
The role of competition for light in interference between ...
95
Retrospective eses and Dissertations Iowa State University Capstones, eses and Dissertations 1989 e role of competition for light in interference between velvetleaf (Abutilon theophrasti) and soybean (Glycine max) Williiam Charles Akey Iowa State University Follow this and additional works at: hps://lib.dr.iastate.edu/rtd Part of the Agricultural Science Commons , Agriculture Commons , and the Agronomy and Crop Sciences Commons is Dissertation is brought to you for free and open access by the Iowa State University Capstones, eses and Dissertations at Iowa State University Digital Repository. It has been accepted for inclusion in Retrospective eses and Dissertations by an authorized administrator of Iowa State University Digital Repository. For more information, please contact [email protected]. Recommended Citation Akey, Williiam Charles, "e role of competition for light in interference between velvetleaf (Abutilon theophrasti) and soybean (Glycine max) " (1989). Retrospective eses and Dissertations. 8909. hps://lib.dr.iastate.edu/rtd/8909
Transcript of The role of competition for light in interference between ...

Retrospective Theses and Dissertations Iowa State University Capstones, Theses andDissertations
1989
The role of competition for light in interferencebetween velvetleaf (Abutilon theophrasti) andsoybean (Glycine max)Williiam Charles AkeyIowa State University
Follow this and additional works at: https://lib.dr.iastate.edu/rtd
Part of the Agricultural Science Commons, Agriculture Commons, and the Agronomy and CropSciences Commons
This Dissertation is brought to you for free and open access by the Iowa State University Capstones, Theses and Dissertations at Iowa State UniversityDigital Repository. It has been accepted for inclusion in Retrospective Theses and Dissertations by an authorized administrator of Iowa State UniversityDigital Repository. For more information, please contact [email protected].
Recommended CitationAkey, Williiam Charles, "The role of competition for light in interference between velvetleaf (Abutilon theophrasti) and soybean(Glycine max) " (1989). Retrospective Theses and Dissertations. 8909.https://lib.dr.iastate.edu/rtd/8909

INFORMATION TO USERS
The most advanced technology has been used to photograph and reproduce this manuscript from the microfilm master. UMI films the text directly from the original or copy submitted. Thus, some thesis and dissertation copies are in typewriter face, while others may be from any type of computer printer.
The quality of this reproduction is dependent upon the quality of the copy submitted. Broken or indistinct print, colored or poor quality illustrations and photographs, print bleedthrough, substandard margins, and improper alignment can adversely affect reproduction.
In the unlikely event that the author did not send UMI a complete manuscript and there are missing pages, these will be noted. Also, if unauthorized copyright material had to be removed, a note will indicate the deletion.
Oversize materials (e.g., maps, drawings, charts) are reproduced by sectioning the original, beginning at the upper left-hand corner and continuing from left to right in equal sections with small overlaps. Each original is also photographed in one exposure and is included in reduced form at the back of the book. These are also available as one exposure on a standard 35mm slide or as a 17" x 23" black and white photographic print for an additional charge.
Photographs included in the original manuscript have been reproduced xerographically in this copy. Higher quality 6" x 9" black and white photographic prints are available for any photographs or illustrations appearing in this copy for an additional charge. Contact UMI directly to order.
University Microfilms International A Bell & Howell Information Company
300 North Zeeb Road, Ann Arbor, Ml 48106-1346 USA 313/761-4700 800/521-0600


Order Number 8920106
The role of competition for light in interference between velvetleaf (Abutilon theophrasti) and soybean {Glycine max)
Akey, William Charles, Ph.D.
Iowa State University, 1989
U M I 300N.ZeebRd. Ann Arbor, MI 48106


The role of competition for light in interference between
velvetleaf (Abutilon theophrastil and soybean (Glycine max")
by
William Charles Ak^
A Dissertation Submitted to the
Graduate Faculty in Partial Fulfillment of the
Requirements for the Degree of
DOCTOR OF PHILOSOPHY
Departments: Agronomy Botany
Co-major: Crop Production and Physiology Botany (Ecology)
/proved:
In Charge of Work
For the Major Departments
Fp^he Graduate College
Iowa State University AmeSj Iowa
1989
Signature was redacted for privacy.
Signature was redacted for privacy.
Signature was redacted for privacy.

ii
TABLE OF CONTENTS
Page
INTRODUCTION 1
LITERATURE REVIEW 4
MATERLUS AND METHODS 13
RESULTS AND DISCUSSION 23
SUMMARY AND CONCLUSIONS 78
LITERATURE CITED 82
ACKNOWLEDGEMENTS 88

1
INTRODUCTION
Velvetleaf (Abutilon theophrasti Medic.) is an important weed throughout
the region where maize and soybean are grown in the United States, and its
geographical range is still expanding (Spencer, 1984). Soybean yield losses caused
by full season interference from velvetleaf can be large depending on the numbers
of velvetleaf present. Velvetleaf densities ranging from 1.6 to 2.5 plants/m^ have
been reported to reduce soybean yields by 13 to 31%, respectively (Dekker and
Meggitt, 1983aj Hagood et al., 1980; Oliver, 1979).
The majority of studies relating to interference between velvetleaf and
soybean have dealt primarily with effects of velvetleaf on soybean morphology
and yield. Velvetleaf interference typically results in reductions in soybean dry
weight, leaf area, numbers of branching and flowering nodes, leaf area index
(LAI), and numbers of pods and seeds produced (Dekker and Meggitt, 1983a and
1983b; Hagood et al., 1980; Higgins et al., 1984; Munger et al., 1987a; Oliver, 1979;
Salisbury, 1985). The effect of velvetleaf interference on branching in soybean
appears to be variable; branching may either be reduced (Dekker and Meggitt,
1983a; Salisbury, 1985) or not affected (Dekker and Meggitt, 1983a; Higgins et al.,
1984) by velvetleaf interference. The height of soybean plants has been reported
to be either increased (Salisbury, 1985) or decreased (Munger et al., 1987a) by
velvetleaf interference.
Comparatively few studies have reported effects of soybean interference on
velvetleaf growth and yield, Higgins et al. (1984) reported that leaf area, number
of nodes with fully developed leaves, canopy width, number of branches, and
number of o^sules of velvetleaf grown in monoculture were all higher con^ared
to those of velvetleaf intercropped with soybean. Similarly, lAI of velvetleaf may

2
be reduced by as much as 75% due to full season interference from soybean
(Oliver, 1979). These results suggest a considerable potential for soybean to
interfere with the vegetative and reproductive growth of velvetleaf.
Despite the fact that velvetleaf is considered a very serious weed in
soybean production and that much research has been devoted to examining
interference between these two species, precisely how velvetleaf interference
reduce? soybean yields is not known. Several investigators have suggested that
velvetleaf interference is due to allelopathy (Bhowmik and Doll, 1982; Colton and
Einhellig, 1980; Retig et al., 1972). Studies on competition between nodulating and
non-nodulating soybeans and three annual weeds suggest that the competitive
ability of soybean is not limited by nitrogen supply (Staniforth, 1962). Dekker and
Meggitt (1983a) concluded that interference for soil nutrients was not involved in
observed reductions in growth of soybean intercropped with velvetleaf.
Competition for water may play a role in interference between velvetleaf and
soybean, especially under conditions of limited soil water availability (Bunce, 1985;
Cheetham 1989; Munger et al., 1987a and 1987b). Much, if not all, of the
interference exerted by velvetleaf on soybean may be due to competition for light
(Stoller and Woolley, 1985). Velvetleaf has an average area of influence of about
0.4 m2 (stoller and Woollqr, 1985). Since velvetleaf plants are taller than soybean
plants over much of the growing season (Frazee and Stoller, 1974), a density of 1
to 2 velvetleaf plants/m^ would produce a canopy that could shade all of the
shorter soybean plants.
Although competition for light appears to play an Important role in
interference between velvetleaf and soybean, little is known about the processes
involved. The objectives of the present study were to; l) examine the

3
development of velvetleaf and soybean grown separately, or together in mixtures,
over the growing season in terms of changes in dry weight and seed yield; and 2)
evaluate the contribution of competition for light to interference between
velvetleaf and soybean. For this purpose, a deWit replacement series (deWit,
1960) was used. Such series have been widely used to interpret competitive
Interactions between two species in mixed populations (Trenbath, 1974; Hall,
1974). Several recent papers have raised objections to the use of the replacement
series design for the following reasons: l) it cannot assess the relative
contributions of inter- and intraspecific interference to the determination of
mixture yields (Jolliffe et al., 1984); 2) it confines attention to one density only
(Firbank and Watkinson, 1985); and 3) it tends to favor larger species in mixtures
(Connolly, 1986). In the present report, a single population density within the
range typical for agronomic production of soybean was chosen. Since the primary
objective was to characterize the relative competitive ability of the two species
rather than to determine the relative importance of intra- versus interspecific
competition in forming mixture yields (Roush and Radosevich, 1985), it was felt
that this design was adequate.

4
LITERATURE REVIEW
Several researchers have examined interference between velvetleaf and
soybean, and the effects of this interference on the vegetative and reproductive
growth of soybean are well documented (Dekker and Meggitt, 1983a and 1983b;
Eaton et al., 1976; Hagood et al., 1980; Higgins et al., 1984; Munger et al., 1987;
Oliver, 1979; Staniforth, 1965). However, the mechanism(s) by which these
effects occur is (are) poorly understood. Interference between velvetleaf and
soybean could potentially involve allelopathy and/or competition for nutrients,
water, or light (Harper, 1977; Zimdahl, 1980).
Allelopathy
Several studies have examined the allelopathic potential of velvetleaf.
Soybean yields were reduced by 14% following incorporation of velvetleaf plant
residues into the soil (Bhowmik and Doll, 1982). Leaf extracts of velvetleaf have
been shown to inhibit soybean seedling growth and chlorophyll production (Colton
and Einhellig, 1980), and velvetleaf seeds are known to affect adversely the
germination and seedling growth of several crop species (Elmore, 1980; Gressel and
Holm, 1964; Retig et al., 1972). However, exudates of glandular trichomes of
velvetleaf do not {^pear to have allelopathic activity under field conditions
(sterling and Putnam, 1887).
Conq>etition for Nutrients
Studies specifically designed to test for con^>etition for nutrients between
velvetleaf and soybean have not been reported in the literature to date.
Competition between nodulating and non-nodulating soybeans and other annual

5
weeds suggest that the competitive ability of soybean is not limited by nitrogen
supply (Staniforth, 1962). From tissue analyses conducted on leaves of
intercropped velvetleaf and soybean, Dekker and Meggitt (1983a) concluded that
interference for soil nutrients was not involved in observed reductions in soybean
growth. However, nutrient competition need not lead to differences in tissue
nutrient concentrations. A species might maintain a fairly constant tissue nutrient
concentration, but increase its biomass and so acquire more nutrients. With a
fixed nutrient pool size, competition for nutrients could occur among species of
differing sizes even if their tissue nutrient concentrations are not different.
The major pulse of nutrient availability in temperate systems occurs in
spring when decomposition activity is stimulated by warmth and moisture and
when fertilizers are applied (Benner and Bazzaz, 1985). Nutrient pulses may also
occur later in the season due to rainfall after a dry period or to senescence of
neighboring plants that were formerly nutrient sinks. Velvetleaf shows an
opportunistic response to nutrient availability, being able to use nutrients supplied
at different times for reproduction (Benner and Bazzaz, 1985). However, seed
production of velvetleaf may be reduced if nutrient availability is low until
fruiting has begun. Soybean may not be as opportunistic as velvetleaf in nutrient
acquisition. Although foliar fertilization with relatively large amounts of
nutrients during early pod filling has shown potentifU for increasing soybean seed
yields, the yield increases have not been consistent or dependable (Shroyer, 1978).
How differential ability to respond to the timing of nutrient availability might
affect con^tition for nutrients between velvetleaf and soybean is presently
unknown.

6
Competition for Water
With normal rainfall, competition for water does not appear to play a role
in interference between velvetleaf and soybean (Cheetham, 1989; Munger et al.,
1987b). However, Cheetham (1989) found that velvetleaf adversely affected
soybean water relations in a drier than normal year. Conversely, Munger et al.
(1987a) reported reduced photosynthesis, transpiration, stomatal conductance, and
leaf water potential in velvetleaf intercropped with soybean, compared to
velvetleaf in monoculture, under drier than normal conditions. Thus, under field
conditions, the role of competition for water in interference between velvetleaf
and soybean is unclear.
Soybean water use efficiency (WUE), the amount of carbon fixed per unit of
water used, appears to be higher when soybean is well watered than when water
is limited (Baldocchi et al., 1983). Under field conditions, carbon dioxide
exchange rates (CER) of soybean are insensitive to water stress early in the
morning, but are severely depressed after mid-moming, primarily due to high
stomatal resistance, high air temperature, and low leaf water potentials. Hatfield
and Carlson (1879) similarly reported an initial time lag after sunrise before leaf
water potentials of soybean began to decrease.
Several researchers have examined water relations and gas exchange
parameters of velvetleaf and soybean under controlled environment conditions.
However, a consistent pattern in reported results is lacking, and the relevance of
these laboratory studies to field situations is not established. Patterson and Flint
(1989) found no differences in net photoqnathesis or WUE between velvetleaf and
soybean under well-watered conditions. Bunce (1985) reported a more rapid
decline in both C02-saturated and normal photoqmthetic rates of soybean

7
compared to velvetleaf, as the relative water content and osmotic potential of
intact leaves were slowly decreased. Neither velvetleaf nor soybean underwent
significant osmotic adjustment during the period of water stress (Bunce, 1985).
Under well-watered conditions in the greenhouse, velvetleaf exhibited
higher stomatal conductance, photosynthesis, and transpiration than soybean
(Munger et al., 1987b). However, these parameters declined more rapidly in
velvetleaf than in soybean when these species were subjected to water stress.
These results suggest that velvetleaf has a higher threshold leaf water potential
for stomatal closure than does soybean. Munger et al. (1987b) concluded that
interspecific con^etition for water would have a greater effect on the
photoqoithetic activity and competitive ability of velvetleaf compared to soybean.
In contrast to these results, Patterson and Flint (1983) found that, compared to
seven dicotyledonous weed species including velvetleaf, stomatal closure in
response to increasing water stress was most abrupt in soybean, occurring over a
range of 2 to 3 bars. Stomatal closure in velvetleaf was also relatively abrupt, but
the threshold leaf water potential at which stomatal closure began was lower
than that for soybean. Therefore, even if soil water availability was sufficiently
limited so that competition for water occurred between velvetleaf and soybean, it
is difficult to predict which species^ growth would be more affected, given the
inconsistency in reported results.

8
Competition for Light
Competition for light occurs in nearly all cropping situations (Donald, 1961),
but little is known about the processes involved, and data to assess unit crop
growth reduction per unit of light lost through competition are generally
unavailable.
A number of plant attributes can be identified as being potentially
important in determining a species^ ability to compete for light, including rate of
partitioning of plant biomass into leaf area (and consequent rate of development of
canopy shade over neighboring species), LAI, leaf arrangement, leaf angle, petiole
length, and height (Donald, 1961: Patterson, 1982; Rhodes and Stem, 1978; Spitters
and Aerts, 1983).
Height
Competition for light occurs ultimately not between q)ecies or plants, but
between leaves (Donald, 1961). Therefore, the physical positioning of the foliage
for light interception is especially important. Any factor that differentially alters
the ability of a plant to intercept light in relation to that of neighboring plants
should influence the plant's ability to compete for light (Rhodes and Stem, 1978).
Although soybean has a very reqpid initial growth rate and attains a height
advantage over many weed species, including velvetleaf, soon after emergence
(Frazee and StoUer, 1974), velvetleaf plants are usually taller later in the growing
season. Maximum shading of soybean occurs during the last 8 to 10 weeks of the
growing season (Stoller and Woolley, 1985). Individual velvetleaf plants have an
area of influence of 0.4 m^ and can intercept 44 to 56% of available RAR (Stoller
and Woollqr, 1985). Therefore, 2 to 3 velvetleaf plants/m^ would be sufficient to
form a full canopy that would intercept most of the available PAR above shorter

8
soybean plants. Based on equivalent levels of soybean yield reductions caused by
natural shading from a velvetleaf canopy or by an equal level of artificial shading,
Stoller and Woolley (1985) concluded that most, if not all, of the interference from
velvetleaf infestations in soybean could be ascribed to con^etition for light.
In addition to reducing the quantity of light received by soybean, shading
by overtopping velvetleaf also suppresses branching in soybean by altering the
quality of light they receive) shading increased the proportion of far-red
wavelengths (Begonia and Aldrich, 1988). This could reduce yields of soybean by
reducing the number of potential fruiting nodes. In addition to suppression of
branching, Myers et al. (1987) reported increased abscission of young reproductive
structures of soybean exposed to enhanced levels of far-red wavelengths,
compared to those exposed to normal light.
Leaf area partitioning and expansion rate
Under growth chamber conditions, velvetleaf had a more rapid leaf area
expansion rate and partitioned biomass into leaf area more rapidly during the first
4 weeks after planting than did soybean (Potter and Jones, 1977). Following a 29
day period of growth, Patterson and Flint (1983) similarly found that velvetleaf
partitioned more of its total biomass into leaf area than did soybean. A more
r^id leaf area expansion rate and greater partitioning of total biomass into leaf
area are both factors that would favor rapid canopy development and enhance the
ability of velvetleaf to compete for light with soybean.
Solar-tracking movements of leaves
Another factor which may influence the ability of a species to coiqpete for
light is the phenomenon of diaheliotropism. Diaheliotropism, or solar-tracking,
involves the active movement of leaf blades during the day to maximize light

10
interception. Diaheliotropic leaf movements have been reported in at least 16
families (Ehleringer and Forseth, 1980) which include crop species such as cotton
(Lang, 1973)j sunflower (Shell et al., 1974), and soybean (Kawashima, 1969a).
Evidence suggests that varietal differences exist in the solar-tracking ability of
soybean (Kawashima, 1969aj Wofford and Allen, 1982). Solar-tracking plants have
more quanta available for photoqmthesis and a greater potential daily
photo^thetic gain than nontracking species (Mooney and Ehleringer, 1978; Shell
and Lang, 1976). Thus, solar-tracking leaf movements may give a species a
competitive advantage in mixture with nontracking or weakly-tracking species
(Ehleringer and Forseth, 1980). Diaheliotropism in velvetleaf has not been
reported to date.
Growth analysis
Mathematical growth analysis provides a convenient means of examining
the processes of total dry matter production and leaf area expansion that
determine a plantas vegetative growth and potential competitiveness under a
variety of environmental conditions (Patterson, 1982). Relative growth rate (RGR),
leaf area ratio (lAR), and net assimilation rate (NAR) are three growth analysis
parameters on a per plant basis which are particularly useful in evaluating the
response of plants to irradiance during growth (Patterson, 1982).
RGR measures the rate of increase in plant dry weight per unit of existing
plant dry weight per unit of time (Hunt, 1982) and represents the efficiency of
the plant as a producer of new material (Blackman, 1919). Any departure from an
adequate supply of light or other environmental resource will decrease RGR (Hunt,
1982). LAR is a measure of the relative leafiness of the plant, i.e., the area of
leaf exposed per unit plant dry weight (Hunt, 1978). Changes in LAR result from

11
changes in the amount of leaf biomass that is partitioned into leaf area (i.e.^
changes in leaf thickness in response to changes in the light environment) or
changes in the amount of total plant biomass that is partitioned into leaf biomass.
LAR is an important component of plant growth dynamics because it reflects the
relative amount of photo^mthetic assimilatory surface (Patterson and Flint, 1983).
With equal rates of photo^thesis per unit leaf area, a species with a high LAR
will have a higher RGR than one with a low LAR.
NAR measures the rate of dry weight increase per unit leaf area. NAR
represents the Efficiency of the whole plant as an assimilating system and
provides a rough estimate of the carbon assimilation c^acity of the leaves (Hunt,
1978).
Under growth room conditions, Potter and Jones (1977) found RGR and NAR
of velvetleaf to be consistently higher than those of soybean during early growth.
Similarly, Patterson and Flint (1983) observed that velvetleaf had higher LAR and
NAR than soybean after 4 weeks of growth in a controlled environment
greenhouse. Under field conditions, Cheetham (1989) also found that velvetleaf
allocated more resources to leaf production than soybean early in the growing
season, but this pattern was reversed later in the season. However, the RGR and
NAR of velvetleaf were consistently higher than those of soybean (Cheetham,
1989).
light wwiysis
On a canopy basis, traditional growth analysis involves evaluation of crop
growth rate (CGR) as a product of LAI and NAR (Hunt, 1982). However, this
approach cannot directly accommodate changes in the crop environment during
growth (Charles-Edwards and Fisher, 1980). Light is the foremost environmental

12
factor affecting growth and is usually the most variable one. Light conversion
analysis (Warren Wilson, 1981), as used in the present stu<ty, directly incorporates
light into the growth analysis equation and, therefore, represents a more direct
way of assessing a species^ ability to compete for light. However, to date, few
researchers have employed this technique (Hunt et al., 1984; Chazdon, 1985).
In summary, from the literature, velvetleaf appears to possess several
attributes which should lend it a strong capability to compete for light.
Moreover, competition for light ^pears to play a prominent role in interference
between velvetleaf and soybean, but the mechanism(s) by which this occurs
remains to be ascertained.

