
The Physical State of Potassium in Frog Skeletal Muscle ... · PDF fileThe Physical State of...
19
Physiol. Chem. Phys. & Med. NMR (2014) 43: 75–93 The Physical State of Potassium in Frog Skeletal Muscle Studied by Ion-Sensitive Microelectrodes and by Electron Microscopy: Interpretation of Seemingly Incompatible Results L. Edelmann Medizinische Biologie, Fachbereich Theoretische Medizin, Universität des Saarlandes, D-6650 Homburg/Saar, FRG Abstract: According to the commonly accepted membrane pump theory most of cellular K + ions are freely dissolved in free cellular water; the alternative association-induction hypothesis postulates that the bulk of cellular K + is adsorbed (weakly bound) to cellular proteins that are maintained in a specific labile state in the cytoplasm of a living cell. K + activities measured with ion-sensitive mi- croelectrodes in the cytoplasm of frog skeletal muscle seem to confirm the claim that most of cel- lular K + ions are free in cellular water. On the other hand, it is evident from electron microscopic ion binding studies that in frog skeletal muscle most of cellular K + ions are adsorbed to cellular pro- teins. The conflicting results can be explained with the assumption that a damage of the cytoplasm caused by the impaling microelectrode leads to a liberation of adsorbed ions. Using the light mi- croscope tests the possibility that microelectrodes damage the muscle cytoplasm. It is found that mi- croelectrodes produce visible traumas that increase with time. Electron microscopic ion binding studies with damaged muscle support the view that monovalent cations are liberated in the disturbed area of a muscle fiber. It is concluded that a K + -sensitive microelectrode is not suited to determine the concentration of free K + ions in intact frog skeletal muscle. KEY WORDS: Membrane pump theory, association-induction hypothesis, potassium binding, ion localization, ion-sensitive microelectrodes, striated muscle, freeze-substitution, low temperature embedding.
Transcript of The Physical State of Potassium in Frog Skeletal Muscle ... · PDF fileThe Physical State of...

Physiol. Chem. Phys. & Med. NMR (2014) 43: 75–93
The Physical State of Potassium in FrogSkeletal Muscle Studied by Ion-Sensitive
Microelectrodes and by Electron Microscopy:Interpretation of Seemingly Incompatible Results
L. Edelmann
Medizinische Biologie, Fachbereich Theoretische Medizin, Universität des Saarlandes, D-6650 Homburg/Saar, FRG
Abstract: According to the commonly accepted membrane pump theory most of cellular K+ ionsare freely dissolved in free cellular water; the alternative association-induction hypothesis postulatesthat the bulk of cellular K+ is adsorbed (weakly bound) to cellular proteins that are maintained in aspecific labile state in the cytoplasm of a living cell. K+ activities measured with ion-sensitive mi-croelectrodes in the cytoplasm of frog skeletal muscle seem to confirm the claim that most of cel-lular K+ ions are free in cellular water. On the other hand, it is evident from electron microscopicion binding studies that in frog skeletal muscle most of cellular K+ ions are adsorbed to cellular pro-teins. The conflicting results can be explained with the assumption that a damage of the cytoplasmcaused by the impaling microelectrode leads to a liberation of adsorbed ions. Using the light mi-croscope tests the possibility that microelectrodes damage the muscle cytoplasm. It is found that mi-croelectrodes produce visible traumas that increase with time. Electron microscopic ion bindingstudies with damaged muscle support the view that monovalent cations are liberated in the disturbedarea of a muscle fiber. It is concluded that a K+-sensitive microelectrode is not suited to determinethe concentration of free K+ ions in intact frog skeletal muscle.
KEY WORDS: Membrane pump theory, association-induction hypothesis, potassiumbinding, ion localization, ion-sensitive microelectrodes, striated muscle, freeze-substitution,low temperature embedding.

