSV-Spn
infection significantly increased the severity of microscopic lesions in comparison to the Spn only
group (p<0.05). Lesions varied among lambs, and RSV lesions consisted of thickening of the
interalveolar wall with inflammatory cellular infiltrates in the airway adventitia and lamina propria
(lymphocytes and plasma cells), alveolar lumen (alveolar macrophages and neutrophils), and
bronchiolar lumen (neutrophils). With RSV, overall, there was a varied degree of epithelial
necrosis and syncytial cell formation. On the other hand, Spn lesions consisted of moderate
interalveolar wall thickening with inflammatory cellular infiltrate mainly in the alveolar septae.
Most of the microscopic lesions seen with RSV overlapped with Spn-induced injury. However,

44
congestion of the interalveolar wall capillaries and hemorrhage was seen only in Spn-infected
lambs.
Immunohistochemistry was used to identify and localize RSV and Spn in tissue sections.
RSV were present multifocally throughout the sections with bronchial and peribronchial
distribution (Figure 4c). Therefore, RSV expression was evaluated in bronchioles and alveoli
separately. There were marked increases in RSV expression in bronchioles in both RSV only and
RSV-Spn groups (p<0.005 and p<0.001, respectively) with high RSV expression in the alveoli of
the RSV-Spn group (p<0.01) (Figure 4a). There were no significant differences between the RSV
only and RSV-Spn groups in the degree of RSV expression in lung tissue sections. Spn was random
and homogenously scattered throughout the lung sections with more intense signal in interalveolar
walls and blood capillaries (Figure 4d). Although not significant, there was a 0.5 fold increase in
Spn in the Spn only group when compared with the RSV-Spn group (Figure 4b).
Discussion
There is a critical need for an animal model to study bacterial pneumonia secondary to an
initial viral infection in the lung in order to study the mechanisms of viral-bacterial co-infection
and to evaluate therapeutic interventions. There are significant advantages of using lambs to model
RSV infection as a correlate for human infants - including the ability to use human viral strains
without adaptation and the similarity of the pathological sequelae [16,17]. In this study, we
demonstrate that Spn readily infects the lungs of lambs and establishes active bacterial pneumonia.
A previous study revealed that the peak of RSV titer and infection in lambs is around day 3 post-
viral nebulization, and we used this time-frame to model early human co-infection [24]. The
results of this study demonstrate consistency in the infection rate of both RSV and Spn, as well as
a good relation to the lesion development induced by either of the infectious agents. Although we

45
used 2 x 106 CFU of Spn for infection, murine studies typically use 5x105 to 107 CFU to establish
productive infections. Moreover, the lung volume of lambs is significantly larger than mice which
suggests that our inoculating dose may be more dispersed throughout the lungs than murine
studies. We believe that we may also be able to reduce the infection dosage to a lower CFU or
potentially use a colonization model to examine co-infection and pneumonia development.
The RSV titer in our study was increased by 0.5 fold in the RSV-Spn group, but bacterial
presence was reduced by 9.4 fold when compared to the RSV and Spn groups, respectively. These
data are consistent with prior studies in mice with influenza/Spn co-infections demonstrating
higher viral loads [11,26], although our observed viral (RSV) increase was quite modest in
comparison. Influenza co-infection studies also predict higher Spn burdens in the lungs due to
damaged epithelial cells serving as anchor points for the opportunistic bacteria. In other studies,
RSV with Spn in mice or cell culture predict that the RSV G protein on the infected epithelial
surface could also serve as an anchor point for Spn in the lungs [27]. In contrast to these murine
models, we found lower bacterial loads in the co-infection group over the Spn only group. These
findings suggest that the immune response might control Spn in the lungs of lambs better than
mice. Importantly, in human clinical studies of co-infection, show in increase in nasal
colonization numbers of Spn upon viral infection but this does not translate into higher invasive
lung disease [28]; suggesting that higher bacterial burdens could be a murine artifact rather than a
mechanism enhancing disease. In human studies of high Spn colonization, RSV disease appeared
less severe [29] suggesting that further using the lamb model to explore mechanistic differences
between Spn colonization and pneumonia during RSV.
The only deaths that occurred in the present study were in the Spn-infected groups, and both
lambs (lamb 11 in the Spn only group, and lamb 23 in the RSV-Spn group) had high lung Spn

46
colony-forming units/gram tissue. These could represent a failure to control bacterial division and
subsequent septicemia.
Lesion severity was consistent with the RSV titer and Spn burden as is shown by the
significant increase in the percent of lung tissue involved by gross lesions, and the increase in the
evaluated histological parameters. Gross lesions were multifocal lesions scattered randomly in all
lung lobes - which is the typical lesion distribution induced by RSV nebulization [18,24].
However, presentation of Spn gross lesions contradicted what was expected by the apparent
development of lesions in all lobes - including the caudal lung lobe, which is not typical for
bacterial pneumonia in lambs. However, the diffuse bacterial lesions and the presence of Spn
lesions in the caudal lobe may be due to the inoculation technique used for Spn infection. For Spn
infection, lambs were held vertically by one person and injected intratracheally by the second
person leading to a fall of inoculum through the bronchial tree into the caudal lobe, which in this
case, was favorable since it gives a bronchopneumonic distribution similar to that found in humans.
It is also possible that Spn spreads across lung lobes after inoculation either by airflow or vascular
flow. RSV-induced microscopic lesions were more prominent in comparison to Spn-induced
lesions and subsequently lead to significant differences between the RSV-Spn and Spn only
groups’ accumulative histologic lesion scores. RSV was more prominent in the bronchioles, while
Spn was diffuse throughout the lung sections.
Although mechanisms are being assessed, the higher observed morbidity in the RSV/Spn
group may derive from an enhanced neutrophil response present in the lungs. Evidence for this is
supported by the histopathology and the lower Spn burdens in these animals. Likely, RSV
infection served as a first activating response to neutrophils that could have then better controlled
the secondary bacterial infection. It is also possible that alveolar macrophages were activated by

47
RSV that, in turn, secreted inflammatory mediators that enhance neutrophil activation. Enhanced
neutrophil/leukocyte activation contrasts with studies in influenza co-infections in mice – which
suggests innate immune exhaustion [30]. While the time of inoculation could be a reason for the
observed differences, another could be the mere difference between influenza and RSV
pathogenesis. In either case, the results suggest further avenues of study using this model. We are
currently evaluating the neutrophil responses in these animals to ascertain how the innate antiviral
response may have played a role in better limiting the bacterial infection. The observed higher
RSV infection rate in the co-infection could also derive from the greater number of neutrophils in
the lungs in this group. There is evidence that RSV can infect neutrophils in humans [31] including
our unpublished data. Thus, if dual infection with Spn leads to enhanced neutrophil recruitment to
the lungs over RSV alone, those cells could become infected and contribute to the higher viral titer
we observed in the dual infection group. The effects of RSV infection on neutrophilic antibacterial
responses would be an interesting further study.
In this study, we have developed an animal model of co-infection for RSV and Spn. We
have determined enhanced disease with co-infection of both pathogens that correlates with human
and murine influenza studies, but this may all be due to a complex enhanced inflammatory/immune
response to co-infection rather than direct damage by either pathogen alone, although we cannot
rule the culpability of co-infection causation out. Future studies will plan to utilize other serotypes
of Spn to determine whether the results we observed are unique to serotype 22F or whether they
are a trend for all Spn strains in general. Additional studies will allow refinement of this model
and will include variations in inoculum volume/concentration, time between infections, and kinetic
analyses.

48
References
1. Nair H, Nokes DJ, Gessner BD, et al. Global burden of acute lower respiratory infections
due to respiratory syncytial virus in young children: a systematic review and meta-
analysis. The Lancet. 2010; 375(9725):1545–1555.
2. Glezen WP, Taber LH, Frank AL, Kasel JA. Risk of Primary Infection and Reinfection
With Respiratory Syncytial Virus. Am J Dis Child. 1986; 140(6):543–546.
3. Thompson WW, Shay DK, Weintraub E, et al. Mortality Associated With Influenza and
Respiratory Syncytial Virus in the United States. JAMA. 2003; 289(2):179–186.
4. Nicholson KG. Impact of influenza and respiratory syncytial virus on mortality in
England and Wales from January 1975 to December 1990. Epidemiol Amp Infect. 1996;
116(1):51–63.
5. Zambon M, Stockton J, Clewley J, Fleming D. Contribution of influenza and respiratory
syncytial virus to community cases of influenza-like illness: an observational study. The
Lancet. 2001; 358(9291):1410–1416.
6. Thorburn K, Harigopal S, Reddy V, Taylor N, Saene HKF van. High incidence of
pulmonary bacterial co-infection in children with severe respiratory syncytial virus
(RSV) bronchiolitis. Thorax. 2006; 61(7):611–615.
7. Madhi SA, Klugman KP. A role for Streptococcus pneumoniae in virus-associated
pneumonia. Nat Med. 2004; 10(8):811–813.
8. Bogaert D, Groot R de, Hermans P. Streptococcus pneumoniae colonisation: the key to
pneumococcal disease. Lancet Infect Dis. 2004; 4(3):144–154.

49
9. O’Brien KL, Wolfson LJ, Watt JP, et al. Burden of disease caused by Streptococcus
pneumoniae in children younger than 5 years: global estimates. The Lancet. 2009;
374(9693):893–902.
10. Klugman KP, Madhi SA, Huebner RE, Kohberger R, Mbelle N, Pierce N. A Trial of a 9-
Valent Pneumococcal Conjugate Vaccine in Children with and Those without HIV
Infection. N Engl J Med. 2003; 349(14):1341–1348.
11. Smith AM, Adler FR, Ribeiro RM, et al. Kinetics of Coinfection with Influenza A Virus
and Streptococcus pneumoniae. PLOS Pathog. 2013; 9(3):e1003238.
12. Demczuk WHB, Martin I, Hoang L, et al. Phylogenetic analysis of emergent
Streptococcus pneumoniae serotype 22F causing invasive pneumococcal disease using
whole genome sequencing. PLOS ONE. 2017; 12(5):e0178040.
13. Jacobs MR, Good CE, Bajaksouzian S, Windau AR. Emergence of Streptococcus
pneumoniae Serotypes 19A, 6C, and 22F and Serogroup 15 in Cleveland, Ohio, in
Relation to Introduction of the Protein-Conjugated Pneumococcal Vaccine. Clin Infect
Dis. 2008; 47(11):1388–1395.
14. Roymans D, Alnajjar SS, Battles MB, et al. Therapeutic efficacy of a respiratory
syncytial virus fusion inhibitor. Nat Commun [Internet]. 2017 [cited 2017 Aug 30]; 8.
Available from: http://www.ncbi.nlm.nih.gov/pmc/articles/PMC5537225/
15. Detalle L, Larios A, Gallup L, Van Geelen A, Duprez L, Stohr T. Delivery of ALX-0171
by inhalation greatly reduces disease burden in a neonatal lamb RSV infection model. 9th
Int Respir Syncytial Virus Symp Cape Town South Afr Abstr OP 72. 2014.
16. Ackermann MR. Lamb Model of Respiratory Syncytial Virus–Associated Lung Disease:
Insights to Pathogenesis and Novel Treatments. ILAR J. 2014; 55(1):4–15.

50
17. Derscheid RJ, Ackermann MR. Perinatal Lamb Model of Respiratory Syncytial Virus
(RSV) Infection. Viruses. 2012; 4(10):2359–2378.
18. Derscheid RJ, Geelen A van, McGill JL, et al. Human Respiratory Syncytial Virus
Memphis 37 Grown in HEp-2 Cells Causes more Severe Disease in Lambs than Virus
Grown in Vero Cells. Viruses. 2013; 5(11):2881–2897.
19. Zaghawa A, Hassan H, El-Sify A. Clinical and Etiological study on respiratory affections
of sheep. Minufiya Vet J. 2010; 7(1):93–103.
20. Garedew L, Ayelet G, Yilma R, Zeleke A, Gelaye E. Isolation of diverse bacterial species
associated with maedi-visna infection of sheep in Ethiopia. Afr J Microbiol Res. 2010;
4(1):014–021.
21. DeVincenzo JP, Wilkinson T, Vaishnaw A, et al. Viral Load Drives Disease in Humans
Experimentally Infected with Respiratory Syncytial Virus. Am J Respir Crit Care Med.
2010; 182(10):1305–1314.
22. Kim Y-I, DeVincenzo JP, Jones BG, et al. Respiratory Syncytial Virus Human
Experimental Infection Model: Provenance, Production, and Sequence of Low-Passaged
Memphis-37 Challenge Virus. PLOS ONE. 2014; 9(11):e113100.
23. Grosz DD, Geelen A van, Gallup JM, Hostetter SJ, Derscheid RJ, Ackermann MR.
Sucrose stabilization of Respiratory Syncytial Virus (RSV) during nebulization and
experimental infection. BMC Res Notes. 2014; 7:158.
24. Larios Mora A, Detalle L, Van Geelen A, et al. Kinetics of Respiratory Syncytial Virus
(RSV) Memphis Strain 37 (M37) Infection in the Respiratory Tract of Newborn Lambs
as an RSV Infection Model for Human Infants. PLoS ONE [Internet]. 2015 [cited 2016
Jan 7]; 10(12). Available from: http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4671688/

51
25. Derscheid RJ, Gallup JM, Knudson CJ, et al. Effects of Formalin-Inactivated Respiratory
Syncytial Virus (FI-RSV) in the Perinatal Lamb Model of RSV. PLOS ONE. 2013;
8(12):e81472.
26. Nguyen DT, Louwen R, Elberse K, et al. Streptococcus pneumoniae Enhances Human
Respiratory Syncytial Virus Infection In Vitro and In Vivo. PLOS ONE. 2015;
10(5):e0127098.
27. Avadhanula V, Wang Y, Portner A, Adderson E. Nontypeable Haemophilus influenzae
and Streptococcus pneumoniae bind respiratory syncytial virus glycoprotein. J Med
Microbiol. 2007; 56(9):1133–1137.
28. Moyes J, Cohen C, Pretorius M, et al. Epidemiology of Respiratory Syncytial Virus–
Associated Acute Lower Respiratory Tract Infection Hospitalizations Among HIV-
Infected and HIV-Uninfected South African Children, 2010–2011. J Infect Dis. 2013;
208(suppl_3):S217–S226.
29. Vissers M, Ahout IM, Kieboom CH van den, et al. High pneumococcal density correlates
with more mucosal inflammation and reduced respiratory syncytial virus disease severity
in infants. BMC Infect Dis. 2016; 16(1):129.
30. McNamee LA, Harmsen AG. Both Influenza-Induced Neutrophil Dysfunction and
Neutrophil-Independent Mechanisms Contribute to Increased Susceptibility to a
Secondary Streptococcus pneumoniae Infection. Infect Immun. 2006; 74(12):6707–6721.
31. Halfhide CP, Flanagan BF, Brearey SP, et al. Respiratory Syncytial Virus Binds and
Undergoes Transcription in Neutrophils From the Blood and Airways of Infants With
Severe Bronchiolitis. J Infect Dis. 2011; 204(3):451–458.

52
Figures and Legends
C
on
tr
ol
S
pn
S
pn
#
9
R
S
V
R
S
V
-S
pn
+
R
S
V
-S
pn
#
22
1 00
1 01
1 02
1 03
1 04
1 05
1 06
1 07
lo
g
CF
U/
1
00
u
g
d
*
* * * * *
* * * * *
* * * * ** * * * *
C o n t r o l S p n R S V R S V - S p n
0
1 0 0 0 0
2 0 0 0 0
3 0 0 0 0
4 0 0 0 0
5 0 0 0 0
FF
U/
mL
B
AL
F
a * * * * *
* * * * *
* * * * *
* * * * *
C o n t r o l S p n R S V R S V - S p n
1 00
1 01
1 02
1 03
1 04
1 05
1 06
1 07
1 08
1 09
1 01 0
Lo
g
vi
ri
on
s/
mL
B
AL
F
b
* * * * *
* * * * *
* * * * *
* * * * *
C o n t r o l S p n R S V R S V - S p n
0
5 1 09
1 1 01 0
1 . 5 1 01 0
2 1 01 0
Lo
g
RS
V
vi
ri
on
s/
g
Lu
ng
c
* * * *
* * *
* * * *
* * *
C o n t r o l S p n R S V R S V - S p n
1 00
1 01
1 02
1 03
1 04
1 05
1 06
1 07
CF
U
/1
00
u
l
e* * * * *
* * * * *
* * * * *
* * * * *
Figure 2-1:RSV and Spn titer in lung tissue and blood
(a) number of infectious RSV particles as measured by IFFU assay, (b) RSV mRNA level in the
BALF, (c) RSV mRNA level in lung tissue (d) Spn colony forming unit per 100 μg lung tissue, (e)
Spn colony forming unit per 100 μl blood, all shown as average + SEM. Animals were either
infected with mock media (control), RSV, Spn, or RSV followed by Spn (RSV-Spn). *P<0.05,
**P<0.01, ***P<0.005, **** P<0.001, ***** P<0.0001.
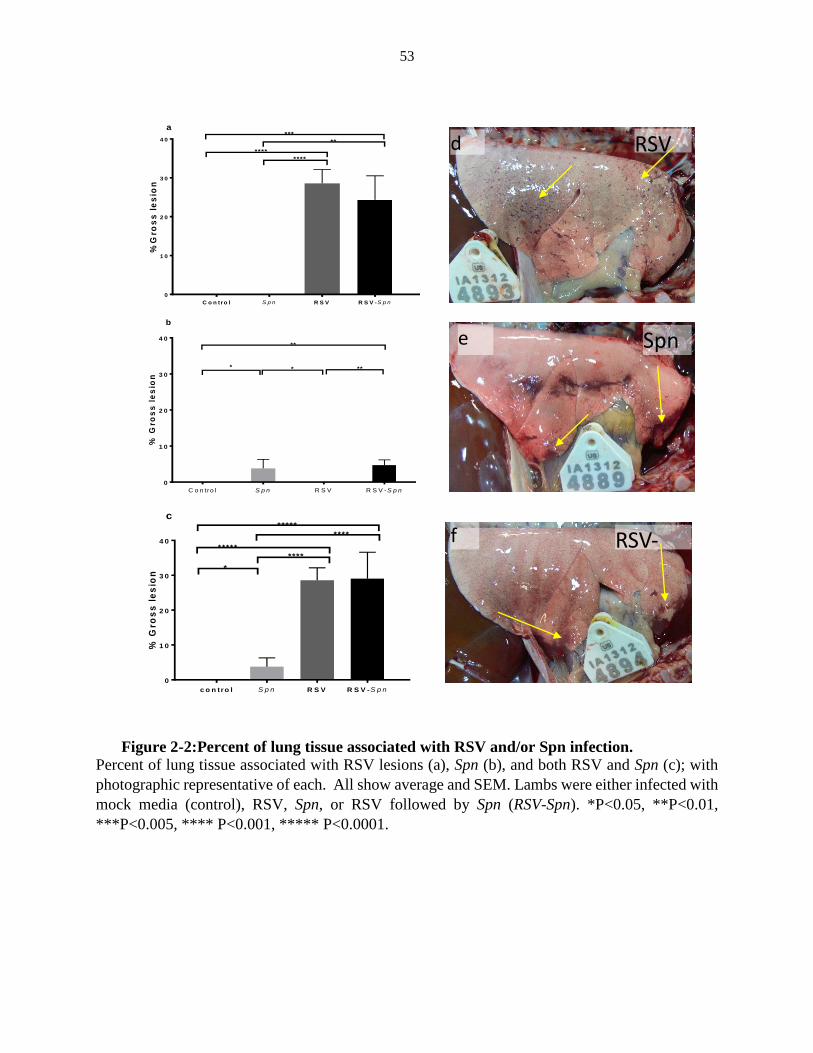
53
Percent of lung tissue associated with RSV lesions (a), Spn (b), and both RSV and Spn (c); with
photographic representative of each. All show average and SEM. Lambs were either infected with
mock media (control), RSV, Spn, or RSV followed by Spn (RSV-Spn). *P<0.05, **P<0.01,
***P<0.005, **** P<0.001, ***** P<0.0001.
C o n tro l S p n R S V R S V -S p n
0
1 0
2 0
3 0
4 0
%G
ros
s l
es
ion
**
***
****
****
a
C o n tro l S p n R S V R S V -S p n
0
1 0
2 0
3 0
4 0
% G
ros
s l
es
ion
**
**
**
b
c o n tro l S p n R S V R S V -S p n
0
1 0
2 0
3 0
4 0
% G
ros
s l
es
ion
****
*****
*********
*
c
RSV
Spn
RSV-Spn
d
e
f
Figure 2-2:Percent of lung tissue associated with RSV and/or Spn infection.

