
The pineal clock affects behavioral circadian rhythms but not photoperiodic induction in the Indian...
10
ORIGINAL ARTICLE Sangeeta Rani Sudhi Singh Vinod Kumar The pineal clock affects behavioral circadian rhythms but not photoperiodic induction in the Indian weaver bird (Ploceus philippinus) Received: 6 April 2005 / Revised: 20 July 2005 / Accepted: 20 July 2005 / Published online: 21 September 2005 ȑ Dt. Ornithologen-Gesellschaft e.V. 2005 Abstract We investigated whether pineal is part of the circadian clock system which regulates circadian rhythms of activity and photosensitivity in the Indian weaver bird (Ploceus philippinus). Two experiments were performed. The first experiment examined the induction of testicular growth, and androgen-dependent beak pigmentation and luteinizing hormone (LH)-specific plumage coloration in pinealectomised (pinx) and sham- operated (sham) birds exposed to short day (8 h light: 16 h darkness, 8L:16D) and long day (16L:8D) for 9 months in the late breeding and early regressive phase (October), or the late regressive and preparatory phase (January) of the annual testicular cycle. As expected, short days were non-stimulatory, and long days stimu- lated testicular growth, beak pigmentation and plumage coloration. There was no difference in the response be- tween pinx and sham birds subjected to short or long days in October, but the response was enhanced in pinx birds that were subjected to long day in January. In the second experiment circadian behavioral rhythms were studied (activity pattern in singly housed birds) in wea- ver birds first exposed at two different phases of the annual testicular cycle to a stimulatory photoperiod (12L:12D in preparatory phase or 13L:11D in early breeding phase) and then released into dim continuous light (LL dim ). All birds showed synchronization to the light period before and after the pinealectomy; there was no difference in the response between pinx and sham birds. When released into LL dim , sham birds exhibited circadian rhythmicity continuously, whereas pinx birds lost circadian rhythmicity after some cycles. Considered together, these results suggest that circadian clock residing within the pineal gland regulates the circadian rhythm in activity, but not the circadian rhythm in- volved in photoperiodic induction of the Indian weaver bird. Keywords Circadian rhythm Melatonin Pineal Pinealectomy Photoperiod Ploceus philippinus Introduction The avian pineal is an autonomous circadian pacemaker with its own input-oscillator-output systems (summary in Gwinner et al. 1997; Kumar et al. 2004). A circadian oscillator is present within each pineal cell (pinealocyte), and the net result is a stable circadian output in mela- tonin secretion (summary in Kumar et al. 2004). Several opsin-based photoreceptive molecules (e.g. pinopsin, Okano et al. 1994; melanopsin, Bailey et al. 2003) and signal transduction mechanisms (Kasahara et al. 2002) present within the pineal represent light-input pathways. Also, the pineal is part of the central clocking system that times circadian functions in birds (Gwinner et al. 1997; Kumar et al. 2004). Several lines of evidence have confirmed disruption of circadian rhythmicity in bird species in the absence of the pineal or after disruption of melatonin rhythmicity (summary in Gwinner et al. 1997; Kumar 2001). In an important experiment done more than 25 years ago, Zimmerman and Menaker (1979) showed that pinealectomised (pinx) arrhythmic house sparrow became rhythmic after a pineal had been transplanted into the anterior chamber of its eye with pineal from another sparrow; and that the restored rhythm had the phase of the donor. Day length regulates seasonal cycles of gonadal growth and regression in many birds (Kumar 1997; Gwinner and Hau 2000). An endogenous circadian photosensitivity rhythm enables birds to identify the time to switch on (photoinduction) the internal mechanisms responsible for the physiological response (Kumar and Follett 1993a; Kumar et al. 1996a). This is achieved by the interaction of daylight with a phase of Communicated by F. Bairlein S. Rani S. Singh V. Kumar (&) Department of Zoology, University of Lucknow, 226 007 Lucknow, India E-mail: [email protected] Tel.: +91-522-2740423 Fax: +91-522-2740423 J Ornithol (2005) 146: 355–364 DOI 10.1007/s10336-005-0005-9
-
Upload
sangeeta-rani -
Category
Documents
-
view
213 -
download
0
Transcript of The pineal clock affects behavioral circadian rhythms but not photoperiodic induction in the Indian...

ORIGINAL ARTICLE
Sangeeta Rani Æ Sudhi Singh Æ Vinod Kumar
The pineal clock affects behavioral circadian rhythmsbut not photoperiodic induction in the Indian weaverbird (Ploceus philippinus)
Received: 6 April 2005 / Revised: 20 July 2005 / Accepted: 20 July 2005 / Published online: 21 September 2005� Dt. Ornithologen-Gesellschaft e.V. 2005
Abstract We investigated whether pineal is part of thecircadian clock system which regulates circadianrhythms of activity and photosensitivity in the Indianweaver bird (Ploceus philippinus). Two experiments wereperformed. The first experiment examined the inductionof testicular growth, and androgen-dependent beakpigmentation and luteinizing hormone (LH)-specificplumage coloration in pinealectomised (pinx) and sham-operated (sham) birds exposed to short day (8 h light:16 h darkness, 8L:16D) and long day (16L:8D) for9 months in the late breeding and early regressive phase(October), or the late regressive and preparatory phase(January) of the annual testicular cycle. As expected,short days were non-stimulatory, and long days stimu-lated testicular growth, beak pigmentation and plumagecoloration. There was no difference in the response be-tween pinx and sham birds subjected to short or longdays in October, but the response was enhanced in pinxbirds that were subjected to long day in January. In thesecond experiment circadian behavioral rhythms werestudied (activity pattern in singly housed birds) in wea-ver birds first exposed at two different phases of theannual testicular cycle to a stimulatory photoperiod(12L:12D in preparatory phase or 13L:11D in earlybreeding phase) and then released into dim continuouslight (LLdim). All birds showed synchronization to thelight period before and after the pinealectomy; there wasno difference in the response between pinx and shambirds. When released into LLdim, sham birds exhibitedcircadian rhythmicity continuously, whereas pinx birdslost circadian rhythmicity after some cycles. Consideredtogether, these results suggest that circadian clockresiding within the pineal gland regulates the circadian
rhythm in activity, but not the circadian rhythm in-volved in photoperiodic induction of the Indian weaverbird.
Keywords Circadian rhythm Æ Melatonin Æ Pineal ÆPinealectomy Æ Photoperiod Æ Ploceus philippinus
Introduction
The avian pineal is an autonomous circadian pacemakerwith its own input-oscillator-output systems (summaryin Gwinner et al. 1997; Kumar et al. 2004). A circadianoscillator is present within each pineal cell (pinealocyte),and the net result is a stable circadian output in mela-tonin secretion (summary in Kumar et al. 2004). Severalopsin-based photoreceptive molecules (e.g. pinopsin,Okano et al. 1994; melanopsin, Bailey et al. 2003) andsignal transduction mechanisms (Kasahara et al. 2002)present within the pineal represent light-input pathways.Also, the pineal is part of the central clocking systemthat times circadian functions in birds (Gwinner et al.1997; Kumar et al. 2004). Several lines of evidence haveconfirmed disruption of circadian rhythmicity in birdspecies in the absence of the pineal or after disruption ofmelatonin rhythmicity (summary in Gwinner et al. 1997;Kumar 2001). In an important experiment done morethan 25 years ago, Zimmerman and Menaker (1979)showed that pinealectomised (pinx) arrhythmic housesparrow became rhythmic after a pineal had beentransplanted into the anterior chamber of its eye withpineal from another sparrow; and that the restoredrhythm had the phase of the donor.
