The serotonin system: a potential target for anti...
10
The serotonin system: a potential target for anti-dyskinetic treatments and biomarker discovery. Ottosson, Daniella Published in: Parkinsonism & Related Disorders DOI: 10.1016/S1353-8020(11)70039-6 Published: 2012-01-01 Link to publication Citation for published version (APA): Rylander, D. (2012). The serotonin system: a potential target for anti-dyskinetic treatments and biomarker discovery. Parkinsonism & Related Disorders, 18 Suppl 1, S126-S128. DOI: 10.1016/S1353-8020(11)70039-6 General rights Copyright and moral rights for the publications made accessible in the public portal are retained by the authors and/or other copyright owners and it is a condition of accessing publications that users recognise and abide by the legal requirements associated with these rights. • Users may download and print one copy of any publication from the public portal for the purpose of private study or research. • You may not further distribute the material or use it for any profit-making activity or commercial gain • You may freely distribute the URL identifying the publication in the public portal Take down policy If you believe that this document breaches copyright please contact us providing details, and we will remove access to the work immediately and investigate your claim.
Transcript of The serotonin system: a potential target for anti...

LUND UNIVERSITY
PO Box 117221 00 Lund+46 46-222 00 00
The serotonin system: a potential target for anti-dyskinetic treatments and biomarkerdiscovery.
Ottosson, Daniella
Published in:Parkinsonism & Related Disorders
DOI:10.1016/S1353-8020(11)70039-6
Published: 2012-01-01
Link to publication
Citation for published version (APA):Rylander, D. (2012). The serotonin system: a potential target for anti-dyskinetic treatments and biomarkerdiscovery. Parkinsonism & Related Disorders, 18 Suppl 1, S126-S128. DOI: 10.1016/S1353-8020(11)70039-6
General rightsCopyright and moral rights for the publications made accessible in the public portal are retained by the authorsand/or other copyright owners and it is a condition of accessing publications that users recognise and abide by thelegal requirements associated with these rights.
• Users may download and print one copy of any publication from the public portal for the purpose of private studyor research. • You may not further distribute the material or use it for any profit-making activity or commercial gain • You may freely distribute the URL identifying the publication in the public portalTake down policyIf you believe that this document breaches copyright please contact us providing details, and we will removeaccess to the work immediately and investigate your claim.

Download date: 27. Sep. 2018

Title page SessionType:PlenarySessionSessiontitle:AndreBarbeauSymposium:StriatalPlasticityPresentation title: The serotonin system: a potential target for anti‐dyskinetictreatmentsandbiomarkerdiscoveryAuthor: Daniella RylanderDate:13.12.2011

Abstract L‐DOPA‐induceddyskinesiaisamajorprobleminthetreatmentofParkinson’sdisease.Todaytherearefewanti‐dyskinetictreatmentsavailableforthepatients,andallofthemhavemajorlimitations.
Recent findings have revealed an important role of the serotonin system in L‐DOPA‐induced dyskinesia. In the parkinsonian brain, serotonin axon terminals cancompensateforthedopaminelossbyconvertingL‐DOPAintodopamineandreleasingitas a false neurotransmitter. However, the terminals represent an aberrant source ofdopaminerelease, increasingtherisk fordyskinesia. In linewith this,arelativelyhighdensityofserotoninaxonfibresinstriatumhasbeenreportedindyskineticanimalsandpatients.Furthermore, serotonin can influencedyskinesiabymodulatingglutamateorGABAsignallinginthebasalgangliaviareceptorslocatedonnon‐serotonergicneurons.Through either mechanism, modulation of certain serotonin receptors has shown toreducetheseverityofdyskineticmovements.
The serotonin system represents an interesting target for developing anti‐dyskinetic treatments. Future therapies may take advantage of the synergistic effectproducedbythemodulationofdifferentserotoninreceptorsorpursuearegion‐specificmodulationofcertainreceptors.Moreover,morphologicalorbiochemicalfeaturesoftheserotonin system could be used to develop biomarkers for patient stratification inclinicaltrialsofanti‐dyskineticcompounds.
