
THE RELATIONSHIP BETWEEN DEVELOPMENT … RELATIONSHIP BETWEEN DEVELOPMENT OF THE BONY FALX AND BONY...
23
THE RELATIONSHIP BETWEEN DEVELOPMENT OF THE BONY FALX AND BONY TENTORIUM IN CETACEANS AND THEIR DIETS TAKAO NOJIMA 1 ) ABSTRACT The correlation between food habits and ossification in the falx cerebri (FC) and tentorium cerebelli (TC) of cetaceans is considered. I have observed the bony falx (BF) and bony tentorium (BT) in a variety of mammals, including cetaceans, and have classified them into the carnivore type and the dolphin type based on their developmental pattern. The carni- vore type develops as part of the skull bones during the fetal period, and these, attaining shapes closely similar to those of adults, are already com- pleted at birth. The dolphin type, on the other hand, is formed by ossifica- tion in the FC and TC during the course of aging. Among cetaceans, the dolphin type BF and BT are seen only in del- phinids and phocoenids. The carnivore type BF and BT are present in all the Ziphiidae, and in the sperm whale (Physeter catodon). In the other cetaceans observed, neither the BF nor BT was seen at all. I have reviewed many reports on food habits of cetaceans and noted that the cetaceans which possess the dolphin type BF and BT are fish-eaters (ichthyophagi). Many fishes contain a large amount of vitamin D which pro- motes ossification of bones, while squid and zooplankton, other main food of toothed whales and the main food of baleen whales, respectively, contain no vitamin D. It appears likely that formation of the dolphin type BF and BT in del- phinids and phocoenids is due to their continual intake of large amounts of fish high in vitamin D. Although this approach to assessing the food habits of cetaceans is indi- rect, it provides consistent results. The general trend of their food habits throughout life appears to be reflected in the degree of ossification in the FC and TC, much as the growth layers formed in teeth and ear plugs of ceta- ceans indicate their ages. Key words: bony falx, bony tentorium, ossification, vitamin D 1) 5-21-1, Ooto, Yono-shi, Saitama, 338 Japan Sci. Rep. Cetacean Res., No. 1, 1990, 39-61
Transcript of THE RELATIONSHIP BETWEEN DEVELOPMENT … RELATIONSHIP BETWEEN DEVELOPMENT OF THE BONY FALX AND BONY...

THE RELATIONSHIP BETWEEN DEVELOPMENT OF THE BONY FALX AND BONY TENTORIUM IN
CETACEANS AND THEIR DIETS
TAKAO NOJIMA 1)
ABSTRACT
The correlation between food habits and ossification in the falx cerebri (FC) and tentorium cerebelli (TC) of cetaceans is considered.
I have observed the bony falx (BF) and bony tentorium (BT) in a variety of mammals, including cetaceans, and have classified them into the carnivore type and the dolphin type based on their developmental pattern. The carnivore type develops as part of the skull bones during the fetal period, and these, attaining shapes closely similar to those of adults, are already completed at birth. The dolphin type, on the other hand, is formed by ossification in the FC and TC during the course of aging.
Among cetaceans, the dolphin type BF and BT are seen only in delphinids and phocoenids. The carnivore type BF and BT are present in all the Ziphiidae, and in the sperm whale (Physeter catodon). In the other cetaceans observed, neither the BF nor BT was seen at all.
I have reviewed many reports on food habits of cetaceans and noted that the cetaceans which possess the dolphin type BF and BT are fish-eaters (ichthyophagi). Many fishes contain a large amount of vitamin D which promotes ossification of bones, while squid and zooplankton, other main food of toothed whales and the main food of baleen whales, respectively, contain no vitamin D.
It appears likely that formation of the dolphin type BF and BT in delphinids and phocoenids is due to their continual intake of large amounts of fish high in vitamin D.
Although this approach to assessing the food habits of cetaceans is indirect, it provides consistent results. The general trend of their food habits throughout life appears to be reflected in the degree of ossification in the FC and TC, much as the growth layers formed in teeth and ear plugs of cetaceans indicate their ages.
Key words: bony falx, bony tentorium, ossification, vitamin D
1) 5-21-1, Ooto, Yono-shi, Saitama, 338 Japan
Sci. Rep. Cetacean Res., No. 1, 1990, 39-61

40 NOJIMA
INTRODUCTION
The brain of mammals is surrounded by three membranes, or meninges: the dura mater, the arachnoid mater and the pia mater. The dura mater is a dense, strong fibrous membrane, and is conventionally described as two layers: the outer (endosteal) and inner (meningeal) layers. The outer layer is the periosteum covering the inner surface of the skull bones. The inner layer is the dura mater proper, and sends four partitions inward, which divide up the cranial cavity into freely communicating spaces lodging the subdivisions of the brain. These partitions operate to restrict the rotatory displacement of the brain.
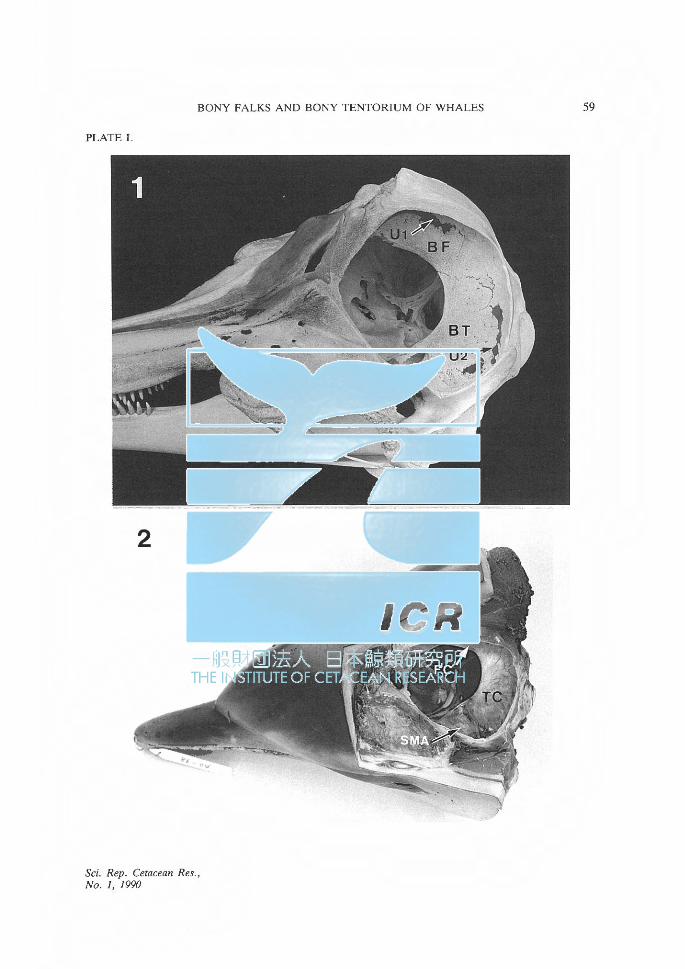
The falx cerebri (FC) is a vertical partition that lies in the longitudinal fissure between the cerebral hemispheres. The tentorium cerebelli (TC) is a partition that inserts between the upper surface of the cerebellum and the occipital lobes of the cerebral hemispheres, and forms a floor supporting the latter (Plate I, Fig. 2). In most mammals, both the FC and TC are composed of only the folded dura mater. In other mammals, a bony plate, developed to varying degrees, is present in the FC and/or TC. The plate in the FC is the bony falx (BF) and that in the TC is the bony tentorium (BT).
There are few studies on the BF and BT of any mammals. I have examined the skulls of 189 species in 54 families from 15 orders (but not Scandentia, Dermoptera, Hyracoidea, Tubulidentata and Macroscelidea) in the class Mammalia (following Corbet and Hill, 1986).
The BT and/or BF were observed in almost all species of Carnivora and in many species of Odontoceti (Cetacea), but were seen only in a few species belonging to orders such as Perissodactyla (Equidae), Sirenia, Primates (Cebidae, genus Ateles) and Marsupialia.
Based on their developmental pattern, I have classified the BF and BT of mammals into two types: the carnivore type and the dolphin type (Nojima, 1988).
The BF and BT of the carnivore type develop as part of the skull bones (the parietal bone and/or occipital bone) during the fetal period, and these attain shapes very similar to those of adults by the time of birth in almost all carnivores (BT, and also the BF in all pinnipeds) and in horses (BT), phalangers (Phalangeridae) (BT), kangaroos (Macropopidae) (BT), sperm whales (Physeteridae) (BF), beaked and bottlenose whales (Ziphiidae) (BF & BT) and sirenians (BF). Morphological characteristics of the BT and BF are useful in the classification of Carnivora (Nojima, 1990a).
On the other hand, the BF and BT of the dolphin type, for example those of spotted dolphins (Stenella attenuata), are not seen in the newborn or juvenile at all, but are gradually formed by ossification in the FC and TC as the animal grows older (Nojima, 1988, 1989). Among cetaceans, clearly formed dolphin type BF and BT are seen only in the Delphinidae and Phocoenidae. The BT of
Sci. Rep. Cetacean Res., No. 1, 1990

