
The ribosome and protein synthesis (part II)faculty.washington.edu/merza/pdf/merz_L26h.pdf ·...
24
Lecture 26 The ribosome and protein synthesis (part II) Noller Group, UCSC • http://rna.ucsc.edu/rnacenter/ribosome_rht.html Alex Merz, Ph.D. [email protected]
Transcript of The ribosome and protein synthesis (part II)faculty.washington.edu/merza/pdf/merz_L26h.pdf ·...

Lecture 26The ribosome and protein synthesis (part II)
Noller Group, UCSC • http://rna.ucsc.edu/rnacenter/ribosome_rht.html
Alex Merz, Ph.D. [email protected]

The ribosome has three tRNA binding sites:
Aminoacyl-tRNAPeptidyl-tRNAExit
To understand initiation, you first must understand the basics of elongation!

Three major steps in elongation:
A site:tRNA selection
P site:peptidyl transfer
translocation:uncharged tRNAexits from E site
Note that the growing polypeptide chain is transferred onto the incoming aa-tRNA!
The aa on the incoming aa-tRNA is not transferred onto the chain!

selection/accomodationpeptidyltransfer
translocation
Elongation cycle: the hybrid state model of elongation
The good news: much simpler than VVP 2/e, fig. 26-37The bad news: you’ve got to know this one cold.
Ef-TuGTP
Ef-GGTP
E P AAE P
50S
30S
AE PE P A E P A
A/T P/E A/P
hybridstate

The N-terminus of the growing polypeptide worms through an exit tunnel in the large subunit
N
5´3´
Macrolide antibiotics plug the exit tunnel from the inside, preventing polypeptide exit!
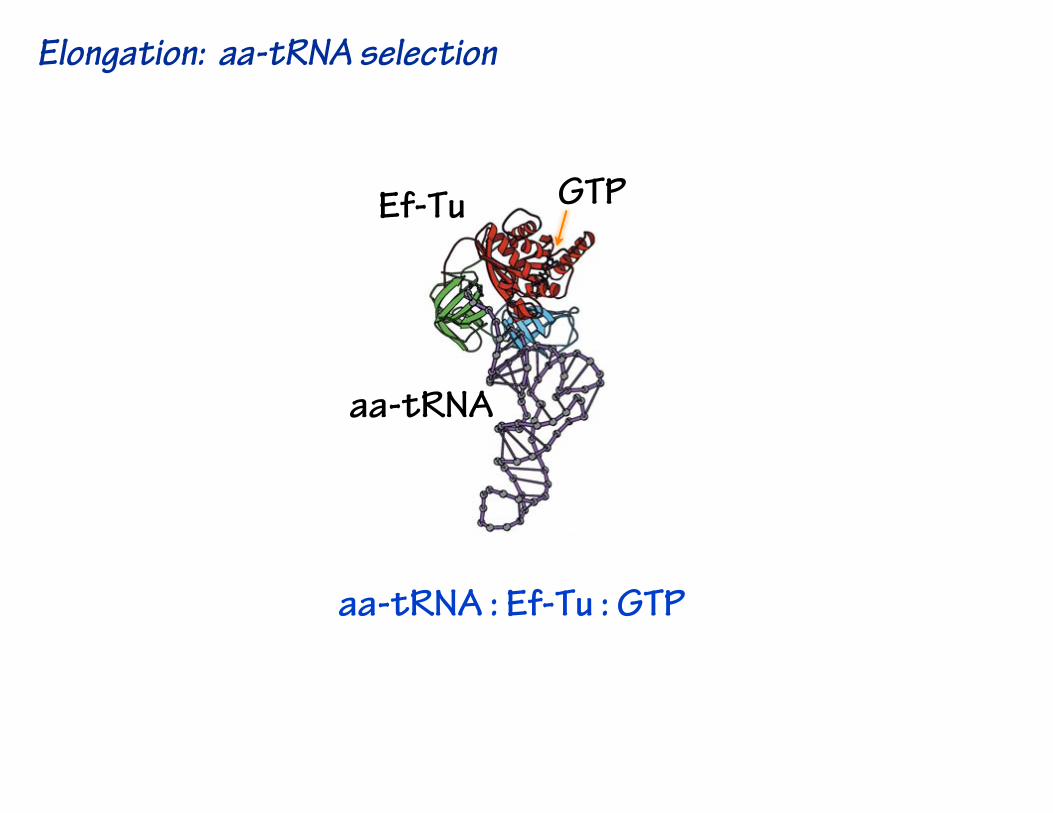
aa-tRNA : Ef-Tu : GTP
Elongation: aa-tRNA selection
Ef-Tu GTP
aa-tRNA