54
(a) accumulative histologic lesion associated with RSV and Spn infection shown as average +
SEM. (b-f) show a representative photograph of lung tissue sections stained with Gram
stain (b), H&E stained tissue section of control (c), RSV only (d), Spn only (e), combined
RSV-Spn (f). Lambs were either infected with mock media (control), RSV, Spn, or RSV
followed by Spn (RSV-Spn). * P<0.05, **P<0.01, ***P<0.005, **** P<0.001, *****
P<0.0001.
C o n tr o l S p n R S V R S V-S p n
0
5
1 0
1 5
ac
cu
mil
ati
ve
his
tolo
gic
le
sio
n
**
*
*a
e
d
f
c
b
Figure 2-3:Histologic lesions associated with RSV, Spn, and RSV-Spn
combined infection.

55
Number of bronchioles and alveoli express the RSV positive signal (a), surface area (mm2)
occupied by Spn IHC positive staining (b), all shown as average + SEM. (c) and (d) show a photo
representation of RSV (c) and Spn (d) IHC positive staining. Animals were either infected with
mock media (control), RSV, Spn, or RSV followed by Spn (RSV-Spn). * P<0.05, **P<0.01,
***P<0.005, **** P<0.001, ***** P<0.0001.
co
ntr
ol
Spn
RS
V
RS
V-S
pn
0
5 0 0
1 0 0 0
1 5 0 0
an
tig
en
ex
prs
sio
n m
m2
* * * ** * *
*
b
C o n tro l S p n R S V R S V -S p n
0
5
1 0
1 5
2 0
2 5
nu
mb
er o
f R
SV
po
sit
ive
bro
nc
hio
les
/ a
lve
oli
B ro n c h io le s
A lv e o li
a
********
***
***
****
c d
Figure 2-4:Immunohistochemistry staining of RSV and Spn in FFPE lung tissue sections.

56
THERAPEUTIC EFFICACY OF JNJ-49214698, AN RSV
FUSION INHIBITOR, IN RSV-INFECTED NEONATAL LAMBS
A paper submitted to Frontiers in Microbiology Journal
Sarhad S. Alnajjar1,2,3, Alejandro Larios-Mora1, Albert Van-Geelen4, Jack M. Gallup1,
Anil Koul5, Peter Rigaux5, Dirk Roymans5, Mark R. Ackermann3
1Department of Veterinary Pathology, College of Veterinary Medicine, Iowa State
University, Ames, Iowa USA
2Department of Veterinary Pathology, College of Veterinary Medicine, Baghdad
University, Baghdad, Iraq
3Biomedical Sciences, Carlson College of Veterinary Medicine, Oregon State
University
4 Agricultural Research Service, United States Department of Agriculture, Ames, Iowa
5Respiratory Infections Discovery, Janssen Infectious Diseases, Beerse, Belgium
Authors Contributions
S.S.A., A. L., J.M.G, and M.R.A carried out several aspects of the neonatal lamb infection
experiment including viral inoculation, treatment deliveries, blood draws, animal monitoring and
assessment of RSV clinical signs, necropsy, IFFU, qRT-PCR, histological, IHC, and RNAscope
assays. A.V. assisted with viral preparation and growth, and IFFU assays. D.R., P.R., and J.M.G
performed data processing. D.R., P.R., and A.K. provided the JNJ-49214698 compound, assayed
the compound for distribution in different body compartments and performed global statistical
analysis of all data. D.R., P.R., A.K., S.S.A., J.M.G, and M.R.A assisted with experimental
design and preparation of this manuscript.

57
ACKNOWLEDGMENTS
The authors graciously thank Laboratory Animal Resources (LAR) veterinarians Dr.
Mary Sauer and Dr. Kathleen Mullin for their expertise with medical intervention and critical
animal care. We also thank Livestock Infectious Disease Isolation Facility (LIDIF)
staff: Diane McDonald, Michelle Tye, Angelia Troutwine, Lori Bonnell, Alan Elsberry, and
Dean Isaacson for animal acquisition and superb daily animal care and data collection. As
well, we thank members of the Histology Laboratory staff: Jennifer Groeltz-Thrush, Toni
Christofferson, Diane Gerjets, and Ronisue Aust for their expert help with preparing sections
for histology and immunohistochemistry. We also kindly thank Dr. Panchan
Sitthicharoenchai for her excellent input in the writing of this manuscript. Finally, we thank
the professional student help we received for this project from Lauren Rowan, Meghann
McCaull, Marissa Kleve, Nicole Lacy, Alanna Hennen, and Drew Grosz.
Conflict of Interests
Sarhad S. Alnajjar, Alejandro Larios-Mora, Albert Van-Geelen, and Jack M. Gallup
worked at Iowa State University for this study and have no disclosures or conflicts. Mark R
Ackermann also worked at Iowa State Univesity for this study served as principal
investigator who received funds from Janssen Pharmaceutica to conduct this study. Anil
Koul, Peter Rigaux, and Dirk Roymans are employees of Janssen Pharmaceutica NV.
Funding
This study was funded by a grant from Janssen Pharmaceutica NV ( Iowa State Univesity
grant # MA14002).

58
Abstract
Respiratory syncytial virus (RSV) is a leading cause of respiratory infection, hospitalization,
and death in infants worldwide. No fully effective RSV therapy using direct antivirals is marketed.
Since clinical efficacy data from naturally infected patients for such antivirals are not available
yet, studies in animals are indispensable to predict if therapeutic intervention. Here we report the
impact of an RSV fusion inhibitor, JNJ-49214698, on severe RSV-associated acute lower
respiratory tract infection (ALRTI) in neonatal lambs. Randomized animals were treated once-
daily with 25 mg/kg JNJ-49214698, starting either before RSV infection, 1 day post-infection, or
as late as peak lung viral load on Day 3 post-infection. The efficacy of treatment was assessed by
scoring clinical signs of illness, development of RSV-induced gross and microscopic lung lesions,
and measuring virus titers in the lungs. Treatment with JNJ-49214698 was very effective in all
treatment groups. Even in animals for which treatment was delayed until peak viral load was
reached, a reduced amount and severity of gross and microscopic lesions as well as RSV titers and
RNA levels were found. These results strongly suggest that treatment with small-molecule fusion
inhibitors is an effective strategy to treat patients who are diagnosed with an RSV-induced ALRTI.
Key Words
Respiratory Syncytial Virus, small molecule fusion inhibitor, RSV Fusion inhibitor,
antiviral, neonatal lamb model, acute lower respiratory tract infection
Introduction
Respiratory syncytial virus (RSV) is one of the leading causes of acute lower
respiratory tract infection (ALRTI). Globally, it is estimated that 33.8 million cases of

59
ALRTI associated with RSV infection occur yearly in children under the age of five.
Subsequently, RSV-associated ALRTI is a major cause of hospital admissions with an
estimated 3.4 million cases annually; and RSV-associated mortality is estimated to be
66,000-199,000 in children under the age of five occurring mostly in developing countries
[1]. Furthermore, post-bronchiolitic development of wheezing syndrome is sometimes
observed, and although still under debate, it was suggested that the development of asthma
could be associated with RSV infection [2,3]. RSV is also one of the leading causes of
respiratory infection in the elderly and high risk adults [4,5]. A recent study showed that 3%
of pneumonia cases in human adults is caused by RSV [6] and with the widespread use of
influenza vaccination, RSV has a similar disease burden in the elderly [4].
Despite decades of investigation, no market-approved vaccine exists [7]. Seasonal
prophylaxis with the monoclonal antibody palivizumab, restricted to high-risk infants in
developed countries, is the only specific antiviral strategy available [8]. The disease
preventive effects of palivizumab in these patients were demonstrated by a significant
reduction in hospitalization rate, hospital residence time, admission to intensive care unit and
days requiring oxygen therapy [9]. However, the therapeutic efficacy of antibodies like
palivizumab in infants hospitalized with an established RSV-associated ALRTI is
questionable at best [10–12]. Other drugs like bronchodilators, corticosteroids, and
antibiotics, often used but not indicated to treat RSV-associated bronchiolitis, do not seem
to confer sufficient therapeutic benefit either, leaving supportive care essentially as the major
treatment option [13,14].
New treatments are clearly needed to decrease the medical consequences related to
RSV-associated ALRTI. Data on the impact of small-molecule direct antivirals on RSV

60
disease manifestation in naturally infected hospitalized patients or outpatients are not
available, and as such, reliable modeling of human RSV disease remains a necessary step in
the search for novel therapies.
The development of an RSV experimental human infection model has provided a new
tool to obtain human proof-of-concept efficacy data of RSV direct antivirals early in the
clinical development pipeline [15–17]. However, volunteers are often young, healthy adults
and infection is studied only in the upper respiratory tract, limiting the information about the
efficacy of therapeutics to treat ALRTI. Therefore, the recent development of a fully-
replicative neonatal lamb model for RSV has presented a significant step forward to generate
reliable preclinical efficacy data and to improve the selection of the proper clinical candidate
molecules. Neonatal lambs are susceptible to human as well as ovine and bovine strains.
Similarities between the pulmonary and immunological systems between lambs and humans
make the neonatal lambs an excellent model to study human RSV disease and the impact of
new potential RSV therapeutics[18,19]. Moreover, neonatal lambs infected with RSV
develop clinical symptoms including but not limited to fever, tachypnea or increased
expiratory effort (wheeze), lethargy, and develop mild to moderate bronchiolitis and
pneumonia. Innate and adaptive immune responses by neonatal lambs are very similar to
those of human infants [20,21].
A promising approach to inhibit RSV is by targeting the viral fusion (F) protein [22,23].
A few small-molecule RSV fusion inhibitors are currently being evaluated in early-stage
clinical trials, but their clinical impact on RSV-associated ALRTI in hospitalized infants is
yet unknown [24–26]. The purpose of this study is therefore to evaluate the therapeutic

61
impact of an experimental RSV fusion inhibitor, JNJ-49214698, on RSV-associated ALRTI
using a lamb model of RSV infection.
Materials and Methods
(see online Supplementary Data for detailed methods)
Compound and Dosing
JNJ-49214698 was discovered and synthesized by Janssen Infectious Diseases (Beerse,
Belgium). The compound was formulated in 10% acidified hydroxypropyl-β-cyclodextrin
[10% HP-β-CD + HCl, pH 2 (vehicle)] at 6.25 mg/mL prior to dosing and stored throughout
the study at 4 °C. The compound was dosed orally by catheter-mediated orogastric gavage
at 4 mL/kg body weight (25 mg/kg) once daily. Dose selection in this study aimed for
reaching the highest possible, safe exposure in the animals in order to maximize the
likelihood to obtain efficacy while avoiding toxic side effects. A daily oral dose of 25 mg/kg
was selected based on the antiviral activity of JNJ-49214698 [EC50 = 0.4 ng/mL (0.8 nM)
and EC90 = 2.4 ng/mL (4.8 nM)], JNJ-49214698 exposure levels obtained in neonatal lambs
at different doses during a separate pharmacokinetic (PK) study and an observed lack of
toxicity of JNJ-49214698 at least until Cmax = 17,167 ng/mL and AUC0-24h = 139,993
ng.h/mL in a 5-day repeated dose rat tolerance study.
Animals
Twenty-one colostrum-deprived neonatal lambs (Suffolk, Polypay, Dorsett cross) aged
1-3 days and 2-7 kg body weights were obtained for this experiment. Animal use was
approved by the Institutional Animal Care and Use Committee of Iowa State University.
RSV-infected lambs were kept in a separate room from the non-infected animals in the
Livestock Infectious Disease Isolation Facility (LIDIF). Lambs were fed iodide-free lamb

62
milk replacer diet (Milk Products Inc., Chilton, WI, USA) [27], and were treated with Naxcel
(Ceftiofur sodium, Pfizer) intramuscular once daily to reduce/prevent secondary bacterial
infections.
Experimental Design
Lambs were randomly assigned to five different groups. Three groups (Px, Tx-1 and
Tx-2) were infected with RSV and treated with JNJ-49214698. The first group (Px, n = 4)
was treated prophylactically, 1 day before RSV challenge and then daily afterward up until
Day 5 post infection (p.i.). The second (Tx-1, n = 5) and the third (Tx-2, n = 5) groups were
treated one day and three days after viral challenge and daily afterward up until Day 5 p.i.,
respectively. The vehicle group (n = 4), serving as a positive RSV control group, was infected
with RSV, but received treatment with vehicle only. The No RSV group (n = 3), served as
negative RSV control group, and was aerosolized with RSV-free, HEp-2 cell-conditioned
media and also received vehicle. All lambs were euthanized at Day 6 p.i. and all endpoints
measured after euthanasia.
RSV Infection
Lambs were infected with RSV strain M37, purchased from Meridian BioSciences
(Memphis, TN, USA). This strain is a wild type A RSV isolated from the respiratory
secretions of an infant hospitalized for bronchiolitis [28,29]. In our laboratory, M37 was
grown in HEp-2 cells and stored at -80°C in media containing 20% sucrose [30]. Six mL of
1.27 x 107 Infectious Forming Unit (IFFU)/mL in media containing 20% sucrose or cell-
conditioned mock media (also containing 20% sucrose) was nebulized using PARI LC
Sprint™ nebulizers to each lamb over the course of 25-30 minutes resulting in the total
inhalation of about 3 mL by each lamb [31].

63
Assessed Parameters
JNJ-49214698 exposure in plasma, BALF, and lung homogenate samples was assessed
using a method based on Protein precipitation and HPLC/MS/MS analysis. Quantitative
reverse transcription polymerase chain reaction (qRT-PCR) was done to measure the RSV
mRNA expression in BALF and lung homogenate samples as previously done in our lab
[30,32,33]. Infectious focus-forming unit (IFFU) assay was used to determine the viable RSV
in BALF samples as previously described [32]. Chlincal score was calculated by the number
of respiratory associated signs present in each animal. For the pathological study, percent of
lung parenchyma involved with RSV induced lesion were measured to evaluate the RSV
gross lesion. Histologically, Hematoxylin-eosin stained sections were examined via light
microscope as described previously [32] with some modification. Lung lesions were scored
according to an integer-based score of 0-4 for each parameter (bronchiolitis, syncytial cells,
epithelial necrosis, epithelial hyperplasia, peribronchial lymphocytic infiltration,
perivascular lymphocytic infiltration, neutrophils), with 4 as the highest score. Then a final
score (accumulative histo lesion score) was assigned by adding up the scores from the seven
individual parameters resulting in final accumulative scores ranging from 0-28 representing
the total RSV-associated lesion in each tissue section. For further evaluation of the JNJ-
49214698 therapeutic efficacy, Immunohistochemical staining was performed to evaluate
RSV antigen expression in lung tissue section as previously described in our laboratory
[32,34,35]. Formalin-fixed paraffin-embedded tissue sections were used for the RNAscope
detection of RSV mRNA in situ. A probe designed to the hRSV M37 nucleoprotein gene
(accession number KM360090) was used (Probe-V-RSV-NP, Advance Cell Diagnostic,
Catalog number 439866). The assay was performed according to the manufacturer’s manual
(user manual document number 320497; RNAscope® 2.0 HD Detection Kit (BROWN) User

64
Manual PART 2). Sections were examined under light microscope, and the number of
bronchioles and alveoli containing the positive signal were counted. The number of positive
bronchioles and alveoli per tissue section was then assigned a score according to the simple
integer-based scale of 0 = no positive alveoli/bronchioles, 1 = 1-10 positive
alveoli/bronchioles, 2 = 11-39 positive alveoli/bronchioles, 3 = 40-99 positive
alveoli/bronchioles, 4 = >100 positive alveoli/bronchioles.
Statistical Analysis
Statistical analysis was completed by using the Kruskall-Wallis test for non-parametric
parameters such as accumulative microscopic lesion scoring, immunohistochemistry and
RNAscope integer-based scores, followed by Dunn’s post-hoc test for multiple comparisons.
One-way ANOVA followed by Dunnett multiple comparisons test was used to compare the
treated groups to the RSV-infected non-treated control group for gross lesion scores and viral
titer analyses by qRT-PCR.
Results
JNJ-49214698 Efficiently Distributes to Different Lung Compartments of Neonatal
Lambs
During a multicyclic RSV infection, small-molecule fusion inhibitors are thought to
inhibit each new infection event at the time of viral fusion. The concentration of JNJ-
49214698 was measured in BALF as well as in homogenized lavaged-lung tissue. To assess
the potential of JNJ-49214698 to distribute from the blood to the lungs of the neonatal lambs,
we also measured its level in plasma. JNJ-49214698 was absorbed very quickly into the
circulation and tended to slightly accumulate over time during the study period. The average
steady-state concentration of JNJ-49214698 in plasma was in the range of 1770 ± 435 and