BONY FALKS AND BONY TENTORIUM OF WHALES 41
spider monkeys is also assigned to this type (Nojima, 1990b). The differences between the two types are not merely due to the difference
in timing of onset of their development. In mammals which possess the carnivore type BT and/or BF, the membranous portions of the TC and FC in which the BT and BF are absent, do not ossify throughout life (Nojima, 1990a). The functional significance of the carnivore type BT and BF is unknown.
In previous studies (Nojima, 1988, 1989), I have suggested that ossification in the FC and TC of delphinids and phocoenids may be due to the continual intake of a large amount of fish high in vitamin D and calcium.
Vitamins D2 ( ergocalciferol) and D3 ( cholecalciferol) operate to maintain calcium homeostasis in mammals after being converted to 25-hydroxyvitamin D. Both are obtained from food. Vitamin D3 is also obtained from exposure of skin sterols to ultraviolet radiation. Deficiencies are more likely to become a problem for housed animals which are not exposed to direct sunlight (Doyle, 1987). Vitamin D deficiencies affect bone development (rickets) in young, growing animals and cause osteomalacia in adults (see for example Coburn, Hartenbower and Norman, 1974; Delucca, 1974; Fraser, 1975; Doyle, 1987).
On the other hand, hypercalcemia, which causes hyperplasia of bones and extensive calcium deposition in soft tissue, especially kidneys and arteries, results from excessive use of the vitamin and calcium in the treatment of humans for rickets (Kure and Sakamoto, 1966; Ishii, 1973; Coburn et al., 1974; Delucca, 1974; Fraser, 1975; Nakayama, 1975; Kobayashi, 1989). According to Schultze and Lubosch (1935), the dura mater of humans may be ossified, although it is unknown whether the condition is caused by hypercalcemia.
Most mammalian food in nature do not contain enough vitamin D to cause toxicity. In fish-eating mammals such as certain pinnipeds and cetaceans, however, there is a danger that excessive intake of vitamin D could produce levels known to be toxic for many terrestrial mammals (Bouillon, Vankerkhove and De Moor, 1976; Keiver, Draper and Ronald, 1988a). Fish contain large amounts of vitamin D, especially in the liver, and are the only source of the vitamin for cetaceans.
Food habits of cetaceans have been studied mainly by the examination of stomach contents. However, with the exception of larger whales whose food habits have been studied extensively using samples obtained from the commercial whaling industry, sufficient data for most toothed whales are not available.
Among cetaceans, clearly ossified BF and BT in the FC and TC, respectively, are seen only in the Delphinidae and Phocoenidae which have been reported to ingest a large amount of fish. On the other hand, little or no ossification occurs in the FC and TC of other cetaceans reported to feed mainly on squid, zooplankton or other invertebrates that lack vitamin D.
Sci. Rep. Cetacean Res., No. 1, 1990

42 NOJIMA
RESULTS
The dolphin type BF and BT were observed in all the Delphinidae and Phocoenidae, and are probably present in the Monodontidae. The carnivore type BF and BT were observed in all the Ziphiidae. The carnivore type BF was also observed in the sperm whale (Physeter catodon). In other cetaceans studied, neither the BF nor BT was seen at all.
All cetacean species and the number of skulls studied, their main food, and the BF and BT types are given in Table 1.
TABLE 1. LIST OF CETACEANS EXAMINED, TYPE OF BF AND BT, AND MAJOR FOOD ITEMS1l.
Family and species Carnivore Dolphin Main food (Sample number in parentheses) type type
Platanistidae Pontoporia blainvillei (2) crustaceans, molluscs, Inia geoffrensis (1) and fish (low vitamin Platanista gangetica (3) D content)
Delphinidae Steno bredanensis ( 4) BF&BT fish & squid Sousa teuszii (1) BF&BT fish & squid Stenella attenuata (35) BF&BT fish Stenella coeruleoalba (12) BF&BT fish Stenella longirostris (3) BF&BT fish Delphinus delphis (4) BF&BT fish & squid Tursiops truncatus (6) BF&BT fish & squid Lissodelphis borealis (2) BF&BT fish & squid Lagenorhynchus obliquidens (4) BF&BT fish & squid Lagenorhynchus albirostris (2) BF&BT fish & squid Lagenorhynchus obscurus (1) BF&BT fish & squid Lagenodelphis hosei (1) BF&BT fish &squid Peponocephala electra (5) BF&BT squid & fish Pseudorca crassidens (2) BF&BT squid & fish Orcinus area (2) BF&BT fish & squid Grampus griseus (3) BF & BT (tiny) squid Globicephala macrorhynchus (4) BF&BT squid & fish Feresa attenuata ( 4) BF&BT squid & fish
Phocoenidae Phocoena phocoena (6) BF&BT fish & squid Phocoenoides dalli (9) BF&BT fish & squid Neophocaena phocaenoides (2) BF & BT (tiny) squid, shrimp, fish
Monodontidae Monodon monoceros (1) BF & BT (tiny) squid & fish
Physeteridae Physeter catodon (2) BF squid Kogia breviceps (2) squid Kogia simus (3) squid
Sci. Rep. Cetacean Res., No. 1, 1990

BONY FALKS AND BONY TENTORIUM OF WHALES
Ziphiidae Berardius bairdii (4) Ziphius cavirostris (7) Mesoplodon ginkgodens (2) Mesoplodon stejnegeri (2) Mesoplodon densirostris (1) Hyperoodon ampullatus (1)
Balaenidae Balaena glacialis (1)
Eschrichtiidae Eschrichtius robustus ( 1)
Balaenopteridae Balaenoptera borealis (l) Balaenoptera edeni (1) Balaenoptera acutorostrata (2) Megaptera novaeangliae (1)
BF&BT BF&BT BF&BT BF&BT BF&BT BF&BT
squid squid squid squid squid squid
zooplankton
amp hi pods
zooplankton zooplankton zooplankton zooplankton
43
1) Classification of cetaceans is basded on Corbet and Hill (1986). The skulls of cetaceans belonging to the collections of Department of Zoology, National Science Museum, Tokyo, were mainly studied. In many species, both adult and juvenile specimens were studied for comparison of the degree of development of the BF and BT. The growth stages (adult or juvenile) and other biological data in the text were copied from museum records. Besides these, a sperm whale of the Whale Museum, Ayukawa, Miyagi, a right whale of the Whale House, Kumomi, Shizuoka. Twenty-seven spotted dolphins including fetuses, five striped dolphins, and three bottlenose dolphins including a newborn are from my own collection.
Main food of each species is based on published reports. "fish & squid" or "squid & fish", respectively, means that the former appears to be more important than the latter.
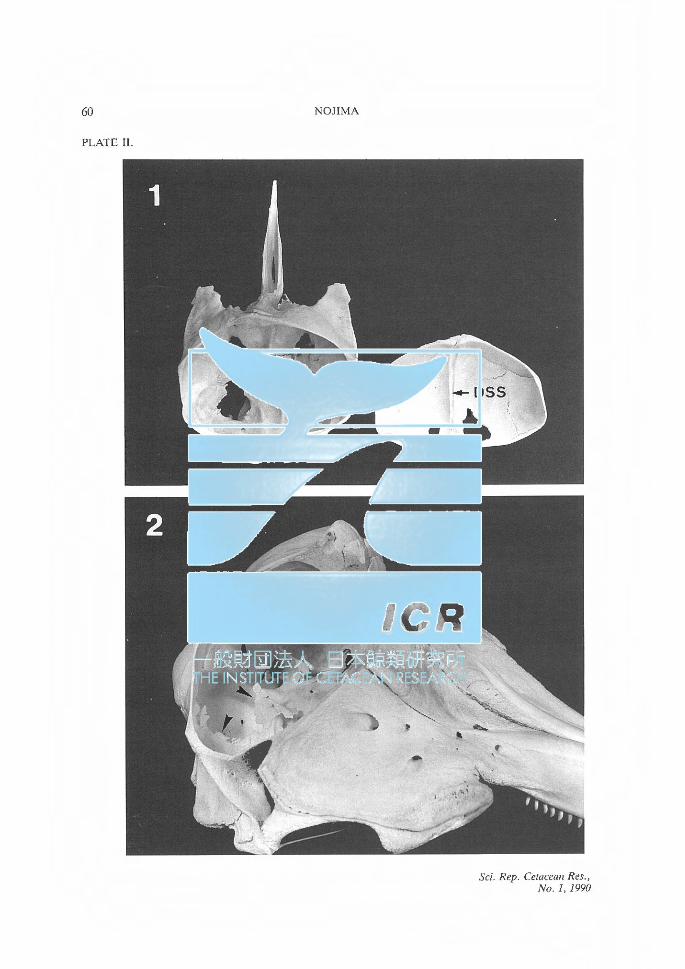
Delphinidae The BF and BT, which had been formed in varying degrees in the FC and TC, were observed in most delphinids. In most of the juvenile animals observed, the BF and BT had not been completed as a continuous bony plate (Plate II), but in some other juveniles, only a small bony process was seen to project from the portion corresponding to the internal occipital protuberance. The BF and BT were not seen at all in fetal or juvenile spotted dolphins (Plate II), or in a newborn bottlenose dolphin (Tursiops truncatus).
The Risso's dolphin (Grampus griseus) was a peculiar species. Even in adult specimens, only a small ossified process projected from the portion corresponding to the internal occipital protuberance.
In the skulls of adult spotted dolphins, in which the BF and BT had been completed (Plate I), the inner surface of the calvaria had been made rough by many laminations formed by ossification in the dura mater (Plate III).
As an example of a delphinid pattern, the order of ossification in the FC and TC of spotted dolphins is shown diagrammatically in Fig. 1.
Sci. Rep. Cetacean Res., No. 1, 1990