13
MATERIALS AM) METHODS
Interference Between Velvetleaf and Soybean: Seasonal Changes in Growth and
Yield
General procedures
Experiments were conducted at the Woodruff and Agronomy farms of Iowa
State University, Ames, Iowa in 1986 and 1987) respectively. The soil type was a
Clarion loam (fine loamy, mixed mesic, typic Hapludoll) with 6.6% organic matter
and pH 7.6 in 1986 and 3.4% organic matter and pH 5.4 in 1987.
Cultural practices were similar in 1988 and 1887. Seeds of the soybean
cultivar, 'BSR 201', used in all studies reported here, were planted on June 4
(1986) or June 5 (1987) with a six row planter in north-south oriented rows
spaced 0.78 m f^art. Velvetleaf seeds were planted directly into the soybean
rows with a Planet Jr. seeder immediately following planting of the soybeans. In
all studies reported here, velvetleaf seeds were obtained locally and acid-
pretreated to break dormancy (Andersen, 1968) prior to planting. Each species
was seeded at a rate of about 500,000 seeds/ha.
Weed control was achieved with a 2.2 kg ai/ha pre-emergence application
of alachlor [2-chloro-N-(2,6-diethylphenyl)-N-(methoxymethyl) acetamide] ^plied
at 94 L/ha and 206 kPa with a CO2 bicycle sprayer, followed by hand weeding for
. the remainder of the growing season.
Experimental layout and design
The experiment was arranged as a randomized con^lete block design with
four replications of five treatments. Each experimental unit consisted of a 7 m
long plot containing six rows. In each plot, the outside two rows and a 0.5 m

14
section at either end of each plot were excluded from sampling. Treatments
consisted of a deWit replacement series (deWit, 1860) containing the following
velvetleaf/soybean species-proportions: 100/0, 50/50, 25/75, 12.5/87.5, 0/100,
generated by counting and thinning both species at 1 and 2 weeks after
emergence. Following the final thinning, the total plant population in both the
mixtures and monocultures was about 370,000 plants/ha.
Sffliptog prQçgdurgs
At 30 (1986) or 24 (1987) days after planting (DAP), 10 sections of row, each
1 m in length, were selected at random in each plot except that no two selected
sections were permitted to be adjacent within or between rows. At
approximately 2 week intervals during the growing season, beginning at 31 DAP
(1986) or 25 DAP (1987), a sampling site was randomly chosen in each plot from
the previously marked row sections and all plants were then harvested from each
row section by cutting at soil level. Six plants were randomly selected from each
of these harvests, three of each species (except in the 12.5 V/87.5 S species
proportion in which only three velvetleaf plants were present, all of which were
used). The height of each plant to the tip of the mainstem axis was recorded and
the leaf area was measured with a leaf area meter^. The number of mainstem
branches/plant and their distribution with height (1987 only) were also
determined at the final san^ling date. Mainstem branch distribution was
determined by dividing the stem into 0.15 m segments and counting the number of
branches off the mainstem in each segment. Each height increment was
subjected to a separate two-way analysis of variance (ANOVA). Dry weights were
^ Model LI-3000. LiCor Inc., Lincoln, NE 68504.

recorded separately by species following oven drying at 60 C. The total dry
weight of the remaining harvested plants of each species was recorded, following
oven drying at 60 C, and combined with those previously determined to obtain a
better estimate of total dry weight/unit ground area for each species proportion.
At maturity (ill DAP, 1986; 120 DAP, 1987 ), all remaining soybean plants in
each plot (except from the border area previously described as excluded from
sampling were harvested by hand and the seeds were removed by threshing.
The moisture-content of each seed sample was determined with a moisture meter^
and sample weights were adjusted to express seed yields on a common (13%)
moisture content basis. In 1987, velvetleaf seed pods were also harvested, as they
matured, from three randomly selected velvetleaf plants in each plot and oven
dried at 35 C. Seeds were subsequently removed from the pods by hand, cleaned,
and weighed.
gtAtlPtW mWyg*5
Results for 1886 and 1987 were subjected to separate statistical analyses.
Except as noted otherwise, a separate two-way ANOVA was performed on results
for each sazqpling date and differences among means were evaluated using Fisher^s
Protected LSD test. Relative yields (RY) of velvetleaf and soybean were analyzed
graphically for each sampling date as described by deWit (1980) and Harper (1977)
to determine the relative con^)etitive ability of the two species. The RY of each
species was calculated separately as RY - yield in mixture/yield in monoculture,
where yield refers to dry weight or seed yield/m^ (relative seed yields were
calculated ia 1887 only). Orthogonal polynomials were employed to determine if
2 Dickqr-john Corporation. Auburn, IL 62615.

16
the response of RY to changes in species proportion differed from the response
expected if two species are of equal competitive ability (a diagonal linear line
from 0 to l).
Results were also analyzed mathematically using relative yield totals (RYT)
as described by Harper (1977) and relative crowding coefficients (RCC) (deWit,
I960). The RYT was calculated for each species proportion as RYT - RY
(velvetleaf) + RY (soybean). The RGCj a measure for comparing the relative
aggressiveness of one species towards another, was calculated separately for each
species. Values of RCC - 1 indicate ^proximately equal con^etitive ability, while
values >1 or < 1 indicate that a species is more or less competitive, respectively,
than another species (deWit, 1960). Data for monocultures were not included in
aiy statistical analysis, but were included in the determination of RYT and RCC.
RCC and RYT values were averaged over the three mixtures for each species.
Deviations of RYT from 1.0 were determined with a Student's t-test and
differences in ROD between species were determined by an F-test.
Con^Tetition for Light: Whole Plant
Although experiments reported here were not expressly designed to test
for competition for water, covered leaf water potentials (LWP) (Meyer and Ritchie,
1980) were measured in 1987 to determine the potential contribution of
competition for water to interference between velvetleaf and soybean. On days
corresponding with plant harvests (25, 41, 53, and 85 DAP), one plant of each
species was chosen at random in each plot from areas adjacent to those sampled
for growth parameters and the most recently fully expanded leaf of each plant
was covered with aluminum foil in the early evening to exclude all light. LWP

17
was measured early the following morning using a portable pressure chamber^
immediately after replacement of the foil with a plastic bag. All LWP
measurements were con^leted by 9 a.m. local time. A preliminary study
confirmed that LWP measured in this manner did not differ from pre-dawn LWP.
growth qnalygjg
Data from individual harvests were used to derive RGR, lARj and NAR
according to methods outlined by Hunt (1882). RGR is the rate of increase in plant
dry weight per unit existing dry weight per unit time. lAR is the ratio of leaf
area to plant dry weight. NAR is the rate of increase in plant dry weight per
unit leaf area. Fisher's LSD test was employed to assess the effect of changing
the proportion of a species in the population on growth analysis parameters of
that species.
Competition for light: Canopy
The lAI was calculated for each species proportion (Hunt, 1982) from the
combined leaf areas of velvetleaf and soybean.
light çfflvgrsion analysis
light conversion analysis (Warren VRlson, 1881) involves the analysis of
crop growth rate (CGF) as a product of light energy incident on a canopy Go), the
efficiency of light interception by the canopy (LIE)^ and the efHciency of
utilization of light for dry matter production by the canopy (LUE). Iq (MJ in~2
day"!) in energy terms was derived from global shortwave radiation data
3 Model 3005. Soilmoisture Equipment Corp.* Santa Barbara^ CA 83105.

18
abstracted from meteorological records of Iowa State University, Ames. In
deriving 1% it was assumed that 50% of total incident shortwave radiation lies
within the 400 to 700 nm waveband.
Since it was impractical to monitor incident light interception continuously^
measurements were taken only at each of the four harvest dates, just prior to
harvesting. A line quantum sensor^, oriented parallel to row direction, was used
to measure the photosynthetic photon flux density (FFFD) at 0.01, 0.19, and 0.38 m
to either side of each selected row section at the soil surface) one reading was
taken directly above the canopy. The six FFFD values measured at the soil surface
were averaged to obtain a mean value of FFFD below the canopy of each plot.
FFFD was recorded automatically at 0.5 s intervals by a micrologger^ and averaged
over a 25 s time period to obtain a better estimate of light interception. All FFFD
measurements were taken at local solar noon i 1 h. The US was then calculated
for each plot using these FFFD measurements.
LUE was calculated in the following manner. A quadratic function was
fitted to a plot of LIE over time (jDAP). Since LIE was not measured at emergence,
an estimated value was used, based on typical expected LAI of soybean or
velvetleaf at emergence. Daily lo and instantaneous daily values of LIE predicted
from the equation of the line were then used to calculate J, the total amount of
incident light (in energy terms) intercepted by canopies of the various species
proportions, summed over each harvest interval from emergence to final harvest^
J - S(LIE X lo). The LUE of each plot was then calculated for each harvest
4 Model LI-191SB. LiCor Inc., Lincoln, NE 68504.
^ Model 2iX. Campbell ScientiHc Inc., Logan, UT 84321.

19
interval as LUE - (W2 - Wi)/Jj where Wi and W2 are the means of dry weight/m^
at the beginning and end of each interval^ respectively.
Data from individual harvest dates were used to calculate CGR for each
species proportion.
Diumal pattern of canopy light interception
Results from a preliminary stucfy in 1886 indicated differences between
canopies of velvetleaf and soybean in their pattern of light interception over the
day. Therefore, in 1987 (July 27), PPFD measurements were taken every 3 h from
800 to 1700 h local solar time above and at several heights within monoculture
velvetleaf and soybean can(^ies in order to quantify their patterns of incident
light interception during the day. Using a line quantum sensor as previously
described, PPFD readings were taken at 0.01 m on each side of the row in 0.2 m
vertical intervals from soil level to a height of 1 m. A single reading of PPFD
incident on each canopy was taken. PPFD readings from each side of the row
were averaged for a better estimate of PPFD at each height.
Percent attenuation of the solar beam by each increment of canopy height
(% PPFD interception) was calculated for each species at each sanq)ling time.
Because height dUfered amongst the canopies the average height of a random
sangle of 10 plants/plot was taken to determine the % of total canopy height the
solar beam had penetrated to reach each sampling position (% canopy penetration).
Solar-trackiny leaf movements and their significance to photosynthesis
Following casual observations in the field of changes in leaf orientation of
velvetleaf and soybean during the day which fç)peared to be diaheliotropic
(Ehleringer and Forseth, 1980), experiments were conducted under greenhouse
(1988) and field (1988) conditions to examine diumal patterns of leaf movement.

20
For the greenhouse study, soyt)ean and velvetleaf were planted (5/pot)
separately in 0.15 m diameter plastic pots containing a greenhouse soil mix
consisting of a 3:1:1 mixture of peat:soil:sand. Pots were watered daily and
fertilized each week with a 20:10:20 (N:P:K) commercial fertilizer mix. At
emergence, both species were thinned to 1 plant/pot. On July 23, when soybean
was at the R2 to R3 growth stage (Fehr et al., 1871) and velvetleaf had 15 to 17
unfolded leaves, the angle of incidence of solar radiation on the most recently
Mly expanded leaf was measured every 2 h from 800 to 1800 h local solar time
with a special inclinometer (Taylor, 1971). A full set of measurements was
typically conq>leted in 30 min. The sine of the angle of incidence, reported here,
is directly proportional to light interception by the leaf.
The Aeld study was conducted at the Agronomy farm on a Clarion loam
having 4.6X organic matter and pH 7.1. Seeds of each species were planted by
hand in sqwate rows at a rate of about 380,000 seeds/ha in six 15 m rows (3
rows per species) q>aced 1.5 m ^art. Hand-weeding was performed as required.
After emergence, plants of both species were thinned so that sampled leaves on
adjacent plants within the rows did not shade each other at the time of
measurement. On August 3, when soybean was at the R4 growth stage and
velvetleaf had 25 unfolded mainstem leaves, 10 plants of each qtecies were chosen
at random and the angle of incidence of solar radiation on the most recently fully
expanded leaf of each plant was measured as described above for the greenhouse
study.
Both experiments were arranged as completely randomized designs with
either eig^t (greenhouse) or ten CTield) replications. The greenhouse experiment
was conducted twice with similar findings] representative results from one

21
experiment are presented here. A separate one-^way ANOVA was conducted on
results from each sampling Urne during the day and differences between species
were evaluated with an F-test.
To assess the significance of the observed leaf movements to the relative
abilities of soybean and velvetleaf to conq>ete for light) the effect of solar-
tracking leaf movements on the CO2 exchange rate (CER) and transpiration rate of
velvetleaf and soybean was investigated under field conditions in 1987 and 1888.
The PPFD incident on the leaves was also measured. Results for 1988j which were
similar to those for 1887^ but from a more con^lete set of measurements, are
rqx>rted here as means ± standard errors.
CERj transpiratioi^ and PPFD were measured for the velvetleaf and soybean
plants used for the 1888 leaf solar-traddng study previously described. However,
these measurements were completed about 2 weeks prior to leaf solar-tracking
measurements on a different set of leaves. The most recently fully expanded leaf
of both species was either constrained in a horizontal position by an enclosure
constructed of wood and monofilament fishing net or allowed to move freely
throughout the day. Measurements were conducted on immobile and freely
moving leaves using a portable infrared gas analysis system consisting of an
infrared gas analyzer^, airflow controller?, and leaf sample chamber^ with
^ Model LCA. Analytical Development Co., Ltd., Hoddesdon, England.
7 Model ASU^4F)-ADA. Analytical Development Co., Ltd.,
Hoddesdon, England.
8 Parkinson Broadleaf Chamber. Analytical Development Co., Ltd.,
Hoddesdoi^ England.

22
attached quantum sensor^. Data were recorded automatically with a micrologger
con^rised of a cooD^uter-controller^^ and Analog-Digital board^l. Measurements
were taken about once per hour on two separate days. Readings were taken
between 600 and 1300 h on July 19, which was cloudy and became overcast after
midday, and between 600 and 1800 h local solar time on July 21, which was
mostly cloudless. Equations of functions derived by polynomial regression and
fitted to transpiration rates and CER over the course of the day were integrated
to obtain total amounts of water transpired and carbon fixed on both the overcast
and clear days. Daily water use efficiencies (WUE) were calculated from the ratio
of total carbon fixed to total water transpired. Data from the clear and cloudy
days were separately analyzed by two-way ANOVA, and differences between
means of horizontal and freely moving leaves for daily totals of water transpired,
carbon fixed, and WUE were determined for each species with an F-test.
Results from different years for solar-tracking studies (leaf movement
studies - 1886, 1988; physiological studies - 1887, 1888) were similarly subjected to
separate statistical analyses. Except as noted otherwise, a separate two-way
ANOVA was performed on results for each sampling date and differences among
means were evaluated using Fisher's Protected LSD test.
9 LiCor Inc., Uncolt^ NE 68504.
^0 Model BGC-52. Micromint, Vemox^ CT 06068.
Model BOC-30. Micromint, Vernon, CT 06066.

23
RESULTS AND DISCUSSION
Interference Between Velvetleaf and Soybean: Seasonal Changes in Growth and
Yield
Two factors must be taken into consideration in the interpretation of the
results of the 1986 interference stu<fy. More than 90% of all soybean seedlings
were infected with tanspot (CorynebaHmrhim famltatnml l to 2 weeks after
emergence. This seedbome bacterial disease caused a temporary but major loss of
leaf area. Considerable leaf area loss also occurred in velvetleaf plants during the
1986 growing season due to infection with verticillium wilt (Verticillium spp.).
Syn^toms (leaf wilting, leaf drop) began appearing 40 to 60 DAP and increased in
severity during the remainder of the growing season.
Relative drv weight
Early in the 1986 growing season, velvetleaf and soybean appeared to escape
competition with each other, since the relative dry weight of each species was
not reduced below its respective monoculture expectation (Figure l). Indeed, at
31 DAP, soybean grew significantly better in mixture than in monoculture. This
may have been an artifact resulting from differential disease levels between
soybeans in mixture and those in monoculture, although the disease appeared to be
fairly evenly distributed over the experimental area. Alternatively, soybean may
somehow have benefitted from its association with velvetleaf or from a reduction
in the level of interference from other soybean plants, i.e., from reduced
intraspecific competition. However, later in the growing season (59 and 75 DAP),
velvetleaf gained resources at the expense of soybean, thereby significantly

Figure 1. Effect of changes in species proportion on the relative dry weight (RDW) of velvetleaf (V) and soybean (S) at 31, 44, 59, and 75 days after planting (DAP) in 1986. Dashed diagonal lines between 0 and 1 depict expected yields in monoculture. The RDW of velvetleaf deviated significantly from its expected yield at 44, 59, and 75 DAP. The RDW of soybean deviated significantly from its expected yield at 31, 59, and 75 DAP
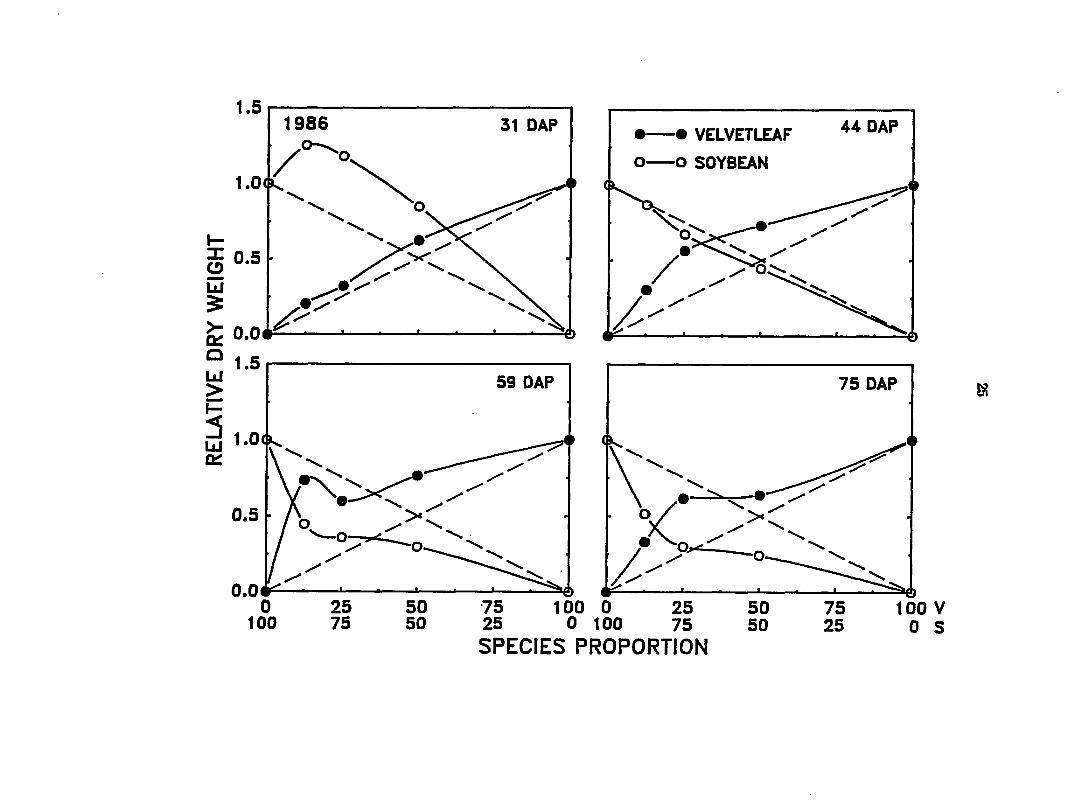
31 DAP
^ 0.5
59 DAP
0.0#: 75 100 0 25 25 0 100 75
SPECIES PROPORTION
44 DAP VELVETLEAF
o o SOYBEAN
75 DAP
100 V
8

