
The Roles of Valine 208 and Histidine 211 in Ligand Binding and Receptor Function of the Ovine...
6
BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS 239, 418–423 (1997) ARTICLE NO. RC977482 The Roles of Valine 208 and Histidine 211 in Ligand Binding and Receptor Function of the Ovine Mel 1ab Melatonin Receptor Shaun Conway,* ,1 Sarah J. Canning,* Perry Barrett,* Beatrice Guardiola-Lemaitre,² Phillipe Delagrange,² and Peter J. Morgan* *Molecular Neuroendocrinology Unit, Rowett Research Institute, Greenburn Road, Bucksburn, Aberdeen, Scotland, United Kingdom AB21 9SB; and ²Institut de recherches Internationales Servier, 6 place de Ple ´iades, F-92415 Courbevoie Cedex, France Received September 8, 1997 in seasonally breeding mammals (2). These effects are Site-directed mutagenesis was used to study two res- achieved via the binding of melatonin to high-affinity idues, valine 208 and histidine 211, in transmembrane cell surface melatonin receptors (3,4,5). Despite our in- domain 5 of the ovine Mel 1ab melatonin receptor. A se- sight into the possible functions of melatonin receptors ries of 4 mutants were constructed (V208A, V208L, little is known about the molecular structure of the H211F, H211L), and each engineered to contain a receptor or receptor-ligand interactions, information FLAG-epitope. Immunocytochemistry demonstrated that could lead to the design of specific agonist and that all the mutants were expressed in COS-7 cells at antagonist modulators of the melatonin signal. The levels comparable to the FLAG-epitope tagged wild- clinical application of such drugs could prove beneficial type Mel 1ab receptor (Ç120 fmol/mg protein). Ligand as administered melatonin has already been shown to binding revealed however that all mutants had re- modulate the sleep-wake cycle in the blind (6,7) and duced affinities for 2-[ 125 I]-iodomelatonin (Kd wild- provides an effective treatment for jet-lag (8). type 139 pM, Kd mutants 320 to 989 pM). Competition studies, with a series of melatonin analogues, identi- The recent cloning of a series of high-affinity melato- fied a probable interaction between histidine 211 and nin receptors has identified them as members of the the 5-methoxy group of melatonin. The wild-type re- GPCR ‘superfamily’ (9,10,11). It has been proposed that ceptor and both valine 208 mutants displayed a dose- melatonin receptors will have a heptahelical architec- dependent melatonin mediated inhibition of cyclic ture containing an intrahelical ligand binding site, in AMP levels in HEK293 cells, with IC 50 values in the common with other GPCRs (9,12). One feature of many same rank-order as their melatonin binding affinities. GPCRs is that the general structure of the ligand bind- Both H211F and H211L, however, did not display any ing site appears to remain essentially conserved. This melatonin mediated effects and may suggest that histi- is illustrated by the alignment of residues that have dine 211 is critical for melatonin mediated receptor been shown to provide ligand binding interactions in activation. q 1997 Academic Press diverse GPCRs. For example two TM5 serine residues in the b2-adrenergic have been proven to interact with Melatonin synchronizes circadian rhythms in mam- the two hydroxyl groups on the aromatic ring of adrena- mals (1), and regulates aspects of reproductive state line (13). Residues which protein sequence align with these serine residues, placing them in the same pre- sumed position within each receptor, have also been 1 Corresponding author. Fax: /44 (0)1224 716653. E-mail: sco@ rri.sari.ac.uk. shown to interact with the substituted ring of 5-HT in Abbreviations: 125 I-Mel, 2-[ 125 I]-iodomelatonin; GPCR(s), G pro- certain 5-HT receptors (14,15). The two residues which tein-coupled receptor(s); TM, transmembrane segment; 5-HT, 5-hy- are in analogous TM5 positions in melatonin receptors droxytryptamine; N-AS, N-acetylserotonin; MSH, melanocyte stimu- are a conserved valine and a conserved histidine (valine lating hormone, DEAE-dextran, diethylaminoethyl-dextran; PBS, phosphate buffered saline (pH 7.4); EGTA, ethylene glycol-bis(b-ami- 208 and histidine 211, ovine Mel 1a receptor). At present noethyl ether) N,N,N*,N*-tetraacetic acid; DMEM, Dulbecco’s modi- the structure of the melatonin binding site is unknown, fied Eagle’s medium; EMEM, Eagle’s minimum essential medium; but if it is essentially conserved with those of adrenergic PEG 8000, polyethylene glycol (average molecular weight 8,000); and 5-HT receptors then these two residues may also IBMX, 3-isobutyl-1-methylxanthine; a-NDP-MSH, [Nle 4 ,D-Phe 7 ]-a- melanocyte stimulating hormone; TCA, trichloroacetic acid. be involved in ligand binding, potentially interacting 0006-291X/97 $25.00 Copyright q 1997 by Academic Press All rights of reproduction in any form reserved. 418
-
Upload
shaun-conway -
Category
Documents
-
view
213 -
download
1
Transcript of The Roles of Valine 208 and Histidine 211 in Ligand Binding and Receptor Function of the Ovine...

BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS 239, 418–423 (1997)ARTICLE NO. RC977482
The Roles of Valine 208 and Histidine 211in Ligand Binding and Receptor Functionof the Ovine Mel1ab Melatonin Receptor
Shaun Conway,*,1 Sarah J. Canning,* Perry Barrett,* Beatrice Guardiola-Lemaitre,†Phillipe Delagrange,† and Peter J. Morgan**Molecular Neuroendocrinology Unit, Rowett Research Institute, Greenburn Road, Bucksburn, Aberdeen,Scotland, United Kingdom AB21 9SB; and †Institut de recherches Internationales Servier,6 place de Pleiades, F-92415 Courbevoie Cedex, France
Received September 8, 1997
in seasonally breeding mammals (2). These effects areSite-directed mutagenesis was used to study two res- achieved via the binding of melatonin to high-affinity
idues, valine 208 and histidine 211, in transmembrane cell surface melatonin receptors (3,4,5). Despite our in-domain 5 of the ovine Mel1ab melatonin receptor. A se- sight into the possible functions of melatonin receptorsries of 4 mutants were constructed (V208A, V208L,
little is known about the molecular structure of theH211F, H211L), and each engineered to contain areceptor or receptor-ligand interactions, informationFLAG-epitope. Immunocytochemistry demonstratedthat could lead to the design of specific agonist andthat all the mutants were expressed in COS-7 cells atantagonist modulators of the melatonin signal. Thelevels comparable to the FLAG-epitope tagged wild-clinical application of such drugs could prove beneficialtype Mel1ab receptor (Ç120 fmol/mg protein). Ligandas administered melatonin has already been shown tobinding revealed however that all mutants had re-modulate the sleep-wake cycle in the blind (6,7) andduced affinities for 2-[125I]-iodomelatonin (Kd wild-provides an effective treatment for jet-lag (8).type 139 pM, Kd mutants 320 to 989 pM). Competition
studies, with a series of melatonin analogues, identi- The recent cloning of a series of high-affinity melato-fied a probable interaction between histidine 211 and nin receptors has identified them as members of thethe 5-methoxy group of melatonin. The wild-type re- GPCR ‘superfamily’ (9,10,11). It has been proposed thatceptor and both valine 208 mutants displayed a dose- melatonin receptors will have a heptahelical architec-dependent melatonin mediated inhibition of cyclic ture containing an intrahelical ligand binding site, inAMP levels in HEK293 cells, with IC50 values in the common with other GPCRs (9,12). One feature of manysame rank-order as their melatonin binding affinities. GPCRs is that the general structure of the ligand bind-Both H211F and H211L, however, did not display any ing site appears to remain essentially conserved. Thismelatonin mediated effects and may suggest that histi- is illustrated by the alignment of residues that havedine 211 is critical for melatonin mediated receptor
been shown to provide ligand binding interactions inactivation. q 1997 Academic Pressdiverse GPCRs. For example two TM5 serine residuesin the b2-adrenergic have been proven to interact with
Melatonin synchronizes circadian rhythms in mam- the two hydroxyl groups on the aromatic ring of adrena-mals (1), and regulates aspects of reproductive state line (13). Residues which protein sequence align with
these serine residues, placing them in the same pre-sumed position within each receptor, have also been1 Corresponding author. Fax: /44 (0)1224 716653. E-mail: sco@
rri.sari.ac.uk. shown to interact with the substituted ring of 5-HT inAbbreviations: 125I-Mel, 2-[125I]-iodomelatonin; GPCR(s), G pro- certain 5-HT receptors (14,15). The two residues which
tein-coupled receptor(s); TM, transmembrane segment; 5-HT, 5-hy-are in analogous TM5 positions in melatonin receptorsdroxytryptamine; N-AS, N-acetylserotonin; MSH, melanocyte stimu-are a conserved valine and a conserved histidine (valinelating hormone, DEAE-dextran, diethylaminoethyl-dextran; PBS,
phosphate buffered saline (pH 7.4); EGTA, ethylene glycol-bis(b-ami- 208 and histidine 211, ovine Mel1a receptor). At presentnoethyl ether) N,N,N*,N*-tetraacetic acid; DMEM, Dulbecco’s modi- the structure of the melatonin binding site is unknown,fied Eagle’s medium; EMEM, Eagle’s minimum essential medium; but if it is essentially conserved with those of adrenergicPEG 8000, polyethylene glycol (average molecular weight 8,000);
and 5-HT receptors then these two residues may alsoIBMX, 3-isobutyl-1-methylxanthine; a-NDP-MSH, [Nle4,D-Phe7]-a-melanocyte stimulating hormone; TCA, trichloroacetic acid. be involved in ligand binding, potentially interacting
0006-291X/97 $25.00Copyright q 1997 by Academic PressAll rights of reproduction in any form reserved.