Introduction L‐DOPA‐induceddyskinesia (LID) is definedas abnormal involuntarymovements thatariseasasideeffectofL‐DOPAtreatment,themosteffectiveandcommondrugusedtotreat Parkinson’s disease (PD). The development of LID can be attributed to apathologicalneuroplasticityofthebrain,attemptingtoadapttoaprogressivedopaminedenervation,(fromthediseaseitself),andaconsecutivedopamineeffluxfrompulsatileL‐DOPA administrations. The brain’s plastic capacity has shown to be variable anddependsonseveral factors, suchas thepatient’sageandgenetics [1].On topof thesefactors,liesanimportantneuromodulatorintheserotonin(5‐HT)system.Serotonergicneurons in the raphe pontis of the brain stemproject axonal fibres tomultiple brainareas including the basal ganglia. The system is particularly vulnerable toneurodegeneration and 5‐HT levels become greatly reduced with age as well as inneurodegenerative diseases, e.g. PD [2]. The variable extent of 5‐HT loss in theparkinsonianbraincouldbeanimportantdeterminantfordyskinesia[3].
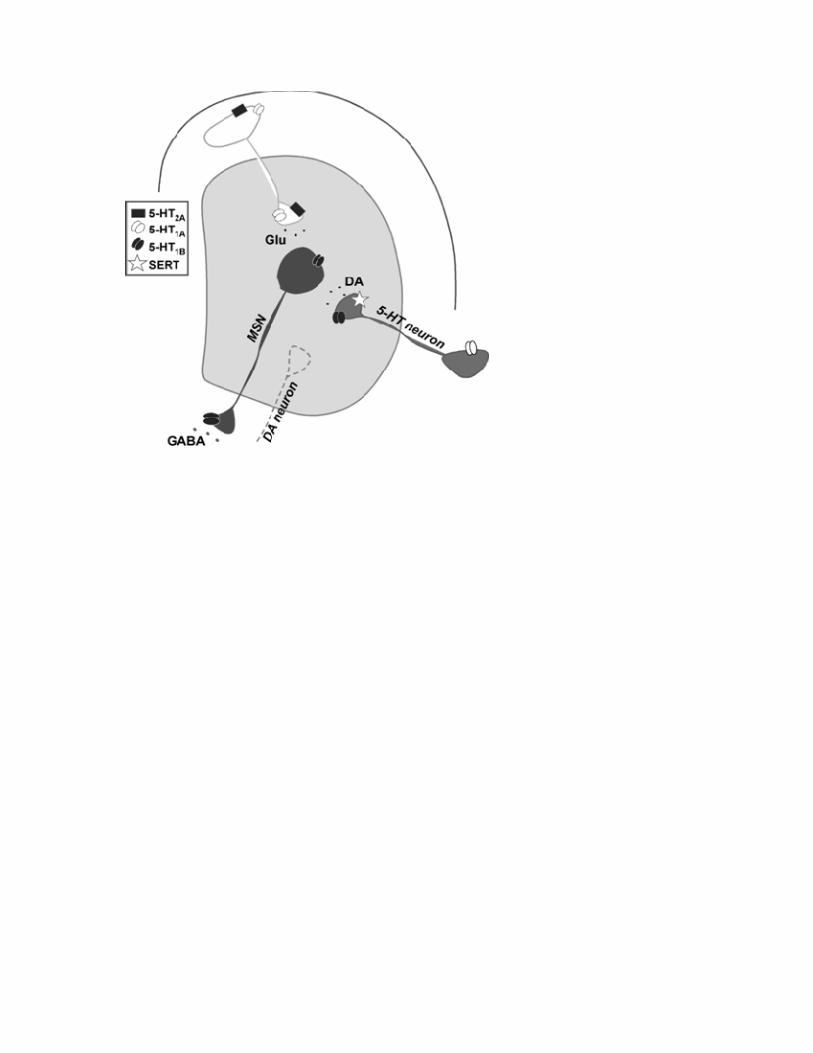
Serotonin’s multiple mechanisms of actions in the basal ganglia The underlyingmechanisms, by which 5‐HT influences dyskinesia, have shown to becomplexandareunderextensiveinvestigation.Thesystemhasseveraldifferenttypesofreceptors distributed in many brain areas, especially the hippocampus, basal gangliaandstriatum.Amongthemoststudiedaretheautoreceptors5‐HT1Aand5‐HT1B.The5‐HT1Areceptorismainly locatedsomatodendriticallyonraphe‐striatalneuronswhereitregulatesneuronal activityand transmitter release.The5‐HT1B receptor,on theotherhand, isexpressedpresynaptically, controlling the terminal release.Thereceptorscanalso be found on non‐serotonergic neurons, such asmedium‐sized spiny neurons (5‐HT1B)andcorticostriatalneurons(5‐HT1A),wherereceptorstimulationreducespallidalGABA‐ or striatal glutamate release, respectively. The corticostriatal neurons also

express the 5‐HT2Areceptor,which regulates striatal glutamate release (See Figure 1)[4].
