
THE TRANSITION FROM TADPOL TO FROEG HAEMOGLOBIN DURING NATURAL AMPHIBIAN METAMORPHOSIS ·...
14
J. Cell Sci. 16, 143-156 (i974) 143 Printed in Great Britain THE TRANSITION FROM TADPOLE TO FROG HAEMOGLOBIN DURING NATURAL AMPHIBIAN METAMORPHOSIS II. IMMUNOFLUORESCENCE STUDIES J. BENBASSAT Department of Medicine A, Hadassah University Hospital, Jerusalem, Israel SUMMARY Rabbits were immunized with frog or tadpole haemoglobin purified either by chromato- graphy or by polyacrylamide gel electrophoresis. The obtained rabbit antisera were shown to be specific for frog or tadpole haemolysates by double diffusion, immunoprecipitation and immunonuorescence. Indirect immunofluorescent staining of peripheral blood smears of Rana catesbeiana tadpoles at the metamorphic climax revealed that 16 % of the red cells were stained with antibodies against frog haemolysates, while almost all of them (98 %) were stained with antibodies against tadpole haemolysates. These results are compatible with the possibility that some of the circulating red cells in metamorphosing tadpoles contain both tadpole and frog haemoglobins. INTRODUCTION During amphibian metamorphosis, the transition from tadpole to frog haemoglobin is associated with the appearance of a poorly haemoglobinized, morphologically immature microcyte in the circulation (Moss & Ingram, 1968; Vankin, Brandt & DeWitt, 1970), which incorporates actively precursors of protein and nucleic acid synthesis (Moss & Ingram, 1968; Benbassat, 1970; Thiel, 1970), and which contains frog haemoglobin (DeWitt, 1968). These findings indicate that the switch from tadpole to frog haemoglobin involves a replacement of the tadpole erythrocytes by a population of cells containing frog haemoglobin. The question, whether these 2 populations belong to different clones or to the same cell line is subject to controversy. Human embryonic and foetal haemoglobins (Kleihauer, Tang & Betke, 1967) as well as human foetal and adult haemoglobins (Betke & Kleihauer, 1958; Dan & Hagiwara, 1967; Gitlin, Sasaki & Vuopio, 1968) may coexist in the same cell. There is also evidence that frog and tadpole haemoglobins are present simultaneously in some of the erythrocytes of metamorphosing and adult anaemic Xenopus laevis (Jurd & Maclean, 1970; Maclean & Jurd, 1971). On the other hand, however, ithasbeen found that red cells of metamorphosing Rana catesbeiana tadpoles contain either tadpole or frog haemoglobins but not both (Rosenberg, 1970; Maniatis & Ingram, 19716). This paper reports the results of immunofluorescence studies of the content of individual erythrocytes of Rana catesbeiana tadpoles and frogs by means of specific rabbit antibodies against tadpole and frog haemolysates. It is shown that during
Transcript of THE TRANSITION FROM TADPOL TO FROEG HAEMOGLOBIN DURING NATURAL AMPHIBIAN METAMORPHOSIS ·...

J. Cell Sci. 16, 143-156 (i974) 143Printed in Great Britain
THE TRANSITION FROM TADPOLE TO FROG
HAEMOGLOBIN DURING NATURAL
AMPHIBIAN METAMORPHOSIS
II. IMMUNOFLUORESCENCE STUDIES
J. BENBASSAT
Department of Medicine A, Hadassah University Hospital,Jerusalem, Israel
SUMMARY
Rabbits were immunized with frog or tadpole haemoglobin purified either by chromato-graphy or by polyacrylamide gel electrophoresis. The obtained rabbit antisera were shown to bespecific for frog or tadpole haemolysates by double diffusion, immunoprecipitation andimmunonuorescence. Indirect immunofluorescent staining of peripheral blood smears of Ranacatesbeiana tadpoles at the metamorphic climax revealed that 16 % of the red cells were stainedwith antibodies against frog haemolysates, while almost all of them (98 %) were stained withantibodies against tadpole haemolysates. These results are compatible with the possibility thatsome of the circulating red cells in metamorphosing tadpoles contain both tadpole and froghaemoglobins.