26
reducing the growth of soybean in mixtures relative to that in monoculture
(Figure l).
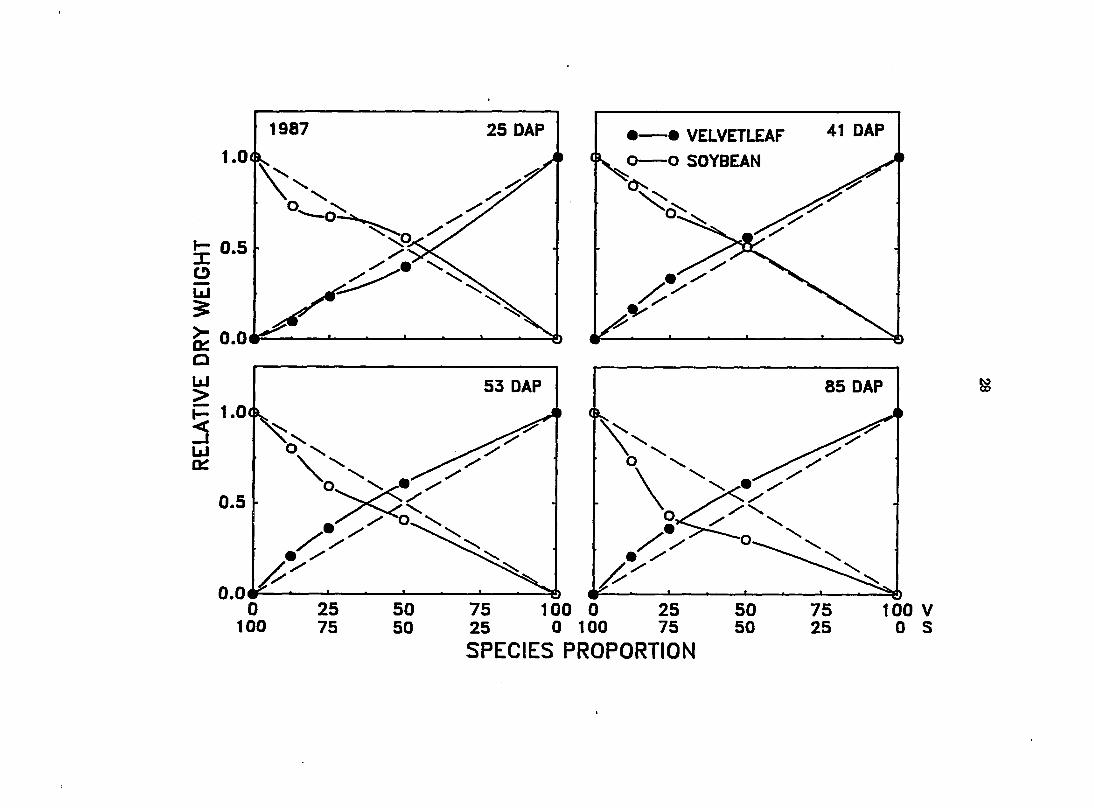
The pattern of Interference development during the early part of the 1987
growing season (Figure 2) differed from that observed in 1986 (Figure l). In
1987, when neither species was diseased, the early season growth of each species
did not seem to be affected by the presence of the other. The relative dry
weight of each species did not deviate significantly from expected monoculture
values. However, as in 1986, later in the 1987 growing season (53 and 85 DAP)
velvetleaf interference significantly reduced the growth of soybean in mixtures
compared to monoculture.
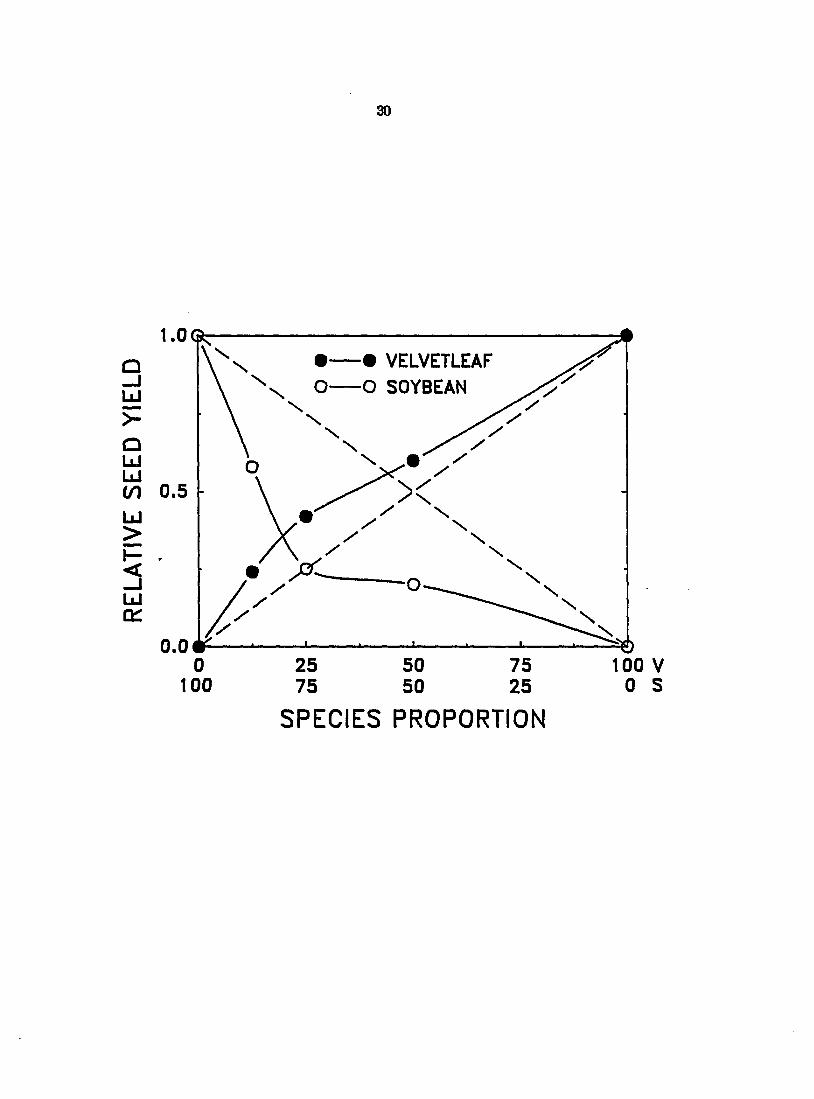
Relative seed yield
When assessed on a relative seed yield basis, velvetleaf again appeared to be
more successful than soybean in acquiring limiting resources (Figure 3). Soybean
seed yields (measured in 1987 only) were reduced below expected monoculture
yields when grown in association with aiy number of velvetleaf. On the other
hand, velvetleaf seed yields were greater in mixture than in monoculture due
either to some beneficial effect of the presence of soybean or to reduced
intraspecific competition.
Relative yield total (RYT)
RYT did not differ significantly from 1.0 at 59 DAP in 1988 (Table 1), when
the reduced growth of soybean in mixture was balanced by the improved growth
of velvetleaf in mixture (Figure l), or on any sampling date in 1987, thus implying
that the two species were making demands on the same limiting resources
(Harper, 1977). Early in the 1988 growing season (31 and 44 DAP), RYT values for
mixtures of velvetleaf and soybean were found to be significantly greater than 1.0

Figure 2. Effect of changes in species proportion on the relative dry weight (RDW) of velvetleaf (V) and soybean (S) at 25, 41, 53, and 85 days after planting (DAP) in 1987. Dashed ^agonal lines between 0 and 1 depict expected yields in monoculture. The RDW of both qpecies deviated significantly from their respective expected yields at 53, and 85 DAP only
25 DAP
53 DAP
N
-e VELVETLEAF
-O SOYBEAN
41 DAP
100 0 0 100
SPECIES PROPORTION
85 DAP
100 V
8

Figure 3. Effect of changes in species proportion on the relative seed yields (RSY) of velvetleaf (V) and soybean (S) in 1987. Dashed diagonal lines between 0 and 1 depict expected yields in monoculture. The EST of both species deviated significantly from their expected yields
30
l .OC^
e VELVETLEAF O SOYBEAN
LU
LU LU (/) 0.5 LU
LU
25 50 75 100 V 100 75 50 25 OS
SPECIES PROPORTION

31
Table 1. Effect of competition on the relative yield total (RYT) and relative crowding coefficient (ROC) of velvetleaf and soybean grown in mixture under field conditions. ROC values are appropriate only when RYT is not significantly different from 1.0
RCCa
Year DAP^ RYT^ Soybean Velvetleaf
1986 31 44 59 75
1.47** 1.19N* 1.07 0.79**
0.22 5.02**
1987 25 41 53 85
0.90 1.03 0.99 1.01
1.19 0.79 0.60 0.35
0.89 1.28** 1.83** 3.21**
^Calculated from dry weights averaged over the three mixtures. Within a sandaling time (DAP)^ asterisks indicate means that are significantly different at the P - 0.01 level according to the F-test.
()Days after planting.
^Calculated from dry weights averaged over the three mixtures. Asterisks indicate means significantly different from 1.0 according to Student's t-test at P - 0.01.

32
(Table l). This may reflect a difference in the use of resources by the two
species or the avoidance of some measure of competition with each other as
earlier indicated from the graphical interpretation of Figure !.. However^ later
in the season (75 DAP), the RYT fell below 1.0 (Table l) when soybean growth
was markedly reduced by velvetleaf interference. This was not compensated for
by improved growth of velvetleaf in mixture.
RYT was also found to be significantly less than 1.0 on a relative seed yield
basis in 1987 (data not shown). This likely resulted from one-sided antagonism in
favor of velvetleaf since soybean yields in mixtures were much less than those in
monoculture while velvetleaf yielded significantly more seeds per plant in mixture
than in monoculture (Figure 3).
Relative crowdini?r coefficient (ROCl
DeWit replacement series may also be analyzed mathematically by means of
RGC values which serve as indices for comparing the relative aggressiveness of one
species towards another (deWit, 1960). In 1988, the RGC of velvetleaf was more
than 20 times greater than that of soybean at 59 DAP (Table l). Therefore,
although velvetleaf was infected with verticillium wilt beginning in the middle
part of the 1986 growing season, it was still many times more conq}etitive than
soybean.
In 1887, the RCC of soybean at 25 DAP was slightly, but not significantly,
greater than that of velvetleaf (Table l). During the remainder of the growing
season, the ROC of velvetleaf progressively increased from 1.6 to 9 times greater
than that of soybean. These results suggest that in the absence of disease,
soybean may be as conq>etitive as velvetleaf in acquiring Hmifing resources early
in the growing season.

33
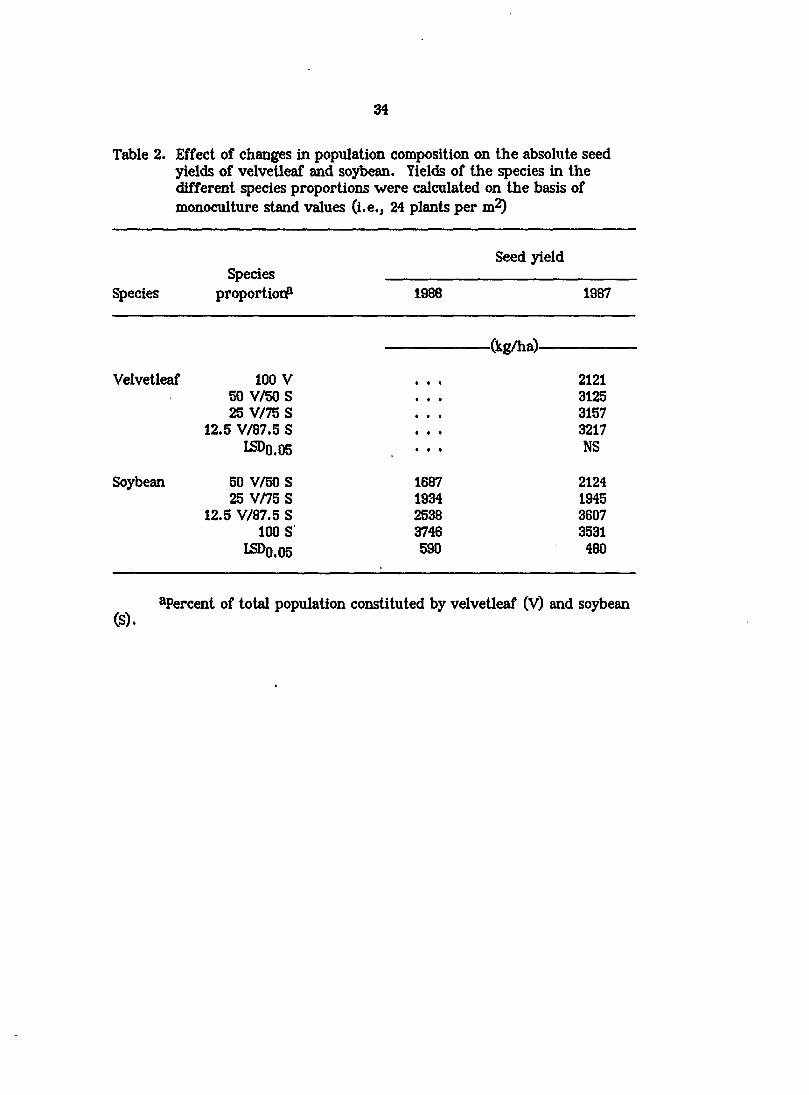
Seed yields of velvetleaf and soybean
On an absolute basisj soybean seed yields in 1986 were reduced by 32, 48,
and 55% as the proportion of velvetleaf in the population was increased from 0 to
12.5, 25, and 50% (Table 2). Similar changes in species composition in 1987
reduced soybean yields by 33, 64, and 81%. Therefore, full season interference
from even a small number of velvetleaf can have a major impact on soybean seed
yields. In contrast, velvetleaf seed yields were lowest in monoculture and were
increased by 47, 49, and 52% as the proportion of soybeans in the mixtures was
raised to 50, 75, and 87.5% (Table 2). The presence of verticillium wilt disease in
velvetleaf resulted in seed yields of soybean that were higher in 1986 than in 1987
(Table 2). In the absence of verticillium wilt, velvetleaf plants had more leaf
area to capture light and to shade soybean plants (due to less leaf drop). This
contributed to the greater reductions in soybean seed yields that were observed in
1987 when the proportion of velvetleaf in the population was increased to 25 or
50%.
From the results of the growth and yield interference studies presented
here, it appears that, although velvetleaf and soybean make demands on the same
resources early in the growing season, thqr do not significantly interfere with
each other's vegetative growth. However, during the middle and latter parts of
the growing season when canopy closure has occurred, velvetleaf gains resources
at the expense of soybean and diminishes soybean vegetative growth. This
ultimately results in a substantial loss of marketable yield.
34
Table 2. Effect of changes in population composition on the absolute seed yields of velvetleaf and soybean. Yields of the species in the different species proportions were calculated on the basis of monoculture stand values (i.e., 24 plants per m^)
Species Species
proportion?
Seed yield
1988 1987
-(kg/ha)-
Velvetleaf
Soybean
100 V 2121 50 V/50S 3125 25 V/75 S 3157
12.5 V/87.5 S • . « 3217 LSDO.05 • • •
NS
50 V/50 S 1687 2124 25 V/75 S 1934 1945
12.5 V/87.5 S 2538 3607 100 S 3746 3531
I5DO.05 590 480
^Percent of total population constituted by velvetleaf (V) and soybean (S).