Recent discoveries from rat and primate models of PD suggest multiplemechanismsofactionbywhichthe5‐HTsysteminfluencesLID.Onegeneralviewisthedopamine‐releasingroleofstriatal5‐HTfibreterminals.Theseterminalsareespeciallygood candidates for converting L‐DOPA into dopamine in the dopamine denervatedbrain,astheyexpressboththeDOPAconvertingenzymeandthevesiculartransporterneeded for processing dopamine [5, 6]. By releasing dopamine into the extracellularspace,the5‐HTterminalsfacilitatethetherapeuticactionofL‐DOPA,butthereleasehasdetrimental consequences. The raphe‐terminals lack auto‐regulatory feedbackmechanisms for dopamine release and cause an aberrant release with excessivelyenhanced dopamine levels in the extracellular space. Such fluctuations between highand lowdopamine levelswill lead to supersensitive responsesof striatalneuronsandtriggerdyskinesia[7,8].
Clinical studies have supported a presynaptic role of the 5‐HT system in LID byshowing excessive swings in striatal dopamine levels in dyskinetic PD patients [9].Similar findings have been reported in dyskinetic rats [8]. Blunting the efflux ofdopamine, by a 5‐HT‐specific lesion, dramatically reduces the severity of LID [6]. Incontrast, enhancing the striatal 5‐HT fibre outgrowth in the dopamine‐denervatedstriatum,bya5‐HTneuronaltransplant,exacerbatesLID[10].
In addition to a presynaptic mechanism, the 5‐HT system also influences LIDthroughitsactiononreceptorslocatedonnon‐serotonergicneurons,i.e.throughapost‐synapticmechanism.Corticalexpressionof5‐HT2Areceptorshasshowntobeenhancedindyskineticmonkeys[4],probablycontributingtothecorticostriataloveractivityandtheexcessivestriatalglutamatereleasethathasbeenassociatedtoLID.Likewise,thereisanincreasedexpressionof5‐HT1Bproteinsonstriatonigralneuronsindyskineticmice[13].Recently,5‐HThasbeenclosely linked toglutamatesignalling in thestriatumbyinducing long‐termdepression,aneffectthatwascoupledtothe5‐HT1Breceptor[11].Thesemechanismsmightbeofparticular interestsincespecificalterations insynapticplasticityhavebeenassociatedtoLID[12].
Growth of raphe‐striatal fibres in dyskinesia: implications for a potential biomarker? Giventhenegativeroleofraphe‐striatalprojections inLIDandtheplasticcharacterofthis system, it seems possible that the variable extent of 5‐HT loss in parkinsonianpatientsmayaffect the individual susceptibility toLID.Recentevidencehaspositivelycorrelatedthedensityof5‐HTaxonal fibresinthestriatumwiththeseverityofLIDinbothratandprimatemodelsaswellasinpost‐mortemtissuefromPDpatients.L‐DOPAtreatmentwasabletoinduceagrowth‐promotingeffectontheraphe‐striatalterminalsalongwithadose‐dependentupregulationofbrain‐derivedneurotrophicfactor(BDNF)[3],promotingregenerativesproutingof5‐HTneurons.Thesproutingeffectwasfurthercoupledtoanincreasedactivity‐dependentdopaminereleaseinratstriatalslices[3].
These findingsdemonstrateapreviouslyunappreciatedmaladaptiveplasticityofthe 5‐HT system in the dyskinetic brain and are in agreement with several clinicalstudies. Indeed, late‐onset PD patients are less susceptible for developing LID whileentailingdegenerativechangesof the5‐HTsystem. Incontrast, youngPDpatientsaremuchmoresusceptiblefordevelopingdyskinesia,andhavealargerplasticitypotentialin the brain along with a higher 5‐HT fibre innervation and BDNF expression [1].Furthermore,morphologicalorbiochemicalfeaturesofthe5‐HTsystemcouldbeused

to devise biomarkers for patient stratification in clinical trials for anti‐dyskineticcompounds.Theextentofthe5‐HTinnervationinthestriatummightalsobeusedasasusceptibility marker for LID. If the degree of striatal 5‐HT innervation could bemeasuredatthetimeofdiagnosis,e.g.bylabellingSERTusingPETimaging,theriskfordevelopingLIDcouldbepredictedandsupport thechoiceoftreatment.Insuchwayitwouldbepossible to initially treatpatientswith a greater risk forLID, i.e. thosewithhigh 5‐HT innervation or young patients, with a less dyskinesiogenic treatment (e.g.dopamineagonistorMAOinhibitors).