Day length regulates seasonal cycles of gonadalgrowth and regression in many birds (Kumar 1997;Gwinner and Hau 2000). An endogenous circadianphotosensitivity rhythm enables birds to identify thetime to switch on (photoinduction) the internalmechanisms responsible for the physiological response(Kumar and Follett 1993a; Kumar et al. 1996a). This isachieved by the interaction of daylight with a phase of
Communicated by F. Bairlein
S. Rani Æ S. Singh Æ V. Kumar (&)Department of Zoology,University of Lucknow, 226 007 Lucknow, IndiaE-mail: [email protected].: +91-522-2740423Fax: +91-522-2740423
J Ornithol (2005) 146: 355–364DOI 10.1007/s10336-005-0005-9

photoinducibility of the circadian photosensitivityrhythm, which occurs some 12 h after dawn each day.Thus, during spring and summer months light periodsextend beyond 12 h of the day; so day is read as a ‘‘longday’’ and result into photoperiodic induction, namely,gonadal growth and development. Conversely, duringautumn and winter months the light periods end before12 h of the day; so day is read as ‘‘short day’’; conse-quently, there is no photoperiodic induction (Kumarand Tewary 1984; Kumar and Follett 1993a).
In mammals, the suprachiasmatic nucleus (SCN) ofthe anterior hypothalamus, which is considered as thesite of principal circadian pacemaker, responds to daylength (Sumova et al. 2002) and translates the photo-periodic message into a seasonal function by regulatingthe phase and duration of the daily rhythm of secretionof melatonin by the pineal gland (Goldman 2001).Hence, pinealectomy abolishes seasonal cycles both inlong-day and short-day breeders; timed melatonininfusions restore them (Goldman 2001). However, inbirds while pineal melatonin is important for circadianrhythmicity (Gwinner et al. 1997; Kumar 2001; Kumaret al. 2004), and circadian rhythms mediate photoperi-odic effects (Kumar and Follett 1993a), the effect ofpinealectomy or melatonin administration in photope-riodic induction of testicular growth is not found in themajority of species investigated thus far (summaries inGwinner and Hau 2000; Kumar 2001; Kumar et al.2002). This is inconsistent with the annual photoperiodicinformation both in the duration and amplitude of itsmelatonin output and the fact that avian pineal decodesdaily (Kumar and Follett 1993b; Brandstatter et al.2000a, b; Kumar 2001). Does this mean that in birds thecircadian photosensitivity rhythm regulating photoperi-odic induction is different from the one that regulatesother circadian functions, for example, rhythm inactivity rest? This has not been investigated. Interest-ingly, several previous studies have measured activityrhythms to validate the involvement of circadian systemin the measurement of day length (Hamner and Enright1967; Elliott et al. 1972) assuming that circadian activityrhythm reflects the characteristics of the circadianphotosensitivity rhythm. The implication of such anassumption is that both the circadian activity andphotosensitivity rhythms are outputs of the same circa-dian oscillator or of the same set of circadian oscillatorsthat are very closely coupled.
Therefore, we sought to investigate the effects of theabsence of pineal clock on circadian photosensitivityand activity rhythms in the Indian weaver bird (Ploceusphilippinus). This is a widely distributed, seasonallybreeding nonmigratory species whose breeding seasonextends from May to September (Ali and Ripley 1974).Saxena (1964) divided annual testicular cycle of thesebirds into four (preparatory (quiescent), progressive,breeding and regressive) phases. During breeding sea-son, males develop luteinizing hormone (LH)-specificbright yellow plumage (Thapliyal and Saxena 1961) anda dark pigmented beak, which is indicative of the
androgen level (Saxena and Thapliyal 1962). Duringnonbreeding (quiescent) phase, males have female-likeplumage and non-pigmented beak (Thapliyal and Sax-ena 1961; Saxena and Thapliyal 1962). Previous studieshave shown that, (1) it is a photoperiodic species (Tha-pliyal and Tewary 1964), (2) circadian photosensitivityrhythm mediates photoperiodic induction (Chandolaet al. 1976) and (3) pinealectomy accelerates photoperi-odic induction under long days, and appears to changethe perception of short days (Balasubramanian andSaxena 1973). In this study, we specifically examined theeffects of the pinealectomy on the photoperiodic re-sponses to short and long days and on circadian activityrhythms at two different phases of the annual testicularcycle.
Methods
Two experiments were performed on male weaver birdsprocured from natural population living at 27�N, 81�E.Before any experimental procedure began, birds wereacclimatized for 1–3 weeks (see below) in the outdooraviary under natural day lengths (NDL). Food andwater were available ad libitum in all experiments.
Experiment 1: Induction under short (8L:16D) and longdays (16L:8D)
This experiment compared induction of photoperiodicresponses in pinx and sham male weaver birds subjectedto short (8L:16D) and long days (16L:8D) for severalweeks (see below). Birds in groups of 4 or 5 were held inlight tight boxes provided with a fluorescent light source(CFL, 14 Watt Philips). The dark phase of an LD cyclehad very dim light at <1.0 lux. Pinealectomy and shamoperations were done as described by Haldar (1986).There is no adverse effect of this surgery on birds asobserved in body mass and activity profile (unpublishedobservations). Photoperiodic induction was assessed byobservations on changes in testis size; beak and plumagecolors were taken at the beginning and end of theexperiment and at monthly intervals. Testicular responsewas measured by unilateral laparotomy under localanesthesia as described by Kumar et al. (2001). Thedimensions of the left testis were recorded and testisvolume (TV) was calculated from 4/3 p ab2, where a andb denote half of the long and short axes, respectively. Wealso did a subjective grading of the testis size to explainthe response: TV=0.33 to <2.35 mm3—no response;2.35 to <9.82 mm3—initiation of response; 9.82 to<18.86 mm3—small response; 18.86 to <41.9 mm3—moderate response; 41.9 mm3 and above—full response.In males, beak color changes over seasons from non-breeding straw to breeding dark black, and feather colorover the head and breast regions from nonbreeding‘‘female-like’’ to breeding (nuptial) bright yellow.Therefore, we recorded changes in the beak and feather
356

colors over the head and in the breast regions, usingsubjective criterion, and considered them an indicator ofendogenous androgen (Saxena and Thapliyal 1962) andLH (Thapliyal and Saxena 1961) levels. For betterillustration of the observations and statistical compari-sons, the beak pigmentation and plumage colors werescored, as described by Trivedi (2004). Briefly, beakcolor was scored on a scale of 0–5 as follows; 0 – straw incolor (S), 1 – straw in color but with a little tinge ofblackness (ratio SSS:B), 2 – slightly blackish in color(ratio SS:B), 3 – straw and black in approximately 50:50patches (ratio S:B), 4 – black with very little straw patchleft (ratio S:BB), 5 – fully black (B). Similarly, plumagecolor was scored on a scale of 1–3 (head: 1=‘‘female-like’’ feathers; 2=mixed ‘‘female-like’’ and yellow(nuptial) feathers; 3=complete yellow feathers) or 0–5(breast: 0=all ‘‘female-like’’ (no yellow) feathers; 5=allyellow (no female-like) feathers; every 0.5 increment inthis scale (e.g. 0.5, 1.0 and so on) meant 10% increasefrom ‘‘female-like’’ to nuptial feathers).