44 NOJIMA
Fig. 1. The order of ossification in the FC and TC of the spotted dolphin. The cranium has been cut along a line slightly lateral and parallel to the midline and the right half has been removed. The BT of the right side has also been cut along the same side of the midline, and the BF and the inferior surface of the left BT are shown.
During 6 months to 1 year of age, ossification begins at the attachment of the FC and TC to the internal occipital protuberance (1), and advances forward along the junction of the FC and TC. Then ossification spreads toward the back part of the FC and toward the anterior portion of the TC (2). At the same time, ossification occurring in the attachment of the TC to the base of the skull also advances upward in the anterior portion of the TC (2), and fuses with the above developing plate to form the anterior half of the BT. From this, ossification spreads backward in the TC (3). At the same time, ossification in the FC advances forward and completes the free margin of the BF (3). Finally, by about 9-11 years of age, the BF and BT are completed in the shape of the entire FC and TC, except for the passage of blood vessels (black arrows) and nerves (V). The most anterior portion of the free margin of the TC is ossified to form a small bony arch above the base of the skull (A). A shallow groove for the continuations of the spinal meningeal artery to the ophthalmic retial complex runs under the arch (see Plate I). The passage of the trigeminal nerve, which occurs at the most medial and inferior portion of the TC, is completed as a foramen on the BT in older specimens (V), but remains as a round notch at the free margin of the BT in younger specimens.
Phocoenidae In both habor and Dall's porpoises (Phocoena phocoena and Phocoenoides dalli), the BT had not been completed as a continuous bony plate to the base of the skull in any specimens. The BF was well developed in older specimens, and a round opening for the passage of the trigeminal nerve, which in the spotted dolphin is completed late during the process of ossification in the FC and TC (see Fig. 1), occurred at the partial BT projecting from the base of the skull. The BF and BT of adult porpoises observed appear to have been damaged during preparation of the skulls. In all young specimens, both the BF and BT were poorly developed.
Even in adult specimens of the finless porpoise (Neophocaena phocaenoides), only a small bony process projected from the portion which corresponded to the internal occipital protuberance.
Sci. Rep. Cetacean Res., No. l, 1990

BONY FALKS AND BONY TENTORIUM OF WHALES
Fig. 2. Schematic drawings of the BF, BT and TC of Ziphiidae; seen from left side slightly anteriorly, and from below (right side). BF: bony falx, BT: bony tentorium, TC: tentorium cerebelli, V: the passage of the trigeminal nerve, which occurs as a foramen or a notch. Attachments of the TC of Mesoplodon spp. are slightly ossified (black arrows).
Monodontidae
45
Only one specimen of adult male narwhal (Monodon monoceros) was examined. The BF and BT were not seen in the skull; however, the portion corresponding to the internal occipital protuberance was rough, probably reflecting damage to the ossified process.
Physeteridae Owen (1866) reported the partial BF projecting from the supraoccipital bone into the back part of the FC in an advanced fetal sperm whale. I also observed the partial BF in both young and adult sperm whales (12.19 m and 16.90 m in body length) at the site corresponding to Owen's descriptions. The BT was absent. The partial BF of sperm whales is assigned to the carnivore type.
Ziphiidae The BF, which appears to have the shape of the entire FC, and the partial BT which inserts into the posterior half of the TC, were seen in all the Ziphiidae, including both young and juvenile specimens (Fig. 2, left side). In the median sagittal section of an adult skull of Baird's beaked whale (Berardius bairdii), it was obvious that the element of the BF and BT was derived from the occipital bone (the supraoccipital bone). The BF and BT of Ziphiidae are assigned to the carnivore type, and characterize the skull of those animals.
In all the Mesoplodon species observed, the attachments of the TC were partially ossified (Fig. 2, right side), but not considered to be ossified in the entire shape of the TC.
Sci. Rep. Cetacean Res., No. 1, 1990

46 NOJIMA
DISCUSSION
Among prey items for wild mammals, some species of fish contain an exceptionally large amount of vitamin D concentrated in the liver. According to Kagawa (1989), vitamin D value in fish meat is fairly high on migratory fishes but is low in most bottom-dwelling fishes and freshwater fishes. Holmes, Tripp and Satterfield (1941) and Inagaki (1970) reported vitamin D value (JU/gram) in fish liver oil of some species (bluefin tuna Thunnus thynnuis, 27,000~70,000 IU; skipjack Euthynnus pelamis, 20,000 IU; mackerel Scomber japonicus, 5,400 IU; swordfish Tetrapturus audax, l,550~25,000 IU; halibut Hippoglossus hippoglossus, 550~20,000 IU). Although available data on vitamin D content in fish liver oil are scarce, this can estimate from the vitamin value in the meat. Vitamin D content in the liver oil tends to increase in parallel with the increase of the vitamin value in the meat. For example, the meat of dogfish Squalus acanthias and blue shark Prionace glauca contain no vitamin D, and liver oil made from these fishes only contain the vitamin of O~ 100 IU per gram. While the meat of bluefin tuna and skipjack, above species respectrively, contains about 600 IU and 420 IU per lOOg.
On the other hand, squid, zooplankton and the other aquatic invertebrates contain no vitamin D (Hirano, 1982; Kagawa, 1989).
In the following sections, the correlation between degree of ossification in the FC and TC and prey items of each group of cetaceans are discussed.
Mysticeti Although the minke whale (Balaenoptera acutorostrata), sei whale (B.borealis), Bryde's whale (B.edeni) and humpack whale (Megaptera novaeangliae) have been reported to ingest considerable amounts of fishes such as sardine, saury and mackerel (Mizue, 1951; Nemoto, 1957, 1959; Mitchell, 1975), in many areas the main food of these cetaceans is zooplankton such as euphausiids and copepods (Nemoto, 1957, 1959, 1962; Nemoto and Nasu, 1958; Nemoto and Yoo, 1970; Kawamura, 1970, 1973, 1977, 1980, 1982). The percentage of fish found in the stomachs of minke and sei whales caught south of 40°S was 0% and 0.1 % , respectively (Horwood, 1987), and those of sei and Bryde's whales caught in the North Pacific was 3.4% and 11.1%, respectively (Nemoto and Kawamura, 1977).
The gray whale (Eschrichtius robustus) feeds primarily on benthic amphipods, especially gammaridean amphipods (Pike, 1962; Nerini, 1984; Wolman 1989).
North Pacific right whale (Balaena glacialis) feeds primarily on copepods such as Ca/anus cristatus and C.plumchrus (Omura, 1958; Omura, Ohsumi, Nemoto, Nasu and Kasuya, 1969).
Sci. Rep. Cetacean Res., No. 1, 1990