INTRODUCTION
During amphibian metamorphosis, the transition from tadpole to frog haemoglobinis associated with the appearance of a poorly haemoglobinized, morphologicallyimmature microcyte in the circulation (Moss & Ingram, 1968; Vankin, Brandt &DeWitt, 1970), which incorporates actively precursors of protein and nucleic acidsynthesis (Moss & Ingram, 1968; Benbassat, 1970; Thiel, 1970), and which containsfrog haemoglobin (DeWitt, 1968). These findings indicate that the switch from tadpoleto frog haemoglobin involves a replacement of the tadpole erythrocytes by a populationof cells containing frog haemoglobin. The question, whether these 2 populationsbelong to different clones or to the same cell line is subject to controversy. Humanembryonic and foetal haemoglobins (Kleihauer, Tang & Betke, 1967) as well ashuman foetal and adult haemoglobins (Betke & Kleihauer, 1958; Dan & Hagiwara,1967; Gitlin, Sasaki & Vuopio, 1968) may coexist in the same cell. There is alsoevidence that frog and tadpole haemoglobins are present simultaneously in some ofthe erythrocytes of metamorphosing and adult anaemic Xenopus laevis (Jurd &Maclean, 1970; Maclean & Jurd, 1971). On the other hand, however, ithasbeen foundthat red cells of metamorphosing Rana catesbeiana tadpoles contain either tadpole orfrog haemoglobins but not both (Rosenberg, 1970; Maniatis & Ingram, 19716).
This paper reports the results of immunofluorescence studies of the content ofindividual erythrocytes of Rana catesbeiana tadpoles and frogs by means of specificrabbit antibodies against tadpole and frog haemolysates. It is shown that during

144 J- Benbassat
metamorphosis some of the circulating red cells are stained by both types of antisera.This rinding is consistent with the possibility that tadpole and frog haemoglobins areproduced by erythroid cells belonging to the same red cell line.
METHODS
The source of the animals and the methods for blood collection, preparation of labelledhaemolysates, polyacrylamide gel electrophoresis, reduction and alkylation of red cell proteinsand electrophoresis in 8 M urea have been described previously (Benbassat, 1974).
Preparation of the immunogen
Red cells oiRana catesbeiana adult frogs and premetamorphic tadpoles (total length 55-70 mm)were washed 3 times with amphibian Ringer solution (Rugh, 1962). The visible buffy coatwas removed after each centrifugation, and the washed cells were lysed in 10 volumes of0-0015 M MgClj. The stroma, nuclei and the ribosomes were removed by centrifugation at105000g for 90 min at 5 °C. About 2 ml of the ribosome-free supernatant were applied to acolumn of CM-cellulose (Serva) and washed through with o-oi M sodium phosphate buffer(pH 70) until the eluate was clear and without absorbance at 280 nm. Then the haemoglobinwas eluted with 0-5 M Na2HPO4 (pH 95), and used as immunogen.
Immunization
One ml of the eluted haemoglobin solution containing about 1 mg of the immunogen wasmixed with complete Freund's adjuvant (Difco Laboratories, Detroit, Michigan), thoroughlyemulsified and injected at different sites in the footpads and skin of adult New Zealand whiterabbits. In some experiments the immunogen consisted of the major tadpole or frog haemo-globin fractions obtained by polyacrylamide gel electrophoresis (Benbassat, 1974). The haemo-globin bands were cutout, thoroughly homogenized as described by Maniatis& Ingram (1971a),and injected into the rabbits; 4 injections were given at 2- to 3-week intervals. One week afterthe last injection, the rabbits were bled by cardiac puncture. The sera were separated and keptfrozen until used.
Immunoprecipitation of labelled haemolysates14C-labelled haemolysates, prepared as detailed previously (Benbassat, 1974) from tadpole
or frog erythrocytes, were incubated at 4 °C for 48 h with antisera prepared as described inImmunization (above). After incubation the mixtures were spun for 10 min at 5000J* in arefrigerated centrifuge. The obtained precipitate was washed 3 times in cold Ringer'ssolution, dissolved in formic acid and mounted on planchets for determination of radioactivity.In other experiments, the immunoprecipitates were dissolved in deionized 8 M urea, reduced,alkylated and electrophoresed on polyacrylamide gels prepared in alkaline 8 M urea. All theimmune precipitations were carried out at antibody excess as judged by standard immuneprecipitation curves (Fig. 6).