418
AID BBRC 7482 / 693b$$$221 09-22-97 21:16:03 bbrcgs AP: BBRC
Vol. 239, No. 2, 1997 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
studies the ovine Mel1ab receptor cDNA was ligated into the vectorpBK-CMV (Stratagene) and the ovine MSH receptor cDNA into vectorpcDNA I (Invitrogen). COS-7 cells and HEK293 cells were providedby ECACC. 125I-Mel (2200 Ci/mmol) was purchased from NEN DuPont.The drugs S20098 and N-NEA (N-[2-(1-naphthyl) ethyl] acetamide)were synthesised by Servier (see Fig. 2 for structures). Other generalreagents were purchased from Sigma or Life Technologies Inc.
Site-directed mutagenesis. Mutations were introduced into theovine Mel1ab melatonin receptor in pBK-CMV using the Chameleonsite-directed mutagenesis kit (Stratagene). Successful construction ofeach mutant was confirmed by DNA sequencing using the Prism dye-deoxy terminator kit (Applied Biosystems Inc.). In this study 4 mutantswere generated, mutants V208A, V208L, H211F, and H211L.
FLAG epitope tagging. A PCR based protocol was used to intro-duce the FLAG-epitope (DYKDDDDK), encoded by the nucleotidesequence 5*»GACTACAAGGACGACGACGATAAG…3 *, between co-don 1 and codon 2 of the ovine Mel1ab receptor cDNA. FLAG-epitopetagged mutant receptors were generated by subcloning fragmentscontaining the desired mutations into the FLAG-epitope taggedwild-type receptor construct. All constructs were confirmed by DNAsequencing.
COS-7 cell tissue culture. COS-7 cells were grown as monolayersin DMEM supplemented with 10% fetal calf serum and 1% antibiotic/antimycotic solution, in 5% CO2 at 377C. Confluent plates were re-seeded at 15,000 cells/cm2 and used 24 hours later for transfection.Cells were transfected with receptor constructs using the DEAE-Dextran method of Cullen (18) and grown for 72 hours. Growth mediawas aspirated and cells washed twice with PBS before being har-vested. Cells were pelleted in 1.5 ml microcentrifuge tubes (13,000g, 1 min, 47C), and stored frozen at 0807C.
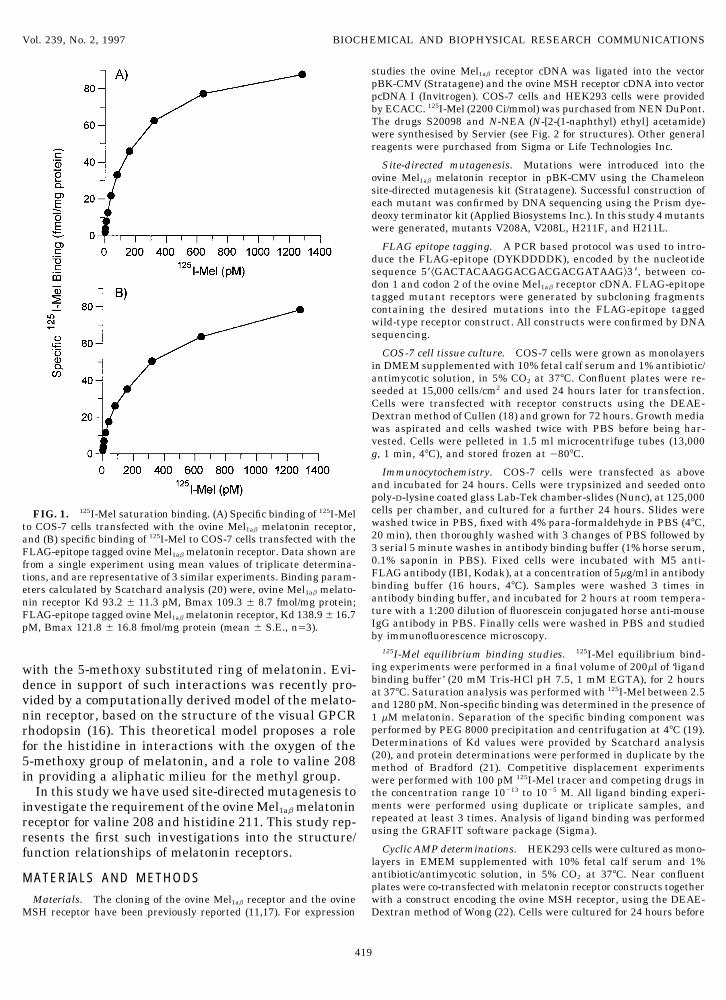
Immunocytochemistry. COS-7 cells were transfected as aboveand incubated for 24 hours. Cells were trypsinized and seeded ontopoly-D-lysine coated glass Lab-Tek chamber-slides (Nunc), at 125,000cells per chamber, and cultured for a further 24 hours. Slides wereFIG. 1. 125I-Mel saturation binding. (A) Specific binding of 125I-Melwashed twice in PBS, fixed with 4% para-formaldehyde in PBS (47C,to COS-7 cells transfected with the ovine Mel1ab melatonin receptor,20 min), then thoroughly washed with 3 changes of PBS followed byand (B) specific binding of 125I-Mel to COS-7 cells transfected with the3 serial 5 minute washes in antibody binding buffer (1% horse serum,FLAG-epitope tagged ovine Mel1ab melatonin receptor. Data shown are0.1% saponin in PBS). Fixed cells were incubated with M5 anti-from a single experiment using mean values of triplicate determina-FLAG antibody (IBI, Kodak), at a concentration of 5mg/ml in antibodytions, and are representative of 3 similar experiments. Binding param-binding buffer (16 hours, 47C). Samples were washed 3 times ineters calculated by Scatchard analysis (20) were, ovine Mel1ab melato-antibody binding buffer, and incubated for 2 hours at room tempera-nin receptor Kd 93.2 { 11.3 pM, Bmax 109.3 { 8.7 fmol/mg protein;ture with a 1:200 dilution of fluorescein conjugated horse anti-mouseFLAG-epitope tagged ovine Mel1ab melatonin receptor, Kd 138.9{ 16.7IgG antibody in PBS. Finally cells were washed in PBS and studiedpM, Bmax 121.8 { 16.8 fmol/mg protein (mean { S.E., nÅ3).by immunofluorescence microscopy.