Hope for new anti‐dyskinetic drugs by 5‐HT receptor modulation Theimportanceofthe5‐HTsysteminLIDhasreceivedgreatattentioninthesearchfornewanti‐dyskineticdrugs.Stimulatingthe5‐HTautoreceptorshasshowntoreducetheactivity of the raphe‐striatal neurons, blunt the extracellular dopamine release in thestriatumandattenuatetheexpressionofLIDinanimalmodelsofPD[7,8].Stimulationof5‐HT1Aand/or5‐HT1BreceptorsisalsoabletoreduceD1‐agonist‐induceddyskinesiathroughapostsynapticmechanism,(although,thisrequirehigherdosesthanthoseforattenuating LID) [14‐16]. In line with this, recent data points to a role of 5‐HT1AreceptorsexpressedinprimarymotorcortexinthemodulationofLID[17].AneffectivealleviationofLIDcanalsobeachievedbyantagonising5‐HT2Areceptors,mostprobablyattenuatingcorticostriatalglutamateornigrostriataldopaminerelease[4].
Serotoninmodulatorsarenowunder investigation inclinical trials.Mostof thetested drugs provide a reduction in LID but at the expense of an attenuated anti‐parkinsonian efficacy of L‐DOPA. In the rat PDmodel, a high concentration of 5‐HT1Areceptor agonist, (stimulating both pre‐ and post‐synaptic receptors), results insomethingcalled ”serotonin syndrome”with flatbodypostureassociatedwithmotordepression.Inpatients,aworseninginparkinsonismhasbeenspeculatedtodependonnon‐specificbindingbythedrugstoextrastriataltargetsortootherreceptortypese.g.theD2receptor.However, theselectivityseemsnottobethesolelyproblemsincethelack of anti‐parkinsonian effect persists also with highly specific substances. Instead,stimulationofanatomicallydistinctsubpopulationsofreceptorshasbeenpostulatedasabetterwayoftreatingdyskinesiawithoutcompromisingtheanti‐parkinsonianefficacy[4].Toexclusivelytargete.g.5‐HT2AreceptorsoncorticostriatalneuronswouldreduceLIDbyattenuating striatal glutamate release,whereas stimulating the same receptorson residual nigrostriatal terminals would impair the anti‐parkinsonian effect byreducing dopamine release [4] (See Figure1). Taken together, less advancedpatientsthat still possess some residual striatal dopamine terminals, might provide bettertargets for anti‐dyskinetic treatmentwith 5‐HT agonists since their dopamine releasewouldnotbeattenuatedasmuchbythe5‐HTagonists[6].
Another solution for avoiding themotor depressant effectwould be to use thesynergisticeffectbetween5‐HT1A/1Breceptoragonists,usingsub‐thresholddoses.ThisregimencanpreventtheappearanceofLIDaswellastheinductionofaLID‐associatedtranscription factor in striatal neurons [7, 16]. Aproof‐of‐concept clinical trial is nowinvestigatingtheefficacyof5‐HT1A/1BreceptoragonistsinadvancedPDpatients.
A therapeutic or a dyskinetic effect in transplantation? Intrastriatal transplantations of ventral mesencephalic tissue are still underdevelopment as a potential future treatment for PD. Serotonin neurons are oftenincludedinthegraftstoavariableextent,andaresuspectedtoworsenLIDbyprovidingan additional source of unregulated dopamine release. Serotonin neurons could also

influence the occurrence of graft‐induced dyskinesia, (i.e. dyskinesia without L‐DOPAadministration).