65
3112 ± 1441 ng/mL across the different treatment groups and was reached within 24-48 hr
after the first dose (Figure 1a). Twenty-four hour after the final dose, the level of JNJ-
49214698 in the BALF of the group receiving prophylactic treatment (Px), or groups in
which compound administration was started one (Tx-1) or three (Tx-2) days after viral
inoculation was 2044 ± 648, 3289 ± 977 and 1109 ± 252 ng/mL, respectively (Figure 1b).
The average lavaged-lung concentrations measured were 2867 ± 403, 5211 ± 841 and 1976
± 175, in Px, Tx-1, and Tx-2 treatment groups, respectively, resulting in an approximate
lung/plasma ratio of 1.4 to 1.8 and a good distribution of JNJ-49214698 in the BALF (Figure
1c). Together, these results indicate a good distribution of JNJ-49214698 to different lung
compartments.
Treatment with JNJ-49214698 Reduces Incidence and Duration of RSV-Associated
Symptoms
Clinical signs of RSV infection in lambs, as in human infants, can be variable and
sometimes difficult to assess. Despite this, RSV-associated respiratory distress can be
observed in the animals as early as one day p.i., developing further in some animals into clear
external signs of RSV-associated illness such as nasal discharge, wheezing or lethargy. By
Day 1 p.i., most of the lambs in this study, except for the animals in the non-infected group,
had already become lethargic as characterized by decreased activity (lower frequency of
getting up and movement in general). The behavior and respiratory-related symptoms in
lambs of the vehicle-treated group (RSV and no treatment) steadily worsened during the
following days (Figure 2a). One lamb started to produce nasal discharge and displayed
episodes of wheezing as from Day 3 p.i. onwards, while its respiratory rate was clearly
increased by Day 6. One lamb on Day 3 p.i. died and was found during post-mortem analysis
with severe gross lesions consistent with RSV infection in its lungs. The other lambs

66
survived until Day 6 when they were euthanized, but by Day 4, all 4 remaining animals from
this group were lethargic.
In contrast, while by Day3 post-infection 3 lambs in the Px group showed fever, the
remaining lambs in Px and Tx-1 groups were free of symptoms by Day 3 post-infection
(Figure 2b and c). Although less prominently, the incidence and duration of RSV-associated
symptoms was also reduced in the lambs of the Tx-2 group as compared to the vehicle-only
treated animals (Figure 2d).
In summary, these findings suggest that treatment with JNJ-49214698 has the potential
to reduce the incidence and duration of the clinical manifestation of RSV disease in lambs.
Treatment with JNJ-49214698 Inhibits a Multicyclic RSV Infection in Neonatal Lambs
The presence of nucleoprotein RNA in BALF and lung samples was quantified by qRT-
PCR. In BALF, The Px group showed a significant 3.4 log10 (p<0.0001) reduction of the
viral RNA level in comparison to the RSV-infected vehicle group (Figure 3a), while the viral
RNA in the Tx-1 and Tx-2 groups was reduced less by approximately 7- and a 3-fold as
compared to the vehicle only treated animals. We could not detect viral RNA in the lung
tissue of prophylactically treated animals, and profound reductions in RSV RNA of
approximately 23-fold in Tx-1 group (p<0.02), and 10-fold in Tx-2 in compared to the
vehicle group (Figure 3b). The infectious viral titer in BALF were done and partially reported
in a separate manuscript [23] in which IFFU were significantly reduced in all treated groups
with no infective virus detected in Px group that receive prophylaxis with JNJ-49214698
similar to what we found in RSV mRNA in lung tissue. There were 34- and a 93-log
reduction in the infectious titer as compared to the RSV-infected control group, was observed

67
in the Tx-1 and Tx-2 groups, respectively (p<0.001) (Figure 3c). To confirm the quantified
viral titers in BALF and lavaged-lung tissue, we determined RSV (M37) antigen and RNA
by immunohistochemical and RNAscope analysis, respectively. RSV antigen was present in
infected airway epithelial cells of bronchi, bronchioles, and alveoli and occasionally in
alveolar macrophages. However, staining of RSV antigen was markedly reduced in all drug-
treated groups in comparison to the RSV-infected control group (Figure 4a, c). RSV antigen
was highly decreased in the Tx-2 group as compared to the vehicle-treated group, and again,
almost no RSV antigen was detected in both Px (p<0.01) and Tx-1 (p<0.05) treatment groups
(Figure 4a, c). When RSV RNA (M37 nucleoprotein) expression in lungs was determined by
RNAscope, concomitant with the RNA levels as determined by qRT-PCR, there was a
marked reduction of RSV RNA expression in all the compound-treated groups in both
bronchioles and alveoli (Figure 4b, d). Heavy brown staining of RSV RNA in RSV-infected
non-treated lung sections was observed which was concentrated in the bronchioles and
alveoli in the consolidated part of the lung (Figure 4d). In contrast, only minimal staining
was seen in the Px (p<0.05) and Tx-1 groups (Figure 4d), aligning with
immunohistochemical and qRT-PCR data.
Treatment with JNJ-49214698 Inhibits RSV-Induced Lung Pathology and Cellular
Immune Response of Neonatal Lambs
Approximately 43% of the lung surface of infected vehicle-treated animals had RSV
associated macroscopic lesions which were multifocal dark red pinpoint foci of consolidation
scattered randomly over the lung surface and deeply throughout the lung tissue (Figure 5a,
b). Moreover, gross lung lesions were evenly distributed over the different left and right lung
lobes. Prophylaxis (Px) or treatment starting twenty-four hours after infection (Tx-1)
completely abolished the formation of RSV associated gross lung lesion (Figure 5a, b). When

68
treatment was delayed even as late as three days after infection (Tx-2), a significant reduction
in the RSV associated gross lesions was observed; dropping from 43 to 16% (p<0.0001)
(Figure 5b). As a negative control, no lesions were observed in the lungs of non-infected,
vehicle-treated animals (Figure 5b).
To investigate the effect of JNJ-49214698 on lung tissue microscopically, we assessed
the impact of the compound on the development of bronchiolitis, syncytial cell formation,
epithelial necrosis and hyperplasia, and inflammatory cell infiltration in lungs. An
accumulative histological lesion score was assigned by combining all the assessed
parameters. Significant reductions of the accumulative score were seen across
prophylactically treated animals or animals that received early treatment starting 1 day p.i.
with JNJ-49214698, (p<0.01 and p<0.05, respectively) (Figure 6a). Prophylaxis with JNJ-
49214698 completely prevented the formation of microscopic lung lesions after RSV
infection (Figure 6a, b). Although the effect on the accumulative histologic score was least
prominent in animals that received treatment with JNJ-49214698 as late as 3 days after
infection, a prominent reduction as compared to the vehicle-treated group was still observed,
with some animals displaying almost no microscopic lesions at all.
Typical RSV lesions that were present in the RSV-infected control group were
multifocal interstitial pneumonia and bronchiolitis. There was thickening of the inter-
alveolar wall due to type II pneumocyte hyperplasia and lymphocyte infiltration (Figure 6b).
There was neutrophil infiltration in the bronchial and bronchiolar lumens and the alveolar
lumen with multifocal and segmental areas of necrosis and sloughing of the epithelial cells
lining bronchioles. Some bronchioles had modest thickening of epithelium due to
hyperplasia. In addition, there were occasional multinucleated syncytial cells present

69
throughout the lung sections. In the lesions formed in the latter Tx-1 group, the formation of
syncytial cells, epithelial necrosis or hyperplasia and infiltrating neutrophils was absent,
while the other parameters assessed were markedly reduced, resulting in a prominent
decrease of the accumulative histological lesion scores.
Discussion
Currently, there are no effective direct antivirals available for the treatment of RSV-
associated ALRTI. Even though a number of RSV inhibitors have reached early-stage
clinical evaluation, no data are available yet demonstrating their clinical benefit in naturally
infected patients suffering from severe RSV-associated ALRTI, and so uncertainty remains
about the therapeutic treatment window and the impact of such molecules on severe RSV
disease. Attempting to minimize the risk for intended late-stage clinical development, we
therefore evaluated an experimental small-molecule RSV fusion inhibitor, JNJ-49214698, in
neonatal lambs, a fully-replicative animal model of RSV infection closely mimicking infant
RSV disease.
The three treatment regimens chosen in the study all reflect realistic potential clinical
drug administration regimens. The first group (Px) began to receive JNJ-49214698 one day
before viral nebulization to test the pre-exposure prophylactic effect of the compound, while
the Day 1 p.i. treatment group (Tx-1) was used to test the effect of very early post-exposure
treatment of an established infection. Previous RSV viral kinetic studies in our laboratory
demonstrated that RSV replication in neonatal lambs is highest between viral inoculation and
Day 3 p.i., reaching peak viral titer at Day 3, while RSV-associated lung pathology peaks at
Day 6 p.i. [32]. Therefore, the Day 3 post viral challenge treatment group (Tx-2) was used

70
to test the effect of JNJ-49214698 on RSV-associated ALRTI when treatment is started close
to peak viral titer, essentially the time patients seek first-line medical assistance and on
average 1 day before they present to hospital [36–40].
The overall assessments demonstrate a strong pre-exposure protective effect of JNJ-
49214698 against RSV infection. The prophylaxis (Px) group showed no gross/microscopic
lung lesions and no viral antigen by IHC. In addition, viral RNA levels significantly reduced
close to undetectable levels and were mirrored by no detectable IFFU[23]. Only very
occasional infection of individual lung cells, as demonstrated by the presence of viral RNA
(by RNAscope) in these cells was observed, consistent with earlier studies which showed
that fusion inhibitors are effective in inhibiting syncytia formation by preventing
transmission of the virus via cell-cell spreading [38,39]. Our data are also consistent with the
prophylactic efficacy of Synagis®, a humanized monoclonal antibody inhibiting the fusion
protein of RSV [8]. In contrast to Synagis®, which lacks therapeutic efficacy[8], early (Tx-
1) and late (Tx-2) therapeutic administration regimens with JNJ-49214698 displayed
unambiguous evidence of efficacy in the neonatal lambs. Administration of JNJ-49214698
after the establishment of a multi-cyclic ALRTI strongly reduced the severity of the infection
as evidenced by the strong and statistically significant reduction of gross and microscopic
changes in the lung, and decreased infectious titer in BALF as compared to vehicle-only
animals [23]. Consistent with these results, a considerable reduction of the viral RNA
expression in lung tissue was measured. However, the significant reduction of infectious
virus in BALF of therapeutically treated animals was not mirrored by a reduction of the viral
RNA to the same extent. This seeming discrepancy between viral RNA expression and
infectious viral titer in the BALF may be explained by the ability of JNJ-49214698 to bind

71
tightly to prefusion RSV F present on the envelope of the virus, resulting in neutralization of
the infectivity of virus particles present in the airways [22]. Although early administration of
JNJ-49214698, starting one day after RSV infection resulted in better reduction and clearing
of RSV infection as compared to the group that received late treatment starting at peak viral
load, the efficacy results obtained in the latter group were clearly noticeable.
Together, the results of this study demonstrate that pre-exposure prophylaxis and
treatment of an established RSV infection with a small-molecule RSV fusion inhibitor results
in significant reduction of viral replication as well as improvement of RSV-associated lung
pathology. Moreover, our data suggest a favorable window within which to treat RSV
infections in a community or hospital setting and contribute to the de-risking of late-stage
clinical compound development pathways.
References
1. Nair H, Nokes DJ, Gessner BD, et al. Global burden of acute lower respiratory infections
due to respiratory syncytial virus in young children: a systematic review and meta-
analysis. The Lancet. 2010; 375(9725):1545–1555.
2. Beigelman A, Bacharier LB. The role of early life viral bronchiolitis in the inception of
asthma. Curr Opin Allergy Clin Immunol. 2013; 13(2):211–216.
3. Régnier SA, Huels J. Association between respiratory syncytial virus hospitalizations in
infants and respiratory sequelae: systematic review and meta-analysis. Pediatr Infect Dis
J. 2013; 32(8):820–826.

72
4. Falsey AR, Hennessey PA, Formica MA, Cox C, Walsh EE. Respiratory Syncytial Virus
Infection in Elderly and High-Risk Adults. N Engl J Med. 2005; 352(17):1749–1759.
5. Widmer K, Zhu Y, Williams JV, Griffin MR, Edwards KM, Talbot HK. Rates of
Hospitalizations for Respiratory Syncytial Virus, Human Metapneumovirus, and
Influenza Virus in Older Adults. J Infect Dis. 2012; :jis309.
6. Ruuskanen O, Lahti E, Jennings LC, Murdoch DR. Viral pneumonia. The Lancet. 2011;
377(9773):1264–1275.
7. Anderson LJ, Dormitzer PR, Nokes DJ, Rappuoli R, Roca A, Graham BS. Strategic
priorities for respiratory syncytial virus (RSV) vaccine development. Vaccine. 2013; 31,
Supplement 2:B209–B215.
8. Brady MT, Byington CL, Davies HD, et al. Updated guidance for palivizumab
prophylaxis among infants and young children at increased risk of hospitalization for
respiratory syncytial virus infection. Pediatrics. 2014; 134(2):415–420.
9. Andabaka T, Nickerson JW, Rojas-Reyes MX, Rueda JD, Vrca VB, Barsic B.
Monoclonal antibody for reducing the risk of respiratory syncytial virus infection in
children. Evid-Based Child Health Cochrane Rev J. 2013; 8(6):2243–2376.
10. Malley R, DeVincenzo J, Ramilo O, et al. Reduction of Respiratory Syncytial Virus
(RSV) in Tracheal Aspirates in Intubated Infants by Use of Humanized Monoclonal
Antibody to RSV F Protein. J Infect Dis. 1998; 178(6):1555–1561.
11. Lagos R, DeVincenzo JP, Muñoz A, et al. Safety and antiviral activity of motavizumab, a
respiratory syncytial virus (RSV)-specific humanized monoclonal antibody, when
administered to RSV-infected children. Pediatr Infect Dis J. 2009; 28(9):835–837.

73
12. Ramilo O, Lagos R, Sáez-Llorens X, et al. Motavizumab treatment of infants hospitalized
with respiratory syncytial virus infection does not decrease viral load or severity of
illness. Pediatr Infect Dis J. 2014; 33(7):703–709.
13. Mazur NI, Martinón-Torres F, Baraldi E, et al. Lower respiratory tract infection caused
by respiratory syncytial virus: current management and new therapeutics. Lancet Respir
Med. 2015; 3(11):888–900.
14. Friedman JN, Rieder MJ, Walton JM, Canadian Paediatric Society, Acute Care
Committee, Drug Therapy and Hazardous Substances Committee. Bronchiolitis:
Recommendations for diagnosis, monitoring and management of children one to 24
months of age. Paediatr Child Health. 2014; 19(9):485–498.
15. DeVincenzo JP, McClure MW, Symons JA, et al. Activity of Oral ALS-008176 in a
Respiratory Syncytial Virus Challenge Study. N Engl J Med. 2015; 373(21):2048–2058.
16. DeVincenzo J, Lambkin-Williams R, Wilkinson T, et al. A randomized, double-blind,
placebo-controlled study of an RNAi-based therapy directed against respiratory syncytial
virus. Proc Natl Acad Sci. 2010; 107(19):8800–8805.
17. DeVincenzo JP, Whitley RJ, Mackman RL, et al. Oral GS-5806 Activity in a Respiratory
Syncytial Virus Challenge Study. N Engl J Med. 2014; 371(8):711–722.
18. Lapin C, Hiatt P, Langston claire, Mason E, Piedra P. A lamb model for human
respiratory syncytial virus infection. - Lapin - 1993 - Pediatric Pulmonology - Wiley
Online Library [Internet]. Wiley Online Libr. 1990 [cited 2018 Jun 20]. Available from:
https://onlinelibrary.wiley.com/doi/abs/10.1002/ppul.1950150305
19. Derscheid RJ, Ackermann MR. Perinatal Lamb Model of Respiratory Syncytial Virus
(RSV) Infection. Viruses. 2012; 4(10):2359–2378.

74
20. Ackermann MR. Lamb Model of Respiratory Syncytial Virus–Associated Lung Disease:
Insights to Pathogenesis and Novel Treatments. ILAR J. 2014; 55(1):4–15.
21. Derscheid RJ, Ackermann MR. The Innate Immune System of the Perinatal Lung and
Responses to Respiratory Syncytial Virus Infection. Vet Pathol Online. 2013; 50(5):827–
841.
22. Battles MB, Langedijk JP, Furmanova-Hollenstein P, et al. Molecular mechanism of
respiratory syncytial virus fusion inhibitors. Nat Chem Biol. 2016; 12(2):87–93.
23. Roymans D, Alnajjar SS, Battles MB, et al. Therapeutic efficacy of a respiratory
syncytial virus fusion inhibitor. Nat Commun [Internet]. 2017 [cited 2017 Aug 30]; 8.
Available from: http://www.ncbi.nlm.nih.gov/pmc/articles/PMC5537225/
24. Mackman RL, Sangi M, Sperandio D, et al. Discovery of an Oral Respiratory Syncytial
Virus (RSV) Fusion Inhibitor (GS-5806) and Clinical Proof of Concept in a Human RSV
Challenge Study. J Med Chem. 2015; 58(4):1630–1643.
25. A Study to Assess the Pharmacokinetics, Safety, and Tolerability of Multiple Doses of
Orally Administered JNJ-53718678 in Infants Hospitalized With Respiratory Syncytial
Virus (RSV) Infection - Full Text View - ClinicalTrials.gov [Internet]. [cited 2017 Mar
22]. Available from: https://clinicaltrials.gov/ct2/show/NCT02593851.
26. A Study of AK0529 in Infants Hospitalized With RSV - Full Text View -
ClinicalTrials.gov [Internet]. [cited 2017 Mar 31]. Available from:
https://clinicaltrials.gov/ct2/show/NCT02460016
27. Derscheid RJ, Geelen A van, Berkebile AR, et al. Increased Concentration of Iodide in
Airway Secretions Is Associated with Reduced Respiratory Syncytial Virus Disease
Severity. Am J Respir Cell Mol Biol. 2013; 50(2):389–397.