125I-Mel equilibrium binding studies. 125I-Mel equilibrium bind-ing experiments were performed in a final volume of 200ml of ‘ligandwith the 5-methoxy substituted ring of melatonin. Evi-binding buffer’ (20 mM Tris-HCl pH 7.5, 1 mM EGTA), for 2 hoursdence in support of such interactions was recently pro- at 377C. Saturation analysis was performed with 125I-Mel between 2.5
vided by a computationally derived model of the melato- and 1280 pM. Non-specific binding was determined in the presence ofnin receptor, based on the structure of the visual GPCR 1 mM melatonin. Separation of the specific binding component was
performed by PEG 8000 precipitation and centrifugation at 47C (19).rhodopsin (16). This theoretical model proposes a roleDeterminations of Kd values were provided by Scatchard analysisfor the histidine in interactions with the oxygen of the(20), and protein determinations were performed in duplicate by the5-methoxy group of melatonin, and a role to valine 208 method of Bradford (21). Competitive displacement experiments
in providing a aliphatic milieu for the methyl group. were performed with 100 pM 125I-Mel tracer and competing drugs inIn this study we have used site-directed mutagenesis to the concentration range 10013 to 1005 M. All ligand binding experi-
ments were performed using duplicate or triplicate samples, andinvestigate the requirement of the ovine Mel1ab melatoninrepeated at least 3 times. Analysis of ligand binding was performedreceptor for valine 208 and histidine 211. This study rep-using the GRAFIT software package (Sigma).resents the first such investigations into the structure/
Cyclic AMP determinations. HEK293 cells were cultured as mono-function relationships of melatonin receptors.layers in EMEM supplemented with 10% fetal calf serum and 1%antibiotic/antimycotic solution, in 5% CO2 at 377C. Near confluentMATERIALS AND METHODSplates were co-transfected with melatonin receptor constructs togetherwith a construct encoding the ovine MSH receptor, using the DEAE-Materials. The cloning of the ovine Mel1ab receptor and the ovine
MSH receptor have been previously reported (11,17). For expression Dextran method of Wong (22). Cells were cultured for 24 hours before
419
AID BBRC 7482 / 693b$$$221 09-22-97 21:16:03 bbrcgs AP: BBRC
Vol. 239, No. 2, 1997 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
TABLE 1
Ligand Binding Affinities of the FLAG-Epitope Tagged Wild-Type and Site-Directed Mutant,Ovine Mel1ab Melatonin Receptors
Kd [nM] Ki [nM]
Receptor 125I-Mel Melatonin S20098 N-NEA N-acetylserotonin
Wild-type-FLAG 0.14 { 0.02 0.63 { 0.05 0.98 { 0.06 86.1 { 3.6 754 { 32H211F 0.83 { 0.10 (05.9) 3.54 { 0.63 (05.6) 5.00 { 1.13 (05.1) 25.0 { 4.0 (/3.4) 832 { 134 (01.1)H211L 0.99 { 0.16 (07.1) 5.01 { 0.63 (07.9) 4.58 { 0.69 (04.7) 5.72 { 0.98 (/15.1) 501 { 49 (/1.5)V208A 0.32 { 0.03 (02.3) 1.08 { 0.18 (01.7) 1.53 { 0.07 (01.6) 146 { 13 (01.7) 1560 { 190 (02.2)V208L 0.71 { 0.09 (05.1) 4.01 { 0.67 (06.4) 11.9 { 2.1 (012.1) 795 { 140 (09.2) Ç10,000 (Ç013.8)
Note. Experiments were performed as stated under Materials and Methods, Kd values were determined by Scatchard analysis (20), andKi values determined using the Cheng-Prusoff correction (25). Mutant receptors are referred to by the abbreviations stated in Materialsand Methods. All values are the mean { S.E. from 3 or 4 identical experiments. The fold difference in affinity for the mutant receptorscompared to the affinity for the FLAG-epitope tagged wild-type receptor is shown in parentheses for each drug.