The experiment consisted of two parts; in differentphases of the annual testicular cycle birds were subjectedto artificial short and long day lengths.
Experiment 1A—late breeding and early regressive phase
Birds procured in the first week of September 2003 hadlarge testes (mean (±SE) testis volume, TV=33.01±3.94 mm3; n=26), dark pigmented beak (mean (±SE)score=4.50±0.50) and bright yellow (nuptial) plumage(mean (±SE) score=3.00±0.00, head; 5.00±0.00,breast). They were kept in the out door aviary, and whilein the NDL, they were pinealectomised (pinx) or sham-operated (sham) in third week of September 2003, andlaparotomised 1 day before the start of the experimenton 01 October 2003. By this time, the birds had regressedtestes (mean (±SE) TV=3.09±0.45 mm3; n=26]) butretained their dark pigmented beak (mean (±SE)score=3.15±0.13) and nuptial plumage (mean (±SE)score=3.00±0.00, head; 5.00±0.00, breast). Groups ofpinx (n=6–7) and sham (n=6–7) birds were exposed toshort day (8 h light: 16 h darkness; 8L:16D) or long day(16L:8D) for 9 months.
Experiment 1B—late regressive and preparatory phase
It was completed in two stages using birds with fullyregressed small testes (mean (±SE)TV=0.34±0.05 mm3; n=16), unpigmented beak and‘‘female-like’’ plumage. In the first stage, birds werecaught in the third week of November 2003, and kept inthe outdoor aviary. Acclimatized birds (n=8 each) werepinealectomised (pinx) or sham-operated (sham) in midDecember 2003, and then exposed to long day (16L:8D)beginning from 29 January 2004. In the second stage,birds procured in second week of January 2004 were firstacclimatized in the outdoor aviary and then pinealecto-mised (pinx) or sham-operated (sham) in mid-February.
They were exposed to short day (8L:16D; pinx, n=8 andsham, n=10) in mid March 2004. At this time, they hadsmall testes (mean (±SE) TV=0.34±0.07 mm3; n=18),unpigmented beak and ‘‘female-like’’ plumage.
Experiment 2: Effects on circadian behavioral rhythm
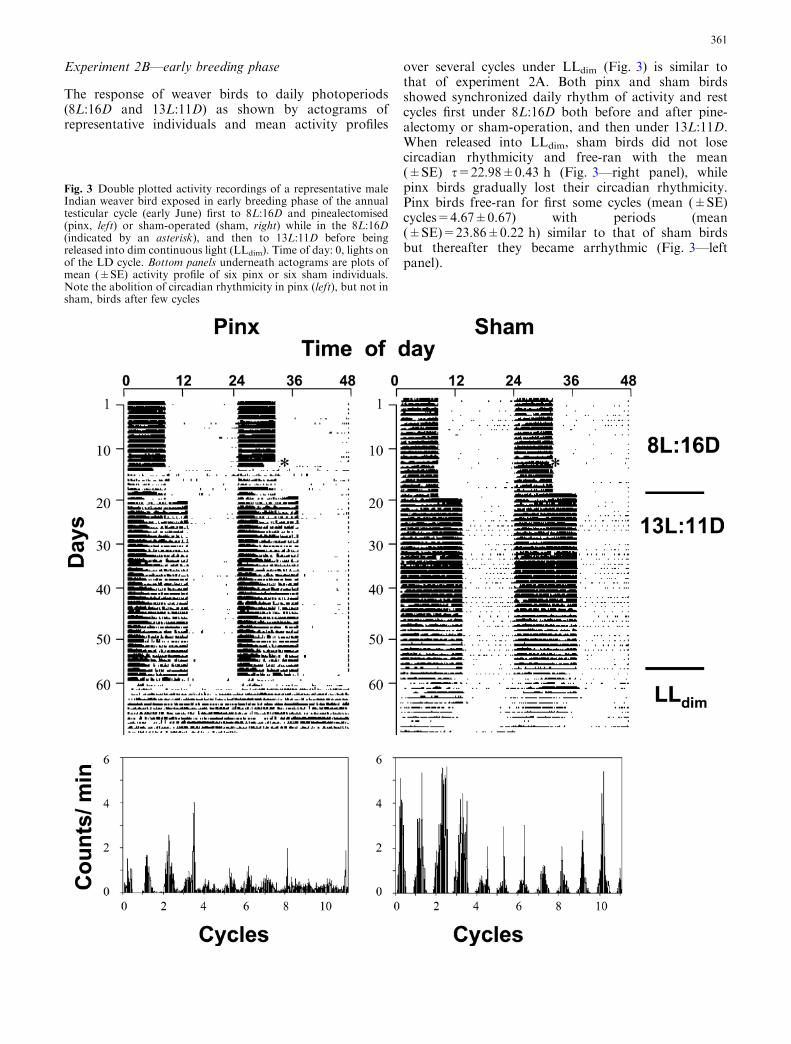
This experiment examined the effects of the absence ofpineal on a circadian behavioral rhythm. We measuredthe bird’s general activity within its cage for analysis ofthe circadian system, as described by Malik et al. (2004).Briefly, each bird was housed in a specially designedactivity cage furnished with two perches and mountedwith a passive infrared motion sensor 12 m (40¢) range(C & K Systems (Intellisense XJ-413T) Conrad Elec-tronic, Germany, Haustier PIR-Melder) which detectsthe movement of a bird within its cage. Each sensor wasconnected to a channel of an IBM-compatible computer,and the recording was done using a software programfrom Stanford Software Systems, Stanford, USA.Activity records in Figs. 2 and 3 are shown as doubleplots. Each day is duplicated along horizontal axis,whereas subsequent days are shown underneath inincreasing order. Under LLdim, the circadian period(tau, s) of activity rhythm and the activity profile tem-porally distributed over 24 h on successive days weredetermined using program of the Stanford SoftwareSystems. We also calculated mean (±SE) activity profileof the pinx (left) sham (right) groups.
The experiment consisted of two parts, which differedin the (1) duration and intensity of daytime light thebirds were subjected to, (2) phase of annual testicularcycle when birds were experimented, (3) length of dailyphotophase and (4) timing and photoperiodic conditionwhen birds were pinealectomised.