35
Competition for nutrients and water
Soil tests conducted on the experimental plots in 1986 and 1887 (data not
shown) indicated sufficient nutrients were available for normal crop growth.
Moreover^ soybean plants are arable of fixing their own nitrogen. Therefore^ it
is likely that competition for nutrients did not contribute significantly to
interference between velvetleaf and soybean in the studies reported here. Dekker
and Meggitt (1983a) similarly concluded that competition for nutrients was not
involved in velvetleaf interference with soybean, based on analysis of leaf tissue
for macro- and micro-nutrients.
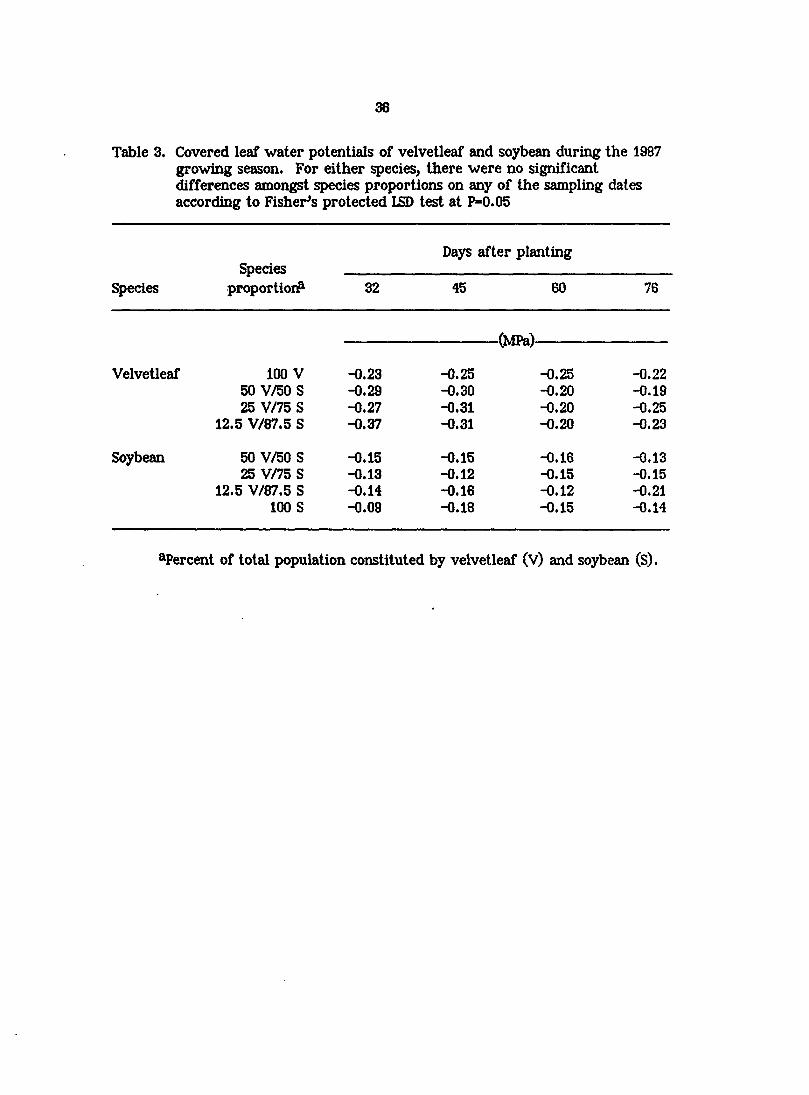
Changing the proportion of species in the population did not significantly
affect covered leaf water potentials of either species on any sampling date in 1987
(Table 3). Leaf water potentials of velvetleaf î^peared to be consistently lower
than those of soybean, but neither species exhibited leaf water potentials
indicative of water stress. Cheetham (1989) similarly reported velvetleaf pre
dawn leaf water potentials that were consistently lower than those of soybean
under field conditions. Although leaf water potential measurements were not
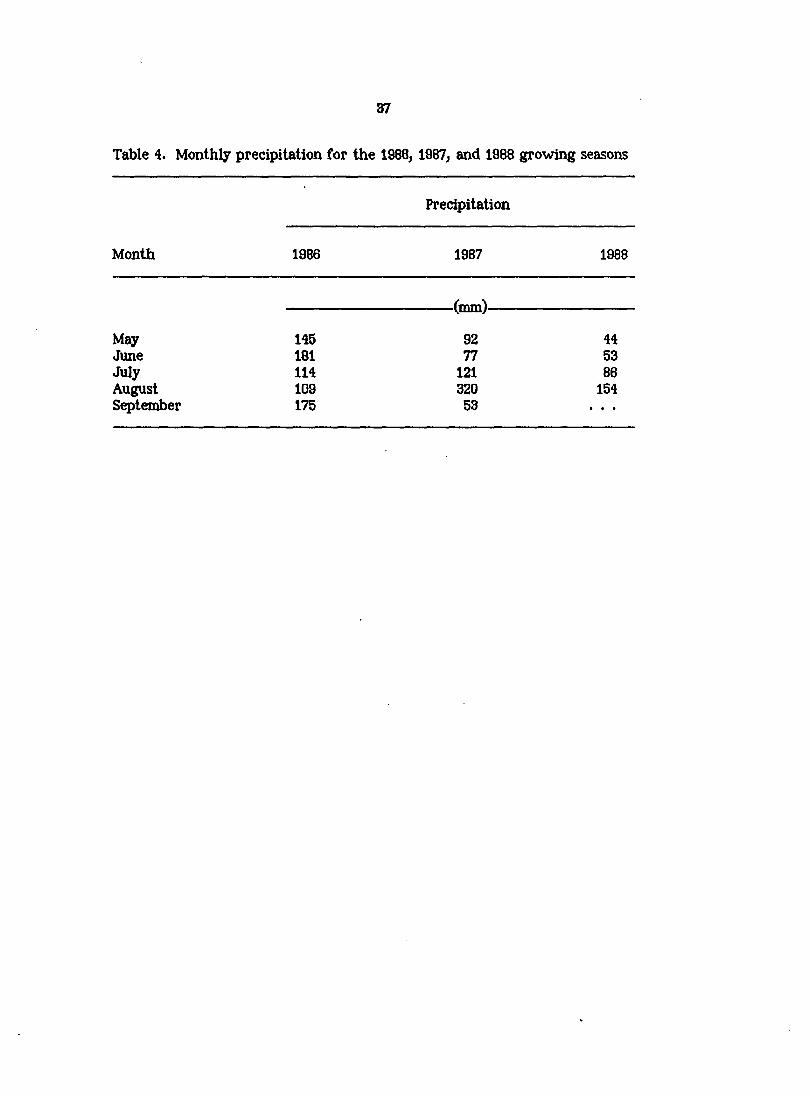
taken in 1986, total monthly precipitation values for 1986 were similar to or
greater than those recorded in 1987 (Table 4) and, therefore, it is likely that soil
water availability in 1986 was adequate to avert water stress in either species.
During years of normal precipitation, velvetleaf and soybean do not t^pear to
con^pete for water (Cheetham, 1989; Munger et al., 1987a).
38
Table 3. Covered leaf water potentials of velvetleaf and soybean during the 1987 growing season. For either species, there were no significant differences amongst species proportions on any of the sampling dates according to Fisher^s protected LSD test at P=0.05
Days after planting Species
Species proportion? 32 45 60 76
(MPa)
Velvetleaf 100 V -0.23 -0.25 -0.25 -0.22 50 V/50 S -0.29 -0.30 -0.20 -0.19 25 V/75 S -0.27 -0.31 -0.20 -0.25
12.5 V/87.5 S -0.37 -0.31 -0.20 -0.23
Soybean 50 V/50 S -0.15 -0.15 -0.16 -0.13 25 V/75 S -0.13 -0.12 -0.15 -0.15
12.5 V/87.5 S -0.14 -0.16 -0.12 -0.21 100 S -0.09 -0.18 -0.15 -0.14
^Percent of total population constituted by velvetleaf (V) and soybean (S).
37
Table 4. Monthly precipitation for the 1988, 1987, and 1988 growing seasons
Precipitation
Month 1986 1987 1988
(mm)
May 145 92 44 June 181 77 53 July 114 121 88 August 109 320 154 Sq)tember 175 53

38
Competition for Light: Whole Plant
Height
Averaged over all species proportions, velvetleaf plants were taller than
soybean plants on all sampling dates both in 1986 and in 1987 (Table 5),
Velvetleaf was significantly taller in mixture than in monoculture only early in
the growing season in 1986 (44 DAP) and in 1987 (25 DAP). In contrast, soybean
plants were significantly shorter in monoculture. Increasing the proportion of
velvetleaf in the mixtures to 25 or 50% resulted in a significant increase in the
height of soybean during the early and middle parts of the 1986 growing season
(Table 5). Similar results were evident in 1987, but the increases in height of
soybeans were not statistically significant. As has been previously reported
(Salisbury, 1985), increased shading by dominant velvetleaf plants may have
resulted in greater than normal intemode elongation in soybean.
Any factor that differentially alters a plantas ability to intercept light in
relation to that of neighboring plants should influence its ability to compete for
light (Rhodes and Stem, 1978). With its consistent height advantage over most of
the growing season as reported here and by other researchers (Stoller and
Woolley, 1985), it is not surprising that velvetleaf gains resources (especially
light) at the expense of soybean when these species co-occur.
Uaf area
Averaged over all species proportions, leaf area per plant was greater for
velvetleaf than for soybean in 1986 until later in the growing season (75 DAP)
when infection with verticiUium wilt resulted In extensive leaf drop in
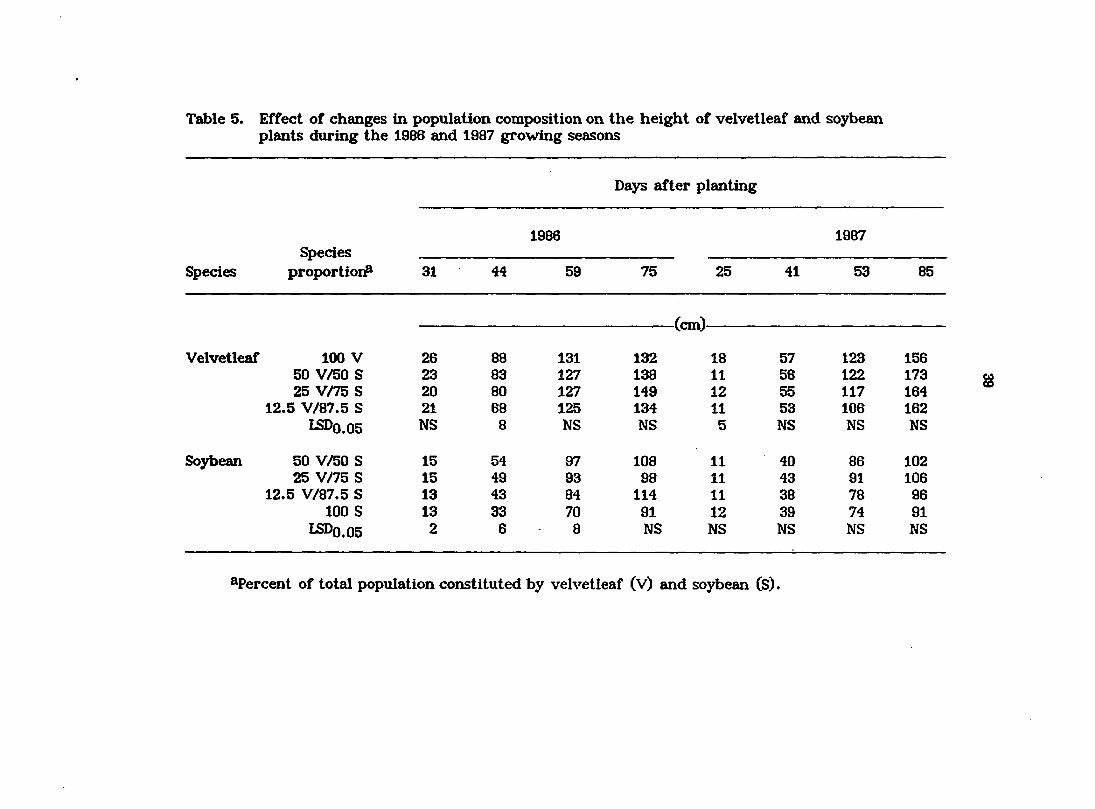
Table 5. Effect of changes in population composition on the height of velvetleaf and soybean plants during the 1986 and 1987 growing seasons
Days after planting
1986 1987 Species
Species proportioi# 31 44 59 75 25 41 53 85
Velvetleaf
Soybean
(cm)-
100 V 26 88 131 132 18 57 123 156 50 V/50 S 23 83 127 138 11 56 122 173 25 V/75 S 20 80 127 149 12 55 117 164
12.5 V/87.5 S 21 68 125 134 11 53 106 162 LSDO.05 NS 8 NS NS 5 NS NS NS
50 V/50 S 15 54 97 108 11 40 86 102 25 V/75 S 15 49 93 98 11 43 91 106
12.5 V/87.5 S 13 43 84 114 11 38 78 96 100 S 13 33 70 91 12 39 74 91
LSDO.05 2 6 8 NS NS NS NS NS
^Percent of total population constituted by velvetleaf (V) and soybean (S).

40
velvetleaf (Table 6). By comparison^ in 1987, when neither species was affected
by disease, soybean leaf area averaged over all species proportions was greater
than that of velvetleaf until later in the growing season (53 and 85 DAP), when
velvetleaf plants displayed greater leaf areas.
In 1986, the leaf area of velvetleaf plants generally decreased in mixtures as
the proportion of velvetleaf in the population was increased (Table 6). A
consistent pattern of change in leaf area of soybean was not observed except on
the final san^ling date when the leaf area of soybean in mixtures was similarly
depressed as the proportion of velvetleaf in the population was increased. In
contrast to the results of the 1986 stu<fy, in 1987, the change in composition of the
species mixtures did not significantly affect the leaf area of velvetleaf or soybean
until late in the season (85 DAP) when the leaf area of soybean was greater when
constituting a larger (lOO or 87.5) rather than a smaller (50) percentage of the
population. In the absence of disease, the greater leaf area of soybean early in
the growing season should have improved its competitive ability relative to that
of velvetleaf by enhancing its ability to capture light. However, by mid- to late-
season the greater leaf area and greater height of velvetleaf would tend to
diminish the impact of any early season gains made by soybean.
Rranff>^ miTpher and distribution
Velvetleaf had many times more mainstem branches than soybean in both
1988 and 1987 (Table 7). While differences amongst species proportions in the
number of mainstem branches per plant in soybean were not significant, there did
appear to be some suppression of branching in soybean in 1986 as the proportion of
velvetleaf in the mixture was increased. However, in 1987, the number of
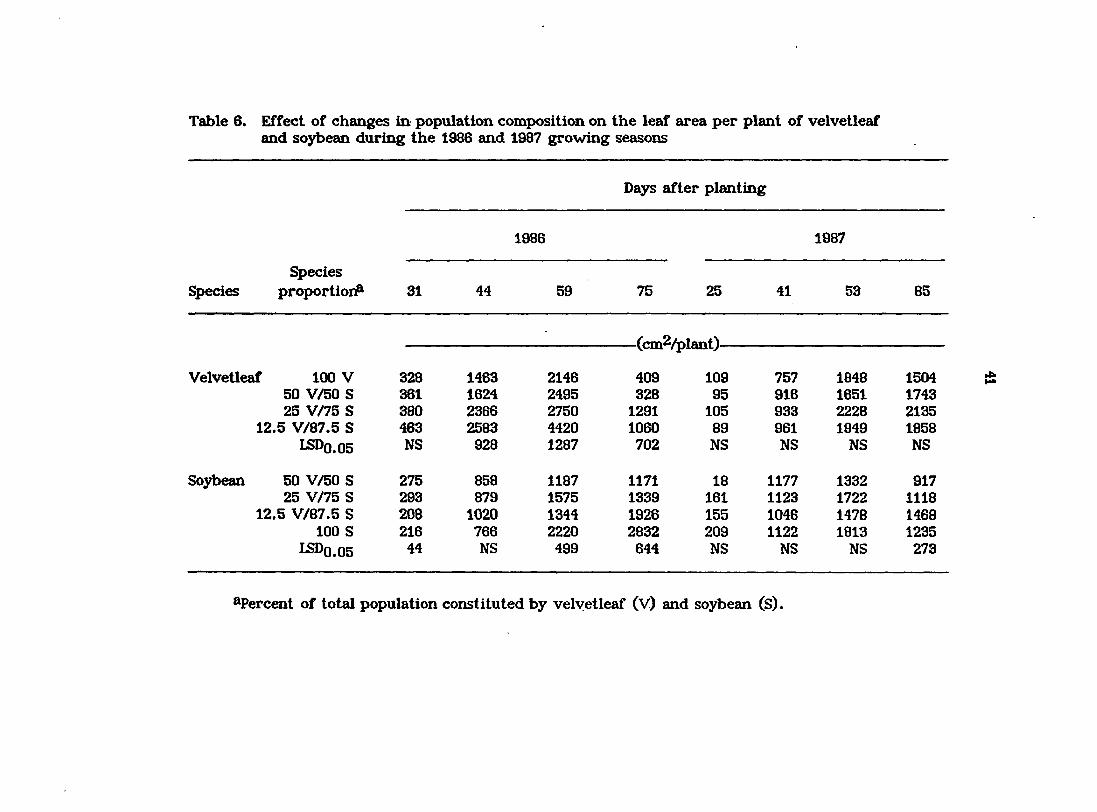
Table 6. Effect of changes in population composition on the leaf area per plant of velvetleaf and soybean during the 1986 and 1987 growing seasons
Days after planting
1986 1987
Species Species proportion? 31 44 59 75 25 41 53 85
(cm2/plant)
Velvetleaf 100 V 328 1463 2146 409 109 757 1848 1504 50 V/50 S 361 1624 2495 328 95 916 1651 1743 25 V/75 S 380 2366 2750 1291 105 933 2228 2135
12.5 V/87.5 S 463 2583 4420 1060 89 961 1949 1858 LSDo.05 NS 928 1287 702 NS NS NS NS
Soyt>ean 50 V/50 S 275 858 1187 1171 18 1177 1332 917 25 V/75 S 293 879 1575 1339 161 1123 1722 1118
12.5 V/87.5 S 208 1020 1344 1926 155 1048 1478 1468 100 S 216 766 2220 2832 209 1122 1813 1235
LSDO.05 44 NS 499 644 NS NS NS 273
^Percent of total population constituted by velvetleaf (V) and soybean (S).
42
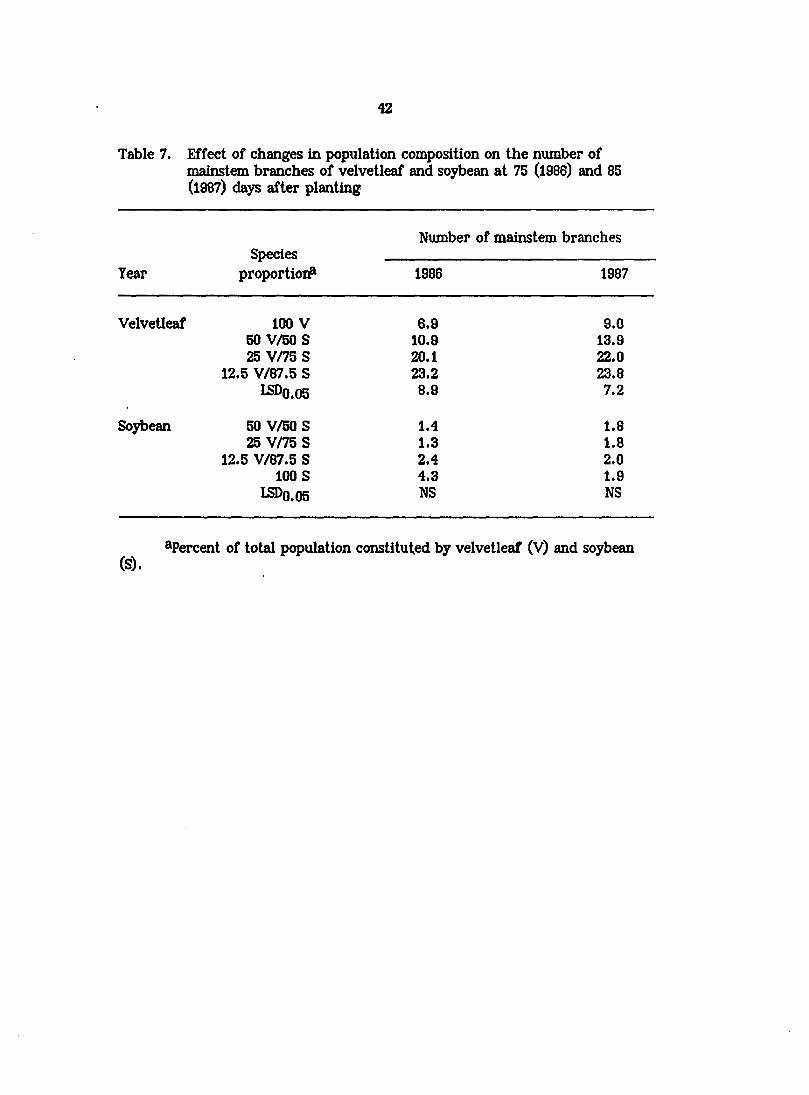
Table 7. Effect of changes in population composition on the number of mainstem branches of velvetleaf and soybean at 75 (1986) and 85 (1987) days after planting
Year Species
proportioi#
Number of mainstem branches
Year Species
proportioi# 1986 1987
Velvetleaf 100 V 6.9 9.0 50 V/50S 10.9 13.9 25 V/75 S 20.1 22.0
12.5 V/87.5 S 23.2 23.8 LSDO.05 8.9 7.2
Soybean 50 V/50 S 1.4 1.8 25 V/75 S 1.3 1.8
12.5 V/87.5 S 2.4 2.0 100 S 4.3 1.9
I5DO.05 NS NS
^Percent of total population constituted by velvetleaf (V) and soybean (S).