Staining of tadpole and frog blood cells with fluorescent antibodies
The indirect method of Weller & Coons (1954) was used. Air-dried smears were preparedfrom washed blood cells of early premetamorphic tadpoles, animals at the metamorphic climax,and adult frogs, and left at —20 °C until used. The smears were covered with antisera againsttadpole or frog haemoglobins and incubated for 30 min at 37 °C with continuous shaking. Serumof non-immunized rabbits was used in control experiments. The coated smears were thenwashed 3 times with amphibian Ringer's solution and covered for 30 min with FluoresceinConjugated Goat anti (Rabbit IgG) globulin (Microbiological Associates, Inc., Bethesda)diluted 1:60 in amphibian Ringer's solution. They were then washed again, dried and examinedin u.v. illumination under glycerol buffered to pH 70 with sodium phosphate. Cell counts wereperformed from photographs of the smears.

Synthesis of red cell protein during metamorphosis. II 145
RESULTS
Determination of the specificity of the antisera
The characterization of the antisera was performed by double diffusion on agar,immunoprecipitation and characterization of the precipitated material by polyacryl-amide gel electrophoresis in 8 M urea, and immunofluorescence.
Double diffusion on agar. Agar-gel diffusion plates were used. The reagents con-sisted of undiluted antisera and of tadpole or frog haemoglobin solutions. The haemo-globin solutions consisted of either unfractionated haemolysates or purified majorhaemoglobin fractions after separation by polyacrylamide gel electrophoresis.
Sera of rabbits immunized with tadpole haemolysates purified by chromatographyproduced one precipitin line when tested against haemolysates of prometamorphictadpoles, animals at the metamorphic climax or against the purified major tadpolehaemoglobin (T-J; the same antisera did not react when tested against frog or mousehaemolysates (Fig. 4). Sera of rabbits immunized with the purified major tadpolehaemoglobin fraction (Tj), produced one precipitation line when tested againstunfractionated tadpole haemolysates, purified major tadpole haemoglobin (Tx) andthe purified minor tadpole haemoglobin fraction (T2). The type of reaction obtainedsuggested an identity between the antigens (Fig. 5).
Rabbit antisera against frog haemoglobin reacted with unpurified haemolysates ofanimals during the metamorphic climax and of adult frogs, as well as with the purifiedfrog haemoglobin fractions (Fx and F3). One precipitation line was observed in allcases. The same antisera did not react when tested against tadpole or mouse haemo-globins (Figs. 6, 7).
These results confirm the findings by Maniatis & Ingram (1971a) and by Jurd &Maclean (1970), that frog and tadpole haemoglobins are relatively strong immunogensfor the rabbit. The absence of any detectable cross-reaction between these haemo-globins is consistent with the findings by Baglioni & Sparks (1963), Moss & Ingram(1968) and Aggarwal & Riggs (1969), that the transition from tadpole to frog haemo-globin involves the beginning of synthesis of a completely different set of polypeptidechains. The major tadpole haemoglobin (Tj) cross-reacted with the minor tadpolehaemoglobin (T2), while the major frog haemoglobin (F3) cross-reacted with the minorone (Fj); this could indicate that these haemoglobin fractions share one or morecommon subunits. Haemolysates of animals at the metamorphic climax reacted withantisera against either tadpole or frog haemoglobin; this is in agreement with thefinding that tadpoles at this stage of metamorphosis contain both types of haemoglobin(Fig. 8).
Immunoprecipitation. The amount of antiserum needed to provide an excess ofantibodies in the reaction mixtures was determined by standard immune precipitationcurves. In these experiments a constant amount (3-4 /tg) of 14C-labelled haemoglobinwas titrated with increasing volumes (5-500 /A) of antiserum. The relative amount ofantiserum which precipitated the maximal amount of radioactivity was used in furtherexperiments (Fig. 1).