BONY FALKS AND BONY TENTORIUM OF WHALES 47
No ossification in the FC and TC occurs in any baleen whales that are feeding mainly upon zooplankton with no vitamin D.
Physeteridae Many reports on the food of sperm whales have been presented (Pike, 1950; Mizue, 1951; Rice, 1963; Okutami and Nemoto, 1964; Roe, 1969; Kawakami, 1976; Clarke, 1980). Clarke (1956) examined a considerable number of stomachs of sperm whales in the Azores, and found eight species of squid which predominated in the stomachs and a small though not insignificant amount of fish. Kawakami (1980) reviewed the published reports on the food of sperm whales taken from different localities of the world. According to this, the percentage of squid and fish found in the stomachs obviously show that the sperm whales are primarily squid-eaters although in some localities their stomachs contained fairly high percentages of fish.
The pygmy sperm whale (Kogia breviceps) feeds mostly on cephalopods (Hale, 1947; Raun, Hoese and Moseley, 1970; Clarke, 1986; Pinedo, 1987). Stomachs of the dwarf sperm whale (Kogia simus) contained large amounts of cephalopods (Ross, 1978; Jones, 1981). Nagorsen and Stewart (1983) also found cephalopod beaks and shrimp in the stomach. On the other hand, Fitch and Brownell (1968) observed otoliths of many fish species (Ichthyococcus sp., Macrouridae, Moridae, Lampadena etc.) besides cephalopod beaks and crustacean remains in the stomachs of three pygmy sperm whales (?, scientific name given in the article is Kogia simus). However, only small amounts of fish appear to be eaten by the species.
In the sperm whale, pygmy sperm whale and dwarf sperm whale, all of which feed mainly on squid with no vitamin D, ossification in the FC and TC does not occur at all, although the carnivore type BF is present in the sperm whale.
Ziphiidae According to reviews on the food of the Ziphiidae by Gaskin (1976), Evans (1987), Balcomb (1989), Heyning (1989) and Mead (1989a,b), the Ziphiidae on the whole appear to be squid-eaters. Heyning (1989) listed many species of cephalopods found in the stomachs of Cuvier's beaked whales (Ziphius cavirostris). Baird's beaked whale (Berardius bairdii) and Cuvier's beaked whale also eat somewhat a considerable amount of deep-sea fishes (Rice, 1963; Nishiwaki and Oguro, 1971). Almost all deep-sea fishes, however, are distinctly low in vitamin D content.
Ross (1984) found 21 fish otoliths identified as species of the genera Cepola? (1), Scopelogadus (1) and Lampanyctus (19) in the stomach of a female adult Blainville's beaked whale (Mesoplodon densirostris) from southern Africa. Mead (1989b) found trace quantities of squid beaks from three Blainville's
Sci. Rep. Cetacean Res., No. 1, 1990

48 NOJIMA
beaked whales and two Stejneger's beaked whales (M.stejnegeri) but no fish remains were found in the stomachs. Concerning the ginkgo-toothed beaked whale (M.ginkgodens), another Mesoplodon species observed in the present study, no information on their food habits appears to be available.
A small amount of ossification in the attachments of the TC was seen in all the Mesoplodon species observed. Because of the scarcity of published data on the food habits of Mesoplodon spp., it is unknown whether these whales feed on a larger proportion of fish than do the Baird's beaked whale and Cuvier's beaked whales, in which ossification in the TC does not occur at all. Mead (1989b) supposes that Mesoplodon species prefer mesopelagic squid and fish.
Benjaminsen and Christensen (1979) examined the stomach contents of 46 bottlenose whales (Hyperoodon ampullatus) caught off Iceland and 108 caught off Labrador. They found that squid was the main food. Fishes such as cusk (Brosmius brosme), lumpsucker (Cyclopterus lumpus), redfish (Sebastes sp.) and halibut (Reinhardtius hippoglossoides) were also eaten by nearly 10% of the animals off Iceland and about 50% off Labrador, respectively. However, fish represented a small proportion of the total volume of stomach contents.
The conditions of the BT and BF of all the Ziphiidae observed indicate that these whales are primarily squid-eaters, though there remains a problem of some ossification in the attachments of the TC of the Mesoplodon species.
Platanistidae Reeves and Brownell (1989) reviewed feeding habits of the susu (Platanista). Among prey items found in the stomachs of susus, prawns, clams, catfishes (Wallago attu and Macrones aor), carp (Catla buchanami) and goby (Glossogobius giuris) appear to be their main food. In the close species to above fishes, the meat of carp (Cyprinus carpio) and catfish (Silurus asotus) contain no viamin D, and that of goby (Acanthogobius flavimanus) only contains the vitamin of 10 IU per 100g (Kagawa, 1989).
Boto (Inia geoffrensis) is known to feed on over 43 species of fish, especially on sciaenids, cichlids and characins (Best and da Silva, 1989). Leatherwood and Reeves (1983) and Evans (1987) mentioned that the dolphin also feeds on armored catfish, and perhaps molluscs and crustaceans. The boto observed in this study had been kept at a aquarium for two years and had been fed exclusively on the Japanese horse mackerel (Trachrus japonicus) (Tobayama and Kamiya, 1989) with low vitamin D content (Kagawa, 1989). Ossification in the FC and TC probably does not occur either in the wild botos that are feeding on prey items low in vitamin D content.
According to Brownell's (1989) review on prey items of franciscana (Pontoporia blainvellei), the dolphins feed on many species of fish and some species of squid and shrimp. Among a wide variety of fishes as food for the dolphins, the most important species was Cynoscion striatus (Sciaenidae) in the studies
Sci. Rep. Cetacean Res., No. 1, 1990

BONY FALKS AND BONY TENTORIUM OF WHALES 49
examined in southern Brazilian and northern Uruguayan waters. Penaeid shrimp of three species were found in the stomachs of dolphin. Squid beaks, Loligo sanpaulensis, were commonly found in the stomachs of dolphins, and the squid was one of the most important prey item for the dolphin in Bahia Samboromdon. The importance of squis as food for the dolphin appears to remain an open question. Although the vitamin D content in most species of sciaenids including the important species, C.striatus, and other bottom-dwelling fishes listed in the review are unknown, bottom-dwelling fishes on the whole are of low vitamin D content. The vitamin is not contained at all in fish meat of Ammodytes personatus, Platycephalus indicus, Paraplagusia japonica, Salangichthys microdon, Lampetra mitsukurii, Trachurus japonicus and Conger myriaster, and is contained only 8 ~ 20 IU per lOOg in fish meat of Acanthopagrus schlegeli, Acanthogobius fiavimanus and Sillago japonica (Kagawa, 1989).
Monodontidae According to a review of prey items on narwhals (Monodon monoceros) by Hay and Mansfield (1989), the narwhal mainly feeds on the Arctic cod (Boreogadus saida), the polar cod (Arctogadus glacialis), the Greenland halibut (Reinhardtius hippoglossoides), the squid (Gonatus fabricii) and the shrimps (Pasiphaea tarda and Hymenodora glacialis). Finley and Gibb (1982) found abundant squid beakes besides a number of cod (above spp.) in the stomachs of 73 nar\Vhals.
Among the fishes eaten by narwhals, the liver oil of halibut (Hippoglossus hippoglossus) contains a large amount of vitamin D, although the value vary according to seasons (550~20,000 IU/g). It is unknown whether the poorly ossified process (might be present) in the animal observed was caused by ingested halibuts.
Delphinidae The genus Stenella feeds mainly on a wide variety of epipelagic and mesopelagic fishes and a substantial amount of squid (Gaskin, 1982; Evans, 1987). Among fish species, myctophids predominated in stomachs of S.longirostris (Fitch and Brownell, 1968), S.coeruleoalba (Wilke, Taniwaki and Kuroda, 1953; Miyazaki, Kusaka and Nishiwaki, 1973) and S.clymene (Perrin, Mitchell, Mead, Caldwell and van Bree, 1981), and they were also important to S.graffmani (Fitch and Brownell, 1968). S.attenuata mainly feeds on epipelagic fishes such as flying fish (Mitchell, 1975).
In the skulls of elderly spotted dolphins, in which the BF and BT are complete, the inner surface of the calvaria is roughened by many laminations formed by ossification in the dura mater. This is considered to be caused by hyperplasia of the bones.
According to Evans (1987) and Mitchell (1975), the genera Delphinus,
Sci. Rep. Cetacean Res., No. 1, 1990