43
mainstem branches per plant was uniformly low in soybean in all species mixtures
(Table 7). Because velvetleaf was taller than soybean throughout much of the
1986 and 1987 growing seasons, soybean plants growing beneath or within their
canopies would have been exposed to a light environment poor in
photosynthetically active radiation (PAR) but enriched in far-red wavelengths
(child et al., 1981). Recent studies have shown that far-red light diminishes the
production of fruiting branches in soybean (Begonia and Aldrich, 1988), and light
interference studies with velvetleaf canopies have shown reduced branching in
velvetleaf-shaded soybean plants under field conditions (Salisbury, 1985).
Velvetleaf experienced reductions in branching of 70% (1988) and 63% (1987)
as its proportion in the population was increased from 12.5 to 100% (Table 7).
Reduced branching in velvetleaf may similarly have been due to changes in quality
and quantity of light as its proportion in the population and, hence, the degree of
intraspecific shading was increased.
The vertical distribution of branches in velvetleaf and soybean was
Investigated in 1987. Branching in velvetleaf tended to be confined to the upper
half of the plantj the largest number of branches occurred in the top one-third of
the plant and no branches were produced below 75 cm, regardless of the
proportion of velvetleaf in the population (Table 8). Additionally, lower leaves of
velvetleaf senesce as the soybean canopy (or velvetleaf canopy in monoculture)
closes, so that by the end of the growing season velvetleaf plants retain leaves
only at or above the top of the soybean canopy (Regnier, 1887). Branching in
soybean, unlike that in velvetleaf, was restricted to lower axillary buds (O to 30
cm above the soil surface). Soybean branch distribution i^peared to be largely
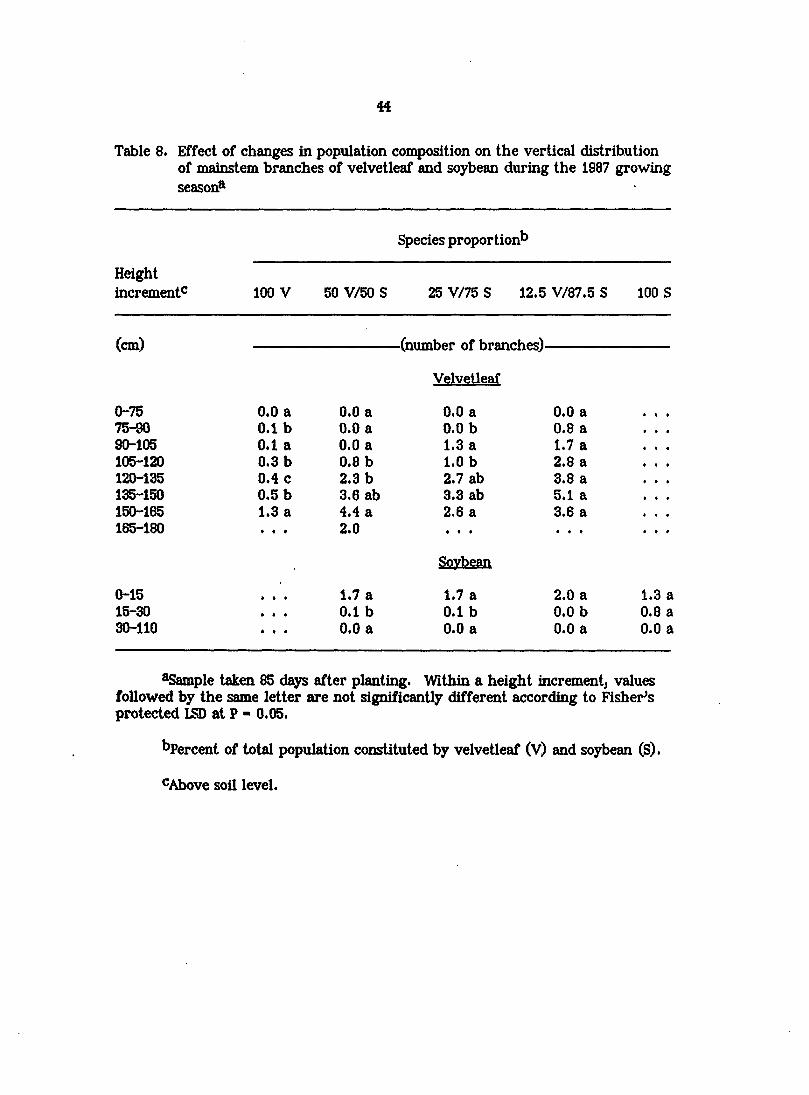
44
Table 8. Effect of changes in population composition on the vertical distribution of mainstem branches of velvetleaf and soybean during the 1987 growing season®
Species proportion^
Height increment^ 100 V 50 V/50 S 25 V/75 S 12.5 V/87.5 S 100 S
(cm) -(number of branches)-
Velvetleaf
0-75 0.0 a 0.0 a 0.0 a 0.0 a 75-90 0.1 b 0.0 a 0.0 b 0.8 a 90-105 0.1 a 0.0 a 1.3 a 1.7 a 105-120 0.3 b 0.8 b 1.0 b 2.8 a 120-135 0.4 c 2.3 b 2.7 ab 3.8 a 135-150 0,5 b 3.6 ab 3.3 ab 5.1 a 150-165 1.3 a 4.4 a 2.6 a 3.6 a 165-180 2.0
Soybean
0-15 1.7 a 1.7 a 2.0 a 1.3 a 15-30 0.1 b 0.1 b 0.0 b 0.8 a 30-110 0.0 a 0.0 a 0.0 a 0.0 a
^Sample taken 85 days after planting. Within a height increment^ values followed by the same letter are not significantly different according to Fisher's protected LSD at P - 0.05.
bpercent of total population constituted by velvetleaf (V) and soybean (S).
CAbove soil level.

45
independent of the number of velvetleaf plants present. Assuming mainstem
branch distribution provides a reasonable approximation of leaf area distribution^
the canopy architecture of velvetleaf would be well suited to shading shorter
understory species such as soybean while maintaining a high capacity to intercept
light.
Growth analysis
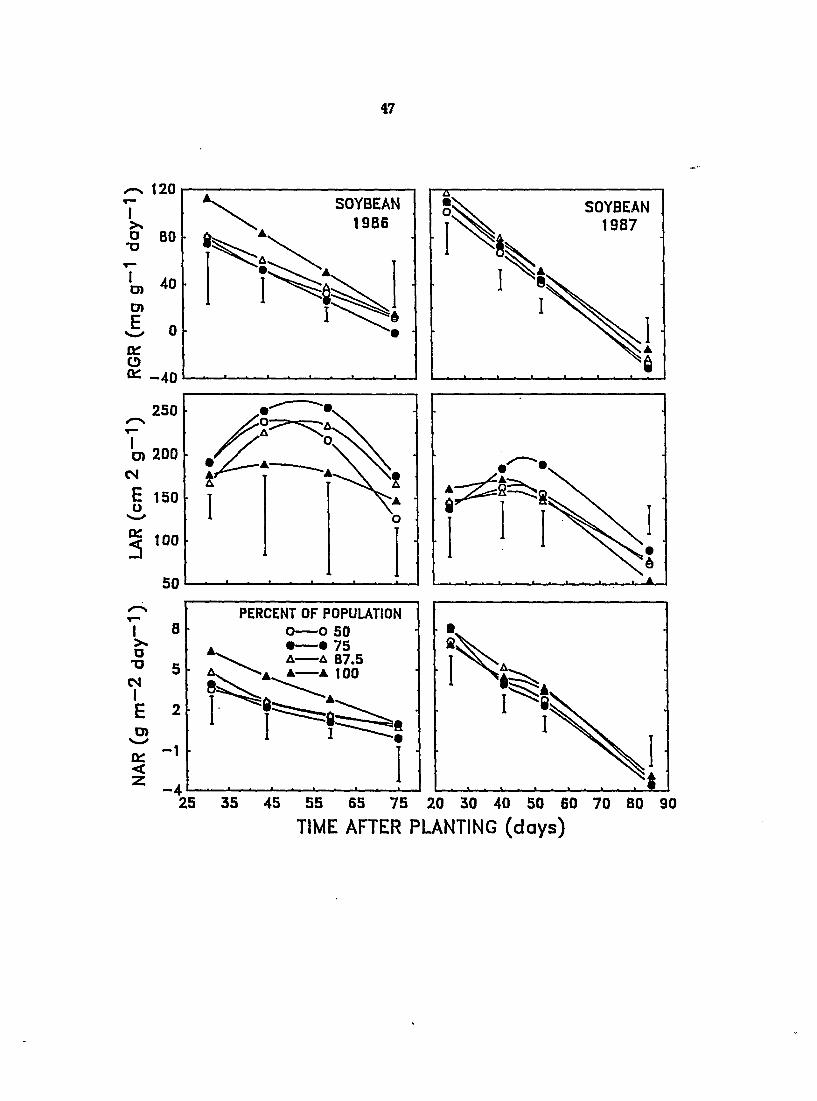
There were no significant differences amongst mixtures in RGRj lARj or
NAR of soybean in 1988 (Figure 4). RGR and NAR of soybean appeared to be fairly
uniformly depressed by all levels of velvetleaf in 1986, although values for these
parameters in mixtures were not always significantly different from those in
monoculture. As the proportion of velvetleaf in the mixtures and, hence, the
degree of shading increased, the LAR of soybean also appeared to increase,
although the results were highly variable and the increases were not statistically
significant. In response to artificial shading under field conditions, LAR in
soybean may be increased by as much as 48%, primarily due to decreased leaf
thickness (Regnier et al., 1988). Differences in RGR, NAR, and LAR between
soybeans grown in mixture with velvetleaf and those grown in monoculture were
not as distinct or consistent in 1987 as in 1988 (Figure 4). Overall, interspecific
competition seemed to have less effect on growth analysis parameters of soybean
during early to mid-season in 1987, perh^s due to a greater competitive ability of
soybean not affected by disease.
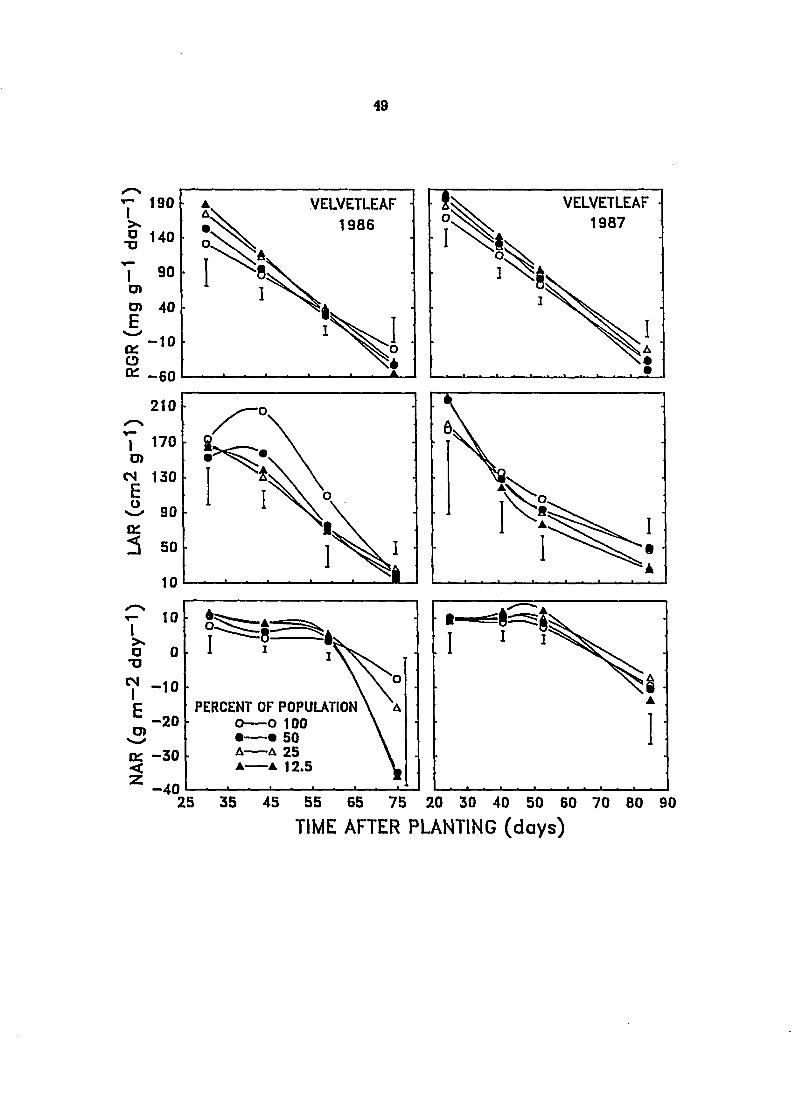
Changes over the growing season in velvetleaf RGR, NAR, and to a lesser
extent lAR were similar in 1986 and 1987 (Figure 5). Generally, velvetleaf grew
better in mixture than in monoculture both years, especially in the early to

Figure 4. Effect of changes in population composition on the relative growth rate (RG^j leaf area ratio (LAlOi and net assimilation rate (NAR) of soybean in monoculture and in mixtures with velvetleaf during the 1986 and 1987 growing seasons. Vertical bars represent LSD at P = 0.05
47
SOYBEAN 1986
PERCENT OF POPULATION O O 50
-• 75 -A 87.5 -A 100
SOYBEAN 1987
«
,
• 1
25 35 45 55 65 75 20 30 40 50 60 70 80 90
TIME AFTER PLANTING (days)

Figure 5. Effect of changes in population composition on the relative growth rate (RGK), leaf area ratio (LAS)^ and net assimilation rate (NAR) of velvetleaf grown in monoculture and in mixtures with soybean during the 1986 and 1887 growing seasons. Vertical bars represent LSD at P - 0.05
49
VELVETLEAF • 1987
190 VELVETLEAF • 1986
40
90
40
210
D)
130
•D
-10
PERCENT OF POPULATION o o 100
• 50 A A 25 A • 12.5
s*./
a: -30
—40 25 35 45 55 65 75 20 60 70 80 90
TIME AFTER PLANTING (days)

50
middle part of the growing season, exhibiting improved RGR as its proportion in
the mixtures was decreased. The lAR of velvetleaf may be increased by up to 50%
when velvetleaf is grown under artificial shade in the field, due primarily to
decreased leaf thickness (Regnier et al., 1988). In the present study, LAR of
velvetleaf was increased only in 1986 between the first and second sampling dates
and only in monoculture and in the mixture containing 50% velvetleaf (Figure 5).
During the remainder of the growing season in those species proportions and over
the entire sampling period in all other mixtures, the LAR of velvetleaf decreased.
In 1987, the LAR of velvetleaf similarly decreased over the course of the
experiment in monoculture and in all mixtures. Similar results were found by
Cheetham (1989). LAR of velvetleaf intercropped with soybean decreased 90%
during the growing season. This was partly due to a progressive decrease in
specific leaf area, i.e., an increase in leaf weight per unit leaf area (Cheetham,
1989). Decreasing LAR in velvetleaf as the growing season progresses may arise
partly from the way its canopy architecture develops. During and after canopy
closure, much of the leaf area in the lower part of the plant is lost due to
senescence and leaf drop. Since branching only occurs near the top of velvetleaf
plants intercropped with soybean, most of the leaf area becomes concentrated in
the upper portion of the plant above the soybean canopy where irradiance is
higher. Leaves of velvetleaf produced in the sun are thicker than those produced
under shaded conditions (Tun and Taylor, 1986). Therefore, as more leaves are
produced in the sun, there would be a progressive decrease in lAR of velvetleaf.
Decreasing lAR over the growing season in velvetleaf may also have been partly
due to a gradual increase in the proportion of total plant weight constituted by
stem growth.

51
Velvetleaf exhibited higher NAR than soybean in mixtures and monoculture
during early to mid-season in 1986 and 1987 (Figures 4 and 5). The greater
carbon assimilation capacity of velvetleaf reflected in the higher NAR values helps
to explain the observed higher RGR of velvetleaf compared to soybean during the
same period. Velvetleaf growth in mixtures may have been enhanced by the
presence of soybean, but the greater LAR of velvetleaf in monoculture rather than
in mixture (Figure 5) suggests that the observed reductions in RGR and NAR of
velvetleaf in monoculture compared to mixtures were due to increased
intraspecific shading and, hence, increased competition for light.
Competition for Light: Canopy
Leaf area indes
At the level of the canopy, LAI is a useful tool for examining the
relationship between plant productivity and light availability (Radosevich and
Holt, 1984) since it represents a broad index of the productive capacity of a
canopy (Hunt, 1978). LAI increased rapidly in all species proportions from early to
midseason in 1988 and in 1987 (Figure 6). Thereafter, lAI decreased in all species
proportions except in monoculture soybean in 1988, where it continued to increase
over the remainder of the growing season.
Incorporation of velvetleaf plants into the canopy apparently compensated
for leaf area loss in soybean due to disease so that the LAI of mixtures was greater
than that of monoculture soybean early in 1988 (Figure 8). After 55 DAP, the LAI

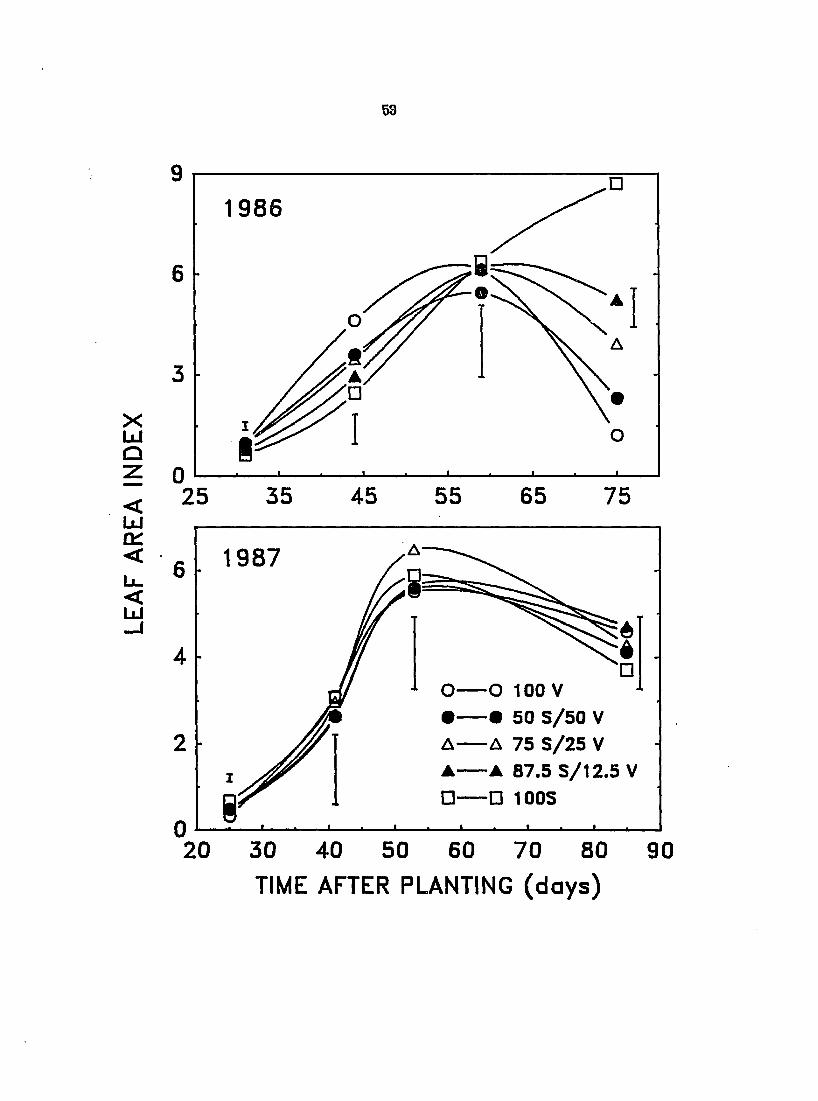
Figure 6. Effect of interference on the leaf area index of velvetleaf (V) and soybean (S) in monoculture and mixtures of different proportions during the 1986 and 1987 growing seasons. Vertical bars represent LSD at P - 0.05
53
X LU O
< LU QC <
Lu < LU
1986
1987
•o 100 V
• 50 S/50 V •A 75 S/25 V •A 87.5 S/12.5 V
-• 100S
20 30 40 50 60 70 80 90
TIME AFTER PLANTING (days)