Introduction
THE commonly taught membrane pump theory (MPT) is based on the assumption thatmost cellular water and ions exist in a physical state that is essentially a dilute aqueoussolution. According to an alternative theory, the association-induction hypothesis (AIH)of Ling [31, 38] the cell represents a metastable coop erative protein-ion-water complex.Proteins, water and solutes exist in a physical state different from that of an aqueous pro-tein salt solution. In particular, it is assumed that most of the cellular K+ ions are adsorbedto β- and γ-carboxyl groups of cellular proteins and that the bulk of the cellular water isdifferently structured than extracellular free water. This controversial issue is addressed ina recently published book entitled, “The State of Water in the Cell” (W. Negendank, L.Edelmann, (eds.), Scanning Microscopy Intl., AMF O’Hare, Chicago 1988, 1–113). Incriticizing the predominance of the AIH throughout this book a reviewer wrote the fol-lowing [49]: “It is amazing that, at a time where ion selective microelectrodes can directlymeasure free ion contents and find a majority of potassium to be in free form, Ling’s hy-pothesis is endorsed and tacitly expanded to a generalized hypothesis by some of the con-tributors without major objections”.It remains, however, an open question as to what extent ion-sensitive microelectrodes
are suited to detect free and adsorbed K+ ions in living cells. According to the AIH the cy-toplasm is a highly sensitive protein-ion-water system that is disturbed by an impaling mi-croelectrode. As a consequence K+ ions may be liberated from adsorption sites and themicroelectrode is then detecting artefactual high K+ ion concentrations [38, 47]. So farthis argument is not widely recognized; and it is usually believed that a K-sensitive mi-croelectrode impaled into the cytoplasm is detecting the same concentration of free K+
ions as it exists in intact cells, provided the microelectrode is perfectly sealing the punc-tured cell membrane [17, 56].It is evident that the above-mentioned critique is justified if it is proven that in first ap-
proximation the cytoplasm is an aqueous protein salt solution as described by the MPT.In the author’s opinion such proof is missing. On the contrary, recent electron microscopicstudies with frog skeletal muscle have led to the conclusion that most of cellular K ionsare adsorbed to cellular proteins of the muscle — a conclusion opposite to the expecta-tions of the MPT [11, 14] and also opposite to conclusions derived from studies with K-sensitive microelectrodes [26–30, 56]. Because of this conflict which eventually may besolved by interpretations provided by the AIH new experiments were devised to test theAIH and to answer the following questions: 1) Do microelectrodes disturb locally the cy-toplasmic structure in skeletal muscle of the frog. 2) Is it possible to detect by electronmicroscopic methods a liberation of alkali-metal ions from adsorption sites of the sar-coplasm, which has been mechanically injured?In order to enable the reader to understand why the AIH is chosen as a working hy-
pothesis an Appendix is presented which contains a critical comparison between AIH andMPT.
76 LUDWIG EDELMAN

Materials and Methods
Experiments with microelectrodes
Single intact skeletal muscle fibers were prepared at room temperature from semitendi-nosus muscles of Northern American leopards frogs (Rana pipiens pipiens, Schreber). Thecaput posticum of a semitendinosus muscle was fixed on a plastic frame kept in Ringer’ssolution in a Petri dish. Under the control of a stereo light microscope most of the fiberswere cut and removed in such a way that only a few single fibers remained fixed at thetendons of the muscle. During the dissection the Ringer’s solution was frequently changedand before and after the microelectrode experiment (see below) the single fibers weremonitored for viability by electrical stimulation. The frame with the mounted single fiberswas transferred (still in the Petri dish) into a small container the bottom of which wasmade of a glass slide normally used for light microscopy (Figure 1a). This container wasthen transferred to a light microscope (Zeiss, Standard 14) and the fibers were inspectedwith phase contrast optics (objective: Ph2, Neofluar 40/075). Glass microelectrodes witha tip diameter of about 0.3 mm were drawn on an electrode puller and mounted to a mi-cromanipulator. Under visual control a microelectrode was inserted into a single musclefiber (Figure 1b). Photographs were taken after different lengths of time. During the ex-periment the Ringer’s solution bathing the muscle fibers was constantly renewed as shownin Figure 1b. Altogether 10 different single fibers were impaled with microelectrodes. Insome experiments the tips of the microelectrodes were sealed with molten wax; theseelectrodes produced the same results as electrodes with open-ended tips.