Experiment 2A—preparatory phase
Birds were procured in the first week of January 2004and kept in outdoor aviary. At this time, they had smalltestes, unpigmented beak and ‘‘female-like’’ plumage.After 1 week of acclimatization in outdoor aviary, birdswere moved indoors under 8L:16D (L=50±2 lux;D=<1.0 lux). On the third day, the LD cycle waschanged to 12L:12D by extending the time of light off.After 12 cycles of 12L:12D, birds were pinealectomised(pinx: n=5) or sham-operated (sham: n=4). Then, fol-lowing 12 cycles of 12L:12D, the birds were released intoconstant dim light (LLdim; <1.0 lux, night time illumi-nation of the LD) for 16 cycles.
Experiment 2B—early breeding phase
Birds were procured in the last week of May 2004. Atthis time, they had large testes, dark pigmented beak andbright yellow (nuptial) plumage. After 1 week of accli-matization in outdoor aviary, birds were moved indoors
357

on 8L:16D (L=160±2 lux; D=<1.0 lux). After 14cycles, birds were pinealectomized (pinx: n=6) or sham-operated (sham: n=6). They were maintained for fur-ther six cycles, and beginning on seventh cycle the lightphase was extended to 13 h (13L:11D). After 38 cycles,all birds were released into LLdim (<1.0 lux, night timeillumination of the LD) for 11 cycles.
The data are presented as means and SEs. One-wayanalysis of variance with repeated measures (one-wayRM ANOVA), followed by post hoc tests if ANOVAindicated the significance of difference, was employedto examine the effect of a photoperiodic treatment as afunction of time. Two-way analysis of variance (two-way ANOVA) was used to determine the significanceof difference when two factors were involved. Themeans of two groups on selected observation werecompared by Student’s t-test. Significance was taken atP<0.05.
Results
Experiment 1: Induction under short (8L:16D) and longdays (16L:8D)
Experiment 1A—late breeding and early regressive phase
At the beginning of the experiment, testes were small butnot fully regressed (mean (±SE) TV=3.09±0.45 mm3),beak was pigmented and head and breast plumage wereyellow (Fig. 1a–d). Under short days, as expected, therewas no stimulation. Instead, there was initially regres-sion of testes and then birds maintained small testes(range of TV=0.34–1.05 mm3); this produced a statis-tically significant variation over the experimental period(pinx: F9,45=5.205, P<0.0001; sham: F9,54=7.056,P<0.0001, one-way RM ANOVA). At the end of theexperiment, four of six pinx and one of seven sham birdshad TV=2.36 mm3. Under long days, testes underwentsignificant growth and regression (pinx: F9,54=9.058,P<0.0001; sham: F9,45=7.542, P<0.0001; one-way RMANOVA). Testes were small and unstimulated in bothgroups at the end of the first month of exposure. At theend of the second month, however, there was initiationof response in four of the seven pinx and five of six shambirds, and after the third month, six of seven pinx and allsham individuals had responded. Of these six pinx re-sponders, the response ranged from gonadal initiation tosmall response; compared to this, among sham birds theresponse ranged from initiation to a moderate (in twobirds) response. A further increase in testis size wasfound for the following 3 months. Testes regressed atthe end of 7 months, and remained so until the end ofthe experiment. Except in the timing of response in fewindividuals, there was no significant difference inphotoperiodic induction of testes under long days be-tween pinx and sham birds (F1,110=0.59, P=0.2704;two-way ANOVA).
Changes in beak pigmentation and plumage colorscorresponded to photoperiod-induced testicular recru-descence (cf. Fig. 1a–d). At the beginning of theexperiment, beaks were dark, and plumage over thehead and breast regions were yellow in all groups sincebirds still had relatively large testes (Fig. 1a). However,during the first 2 months of exposure, beak becamesignificantly (P<0.05, Newman–Keuls test) de-pig-mented in both short and long day groups (Fig. 1b).Under continued short day exposure, beaks remainedalmost straw colored until 7 months of exposure. Therewas small gain in color in few individuals (pinx—SSS/Bin two and SS/B in one bird; sham—SSS/B in fourbirds). There was also a significant loss of beak pig-mentation (pinx: F9,45=9.09, P<0.0001; sham:F9,54=31.09, P<0.0001, one-way RM ANOVA). Un-der long days, beak began darkening by the thirdmonth, and pigmented beak (mean (SE)score=3.7±0.3) was attained by 6 months of exposure(Fig. 1b; pinx: F9,54=5.964, P<0.0001; sham:F9,45=6.769, P<0.0001; one-way RM ANOVA). Theeffect on plumage colors was similar. Gradual increasesin the proportion of newly generated feathers lightenedthe plumage color of both the head and breast regions.There was a significant decrease in the color of headplumage of birds under long days (pinx: F9,54=4.124,P=0.0004; sham: F9,45=2.092, P<0.05; one-way RMANOVA) but the plumage did not become ‘‘female-like’’ during this experiment. Under short days,however, head plumage became significantly lighter(pinx: F9,45=23.99, P<0.0001; sham: F9,54=50.22,P<0.0001, one-way RM ANOVA), and birds attained‘‘female-like’’ plumage by the third month, which theymaintained till the end of the experiment (Fig. 1c). Notmuch different was the response of feathers of breastregion except that under short days pinx birds did notbecome as light as sham birds, and difference betweenthem was significant (P<0.05; student’s t-test) at theend of 8- and 9-month exposure. There was significantloss of color of breast feathers in all groups (short day:pinx—F9,45=6.372, P<0.0001; sham—F9,54=16.54,P<0.0001; long day: pinx—F9,54=5.614, P<0.0001;sham—F9,45=3.871, P=0.011; one-way RM ANOVA)but short day birds lost relatively more than long daybirds (cf. Fig. 1d). As with testes, there was no signif-icant difference in response between pinx and shambirds.