50 NOJIMA
Steno, Tursiops, Lagenorhynchus, Sousa, Lissodelphis and Lagenodelphis feed on a wide variety of epipelagic and mesopelagic fishes as well as squid and other molluscs.
The percentage of fish and squid in the stomachs of bottlenose dolphins and Pacific white-sided dolphins (Lagenorhynclus obliquidens) caught in waters adjacent to Kyushu, Japan, during 1968-1969, were reported by the Research Department Fisheries Agency, Japan (1969). Although stomach contents of catches varied a little according to different localities and different seasons, these dolphins had fed mainly on epipelagic and bottom-dwelling fishes, and the percentage of squid was only 15-30% in the bottlenose dolphin and less than 40% in the white-side dolphin.
According to Baker (1981), Torres and Aguayo (1979) found 1,261 lantern fish otoliths and 46 squid beaks in the stomach of one southern right whale dolphin (Lissodelphis peronii).
Nishiwaki and Randa (1958) reviewed the published reports of the stomach contents of 567 killer whales ( Orcinus area) caught in the coastal waters off Japan from 1948 to 1957. Although the killer whale eats other warmblooded animals, most stomachs of the animals contained a large amount of fishes such as cod, flatfish, sardine, salmon and tuna as well as squid. In the younger animals, the intake ratio of fish was higher than those of adults. Macintyre (1934) described that the killer whale is an important predator of the salmon.
Nishiwaki (1965) described that the captive Risso's dolphin showed little interest in any food except squid. According to Evans (1987), squid and octopus are main foods of the Risso's dolphin, but fish is occasionally taken. No fish was found in any Risso's dolphins in a study in Washington State (Stroud, 1968). Poorly ossified BF and BT of the Risso's dolphins observed also suggest that this species is primarily a squid-eater.
On the whole, the correlation between the degrees of ossification in the FC and TC and known food items of cetaceans agrees with the hypothesis in this study. However, there are several species in which the data do not support the hypothesis, although the details of their feeding habits are not well known.
Pilot whale generally prefer squid. Sergeant (1962) observed the longfinned pilot whale (Globicephala melaena) feeding on short-finned squid (Illex illecebrosus) almost exclusively during the July-October interval at Trinity Bay, Newfoundland; cod was only taken when squid was scarce. He pointed out a close relationship between the distribution of the pilot whale population off the east coast of Newfoundland and that of the squid. He also quoted Kritzler's (1952) observations that a young captive short-finned pilot whale ( Globicephala macrorhynchus) had refused to feed on fish at first, although it eventually learned to take them. Sergeant (1962) concluded that squid is the preferred food of pilot whales, with fish as an alternative.
Sci. Rep. Cetacean Res., No. 1, 1990

BONY FALKS AND BONY TENTORIUM OF WHALES 51
Keiver, Ronald and Draper (1988b) showed that plasma levels of 25-0HD, 24,25-(0H)2D and calcium in the long-finned pilot whales (stranded, place and season are not mentioned) were similar to those of nonmarine mammals, while in the white whale (Delphinapterus leucas) and bottlenose dolphin (feeding mainly on fish), levels of 25-0HD and 24,25-(0H)2D were extremely high. The results suggest that the long-finned pilot whale had fed mainly on squid (at least during the time immediately preceding the stranding) and indicate high plasma levels of vitamin D in fish-eating dolphins.
In this study, the long-finned pilot whale was not studied. If the animals are truly squid-eaters, both the BF and BT should be absent according to my hypothesis. However, the BF and BT of the short-finned pilot whale, another Globicephala species, and those of the false killer whale (Pseudorca crassidens) and pygmy killer whale (Feresa attenuata), were ossified in the shape of the entire FC and TC. The stomach of a false killer whale caught off western Japan contained 12 common sea basses Lateolabrax japonicus (about 60 cm in length) as well as many squid beaks (the Research Department Fisheries Agency, Japan, 1969). These whales may ingest much larger amounts of fish than is generally believed.
Phocoenidae In the finless porpoise which feeds on small squid, cuttlefish and shrimp but also takes small sandeels and other fishes (Mitchell et al., 1975), ossification in the FC and TC (even in a lactating adult) has occurred much less frequently than in other phocoenids. Poorly developed BF and BT in these porpoises suggest that they feed mainly on crustaceans and molluscs or that fishes eaten have a low vitamin D content. The meat of sandeel Ammodytes personatus contains no vitamin D (Kagawa, 1989).
The harbor and Dall's porpoises feed on a large number of fish species. Rae (1965) reviewed food habits of the harbor porpoise reported in an earlier time in the North Sea, and more recently in the Baltic Sea (Lindroth, 1962), off eastern Canada (Sergeant and Fisher, 1957) and off Scotland by Rae himself. The stomach contents showed that the harbor porpoise feeds mainly on fishes such as clupeoids, gadoids and pleuronectids such as herring, whiting and mackerel, but squid were occasionally found in the stomachs in some quantity. Smith and Gaskin (1974) examined stomach contents of P.phocoena taken from coastal waters of New Brunswick, Nova Scotia, and Prince Edward Island during 1969-1972. The animals fed mainly on fishes such as herring, gadoids and mackerel, but squid was occasionally taken. Wilke and Nicholson (1958) examined the stomachs of 86 truei-type and seven dalli-type Dall's porpoises (Phocoenoides dalli) collected off Japan, March to June 1952. Various myctophids formed 70%, by volume, of the stomach contents in the True's porpoises and 73% in the Dall's porpoises; the percentages of squid were 11 and
Sci. Rep. Cetacean Res., No. I, 1990

52 NOJIMA
5%, respectively. Wilke et al. (1953) found that squid formed 98%, by volume, of the stomach contents in four Dall's porpoises taken from March to May off northern Japan. This result contradicts Cowan's (1944) statement that Dall's porpoise feeds exclusively on herring in the northern North Pacific.
In a previous study (Nojima, 1988) of the BF and BT of spotted dolphins, I prepared the skulls very carefully so as not to damage the BF and BT. When ossification of the BF and BT has not been completed, these are very fragile, especially the BT. It is likely that the BF and BT of all harbor and Dall's porpoises observed had been damaged during preparation of the skulls. I observed no specimen in which the BT was complete, but the BF was relatively well developed and the passage of the trigeminal nerve occurred as a round foramen in most older specimens.
These porpoises probably ingest a large amount of fish, as has been reported.
Consumption of Vitamin D by Fish-eating Dolphins In the following section, the amounts of vitamin D that are taken by fish-eating dolphins are estimated.
Sergeant (1969) examined feeding rates in captivity of nine dolphin species. He estimated that of bottlenose dolphins to be 4.2 % body weight. Nakajima (1973) also reported the average volume of daily food of a 300 kg captive bottlenose dolphin as about 15 kg (mackerel Scomber japonicus).
The liver of a 410 g mackerel (Scomber japonicus) weighs about 10 g. If a 10 g mackerel liver is estimated conservatively to yield 1 g liver-oil (the average liver-oil content of cod liver is more than 50%, by weight: Omura, Matsuura and Miyazaki, 1942; although the values for fish vary according to season: Pugsley, 1939), total vitamin Din 15 kg of mackerel can be estimated as at least 230,000 IU. This amount corresponds to about 120 times the recommended daily dose for a human in weight-specific intake. With the exception of 1 a-OH-D and 1 a, 25-(0H)iD, the active form of vitamin D used to treat humans for vitamin deficiencies, toxicity rarely results from an excessive intake of vitamins D2 and D3 contained in food. However, a repeated intake of more than 100,000 IU per day of these vitamins may cause toxicity in humans (Krupp and Chatton, 1977; Kobayashi, 1989). Although the estimated vitamin D values taken by the dolphin do not approach the above dangerous level for humans in weightspecific intake, such large values cannot be ignored. Further consideration is needed to extend this calculation to the wild dolphins, because of uncertainties in the feeding rate (probably higher in the wild) and the consumption of variable food items with lower vitamin D values such as squids and many fish species other than mackerel.
Keiver et al. (1988a) studied vitamin D metabolism in hooded seals (Cystophora cristata), which ingest large amounts of fish, as do dolphins of the genus
Sci. Rep. Cetacean Res., No. 1, 1990