Studiesusingspecialdissectionprotocol,yieldinggraftsrichineitherdopamineor5‐HTneurons,haveshownthattransplantsenrichedindopamineneuronsinducegoodfunctionalrecovery,whilethoseenrichedin5‐HTneuronscauseadramaticworseningofLIDandnoimprovementofphysiologicalmotortasks[10].Inamixedgrafthowever,thebehaviouraloutcomeisnotaffectedbytheamountof5‐HTneuronsas longasthegraft also contains only a small amount of dopamine neurons. Thus, the relativedensities,andnottheabsolutenumberof5‐HTneuronsinthegraft,seemtodeterminewhether 5‐HT grafted neurons have detrimental effects or not [18]. Taken into theclinicalsituation,thesefindingssuggestlessseverePDpatients,whostillpossesssomeresidualdopaminefibresinstriatum,astheoptimalpatientsforneuraltransplantation.
Theroleof5‐HTneuronsingraft‐induceddyskinesiaismorevaguethantheirroleinLID.Also for thesecomplications, thedopaminecontentseems tohave thegreatestimpact even if the contribution of 5‐HT system cannot be ruled out. Graft‐induceddyskinesia is worsened by SERT inhibition and attenuated by 5‐HT1A receptorstimulation[19],butapossiblecontributionofthe5‐HTsystemseemstodependontheendogenoussystemas theremovalof thegrafted5‐HTneuronshasnoeffect [19].Ontheotherhand, a recent studyof two transplantedPDpatients showedapronouncedroleofgrafted5‐HTneuronsingraft‐induceddyskinesia.Inthesepatients,graft‐induceddyskinesiawasassociatedwithanabnormally largestriatal5‐HTinnervationandwaseffectivelyattenuatedbytheadministrationofa5‐HT1Aagonist[20].
Taken together, the role of the 5‐HT system in the occurrence of graft‐induceddyskinesia remains an intriguing question that needs further evaluation in order todevelopthispotentialfuturetherapyforPD.
Conclusion Vast evidences are today clearlydemonstrating the importanceof the5‐HT system inthepathophysiologyof LID, acting eitherasadopamine‐releasing compartmentoronreceptors located on non‐serotonergic neurons. By influencing striatal dopamine andglutamate signalling, the 5‐HT system does provide major influence on the aberrantstriatal signalling that underlies LID. A growth‐promoting effect on the raphe‐striatalfibres in L‐DOPA‐treated dyskinetic subjects, along with its dopamine‐releasingproperties,furtherputsthebrainatgreaterriskfordevelopingdyskinesia.Inlinewiththis, a graft‐induced 5‐HT hyperinnervation from intrastriatal grafts would expect toworsen thedyskinesia.Yet, the respective roleof endogenousversus exogenous5‐HTcontributiontothisrequiresfurtherevaluation.
The5‐HTsystemcanbeseenasaparticularinterestingtargetforanti‐dyskinetictreatment andnewevidenceofplastic responses in this systemcould furtherprovidetoolsforhowtoindividuallytreatdyskineticcomplicationsinPD.
References 1. LinazasoroG.NewideasontheoriginofL‐dopa‐induceddyskinesias:age,genesandneuralplasticity.TrendsPharmacolSci.2005Aug;26(8):391‐7.2. NicholsonSL,BrotchieJM.5‐hydroxytryptamine(5‐HT,serotonin)andParkinson'sdisease ‐ opportunities for novel therapeutics to reduce the problems of levodopatherapy.EurJNeurol.2002Nov;9Suppl3:1‐6.3. RylanderD,ParentM,O'SullivanSS,DoveroS,LeesAJ,BezardE,etal.Maladaptiveplasticityofserotoninaxonterminalsinlevodopa‐induceddyskinesia.AnnNeurol.2010Nov;68(5):619‐28.