Experiment 1B—late regressive and preparatory phase
There was no stimulation under short days, and hencetestes remained small, the beak unpigmented, andhead and breast plumage ‘‘female-like’’ type, as theyhad been at the beginning of the experiment (Fig. 1e–h). However under long days, testes underwent sig-nificant recrudescence and regression in both the pinxand sham groups (pinx: F9,63=7.503, P<0.0001,sham: F9,63=5.005, P<0.0001; one-way RM ANO-
358
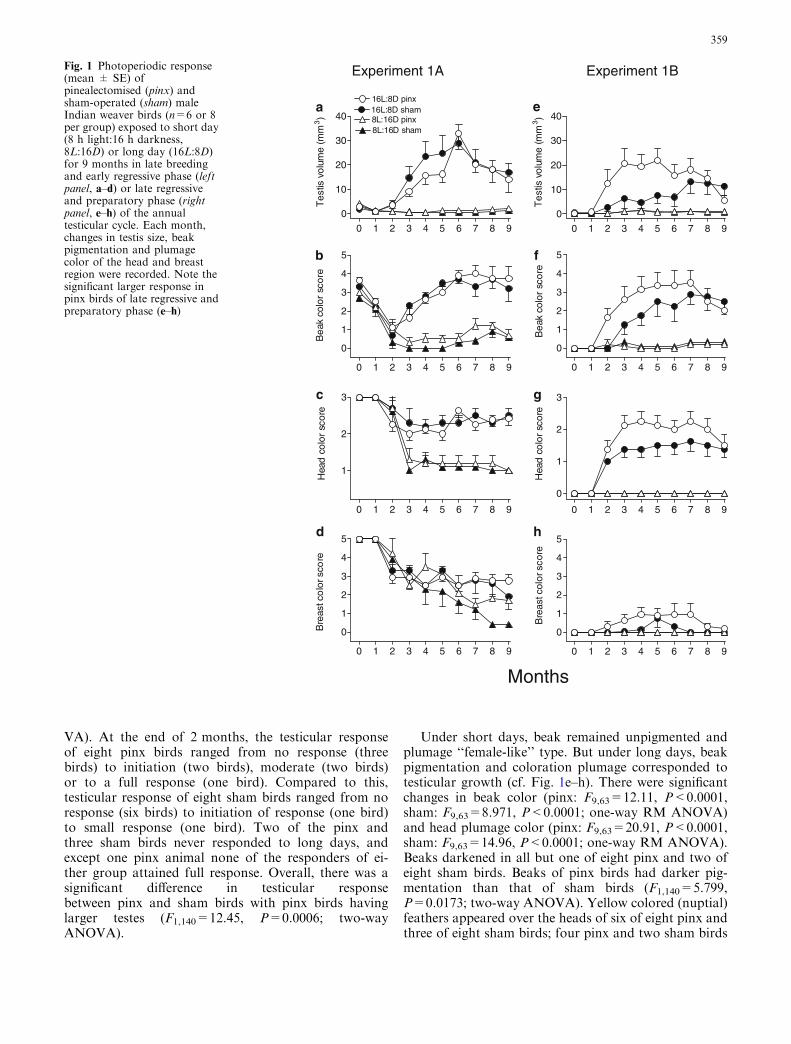
VA). At the end of 2 months, the testicular responseof eight pinx birds ranged from no response (threebirds) to initiation (two birds), moderate (two birds)or to a full response (one bird). Compared to this,testicular response of eight sham birds ranged from noresponse (six birds) to initiation of response (one bird)to small response (one bird). Two of the pinx andthree sham birds never responded to long days, andexcept one pinx animal none of the responders of ei-ther group attained full response. Overall, there was asignificant difference in testicular responsebetween pinx and sham birds with pinx birds havinglarger testes (F1,140=12.45, P=0.0006; two-wayANOVA).
Under short days, beak remained unpigmented andplumage ‘‘female-like’’ type. But under long days, beakpigmentation and coloration plumage corresponded totesticular growth (cf. Fig. 1e–h). There were significantchanges in beak color (pinx: F9,63=12.11, P<0.0001,sham: F9,63=8.971, P<0.0001; one-way RM ANOVA)and head plumage color (pinx: F9,63=20.91, P<0.0001,sham: F9,63=14.96, P<0.0001; one-way RM ANOVA).Beaks darkened in all but one of eight pinx and two ofeight sham birds. Beaks of pinx birds had darker pig-mentation than that of sham birds (F1,140=5.799,P=0.0173; two-way ANOVA). Yellow colored (nuptial)feathers appeared over the heads of six of eight pinx andthree of eight sham birds; four pinx and two sham birds
0 1 2 3 4 5 6 7 8 9
0
1
2
3
Hea
d co
lor
scor
e
0 1 2 3 4 5 6 7 8 9
1
2
3
Hea
d co
lor
scor
e
0 1 2 3 4 5 6 7 8 9
0
1
2
3
4
5B
reas
t col
or s
core
0 1 2 3 4 5 6 7 8 9
0
1
2
3
4
5
Bre
ast c
olo
r sco
re
0 1 2 3 4 5 6 7 8 9
0
1
2
3
4
5
Bea
k co
lor
scor
e
0 1 2 3 4 5 6 7 8 9
0
1
2
3
4
5
Bea
k co
lor
scor
e
0 1 2 3 4 5 6 7 8 9
0
10
20
30
40
Tes
tis v
olum
e (m
m3 )
0 1 2 3 4 5 6 7 8 9
0
10
20
30
40a e
b f
c g
d h
16L:8D pinx
8L:16D pinx16L:8D sham
8L:16D sham
Tes
tis v
olum
e (m
m3)
Months
Experiment 1A Experiment 1BFig. 1 Photoperiodic response(mean ± SE) ofpinealectomised (pinx) andsham-operated (sham) maleIndian weaver birds (n=6 or 8per group) exposed to short day(8 h light:16 h darkness,8L:16D) or long day (16L:8D)for 9 months in late breedingand early regressive phase (leftpanel, a–d) or late regressiveand preparatory phase (rightpanel, e–h) of the annualtesticular cycle. Each month,changes in testis size, beakpigmentation and plumagecolor of the head and breastregion were recorded. Note thesignificant larger response inpinx birds of late regressive andpreparatory phase (e–h)
359

had bright yellow head feathers (score=3). As with beakpigmentation, head plumage color was more brightlycolored in pinx than in sham birds (F1,140=14.46,P=0.0002; two-way ANOVA). The breast plumage of afew individuals gained color but the mean score did notvary significantly over the experimental period (pinx:F9,63=1.855, P=0.0756, sham: F9,63=1.890, P=0.0698;one-way RM ANOVA). Feathers in the breast regionbecame yellow in three of eight pinx and one of eightsham birds. Similar to head feathers, breast plumage ofpinx birds appeared was significantly brighter in color
than in sham birds (F1,140=7.795, P=0.0006; two-wayANOVA).
Experiment 2: Effects on circadian behavioral rhythm
Experiment 2A—preparatory phase
Actograms of representative individuals and meanactivity profiles over several cycles under LLdim areshown in Fig. 2. As expected, birds showed synchroni-zation to the light period under 12L:12D both beforeand after pinealectomy or sham-operation. There wereno differences in activity behavior under long days be-tween pinx and sham birds (cf. Fig. 2). When releasedinto LLdim, sham birds did not lose circadian rhyth-micity and free-ran with mean (±SE) s=23.44±0.26 h(Fig. 2—right panel). Pinx birds also free-ran with sim-ilar periods (mean (±SE) s=23.56±0.60 h; Fig. 2—leftpanel) for some cycles (mean (±SE) cycles=6.40±1.72)but then they lost their circadian rhythmicity(Fig. 2—left panel).