54
of all species proportions containing velvetleaf r^idly decreased due to
verticillium induced leaf drop in velvetleaf. Differences in LAI among
monocultures and mixtures in the absence of leaf area-depleting diseases in 1987
were generally small and nonsignificant, especially early in thé growing season
(Figure 6). Because the LAI of the canopies did not change significantly with
changes in population composition in 1987, this suggests that, in terms of LAI,
velvetleaf and soybean were approximately equivalent and were freely
interchangeable without much effect on a canopy^s LAI and perhaps on its
productivity.
Lifyht conversion analysis
There were no discernable differences in total seasonal I© between growing
seasons (data not shown).
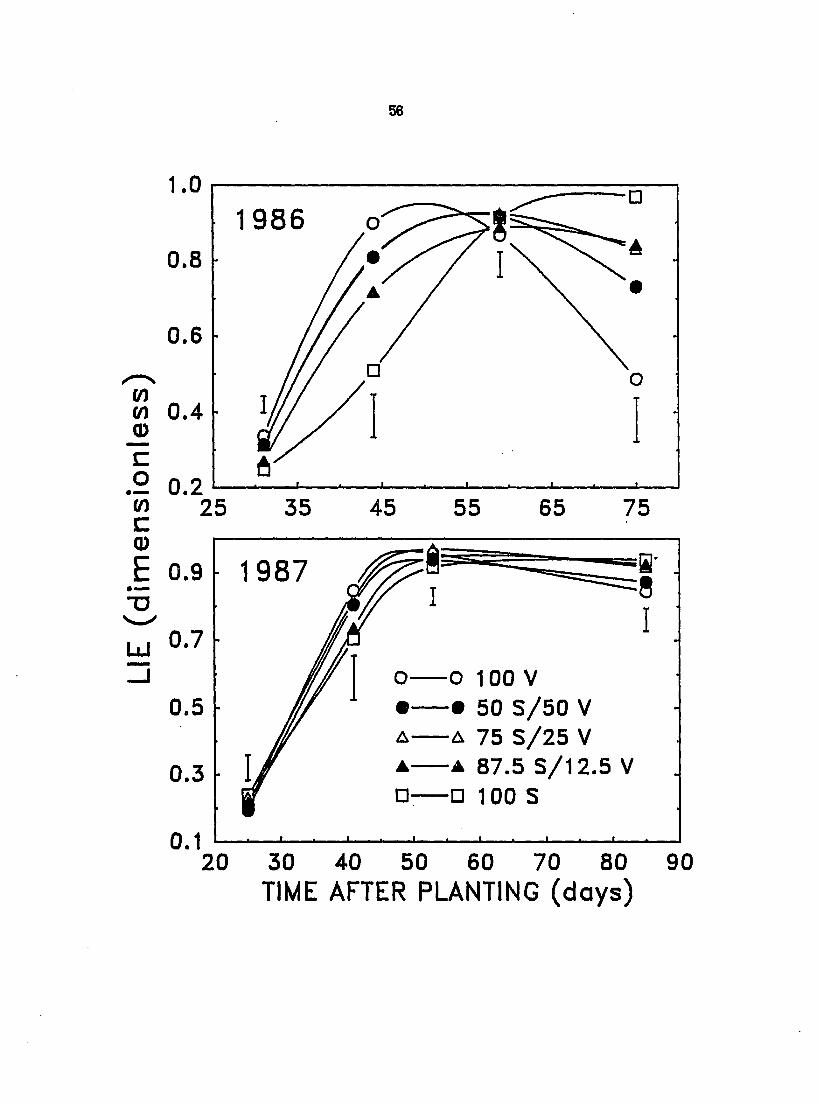
Changes in LIE (Figure 7), which is a measure of a canopy^s ability to
intercept PAR, correlated fairly well with observed changes in LAI over the
growing season both in 1988 and 1987 (cf. Figure 8). This close correlation
emphasizes the io^wrtance of LAI to ability to compete for light. Due to early
season leaf area loss in soybean in 1988 (31 and 44 DAP), the LIE of velvetleaf in
monoculture exceeded that of monoculture soybeans by as much as 35 to 77%.
However, with premature leaf drop in velvetleaf infected with verticillium wilt,
by 75 DAP the LIE of monoculture velvetleaf was 50% less than that of soybean in
monoculture (Figure 7). In 1987, LIE increased rapidly in all species proportions to
a maximum over 90% by midseason (53 DAP). Although velvetleaf in monoculture
speared to have a greater LIE than monoculture soybean over much of the 1987
growing season, the differences were not significant until late in the growing

Figure 7. Effect of interference on the light interception efficiencies (LIE) of canopies of velvetleaf (V) and soybean (S) in monoculture and mixtures (V+S) of varying proportions during the 1986 growing season. Vertical bars rq>resent LSD at P - 0.05
56
986
M 0.4
a
o 100 V 50 S/50 V
A A 75 S/25 V A A 87.5 S/12.5 V D D 1 00 S
20 30 40 50 60 70 80 90 TIME AFTER PLANTING (days)

57
season when normal leaf abscission in velvetleaf significantly reduced its LIE
below that of soybean (Figure 7). However, since the canopy of velvetleaf is
taller than that of soybean for much of the growing season, in mixtures even a
slightly greater LIE in velvetleaf would substantially reduce light availability for
soybean. Replacement of soybean with velvetleaf significantly improved canopy
LIE of mixtures compared to monoculture soybean before canopy closure in 1886
(Figure 7). A similar trend was evident in 1987 although the differences in LIE
were not significant (Figure 7).
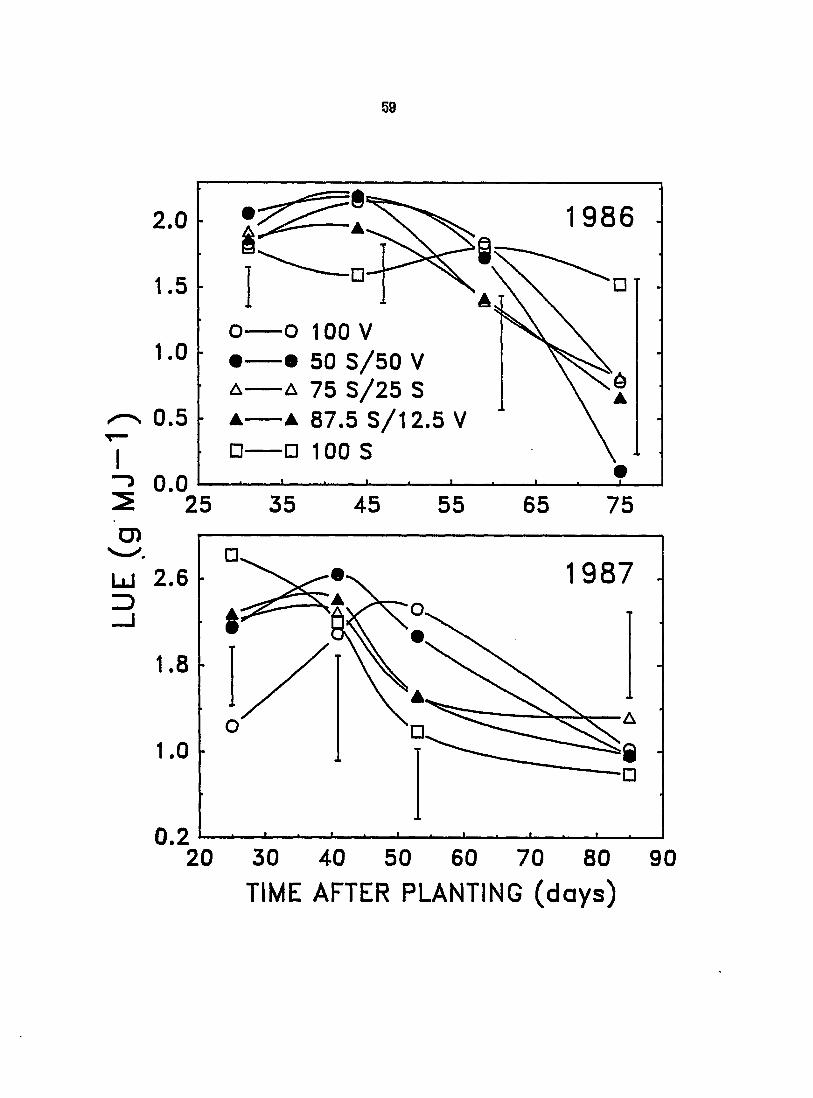
The LUE of velvetleaf in pure stand did not significantly exceed that of
soybean grown alone in 1986 except at 44 DAP (Figure 8). Verticillium wilt
speared to diminish the LUE of velvetleaf before leaf drop resulted in decreased
LIE (cf. Figure 7). The LUE of monoculture soybeans remained nearly constant in
1988 despite substantial early leaf area loss. Under more normal conditions in
1987, soybean displayed a high LUE early in the season (25 DAP) despite having a
somewhat lower NARj but, by 53 DAP, the LUE of velvetleaf was nearly twice that
of soybean (Figure 8). The somewhat greater light-capture efficiency of
velvetleaf during a portion of the early part of the growing season and a superior
LUE during the middle and latter parts of the growing season, compared to those
of soybean, combined to give velvetleaf a greater ability to con^ete for light.
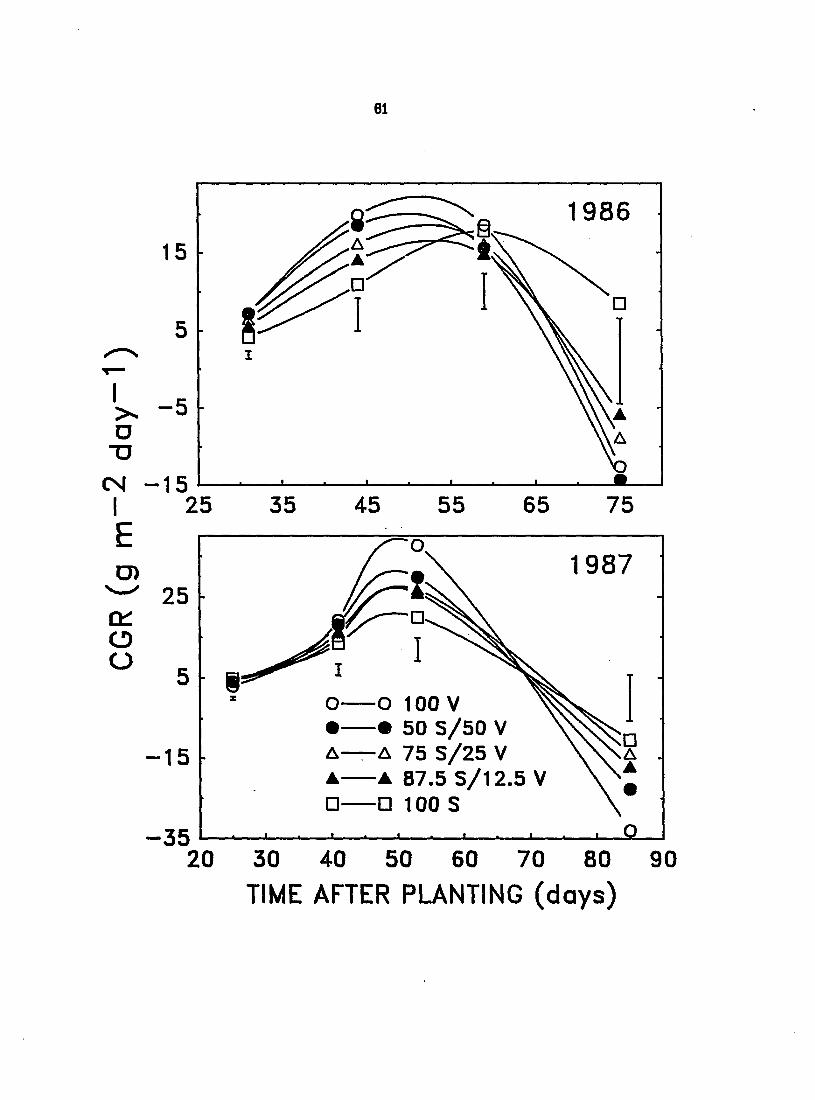
CGR, the rate of dry matter production per unit area of land (including
that of velvetleaf in mixtures), followed ^^proximately the same trends over the
growing season as UE, especially in 1986 (Figure 9). This emphasizes the
importance of efficient light cf^pture to overall growth and conq)etitive ability.
The CGR of monoculture velvetleaf was nearly twice that of monoculture soybean

Figure 8. Effect of interference on the light utilization efficiencies (LUE) of canopies of velvetleaf (V) and soybean (S) in monoculture and mixtures (V+S) of varying proportions during the 1987 growing season. Vertical bars represent LSD at P - 0.05
59
2.0
1.5
1.0
0.5
0.0
1986
o o 100 V e • 50 S/50 V A A 75 s/25 S A A 87.5 S/12.5 V
• • 100 S
25 35 45 55 65 75
LU 2.6
20 30 40 50 60 70 80 90
TIME AFTER PLANTING (days)

Figure 9. Effect of interference on the crop growth rate (CGR) of canopies of velvetleaf (V) and soybean (S) in monoculture and mixtures (V+S) of varying proportions during the 1987 growing season. Vertical bars represent LSD at P - 0.05
61
1986
CN -15 I £
vS
Q1
O
o
25 35 45 55 65 75
25
-15
-35
—O 100 V • e 50 S/50 V A — A 75 S/25 V A A 87.5 S/12.5 V • • 100 S
20 30 40 50 60 70 80
TIME AFTER PLANTING (days)
90

62
early in the 1986 growing season (31 and 44 DAP) and mixtures generally displayed
CGR values intermediate between those of pure stands of velvetleaf and soybean
(Figure 9). The CGR of the various species proportions was generally higher in
1987 than in 1986 in the absence of disease in either species. In 1987, the CGR of
velvetleaf in monoculture at 25 DAP was 57% less than that of soybean grown in
monoculture (Figure 9). This reflects the significantly greater LUE and slightly
greater LIE of soybean early in the 1987 growing season . However, the CGR of
velvetleaf rapidly increased to 42 to 89% greater than that of soybean at midseason
(41 and 53 DAP). This, in turn, reflects the superior ability of velvetleaf to
capture and convert light to dry matter. Although altering the composition of
mixtures did not significantly alter their CGR, each increase in the proportion of
velvetleaf in the mixtures tended to boost their CGR over most of the growing
season relative to that of monoculture soybean (Figure 9).
Diurnal pattern of canopy light interception
At midseason in 1987, both velvetleaf and soybean in monocultures
intercepted 90% or more of incident PPFD by the time the solar beam had
penetrated 40 to 60% of the way (based on height of the canopy) through their
respective canopies (Figure 10). While ftiUy developed canopies of both species
were efficient at intercepting nearly all available PAR, differences were apparent
in the vertical distribution of PAR interception during the day. A larger portion
of total PAR c^tured by velvetleaf was intercepted near the top of the canopy
profile CFigure 10), partly because leaf area is concentrated in the upper portion
of the canopy in velvetleaf, especially in monoculture. In soybean, leaf area is

Figure 10. Diurnal pattern of light interception by monoculture velvetleaf (VEL) and soybean (SOY) canopies. Hour designations are based on local solar time. Curves on graphs are predicted values generated from the following regression equations where Y = percent photo^ynthetic photon flux density (% PPFD) intercepted and X = percent of total canopy height penetrated by the incident solar beam (O «= top of canopy; 100 = base of canopy). 800 h (VEI, Y = 8.16 + 0.58 - 0.005x2 + O.OOOOZx^j SOYj Y = 3.17 + 2.75x - 0.04x2 + 0.0002x3). 1100 h (VEL, Y = 6.88 + 1.25x - 0.02x2 + 0.00008x3; SOYj Y = 3.39 + 2.38x -0.03x2 + 0.0001x3). 1400 h (VEL, Y = 5.29 + 1.88x - 0.03x2 + 0.0001x3; SOY, Y = 4.35 + 1 .25x - 0.008x2 + 0.000007x3). 1700 h (VEL, Y = 8.29 + 0.52x - 0.006x2 + 0.00003x3; SOY, Y - 3.73 + 2.28X - 0.03x2 + 0.0001x3)
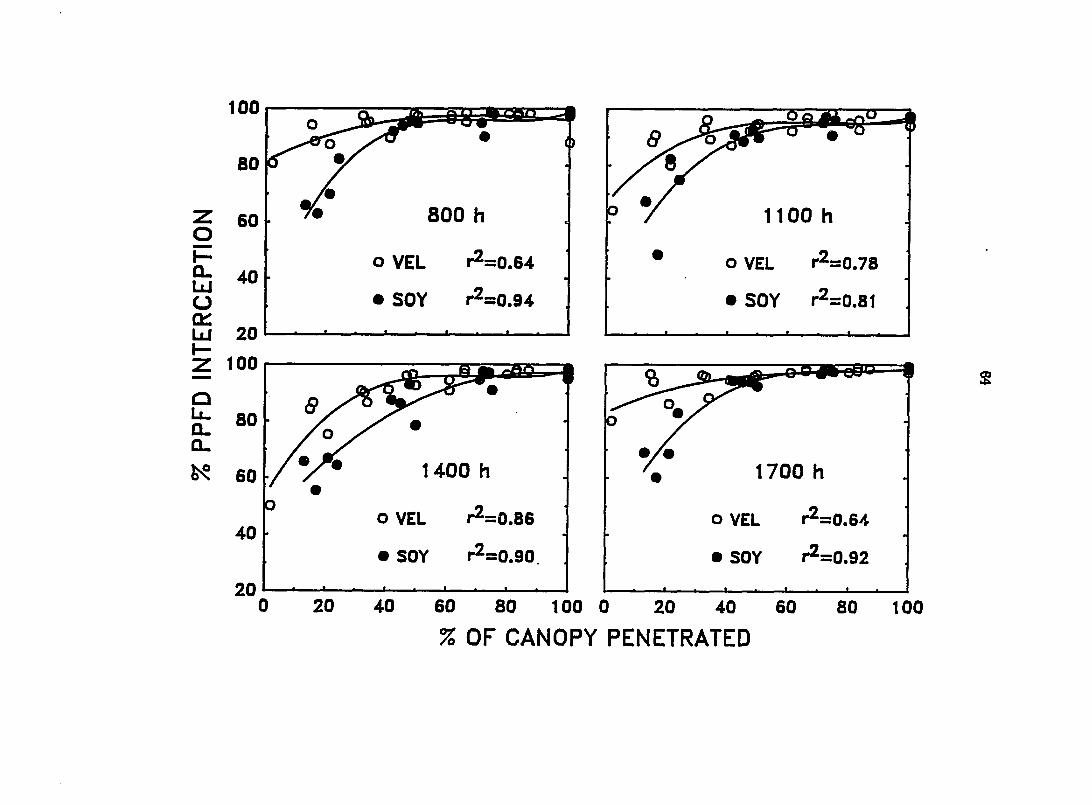
100
80
60 800 h
o VEL r2=o.64
• SOY r2=;0.94
1400 h
o VEL r2=0.86
SOY r^=0.90
1100 h
o VEL r2=o.78
100 0 20
oQO i
1700 h
o VEL r2=o.64
• SOY r2=0.92
40 60 80 100
2
% OF CANOPY PENETRATED

65
more evenly distributed throughout the canopy profile.
Light interception by soybean canopies did not change much during the
day, but velvetleaf appeared to intercept more of the available PAR at the top of
the canopy early (800 h) and late (1700 h) in the day. Since velvetleaf plants
were taller than soybean plants throughout most of the 1987 growing season^
these observed differences in diurnal light interception patterns may help to
explain the superior ability of velvetleaf to compete for light (i.e., daily LIE of
velvetleaf may be greater than indicated by the mid-day measurements shown in
Figure 7).
Solar-trackiny leaf movements and their significance to photosynthesis
With adequate water under field conditions, soybean leaves maintain a 30
degree angle of incidence between the leaf blade and the sun^s raysj this reduces
their PAR interception by 50% as compared to leaves with their blades
perpendicular to the sun's rays (Kawashima, 1969a and 1969b). Under conditions
of limited soil water availability, soybean leaves exhibit light-avoiding movements
(Meyer and Walker, 1981), as do the leaves of kidn^ bean seedlings (Berg and
Hsiao, 1986).
Differences in solar-tracking behavior between velvetleaf and soybean
(Figure 11) may also contribute to the previously noted differences in light
interception by canopies of these species, since enhanced light interception early
and late in the day is typical of solar-tracking species (Ehleringer and Forseth,
1980). Under well-watered conditions in the greenhouse, velvetleaf actively re
oriented its leaves to maintain a nearly constant angle between their lamina and
the incident solar beam, thus ensuring a fairly constant level of PAR on their