4. Huot P, Fox SH, Newman‐Tancredi A, Brotchie JM. Anatomically‐selective 5‐HT1Aand 5‐HT2A therapies for Parkinson's disease ‐ an approach to reducing dyskinesiawithoutexacerbatingparkinsonism?JPharmacolExpTher.2011Jul22.5. Arai R, Karasawa N, Geffard M, Nagatsu I. L‐DOPA is converted to dopamine inserotonergic fibers of the striatum of the rat: a double‐labeling immunofluorescencestudy.NeurosciLett.1995Aug11;195(3):195‐8.6. Carta M, Bezard E. Contribution of pre‐synaptic mechanisms to l‐DOPA‐induceddyskinesia.Neuroscience.2011Aug5.7. CartaM,CarlssonT,KirikD,BjorklundA.Dopaminereleasedfrom5‐HTterminalsisthe cause of L‐DOPA‐induced dyskinesia in parkinsonian rats. Brain. 2007 Jul;130(Pt7):1819‐33.8. LindgrenHS,AnderssonDR,LagerkvistS,NissbrandtH,CenciMA.L‐DOPA‐induceddopamineefflux inthestriatumandthesubstantianigra inaratmodelofParkinson'sdisease: temporal and quantitative relationship to the expression of dyskinesia. JNeurochem.2010Mar;112(6):1465‐76.9. de la Fuente‐Fernandez R, Sossi V, Huang Z, Furtado S, Lu JQ, Calne DB, et al.Levodopa‐induced changes in synaptic dopamine levels increase with progression ofParkinson'sdisease:implicationsfordyskinesias.Brain.2004Dec;127(Pt12):2747‐54.10. CarlssonT,CartaM,WinklerC,BjorklundA,KirikD.Serotoninneurontransplantsexacerbate L‐DOPA‐induced dyskinesias in a rat model of Parkinson's disease. JNeurosci.2007Jul25;27(30):8011‐22.11. Mathur BN, Capik NA, Alvarez VA, Lovinger DM. Serotonin induces long‐termdepressionatcorticostriatalsynapses.JNeurosci.2011May18;31(20):7402‐11.12. PicconiB,BagettaV,GhiglieriV,PailleV,DiFilippoM,PendolinoV,etal.Inhibitionof phosphodiesterases rescues striatal long‐term depression and reduces levodopa‐induceddyskinesia.Brain.2011Feb;134(Pt2):375‐87.13. ZhangX,AndrenPE,GreengardP,SvenningssonP.Evidenceforaroleofthe5‐HT1Breceptor and its adaptor protein, p11, in L‐DOPA treatment of an animal model ofParkinsonism.ProcNatlAcadSciUSA.2008Feb12;105(6):2163‐8.14. DupreKB,OstockCY,EskowJaunarajsKL,ButtonT,SavageLM,WolfW,etal.Localmodulation of striatal glutamate efflux by serotonin 1A receptor stimulation indyskinetic,hemiparkinsonianrats.ExpNeurol.2011Jun;229(2):288‐99.15. JaunarajsKL,DupreKB,SteinigerA,KliouevaA,MooreA,KellyC,etal.Serotonin1Breceptor stimulation reduces D1 receptor agonist‐induced dyskinesia. Neuroreport.2009Sep23;20(14):1265‐9.16. Munoz A, Carlsson T, Tronci E, Kirik D, Bjorklund A, Carta M. Serotonin neuron‐dependentand ‐independentreductionofdyskinesiaby5‐HT1Aand5‐HT1BreceptoragonistsintheratParkinsonmodel.ExpNeurol.2009Sep;219(1):298‐307.17. OstockCY,DupreKB,EskowJaunarajsKL,WaltersH,George J,KrolewskiD,etal.Roleoftheprimarymotorcortexinl‐DOPA‐induceddyskinesiaanditsmodulationby5‐HT1Areceptorstimulation.Neuropharmacology.2011Sep;61(4):753‐60.18. Garcia J, Carlsson T, Dobrossy M, Nikkhah G, Winkler C. Impact of dopamine toserotonin cell ratio in transplants on behavioral recovery and L‐DOPA‐induceddyskinesia.NeurobiolDis.2011Sep;43(3):576‐87.19. LaneEL,BjorklundA,DunnettSB,WinklerC.NeuralgraftinginParkinson'sdiseaseunraveling the mechanisms underlying graft‐induced dyskinesia. Prog Brain Res.2010;184:295‐309.

20. Politis M, Wu K, Loane C, Quinn NP, Brooks DJ, Rehncrona S, et al. SerotonergicneuronsmediatedyskinesiasideeffectsinParkinson'spatientswithneuraltransplants.SciTranslMed.2010Jun30;2(38):38ra46.
Figure legend Figure 1: A simplified view illustrating the location of potential anti-dyskinetic targets (i.e. the 5-HT receptors) and potential biomarker (i.e. SERT). An anti-dyskinetic effect can be achieved by i) modulating serotonin autoreceptors on raphe-terminals inhibiting dopamine release, ii) on corticostriatal neurons inhibiting glutamate release or iii) on striatonigral medium-sized spiny neurons inhibiting GABA release. Abbreviations: Glu = glutamate, DA = dopamine, 5-HT = serotonin, SERT = serotonin uptake transporter, MSN = medium-sized spiny neurons.