In three different experiments, 76, 83 and 89% of the total radioactivity ofIO C EL 16

146 J. Benbassat
100 -
20
200 300 400 500it\ antiserum added
900 1000
Fig. 1. Immunoprecipitation of tadpole (solid line) and frog (dashed line) 14C-labelled haemolysates by rabbit anti-tadpole hb and anti-frog hb antisera, respectively.The indicated amount of antiserum was incubated for 48 h at 4 CC with a constantamount of 14C-labelled haemolysate containing 3 fig haemoglobin. After incubationthe immunoprecipitate was washed, dissolved in formic acid and the radioactivitydetermined in a Nuclear Chicago Gas Flow Counter.
Table 1. Immunoprecipitation of tadpole and frog labelled haemolysates
Antigen
Unfractionatedtadpole haemo-lysate
Purified majortadpole haemo-globin (Tj)
Unfractionatedfrog haemo-lysate
Purified majorfrog haemo-globin (F3)
T T
globin,/*g
3 2
1 7
4 i
2-O
TCA-precipitable
cpm
640 (100%)
137(100%)
318(100%)
112(100%)
Anti-T*
356(56%)
82(60%)
21(6%)
5(4%)
cpm precipitated by
Anti-F*
14 (3 %)
6(4%)
240 (76 %)
79(7O%)
Controlserum*
10(2%)
7 (5 %)
24(7%)
3 (3 %)
* Anti-T and anri-F refer to non-absorbed rabbit antisera against tadpole or frog haemo-lysates purified by chromatography on CM-cellulose. The control consisted of the serum ofa non-immunized rabbit.

Synthesis of red cell protein during metamorphosis. II 147
unfractionated frog haemolysates could be precipitated by antisera against frog haemo-globin. The highest amounts of radioactivity, which could be precipitated by antiseraagainst tadpole haemoglobin from unfractionated haemolysates were 56, 53 and 38%.Similar results were obtained when the purified major haemoglobin fractions wereused as antigens in the reaction mixture. The amounts of tadpole haemoglobin pre-cipitated by antibodies against frog haemoglobin and vice versa did not exceed thoseprecipitated by sera of non-immunized rabbits. The results of a representativeexperiment are given in Table 1.
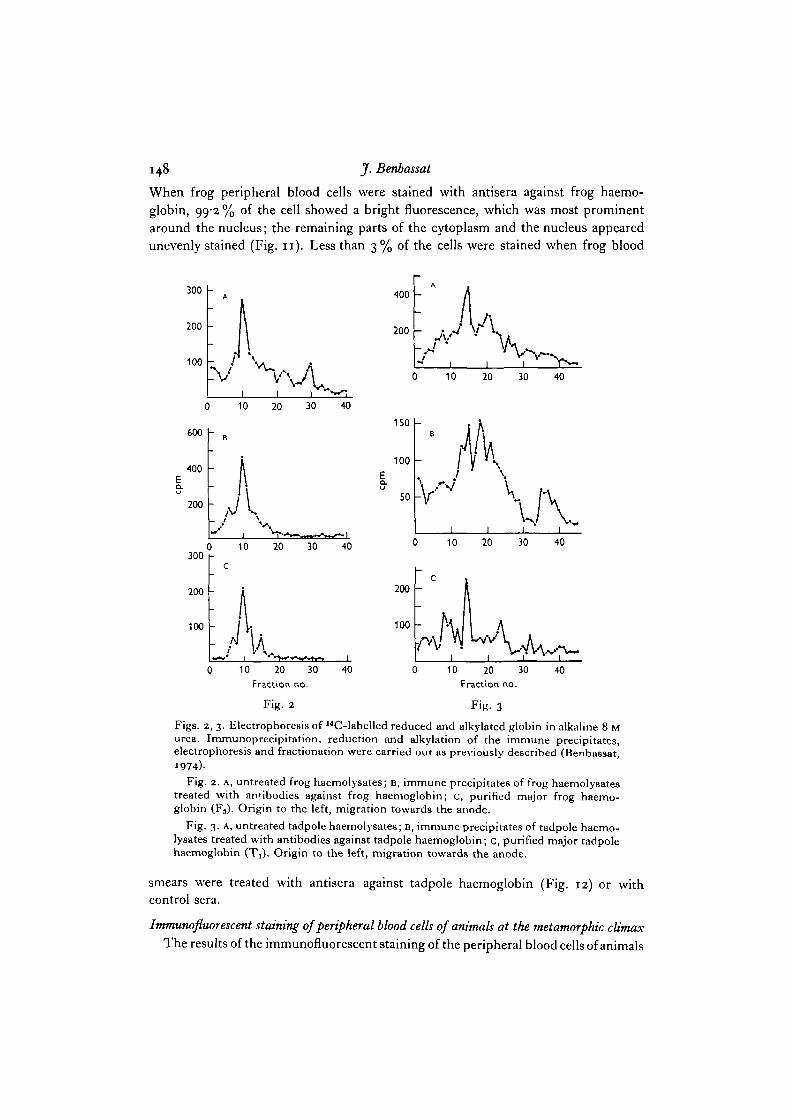
In order to confirm that the antisera reacted with haemoglobin and not with anyother red cell protein, which might have contaminated the immunogen, the immuno-precipitated material was characterized by polyacrylamide gel electrophoresis. Forthis purpose, immunoprecipitated haemolysates were dissolved in 8 M urea, in orderto dissociate between the antibody and the labelled antigen. Possible S—S bonds werereduced by the addition of mercaptoethanol and blocked by amidation. Then thedissociated protein subunits were resolved by electrophoresis on polyacrylamide gelsprepared in alkaline 8 M urea. After electrophoresis the gels were sliced and the radio-activity