75
28. DeVincenzo JP, Wilkinson T, Vaishnaw A, et al. Viral Load Drives Disease in Humans
Experimentally Infected with Respiratory Syncytial Virus. Am J Respir Crit Care Med.
2010; 182(10):1305–1314.
29. Kim Y-I, DeVincenzo JP, Jones BG, et al. Respiratory Syncytial Virus Human
Experimental Infection Model: Provenance, Production, and Sequence of Low-Passaged
Memphis-37 Challenge Virus. PLOS ONE. 2014; 9(11):e113100.
30. Derscheid RJ, Geelen A van, McGill JL, et al. Human Respiratory Syncytial Virus
Memphis 37 Grown in HEp-2 Cells Causes more Severe Disease in Lambs than Virus
Grown in Vero Cells. Viruses. 2013; 5(11):2881–2897.
31. Grosz DD, Geelen A van, Gallup JM, Hostetter SJ, Derscheid RJ, Ackermann MR.
Sucrose stabilization of Respiratory Syncytial Virus (RSV) during nebulization and
experimental infection. BMC Res Notes. 2014; 7:158.
32. Larios Mora A, Detalle L, Van Geelen A, et al. Kinetics of Respiratory Syncytial Virus
(RSV) Memphis Strain 37 (M37) Infection in the Respiratory Tract of Newborn Lambs
as an RSV Infection Model for Human Infants. PLoS ONE [Internet]. 2015 [cited 2016
Jan 7]; 10(12). Available from: http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4671688/
33. Derscheid RJ, Gallup JM, Knudson CJ, et al. Effects of Formalin-Inactivated Respiratory
Syncytial Virus (FI-RSV) in the Perinatal Lamb Model of RSV. PLOS ONE. 2013;
8(12):e81472.
34. Olivier A, Gallup J, De Macedo MMMA, Varga SM, Ackermann M. Human respiratory
syncytial virus A2 strain replicates and induces innate immune responses by respiratory
epithelia of neonatal lambs. Int J Exp Pathol. 2009; 90(4):431–438.
35. Meyerholz DK, Grubor B, Fach SJ, et al. Reduced clearance of respiratory syncytial virus
infection in a preterm lamb model. Microbes Infect. 2004; 6(14):1312–1319.

76
36. Saleeby E, M C, Bush AJ, Harrison LM, Aitken JA, DeVincenzo JP. Respiratory
Syncytial Virus Load, Viral Dynamics, and Disease Severity in Previously Healthy
Naturally Infected Children. J Infect Dis. 2011; 204(7):996–1002.
37. Buckingham SC, Bush AJ, Devincenzo JP. Nasal quantity of respiratory syncytical virus
correlates with disease severity in hospitalized infants. Pediatr Infect Dis J. 2000;
19(2):113–117.
38. DeVincenzo JP, Saleeby E, M C, Bush AJ. Respiratory Syncytial Virus Load Predicts
Disease Severity in Previously Healthy Infants. J Infect Dis. 2005; 191(11):1861–1868.
39. Bagga B, Woods CW, Veldman TH, et al. Comparing influenza and RSV viral and
disease dynamics in experimentally infected adults predicts clinical effectiveness of RSV
antivirals. Antivir Ther. 2013; 18(6):785–791.
40. DeVincenzo JP, Aitken JB, Harrison LG. Opportunities for early therapy of respiratory
syncytial virus (RSV) infection: what happens before hospitalization. Antiviral Res.
2004; 62(1):47–51.

77
Figures and Legends
a) shows the daily plasma concentration (ng/mL) 24 hr after the first dose and daily just before the next
dose (Ctrough) to the end of the study. The compound reached a steady state concentration in the plasma
within 24 h of treatment and maintained that level throughout the study. b), c) show JNJ-49214698 exposure
in BALF (b) and lavaged lung (c) at necropsy (day 6 p.i.).
0 2 4 4 8 7 2 9 6 1 2 0 1 4 4
1
1 0
1 0 0
1 0 0 0
1 0 0 0 0
T im e (h )
Co
nc
en
tra
tio
n J
NJ
-49
21
46
98
(n
g/m
L)
Px
Tx-1
Tx-2
P la s m a
P x T x -1 T x -2
1
1 0
1 0 0
1 0 0 0
1 0 0 0 0
T r e a tm e n t g r o u p
Co
nc
en
tra
tio
n J
NJ
-49
21
46
98
(n
g/m
L) B A L F
P x T x -1 T x -2
1
1 0
1 0 0
1 0 0 0
1 0 0 0 0
T r e a tm e n t g r o u p
Co
nc
en
tra
tio
n J
NJ
-49
21
46
98
(n
g/g
lu
ng
)
L a va g e d -lu n g
a c b
Figure 3-1:Exposure of JNJ-49214698 in different body compartments of neonatal lambs.

78
1 2 3 4 5
0 .0
0 .5
1 .0
1 .5
D a y p o s t in fe c tio n
Cli
nic
al
sc
ore
V e h ic le a n d N o R S V G ro u p
a
V e h ic le
1 2 3 4 5
0 .0
0 .2
0 .4
0 .6
0 .8
1 .0
D a y p o s t in fe c tio n
Cli
nic
al
sc
ore
P x g ro u p
b
Px
1 2 3 4 5
0 .0
0 .1
0 .2
0 .3
0 .4
0 .5
D a y p o s t in fe c tio n
Cli
nic
al
sc
ore
T x 1 g ro u p
c
Tx1
1 2 3 4 5
0 .0
0 .1
0 .2
0 .3
0 .4
0 .5
D a y p o s t in fe c tio n
Cli
nic
al
sc
ore
T x 2 G ro u p
d
Tx2
Figure 3-2: Clinical signs score in different groups.
Data shown as average number of RSV-associated clinical signs observed in each group
(wheeze, increased respiratory rate and expiratory effort). Animals were infected with RSV
and received either vehicle only (Vehicle) and animal not infected with RSV (a), prophylaxis
(Px) (b) or therapeutic treatment with JNJ-49214698, starting one (Tx-1) (c) or three (Tx-2)
(d) days post infection. See online supplement for detailed table of the clinical signs
observed.

79
a) the viral RNA titer as measured by qRT-PCR in BALF, and b) the viral RNA titer as
measured by qRT-PCR in lung tissue, c) number of infectious particles as measured by IFFU
assay [23] with modification, all shown as average + SEM. Animals were infected with RSV
and received either vehicle only (Vehicle), prophylaxis (Px) or therapeutic treatment with
JNJ-49214698, starting one (Tx-1) or three (Tx-2) days post infection. Non-infected, vehicle-
treated animals were indicated as No RSV. JNJ-49214698 reduces the viable viral particles
in BALF, as well as RSV RNA in both BALF and lung tissue. Statistical analysis was
performed by one-way ANOVA, followed by Dunnett’s post-hoc test for multiple
comparison correction. *p-value < 0.02; **p-value < 0.002; ***p-value < 0.001; ****p-
value < 0.0001. LLOD = lower limit of detection.
N o R S V Ve h ic le P x T x -1 T x -2
0
1
2
3
4
5
Vir
al
tite
r (
log
10 F
FU
/mL
)
***
***
*****
****
L L O D
B A L Fc
N o R S V Ve h ic le P x T x -1 T x -2
2
3
4
5
6
7
8
Vir
al
lite
r (
log
10 R
NA
co
pie
s/m
l)
L L O D
****
****
B A L Fa
N o R S V Ve h ic le P x T x -1 T x -2
3
4
5
6
7
Vir
al
lite
r (
log
10 R
NA
co
pie
s/g
lu
ng
)L L O D
*
***
**
lungb
Figure 3-3:Effect of JNJ-49214698 on viral titers in BALF and lung tissue at Day 6 p.i.

80
a) integer-based scoring of M37 antigen in lung tissue detected by immunohistochemistry,
b) integer-based scoring of M37 RNA in lung tissue detected by RNAscope. Scoring on y-
axis represents the number of bronchioles or alveoli stained positive for M37 antigen (a) or
RNA (b) Bars represent the average score per group + SEM. Animals were infected with
RSV and received either vehicle only (Vehicle), prophylaxis (Px) or therapeutic treatment
with JNJ-49214689, starting one (Tx-1) or three (Tx-2) days post-infection. Non-infected,
vehicle-treated animals were indicated as No RSV. c) and d) show representative images of
M37 antigen (c) and M37 RNA (d) in the lung tissue of animals allocated to the different
treatment groups. JNJ-49214698 reduced the RSV M37 antigen and RNA in the lung of all
JNJ-49214698-treated animals. Statistical analysis was performed by Kruskall-Wallis non-
parametric test, followed by Dunn’s post-hoc test for multiple comparison correction. p-
value bars are representative for both bronchioles and alveoli. *p-value < 0.05; **p-value <
0.01.
N o R S V Ve h ic le P x T x -1 T x -2
0
1
2
3
4
M3
7 a
nti
ge
n s
co
re
B ro n c h io li
A lveo li
**
*
*
a
N o R S V Ve h ic le P x T x -1 T x -2
0
1
2
3
4
5
B ro n c h io li
A lveo li
M3
7 R
NA
sc
ore
*
**
b
RSV Px
Tx-1 Tx-2
RSV Px
Tx-1 Tx-2
c d
Figure 3-4: Effect of JNJ-49214698 on viral antigen and RNA in lung tissue at Day 6 p.i.

81
a) shows the average + SEM % of lung surface affected by RSV-induced gross lesions in the
different treatment groups. Animals were infected with RSV M37 and received either vehicle
only (Vehicle), prophylaxis (Px) or therapeutic treatment with JNJ-49214689, starting one
(Tx-1) or three (Tx-2) days post infection. Non-infected, vehicle-treated animals were
indicated as No RSV. JNJ-49214698 significantly reduces the RSV gross lesions in the
treated groups in comparison to the vehicle group. Statistical analysis was performed by one-
way ANOVA, followed by Dunnett’s post-hoc test for multiple comparison correction.
****p-value < 0.0001. b) shows representative images of lungs from an animal infected with
RSV receiving vehicle only, prophylaxis with JNJ-49214698, or therapeutic treatment,
starting one (Tx-1) or three (Tx-2) days post-infection and Non-infected, vehicle-treated
animals were indicated as Baseline. Yellow arrows indicate variably sized dark red
consolidated areas on the lung surface.
B a s e lin e Ve h ic le P x T x -1 T x -2
0
1 0
2 0
3 0
4 0
5 0
Av
era
ge
% o
f to
tal
lun
g w
ith
le
sio
ns
********
********
a
Baseline
Tx-1
Px Vehicle
Tx-2
b
Figure 3-5: Effect of JNJ-49214698 on development of gross lung lesions at Day 6 p.i.

82
a) shows the average + SEM of the accumulative RSV-induced microscopic lesions in the different
treatment groups. Animals were infected with RSV M37 and received either vehicle only
(Vehicle), prophylaxis (Px) or therapeutic treatment with JNJ-49214689, starting one (Tx-1) or
three (Tx-2) days post infection. Non-infected, vehicle only-treated animals were indicated as No
RSV. JNJ-49214698 reduces the pathological changes associated with RSV infection. Statistical
analysis was performed by Kruskall-Wallis non-parametric test, followed by Dunn’s post-hoc test
for multiple comparison correction. *p-value < 0.05; **p-value < 0.01. b) shows representative
images of the lung tissue sections from animals allocated to the different treatment groups.
Px
Tx-2
RSV
Tx-1
a b
N o R S V Ve h ic le P x T x -1 T x -2
0
2
4
6
8
1 0
ac
um
ila
tiv
e l
un
g l
es
ion
sc
ore
**
***
No RSV
Figure 3-6:Effect of JNJ-49214698 on RSV-induced accumulated lung histopathology at Day
6 p.i.

83
Supplementary Data
Materials and Methods
Compound and Dosing
JNJ-49214698 was discovered and synthesized by Janssen Infectious Diseases (Beerse,
Belgium). The compound was formulated in 10% acidified hydroxypropyl-β-cyclodextrin
[10% HP-β-CD + HCl, pH 2 (vehicle)] at 6.25 mg/mL prior to dosing and stored throughout
the study at 4°C. The compound was dosed orally by catheter-mediated orogastric gavage at
4 mL/kg body weight (25 mg/kg) once daily. Dose selection in this study aimed for reaching
the highest possible, safe exposure in the animals in order to maximize the likelihood to
obtain efficacy while avoiding toxic side effects. A daily oral dose of 25 mg/kg was selected
based on the antiviral activity of JNJ-49214698 [EC50 = 0.4 ng/mL (0.8 nM) and EC90 = 2.4
ng/mL (4.8 nM)], JNJ-49214698 exposure levels obtained in neonatal lambs at different
doses during a separate pharmacokinetic (PK) study and an observed lack of toxicity of JNJ-
49214698 at least until Cmax = 17,167 ng/mL and AUC0-24h = 139,993 ng.h/mL in a 5-day
repeated dose rat tolerance study.
Animals
Twenty-one colostrum-deprived neonatal lambs (Suffolk, Polypay, Dorsett cross) aged
1-3 days and 2-7 kg body weights were obtained for this experiment. Animal use was
approved by the Institutional Animal Care and Use Committee of Iowa State University.
RSV-infected lambs were kept in a separate room from the non-infected animals in the
Livestock Infectious Disease Isolation Facility (LIDIF). These rooms have separate
ventilation units as well as separate entrances and exits to avoid any cross-contamination
between infected and non-infected lambs. Lambs were fed iodide-free lamb milk replacer

84
diet (Milk Products Inc., Chilton, WI, USA) [1], and were treated with Naxcel (Ceftiofur
sodium, Pfizer) intramuscular once daily to reduce/prevent secondary bacterial infections.
Experimental Design
Lambs were randomly assigned to five different groups. Three groups (Px, Tx-1 and
Tx-2) were infected with RSV and treated with JNJ-49214698. The first group (Px, n = 4)
was treated prophylactically, 1 day before RSV challenge and then daily afterward up until
Day 5 post infection (p.i.). The second (Tx-1, n = 5) and the third (Tx-2, n = 5) groups were
treated one day and three days after viral challenge and daily afterward up until Day 5 p.i.,
respectively. The vehicle group (n = 4), serving as a positive RSV control group, was infected
with RSV, but received treatment with vehicle only. The No RSV group (n = 3), served as
negative RSV control group, and was aerosolized with RSV-free, HEp-2 cell-conditioned
media and also received vehicle. All lambs were euthanized at Day 6 p.i. and all endpoints
measured after euthanasia.
RSV Infection
Lambs were infected with RSV strain M37, purchased from Meridian BioSciences
(Memphis, TN, USA). This strain is a wild type A RSV isolated from the respiratory
secretions of an infant hospitalized for bronchiolitis [2,3]. In our laboratory, M37 was grown
in HEp-2 cells and stored at -80°C in media containing 20% sucrose [4]. PARI LC Sprint™
nebulizers were used to administer virus or cell-conditioned control media (lacking RSV) to
each lamb [5]. Six mL of 1.27 x 107 Infectious Forming Unit (IFFU)/mL in media containing
20% sucrose or cell-conditioned mock media (also containing 20% sucrose) was nebulized
to each lamb over the course of 25-30 minutes resulting in the total inhalation of about 3 mL
by each lamb.

85
Animal Monitoring for Appearance of Clinical Signs and the Clinical Score
Animals were monitored for clinical signs in the beginning of the study right before
viral inoculation (Day 0: all animals scored 0) and immediately after each administration of
vehicle or test article throughout the course of the study. In addition to the monitoring of
their behavior, respiratory associated clinical signs (respiratory rate, wheezing, expiratory
effort) were measured. Because clinical signs are variable in lambs and difficult to score in
terms of severity, an accumulative clinical score to summarize the overall distress per group
was applied by adding up the number of the scored clinical sings observed in each of the
individual lambs and then averaging the daily obtained sums for the respective groups to
normalize for group size.
Blood sampling for PK Analysis
Blood samples (1.5 - 2 mL) were collected from the jugular vein pre-dose (just before
the first dose) and at 24 h following each dose until 144 h for PK analyses. Blood was
dispensed into 3 mL blood collection tubes containing K2EDTA anticoagulant. Blood
samples were kept at room temperature prior to centrifugation. The blood samples were then
centrifuged at 1,600 x g for 10 min at 4°C to obtain the plasma. Plasma was stored in 2 mL
cryovials at -80°C.
Lung Collection and Processing
The thorax was opened, lungs removed and gross lesions were scored as performed
previously (40). The lungs were also photographed in situ and ex vivo. After removal,
percentage parenchymal involvement was scored for each lung lobe before the
bronchoalveolar lavage fluid (BALF) collection procedure. Left and right lungs were then
separated and each lobe excised. Tissue samples were collected from each lung lobe of all

86
animals. In brief, one sample from each lobe not destined for BALF collection (i.e. 4 lobes -
Right Cranial, Left Cranial, Left Middle and Left Caudal) were snap-frozen in liquid nitrogen
for qRT-PCR. Two samples from each of these lobes were placed in tissue cassettes and put
in 10% neutral-buffered formalin (NBF) for histological and immunohistochemical analysis.
Representative lung samples from each of these lobes were also placed into a cryovial and
immediately snap-frozen in liquid nitrogen, then transferred to -80°C for storage and shipped
on dry ice pellets for PK analysis of JNJ-53718678 compound at Janssen.
BALF Collection
BALF samples from each animal were collected immediately after euthanasia on Day
6 p.i. from right middle and right caudal lobes as performed previously in our laboratory [6].
Briefly, the excised lung lobes were instilled with 5 mL of cold DMIM (42.5% Iscove's
modified Dulbecco's medium, 7.5% glycerol, 1% heat-inactivated FBS, 49% DMEM, and 5
μg/ml kanamycin sulfate). 100 μL of the right caudal lobe BALF was added to 1 mL TRIzol
(Invitrogen) and kept in – 80 C for the qRT-PCR assay to assess RSV mRNA, and the rest
of BALF sample was placed on ice and used within 2 h for infectious focus-forming unit
(IFFU) assay to assess the infectious RSV titer. The right middle lobe BALF sample placed
-80°C for storage and shipped on dry ice pellets for determination of JNJ-49214698 and
blood urea nitrogen concentrations at Janssen.