Elongation: aa-tRNA selection
Notice: there are two steps at which the aa-tRNA can dissociate (fall off) before peptidyl transfer!
This is a true proofreading mechanism.
The antibiotic tetracycline binds the 30S subunit’s A-site, blocking entry of the aa-tRNA-Ef-Tu complex!

selection/accomodationpeptidyltransfer
translocation
Elongation cycle: selection & accomodation
The good news: much simpler than VVP 2/e, fig. 26-37The bad news: you’ve got to know this one cold.
Ef-TuGTP
Ef-GGTP
E P AAE P
50S
30S
AE PE P A E P A
A/T P/E A/P
hybridstate

Valle et al. (2002) EMBO Journal
21:3557
aa-tRNA : Ef-Tu : GTPadopts a bent conformaton in the decoding site.
Elongation: aa-tRNA selection

examine and discard an incorrectaminoacyl-tRNA (Figure 3). Thismeans that the binding energybetween the ribosome and theternary complex can be sampledtwice and the specificity thusincreased. While the idea of havingconsecutive selective steps issimilar to the ‘double-sieveediting’ mechanism discussedabove, it is distinguished by thefact that kinetic proofreadingapplies the same basic selectivestep twice, whereas editinggenerally relies on a seconddistinct site or activity thatmonitors different properties thanthe first selective step.
Kinetic proofreading duringtRNA selection is made possibleby the fact that aminoacyl-tRNAsare delivered to the ribosome in aternary complex with the GTPaseelongation factor Tu (EF-Tu inbacteria, EF1A in eukaryotes) andGTP. In an encounter betweenternary complex and the ribosome(initial selection), a cognate ternarycomplex is more likely to triggerGTP hydrolysis than to dissociate,whereas a near-cognate ternarycomplex is more likely todissociate. Simply put, the cognatespecies partitions forward in thestepwise scheme whereas thenear-cognate partitions backward.This initial selection step isfollowed by the proofreading stepwhere inherent binding differencesbetween codon and anticodon areagain sampled. As before, thecognate aminoacyl-tRNA speciesis more likely to partition forward(and ‘accommodate’ into the A siteand participate in peptide bond
formation), while the near-cognateaminoacyl-tRNAs are more likely topartition backward (and berejected from the ribosome).
The relative contribution of eachof these selective steps, initialselection and proofreading, hasbeen measured in vitro in multipleways, where overall error rates of~1 in 450 to 1 in 1600 approachthe overall fidelity measured invivo. In these systems, essentiallyall non-cognate aminoacyl-tRNAsare rejected during initialselection. Near-cognateaminoacyl-tRNAs, however, canpass through initial selection andtrigger GTP hydrolysis with afrequency of ~1 in 30. Thesesneaky aminoacyl-tRNAs aregenerally rejected during thesecond stage, thus increasingselectivity by ~15–45-fold.
Interestingly, the maximaltheoretical selectivity of kineticproofreading is not realized herebecause of the processive natureof translation and associatedrequirement for speed. Tomaximize each selective step, theforward rates should be slowenough that differences indissociation rates can beexploited. Indeed, experimentalevidence shows that, when GTPhydrolysis is made extremelyslow, the selectivity observed inthis initial selection step issubstantially increased. It waslong ago suggested that ribosomemutations which affect the fidelityof tRNA selection act similarly byincreasing or decreasing the ratesof individual steps in the selectionprocess.
While separating the processinto two steps — kineticproofreading — does provide anadvantage during tRNA selection,it is not because differences indissociation rates betweencognate and near-cognateaminoacyl-tRNAs are exploitedtwice, as previously thought.Rather, during each stage oftRNA selection a second strategycomes into play that introduces alarge difference in the rates oftwo critical forward steps. Duringinitial selection, the rate ofGTPase activation (k3) issignificantly faster for thecognate than for near-cognateaminoacyl-tRNAs, and duringproofreading, there are similardifferential rates ofaccommodation (k5) (Figure 3).These differences in forwardrates have been attributed to amechanism historically termedinduced fit, which is used by thetranslation machinery,polymerases and a number ofother enzymes. Induced fit refersto the ability of a correctsubstrate, but not an incorrectone, to cause conformationalchanges in the enzyme and/orthe substrate which havedownstream effects on catalysis.During tRNA selection on theribosome a series ofconformational changes inducedby binding of the cognateaminoacyl-tRNA, but not a near-cognate one, result in a numberof rearrangements in theribosome and the tRNA itself thatresult in the kinetic effectsdiscussed above.
Magazine R539
Figure 3. Detailed kinetic scheme for tRNA selection highlighting the two stages of the process, initial selection and proofreading. The selectivity of the initial selection stage is determined by the difference in rate of GTPase activation (k3) between the cognate and anear-cognate tRNA. The selectivity of the proofreading stage is determined primarily by the difference in rate of accommodation (k5)between the cognate and a near-cognate tRNA. EF-Tu (green) is shown in two different conformations before and after GTP hydrolysis.
Initialbinding
Codonrecognition
GTPase activation/GTP hydrolysis
Accommodation/peptidyl transfer
Rejection
Initial Selection
Proofreading
k1
k–1
k2 k3 + kGTP
k5 + kPEP
k7k–2
Current Biology
Ef-Tu:GTP Ef-Tu:GDP
Rachel Green
Kinetic proofreading: the timer function of Ef-Tu is critical for accuracy
Elongation: aa-tRNA selection
A slowly-hydrolyzed GTP analog increases accuracy of translation — but at the expense of speed.