BONY FALKS AND BONY TENTORIUM OF WHALES 53
Stenella. They concluded that rapid conversion of vitamin D and a high capacity for vitamin D storage in the large blubber mass appear to be factors in the resistance of seals to vitamin D toxicity. It is unknown whether similar factors operate in the fish-eating cetaceans.
Concluding Remarks Besides delphinids and phocoenids, I have observed the dolphin type BT only in spider monkeys (Ateles geoffroyi and A.paniscus) among all mammals studied, and in those species the BF was absent although the FC was identical with the TC histologically. The factors involved in ossification only in the TC are unknown. Most of the animals studied had been born and bred in a zoo and they had been fed the usual diets, with no special items high in vitamin D. Formation of the dolphin type BT in spider monkeys cannot be related to their diet (Nojima, 1990b), and it obviously differs from those of delphinids and phocoenids in which ossification occurs in both the FC and TC.
Although I have not systematically examined hyperplasia of bones and calcium deposition in soft tissue (which may occur in the fish-eating mammals), the degree of ossification in the FC and TC of cetaceans obviously proceeds in parallel with the increased proportion of fish in the diet.
Stomach contents provide direct evidence of the food eaten by an animal. However, they do not necessarily reveal much about its longterm diet.
Although many cetaceans have been described as opportunistic feeders, the degree of ossification in their FC and TC appears to reflect the general trend of their food habits throughout life in the same way that the growth layers formed in the teeth and ear plugs of cetaceans indicate their ages. A comparison of the degrees of ossification in the FC and TC between captive dolphins fed mainly squid and others fed mainly fish would provide a further test of this hypothesis.
Recommended Preparation Procedures As already mentioned in the text, the BF and BT of delphinids and phocoenids are fragile during preparation, especially when ossification has not been completed in the shape of the entire FC and TC. Therefore, close attention should be paid to preparation of the skulls. Boiling the skull is not recommended because the incomplete BF and BT are often fractured owing to strong contraction of the FC and TC by heat. Furthermore, the BF and BT are easily damaged when the brain and its membranes are extracted through the foramen magnum. In order to keep the BF and BT complete during preparation of the skulls, it is best to ensure that the head (after removing the soft tissue as completely as possible) is submerged in fresh water for several months, or until the brain and its membranes have completely rotted and can be washed away.
Sci. Rep. Cetacean Res., No. I, 1990

54 NOJIMA
ACKNOWLEDGMENTS
I wish to express sincere gratitude to Dr N. Miyazaki and Dr M. Yoshiyuki, Department of Zoology, National Science Museum, Tokyo, for giving me access to the valuable collections, and to Dr M. Nakajima, Izu-Mito Sea Paradise, Shizuoka, who supplied me with materials and useful information. My sincere thanks are also due to Dr K. Shigemasa, Department of Anatomy, Nihon University School of Dentistry, Tokyo, who kindly helped me to collect references. I particulary thank Dr T. Kasuya and two anonymous reviewers for their comments on the manuscript. One of the two reviewers is greatly acknowledged for his kindly check of English style and grammar. I want to add a few words of gratitude to my wife Miwa for her help in all respects throughout this study.
REFERENCES
BAKER, A. N. 1981. The southern right whale dolphin Lissodelphis peronii (Lacepede) in Australasian waters. Nat. Mus. N.Z. Ree. 2(4): 17-34.
BALCOMB, K. C. 1989. Baird's beaked whale Berardius bairdii Stejneger, 1883: Arnoux's beaked whale Berardius arnuxii Duvernoy, 1851. pp. 261-288. In: S.H. Ridgway and R.J. Harrison (eds) Handbook of Marine Mammals, vol. 4. Academic Press, London. 442pp.
BENJAMINSEN, T. and CHRISTENSEN, I. 1979. The natural history of the bottlenose whale, Hyperoodon ampullatus (Forster). pp. 143-164. In: H.E. Winn and B.L. Olla (eds) Behavior of Marine Animals, vol. 3: Cetaceans. Plenum Press, New York. 438pp.
BEST, R.C. and da SILVA, V.M.F. 1989. Amazon river dolphin, boto Inia geoffrensis (de Blainville, 1817). pp. 1-23. In: S.H. Ridgway and R.J. Harrison (eds) Handbook of Marine Mammals, vol. 4. Academic Press, London. 442pp.
BOUILLON, R., VANKERKHOVE, P. and De MooR, P. 1976. Measurement of 25-hydroxyvitamin D3 in serum. Clin. Chem. (Winston-Salem, NC) 22: 364-368.
BROWNELL, R. L. 1989. Franciscana Pontoporia blainvillei (Gervais and d'Orbigny, 1844). pp.45-67. In: S.H. Ridgway and R.J. Harrison (eds) Handbook of Marine Mammals, vol. 4. Academic Press, London. 442pp.
CLARKE, R. 1956. Sperm whales of the Azores. Discovery Rep. 28: 237-298. CLARKE, M. R. 1980. Cephalopoda in the diet of sperm whales of the southern hemisphere and their bearing
on sperm whale biology. Discovery Rep. 37: 1-324. CLARKE, M. R. 1986. Cephalopods in the diet of odontocetes. pp.281-321. In: M. M. Bryden and R.J. Harri
son (eds) Research on Dolphins. Clarendon Press, Oxford. 478pp. COBURN, J. W., HARTENBOWER, D.L. and NORMAN, A. w. 1974. Metabolism and action of the hormone vita
min D: Its relation to diseases of calcium homeostasis. Western J. Med. 121: 22. CORBET, G. B. and HILL, J.E. 1986. A World List of Mammalian Species. Facts On File Publications, New
York. 254pp. Cow AN, I. M. 1944. The Dall porpoise, Phocoenoides dalli (True) of the northern Pacific Ocean. J. Mammal.
25: 295-306. DELUCCA, H.F. 1974. Vitamin D: 1973. Am. J. Med. 57: 1. DoYLE, P. T. 1987. Supplements other than forages. pp.429-464. In: J.B. Hacker and J.H. Ternouth (eds)
The Nutrition of Herbivores. Academic Press, Sydney. 552pp. EVANS, P. G. H. 1987. The Natural History of Whales and Dolphins. Christopher Helm, London. 343pp. FINLEY, K. J. and GrnB, E. J. 1982. Summer diet of the narwhal (Monodon monoceros) in Pond Inlet, North
ern Baffin Island. Can. J. Zoo!. 60: 3353-3363.
Sci. Rep. Cetacean Res., No. I, 1990