87
Quantification of JNJ-49214698 Exposure
Samples of plasma (50 µL), lung homogenate (50 µL) and BALF (50 µL) were analyzed for
JNJ-49214698 using a method based on protein precipitation and HPLC/MS/MS analysis. To each
sample, DMSO (50 µL) and acetonitrile (500 µL) was added. Samples were mixed thoroughly
(mechanical shaking for 10 min), and then centrifuged (5000 g for 10 min at 15C). An aliquot
(400 µL) of the resulting supernatant was transferred to a 96-well plate and assayed for JNJ-
49214698 concentrations using HPLC/MS/MS employing positive-ion electrospray ionization
(Sciex API 4000) and a Waters ACQUITY UPLC C18 1.7um (50 x 2.1 mm i.d.) column. Elution
was achieved at a flow rate of 0.8 mL/min with a gradient of 0.1% FA and acetonitrile. The lower
limit of quantification was 1 ng/mL for plasma, 50 ng/g for lung and 1 ng/mL for BALF. The
assay was linear up to 20,000 ng/mL for plasma, 5,000 ng/mL for BALF and 100,000 ng/g for
lung. Samples of plasma and BALF were also analyzed for concentrations of urea in order to
calculate the dilution of BALF on sample collection. To a separate sample of plasma and BALF
(50 µL), 50 µL H2O/acetonitrile (50/50), 50 µL internal standard, 200 µl acetonitrile was added.
Samples were mixed thoroughly (mechanical shaking for 10 min), and then centrifuged (5000 g
for 10 min at 15C). An aliquot (200 µL) of the supernatant was transferred to a 96-well plate and
evaporated at 40C with nitrogen. An aliquot (200 µL) of camphanic chloride (1 mg/mL in
acetonitrile) was added for derivatization. Samples were mixed thoroughly (mechanical shaking
for 10 min) and incubated for 90 min at 37C. After incubation, 100 µL H2O was added and assayed
for urea concentrations using HPLC/MS/MS employing positive-ion electrospray ionization
(Sciex API 4000) and a Waters ACQUITY UPLC HSS T3 (50 x 2.1 mm i.d.) column. Elution
was achieved at a flow rate of 1 mL/min with a gradient of 0.01M Ammonium formate (pH = 3)

88
and acetonitrile. The lower limit of quantification was 2 µg/mL for plasma and BALF. The assay
was linear up to 10,000 µg/mL.
Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR)
Tissue samples from right and left cranial, left middle and left caudal lung lobes (0.3–
0.4 g of each lobe) were homogenized in TRIzol (Invitrogen, Carlsbad , CA) for total RNA
isolation according to manufacturer’s instructions and as previously described [4,6,7].
Briefly, RNA isolation was followed by DNase treatment (Ambion, TURBO DNase, Austin,
TX) at 1:10 dilution in combination with RNaseOUT (Invitrogen) and nuclease-free water
(GIBCO/Life Technologies, Carlsbad , CA). Spectrometry (Beckman Scientific,
Indianapolis, IN) was used to assess each RNA sample isolate at a dilution of 1:50 to measure
sample purity and quantity (A260nm/A280nm all >1.95). Agilent Bioanalyzer 2100 analyses
of the RNA isolates gave RNA Integrity Number values > 8.0. qRT-PCR was carried out
using One-Step Fast qRT-PCR Kit master mix (Quanta, BioScience, Gaithersburg, MD) in
a StepOnePlus™ qPCR machine (Applied Biosystems, Carlsbad, CA) in conjunction with
PREXCEL-Q assay-optimizing calculations [8,9]. Primers and probe for RSV M37
nucleoprotein were designed based on RSV accession number M74568. Forward primer: 5′-
GCTCTTAGCAAAGTCAAGTTGAACGA; reverse primer: 5′-
TGCTCCGTTGGATGGTGTATT; hydrolysis probe: 5′-6FAM-
ACACTCAACAAAGATCAACTTCTGTCATCCAGC-TAMRA. Each 1:10-diluted total
RNA sample was further diluted so that each final qRT-PCR contained 0.784 ng total
RNA/µL; as determined to be optimal by PREXCEL-Q [10]. Thermocycling conditions were
5 minutes at 50°C; 30 seconds at 95°C; and 45 cycles of 3 seconds at 95°C and 30 seconds
at 60°C. Samples and standards were assessed in duplicate and each qRT-PCR quantification

89
cycle (Cq) was converted to a relative initial quantity (Xo) based on a standard curve using
the following equation: Xo = EAMP(b-Cq), where EAMP and b are the PCR exponential
amplification efficiency value and the y-intercept, respectively, from a sample mixture
derived standard curve for RSV M37 nucleoprotein mRNA. The efficiency-corrected delta
Cq (EAMPΔCq) method was used for qRT-PCR quantification calculations. Results were
normalized to total tissue RNA loaded per reaction (identical for all reactions). No-RT
control reactions proved negative for RSV M37. qRT-PCR was demonstrably free of
inhibition based on preliminary dilution threshold analyses as per the PREXCEL-Q method
for qPCR [10,11].
Hematoxylin-Eosin Staining and Histological Scoring of Lung Sections
Hematoxylin-eosin stained sections were examined via light microscope as described
previously [6] with some modification. Lung lesions were scored according to an integer-
based score of 0-4 for each parameter (bronchiolitis, syncytial cells, epithelial necrosis,
epithelial hyperplasia, peribronchial lymphocytic infiltration, perivascular lymphocytic
infiltration, neutrophils), with 4 as the highest score. Then a final score (accumulative histo
lesion score) was assigned by adding up the scores from the seven individual parameters
resulting in final accumulative scores ranging from 0-28 representing the total RSV-
associated lesion in each tissue section. The scale used to assess the pathological changes
briefly, for bronchiolitis scale: 0 = no remarkable lesions, 1 = minimal detectable lesion
(epithelial degeneration in one or a few bronchioles per 20 X field), 2 = epithelial
degeneration involving less than 10% of the airway lumen; minimal neutrophils and cell
debris; adventitial lymphocytes in multiple bronchioles, 3 = epithelial degeneration involving
more than 10-50% of the airway lumen with cell debris and neutrophils; adventitial

90
lymphocytes; multiple bronchioles, 4 = circumferential bronchiolitis with dense adventitial
lymphocytes; multiple bronchioles. For syncytial cells scale: 0 = none, 1 = one distinct
syncytial cell, 2 = up to three in three 20x fields, 3 = more than three in three fields, 4 =
numerous. Epithelial necrosis and epithelial hyperplasia scale: 0 = none, 1 = minimally
detectable in one or a few bronchioles per 20x per field, 2 = 10% of the bronchioles in
multiple airways per field, 3 = 10-50% of the bronchioles in multiple airways per field, 4 =
circumferential in multiple airways. Neutrophils scale (in bronchi, bronchioles or alveoli): 0
= none, 1 = minimally detectable, 2 = 10 or less neutrophils in one or a few airways/alveoli,
3 = 10 or more neutrophils in several airways/alveoli, 4 = 10 or more involving many or most
airways/alveoli. The same scale was used for peribronchiolar lymphocytic infiltrates and
perivascular lymphocytic infiltrates: 0 = none, 1 = earliest detectable lymphocytic infiltration
in the adventitia, 2 = segmental to circumferential infiltration, 3 = circumferential infiltrates
that expand more than three cells wide, 4 = circumferential infiltrates that form nodules.
Immunohistochemistry (IHC) of Lung Sections
IHC for determining the distribution of RSV antigen was performed as described
previously [6,12,13]. Briefly, after deparaffinization and rehydration of the formalin fixed
paraffin-embedded tissue sections, antigen retrieval was performed by placing the slides with
tissue sections in pH 9.0 10mM TRIZMA base, 1mM EDTA buffer and 0.05% Tween 20
and boiling under pressure for up to 15 minutes. Polyclonal goat anti-RSV antibody
(Millipore/Chemicon, Temecula, CA; Cat. No. AB1128) was used as the primary antibody
after two blocking steps, the first with 3% bovine serum albumin in Tris-buffered saline +
0.05% Tween 20 (TBS-T) and the second with 20% normal swine serum in TBS-T, 15
minutes each. Primary antibody was followed by biotinylated rabbit anti-goat secondary

91
antibody (KP&L; Cat. No. 16-13-06). Signal development was done by using 1:200 dilution
of streptavidin-horseradish peroxidase (Invitrogen; Cat. No. 43-4323) for 30 minutes
followed by incubation with Nova Red chromagen solution (Vector; Cat. No. SK-4800). The
positive signal was quantified in both bronchioles and alveoli for each tissue section, and a
score of 0-4 was assigned according to an integer-based scale of 0 = no positive
alveoli/bronchioles, 1 = 1-10 positive alveoli/bronchioles, 2 = 11-39 positive
alveoli/bronchioles, 3 = 40-99 positive alveoli/bronchioles, 4 = >100 positive
alveoli/bronchioles.
RNAscope
Formalin fixed paraffin embedded (FFPE) IHC tissue sections were used for the
RNAscope detection of RSV mRNA in situ. A probe designed to the hRSV M37
nucleoprotein gene was used (Probe-V-RSV-NP, Advance Cell Diagnostic, Catalog number
439866). This probe was designed to target the 8-1111 base region of the nucleoprotein gene
of accession number KM360090.1 CDS sequence (1114-2289). The assay was performed
according to the manufacturer’s manual (user manual document number 320497;
RNAscope® 2.0 HD Detection Kit (BROWN) User Manual PART 2). Sections were
examined under light microscope, and the number of bronchioles and alveoli containing the
positive signal were counted. The number of positive bronchioles and alveoli per tissue
section was then assigned a score according to the simple integer-based scale of 0 = no
positive alveoli/bronchioles, 1 = 1-10 positive alveoli/bronchioles, 2 = 11-39 positive
alveoli/bronchioles, 3 = 40-99 positive alveoli/bronchioles, 4 = >100 positive
alveoli/bronchioles.

92
Infectious Focus-Forming Unit (IFFU) Assay
Viral titers in BALF from the right caudal lobe were determined by IFFU assay [6].
BALF samples were spun down for 5 minutes in a centrifuge at 3,000 x g to pellet large
debris. Approximately 800–850 μL of each supernatant was collected and then spun through
850 μL-capacity 0.45 μm Costar SPIN-X filter (microcentrifuge 15,600 x g) for 5 minutes.
The resulting clear BALF samples were applied to HEp-2 cells grown to 70% confluence in
12-well culture plates (Fisher Scientific, Hanover Park, IL) at full strength and three serial
dilutions (1:10, 1:100,and 1:1000), all tested in triplicate. The BALF samples in the wells
were diluted with DMEM media (Mediatech, Inc., Manassas, VA) supplemented with 10%
with heat-inactivated fetal bovine serum (FBS) (Atlanta Biologicals, Atlanta, GA) and 50
μg/mL kanamycin sulfate (Invitrogen/Life Technologies). Plates were incubated for 1 hr at
37°C, 5% CO2, then 1 mL of media was added to each well and plates were returned to the
incubator. After a 48 hr incubation, wells were fixed with cold 60% acetone/40% methanol
solution for 1 minute. Wells were then rehydrated with TBS-T, then blocked with 3% BSA
solution for 15 minutes followed by overnight incubation with primary polyclonal goat anti-
RSV (all antigens) antibody (EMD Millipore Corporation, Billerica, MA, USA). The next
day, plates were washed with TBS-T then incubated for 1 hr with secondary antibody (Alexa
Fluor® 488 F(ab’)2 fragment of rabbit anti-goat IgG (H+L) (Molecular Probes/Life
Technologies). Plates were rinsed and examined under inverted fluorescence microscopy
using the FITX/GFP filter (Olympus CKX41, Center Valley, PA), and total number of IFFUs
(which is defined here as 3 or more fluorescing cells) were counted for each well. IFFU/mL
calculations were obtained by multiplying the average number resulting from triplicate well
counts by the initial BALF sample dilution factor and multiplying that value by 5 to obtain
counts/mL since 1,000 μL/200 μl (the actual sample applied for each well) equals 5.

93
Statistical Analysis
Statistical analysis was completed by using the Kruskall-Wallis test for non-parametric
parameters such as accumulative microscopic lesion scoring, immunohistochemistry and
RNAscope integer-based scores, followed by Dunn’s post-hoc test for multiple comparisons.
One-way ANOVA followed by Dunnett multiple comparisons test was used to compare the
treated groups to the RSV-infected non-treated control group for gross lesion scores and viral
titer analyses by qRT-PCR and IFFU assays.

94
Table 1: clinical signs observed in lambs in different groups throughout the
study period
RSV-associated symptoms
G
roup
Animal ID Day 1 Day 2 Day 3 Day 4 Day 5
Vehicle
1 W L R
2 D
3 N
D
L
4 L
5 L
Px 6 R
7 R
8
9
Tx-1
1
1
W R
1
2
1
3
1
4
1
5
Tx-2
1
6
1
7
1
8
W
1
9
2
0
E
e
No
RSV
2
1
2
2
2
3
W = wheeze, L = lethargy, R = increased respiratory rate, Ee = expiratory effort, D = Dead,
ND = nasal discharge.

95
Panels show the average + SEM of integer-based scoring of the lung microscopic criteria
induced by RSV infection. Animals were infected with RSV and received either vehicle only
(Vehicle), prophylaxis (Px) or therapeutic treatment with JNJ-49214689, starting one (Tx-1)
or three (Tx-2) days post infection. Non-infected, vehicle-treated animals were indicated as
No RSV. Assessed criteria were a) bronchiolitis, b) syncytial cell formation, c) epithelial
necrosis, and d) neutrophils. (see supplementary data for Figure 8, which shows epithelial
hyperplasia, peribronchiolar nodules, and perivascular nodules. JNJ-49214698 greatly reduces
all assessed criteria in all treated lambs in comparison to the RSV-infected vehicle-treated lambs.
Statistical analysis was performed by Kruskall-Wallis non-parametric test, followed by
Dunn’s post-hoc test for multiple comparison correction. *p-value < 0.05; **p-value < 0.01;
***p-value < 0.001.
N o R S V V e h ic le P x T x -1 T x -2
0 .0
0 .5
1 .0
1 .5
2 .0
Bro
nc
hio
liti
s s
co
re
**
**
N o R S V V e h ic le P x T x -1 T x -2
0 .0
0 .5
1 .0
1 .5
2 .0
Sy
nc
yti
al
ce
ll s
co
re
**
**
***
***
N o R S V V e h ic le P x T x -1 T x -2
0 .0
0 .5
1 .0
1 .5
2 .0
Ep
ith
eli
al
ne
cro
sis
sc
ore
**
**
**
*
N o R S V V e h ic le P x T x -1 T x -2
0 .0
0 .5
1 .0
1 .5
2 .0
Ne
utr
op
hil
sc
ore
**
*
**
a
dc
b
Figure 3-7: Effect of JNJ-49214698 on RSV-induced lung histopathology at Day 6 p.i.

96
Panels show the average + SEM of integer-based scoring of the lung microscopic criteria
induced by RSV infection. Animals were infected with RSV and received either vehicle only
(Vehicle), prophylaxis (Px) or therapeutic treatment with JNJ-49214689, starting one (Tx-1)
or three (Tx-2) days post infection. Non-infected, vehicle-treated animals were indicated as
No RSV. Assessed criteria were a) epithelial hyperplasia, b) peribronchiolar nodules, and c)
perivascular nodules. JNJ-49214698 greatly reduces all assessed criteria in all treated lambs in
comparison to the RSV-infected vehicle-treated lambs. Statistical analysis was performed by
Kruskall-Wallis non-parametric test, followed by Dunn’s post-hoc test for multiple
comparison correction. *p-value < 0.05; **p-value < 0.01; ***p-value < 0.001.
N o R S V V e h ic le P x T x -1 T x -2
0 .0
0 .5
1 .0
1 .5
2 .0
Ep
ith
eli
al
hy
pe
rp
las
ia s
co
re
**
**
**
*
N o R S V V e h ic le P x T x -1 T x -2
0 .0
0 .5
1 .0
1 .5
2 .0
Pe
rib
ro
nc
hia
l ly
mp
ho
cy
tic
in
filt
ra
te s
co
re
**
**
*
N o R S V V e h ic le P x T x -1 T x -2
0 .0
0 .5
1 .0
1 .5
2 .0
Pe
riv
as
cu
lar l
ym
ph
oc
yti
c i
nfi
ltra
te s
co
re
**
**
a
c
b
Figure 3-8:Effect of JNJ-49214698 on RSV-induced lung histopathology at Day 6 p.i.

97
References
1. Derscheid RJ, Geelen A van, Berkebile AR, et al. Increased Concentration of Iodide in
Airway Secretions Is Associated with Reduced Respiratory Syncytial Virus Disease
Severity. Am J Respir Cell Mol Biol. 2013; 50(2):389–397.
2. DeVincenzo JP, Wilkinson T, Vaishnaw A, et al. Viral Load Drives Disease in Humans
Experimentally Infected with Respiratory Syncytial Virus. Am J Respir Crit Care Med.
2010; 182(10):1305–1314.
3. Kim Y-I, DeVincenzo JP, Jones BG, et al. Respiratory Syncytial Virus Human
Experimental Infection Model: Provenance, Production, and Sequence of Low-Passaged
Memphis-37 Challenge Virus. PLOS ONE. 2014; 9(11):e113100.
4. Derscheid RJ, Geelen A van, McGill JL, et al. Human Respiratory Syncytial Virus
Memphis 37 Grown in HEp-2 Cells Causes more Severe Disease in Lambs than Virus
Grown in Vero Cells. Viruses. 2013; 5(11):2881–2897.
5. Grosz DD, Geelen A van, Gallup JM, Hostetter SJ, Derscheid RJ, Ackermann MR.
Sucrose stabilization of Respiratory Syncytial Virus (RSV) during nebulization and
experimental infection. BMC Res Notes. 2014; 7:158.
6. Larios Mora A, Detalle L, Van Geelen A, et al. Kinetics of Respiratory Syncytial Virus
(RSV) Memphis Strain 37 (M37) Infection in the Respiratory Tract of Newborn Lambs
as an RSV Infection Model for Human Infants. PLoS ONE [Internet]. 2015 [cited 2016
Jan 7]; 10(12). Available from: http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4671688/
7. Derscheid RJ, Gallup JM, Knudson CJ, et al. Effects of Formalin-Inactivated Respiratory
Syncytial Virus (FI-RSV) in the Perinatal Lamb Model of RSV. PLOS ONE. 2013;
8(12):e81472.