Experiments with freeze substituted and low temperature embedded muscles
Under sterile conditions frog sartorius muscles (Rana pipiens pipiens, Schreber) were in-cubated at room temperature for 4 days in K+- free Ringer-GIB medium [40] containing2.5 mM Cs+ as described by Ling and Bohr [41]. About 80% of cellular K+ was then re-placed with Cs+ [6]. A muscle was transferred to a humidity chamber (100% humidity) inwhich adhering fluid was gently removed with a wet filter paper. The muscle was cut with
PHYSICAL STATE OF POTASSIUM IN FROG SKELETAL MUSCLE 77
FIGURE 1. Muscle preparation for microelectrode experiments. (a) Top view of a plastic frame Fwith mounted single fibers M in a small container C. (b) Cross-section of container C with musclepreparation under the objective O of a light microscope; a microelectrode E is inserted into a musclefiber. Arrows indicate a flow of Ringer’s solution S through stainless steel tubes T into and out ofcontainer C.

a razor blade perpendicular to the direction of the muscle fibers. After 30 min the twopieces of the cut muscle were rapidly frozen as described elsewhere [15]. The 30 min waitbetween cutting and freezing was arbitrarily chosen to produce a visible damage of themuscle cytoplasm at the cut end (see Discussion). Frozen muscle pieces were freeze sub-stituted in pure acetone at −80°C for one week and low temperature embedded at −60°Cin Lowicryl K11M as described elsewhere [15]. Intact K+-containing and Cs-loaded mus-cles were cryofixed, freeze substituted and low temperature embedded in the same way.Diatome diamond knives were used for obtaining ultrathin, 0.1 µm thick and 0.2 µm thickwet-cut sections, glass knives for 0.3 µm thick dry-cut sections. Ultrathin sections werestained with uranyl acetate and lead citrate. 0.1 µm thick and 0.2 µm thick sections wereexposed to a staining solution containing 100 mM LiCl and 10 mM CsCl as describedelsewhere [11].
Results
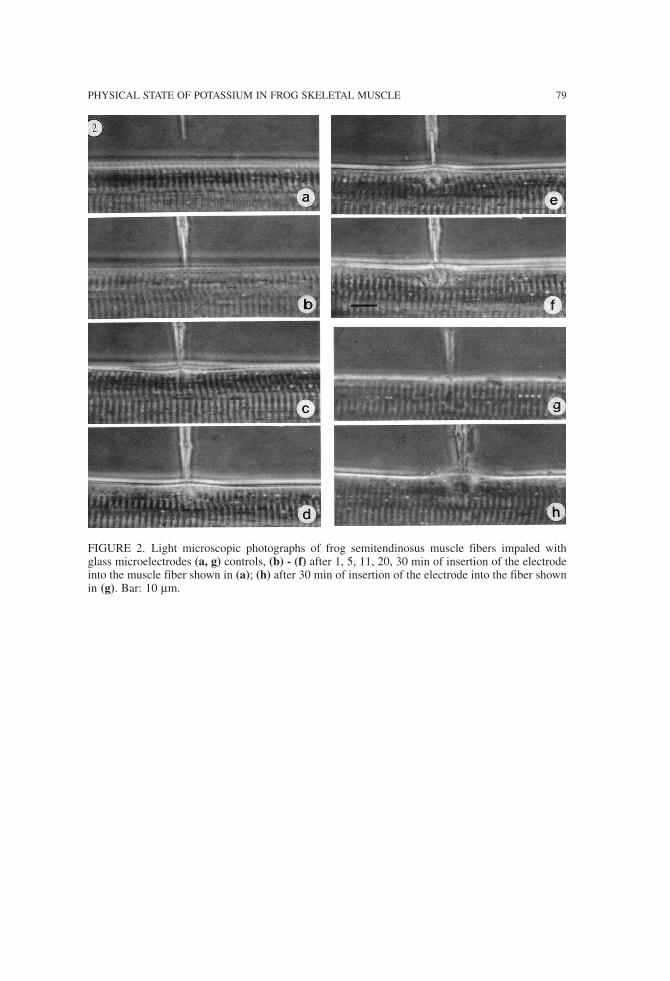
Typical results of the microelectrode experiments are shown in Figure 2. The impaledfibers show a spreading disturbance of the muscle structure. After 30 min (Figures 2f, h)the disturbed area has a diameter of several sarcomeres. Specific features of the artifactsproduced by the microelectrode are:
1) The impaled muscle fibers show signs of local contraction (e.g., Figures 2d, e). 2) At the place of the impalement one observes a local swelling of the disturbed area(e.g., Figures 2e, f, and h).