BONY FALKS AND BONY TENTORIUM OF WHALES 55
FITCH, J.E. and BROWNELL, R. L. 1968. Fish otoliths in cetacean stomachs and their importance in interpreting feeding habits. J. Fish. Res. Bd Can. 25(12): 2561-2574.
FITCH, J.E. and BROWNELL, R. L. 1971. Food habits of the franciscana Pontoporia blainvillei (Cetacea: Platanistidae) from South America. Bull. Mar. Sci. 21(2): 626-636.
FRASER, D.R. 1975. Advances in the knowledge of metabolism of vitamin D. Proc. Nutr. Soc. 34: 139. GASKIN, D.E. 1976. The evolution, zoogeography and ecology of Cetacea. Oceanogr. Mar. Biol. Ann. Rev.
14: 247-346. GASKIN, D.E. 1982. The Ecology of Whales and Dolphins. Heinemann Educational Books Ltd., London.
459pp. HALE, H.M. 1947. The pigmy sperm whale (Kogia breviceps, Blainvelle) on South Australian coasts. Ree. S.
Aust. Mus. 8: 531-546. HAY, K. A. and MANSFIELD, A. W. 1989. Narwhal Monodon monoceros Linnaeus, 1758. pp.145-176. Jn: S.H.
Ridgway and R.J. Harrison (eds) Handbook of Marine Mammals, vol. 4. Academic Press, London. 442pp.
HEYNING, J.E. 1989. Cuvier's beaked whale Ziphius cavirostris G.Cuvier, 1823. pp.289-308. In: S.H. Ridgway and R.J. Harisson (eds) Handbook of Marine Mammals, vol. 4. Academic Press, London. 442pp.
HIRANO, T. 1982. Vitamin. p.251. In: Editorial Committee of the 8th Open Lecture of Tokyo Fishing College (ed.) Livelihood and fish. Seizando, Tokyo. 381pp. (in Japanese).
HoLMES, A. D., TRIPP, F. and SATTERFIELD, G. H. 1941. Fish liver and body oils: Chemical characteristics, physical properties, and vitamin content. Ind. Eng. Chem. 33: 944-949.
HORWOOD, J. W. 1987. The sei whale: Population Biology, Ecology and Management. Croom Helm, London, New York and Sydney. 375pp.
INAGAKI, N. 1970. Vitamin. pp.36-43. In: H. Toshikawa (ed.) Handbook of nutrition (Popular Medicine, special issue 1). Nihonhyoronsha, Tokyo. 180pp. (in Japanese).
ISHII, M. 1973. Dictionary of Nutrition pp.262-263. Ishiyaku Shuppan, Tokyo. 458pp. (in Japanese). JoNES, R. E. 1981. Food habits of smaller marine mammals from northern California. Proc. Calif. Acad. Sci.
42: 409-433. KAGAWA, A. (ed.) 1989. Standard Tables of Food Composition in Japan (by Resources Council, Science and
Technology Agency, Japan). Kagawa Nutrition College, Tokyo 368pp. KAWAKAMI, T. 1976. Squids found in the stomach of sperm whales in the northwestern Pacific. Sci. Rep.
Whales Res. Inst., Tokyo 28: 145-151. KAWAKAMI, T. 1980. A review of sperm whale food. Sci. Rep. Whales Res. Inst., Tokyo 32: 199-218. KAWAMURA, A. 1970. Food of sei whale taken by Japanese whaling expeditions in the Antarctic season 1967/
68. Sci. Rep. Whales Res. Inst., Tokyo 22: 127-152. KAWAMURA, A. 1973. Food and feeding of sei whale caught in the waters south of 40° Nin the North Pacific.
Sci. Rep. Whales Res. Inst., Tokyo 25: 219-236. KAWAMURA, A. 1977. On the food of Bryde's whales caught in the South Pacific and Indian Oceans. Sci. Rep.
Whales Res. Inst., Tokyo 29: 49-58. KAWAMURA, A. 1980. A review of food of balaenopterid whales. Sci. Rep. Whales Res. Inst., Tokyo 32: 155-
197. KAWAMURA, A. 1982. Food habits and prey distributions of three rorqual species in the North Pcific Ocean.
Sci. Rep. Whales Res. Inst., Tokyo 34: 59-91. KEIVER, K. M., DRAPER, H. H. and RONALD, K. 1988a. Vitamin D metabolism in the hooded seal ( Cystophora
cristata). J. Nutr. 118: 332-341. KEIVER, K. M., RoNALD, K. and DRAPER, H. H. 1988b. Plasma levels of vitamin D and some metabolites in
marine mammals. Can. J. Zoo/. 66: 1297-1300. KOBAYASHI, T. 1989. Vitamin D. pp. 31-48. In: The Vitamin society of Japan (ed.) Vitamin Handbook. vol.
1, Kagakudojin, Kyoto. 127pp. (in Japanese). KRITZLER, H. 1952. Observations on the pilot whale in captivity. J. Mammal. 33: 321-334. KRUPP, M.A. and CHATTON, M.J. (eds) 1977. Current Medical Diagnosis and Treatment. Lange Medical
Publications, California. Japanese translation published by Maruzen, Tokyo in 1977. 1071pp. KuRE, K. and SAKAMOTO, T. 1966. Avitaminosis D. pp.52-57. In: S. Okinaka (ed.) Internal Medicine. vol. 2,
Nanzando, Tokyo. 818pp. (in Japanese).
Sci. Rep. Cetacean Res., No. 1, 1990

56 NOJIMA
LEATHERWOOD, S. and REEVES, R. 1983. Sierra Club Handbook of Whales and Dolphins. Sierra Club Books, San Francisco. 302pp.
LINDROTH, A. 1962. Baltic salmon fluctuations 2: Porpoise and salmon. Rep. Inst. Freshwat. Res. Drottningholm. 44: 105-112.
MACINTYRE, D. 1934. Sea enemies of salmon. Salm. Trout Mag. 74: 38-42. MATSUURA, Y. 1942. On the Northern Porpoise Whale, Berardius bairdii Stejneger in the adjacent waters of
"Bosyu". Zoo[ Mag., Tokyo 54: 466-473. (in Japanese). MEAD, J. G. and PAYNE, R. S. 1975. A specimen of the Tasman beaked whale, Tasmacetus shepherdi, from
Argentina. J. Mammal. 56: 213-218. MEAD, J. G. 1981. First records of Mesoplodon hectori (Ziphiidae) from the northern hemisphere and a
description of the adult male. J. Mammal. 62: 430-432. MEAD, J.G. 1989a. Bottlenose whales Hyperoodon ampullatus (Forster, 1770) and Hyperoodon planifrons
Flower, 1882. pp.321-348. In: S.H. Ridgway and R.J. Harrison (eds) Handbook of Marine Mammals, vol. 4. Academic Press, London. 442pp.
MEAD, J. G. 1989b. Beaked whales of the genus Mesoplodon. pp.349-430. In: S.H. Ridgway and R.J. Harrison (eds) Handbook of Marine Mammals, vol. 4. Academic Press, London. 442pp.
MITCHELL, E. (ed.) 1975. Review of biology and fisheries for small cetaceans. J. Fish. Res. Bd Can. 32: 889-983.
MIYAZAKI, N., KusAKA, T. and N1sHIWAKI, M. 1973. Food of Stenella coeruleoalba. Sci. Rep. Whales Res. Inst., Tokyo 25: 265-275.
M1zuE, K. 1951. Food of whales. Sci. Rep. Whales Res. Inst., Tokyo 5: 81-90. NAGORSEN, D. W. and STEWART, G. E. 1983. A dwarf sperm whale (Kogia simus) from the Pacific coast of
Canada. J. Mammal. 64(3): 505-506. NAKAJIMA, M. 1973. A dolphin named Echo. Akaneshobo, Tokyo. 146pp. (in Japanese). NAKAYAMA, K. 1975. Dose. pp.650-661. In: K. Nakayama (ed.) Pediatrics. Bunkodo, Tokyo 688pp. (in
Japanese). NEMOTO, T. 1957. Food of baleen whales in the northern Pacific. Sci. Rep. Whales Res. Inst., Tokyo 12:
33-89. NEMOTO, T. 1959. Food of baleen whales with reference to whale movements. Sci. Rep. Whales Res. Inst.,
Tokyo 14: 149-290. NEMOTO, T. 1962. Food of baleen whales collected in recent Japanese Antarctic whaling expeditions. Sci. Rep.
Whales Res. Inst., Tokyo 16: 80-103. NEMOTO, T. and NAsu, K. 1958. Thysanoessa macrura as a food of baleen whales in the Antarctic. Sci. Rep.
Whales Res. Inst., Tokyo 13: 193-199. NEMOTO, T. and Yoo, K. 1970. An amphipod, (Parathemisto gaudichaudii) as a food of the Antarctic sei
whale. Sci. Rep. Whales Res. Inst., Tokyo 22: 153-158. NEMOTO, T. and KAWAMURA, A. 1977. Characteristics of food habits and distribution of baleen whales with
special reference to the abundance of North Pacific sei and Bryde's whales. Rep. int. Whal. Commn (special issue 1): 80-87.
NERINI, M. 1984. A review of gray whale feeding ecology. pp.423-450. In: M.L. Jones, S.L. Swartz and S. Leatherwood (eds) The gray whale Eschrichtius robustus. Academic Press, Florida. 600pp.
NisHIWAKI, M. and RANDA, C. 1958. Killer whales caught in the coastal waters off Japan for recent 10 years. Sci. Rep. Whales Res. Inst., Tokyo 13: 85-96.
N1sHIWAKI, M. 1963. Taxonomical consideration on genera of Delphinidae. Sci. Rep. Whales Res. Inst., Tokyo 17: 93-103.
NrsHIWAKI, M. 1965. Whales and Pinnipeds. University of Tokyo Press, Tokyo. 439pp. (in Japanese). NrsHIWAKI, M. and OGURO, N. 1971. Baird's beaked whales caught on the coast of Japan in recent 10 years.
Sci. Rep. Whales Res. Inst., Tokyo 23: 111-122. NonMA, T. 1988. Developmental pattern of the bony falx and bony tentorium of spotted dolphins (Stenella
attenuata) and the relationship between degree of development and age. Mar. Mammal. Sci. 4: 313-322.
NonMA, T. 1989. Mairuka-ka no Kotsusei-dainokama to Kotsusei-shonotento no Keisei ni tsuite. [On the formation of the bony falx and bony tentorium of Delphinidae]. Geiken Tsushin, Whales Res. Inst., Tokyo 375: 9-14. (in Japanese).
Sci. Rep. Cetacean Res., No. 1, 1990