Fig. 2 Double plotted activity recordings of a representative maleIndian weaver bird exposed first to 12L:12D and then released intodim continuous light (LLdim) in preparatory phase of the annualtesticular cycle (mid January). Time of day: 0, lights on of the LDcycle. Asterisk within the actograms indicates when birds werepinealectomised (pinx, left) or sham-operated (sham, right). Bottompanels underneath actograms are plots of mean (±SE) activityprofile of five pinx (left) or four sham (right) individuals. Noteabolition of circadian rhythmicity in pinx (left), but not in sham,birds after few cycles
360
Experiment 2B—early breeding phase
The response of weaver birds to daily photoperiods(8L:16D and 13L:11D) as shown by actograms ofrepresentative individuals and mean activity profiles
over several cycles under LLdim (Fig. 3) is similar tothat of experiment 2A. Both pinx and sham birdsshowed synchronized daily rhythm of activity and restcycles first under 8L:16D both before and after pine-alectomy or sham-operation, and then under 13L:11D.When released into LLdim, sham birds did not losecircadian rhythmicity and free-ran with the mean(±SE) s=22.98±0.43 h (Fig. 3—right panel), whilepinx birds gradually lost their circadian rhythmicity.Pinx birds free-ran for first some cycles (mean (±SE)cycles=4.67±0.67) with periods (mean(±SE)=23.86±0.22 h) similar to that of sham birdsbut thereafter they became arrhythmic (Fig. 3—leftpanel).
Fig. 3 Double plotted activity recordings of a representative maleIndian weaver bird exposed in early breeding phase of the annualtesticular cycle (early June) first to 8L:16D and pinealectomised(pinx, left) or sham-operated (sham, right) while in the 8L:16D(indicated by an asterisk), and then to 13L:11D before beingreleased into dim continuous light (LLdim). Time of day: 0, lights onof the LD cycle. Bottom panels underneath actograms are plots ofmean (±SE) activity profile of six pinx or six sham individuals.Note the abolition of circadian rhythmicity in pinx (left), but not insham, birds after few cycles
361

Discussion
The response of both pinx and sham birds under shortand long days was essentially similar in both theexperiments except in experiment 1B under long days. Inthis case, the rate of photoperiodic induction was fasterand the magnitude of response greater in pinx than insham birds (cf. Fig. 1e–h). Birds darkened and lightenedtheir beak as they grew and regress their testes, respec-tively (Fig. 1a–h), which confirms a previous finding ofThapliyal and Saxena (1961). Also, there were increasednumbers of bright yellow feathers over the head andbreast regions in birds that were stimulated under longdays, reflecting activity of the hypothalamo-hypophysealaxis (Fig. 1c, d, g, h). This is consistent with the findingof Saxena and Thapliyal (1962) suggesting LH-specificplumage coloration in the Indian weaver bird. However,plumage color of birds in experiment 1A did not remainas bright as they were at the beginning of the experimentalthough their testes regrew in size and beak repig-mented after 3 months of long days (cf. Fig. 1a–d). Wedid not record body molt in this experiment, but this isthe time of the year when birds undergo post-nuptialmolt (our unpublished observation) and hence newlygenerated feathers at this time might have produced alighter appearance to the plumage. A comparison of thedata on plumage color scores from both the experimentsindicates that under long days plumage over the headand breast regions had mixture of colored and ‘‘female-like’’ feathers (cf. Fig 1c, d, g, h). Interestingly, plumagecolors in long day birds were brighter than short daybirds, and among the former birds of experiment 1B,pinx individuals had brighter plumage than shams, aswas the case with the testicular growth and beak pig-mentation (Fig. 1).
Our results are only partially consistent with a pre-vious study on Indian weaver bird by Balasubramanianand Saxena (1973). There it was shown that (1) underartificial long days (18L:6D), or naturally increasingday lengths, pinx birds showed acceleration of testisgrowth, darkening of androgen-dependent beak colorand yellow pigmentation of the plumage and (2) undershort days (9L:15D), reproductively mature pinx birdsshow transient partial regression of testes but, unlikesham birds, they never fully regress, lighten their beakto straw color or depigment their plumage to non-breeding ‘‘female-like’’ type. In the present study, therewas an enhanced testis growth, beak pigmentation andplumage coloration in pinx birds subjected to long daysin January but not in October (Fig. 1a–h), and bothpinx and sham birds under short days had fully re-gressed testes (Fig. 1a, e). Thus, the effect of pinealec-tomy on photoperiodic induction in the Indian weaverbird under long day appears dependent on the phase ofthe birds’ annual gonadal cycle (cf. experiments 1A and1B; Fig. 1a–h). In another relatively recent study per-formed in late August on weaver birds with large testesand dark beak, exogenous melatonin received via
silastic implants failed to show antigonadal effects ei-ther on testes or on androgen-dependent beak color(Kumar et al. 2002). However, a reproductive phase-dependent effect of the pinealectomy and of exogenousmelatonin, respectively, is reported in the Indian junglebush quail (Perdicula asiatica, Haldar and Ghosh 1990)and roseringed parakeet (Psittacula krameri, Maitraand Dey 1992).
Figures 2 and 3 show consistent effects of pinealec-tomy on circadian activity rhythms in both the experi-ments. Pinealectomy abolished circadian rhythmicitywhen weaver birds were kept in LLdim and free-ran, butnot when they were kept under LD. Further, pinx birdsdid not become arrhythmic immediately; rather theygradually lost their circadian rhythmicity. There was al-ways a residual rhythmicity for-a-while in pinx individ-uals. Similar results of the pinealectomy on circadianrhythms are reported in other song birds (summary inKumar 2001). In the house sparrow, for example, pine-alectomy disrupts the circadian rhythmicity in locomo-tion (Gaston and Menaker 1968), body temperature(Binkley et al. 1971) and feeding (Heigl and Gwinner1994), but the effects are seen only after three to sevencycles (Gwinner et al. 1997). All this means that pinealclock interacts with one or more clocks of the aviancentral clocking system regulating circadian rhythms. Inall probability, pineal interacts with the hypothalamicoscillator, the avian equivalent of the mammalian SCN,and possibly the retinal oscillator (Kumar et al. 2004).Lesions of the hypothalamic pacemaker result in severeimpairment of rhythmicity in the pineal intact housesparrow (Takahashi and Menaker 1982). Similarly,complete removal of eyes (blinding) produces ar-rhythmicity in the Japanese quail (Underwood 1994).
A circadian photosensitivity rhythm is suggested formediating the measurement of day length, hence photo-periodic induction, in birds (Kumar and Follett 1993b;Kumar et al. 1996a). Because pineal is part of the aviancentral clock system regulating circadian rhythms(Gwinner et al. 1997; Kumar 2001; Kumar et al. 2004),the pinealectomy should disrupt the circadian rhythmthat helps distinguish weaver birds between short andlong days. However, the present results on photoperiodicresponse (Fig. 1) show that pinealectomy, which disruptsthe circadian activity rhythm (Figs. 2, 3), does not affectphotoperiodic time measurement in weaver birds; pinxbirds continued showing responses characteristic of shortand long days. Studies on several other species includingEuropean starling (Sturnus vulgaris, Gwinner and Dit-tami 1980), spotted munia (Lonchura punctulata, Chan-dola-Saklani et al. 1988), tree sparrow (Spizella arborea,Wilson 1991), Japanese quail (Coturnix c. japonica,Kumar et al. 1993), blackheaded bunting (Emberizamelanocephala, Kumar 1996; Kumar et al. 1996b) andredheaded bunting (Emberiza bruniceps, Kumar et al.2002) also show the absence of a role of melatonin inphotoperiodic induction of testicular recrudescence.