98
8. Gallup JM, Ackermann MR. The ‘PREXCEL-Q Method’ for qPCR. Int J Biomed Sci
IJBS. 2008; 4(4):273–293.
9. Sow FB, Gallup JM, Sacco RE, Ackermann MR. Laser Capture Microdissection
Revisited as a Tool for Transcriptomic Analysis: Application of an Excel-Based qPCR
Preparation Software (PREXCEL-Q). Int J Biomed Sci IJBS. 2009; 5(2):105–124.
10. Polack FP, Teng MN, L.Collins P, et al. A Role for Immune Complexes in Enhanced
Respiratory Syncytial Virus Disease. J Exp Med. 2002; 196(6):859–865.
11. Delgado MF, Coviello S, Monsalvo AC, et al. Lack of antibody affinity maturation due to
poor Toll-like receptor stimulation leads to enhanced respiratory syncytial virus disease.
Nat Med. 2009; 15(1):34–41.
12. Olivier A, Gallup J, De Macedo MMMA, Varga SM, Ackermann M. Human respiratory
syncytial virus A2 strain replicates and induces innate immune responses by respiratory
epithelia of neonatal lambs. Int J Exp Pathol. 2009; 90(4):431–438.
13. Meyerholz DK, Grubor B, Fach SJ, et al. Reduced clearance of respiratory syncytial virus
infection in a preterm lamb model. Microbes Infect. 2004; 6(14):1312–1319.

99
THERAPEUTIC EFFICACY OF RESPIRATORY
SYNCYTIAL VIRUS NON-FUSION INHIBITOR (RSV-NFI) IN
NEONATAL LAMBS INFECTED WITH A HUMAN STRAIN OF RSV
A paper will be submitted to virology journal.
Sarhad S. Alnajjar1,2, Panchan Sitthicharoenchai, Jack M. Gallup1, David Lançois3, Peter Rigaux3,
Dirk Roymans3, Mark R. Ackermann1
1Department of Veterinary Pathology, College of Veterinary Medicine, Iowa State University,
Ames, Iowa USA
2Department of Veterinary Pathology, College of Veterinary Medicine, Baghdad University,
Baghdad, Iraq
3Janssen Infectious Diseases, Janssen Pharmaceutica NV, Beerse, Belgium
Summary
Respiratory Syncytial Virus (RSV) infects individuals of all ages and can cause severe lower
respiratory tract infection in infants and elderly. Given the burden of RSV disease and the lack of
any vaccine or efficacious therapeutic intervention, the discovery of new antiviral compounds are
in critically need. Therefore, the efficacy of RSV-NFI, a non-fusion RSV inhibitor, was tested for
efficacy in this study in the neonatal lamb model. Five groups of lambs were used to test the
efficacy of this compound at different concentrations. Three groups, TX1, TX2, and TX3, were
treated with RSV-NFI 24hr after RSV inoculation and once daily with 16, 4, or 1 mg/kg body
weight, respectively. The two other groups were treated with vehicle only after nebulization with
RSV or mock media. All lambs were euthanized at day 6 post-infection, which corresponds to the
expression of severe disease features in untreated animals. Pharmacokinetic analysis indicated that
RSV-NFI reached a suitable plasma concentration within 24 hr after administration, and this level
persist throughout the study that helped relate the plasma level to the antiviral efficacy. All tested

100
doses drastically reduced the appearance of RSV-induced gross and microscopic lesions. There
was a significant reduction in the viral load in the lung in both TX1 and Tx2 groups. Together;
these results demonstrate efficacy of the RSV-NFI in reducing RSV infection and inhibiting RSV-
induced lung pathology.
Introduction
Respiratory Syncytial Virus (RSV) is a common respiratory virus in all ages and leads to
moderate upper respiratory tract infection. However, RSV can progress to severe acute lower
respiratory tract infection (ALRTI) in children under the age of five, older adults, and immune
compromised individuals[1,2]. A systematic review and meta-analysis of RSV infection
worldwide estimated that 2.4 million episodes of severe RSV associated ALRTI of 33.8 million
total RSV infection each year[3]. Currently, there is no fully effective treatment or vaccine
available for RSV infection.
Recently, several antiviral compounds showed promising antiviral activity against RSV
ALRTI and some of these have been approved for human clinical trials [4]. Only two approved
treatments are commercially available including Palivizumab, which is a monoclonal antibody
against RSV fusion protein administered as prophylaxis for preterm infants and high-risk
individuals, and Ribavirin, a nucleoside analog antiviral drug [5,6]. Both of these compounds have
limited indication and therapeutic efficacy [7–10]. Developing RSV antiviral drugs generally fall
into three different categories: nucleoside inhibitors, fusion inhibitors, and non-fusion non-
nucleoside inhibitors [4]. A recent study with a fusion protein inhibitor indicated the possibility of
treating existing RSV infection even after peak viral load and lung pathology and demonstrate a
wide therapeutic intervention window for RSV [11].

101
Several models for RSV infection have been developed, and of these the neonatal lamb
model has some characteristics similar to those in infants with RSV lower respiratory tract
infection [12]. Lambs are susceptible to the human strain of RSV that causes moderate lower
respiratory tract infection [13], have similar lung size, lung anatomy, and cellular composition of
human infants, with similarity in immune responses and lung development. In this study, we
evaluate the therapeutic efficacy of RSV-NFI, which is a non-fusion non-nucleoside RSV
inhibitor, against RSV lower respiratory tract infection in the neonatal lamb model of RSV
infection.
Materials and Methods
Experimental Design
This study was completed in two parts in an experimentally identical environment. Animal
use was approved by the Institutional Animal Care and Use Committee of Iowa State University.
In the first part of the study, 2 high doses of RSV-NFI were tested with 4 experimental groups:
RSV infected group; RSV (n=3) treated with vehicle only, TX1 (n=2) treated with RSV-NFI
16mg/kg body weight and TX2 (n=4) treated with RSV-NFI 4 mg/kg body weight. The fourth
group (n=4) infected with mock media and treated with vehicle to serve as a negative control
group. In the second part of the study, the efficacy of RSV-NFI low dose (1mg/kg body weight)
was tested. 21 lambs were divided into 3 groups. There were two RSV infected groups: RSV (n=6)
treated with vehicle only, and TX3 (n=7) treated with 1mg/kg RSV-NFI one day after viral
nebulization, and a third group with no RSV (n=8) were nebulized with mock media and treated
with vehicle only (Figure 1). Lambs were nebulized with 6 ml hRSV strain M37 (1.27 x 107
Infectious Forming Unit (IFFU)/mL) on day 0. M37 is wild-type RSV A strain isolated from
human infant and commercially provided by Meridian BioSciences (Memphis, TN, USA)[14,15].

102
Treatments either with RSV-NFI or vehicle were started 24 hr after viral nebulization, then once
daily for 5 days. Lambs were monitored daily for the presence of clinical signs, and then all lambs
were humanely euthanized at day 6 post nebulization.
Tissue and Sample Collection
At necropsy (day 6 post nebulization), lambs were euthanized, and lungs were examined in
situ, removed, and the percent of the lung with lesion were determined. Lung tissues were collected
from right cranial, right middle, right caudal, and left cranial and placed in 10% neutral buffered
formalin for histologic evaluation, or snap frozen in liquid nitrogen for qRT-PCR and
pharmacokinetic study. Also, BALF samples were collected for left caudal lobe for infectious
focus forming unit assay (IFFU) and RT-qPCR assays, and from left middle lobe for compound
pharmacokinetic level assessment.
Infectious Focus-Forming Unit (IFFU) Assay
IFFU was completed on caudal lobe BALF, and homogenized lung tissue samples from the
4 collected lung lobes, and assays were performed as previously described in our laboratory [16].
Briefly, samples were pelleted and resultant supernate was spin again through a 0.45 μm Costar
SPIN-X filter (microcentrifuge 15,600 x g) for 5 minutes. The filtered samples were applied to
70% confluent HEp-2 cells in a 12-well culture plates (Fisher Scientific, Hanover Park, IL) at full
strength and three serial dilutions (1:10, 1:100, and 1:1000)in triplicate. After initial incubation at
37°C, 5% CO2 for 1 hr, 1 mL of media was added to each well and plates were returned to the
incubator and incubated for 48 hrs. plates, then, fixed with cold 60% acetone/40% methanol
solution for 1 minute followed by immunofluorescent staining. Primary and secondary antibody
were used. The primary was polyclonal goat anti-RSV (all antigens) antibody (EMD Millipore
Corporation, Billerica, MA, USA), and the secondary antibody was (Alexa Fluor® 488 F(ab’)2

103
fragment of rabbit anti-goat IgG (H+L) (Molecular Probes/Life Technologies). Plates examined
under inverted fluorescence microscopy using the FITX/GFP filter (Olympus CKX41, Center
Valley, PA), and the total number of IFFUs (which is defined here as 3 or more fluorescing cells)
were counted for each well. IFFU/mL calculations were obtained by multiplying the average
number resulting from triplicate well counts by the initial BALF sample dilution factor and
multiplying that value by 5 to obtain counts/mL since 1,000 μL/200 μl (the actual sample applied
for each well) equals 5.
Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR)
RSV RT-qPCR were completed on BALF and lung tissue. Lung samples from the 4 collected
lung lobe were homogenized in TRIzol and extracted for total RNA according to the
manufacturer's instruction. RNA samples were used for qRT-PCR as described previously[13,16–
18] to detect RSV, IP-10, MCP-1, MIP-1α, PDL-1, IFN-λ, IL-13, and RANTES mRNA
expression.
Histologic Evaluation
Microscopic evaluation of the tissue section were completed with hematoxylin and eosin
stained tissue sections, slides stained RSV antigen by immunohistochemistry, and slides processed
by RSV RNAscope to detect RSV mRNA. In H and E sections, RSV lesions were scored on a
scale of 0-4. These changes include bronchiolitis, syncytial cells, epithelial cells necrosis,
epithelial cells hyperplasia, neutrophils, bronchiolar nodules, and perivascular nodules. Then an
accumulative histological lesion score was determined by adding up the individualized score
resulting in a score of 0-28 that represent the overall pathological changes in each tissue section
(FI paper). Immunohistochemistry was completed on formalin fixed paraffin embedded tissue
sections and as described previously [16,19,20]. Briefly, the slides were deparaffinized and

104
rehydrated; then antigen retrieval step were completed using PH 9 10mM TRIZMA base, 1mM
EDTA buffer and 0.05 tween 20, then boiled and put in a steamer to maintain the temperature for
at least 30 minutes. Tissues were blocked with 3% BSA and 20% NSS for 15 minutes each,
followed by 1:500 dilution goat anti-RSV polyclonal antibody (Millipore/Chemicone Cat. No.
AB1128) and incubated in 4C overnight. The next day, tissue were washed 3 times with TBS-tw
followed by 1 hr incubation with 1:300 dilution of biotinylated Rabbit anti Goat secondary
antibody (KP&L Cat. No. 16-13-06). After 3 washes with TBS-tw, tissue were incubated with 3%
H2O2 for 25 minutes in room temperature followed by 2 TBS-tw washes and signal detection by
incubation with 1:200 dilution streptavidin-horseradish peroxidase (Invitrogen Cat. No. 43-4323)
followed by Nova Red staining solution. Slides were examined under light microscope and signal
were quantified in both alveoli and bronchioles. Score of 0-4 were applied according to an integer
based scale (of 0 = no positive alveoli/bronchioles, 1 = 1-10 positive alveoli/bronchioles, 2 = 11-
39 positive alveoli/bronchioles, 3 = 40-99 positive alveoli/bronchioles, 4 = >100 positive
alveoli/bronchioles). The RNAscope assay was also done on FFPE tissue sections for the detection
of RSV mRNA in situ by using a probe for RSV M37 nucleoprotein gene (Probe-V-RSV-NP,
Advance Cell Diagnostic, Catalog number 439866). The assay were done as described previously
( FI paper ) and according to the manufacturer protocol ((user manual document number 320497;
RNAscope® 2.0 HD Detection Kit (BROWN) User Manual PART 2). Microscopic tissue sections
were examined, and RNA signal were counted in both alveoli and bronchioles and applied for the
same integer based scale used to assess the IHC, which is 0 = no positive alveoli/bronchioles, 1 =
1-10 positive alveoli/bronchioles, 2 = 11-39 positive alveoli/bronchioles, 3 = 40-99 positive
alveoli/bronchioles, 4 = >100 positive alveoli/bronchioles.).

105
Statistics
When appropriate and unless specified otherwise, due to the limited number of data
observations in some treatment groups, the non-parametric Kruskal-Wallis test is applied to test
whether there is difference between the compound groups and the vehicle group. P-values are
adjusted with Bonferroni's multiple comparisons method. For the histological analysis of RSV
antigen producing cells and for the histopathological analysis, treatment group at a value of zero
were remove from the analysis (on/off patterns cannot be appropriately analyzed by statistics.
However, this pattern indicates the presence of a treatment effect). In the case of the histological
analysis of RSV antigen producing cells, scores of at least 1 were recoded to 1 and logistic
regression with random effects was applied for Slide and Animal to compare differences between
treatment Tx3 and Vehicle. In the case of the histopathology analysis, scores of at least 1 are
recoded to 1 and logistic regression with random effects for Animal was used to compare
differences between treatments and Vehicle results
Results
Pharmacokinetics of Different Doses of RSV-NFI in the Neonatal Lambs
Antiviral compounds need to reach an adequate and consistent exposure in the lung tissue in
order to protect against multicyclic RSV infection. First, we assessed RSV-NFI levels in the
plasma, then the compound distribution to the lung was evaluated. Compound absorption allowed
a rapid increase in plasma exposure within 1 hr. Steady state was not reached at 24h probably due
to the long half-life and slow elimination/metabolism of the compound. RSV-NFI plasma level at
24 hr after the first dose were respectively :5902ng/ml (16 mg/kg group), 871ng/ml (4 mg/kg
group), and 265ng/ml (1 mg/kg group:) with more than dose proportional increase of 5.0 and 4.6
was observed when comparing 1 to 4 mg/kg doses and 4 to 16 mg/kg doses, respectively (Figure:

106
2a). Once-daily treatment with 1, 4 or 16 mg/kg of RSV-NFI lead to an increase of Ctrough over
time across all different dose groups (3.6 fold at 1 mg/kg, 3.5 fold at 4 mg/kg, and 3.0 fold at 16
mg/kg). At necropsy, the RSV-NFI concentration was measured in both BALF representing the
airway lumen/ secretion, and in the whole lung homogenate representing lung tissue (cells,
interstitial tissue, and vasculature). The compound exposure in bronchoalveolar lavage fluid
(BALF) and homogenized-lungs tissue were the highest in the 16 mg/kg group (24431ng/ml,
36213ng/g, respectively). BALF exposure in 4 and 1 mg/kg group was (2091 ng/ml, 299 ng/ml,
respectively), and in the lung were (9740 ng/g, 2650 ng/g, respectively) (Figure 2 b & c).
Subsequently, lung/ plasma ratio of 1.3, 0.9, 0.9 in animal treated with 1, 4, and 16 mg/kg,
respectively, indicating a dose-dependent distribution of the compound to the lung compartments
and optimal conditions for investigating therapeutic exposure levels.
Viral Load in BALF and Lavaged-Lung Samples of Neonatal Lambs
The viral titer was determined at day 6 post-infection in BALF and homogenized lung tissue
samples from animals that were treated once-daily as from Day 1 post-infection with either vehicle
or different concentrations of RSV-NFI by the IFFU assay. There was a strong concentration-
dependent decrease of the production of infectious virions in BALF with 1.5 log10 reduction at 1
mg/kg, 4.2 log10 reduction at 4 mg/kg (p<0.01), and 4.5 log10 reduction at 16 mg/kg(p<0.01) (Figure
3 a). Concomitantly, lung titers of infectious virions were drastically reduced in all compound
treated groups. There was a 2.0 log10 reduction in the viral titer in 1 mg/kg treated group(p<0.01),
while full effect of 2.2 log10 reduction in groups treated with 4, and 16 mg/kg (p<0.01) & (p<0.05),
respectively (Figure 3b). To assess whether the reduction in the viable viral particle was
accompanied by a reduction in the viral RNA, a qRT-PCR was done on the same BALF samples
and lung tissues representative from four lung lobes. There was a great reduction in viral RNA

107
copies in both BALF and lung (Figure 3c and 2d). There was a significant 1.4 log10 decrease in
viral RNA at 1 mg/ml in BALF (p<0.0001) accompanied by 1 log10 reduction lung tissue. Superior
effects were observed at 4 mg/kg and 16 mg/kg treatment as is shown by 2.8 log10 reduction in
BALF(p<0.0001), accompanied by 4.4 log10 and 4 log10 reduction in lung tissue, respectively
((p<0.001) (p<0.0001) respectively). These results prove that the compound was efficient in
reducing the viral load in the treated lambs.
Treatment with RSV-NFI Significantly Reduced RSV-Induced Lung Lesion
Six days after infection, lungs from animals across different study groups were removed, and
the gross lesion developed by the lung upon RSV infection were scored. The RSV gross lesion
which was prominent in the RSV infected vehicle-treated group characterized by multifocal dark
red areas of consolidation on the lung surface and extend deep into the lung parenchyma in cut
section. In lungs of non-infected vehicle-treated animals, no gross lesions compatible with RSV
infection was seen. In contrast, in infected vehicle-treated animals, a significant level of the lung
surface area (32% on average) had gross lesions compatible with RSV infection (Figure 4 a and
b). Treatment with different doses of RSV-NFI drastically reduced the level of gross lesions
observed in all treated group when compared to the RSV infected vehicle-treated group. Gross
lesions were decreased to 0.76 % (p<0.05) or 0% (p<0.01) of lung surface when animals were
treated with 1 mg/kg or higher doses (4 and 16 mg/kg), respectively (Figure 4 a and b).
Microscopically, different concentrations of RSV-NFI resulted in a drastic reduction of all
assessed parameters and subsequently the accumulative histopathologic lesion score. The
histopathological lesion observed in the RSV infected vehicle-treated group was severe interstitial
pneumonia with bronchiolitis. There was inter-alveolar wall thickening due to inflammatory cell
infiltration, and epithelial cell hyperplasia with occasional necrotic epithelial cells and syncytia

108
formation leading to an accumulative histopathologic score of 11 (figure 5 a and b and figure 8).
The compound treatments significantly reduce the accumulative histopathologic lesion score in all
treated group (P<0.01) (Figure 5 a and b). 16 mg/kg reduced the accumulative histopathologic
score to 0.12, while 4mg/k and 1mg/kg treatments reduced the accumulative histopathologic score
to 0.6 and 1.2 (Figure 5 a and b). To further evaluate the degree of RSV infection in the lung tissue,
RSV immunohistochemical stain was done on FFPE tissue sections. RSV M37 antigen
demonstrated a dose-dependent decrease in the density of M37 antigen in the different lung lobes.
No RSV antigen detected in animals treated with 4 and 16 mg/kg (P<0.01 and P<0.05,
respectively) (Figure 6 a). There was a significant decrease in the RSV antigen expression in
bronchioles of animal treated with the low dose RSV-NFI (1 mg/kg)(p<0.05), with a high
reduction in the alveoli (P=0.059) indicating the efficiency of treatment with the study compound
to reduce RSV dissemination. Similar to what found in qRT-PCR of the BALF and lung tissue,
there was a significant reduction in RNA expression in situ in all RSV-NFI treated groups (P<0.01)
(Figure 7 b) as tested by RNAscope technique. The compound treatment drastically reduces RSV
mRNA in all treated groups with the most reduction was in 16 and 4 mg/kg treatment. Hence, the
RSV-NFI showed a strong effect in reducing RSV antigen and mRNA and subsequently RSV
associated lung lesion.
Treatment with RSV-NFI Significantly Decrease Inflammatory Chemokines
By qRT-PCR in lung at day 6 p.i. RSV-NFI greatly reduce mRNA expression of IP-10, MCP-
1, MIP-1α, PDL-1, and IFN-λ in comparison to the RSV infected vehicle-treated group in which
the expression of these chemokines were elevated (Figure 9 a-e). There was no difference shown
in the endogenous levels of IL-13 and RANTES mRNA expression (figure 9 f, g).