being re-seeded into poly-D-lysine coated 24 well tissue culture plates in the range 10012 to 1007 M, added for 30 min at 377C. Reactionswere stopped by removal of the assay medium and addition of 0.4 mlat 21105 cells per well. Following incubation for a further 24 hours
the cells were assayed. Medium was aspirated and the cells washed 5% TCA solution. Cyclic AMP determinations were performed on theTCA extract by radio-immuno assay as previously described (23). Dataonce with 0.5 ml of supplement free EMEM. The wash medium was
removed and 0.5 ml EMEM (containing 1 mM IBMX) and appropriate from individual experiments were averaged from duplicate determina-tions of 3 separate wells treated under identical conditions.concentrations of drugs, routinely 1 mM a-NDP-MSH and melatonin
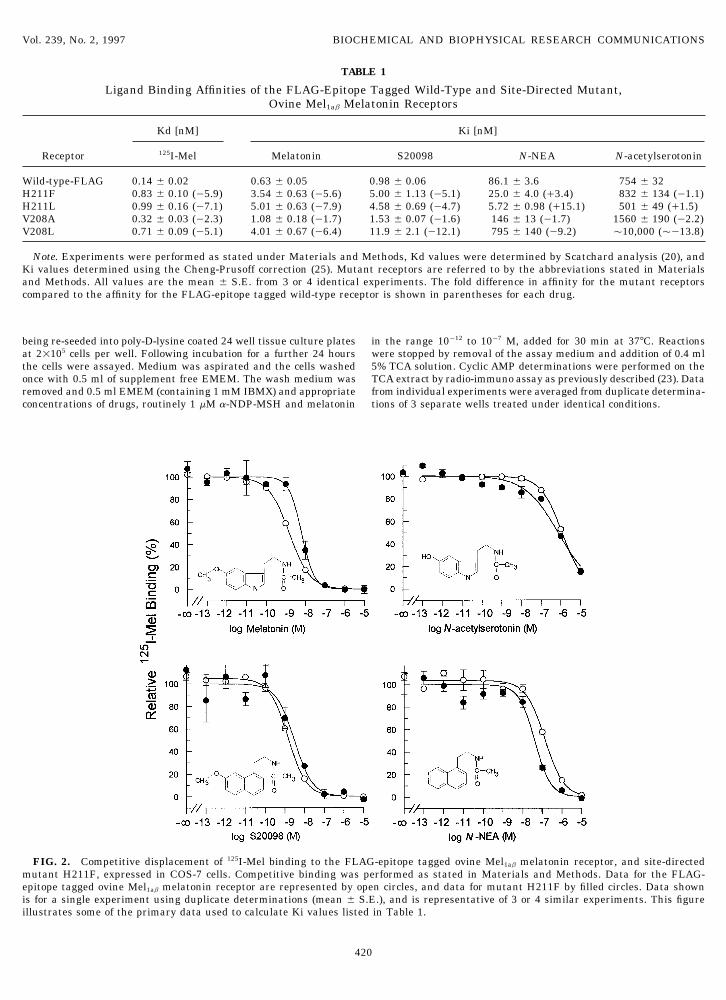
FIG. 2. Competitive displacement of 125I-Mel binding to the FLAG-epitope tagged ovine Mel1ab melatonin receptor, and site-directedmutant H211F, expressed in COS-7 cells. Competitive binding was performed as stated in Materials and Methods. Data for the FLAG-epitope tagged ovine Mel1ab melatonin receptor are represented by open circles, and data for mutant H211F by filled circles. Data shownis for a single experiment using duplicate determinations (mean { S.E.), and is representative of 3 or 4 similar experiments. This figureillustrates some of the primary data used to calculate Ki values listed in Table 1.
420
AID BBRC 7482 / 693b$$$221 09-22-97 21:16:03 bbrcgs AP: BBRC
Vol. 239, No. 2, 1997 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
RESULTS AND DISCUSSION
The melatonin receptor used in this study was theovine Mel1ab receptor (11), which had been modifiedby the inclusion of a FLAG-epitope within the amino-terminal domain prior to TM1 (data not shown). Whentested against an unmodified ovine Mel1ab receptor theaddition of the FLAG-epitope did not alter the expres-sion level or 125I-Mel binding affinity of transfectedCOS-7 cells (Fig. 1). The FLAG-epitope tagged receptorwas further characterized by competitive displacementof 125I-Mel by melatonin and several related drugs, pro-ducing affinity values consistent with those previouslyreported for other Mel1a and Mel1b melatonin receptors(Table 1 cf. refs. 10,24). Also the FLAG-tagged receptorwas fully functional, displaying a melatonin mediatedinhibition of stimulated cAMP levels in transfectedHEK293 cells (Fig. 3). Therefore the FLAG-epitopetagged ovine Mel1ab receptor would appear a suitablemodel for investigations into melatonin receptor struc-ture and function.