The pineal appears exerting a modulatory effect onthe timing of testicular response. Removal of the pineal
362

augmented testicular growth in birds subjected to longdays in the preparatory phase of the annual testicularcycle (Fig. 1e). This might mean that pineal, hence dailymelatonin signal from it, plays a role in temporalphasing of the annual testicular cycle correspondingwith the annual photoperiodic cycle. Kumar et al. (2002)drew a similar conclusion from their study on redheadedbunting in which the abolition of daily rhythm of mel-atonin by exogenous melatonin administered via silasticimplant had altered the timing of the peak testicularresponse. How is it that the absence of pineal or theabolition of daily rhythmicity in melatonin modulatephotoperiod-induced response is not clearly known? Aplausible explanation is that the absence of pineal or ofrhythmic melatonin signal enhances sensitivity of thecircadian system to light, and as a consequence theamplitude of perceived LD cycle is amplified. Studies onrodents support such an explanation (Cassone et al.1993). At an identical light intensity in LL, circadianrhythms of pinx, but not of intact, rats are disrupted;this meant that pinx animals perceived ambient intensityhigher than did intacts (Cassone 1992).
In conclusion, pineal clock regulates circadianrhythms, but not the circadian rhythm that mediatesmeasurement of day length, hence photoperiodicinduction, in the Indian weaver bird. The present studythus supports the view that in birds the circadian clockregulating photoperiodic induction apparently functionsdifferently from the one that regulates other circadianrhythms, for example, rhythms in activity-rest. In fact, ata physiological level, we have found that in certainexperimental situations, the circadian rhythm mediatingphotoperiodic induction of LH release dissociates fromthe circadian locomotor rhythm (Juss et al. 1995). Fur-ther, there is evidence from ring doves (Streptopeliaroesogrisea) that deep brain photoreceptors, which arenecessary and sufficient for the detection of changes inday length, are directly linked to GnRH neurons(Saldanha et al. 2001). If these photoreceptors them-selves contain circadian clock as pineal does, thephotoperiodic environment can be measured andtransduced to reproductive system by them bypassingthe central clocking system (Kumar et al. 2004) thatregulates several circadian functions including circadianactivity rhythm.
Zusammenfassung
Die Epiphysen-Uhr reguliert circadiane Rhythmen, abernicht denjenigen, der photoperiodische Anregung beimBayaweber (Ploceus philippinus) vermittelt
Wir untersuchten die Rolle der Epiphyse (Pinealorgan)in der Regulation photoperiodischer und circadianerVerhaltensrhythmen beim Bayaweber (P. philippinus).Zwei Experimente wurden durchgefuhrt. Das eine un-tersuchte die Induktion des Hodenwachstums, die an-drogenabhangige Schnabelpigmentierung und die von
luteinisierendem Hormon (LH) abhangige Gefiederfar-bung bei pinealektomierten Vogeln (pinx) und solchenmit einer Scheinoperation (sham). Dazu wurden dieVogel wahrend der spaten Brut- und fruhen Regres-sionsphase (Oktober) oder wahrend der spaten Regres-sions- und Vorbereitungsphase (Januar) des jahrlichenHodenzyklus fur 9 Monate entweder in einem Kurztag(8 Stunden Licht, 16 Stunden Dunkelheit; 8L:16D) oderin einem Langtag (16L:8D) gehalten. Wie erwartet, zei-gten kurze Tag keine stimulierende Wirkung, lang Tagedagegen stimulierten Hodenwachstum, Schnabelpig-mentierung und Gefiederfarbung. Es gab keinen Un-terschied zwischen pinx- und sham-Vogeln im Oktober,im Januar jedoch war die Reaktion der pinx-Vogel imLangtag ausgepragter. Das zweite Experiment un-tersuchte circadiane Aktivitatsmuster einzeln gekafigterWebervogel, die zunachst zu zwei verschiedenen Phasendes jahrlichen Hodenzyklus einem stimulierenden Licht-regime (12L:12D in der Vorbereitungsphase und13L:11D in der fruhen Brutphase) ausgesetzt und dannin eine kontinuierliche Dammerung (LL_dim) umgesetztwurden. Alle Vogel zeigten sowohl vor wie nach derPinealektomie eine Synchronisierung mit den Lichtzyk-len und pinx—und sham-Vogel verhielten sich gleich.Nach der Umsetzung in Dauerdammerung entwickeltendie sham-Vogel eine andauernde circadiane Rhythmik,wogegen pinx-Vogel die circadiane Rhythmik nach ei-nigen Zyklen verloren. Diese Ergebnisse deuten daraufhin, dass die circadiane Uhr im Pinealorgan den circa-dianen Rhythmus von Aktivitat steuert, nicht jedochden circadianen Rhythmus, der an Prozessen der pho-toperiodischen Induktion des Hodenwachstums beimBayaweber beteiligt ist.
Acknowledgments The experiments included in this paper conformto Indian laws, and were done using the facility exclusively gener-ated from the SERC research grant to VK by the Department ofScience and Technology, Government of India.