109
Discussion
Given the burden of RSV infection in infant and elderly, the development of active anti-RSV
therapeutic is urgently needed. Several antiviral drugs that target RSV have reached advanced
clinical trials. Most of these antiviral drugs are f monoclonal antibodies that inhibit fusion [4].
However, RSV resistant mutants can potentially form after few passages [21] and thus, additional
antiviral compounds with different mode of action that targets other viral structural component is
needed. In this study, we evaluate a non-fusion RSV inhibitor RSV-NFI, which is a non-
nucleoside replication inhibitor, in the neonatal lamb model. This study aims to assess the
therapeutic effect of different doses of RSV-NFI on established RSV associated acute lower
respiratory tract infection with treatment started one day after the viral nebulization and repeated
once daily up to 5 days. Our results demonstrated a potent antiviral activity as reflected by reduced
viral titer, mRNA expression, and RSV associated lesions in infected lambs.
Different concentrations of the compound resulted in different plasma exposure with more
than dose proportional when comparing each treatment with the one higher. However, the plasma
exposure was stable throughout the study with a tendency to increase over time. All the treatment
groups reached a concentration above the EC90 after 24 hr post-treatment except for 1mg/kg
treatment group which reached a concentration above the EC90 48hr after the first dose. This
plasma concentration leads to adequate lung exposure (at day 6 post-infection) which reflect the
concentration pattern seen in the plasma, i.e., high dose group produced a higher RSV-NFI
concentration in the lung tissue. Consistent RSV-NFI presence in the lung gives the compound the
ability to act on each new viral cycle reducing the RSV replication and cellular reinfection.
Lambs are permissive for hRSV and the human strains replicates well in lambs airways
causing acute lower respiratory tract infection [19,20,18,22]. Lambs nebulized with hRSV have

110
the peak viral load by day 3 of infection with the most lung lesions developing at day 4 to 6 post
nebulization [16]. RSV infected and vehicle-treated lambs in this study had high infective virus
and mRNA levels with distinct lesions in lung tissue characterized by interstitial pneumonia and
bronchiolitis. In contrast, treatment with different concentrations of RSV-NFI resulted in
consistent and significant reductions of all the assessed endpoints such as the reduction of lung
viral titers as measured by IFFU assay or qRT-PCR and reduced RSV associated lung lesions. The
results of both IFFU and qRT-PCR indicate the presence of viable RSV and whole viral load,
respectively, and were consistent and had the same pattern in BALF and lung tissue, and both had
a great reduction in the viral load with treatment that correlated with the RSV-NFI concentration.
Treatment with RSV-NFI as low as 1mg/kg caused a drastic reduction in the viral load in lung.
However, the most pronounced reduction occurred in 4 and 16 mg/kg groups. Several studies
indicated the positive relation between viral load and disease severity in infant [23–25]. Thus, the
observed reduction in the viral load in this study predicted to have a substantial effect in reducing
the pathological lesions associated with RSV by giving the lambs the ability to respond and repair
the tissue damage.
RSV associated gross and microscopic lesions were significantly decreased in all treated
animals. Almost no RSV lesions were seen at day 6 post-infection in the 4 and 16 mg/kg groups
with a great decline the lesions in the 1 mg/kg treated group. These results were consistent with
RSV immunohistochemistry and RNAscope results that had similar reduction trend in RSV
antigen and RSV RNA expression. Additionally, the increased inflammatory cell infiltration in
lung tissue of RSV infected vehicle treated lambs were accompanied by elevated level of MCP-1
and MIP-1α, and consistent with an earlier study in lamb indicated increase level of these
chemokines in RSV infected lambs at day 6 p.i. [18]. Several studies on RSV lower respiratory

111
tract infection in infants indicated that elevated levels of IP10, MIP-1α, and MCP-1 act as a
chemoattractant for inflammatory cells and subsequently associated with severe bronchiolitis and
hospitalization [26,27]. In contrast, an elevated level of PDL-1 in RSV infected vehicle treated
lambs may suggest a vital role in inhibiting lymphocytes antiviral response by modulating PDL-1
expression in infected epithelial cells [28,29]. The presence of minimal RSV antigen and mRNA
in the lung tissue section of the treated group indicate the ability of RSV-NFI to halt RSV
replication and infection of adjacent cells.
In summary, the compound under investigation had a strong effect on reducing RSV viral
load and lesions in all treated groups in comparison to the RSV infected vehicle-treated group.
This strong treatment effect occurred even when RSV-NFI was delivered to lambs one day after
viral nebulization, indicating the efficacy of the compound as a post infection therapeutic modality
to decrease the severity of lower respiratory tract infection after established RSV infection.
References
1. Falsey AR, Hennessey PA, Formica MA, Cox C, Walsh EE. Respiratory Syncytial Virus
Infection in Elderly and High-Risk Adults. N Engl J Med. 2005; 352(17):1749–1759.
2. Widmer K, Zhu Y, Williams JV, Griffin MR, Edwards KM, Talbot HK. Rates of
Hospitalizations for Respiratory Syncytial Virus, Human Metapneumovirus, and
Influenza Virus in Older Adults. J Infect Dis. 2012; :jis309.
3. Nair H, Nokes DJ, Gessner BD, et al. Global burden of acute lower respiratory infections
due to respiratory syncytial virus in young children: a systematic review and meta-
analysis. The Lancet. 2010; 375(9725):1545–1555.

112
4. Jorquera PA, Tripp RA. Respiratory syncytial virus: prospects for new and emerging
therapeutics. Expert Rev Respir Med. 2017; 11(8):609–615.
5. Thompson DF, Raebel MA, Hebert MF, Guglielmo BJ. What is the Clinical Role of
Aerosolized Ribavirin? DICP. 1990; 24(7–8):735–738.
6. Hynicka LM, Ensor CR. Prophylaxis and Treatment of Respiratory Syncytial Virus in
Adult Immunocompromised Patients. Ann Pharmacother. 2012; 46(4):558–566.
7. Chemaly RF, Shah DP, Boeckh MJ. Management of Respiratory Viral Infections in
Hematopoietic Cell Transplant Recipients and Patients With Hematologic Malignancies.
Clin Infect Dis. 2014; 59(suppl 5):S344–S351.
8. Ramilo O, Lagos R, Sáez-Llorens X, et al. Motavizumab treatment of infants hospitalized
with respiratory syncytial virus infection does not decrease viral load or severity of
illness. Pediatr Infect Dis J. 2014; 33(7):703–709.
9. Andabaka T, Nickerson JW, Rojas-Reyes MX, Rueda JD, Vrca VB, Barsic B.
Monoclonal antibody for reducing the risk of respiratory syncytial virus infection in
children. Evid-Based Child Health Cochrane Rev J. 2013; 8(6):2243–2376.
10. Lagos R, DeVincenzo JP, Muñoz A, et al. Safety and antiviral activity of motavizumab, a
respiratory syncytial virus (RSV)-specific humanized monoclonal antibody, when
administered to RSV-infected children. Pediatr Infect Dis J. 2009; 28(9):835–837.
11. Roymans D, Alnajjar SS, Battles MB, et al. Therapeutic efficacy of a respiratory
syncytial virus fusion inhibitor. Nat Commun [Internet]. 2017 [cited 2017 Aug 30]; 8.
Available from: http://www.ncbi.nlm.nih.gov/pmc/articles/PMC5537225/

113
12. Ackermann MR. Lamb Model of Respiratory Syncytial Virus–Associated Lung Disease:
Insights to Pathogenesis and Novel Treatments. ILAR J. 2014; 55(1):4–15.
13. Derscheid RJ, Geelen A van, McGill JL, et al. Human Respiratory Syncytial Virus
Memphis 37 Grown in HEp-2 Cells Causes more Severe Disease in Lambs than Virus
Grown in Vero Cells. Viruses. 2013; 5(11):2881–2897.
14. DeVincenzo JP, Wilkinson T, Vaishnaw A, et al. Viral Load Drives Disease in Humans
Experimentally Infected with Respiratory Syncytial Virus. Am J Respir Crit Care Med.
2010; 182(10):1305–1314.
15. Kim Y-I, DeVincenzo JP, Jones BG, et al. Respiratory Syncytial Virus Human
Experimental Infection Model: Provenance, Production, and Sequence of Low-Passaged
Memphis-37 Challenge Virus. PLOS ONE. 2014; 9(11):e113100.
16. Larios Mora A, Detalle L, Van Geelen A, et al. Kinetics of Respiratory Syncytial Virus
(RSV) Memphis Strain 37 (M37) Infection in the Respiratory Tract of Newborn Lambs
as an RSV Infection Model for Human Infants. PLoS ONE [Internet]. 2015 [cited 2016
Jan 7]; 10(12). Available from: http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4671688/
17. Derscheid RJ, Gallup JM, Knudson CJ, et al. Effects of Formalin-Inactivated Respiratory
Syncytial Virus (FI-RSV) in the Perinatal Lamb Model of RSV. PLOS ONE. 2013;
8(12):e81472.
18. Sow FB, Gallup JM, Olivier A, et al. Respiratory syncytial virus is associated with an
inflammatory response in lungs and architectural remodeling of lung-draining lymph
nodes of newborn lambs. Am J Physiol-Lung Cell Mol Physiol. 2010; 300(1):L12–L24.
19. Olivier A, Gallup J, De Macedo MMMA, Varga SM, Ackermann M. Human respiratory
syncytial virus A2 strain replicates and induces innate immune responses by respiratory
epithelia of neonatal lambs. Int J Exp Pathol. 2009; 90(4):431–438.

114
20. Meyerholz DK, Grubor B, Fach SJ, et al. Reduced clearance of respiratory syncytial virus
infection in a preterm lamb model. Microbes Infect. 2004; 6(14):1312–1319.
21. Cianci C, Yu K-L, Combrink K, et al. Orally Active Fusion Inhibitor of Respiratory
Syncytial Virus. Antimicrob Agents Chemother. 2004; 48(2):413–422.
22. Derscheid RJ, Geelen A van, Gallup JM, et al. Human Respiratory Syncytial Virus
Memphis 37 Causes Acute Respiratory Disease in Perinatal Lamb Lung. BioResearch
Open Access. 2014; 3(2):60–69.
23. Buckingham SC, Bush AJ, Devincenzo JP. Nasal quantity of respiratory syncytical virus
correlates with disease severity in hospitalized infants. Pediatr Infect Dis J. 2000;
19(2):113–117.
24. Saleeby E, M C, Bush AJ, Harrison LM, Aitken JA, DeVincenzo JP. Respiratory
Syncytial Virus Load, Viral Dynamics, and Disease Severity in Previously Healthy
Naturally Infected Children. J Infect Dis. 2011; 204(7):996–1002.
25. Hasegawa K, Jartti T, Mansbach JM, et al. Respiratory Syncytial Virus Genomic Load
and Disease Severity Among Children Hospitalized With Bronchiolitis: Multicenter
Cohort Studies in the United States and Finland. J Infect Dis. 2015; 211(10):1550–1559.
26. Tabarani CM, Bonville CA, Suryadevara M, et al. Novel Inflammatory Markers, Clinical
Risk Factors, and Virus Type Associated with Severe Respiratory Syncytial Virus
Infection. Pediatr Infect Dis J. 2013; 32(12):e437–e442.
27. Walsh EE, Peterson DR, Kalkanoglu AE, Lee FE-H, Falsey AR. Viral Shedding and
Immune Responses to Respiratory Syncytial Virus Infection in Older Adults. J Infect Dis.
2013; 207(9):1424–1432.

115
28. Telcian AG, Laza-Stanca V, Edwards MR, et al. RSV-Induced Bronchial Epithelial Cell
PD-L1 Expression Inhibits CD8+ T Cell Nonspecific Antiviral Activity. J Infect Dis.
2011; 203(1):85–94.
29. Zdrenghea MT, Johnston SL. Role of PD-L1/PD-1 in the immune response to respiratory
viral infections. Microbes Infect. 2012; 14(6):495–499.

116
Figures and Legends
Figure 4-1:Experimental design.
a) RSV-NFI dosing schedule. b) RSV nebulization, treatment, and blood draw timeline.
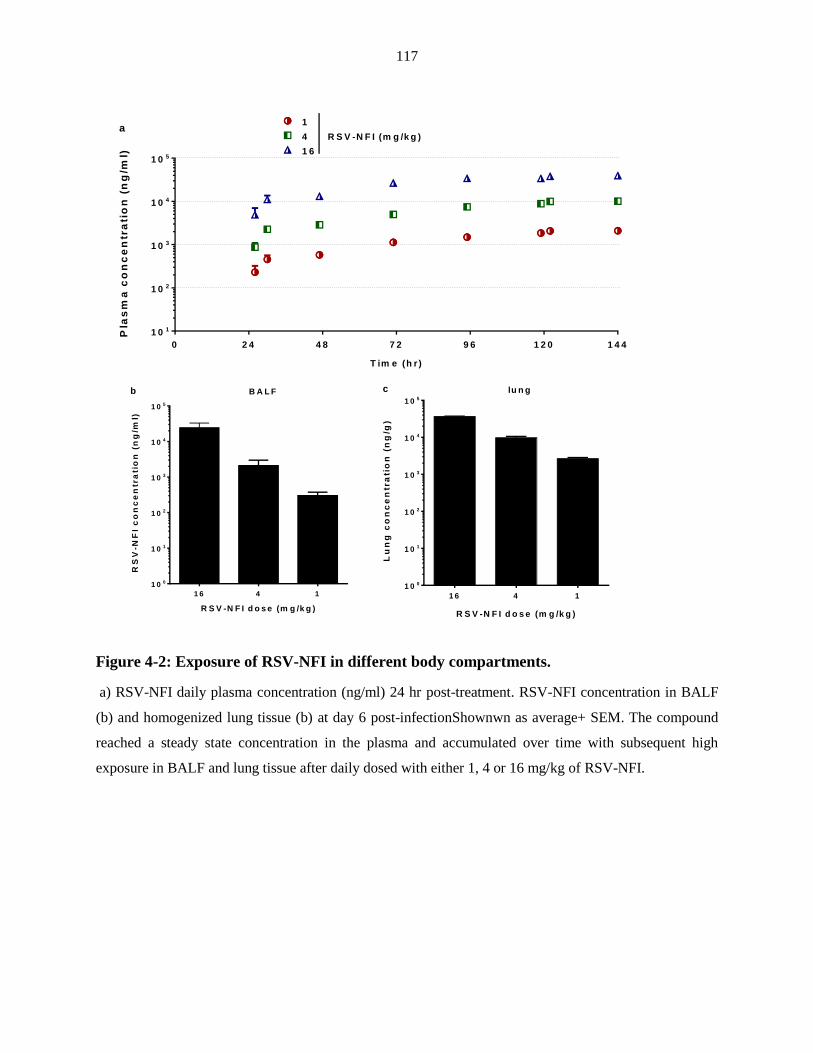
117
0 2 4 4 8 7 2 9 6 1 2 0 1 4 4
1 0 1
1 0 2
1 0 3
1 0 4
1 0 5
T im e (h r )
Pla
sm
a c
on
ce
ntr
ati
on
(n
g/m
l)
R S V -N F I (m g /k g )
1
4
1 6
a
1 6 4 1
1 0 0
1 0 1
1 0 2
1 0 3
1 0 4
1 0 5
R S V -N F I d o s e (m g /k g )
RS
V-N
FI
co
nc
en
tra
tio
n (
ng
/ml)
B A L Fb
1 6 4 1
1 0 0
1 0 1
1 0 2
1 0 3
1 0 4
1 0 5
R S V -N F I d o s e (m g /k g )
Lu
ng
co
nc
en
tra
tio
n (
ng
/g)
lu n gc
Figure 4-2: Exposure of RSV-NFI in different body compartments.
a) RSV-NFI daily plasma concentration (ng/ml) 24 hr post-treatment. RSV-NFI concentration in BALF
(b) and homogenized lung tissue (b) at day 6 post-infectionShownwn as average+ SEM. The compound
reached a steady state concentration in the plasma and accumulated over time with subsequent high
exposure in BALF and lung tissue after daily dosed with either 1, 4 or 16 mg/kg of RSV-NFI.

118
B a s e lin e 0 1 4 1 6
0
1
2
3
4
5
6
7V
iru
s t
ite
r (l
og
10 F
FU
/ml)
R S V -N F I (m g /k g )
n .s .
**
**
L L O D
1 .9
4 .2
4 .5a
B A L F
L L O D
B a s e lin e 0 1 4 1 6
1
2
3
4
5
6
7
Vir
us
tit
er
(lo
g1
0 F
FU
/g)
R S V -N F I (m g /k g )
**
**
*
2 .0
2 .2
2 .2
blung
B a s e lin e 0 1 4 1 6
6
8
1 0
1 2
Vir
us
tit
er
(lo
g1
0 R
NA
co
pie
s/m
l)
R S V -N F I(m g /k g )
n .s .***
*
1 .4
2 .8
2 .8
L L O Q
c B A L F
B a s e lin e 0 1 4 1 6
7
8
9
1 0
1 1
1 2
Vir
us
tit
er
(lo
g1
0 R
NA
co
pie
s/g
)
R S V -N F I (m g /k g )
n .s .
**
*
1 .0
4 .4
4 .0
L L O Q
dlung
Figure 4-3: Effect of RSV-NFI on viral titer in BALF and lung tissue homogenates at day 6
p.i.
a and b show the infectious viral titer as measured by IFFU assay in BALF (a) and in lung tissue (b). c and
d show the RSV mRNA titer as measured by qRT-PCR in BALF (c) and in lung tissue (d)All shown as
individual titer and mean. Treatment with RSV-NFI reduces the infectious viral titer and RNA in BALF
and Lung tissue. Animal infected with RSV and treated with either vehicle only (0) or different
concentration of RSV-NFI (1, 4, 16 mg/kg). Baseline lambs are not infected and treated with vehicle only.
*p<0.05, **p<0.01, ***p<0.001,****p<0.0001. n.s.: not significant.