A series of 4 site-directed FLAG-epitope tagged mu-tants were constructed, V208A, V208L, H211F, and
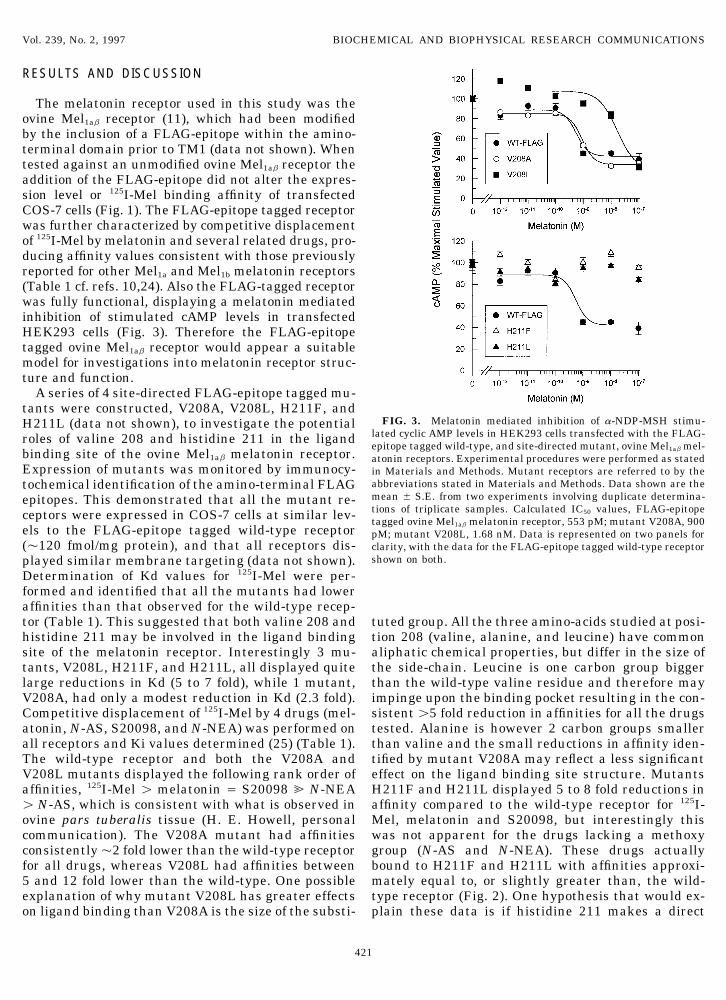
FIG. 3. Melatonin mediated inhibition of a-NDP-MSH stimu-H211L (data not shown), to investigate the potentiallated cyclic AMP levels in HEK293 cells transfected with the FLAG-roles of valine 208 and histidine 211 in the ligandepitope tagged wild-type, and site-directed mutant, ovine Mel1ab mel-
binding site of the ovine Mel1ab melatonin receptor. atonin receptors. Experimental procedures were performed as statedExpression of mutants was monitored by immunocy- in Materials and Methods. Mutant receptors are referred to by the
abbreviations stated in Materials and Methods. Data shown are thetochemical identification of the amino-terminal FLAGmean { S.E. from two experiments involving duplicate determina-epitopes. This demonstrated that all the mutant re-tions of triplicate samples. Calculated IC50 values, FLAG-epitopeceptors were expressed in COS-7 cells at similar lev- tagged ovine Mel1ab melatonin receptor, 553 pM; mutant V208A, 900
els to the FLAG-epitope tagged wild-type receptor pM; mutant V208L, 1.68 nM. Data is represented on two panels for(Ç120 fmol/mg protein), and that all receptors dis- clarity, with the data for the FLAG-epitope tagged wild-type receptor
shown on both.played similar membrane targeting (data not shown).Determination of Kd values for 125I-Mel were per-formed and identified that all the mutants had loweraffinities than that observed for the wild-type recep-tor (Table 1). This suggested that both valine 208 and tuted group. All the three amino-acids studied at posi-
tion 208 (valine, alanine, and leucine) have commonhistidine 211 may be involved in the ligand bindingsite of the melatonin receptor. Interestingly 3 mu- aliphatic chemical properties, but differ in the size of
the side-chain. Leucine is one carbon group biggertants, V208L, H211F, and H211L, all displayed quitelarge reductions in Kd (5 to 7 fold), while 1 mutant, than the wild-type valine residue and therefore may
impinge upon the binding pocket resulting in the con-V208A, had only a modest reduction in Kd (2.3 fold).Competitive displacement of 125I-Mel by 4 drugs (mel- sistent ú5 fold reduction in affinities for all the drugs
tested. Alanine is however 2 carbon groups smalleratonin, N-AS, S20098, and N-NEA) was performed onall receptors and Ki values determined (25) (Table 1). than valine and the small reductions in affinity iden-
tified by mutant V208A may reflect a less significantThe wild-type receptor and both the V208A andV208L mutants displayed the following rank order of effect on the ligand binding site structure. Mutants
H211F and H211L displayed 5 to 8 fold reductions inaffinities, 125I-Mel ú melatonin Å S20098 @ N-NEAú N-AS, which is consistent with what is observed in affinity compared to the wild-type receptor for 125I-
Mel, melatonin and S20098, but interestingly thisovine pars tuberalis tissue (H. E. Howell, personalcommunication). The V208A mutant had affinities was not apparent for the drugs lacking a methoxy
group (N-AS and N-NEA). These drugs actuallyconsistentlyÇ2 fold lower than the wild-type receptorfor all drugs, whereas V208L had affinities between bound to H211F and H211L with affinities approxi-
mately equal to, or slightly greater than, the wild-5 and 12 fold lower than the wild-type. One possibleexplanation of why mutant V208L has greater effects type receptor (Fig. 2). One hypothesis that would ex-
plain these data is if histidine 211 makes a directon ligand binding than V208A is the size of the substi-
421
AID BBRC 7482 / 693b$$$221 09-22-97 21:16:03 bbrcgs AP: BBRC

Vol. 239, No. 2, 1997 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