was determined in each of the gel fractions. The radioactive profiles of theimmunoprecipitated material were compared to those of the reduced and alkylatedpurified major haemoglobin components. As shown in Figs. 2 and 3, the purifiedmajor haemoglobin components of both tadpoles and frogs were resolved into onemajor and several smaller peaks of radioactivity (Figs. 2C and 3 c). The major peakconsisted of both globin chains, which could not be separated electrophoretically underthese conditions. The smaller peaks represented probably small amounts of con-taminating red cell (haemoglobin or non-haemoglobin) protein. The pattern ofresolution of the immunoprecipitated protein of frog haemolysates was almost identicalwith that of reduced and alkylated globin prepared from the purified major frog haemo-globin fraction (Fig. 2). The immunoprecipitated protein of tadpole haemolysatesresolved into several peaks of radioactivity, one of which coelectrophoresed with themajor tadpole reduced and alkylated globin (Fig. 3). The nature of the remainingpeaks of immunoprecipitated radioactivity remains unclear: they could be globinsubunits of the minor tadpole haemoglobins, or non-haemoglobin red cell proteins.Similar results were obtained whether the immunogen employed in the preparationof the antisera was purified by chromatography or by polyacrylamide gel electro-phoresis.
Immunofiuorescence. The antisera against frog or tadpole haemoglobins character-ized in the two preceding sections, were used for immunofluorescent staining aftercross-absorption with tadpole and frog lysed red cells, respectively. The degree ofspecificity of the antisera employed is shown in Figs. 9-12 and in Table 2. When peri-pheral blood smears of premetamorphic tadpoles were stained with antisera againsttadpole haemoglobin, 98-7 % of the cells showed a bright fluorescence, which was evenlydistributed throughout the cytoplasm; the nucleus was not stained (Fig. 9). Onlyoccasional cells (less than 2 %) were stained when tadpole blood smears were treatedwith antisera against frog haemoglobin (Fig. 10), normal rabbit serum or antiserumagainst tadpole haemoglobin, which had been absorbed with tadpole haemoglobin.
148 J. Benbassat
When frog peripheral blood cells were stained with antisera against frog haemo-globin, 99-2 % of the cell showed a bright fluorescence, which was most prominentaround the nucleus; the remaining parts of the cytoplasm and the nucleus appearedunevenly stained (Fig. 11). Less than 3% of the cells were stained when frog blood
300
200
100
0
600 |-
10 20 30
400 -EQ.
200 -A300
200
100
)
c
-
-
/
10
lift
20 30 40
I
10 20 30Fraction no.