Figure 11. Diurnal course of solar-tracking movements of velvetleaf leaves and of the terminal leaflet of soybean leaves under greenhouse (July 23j 1986) and field conditions (August 3, 1988). A leaf blade maintained perpendicular to the sun's rays would have a sine of angle of incidence of 1.0 ^perfect tracking, while a leaf blade held parallel to the sun's rays would have a sine of angle of incidence of 0 (perfect avoidance). Asterisks signify significant differences in tracking between q)ecies at each sampling time according to an F-test at P- 0.05
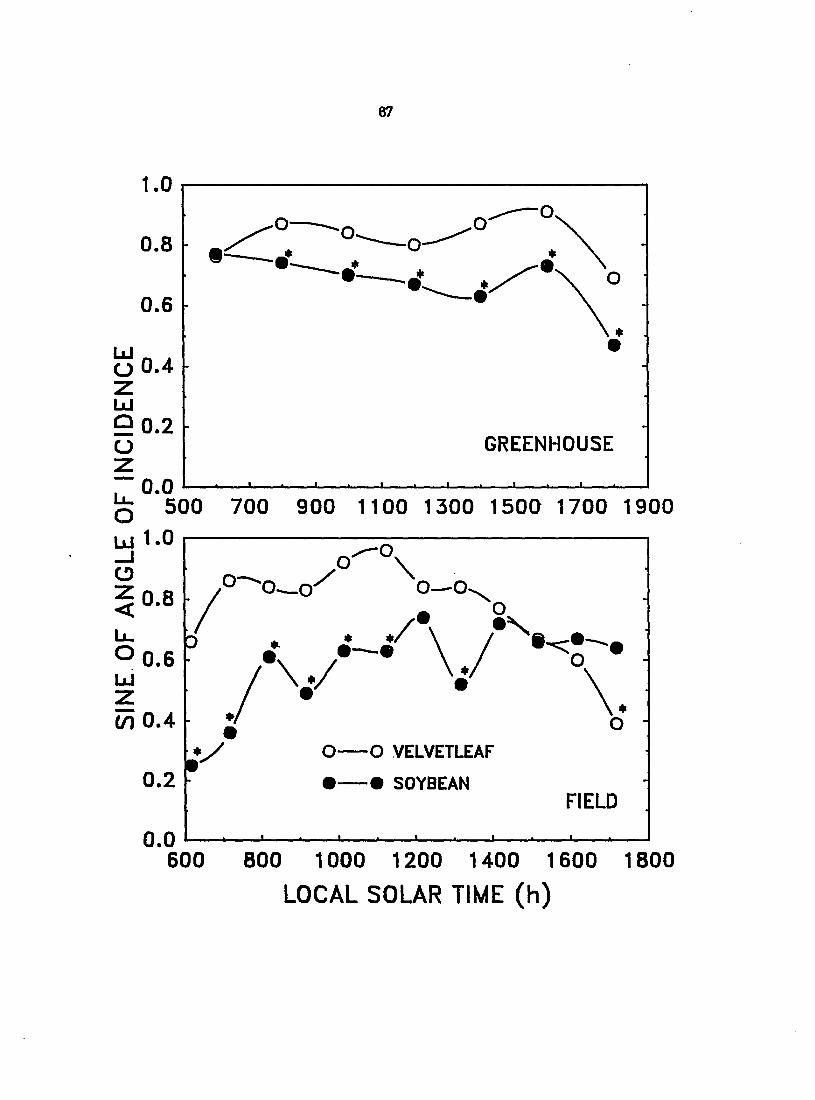
67
O 0.4
GREENHOUSE
^ 500 700 900 1100 1300 1500 1700 1900
LU
CD
1.0
$ 0 . 8
Lu
O 0.6 LU z
(/) 0.4
0.2
O^o o—o
. y
\y \y
•N o
* O
o o VELVETLEAF
e e SOYBEAN FIELD
0.0 600 800 1000 1200 1400 1600 1800
LOCAL SOLAR TIME (h)

68
surfaces through most of the day (Figure 11). In contrast, the terminal leaflet of
soybean leaves became oriented progressively more nearly parallel to the sun's
rays over the course of the day.
Under field conditions with greater insolation and reduced soil available
water (1888 was a very dry year con^ared to 1888 and 1887 - see Table 4)
velvetleaf again tracked the sun more closely than did soybean until about 1300 h
(Figure ll). Thereafter, leaves of velvetleaf were oriented either similarly to
those of soybean or had even lower angles of incidence. Some wilting of the
leaves of velvetleaf later in the day was evident, which would have adversely
affected these turgor driven leaf movements.
Since the sine of the angle of incidence is directly proportional to PAR
interception by the leaves, it is clear that velvetleaf intercepted a significantly
greater portion of available PAR during the day than did soybean under both field
and greenhouse conditions (Figure 11). Thus, solar-tracking could significantly
enhance the daUy LIE of velvetleaf compared to soybean. This was further
demonstrated by a second experiment in the field.
Leaves permitted to move freely (solar-track) during the day intercepted
more PAR during the early morning and late afternoon than those constrained to a
horizontal position (Figure 12). In addition to Improving light interception, solar-
tracking leaf movements also increase shading of other shorter species, especially
early and late in the day when solar elevations are low.
The CER of solar-tracking leaves of soybean was higher than that of
horizontal leaves early and late on July 21, 1888, a sunny day (Figure 13). During
the middle of the day, tracking leaves exhibited lower CER than nontracking

Figure 12. Effect of solar-tracking leaf movements on the diurnal course of interception of photosynthetically active radiation (PPFD) by velvetleaf and soybean plants under field conditions (July 21^ 1988). Leaves were either restrained in a horizontal position (leaf horizontal) or allowed to move freely (leaf free) throughout the day. Vertical bars represent one standard error of the mean
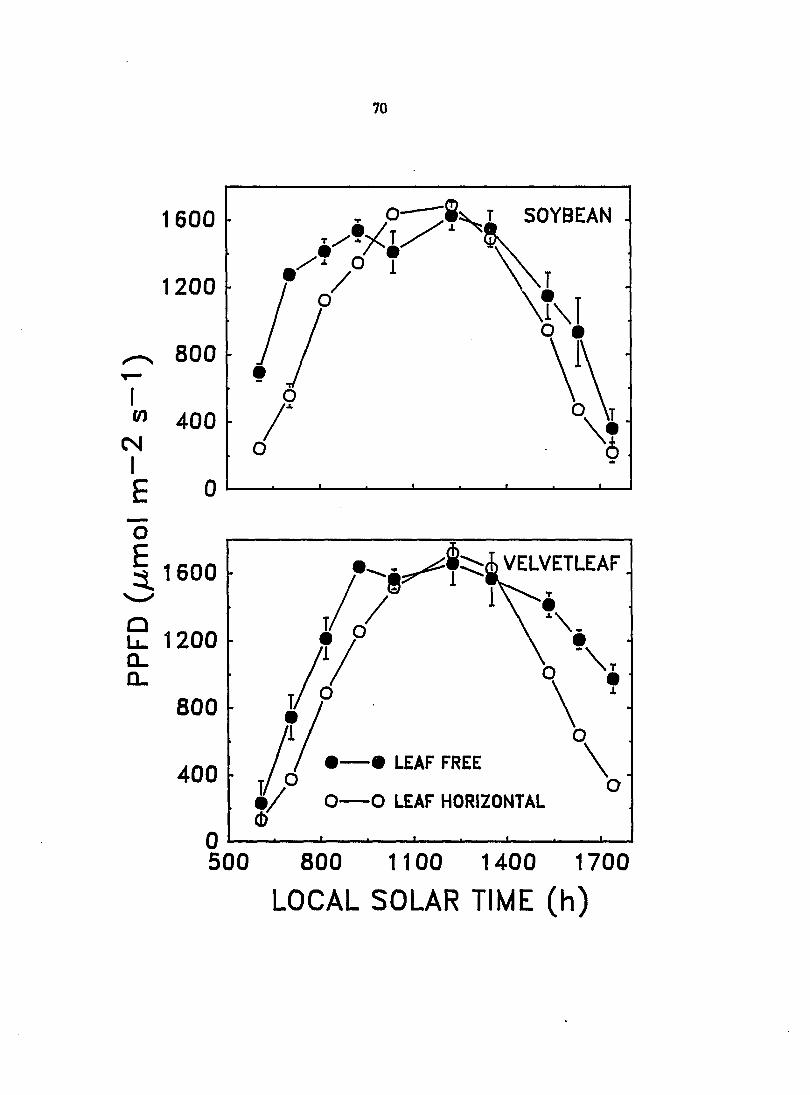
70
, O — S O Y B E A N m /_ / i
V
200 \T\T
O
1600
£ 1200 û-0_
800
400
VELVETLEAF
LEAF FREE
O LEAF HORIZONTAL
0 500 800 1100 1400 1700
LOCAL SOLAR TIME (h)

Figure 13. Effect of solar-tracking leaf movements on the diurnal course of carbon dioxide exchange rates (CER) of velvetleaf and soybean plants under field conditions (July 21, 1988). Leaves were either restrained in a horizontal position (leaf horizontal) or allowed to move freely (leaf free) throughout the day. Vertical bars represent one standard error of the mean
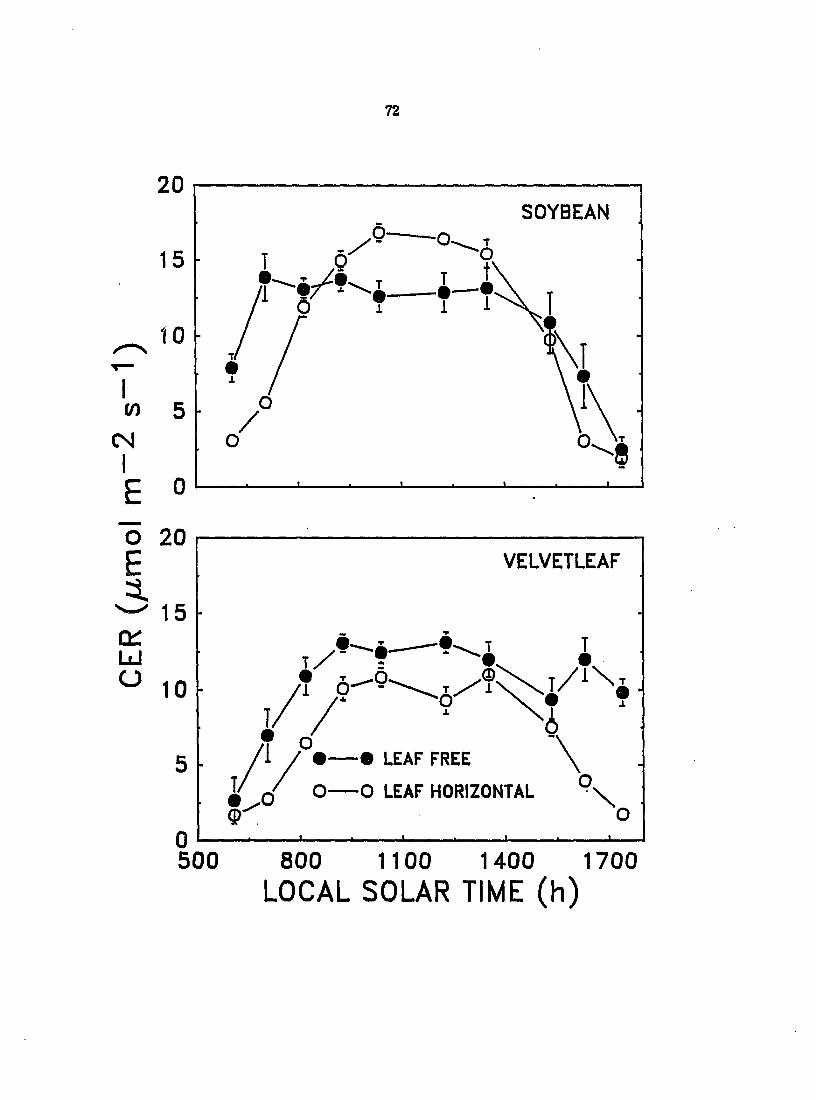
72
I to
CM I E
20
15 •
10
5 •
SOYBEAN
\i
O 20 VELVETLEAF
m LU O
15 •
10
Q'
1/
• LEAF FREE
•O LEAF HORIZONTAL
500 800 1100 1400 1700 LOCAL SOLAR TIME (h)

73
horizontal leaves (Figure 13) even though thqr received only slightly lower PAR
(Figure 12). Thus, freely moving soybean leaves did not qipear to avoid light
actively as occurs when they are under water stress (Meyer and Walker, 1981).
The experimental plots received 7.3 cm of rain during the three days preceding
the CER measurements, so it is unlikely that the plants were water stressed. This
may explain why th^ did not display the same pattern of leaf movements as
measured on August 5, 1988 (Figure 11), after two weeks without precipitation.
In general, CER values were higher for tracking leaves of velvetleaf,
compared to horizontal leaves, during most of the day and especially in the late
afternoon and early morning (Figure 13). Although PAR interception by tracking
and horizontal velvetleaf leaves was not significantly different during the middle
of the day, photosynthetic rates of freely moving leaves were somewhat higher
than those of horizontal leaves.
The diurnal course of tranq)iration of solar-tracking and horizontal
velvetleaf and soybean leaves (Figure 14) generally followed that of CER (Figure
13), although differences in transpiration rates between tracking and horizontal
leaves of either species were generally smaller than differences in CER.
Despite improved light interception and higher CER early and late in the
day, tracking (free) leaves of soybean did not have significantly increased amounts
of carbon fixed or WUE on either the cloudy or the clear day, as compared to
horizontal leaves (Table 8). Conversely, solar-tracking leaves of velvetleaf fixed
significantly more carbon and exhibited greater WUE than nontracking leaves on
both the cloudy and the clear day (Table 8). Even though tracking leaves of

Figure 14. Effect of solar-trackii^ leaf movements on the diurnal course of transpiration of velvetleaf and soybean plants under field conditions (July 21j 1988). Leaves were either restrained in a horizontal position (leaf horizontal) or allowed to move freely (leaf free) throughout the day. Vertical bars represent one standard error of the mean
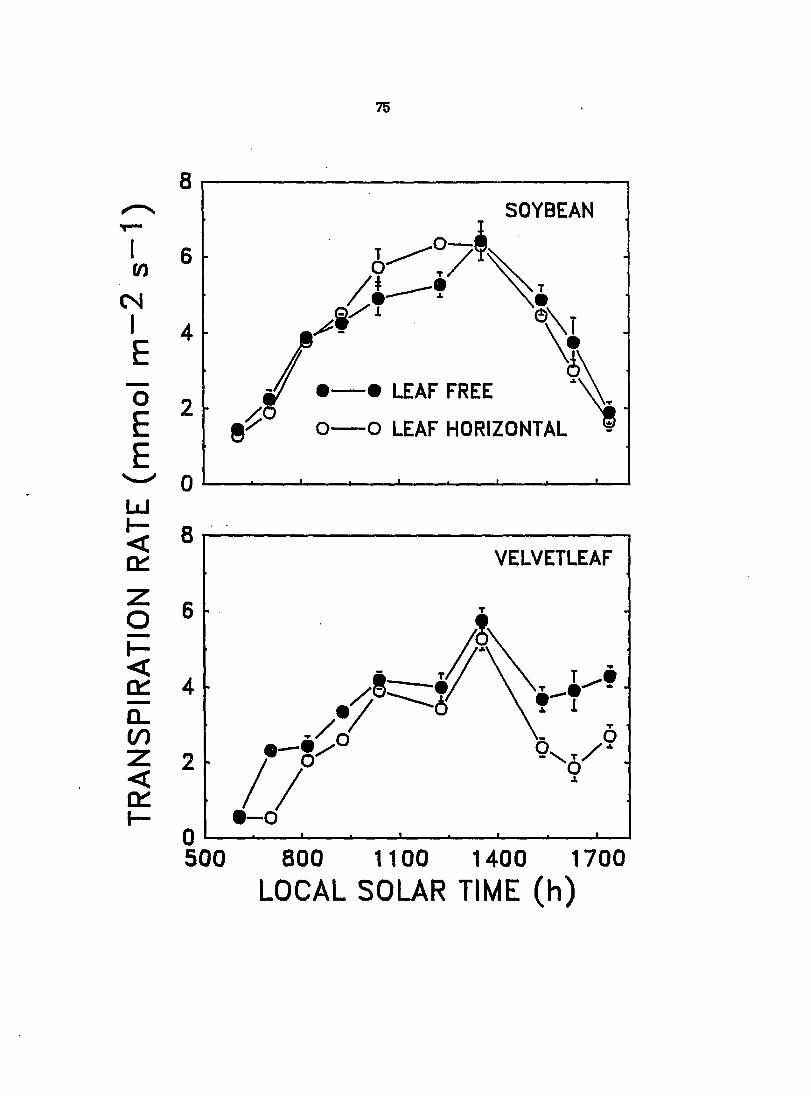
75
8
I w
CM I E "o
E
LU
< et:
h-< ck:
ÛL 10
z < n:
2 - /r
SOYBEAN
LEAF FREE
O O LEAF HORIZONTAL
VELVETLEAF
; /
500 800 1100 1400 1700
LOCAL SOLAR TIME (h)
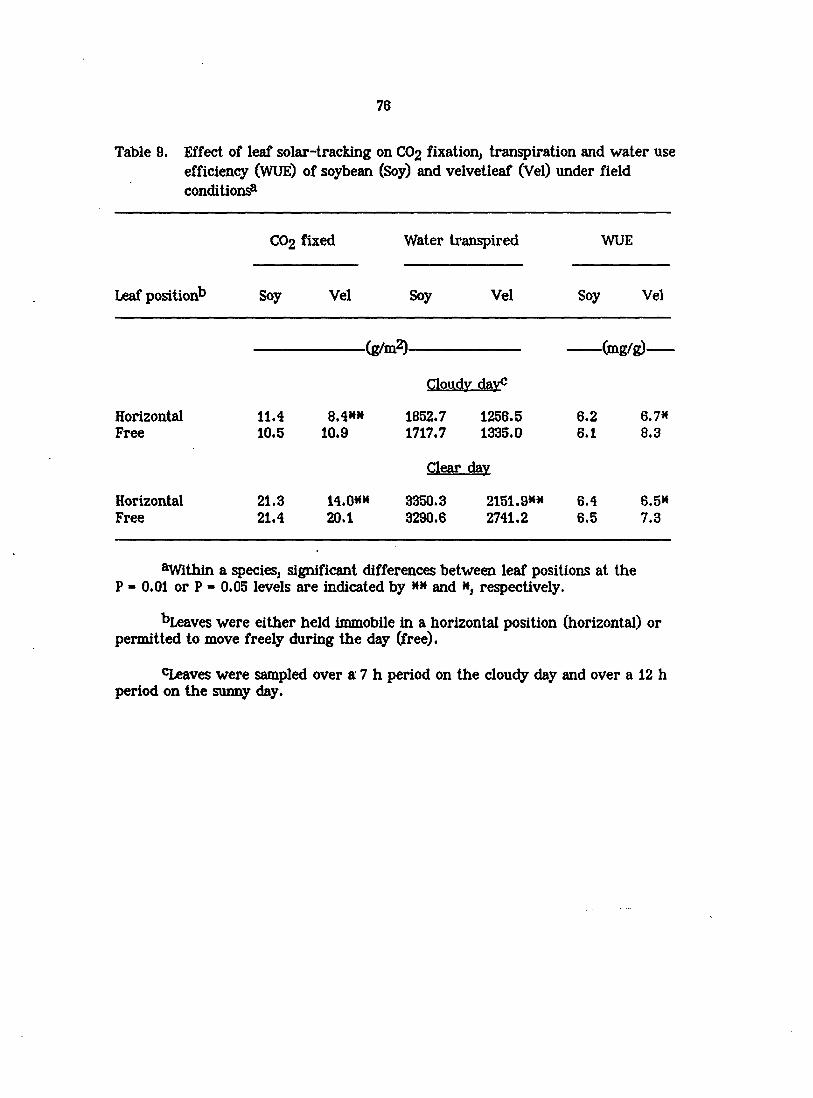
76
Table 9. Effect of leaf solar-tracking on CO2 fixation, transpiration and water use efficiency (WUE) of soybean (Soy) and velvetleaf (Vel) under field condition#
CO2 fixed Water transpired WUE
Leaf position^ Soy Vel Soy Vel Soy Vel
-(g/m2)- -(mg/g)-
Cloudv dayC
Horizontal Free
11.4 10.5
(L4M* 10.9
1852.7 1717.7
1256.5 1335.0
6.2 6.1
6.7* 8.3
Clear day
Horizontal Free
21.3 21.4
14.0** 20.1
3350.3 3290.6
2151.9** 2741.2
6.4 6.5
6.5* 7.3
^Within a species, significant differences between leaf positions at the P - 0.01 or P - 0.05 levels are indicated by ** and *, respectively.
^Leaves were either held immobile in a horizontal position (horizontal) or permitted to move freely during the day (free).
^Leaves were sampled over a 7 h period on the clou<fy day and over a 12 h period on the sunny day.