References
Ali S, Ripley SD (1974) Handbook of birds of India and Pakistan,vol 10, 2nd edn. Oxford University Press, Delhi, Bombay,London, New York
Bailey MJ, Beremand PD, Hammer R, Bell-Pedersen D, ThomasTL, Cassone VM (2003) Transcriptional profiling of the chickenpineal gland, a photoreceptive circadian oscillator and pace-maker. Mol Endocrinol 17:2084–2095
Balasubramanian KS, Saxena RN (1973) Effect of pinealectomyand photoperiodism in the reproduction of the Indian weaverbirds, Ploceus philippinus. J Exp Zool 185:333–348
Binkley S, Kluth E, Menaker M (1971) Pineal function in sparrows:circadian rhythm and body temperature. Science 174:311–314
Brandstaetter R, Kumar V, Abraham U, Gwinner E (2000a)Photoperiodic information acquired and stored in vivo is re-tained in vitro by a circadian oscillator, the avian pineal gland.Proc Natl Acad Sci USA 97:12324–12328
Brandstaetter R, Kumar V, Van’t Hof TJ, Gwinner E (2000b)Seasonal variations of in vivo and in vitro melatonin produc-tion in a passeriform bird, the house sparrow (Passer domesti-cus). J Pineal Res 31:120–126
Cassone VM (1992) The pineal gland influences rat circadianactivity rhythm in constant light. J Biol Rhythms 7:27–40
363

Cassone VM, Warren WS, Brooks DS, Lu J (1993) Melatonin, thepineal gland, and circadian rhythms. J Biol Rhythms 8(Sup-pl):573–581
Chandola-Saklani A, Singh R, Thapliyal JP (1976) Evidence for acircadian oscillation in the gonadal response of the tropicalweaver bird (Ploceus philippinus) to programmed photoperiods.Chronobiologia 3:219–227
Chandola-Saklani A, Bhatt D, Lakhera P (1988) Effect of pineal-ectomy on free-running reproductive cycle of tropical spottedmunia. J Comp Physiol A 164:117–121
Elliott JA, Stetson MH, Menaker M (1972) Regulation of testisfunction in golden hamsters: a circadian clock measuresphotoperiodic time. Science 178:771–773
Gaston S, Menaker M (1968) Pineal function: the biological clockin the sparrow? Science 160:1125–1127
Goldman BD (2001) Mammalian photoperiodic systems: formalproperties and neuroendocrine mechanisms of photoperiodictime measurement. J Biol Rhythm 16:283–301
Gwinner E, Dittami J (1980) Pinealectomy affects the circannualtesticular rhythm in European starlings (Sturnus vulgaris). JComp Physiol A 136:345–348
Gwinner E, Hau M (2000) The pineal gland, circadian rhythms andphotoperiodism. In: Whittow GC (ed) Sturkie’s avian physiol-ogy. Academic Press, New York, pp 557–568
Gwinner E, Hau M, Heigl S (1997) Melatonin: generation andmodulation of avian circadian rhythms. Brain Res Bull 44:439–444
Haldar C (1986) Methods of pinealectomy in vertebrates. Indian JExp Biol 24:319–322
Haldar C, Ghosh M (1990) Annual pineal and testicular cycle inthe Indian jungle bush quail, Perdicula asiatica, with referenceto the effect of pinealectomy. Gen Comp Endocrinol 77:150–157
Hamner WM, Enright JT (1967) Relationships between photo-periodism and circadian rhythms of activity in the house finch. JExp Biol 46:43–61
Heigl S, Gwinner E (1994) Periodic melatonin in the drinking watersynchronizes circadian rhythms in sparrows. Naturwissens-chaften 81:83–85
Juss TS, King VM, Kumar V, Follett BK (1995) Does an unusualentrainment of the circadian system under T36 h photocyclesreduce the critical day length for periodic induction in Japanesequail. J Biol Rhythms 10:17–32
Kasahara T, Okano T, Haga T, Fukada Y (2002) Opsin-G11-mediated signaling pathway for photic entrainment of thechicken pineal circadian clock. J Neurosci 22:7321–7325
Kumar V (1996) Effect of melatonin in blocking the response to askeleton photoperiod in the blackheaded bunting. Physiol Be-hav 59:617–620
Kumar V (1997) Photoperiodism in higher vertebrates—An adap-tive strategy in temporal environment. Indian J Exp Biol35:427–437
Kumar V (2001) Melatonin and circadian rhythmicity in birds. In:Dawson A, Chaturvedi CM (eds) Avian endocrinology. NarosaPublishing House, New Delhi, pp 93–112
Kumar V, Tewary PD (1984) Circadian rhythmicity and the ter-mination of refractoriness in the blackheaded bunting (Ember-iza melanocephala). Condor 86:27–29
Kumar V, Follett BK (1993a) The nature of photoperiodic clock invertebrates. Proc Zool Soc Calcutta; JBS Haldane Com-.memoration Vol. pp 217–227
Kumar V, Follett BK (1993b) The circadian nature of melatoninsecretion in Japanese quail (Coturnix coturnix japonica). JPineal Res 14:192–200
Kumar V, Juss TS, Follett BK (1993) Melatonin secretion in quailprovides a seasonal calendar but not one used for photoperiodictime measurement. In: Touitou Y, Arendt J, Pevet P (eds)
Melatonin and the pineal gland from basic sciences to clinicalapplications. Elsevier Science Publications, Amsterdam, pp163–168
Kumar V, Jain N, Follett BK (1996a) The photoperiodic clock inblackheaded buntings (Emberiza melanocephala) is mediatedby self-sustaining circadian system. J Comp Physiol A 179:59–64
Kumar V, Kumar BS, Jain N (1996b) Effect of late afternoonadministration of melatonin on the photoperiod-induced re-sponses in buntings. Indian J Exp Biol 34:20–225
Kumar V, Singh S, Misra M, Malik S (2001) Effects of durationand time of food availability on photoperiodic responses in themigratory male blackheaded bunting (Emberiza melanocepha-la). J Exp Biol 204:2843–2848
Kumar V, Singh S, Misra M, Malik S, Rani S (2002) Role ofmelatonin in photoperiodic time measurement in the migratoryredheaded bunting (Emberiza bruniceps) and the non-migratoryIndian weaver bird (Ploceus philippinus). J Exp Zool 292:277–286
Kumar V, Singh BP, Rani S (2004) The bird clock: a complexmulti-oscillatory and highly diversified system. Biol RhythmRes 35:121–144
Maitra SK, Dey M (1992) Testicular responsiveness to exogenousmelatonin during different phases of the annual testicular cyclein roseringed parakeet, Psittacula krameri. Eur Arch Biol103:157–164
Malik S, Rani S, Kumar V (2004) Wavelength dependency of light-induced effects on photoperiodic clock in the migratory black-headed bunting (Emberiza melanocephala). Chronobiol Int21:367–384
Okano T, Yoshizawa T, Fukada Y (1994) Pinopsin is a chickenpineal photoreceptive molecule. Nature 372:94–97
Saldanha CJ, Siverman AJ, Silver R (2001) Direct innervation ofGnRH neurons by encephalic photoreceptors in birds. J BiolRhythms 16:39–49
Saxena RN (1964) Sexual cycle and secondary sex characters ofIndian weaver bird (Ploceus philippinus). PhD Thesis, BanarasHindu University, Varanasi, India
Saxena RN, Thapliyal JP (1962) Male hormone and bill pigmen-tation in baya, Ploceus philippinus (Linn.). Sec Cong Zool(Abstr 29)
Sumova A, Sladek M, Jac M, Illnerova H (2002) The circadianrhythm of Per1 gene product in the rat suprachiasmatic nucleusand its modulation by seasonal changes in day length. BrainRes 947:260–270
Takahashi JS, Menaker M (1982) Role of suprachiasmatic nuclei inthe circadian system of the house sparrow, Passer domesticus. JNeurosci 2:815–828
Thapliyal JP, Saxena RN (1961) Plumage control in Indian weaverbird (Ploceus philippinus). Naturwissenschaften 24:741–742
Thapliyal JP, Tewary PD (1964) Effect of light on the pituitary,gonad and plumage pigmentation in the avadavat, Estrildaamandava, and Baya weaver, Ploceus philippinus. Proc Zool SocLond 142:67–71
Trivedi AK (2004) Seasonal responses of house sparrow (Passerdomesticus) Linnaeus at 27�N. PhD Thesis, University of Luc-know, Lucknow, India
Underwood H (1994) The circadian rhythm of thermoregulation inJapanese quail. I. Role of the eyes and pineal. J Comp PhysiolA 175:639–653
Wilson FE (1991) Neither retinal nor pineal photoreceptorsmediate photoperiodic control of seasonal reproduction inAmerican tree sparrows. J Exp Zool 259:117–127
Zimmerman NH, Menaker M (1979) The pineal gland: a pace-maker within the circadian system of the house sparrow. ProcNatl Acad Sci USA 76:999–1003
364


