Fig. 2
40
400
200
10 20 30
10 20 30
200
100
10 20 30Fraction no.
Fig. 3
40
150
100
50
1
k
i i i
40
40
Figs. 2, 3. Electrophoresis of I4C-labelled reduced and alkylated globin in alkaline 8 Murea. Immunoprecipitation, reduction and alkylation of the immune precipitates,electrophoresis and fractionation were carried out as previously described (Benbassat,
)Fig. 2. A, untreated frog haemolysates; B, immune precipitates of frog haemolysates
treated with antibodies against frog haemoglobin; c, purified major frog haemo-globin (F3). Origin to the left, migration towards the anode.
Fig. 3. A, untreated tadpole haemolysates; B, immune precipitates of tadpole haemo-lysates treated with antibodies against tadpole haemoglobin; c, purified major tadpolehaemoglobin (T^). Origin to the left, migration towards the anode.
smears were treated with antisera against tadpole haemoglobin (Fig. 12) or withcontrol sera.
Imtnunofluorescent staining of peripheral blood cells of animals at the metamorphic climax
The results of the immunofluorescent staining of the peripheral blood cells of animals

Synthesis of red cell protein during metamorphosis. II 149
at the metamorphic climax are given in Table 2 and Figs. 13 and 14. Of these cells,98'1 % exhibited a bright fluorescence after treatment with antisera against tadpolehaemoglobin. The pattern of fluorescence of the cells was identical with that of tadpolered cells stained with antisera against tadpole haemoglobin (Fig. 13). In contrast tothe homogeneity of the smears stained with antisera against tadpole haemoglobin,smears treated with antibodies against frog haemoglobin exhibited a mosaic-likepattern of 16% positively stained cells (Fig. 14).
Table 2. Staining of frog tadpole peripheral blood smears withfluorescent antibodies
Total no. of % cells showingStage Antiserum used* cells counted fluorescence
Premetamorphosis Anti-tadpole Hb 304 987Premetamorphosis Anti-frog Hb 228 17Metamorphic climax Anti-tadpole Hb 527 98-1Metamorphic climax Anti-frog Hb 231 160Adult Anti-tadpole Hb 287 24Adult Anti-frog Hb 240 99-2
• The antisera were prepared by immunizing rabbits with frog or tadpole haemolysatespurified by chromatography on CM-cellulose.
DISCUSSION
The degree of specificity of the antisera employed was determined by doublediffusion on agar, immunoprecipitation of labelled haemolysates and fluorescentstaining of peripheral blood smears. The amounts of labelled tadpole and frog haemo-globin, precipitated by antisera against frog and tadpole haemoglobin, respectively,did not exceed those precipitated by control sera of non-immunized rabbits. Less than3 % of the tadpole or frog blood cells were stained with antisera against frog or tad-pole haemoglobin, respectively, while more than 98 % of the cells in tadpole and frogperipheral blood smears were stained with antibodies against the haemoglobin of thesame developmental stage. Therefore, the finding that 16% of the peripheral bloodcells of metamorphosing animals were stained with antisera against frog haemoglobin,while almost all of them were stained with antisera against tadpole haemolysates,suggests the coexistence of both types of haemoglobin in some of the erythrocytes.
These results conform with the evidence presented by Shukuya (1966) and by Jurd& Maclean (1970) that some of the erythroid cells of metamorphosing tadpoles con-tain both tadpole and frog haemoglobins. The presented observations are consistentalso with the coexistence of human embryonic and foetal haemoglobins (Kleihaueret al. 1967) and of human foetal and adult haemoglobins (Dan & Hagiwara, 1967)within the same erythrocyte. They are at variance, however, with the conclusions ofRosenberg (1970) and Maniatis & Ingram (19716), that during metamorphosis theerythroid cells carry either frog or tadpole haemoglobin but not both.