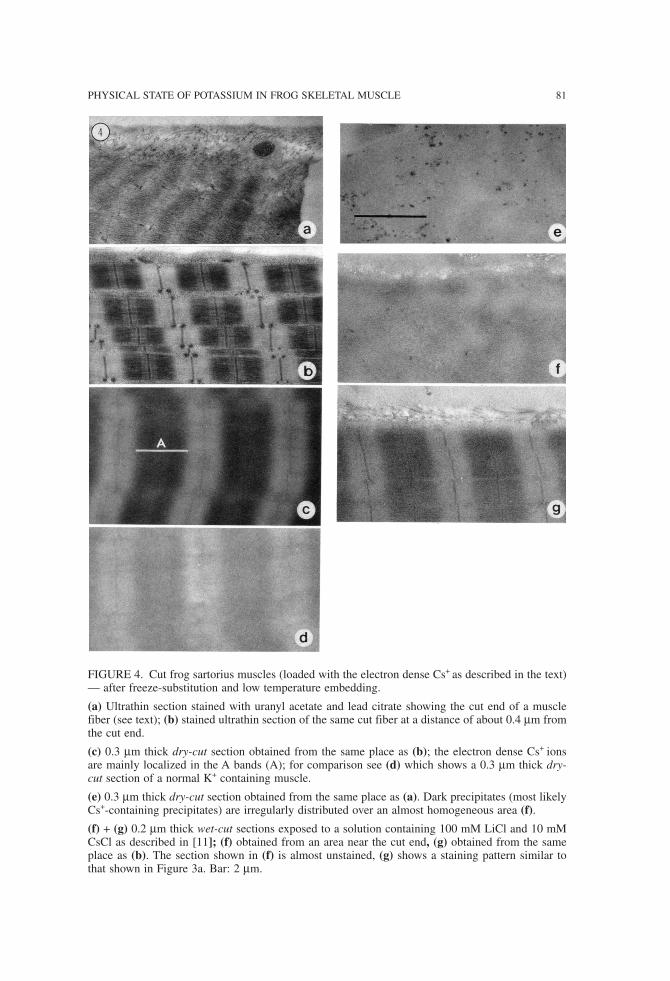
Figure 3 shows a typical result of a 0.1 µm thick section stained with a solution con-taining 100 mM LiCl and 10 mM CsCl. The electron-dense Cs ions preferentially stainproteins of the A band and of the Z line. The ultrastructure of the muscle cannot be seenin unstained sections (Figure 3b). Results obtained with a cut sartorius muscle are shown in Figure 4. Muscle contrac-
tion can be observed near the cut end (Figure 4a) whereas most or the fiber appears in aperfect resting state (Figure 4b). In the undisturbed parts of the muscle the electron-denseCs+ ions are localized preferentially in the A bands and at Z lines as can be demonstratedwith dry-cut sections (Figure 4c). For comparison see Figure 4d, which has been obtainedfrom an intact K+-containing muscle and which shows a rather poor contrast between Aband and I band regions. A dry-cut section of a contracted part of the cut sartorius muscleshows dark precipitates irregularly distributed over an almost homogeneous area, the pe-riodicity of the contracted sarcomeres is barely visible (Figure 4e). Figures 4f and g showwet-cut 0.2 µm thick sections “stained” with the LiCl-CsCl solution. The section obtainedfrom the damaged part near the cut end (Figure 4f) appears almost unstained (no Cs+-binding) whereas the section of an intact area of the same muscle fiber (Figure 4g) isstained similar to the section shown in Figure 3a.