FIG. 4. Model of the ovine Mel1ab melatonin receptor. The 7 TMs of the ovine Mel1ab melatonin receptor are shown as if viewed fromthe cytoplasmic side of the cell membrane. The model is based on the GPCR model of the adrenergic receptor proposed by Donnelly et al.(27), and highlights valine 208 and histidine 211 on the ligand binding face of TM 5.
interaction with the methoxy groups of 125I-Mel, mela- or any of the 4 mutant receptors by either ligand bind-ing or immunocytochemistry techniques. Nevertheless,tonin and S20098, causing the wild-type receptor to
bind these drugs with increased affinity compared to given that all receptors were expressed equally well inCOS-7 cells it would appear unlikely that the lack ofmutants H211F and H211L. However when structur-
ally related drugs, which do not have a methoxy an observed response of H211F and H211L in HEK293is due to a selective lack of expression of these mutantsgroup, are tested the absence of this putative interac-
tion results in the wild-type receptor displaying simi- in this cell line. The lack of a functional response bythese mutants is very interesting in the light of thelar affinities for N-NEA and N-AS as those observed
for mutants H211F and H211L. Such an interaction proposal that a methoxy group is essential for agonistinduced functional responses in melatonin receptorsbetween histidine 211 and the 5-methoxy group of
melatonin supports the recent theoretical melatonin (26). Potentially the replacement of histidine 211 mim-ics the effect of a drug not having a methoxy groupreceptor model of Navajas et al. (16).
Functional analysis of mutant receptors was per- when bound to the wild-type receptor, i.e. the absenceof the normal histidine 211-methoxy group interactionformed in transiently transfected HEK293 cells, by
measuring the melatonin mediated inhibition of a- results in the receptor maintaining an inactive state.As valine 208 and histidine 211 are totally conservedNDP-MSH stimulated cAMP levels (Fig. 3). HEK293
cells were chosen for functional studies as COS-7 cells in all identified melatonin receptors, it is most probablethat the effects identified in this study will be commonare not suitable for analysis of GPCRs which lower
cAMP levels via a Gia G protein subunit (22). The wild- to the entire melatonin receptor family.This study represents the first investigations intotype receptor and mutants V208A and V208L all inhib-
ited cAMP levels in transfected HEK293 cells, with IC50 melatonin receptor structure and function using site-directed mutagenesis. These data identify that the mel-values in the same rank order as their respective mela-
tonin binding affinities (Fig. 3). This confirms that va- atonin receptor has a binding site involving residuesin TM5, in common with other GPCRs such as the b2-line 208 appears to be in the melatonin receptor ligand
binding pocket, but is not absolutely required for recep- adrenergic receptor. A model of the ovine Mel1ab recep-tor, based on the adrenergic GPCR model of Donnellytor function. Both mutants H211F and H211L did not
show any melatonin mediated effects in HEK293 cells, et al. (27) is shown, and highlights valine 208 and histi-dine 211 on the predicted ligand binding face of TM 5even at concentrations of 1005 M (data not shown), sug-
gesting that histidine 211 may be essential for melato- (Fig. 4). The use of this model will allow further experi-ments to be designed, targeting other residues on othernin mediated receptor function. Due to the low tran-
sient expression levels in HEK293, however, we were helices which may be involved in melatonin receptorligand binding interactions.unable to determine expression levels of the wild-type
422
AID BBRC 7482 / 693b$$$221 09-22-97 21:16:03 bbrcgs AP: BBRC