The reason for this inconsistency is uncertain. One possibility could be, that the

150 J.Benbassat
antisera against tadpole and frog haemoglobins employed by Maniatis & Ingram (1971 a)may have had a higher degree of monospecificity than those used in the presentinvestigation. As shown in Fig. 3, treatment of tadpole haemolysates with antiseraagainst tadpole haemoglobin resulted in the precipitation of several protein subunitsin addition to the main tadpole globin; this additional immunoprecipitate could haveconsisted either of minor tadpole globin fractions or of non-haemoglobin red cellproteins. It is possible, therefore, that the antisera against tadpole haemoglobinemployed in the present study were not monospecific. Thus, the antigen involved inthe immunofluorescent staining of the red cells with antisera against tadpole haemo-globin, could have been a non-haemoglobin protein, present in the red cells of pre-metamorphic and metamorphosing tadpoles, but not in mature frog erythrocytes.
Alternatively, the discrepancy in results could be explained by differences in thesensitivity of the methods employed for immunofluorescent staining. The indirect'sandwich' method for immunofluorescent staining employed in this paper is moresensitive than the direct 'double label' method used by Maniatis & Ingram (1971).Conceivably, small amounts of tadpole haemoglobin in a cell containing principally froghaemoglobin may have escaped detection by direct immunofluorescent staining,particularly when applied to cells already pretreated with fluorescent antibodiesagainst frog haemoglobin.
I am grateful to Dr I. M. London for advice and encouragement and to Dr S. Baum forhelp in the staining with fluorescent antibodies. This investigation was supported by a PublicHealth Service International Research Fellowship (FOTW5-1224) while the author was a post-doctoral fellow in the Department of Medicine, Albert Einstein College of Medicine, New York,and by a grant from the Joint Research Fund of the Hebrew University and Hadassah.
REFERENCES
AGGARWAL, S. J. & RIGGS, A. (1969). The hemoglobins of the bullfrog, Rana catesbeiana.I. Purification, amino acid composition and oxygen equilibria.^, biol. Chem. 244, 2372-2383.
BAGLIONI, C. & SPARKS, C. E. (1963). A study of hemoglobin differentiation in Rana catesbeiana.Devi Biol. 8, 272-285.
BENBASSAT, J. (1970). Erythroid cell development during natural amphibian metamorphosis.Devi Biol. 21, 557-583.
BENBASSAT, J. (1974). The transition from tadpole to frog haemoglobin during natural amphibianmetamorphosis. I. Protein synthesis by peripheral blood cells in vitro. J. Cell Sci. 15,347-357.
BETKE, K. & KLEIHAUER, E. (1958). Fetaler und blebender Blutfarbstoff in Erythrozyten undErythroblasten. Blut 4, 241.
DAN, M. & HAGIWARA, A. (1967). Detection of two types of hemoglobin (HbA and HbF) insingle erythrocytes by fluorescent antibody technique. Expl Cell Res. 46, 596-598.
DEWITT, W. (1968). Microcytes response to thyroxine administration. J. molec. Biol. 32,502-504.
GITLIN, D., SASAKI, T. & VUOPIO, P. (1968). Immunochemical quantitation of proteins in singlecells. Blood 32, 796.
JURD, R. D. & MACLEAN, N. (1970). An immuno-fluorescent study of the haemoglobins inmetamorphosing Xenopus laevis. J. Embryol. exp. Morph. 23, 299—309.
KLEIHAUER, E., TANG, T. E. & BETKE, K. (1967). Die intrazellnare Verteilung von embryo-nalem Hamoglobin in roten Blutzellen menschlicher Embryonen. Ada haemat. 38, 264.
MACLEAN, N. & JURD, R. D. (1971). The haemoglobins of healthy and anaemic Xenopus laevis.J. CeU Sci. 9, 509-528.

Synthesis of red cell protein during metamorphosis. II 151
MANIATIS, G. M. & INGRAM, V. M. (1971a). Erythropoiesis during amphibian metamorphosis.II. Immunochemical study of larval and adult hemoglobins of Rana catesbeiana. J. Cell Biol.49- 380-389.
MANIATIS, G. M. & INGRAM, V. M. (19716). Erythropoiesis during amphibian metamorphosis.III. Immunochemical detection of tadpole and frog hemoglobins in single erythrocytes.J. Cell Biol. 49, 390-404.