selection/accomodationpeptidyltransfer
translocation
The good news: much simpler than VVP 2/e, fig. 26-37The bad news: you’ve got to know this one cold.
Ef-TuGTP
Ef-GGTP
E P AAE P
50S
30S
AE PE P A E P A
A/T P/E A/P
hybridstate
GTPhydrolysis
Elongation: peptidyl transfer
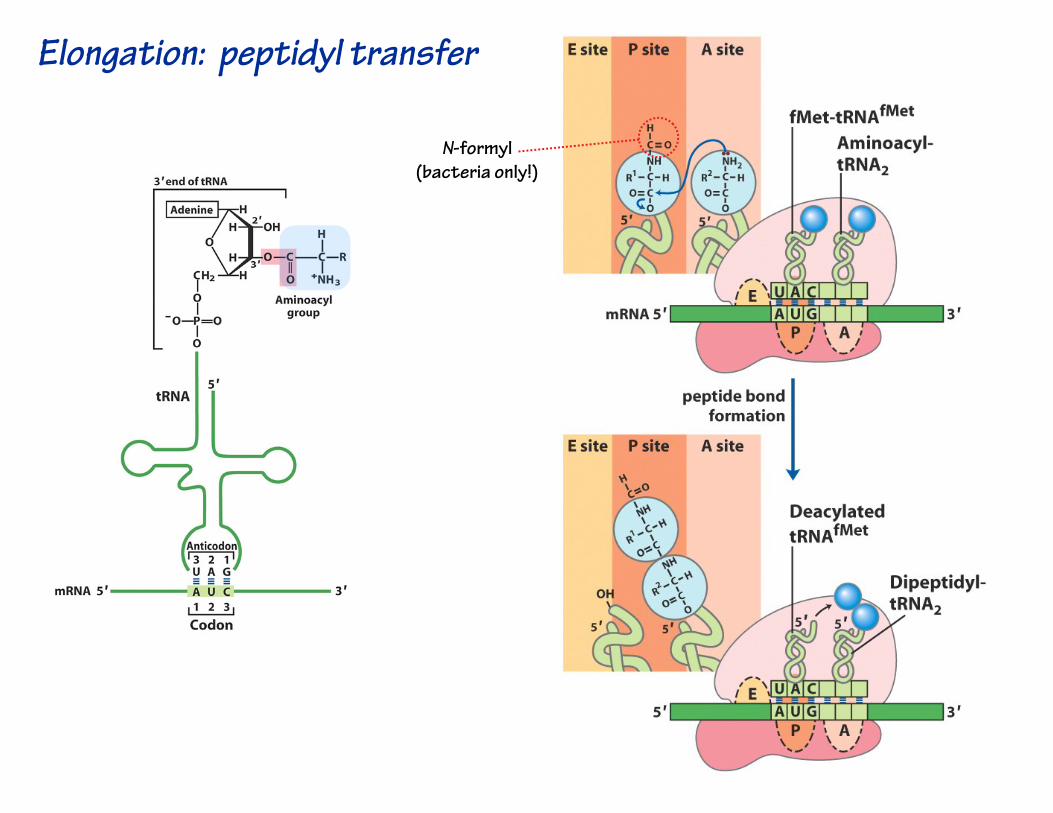
N-formyl(bacteria only!)
Elongation: peptidyl transfer

The antibiotic Puromycin mimics anaminoacylated tRNA
E P A E P A
P/E A/P
…but it is a polypeptide chain terminator!
Elongation: Puromycin is a chain terminator

“Fragment reaction” mimics peptidyltransfer
f-[35S]Met is a radioactive form of the amino acid
formylmethionine
E
50S
50S
*
P A
E P A
*
REACTANTS
PRODUCTS
*

“Fragment reaction” reveals: peptidyltransferase enzyme is RNA — not protein!!!

Peptidyltransferase center (PT) within the large ribosomal subunit
Similar view as left, but a space-filling model
Structure confirms Noller result: Ribosome is a ribozyme — no proteins within 18 Ångstroms of peptidyltransfer active site!