78 LUDWIG EDELMAN
PHYSICAL STATE OF POTASSIUM IN FROG SKELETAL MUSCLE 79
FIGURE 2. Light microscopic photographs of frog semitendinosus muscle fibers impaled withglass microelectrodes (a, g) controls, (b) - (f) after 1, 5, 11, 20, 30 min of insertion of the electrodeinto the muscle fiber shown in (a); (h) after 30 min of insertion of the electrode into the fiber shownin (g). Bar: 10 µm.

Discussion
Experimental determination of the physical state of K+ in living frog skeletal muscle
Results obtained with ion-sensitive microelectrodes. Experimental studies on frogskeletal muscle of Hill [22], Fenn [19] and Gersh [20] provided indirect evidence for theview that virtually all water and K+ ions exist in the free state in living cells. With the de-velopment of ion sensitive microelectrodes it was expected that free and bound cellularK+ could be determined unequivocally [18]. Experimental testing of nerve and musclecells yielded the following results: the intracellular K+ activity agreed in first approxima-tion with the intracellular K+ concentrations multiplied by an activity coefficient equal tothat of K+ in an aqueous solution of ionic strength similar to that expected in living cells(e.g., [23, 28, 30]. The basic tenet of the MPT — virtually all K+ is in free solution —seemed to be confirmed. However, Ling criticized the results in the following way [33]:An ion-sensitive microelectrode can monitor only the ionic activity in a microscopicallythin layer of fluid in immediate contact with the microelectrode tip. Thus even though thebulk of the cytoplasm may be in a perfectly good state of health and its K+ in a normalphysiological state, the inserted ion- sensitive microelectrode cannot “see” that K+. It canonly detect the activity of K+ in the microscopic portion of the cytoplasm that must havebeen forcibly torn apart to make room for the impaling ion-sensing electrode. Therecorded activity is therefore that of a disturbed cytoplasm and not that of normal cyto-plasm. In response to this argument Dick and McLaughlin [4] agreed that trauma couldindeed liberate K+ ions but they pointed out that such liberated K+ would soon diffuseaway while in actual measurements the K+ activity remained more or less the same for aslong as 30 min. This view was not accepted because of the following two reasons [47]: 1)Experiments with K+ containing droplets injected into squid axons show that the diffusionof free K+ inside the cytoplasm is so slow that it can hardly be detected within 30 min.2) The disintegration of the cytoplasm caused by the impaling electrode is spreading pro-gressively to healthier regions during the experiment; thereby additional liberation of K is
80 LUDWIG EDELMAN
FIGURE 3. −0,1 µm thick wet-cut sections of frog sartorius muscles after freeze-substitution andlow temperature embedding, (a) without any staining, (b) exposed to a solution containing 100 mMLiCl and 10 mM CsCl as described in [11] p. 884. Mainly myosin filaments of the A band (A) andproteins of the Z line (Z) bind the electron dense Cs+. Bar: 1 µm.
PHYSICAL STATE OF POTASSIUM IN FROG SKELETAL MUSCLE 81
FIGURE 4. Cut frog sartorius muscles (loaded with the electron dense Cs+ as described in the text)— after freeze-substitution and low temperature embedding.
(a) Ultrathin section stained with uranyl acetate and lead citrate showing the cut end of a musclefiber (see text); (b) stained ultrathin section of the same cut fiber at a distance of about 0.4 µm fromthe cut end.