77
velvetleaf transpired more water than nontracking leaves on the clear (but not
the cloudy) day, their greater carbon fixation resulted in a higher WUE.

78
SUMMARY AND CONCLUSIONS
The role of competition for light in interference between velvetleaf and
soybean was evaluated at the whole plant and canopy levels in field and
greenhouse studies conducted from 1986 to 1988 at Ames, Iowa. In field studies, a
deWit replacement series, consisting of velvetleaf/soybean species proportions of
lOO/Oj 50/50, 25/75, 12.5/87.5, and 0/100, was used to examine the pattern of
interference between the two species during the course of the growing season.
Growth analysis harvests were taken at approximately 2 week intervals between 5
and 12 weeks after emergence. Leaf areas, plant heights, numbers and vertical
distribution of mainstem branches, and plant dry weights were determined. Seed
yields were also determined at maturity.
In conjunction with growth analysis harvests, the amount of PAR
intercepted at solar noon by the canopies of the various species proportions was
determined. Light conversion analysis (Warren Wilson, 1981) was employed to
assess each species^ efficiency of interception and utilization of light for dry
matter production. FAR interception was also monitored at several heights in
monoculture velvetleaf and soybean canopies throughout the day in order to
compare their diurnal patterns of light interception. Additionally, the diurnal
patterns of leaf movement of isolated velvetleaf and soybean plants were examined
under field conditions. The significance of these movements to light interception,
CER, transpiration, and WUE of these species was also determined.
Analysis of replacement series results indicated that interference between
velvetleaf and soybean does not occur early in the growing season when individual
plants are fairly widely separated. However, as canopy closure occurs, velvetleaf
gains resources at the expense of soybean and thereby reduces the vegetative and

79
reproductive growth of the crop. The presence of verticillium wilt disease in
velvetleaf appeared to reduce its ability to interfere with soybean. This was
partly due to premature leaf drop associated with the disease, which markedly
reduced the leaf area of velvetleaf and, hence, its ability to intercept and convert
light to dry matter.
Velvetleaf plants were taller than soybean plants on all sampling dates in
1986 and 1987. Velvetleaf displayed greater leaf area than soybean in 1986 until
later in the growing season. However, in 1987, the leaf area of soybean plants
was greater than that of velvetleaf plants until mid- to late-season. Velvetleaf
also had many times more mainstem branches than soybean. Branching in
velvetleaf was concentrated near the top of the plant while the limited branching
that occurred in soybean was confined to lower axillary buds. Although soybean
plants are initially taller and have more leaf area than velvetleaf plants,
velvetleafis more rapid early growth rate, greater leaf area expansion rate (Potter
and Jones, 1977), and greater allocation of dry matter into stem growth in
velvetleaf, rapidly produce a taller plant with a larger leaf surface area available
for PAR interception.
The MAR and RGR of velvetleaf were higher than those of soybean during
early to midseason in 1986 and 1987. Velvetleaf interference had less effect on
growth analysis parameters of soybean in 1987 than in 1986 due to a greater
competitive ability of soybean not affected by disease. In the absence of leaf
area-depleting diseases in either species, there were no significant differences in
lAI among species proportions. Such results suggest roughly equivalent light
interception and productivity capabilities of both monoculture and mixed canopies
of these species.

80
The midday LIE of monoculture canopies of both species was also markedly
reduced by disease. However, in the absence of disease, the LIE of all species
proportions were roughly equivalent, a reflection of the earlier noted similarities
in LAI of canopies of these species. Fully developed monoculture canopies of both
velvetleaf and soybean intercept nearly all available PAR. However, a greater
percentage of c^tured PAR is intercepted closer to the top of the canopy in
velvetleaf than in soybean. Furthermore, the daily LIE of velvetleaf may be
greater than that of soybean. While the distribution of PAR interception
throughout a soybean canopy profile changes little during the day, velvetleaf
appears to capture a greater proportion of the PAR it intercepts early and late in
the day near the top of the canopy. This would effectively limit the PAR
available to soybean plants infested with taller velvetleaf plants, especially during
the early morning and late afternoon.
Although both species exhibited leaf solar-tracking movements under well-
watered conditions in the field and greenhouse, leaves of velvetleaf tracked the
sun more closely than those of soybean. Under field conditions, solar-tracking
movements increased the PAR interception and CER of leaves of both species early
and late in the day compared to immobile leaves. However, while such movements
significantly enhanced the daily carbon fixation c^acity and improved the WUE of
velvetleaf, they did not do so in soybean.
The results of the studies reported here and those previously reported
elsewhere in the literature support the conclusion that much of the interference
between velvetleaf and soybean is due to conqïetition for light. Velvetleaf
possesses a number of attributes which enhance its ability to compete with
soybean for light. These include a rapid early growth ratej rapid, extensive stem

81
growth which leads to a tall stature; a high leaf area expansion rate; and a leaf
senescence and branching pattern that situates much of the leaf area near the top
of the plant, above the leaves of shorter competing species. Additionally, active
leaf movements which improve the WUE, light interception and photosynthetic
activity of velvetleaf (and concomitantly increase shading of associated shorter
species) also contribute to its greater ability to compete for light.

82
LITERATURE CITED
Andersei^ R. N. 1968. Germination and establishment of weeds for experimental Purposes. W. F. Humphry Pressj Inc., Geneva, New York.
Baldocchij D. D., S. B. Vermiy N. J. Rosenberg, B. L. Blad> A. Garay, and J. E. S^>echt. 1983. Influence of water stress on the diurnal exchange of mass and energy between the atmo^here and a soybean canopy. Agron. J. 75:543-548.
Begonia, G. B., and R. J. Aldrich. 1888. Responses of soybean to light quality altered by velvetleaf (Abutilon theophrasti Medic.). Abstr. Weed Sci. Soc. Am. 28:95.
Benner, B. L., and F. A. Bazzaz. 1885. Response of the annual Abutilon theophrasti Medic. (Malvaceae) to timing of nutrient availability. Am. J. Bot. 72:320-323.
Berg, V. S.j and T. C. Hsiao. 1888. Solar tracking: light avoidance induced by water stress in leaves of kidn^ bean seedlings in the field. Crop Sci. 28:980-986.
Bhowmik, P. C., and J. D. Doll. 1982. Com and soybean response to allelopathic effects of weed and crop residues. Agron. J. 74:801-606.
Blackman, N. H. 1919. The con^und interest law and plant growth. Ann. Bot. 33:353-380.
Bunce, J. A. 1985. Volume and osmotic potential changes in relation to inhibition of photoqmthesis by water stress in intact leaves. Can. J. Bot. 64:557-560.
Charles-Edwards, D. A., and M. J. Fisher. 1980. A philosophical approach to the analysis of cr<^ growth data. I. Theoretical considerations. Ann. Bot. 48:413-423.
Chazdon, R. L. 1885. Leaf diq>lay, canopy structure, and light interception of two understory palm species. Am. J. Bot. 72:1493-1502.
Cheetham, J. Z. 1889. Growth, water relations, and gas exchange of intercropped soyt)ean and velvetleaf. Ph.D. Dissertation. Iowa State Univ., Ames.
Child, R., D« C. Morgan, and H. Smith. 1981. Morphogenesis in simulated shadelight quality, p. 409-420. In H. Smith (ed.) Plants and the daylight spectrum. Academic Pres^ New York.

83
ColtoDj C, E.) and F. A. Einhellig. 1980. AUelopathic mechanisms of velvetleaf (AbtttUon theophrasti Medic.^ Malvaceae) on soybean. Am. J. Bot. 67:1407-1413.
Connolly) J. 1988. On difficulties with replacement-series methodology in mixture experiments. J. Appl. Ecol. 23:125-137.
Dekker^ J. H., and W. F. Meggitt. 1883a. Interference between velvetleaf fAbutilon theophrasti Medic.) and soybean fGlycine quo. CL>) Merr. ]. I. Growth. Weed Res. 23:91-101;
DeUeer^ J. H., and W. F. Meggitt. 1983b. Interference between velvetleaf fAbutilon theophrasti Medic.) and soybean fGlycine (L.) Merr.]. II. Population dynamics. Weed Res. 23:103-107.
deWltj C. T. 1960. On con^tition. Versl. Landbouvtd. Onderz. No. 66:1-82.
Donaldj C. M. 1961. Gon^)etition for light in crops and pastures, p. 282 313. bk Mechanisms in biological con^tition. Soc. Exp. Biol. Syn^>osia 15. Academic Fres^ New York.
Eatonj B. J.) 0. G. Russ^ and K. C. Feltner. 1976. Competition of velvetleaf^ pridcly sida, and Venice mallow in soybeans. Weed Sci. 24:224-228.
Ehleringerj and I. Forseth. 1980. Solar tracking by plants. Science 210:1084-1098,
Elmore, C. D. 1880. Inhibition of turnip (Brassica rapa) seed germination by velvetleaf (Abutilon theophrasti) seed. Weed Sci. 28:658-660.
FehTj W. R.j C. E. Caviness, D. T. Burmood, and J. S. Pennington. 1871. Stage of development descriptions for soybeans fGlycine max CQ. Merr.]. Crop Sci. 11:828-831.
Firbankj G., and A. R. Watkinson. 1885. On the analysis of competition. J. Appl. Ecol. 22:503-617.
Frazee, R. W., and E. W. StoUer. 1874. Differential growth of com, soybean, and seven dicotyledonous weed seedlings. Weed Sci. 22:336-338.
Gressel, J. B., and L. G. Holm. 1864. Chemical inhibition of crop germination by weed seeds and the nature of inhibition by Abutilon theophrasti. Weed Res. 4:44-63.

84
Hagoodj E, S.) Jr., T. J. Bauman, J. L V^Uiams, Jr., and M. M. Schreiber. 1980. Growth analysis of soybeans (Glycine max) in competition with velvetleaf (Abutilon theophrasti). Weed Sci. 28:728-734.
Hall; R, L. 1974. Analysis of the nature of interference between plants of different species. I. Concepts and extension of the deWit analysis to examine effects. Aost. J. Agric. Res. 25:739-747.
Harper, J. L. 1977. Population biology of plants. Academic Press, New York.
Hatfield, J. L, and R. E. Carlson. 1979. Leaf-'water-potential patterns within two soybean cultivars. Iowa State J. Res. 53:259-287.
Higgins, R. A., D. W. Staniforth, and L. P. Pedigo. 1984. Effects of weed density and defoliated or undefoliated soybeans (Glycine max) on velvetleaf (Abutilon theophrasti) development. Weed Sci. 32:511-519.
Hunt, R. 1878. Plant growth analysis. Studies in Biology Series no. 96. Edward Arnold, London.
Hunt, R. 19%. Plant growth curves. The functional approach to plant growth analysis. Edward Arnold, London.
Hunt, R., J. Warren Wilson, D. W. Hand, and D. G. Sweeney. 1984. Integrated analysis of growth and light intercepti<m in winter lettuce. I. Analytical methods and environmental influences. Ann. Bot. 54:743-757.
Jolliffe, P. A., A. N. Minjas, and V. C. Runeddes. 1984. A reinterpretation of yield relationships in replacement series experiments. J. Appl. Ecol. 21:227-243.
Eawashima, R. 1969a. Studies on the orientation-adjusting movement in soybean plants. I. The leaf orientation-adjusting movement and light intensity on leaf surface. Proc. Crop Sci, Soc. Japan 38:718-729.
Kawashima* R. 1968b. Studies on the leaf orientation-adjusting movement in soybean plants. H. Fundamental pattern of leaf orientation-adjusting movement and its significance for the diy matter production. Proc. Crop Sd. Soc. Japan 38:730-742.
Lan& A. R. G. 1973. Leaf orientation of a cotton plant. Agric. Meteorol. 11:37-51.
Mqrer, W. S., and J. T. Ritchie. 1980. Resistance to water flow in the sorghum plant. Plant Physiol. 85:33-39.

85
Meyer^ W. S.j and S. Walker. 1981. Leaflet orientation in water-stressed soybeans. Agron. J. 73:1071-1074.
Mooney, H. A., and J. R. Ehleringer. 1878. The carbon gain benefits of solar tracking in a desert annual. Plant* Cell Environ. 1:307-311.
Morgan* D. C., and H. Smith. 1978. The relationship between phytochrome photoequilibrium and development in light grown Chenopodimn alhnm L. Planta 142:187-193.
Hunger* P. H.* J. M. Chandler* and J. T. Cothren. 1987a. Effect of water stress on photosynthetic parameters of soybean (Glycine msz) and velvetleaf (Abutilon theophrasti). Weed Sci. 35:15-21.
Munger* P. H.* J. M. Chandler* J. T. Cothren* and F. M. Hons. 1887b. Soybean (SWns. max) - velvetleaf (Abutilon theophrasti) interspecific competition. Weed Sci. 35:647-653.
Myers* R. L.* W. A. Brun* and M. L. Brenner. 1987. Effect of raceme-localized supplemental light on soybean reproductive abscission. Crop Sci. 27:273-277.
Oliver* L. R. 1979. Influence of soybean (Glycine s^) planting date on velvetleaf (Abutilon theophrasti^ competition. Weed Sci. 27:183-188.
Patterson* D. T. 1982. Effects of light and ten^rature on weed/crop growth and competition, p. 407-420. ^ J. L. Hatfield and I. J. Thomason (ed.). Biometeorology in integrated pest management. Academic Press* New York.
Patterson, D. T.* and E. P. Flint. 1983. Comparative water relations* photoqmthesis* and growth of soybean (Glycine msO and seven associated weeds. Weed Sci. 31:318-323.
Potter* J. R.* and J. W. Jones. 1877. Leaf area partitioning as an important factor in growth. Plant Physiol. 59:10-14.
Radosevich* S. R.* and J. S. Holt. 1984. Weed ecology: Implications for vegetation management. John WU^ and Sons* New York.
Regoier* E. E. 1987. Aspects of velvetleaf (Abutilon theophrasti Medic.)* Jimsonweed (Datura stramonium L,)* and common cocklebur (Xanthium gtninmriiiin L.) interference with soybeans. Ph.D. Dissertati<m. Univ. Illinois* Urbana-Champaign (Diss. Abstr. Int. 48:306).

86
Regnier, E. E.j M. E, Salvucci, and E. W. StoUer. 1888. Photosynthesis and growth responses to irradiance in soybean (Glycine max) and three broadleaf weeds. Weed Sci. 38;487-486.
Retig, B.) L. G. Holm, and B. E. Struckmeyer. 1872. Effects of weeds on the anatomy of roots of cabbage and tomato. Weed Sci. 20:33-36.
RhodeSj I., and W. R. Stem. 1878. Coiiq>etition for light, p. 175-188. In J* R, WUsoD) (ed.). Plant relations in pastures. CSIROj Australia.
Roushj M. L.j and S. R. Radosevich. 1885. Relationships between growth and competitiveness of four annual weeds. J. Appl. Ecol. 22:885-905.
Salisbury, C. D. 1885. The effects on soybeans of velvetleaf competition for light. Masters Thesis. Univ. Missouri, Columbia. (Masters Theses in Pure and implied Sciences 30:27).
Shell) G. S. G., and A. R. G. Lang. 1876. Movements of sunflower leaves over a 24-h period. Agric. Meteorol. 16:161-170.
Shell) G. S. G.) A. R. G. Lang, and J. M. Sale. 1874. Quantitative measures of leaf orientation and hellotr<^ic response in sunflower, bean, pepper and cucumber. Agric. Meteorol. 13:25-37.
Shroyer, J. P. 1878 Profitable soybean production. Rn-441. Cooperative Extension Service. Iowa State University, Ames.
Shurtleff, J. L., and H. D. Coble. 1885. The interaction of soybean (Glycine maxl and five weed species in the greenhouse. Weed Sci. 33:668-672.
Silvertown, J. W. 1882. Introduction to Plant Population Biology. Longman, New York.
Spencer, N. R, 1884. Velvetleaf, Abutilon theophrasti (Malvaceae), history and ecwomic impact in the United States. Econ. Bot. 38:407-416.
Spittersi, C. J. T., and R. Aerts. 1883. Simulation of conq>etition for light and water in crop-weed associations. Aspects Appl. Biol. 4:467-483.
Staniforth, D. W. 1862. Responses of nodolating and non-nodulating soybeans to weed competition. Proc. North Cent. Weed Cont. Conf. 18:48.
Staniforth, D. W. 1865. Competitive effects of three foztaU q)ecies on soybeans. Weeds 13:181-183.

87
Sterling, T. M., and A. R. Putnam. 1987. Possible role of glandular trichome exudates in interference by velvetleaf (Abutilon theophrastil. Weed Sci. 35:308-314.
Sterling, T. M., R. L. Houtz> and A. R. Putnam. 1887. Fhytotoxic exudates from velvetleaf (Abutilon theophrasti Medic.) glandular trichomes. Am. J. Bot. 74:543-550.
StoUer* E. W.j and J. T. Woolley. 1885. Competition for light by broadleaf weeds in soybean (Glycine Weed Sci. 33:198-202.
Taylor, S. E. 1971. Ecological in^lications of leaf morphology considered from the stan(!^int of energy relations and productivity. Ph.D. Dissertation. Washington Univ., St. Louis (piss. Abstr. Int. 71:150).
Trenbath, B. R. 1974. Biomass productivity of mixtures. Adv. Agron. 26:177-210.
Tucker, D. J. 1976. Effects of far-red light on the hormonal control of side shoot growth in the tomato. Ann. Bot. 40: 1033-1042.
Tucker, D. J., and T. A. Mansfield. 1972. Effects of light quality on apical dominance in Xanthium strumariwm and the associated changes in endogenous levels of abscisic acid and cytokinins. Planta 102: 140-151.
Warren Mlson, J. 1981. Analysis of growth, photo^thesis and light interception for single plants and stands. Ann. Bot. 48:507-512.
Warren Wlson, J., R. Hunt, and D. W. Hand. 1988. Philosophical aq>ects of measurements, equations and inferences in plant growth studies. Ann. Bot. 58:73-80.
Wofford, T. J., and F. L. Allen. 1982. Variation in leaflet orientation among soybean cultivars. Crop Sci. 22:999-1004.
Tun, J. I., and S. E. Taylor. 1986. Adaptive implications of leaf for sun- and shade-grown Abutilon theophrasti. Ecology 67:1314-1318.
Zimdahl, R. L. 1980. Weed-crop competition: A review. Int. Plant Prot. Ctr., Oreg(m State Univ., Corvallis.

88
ACKNOWLEDGEMENTS
Sincere thanks are extended to all members of my committee for their help
in designing and executing the experiments, and for their thorough review of the
dissertation. I also wish to acknowledge Dr. Dekker^s friendly "^open door policy*'
which made the whole process seem easier. The efforts of Chris Dionigi in
assisting with field work under the searing sun of lowan summers are gratefully
«appreciated. My deepest thanks go to my wife for her steadfast love and abiding
endurance during the time she spent as a "^dissertation widow^.


