119
a) Show the average % of lung tissue parenchyma involved with RSV gross lesion shown as individual %
and mean. b) Shows picture representative of gross lesion observed in different treatment group. RSV-NFI
greatly prevent the development of RSV associated gross lesion. Animal infected with RSV and treated
with either vehicle only (0) or different concentration of RSV-NFI (1, 4, 16 mg/kg). Baseline lambs are not
infected and treated with vehicle only.*p<0.05, **p<0.01.
Vehicle
4 mg/kg
1 mg/kg
16 mg/kg
Baseline
B a s e lin e 0 1 4 1 6
0
2 0
4 0
6 0
8 0
1 0 0
Pro
po
rtio
n o
f lu
ng
are
a
dis
pla
yin
g g
ro
ss
le
sio
ns
(%
)
R S V -N F I (m g /k g )
*
**
**
a b
Figure 4-4: Effect of RSV-NFI on the development of RSV associated gross lesion.

120
a) the accumulative histological lesion associated with RSV infection, which represents the sum of lesions
scored in RSV infected lung tissue which includes Bronchiolitis, Syncytial cells, Epithelial cells necrosis,
epithelial cells hyperplasia, neutrophils, Bronchiolar nodules, and perivascular nodules. b) a photo
representative for each treatment group. RSV-NFI greatly reduced the RSV associated microscopic lesion
in lung tissue. Animal infected with RSV and treated with either vehicle only (0) or different concentration
of RSV-NFI (1, 4, 16 mg/kg). Baseline lambs are not infected and treated with vehicle only. *p<0.05,
**p<0.01.
b 0mg/kg
4 mg/kg 16 mg/kg
Baseline
1 mg/kg
B a s e lin e 0 1 4 1 6
0
5
1 0
1 5
2 0
Ac
cu
mu
lati
ve
lu
ng
le
sio
n s
co
re
R C r
L C r
LM
L C d
R S V -N F I (m g /k g )
****
****
****
a
Figure 4-5: Effect of RSV-NFI on the development of RSV associated histologic lesion.

121
a) integer-based scoring of RSV M37 antigen in lung tissue detected by IHC had as average + SEM.
Treatment with RSV-NFI greatly reduces RSV antigen expression in lung tissue. b) shows photo
representation antigen distribution from different treatment group. Lung from lamb infected with RSV and
treated with either vehicle only (0) or different concentration of RSV-NFI (1, 4, 16 mg/kg). Baseline lambs
are not infected and treated with vehicle only. Estimated odds ratios: ***p<0.001. $: on/off pattern
indicating an obvious effect. n.s.: not significant.
0 mg/kg
Baseline
1 mg/kg
4 mg/kg
mg/kg
16 mg/kg
b
B a s e lin e 0 1 4 1 6
0
5 0
1 0 0
1 5 0
M3
7 a
ntig
en
sc
ore
R S V -N F I (m g /k g )
B ro n c h i
A lveo li
n .s .
* * *
$
$
$
$
a
Figure 4-6:Effect of RSV-NFI on RSV antigen.

122
.
a) integer-based scoring of RSV M37 RNA in lung tissue detected by RNAscope had as average + SEM.
Treatment with RSV-NFI greatly reduces RSV RNA expression in lung tissue. b) Picturesdepict findings
representation from different treatment group. Animal infected with RSV and treated with either vehicle
only (0) or different concentration of RSV-NFI (1, 4, 16 mg/kg). Baseline lambs are not infected and treated
with vehicle only. *p<0.05, **p<0.01. n.s.: not significant.
0 mg/kg
Baseline
1 mg/kg
4 mg/kg 16 mg/kg
b
B a s e lin e 0 1 4 1 6
0
5 0 0
1 0 0 0
1 5 0 0
Vir
al
RN
A+ c
ell
s
(c
ou
nts
/ 3
52
mm
2)
R S V -N F I (m g /k g )
B ro n c h i
A lveo li
n .s .* *
*
n .s .
*n .s .
a
Figure 4-7: Effect of RSV-NFI on RSV M37 RNA expression.

123
B a s e lin e 0 1 4 1 4
0 .0
0 .5
1 .0
1 .5
2 .0
2 .5
Bro
nc
hio
liti
s s
co
re
$
$
**
R S V -N F I (m g /k g )
B a s e lin e 0 1 4 1 4
0 .0
0 .1
0 .2
0 .3
0 .4
0 .5
Sy
nc
yti
al
ce
lls
sc
ore
$
$
$
R S V -N F I (m g /k g )
B a s e lin e 0 1 4 1 4
0 .0
0 .5
1 .0
1 .5
2 .0
Ep
ith
eli
um
ne
cro
sis
sc
ore
$
$
**
R S V -N F I (m g /k g )
B a s e lin e 0 1 4 1 4
0 .0
0 .5
1 .0
1 .5
2 .0
2 .5
Ep
ith
eli
um
hy
pe
rp
las
ia s
co
re
$
$
$
R S V -N F I (m g /k g )
B a s e lin e 0 1 4 1 4
0 .0
0 .5
1 .0
1 .5
2 .0
Ne
utr
op
hil
s s
co
re
$
*
**
R S V -N F I (m g /k g )
B a s e lin e 0 1 4 1 4
0 .0
0 .5
1 .0
1 .5
2 .0
2 .5
Pe
rib
ro
nc
hio
lar l
ym
ph
oc
yti
c
infi
ltra
tes
sc
ore
n .s .
*
R S V -N F I (m g /k g )
n .s .
B a s e lin e 0 1 4 1 4
0 .0
0 .5
1 .0
1 .5
2 .0
Pe
riv
as
cu
lar l
ym
ph
oc
yti
c
in
filt
ra
tes
sc
ore
n .s .
R S V -N F I (m g /k g )
$
n .s .
a b
d e
c
f
g
Figure 4-8: Effect of RSV-NFI on the development of RSV associated histologic lesion.
Graphs show an integer-based score of Bronchiolitis (a), Syncytial cells (b), Epithelial cells necrosis (c),
epithelial cells hyperplasia (d), neutrophils (e), Bronchial nodules (f), and perivascular nodules (g) RSV-
NFI greatly reduce the RSV associated microscopic lesion in lung tissue. Animal infected with RSV and
treated with either vehicle only (0) or different concentration of RSV-NFI (1, 4, 16 mg/kg). Baseline lambs
are not infected and treated with vehicle only. *p<0.05, **p<0.01, ***p<0.001,****p<0.0001. n.s.: not
significant. . $: on/off pattern indicating an obvious effect.

124
B a s e lin e 0 1 4 1 6
0
1 0 0 0
2 0 0 0
3 0 0 0
4 0 0 0
5 0 0 0
Re
lati
ve
IP
-10
ex
pre
ss
ion
(%
)
R S V -N F I (m g /k g )
n .s .
**
*
B a s e lin e 0 1 4 1 6
0
5 0 0
1 0 0 0
1 5 0 0
Re
lati
ve
MC
P-1
ex
pre
ss
ion
(%
)
R S V -N F I (m g /k g )
n .s .
**
*
B a s e lin e 0 1 4 1 6
0
1 0 0 0
2 0 0 0
3 0 0 0
4 0 0 0
Re
lati
ve
MIP
-1
ex
pre
ss
ion
(%
)
R S V -N F I (m g /k g )
n .s .
**
*
B a s e lin e 0 1 4 1 6
0
5 0 0
1 0 0 0
1 5 0 0
2 0 0 0
Re
lati
ve
PD
L-1
ex
pre
ss
ion
(%
)
R S V -N F I (m g /k g )
n .s .
**
*
B a s e lin e 0 1 4 1 6
0
1 0 0 0
2 0 0 0
3 0 0 0
21 0 0 6
41 0 0 6
61 0 0 6
Re
lati
ve
IF
N-
ex
pre
ss
ion
(%
)
R S V -N F I (m g /k g )
*
*
**
B a s e lin e 0 1 4 1 6
0
5 0
1 0 0
1 5 0
2 0 0
Re
lati
ve
IL
-13
ex
pre
ss
ion
(%
)
R S V -N F I (m g /k g )
n .s .
n .s .
n .s .
B a s e lin e 0 1 4 1 6
0
5 0
1 0 0
1 5 0
Re
lati
ve
CC
10
ex
pre
ss
ion
(%
)
R S V -N F I (m g /k g )
n .s .
n .s .
n .s .
B a s e lin e 0 1 4 1 6
0
1 0 0
2 0 0
3 0 0
4 0 0
5 0 0
Re
lati
ve
RA
NT
ES
ex
pre
ss
ion
(%
)
R S V -N F I (m g /k g )
n .s .
n .s .
**
a b c
d ef
g h
Figure 4-9: Effect of RSV-NFI inflammatory chemokine RNA expression in lung tissue.
Chemokine RNA expression IP-10 (a), MCP-1 (b), MIP-1α (c), PDL-1 (d), INF-λ (e), IL-13 (f), RANTES
(g). All are shown as average + SEM. Treatment with RSV-NFI greatly reduces IP-10, MCP-1, MIP-1α,
PDL-1, INF-λ. Animal infected with RSV and treated with either vehicle only (0) or different concentration
of RSV-NFI (1, 4, 16 mg/kg). Baseline lambs are not infected and treated with vehicle only. *p<0.05,
**p<0.01, ***p<0.001,****p<0.0001. n.s.: not significant.

125
CHAPTER 5. GENERAL CONCLUSION
Secondary Bacterial Co-Infection
The neonatal lamb model of RSV infection is well established, and many aspects of the
model were investigated in our laboratory such as different viral strains, different viral cultures,
different inoculation techniques, viral kinetics, immune response, and associated clinical signs [1–
6]. Investigating mechanisms of bacterial infections secondary to RSV infection is the next step to
understand the pathogenesis of bacterial coinfection and utilize the model to test the efficacy of
therapeutic agents in complex infections which are associated with enhanced disease severity and
death. This study demonstrated the susceptibility of lamb to Streptococcus pneumoniae (Spn)
infection, and the enhanced disease severity when lambs challenged with Spn 3 days after primary
RSV nebulization in comparison to lambs receiving either RSV or Spn alone. The titer of RSV
were higher in the RSV-Spn infected group than RSV alone, while there were no differences
between Spn infected lambs in Spn lung titer. There was an increase in severity of lung lesions in
the RSV-Spn group, which was characterized by a higher number of infiltrating neutrophils, and
increased epithelial necrosis and degeneration. These findings were in agreement other studies of
viral-bacterial co-infection, and demonstrated the enhanced infection by both pathogens.
Efficacy of Anti-RSV Fusion Inhibitor
The neonatal lamb model of RSV infection which mimics RSV infection in infants provides
an excellent tool to evaluate the efficacy of antiviral compound on the development of RSV-
associated acute lower respiratory tract infection. This study demonstrated the efficacy of JNJ-
49214698, which is an RSV small molecule fusion inhibitor that binds to the RSV prefusion
conformation preventing critical F protein transformation to postfusion conformation and

126
subsequently preventing multicyclic RSV infection. In three treatment regimen,JNJ-49214698
had a very protective effect when given prophylactically 1 day before RSV nebulization. These
lambs had no infectious RSV or mRNA in the lung, and had minimal clinical signs, and almost no
RSV associated lung lesion. Although prophylactic administration of JNJ-49214698 is not its
intended use, the finding demonstrates the ability of fusion inhibitor to prevent RSV infection.
When JNJ-49214698 given 1 day and 3 days post-RSV nebulization, it had significant decrease in
infectious RSV in the BALF with a less pronounced reduction in the mRNA. There were
reductions in RSV associated clinical signs, gross and microscopic lung lesion, and RSV protein
and mRNA expression in lung tissue. The effect of JNJ-49214698 was more evident when given
1 day after RSV nebulization. These data suggest a protective effect of JNJ-49214698 in reducing
RSV replication and subsequently RSV associated signs and lesions. Therefore, the compound is
a promising antiviral candidate against RSV infection in infants.
Efficacy of Anti-RSV Non-Fusion Inhibitor
Another mechanistic type of anti-RSV compounds are the non-fusion inhibitors, which bind
and inhibit viral structural protein other than F protein. In this study, we evaluated the efficacy of
the non-fusion inhibitor JNJ-64417184 in the neonatal lamb model of RSV infection. In this study,
3 different doses were used to treat lambs 1 day after RSV nebulization (16, 4, or 1 mg/kg body
weight). The study demonstrated that JNJ-64417184 markedly reduced all RSV associated lesions
in treated lambs in comparison to RSV infected vehicle-treated lambs. Additionally, the compound
significantly reduced infectious RSV titer and mRNA expression in BALF and lung tissue
homogenate and markedly reduced RSV antigen and mRNA expression in lung tissue sections.
These results were accompanied by a reduction in immunomodulatory cytokines IP-10, MCP-1,

127
MIP-1α, PDL-1, and IFN-λ, and correlated with the reduction of inflammatory cells in the lung
tissue observed in histological evaluation. These results indicate the ability of JNJ-64417184 to
reduce RSV replication and disease progress.
Future Directions
The neonatal lamb model of RSV infection in infants is a well-characterized model and
currently used to evaluate anti-RSV therapeutics[7,8]. We demonstrated that neonatal lambs
infected with RSV are also susceptible for infection with Spn and lead to enhance disease severity
in lambs infected with both RSV-Spn. Several aspects of this model can be modified and further
developed in future studies of viral-bacterial co-infection such as: expanding the length of time of
infection, assessing different inoculation time points of Spn following RSV, assessing different
doses of Spn, and different types of Spn delivery (e.g., intranasal, intrabronchial, aerosolization).
Also regarding Spn, a colonization model similar to human and mice could be tested to evaluate
whether such mechanism of infection is a possibility in the lamb model. A third consideration is
to try assess other bacterial pathogens or combinations of:Staphylococcus aureus, Streptococcus
pyogenes, and Haemophilus influenza. These three bacterial pathogens are the most common
bacteria identified secondarily to viral infections in human.
Several characteristics make the neonatal lamb model an interest and compelling option for
pharmaceutical companies to assess their compounds. Although our laboratory has inoculated
lambs by various routes including intra-tracheal, bronchoscopy guided intra-bronchial inoculation,
nebulization of the virus has several advantages [2,5,6]. RSV nebulization provides more
homogenous distribution of the virus to different lung lobes and subsequently homogenized lung
pathology[5,9]. RSV nebulization is one feature of the neonatal lamb model that provides
consistency for RSV associated lower respiratory tract infection and does overwhelm areas of RSV

128
infection as can occur with intrabronchial deposition. We demonstrated that two antiviral
compounds which target fusion protein (entry of the virus) and non-fusion protein (replication of
the virus) had a strong effect in reducing RSV replication and associated pathology. In this same
fashion, other viral targets could be tested in the lamb model such as RSV N, SH, and L proteins.
Other potential studies could include a combination of compound that target 2 or more RSV protein
and different time points for treatment administration. However, treating within 2 days post viral
inoculation may be ideal because this likely is the time an infant may seek medical
intervention[10].
References
1. Derscheid RJ, Gallup JM, Knudson CJ, et al. Effects of Formalin-Inactivated Respiratory
Syncytial Virus (FI-RSV) in the Perinatal Lamb Model of RSV. PLOS ONE. 2013;
8(12):e81472.
2. Larios Mora A, Detalle L, Van Geelen A, et al. Kinetics of Respiratory Syncytial Virus
(RSV) Memphis Strain 37 (M37) Infection in the Respiratory Tract of Newborn Lambs
as an RSV Infection Model for Human Infants. PLoS ONE [Internet]. 2015 [cited 2016
Jan 7]; 10(12). Available from: http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4671688/
3. Meyerholz DK, Grubor B, Fach SJ, et al. Reduced clearance of respiratory syncytial virus
infection in a preterm lamb model. Microbes Infect. 2004; 6(14):1312–1319.
4. Sow FB, Gallup JM, Olivier A, et al. Respiratory syncytial virus is associated with an
inflammatory response in lungs and architectural remodeling of lung-draining lymph
nodes of newborn lambs. Am J Physiol-Lung Cell Mol Physiol. 2010; 300(1):L12–L24.

129
5. Derscheid RJ, Geelen A van, McGill JL, et al. Human Respiratory Syncytial Virus
Memphis 37 Grown in HEp-2 Cells Causes more Severe Disease in Lambs than Virus
Grown in Vero Cells. Viruses. 2013; 5(11):2881–2897.
6. Olivier A, Gallup J, De Macedo MMMA, Varga SM, Ackermann M. Human respiratory
syncytial virus A2 strain replicates and induces innate immune responses by respiratory
epithelia of neonatal lambs. Int J Exp Pathol. 2009; 90(4):431–438.
7. Detalle L, Larios A, Gallup L, Van Geelen A, Duprez L, Stohr T. Delivery of ALX-0171
by inhalation greatly reduces disease burden in a neonatal lamb RSV infection model. 9th
Int Respir Syncytial Virus Symp Cape Town South Afr Abstr OP 72. 2014.
8. Olivier AK, Gallup JM, Geelen A van, Ackermann MR. Exogenous administration of
vascular endothelial growth factor prior to human respiratory syncytial virus a2 infection
reduces pulmonary pathology in neonatal lambs and alters epithelial innate immune
responses. Exp Lung Res. 2011; 37(3):131–143.
9. Grosz DD, Geelen A van, Gallup JM, Hostetter SJ, Derscheid RJ, Ackermann MR.
Sucrose stabilization of Respiratory Syncytial Virus (RSV) during nebulization and
experimental infection. BMC Res Notes. 2014; 7:158.
10. Saleeby E, M C, Bush AJ, Harrison LM, Aitken JA, DeVincenzo JP. Respiratory
Syncytial Virus Load, Viral Dynamics, and Disease Severity in Previously Healthy
Naturally Infected Children. J Infect Dis. 2011; 204(7):996–1002.


