BONY FALKS AND BONY TENTORIUM OF WHALES 57
NoJIMA, T. 1990a. A morphological consideration of the relationships of Pinnipedia to other carnivorans based on the bony tentorium and bony falx. Mar. Mamm. Sci. 6: 54-74.
NonMA, T. 1990b. Developmental pattern of the bony tentorium of spider monkeys (Ateles). Primates 31: 137-141.
0KUTANI, T. and NEMOTO, T. 1964. Squids as the food of sperm whales in the Bering Sea and Alaskan Gulf. Sci. Rep. Whales Res. Inst., Tokyo 18: 111-122.
OMURA, H., MATSUURA, Y. and MIYAZAKI, I. 1942. [Whales - their science and practice of whaling]. Suisansha, Tokyo. 319pp. (in Japanese).
OMURA, H. 1958. North Pacific right whale. Sci. Rep. Whale Res. Inst., Tokyo 13: 1-52. OMURA, H., 0HSUMI, S., NEMOTO, T., NAsu, K. and KASUYA, T. 1969. Black right whales in the North Pacific.
Sci. Rep. Whales Res. Inst., Tokyo 21: 1-78. OWEN, R. 1866. On the Anatomy of Vertebrates. vol. 3, Reprinted in 1973, AMS Press, New York. 915pp. PERRIN, W.F., WARNER, R.R., Fiscus, C.H. and HOLTS, D.B. 1973. Stomach contents of porpoise, Stenella
spp., and yellowfin tuna, Thunnus albacares, in mixed-species aggregations. Fish. Bull. 71: 1077-1092. PERRIN, W. F., MITCHELL, E. D., MEAD, J. G., CALDWELL, D. K. and van BREE, P. J. H. 1981. Stenella clymene,
a rediscovered tropical dolphin of the Atlantic. J. Mammal. 62(3): 583-598. PIKE, G. C. 1950. Stomach contents of whales caught off the coast of British Columbia. Prag. Rep. Pacif. Cst.
Stns. 83: 27-28. PIKE, G. C. 1962. Migration and feeding of the gray whale (Eschrichtius gibbosus). J. Fish. Res. Bd Can.
19(5): 815-838. PINEDO, M. C. 1987. First record of a dwarf sperm whale from southwest Atlantic, with reference to osteology,
food habits and reproduction. Sci. Rep. Whales Res. Inst., Tokyo 38: 171-186. PUGSLEY, L. I. 1939. Vitamin A and D potencies of liver and intestinal oils of halibut. J. Fish. Res. Bd Can. 4:
396-404. RAE, B.B. 1965. The food of the common porpoise (Phocaena phocaena). J. Zool. 146: 114-122. RAUN, G.G., HOESE, H.D. and MosELEY, F. 1970. Pygmy sperm whales, genus Kogia, on the Texas coast.
Tex. J. Sci. 21: 269-274. REEVES, R.R. and BROWNELL, R. L. 1989. Susu Platanista gangetica (Roxburgh, 1801) and Platanista minor
Owen, 1853. pp.69-99. In: S.H. Ridgway and R.J. Harrison (eds) Handbook of Marine Mammals, vol. 4. Academic Press, London. 442pp.
Research Department Fisheries Agency, Japan, 1969. [Reports on investigation of fishery damage by small toothed whales in western waters of Japan during 1967-68.] 108pp. (in Japanese).
RICE, D. W. 1963. Progress report on biological studies of the larger Cetacea in the waters off California. Norsk Hvalfangst-Tid. 52(7): 181-187.
RoE, H.S.J. 1969. The food and feeding habits of the sperm whales (Physeter catodon, L) taken off the west coast of Iceland. J. Cons. Int. Explor. Mer. 33: 93-102.
Ross, G.J.B. 1978. Records of pygmy and dwarf sperm whales, genus Kogia from Southern Africa, with biological notes and some comparisons. Cape Prov. Mus., Nat. Hist. 11: 259-327.
Ross, G.J.B. 1984. The smaller cetaceans of the south east coast of southern Africa. Ann. Cape. Prov. Mus., Nat. Hist. 15: 173-410.
SCHULTZE, 0. and LuBoscH, W. 1935. Atlas und kurzgefasstes Lehrbbuch der topographischen und angewandten Anatomie. (Japanese translation by Nishi, S., 1941. Marzen Co., Tokyo, 353pp.)
SERGEANT, D.E. and FISHER, H.D. 1957. The smaller cetacea of eastern Canadian waters. J. Fish. Res. Bd Can. 14: 83-115.
SERGEANT, D. E. 1962. The biology of the pilot or pot-head whale Globicephala melaena (Traill) in Newfoundland waters. Bull. Fish. Res. Bd Can. 131: 1-84.
SERGEANT, D. E. 1969. Feeding rates of Cetacea. FiskDir. Skr. Ser. HavUnders. 15: 246-258. SMITH, G.J.D. and GASKIN, D.E. 1974. The diet of harbor porpoise (Phocoena phocoena (L.)) in coastal
waters of Eastern Canada, with special reference to the Bay of Fundy. Can. J. Zoo!. 52: 777-782. STROUD, R. K. 1968. Risso dolphin in Washington State. J. Mammal. 49: 347-348. ToBAYAMA T. and KAMIYA, T. 1989. Observations on Inia geoffrensis and Platanista gangetica in captivity at
Kamogawa Sea World, Japan. pp.42-45. In: W.F. Perrin, R.L. Brownell, Jr., Zhou Keiya and Lin Jiankang (eds) Biology and conservation of the river dolphins. (IUCN Species Survival Commission Occasional Papers No.3) IUCN Publications Services Unit, Cambreidge. vi + 173pp.
Sci. Rep. Cetacean Res., No. 1, 1990

58 NOJIMA
TORRES, D. N. and AGUAYO, L. A. 1979. Habitors alimentaries de Lissodelphis peronii en Chile central (Cetacea: Delphinidae). Revista de Biologia Marina 16(3): 221-224.
WILKE, F., TANIWAKI, T. and KURODA, H. 1953. Phocoenoides and Lagenorhynchus in Japan with notes on hunting. f. Mammal. 34: 488-497.
WILKE, F. and NICHOLSON, A. J. 1958. Food of porpoises in waters off Japan. f. Mammal. 39(3): 441-443. WoLMAN, A.A. 1989. Gray whale Eschrichtius robustus (Lilljeborg, 1861). pp.67-90. In: S.H. Ridgway and
R.J. Harrison (eds) Handbook of Marine Mammals, vol. 3. Academic Press, London. 362pp.
EXPLANATIONS OF PLATES
PLATE I. Lateral view of the skull and head of adult spotted dolphins. Fig. 1. BF: bony falx, BT: bony tentorium, U 1: unossified portion for the passage of the
cerebral veins into the dorsal sagittal sinus, U 2: unossified portion for the passage of the continuation of the spinal meningeal artery (SMA), G: groove for the continuation of SMA.
Fig. 2. FC: falx cerebri, TC: tentorium cerebelli, SMA: continuation of the SMA (intradural).
PLATE II. Fig. 1. Inner surface of the cranial cavity of a juvenile spotted dolphin. DSS: dorsal
sagittal sulcus. Fig. 2. Lateral view of the skull of a young striped dolphin. The BF and BT have not
been completed as a continuous bony plate. Black arrows point to the edges of developing BT.
PLATE III. Interior of the posterior half of the cranial cavity of an elderly spotted dolphin. Arrows indicate ossified laminations. FM: foramen magnum.
Sci. Rep. Cetacean Res., No. 1, 1990
6~
066! '[・ON’・Si!UUVii:JVJil:J ・diiU・!:JS
Iヨ工.V!d
Sヨ!VHMdO lぺ凡nIO.LNヨ.L人NOHONV S)f!Vd入NOH
60
PLATE II.
NOJIMA
Sci. Rep. Cetacean Res., No. I, 1990

19
0661 '[ 'ON "SJ~ UViJ:llJIJ:) ·da~ ・1:1s
・m 3lV'1d
SヨTv'HM.'.:10 lぺ凡nIOlNヨi入NOSON¥i $)11¥i.cl入NOS


