Elongation: many antibiotics bind in peptidyltransfer center (PTC)
TA Steitz and PB Moore (2003) TiBS 28, 411–418
Tetracyclinebinds 30S A-site, prevents aa-tRNA entry
• Puromycinbinds in PTC, chain terminator
• Chloramphenicolbinds in PTC, inhibits peptidyltransfer reaction and blocks exit tunnel
• Macrolidesbind in PTC, block exit tunnel

selection/accomodationpeptidyltransfer
translocation
Elongation cycle: the hybrid state model of elongation
The good news: much simpler than VVP 2/e, fig. 26-37The bad news: you’ve got to know this one cold.
Ef-TuGTP
Ef-GGTP
E P AAE P
50S
30S
AE PE P A E P A
A/T P/E A/P
hybridstate
GTPhydrolysis
GTPhydrolysis

Ef-Tu:GTP+ aa-tRNA
Ef-G:GTP
decoding translocation
Elongation cycle: translocation is powered by Ef-G, a GTP-powered motor

30S subunit viewed from the perspective of the 50S subunit.
Shifts between RNA phosphorus atoms and protein Cα atoms in unrotated and hybrid states color coded as indicated by scale.
Dunkle et al., Science 332:981 (=2 nm)
(hybrid state)
Elongation cycle: the hybrid state model of elongation
NOTE - add movie from
Valle & Frank, Cell 2003

Termination — release of polypeptide from tRNA in P site
5´3´
folded protein domains
50S subunit
tRNA (inside ribosome, in P site)

decoding…
translocation…
tRNA : Ef-Tu
Ef-G
Termination — release factors shaped like tRNA!
…termination.
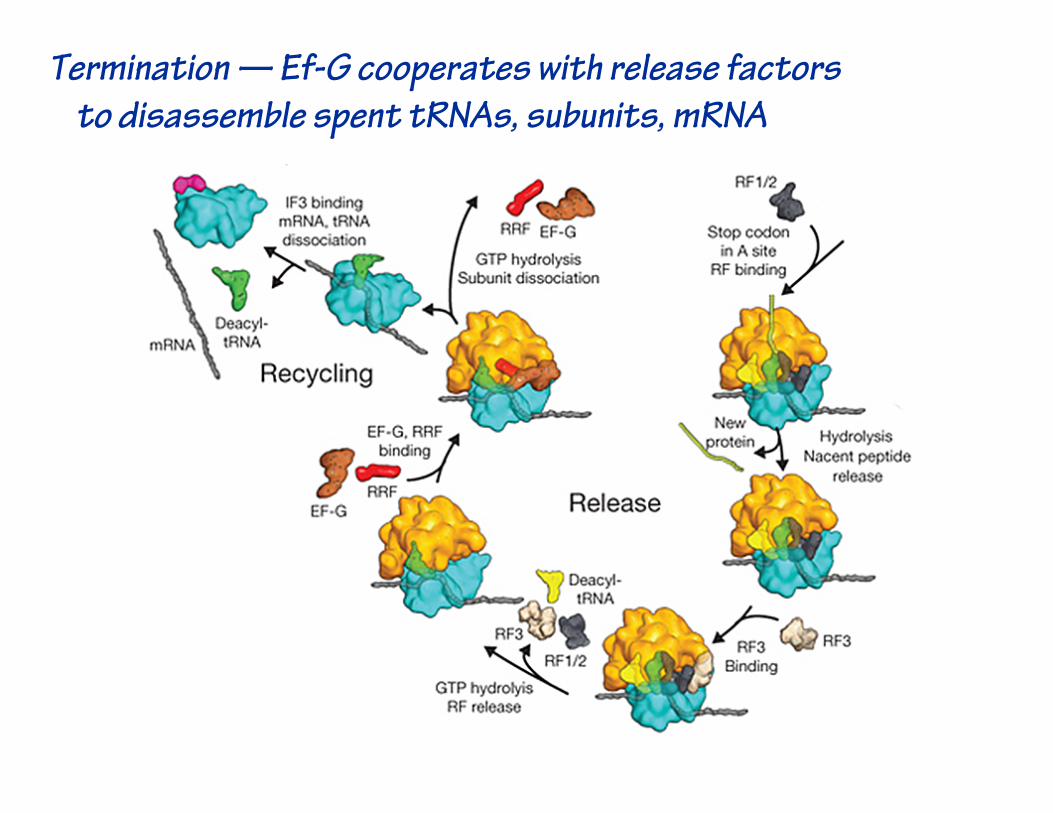
Termination — Ef-G cooperates with release factorsto disassemble spent tRNAs, subunits, mRNA





![Ribosome Stoichiometry: From Form to Function · Ribosome abundance: A major model, also termed the ribosome concentration hypothesis [3], that explains how ribosomes could exert](https://static.fdocuments.us/doc/165x107/60de31e56d30fc4fb30719b8/ribosome-stoichiometry-from-form-to-function-ribosome-abundance-a-major-model.jpg)