Moss, B. & INGRAM, V. M. (1968). Hemoglobin synthesis during amphibian metamorphosis.I. Chemical studies on the hemoglobins from larval and adult stages of R. catesbeiana.J. molec. Biol. 32, 481-492.
ROSENBERG, M. (1970). Electrophoretic analysis of hemoglobin and isozymes in individualvertebrate cells. Proc. natn. Acad. Set. U.S.A. 67, 32-36.
RUGH, R. (1962). Experimental Embryology. Minneapolis: Burgess.SHUKUVA, R. (1966). As cited by Frieden, E. in Metamorphosis (ed. W. Etkin & L. I. Gilbert).
New York: Appleton-Cenrury-Crofte.THIEL, E. (1970). Red blood cell replacement during the transition from tadpole to frog
hemoglobin in Rana catesbeiana. Comp. Biochem. Physiol. 33, 717-720.VANKIN, G. L., BRANDT, E. M. & DEWITT, W. (1970). Fine structure of erythroid cells during
thyroxjn-induced metamorphosis of bullfrog tadpoles. Am. Zool. 10, 321.WELLER, T. H. & COONS, A. H. (1954). Fluorescent antibody studies with agents of Varicella
and Herpes Zoster propagated in vitro. Proc. Soc. exp. Biol. Med. 86, 789-794.
{Received 15 February 1974)

is* J. Benbassat
Figs. 4-7. Double diffusion in 1 % agarose. The reagents consisted of undiluted rabbitantisera (central wells) and haemoglobin solutions in concentrations varying betweenO-O2-O-3 mg/ml (peripheral wells).
Central wells: Figs. 4 and 5, rabbit anti-tadpole hb serum; Figs. 6 and 7, rabbit anti-frog hb serum; the antisera in Figs. 4 and 6 were prepared by immunization of rabbitswith haemolysates purified by chromatography on CM-cellulose; the immunogen usedfor the preparation of the antisera in central wells Figs. 5 and 7 consisted of the purifiedmajor hb fraction.
Peripheral wells: T, unfractionated haemolysate of a prometamorphic tadpole;Tj, purified major tadpole hb fraction; T2, purified minor tadpole hb fraction (seeFig. 8); E, haemolysate of an animal at emergence of the front legs (beginning ofmetamorphic climax); C, haemolysate of an animal at the end of the metamorphicclimax; F, haemolysate of an adult frog; Flt purified minor frog hb fraction; F3, purifiedmajor frog hb fraction (see Fig. 8); M, mouse hb.

Synthesis of red cell protein during metamorphosis. II
8
A B E C FFig. 8. Polyacrylamide gel electrophoresis (discontinuous Tris-glycine/Tris-HCl alka-line system) of haemolysates of R. catesbeiana tadpoles and frogs. Unstained gels.A, early premetamorphic tadpole (total length, 35mm, estimated age 2-3 weeks);B, prometamorphic tadpole (total length, 85 mm, hind legs 2 mm, estimated age, atleast 1 year); E, tadpole at the beginning of the metamorphic climax (total length,98 mm, hind legs 45 mm, 2 days after emergence of front legs); C, animal at the endof metamorphic climax (tail almost completely reabsorbed, 12 days after emergenceof front legs); F, adult frog.

154 J. Benbassat
Figs. 9-12. R. catesbeiana tadpoles and frogs. Peripheral blood smears treated withfluorescent antibodies, x 750.
Fig. 9. Tadpole blood cells stained with antibodies against tadpole haemoglobin.Fig. 10. Tadpole blood cells stained with antibodies against frog haemoglobin. The
fluorescent round body is probably either a white cell or an artifact.

Synthesis of red cell protein during metamorphosis. II
Fig. I I . Frog blood cells stained with antibodies against frog haemoglobin.Fig. 12. Frog blood cells stained with antibodies against tadpole haemoglobin.

156 J. Benbassat
Figs. 13, 14. R. catesbeiana metamorphosing tadpoles. Peripheral blood smearstreated with fluorescent antibodies, x 750.
Fig. 13. Stained with antisera against tadpole haemoglobin.Fig. 14. Stained with antisera against frog haemoglobin.


