Vol. 239, No. 2, 1997 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
10. Reppert, S. M., Weaver, D. R., and Ebisawa, T. (1994). NeuronACKNOWLEDGMENTS13, 1–20.
11. Barrett, P., Conway, S., Jockers, R., Strosberg, A. D., Guardiola-We thank April Newman for DNA sequencing, and Diane Slater, Lemaitre, B., Delagrange, P., and Morgan, P. J. (1997) Biochim.Lynda Williams, Janice Drew, Nigel Bunnett, and Patrick Gamp for Biophys. Acta. 1356, 299–307.technical information and assistance with the immunocytochemistry12. Sugden, D., Chong, N. W. S., and Lewis, D. F. V. (1995) Br. J.studies. This study was supported by SOAEFD and a grant from
Pharmacol. 114, 618–623.Servier, France.13. Strader, C. D., Candelore, M. R., Hill, W. S., Sigal, I. S., and
Dixon, R. A. F. (1989) J. Biol. Chem. 264, 13572–13578.14. Ho, B. Y., Karschin, A., Branchek, T., Davidson, N., and Lester,REFERENCES
H. A. (1992) FEBS Lett. 312, 259–262.15. Kao, H.-T., Adham, N., Olsen, M. A., Weinshank, R. L., Bran-1. Cassone, V. M. (1990) Trends Neurosci. 13, 457–464.
chek, T. A., and Hartig, P. R. (1992) FEBS Lett. 307, 324–328.2. Williams, L. M., Lincoln, G. A., and Morgan P. J. (1995) in Rumi-
16. Navajas, C., Kokkola, T., Poso, A., Honka, N., Gynther, J., andnant Physiology: Digestion, Metabolism, Growth and Reproduc-Laitinen, J. T. (1996) Eur. J. Pharmacol. 304, 173–183.tion: Proceedings of the Eigth International Symposium on Ru-
17. Barrett, P., MacDonald, A., Helliwell, R., Davidson, G., and Mor-minant Physiology (Engelhardt, W. V., Leonhard-Marck, S.,gan, P. J. (1994) J. Mol. Endocrinol. 12, 203–213.Breves, G., and Giesecke D., Eds.), pp. 541–561, Ferdinand Enke
18. Cullen, B. R. (1987) Methods Enzymol. 152, 684–704.Verlag, Stuttgart, Germany.19. Conway, S., Drew, J. E., Canning, S. J., Barrett, P., Jockers, R.,3. Reppert, S. M., Weaver, D. R., Rivkees, S. A., and Stopa, E. G.
Strosberg, A. D., Guardiola-Lemaitre, B., Delagrange, P., and(1988) Science 242, 78–81.Morgan. P. J. (1997) FEBS Lett. 407, 121–126.
4. Morgan, P. J., Williams, L. M., Davidson, G., Lawson, W., and20. Scatchard, G. (1949) Ann. NY Acad. Sci. 51, 660–672.Howell, E. (1989) J. Neuroendocrinol. 1, 1–4.21. Bradford, M. M. (1976) Anal. Biochem. 72, 248–254.
5. Morgan, P. J. and Williams, L. M. (1996) Rev. Reprod. 1, 153– 22. Wong, Y. H. (1994) Methods Enzymol. 238, 81–94.161.23. Morgan, P. J., Davidson, G., Lawson, W., and Barrett, P. (1990)
6. Arendt, J., Aldhous, M., and Wright, J. (1988) Lancet 8588, 772– J. Neuroendocrinol. 2, 773–776.773. 24. Reppert, S. M., Weaver, D. R., and Godson, C. (1996) Trends
7. Palm, L., Blennow, G., and Wetterberg, L. (1991) Annals Neurol- Pharmacol. Sci. 17, 100–102.ogy 29, 193–198. 25. Cheng, Y.-C., and Prusoff, W. H. (1973) Biochem. Pharmacol. 22,
8. Arendt, J., Aldhous, M., and Marks, V. (1986) Br. Med. J. 292, 3099–3108.1170. 26. Heward, C. B., and Hadley, M. E. (1975) Life Sci. 17, 1167–1178.
27. Donnelly, D., Findlay, J. B. C., and Blundell, T. L. (1994) Recep-9. Ebisawa, T., Karne, S., Lerner, M. R., and Reppert, S. M. (1994)Proc. Natl. Acad. Sci. USA 91, 6133–6137. tors and Channels 2, 61–78.
423
AID BBRC 7482 / 693b$$$221 09-22-97 21:16:03 bbrcgs AP: BBRC













![Strategie Ovine [Compatibility Mode]](https://static.fdocuments.us/doc/165x107/577cd4611a28ab9e78985bbf/strategie-ovine-compatibility-mode.jpg)