(c) 0.3 µm thick dry-cut section obtained from the same place as (b); the electron dense Cs+ ionsare mainly localized in the A bands (A); for comparison see (d) which shows a 0.3 µm thick dry-cut section of a normal K+ containing muscle.
(e) 0.3 µm thick dry-cut section obtained from the same place as (a). Dark precipitates (most likelyCs+-containing precipitates) are irregularly distributed over an almost homogeneous area (f).
(f) + (g) 0.2 µm thick wet-cut sections exposed to a solution containing 100 mM LiCl and 10 mMCsCl as described in [11]; (f) obtained from an area near the cut end, (g) obtained from the sameplace as (b). The section shown in (f) is almost unstained, (g) shows a staining pattern similar tothat shown in Figure 3a. Bar: 2 µm.

to be expected. These arguments, however, were not further considered and Edzes andBerendsen wrote in 1975 [17]: “The experimental results point to little or no binding ofthe alkali cations and certainly not to a strong preferential K+ binding”. This statementincludes results obtained with K+-sensitive microelectrodes and frog skeletal muscle[26–30].Results obtained with electron microscopic methods. Stimulated by the controversy
between AIH and MPT electron microscopic experiments were designed to investigate thephysical state of K+ in living cells by methods that avoid mechanical disturbance of thecytoplasm. The idea was to test the following predictions of the MPT and the AIH con-cerning the localization of K+ in the striated muscle: According to the MPT K+ ions arefreely dissolved in the free water of the striated muscle cells, and their localization fol-lows the water distribution. Since the water content in the I band is higher than in the Aband [25] the membrane theory would predict a higher amount of free K+ ions in the Iband than in the A band. The AIH, on the other hand, predicts a higher accumulation ofK+ in the A band compared to the I band because β- and γ-carboxyl groups are primarilyfound on myosin in the A bands [35]. These opposing predictions should also hold for theelectron dense Rb+, Cs+, and Tl+ as these ions accumulate in frog skeletal muscle bymeans of the same mechanism as K+; they replace each other reversibly in a mole-for-mole fashion under physiological conditions [34, 41]. This implies that we can tackle thebasic problem of cation binding in muscle with all four different cations. For example, ifabout 80% of cellular K+ has been replaced by Cs+ or Tl+, which are then found to bebound at cellular proteins, we must conclude that also K+ is bound to the same proteinsin a normal K+ containing muscle.Starting in 1976 several newly developed cryotechniques have been used to localize al-
kali- metal ions and Tl+ in the striated muscle of the frog by electron microscopic meth-ods. Either muscles with their normal K+ content or muscles in which about 80% of thecellular K was replaced by Rb+, Cs+ or Tl+ have been investigated. The methods used in-clude analysis of sections of freeze-dried [6] or freeze- substituted [14] and embeddedmuscle, autoradiography of frozen-hydrated single fibers using 86Rb and 134Cs [7], elec-tron probe X-ray microanalysis of freeze-dried cryosections [10], and visualization of Tl+
in frozen-hydrated cryosections [12], (for reviews see [11, 14]).The main findings and conclusions of these studies are the following: In the normal K+
containing frog skeletal muscle and in muscle loaded with the electron-dense surrogatesRb+, Cs+ or Tl+ the accumulated ions are preferentially localized within the A bands, es-pecially at the 2 marginal regions and at the Z lines. These results fulfill the expectationsof the AIH and are not in accordance with the predictions of the MPT. Of particular arethe results obtained with frozen-hydrated preparations:Autoradiographs visualizing the Cs+-distribution in a Cs+-loaded muscle cell (Figure
5a, b) show that the concentration of Cs+ is low in the I bands and high in the A bands[7]. Since the concentrations of free Cs+ in the cellular water must be equal in the A bandsand in the I bands one can only conclude that the concentration of the cellular alkali-metalion in the cellular water is low. 2). Micrographs of frozen hydrated cryosections of T1loaded muscle (Figure 6b) show that individual filaments (mainly myosin filaments in theA band, but also I band filaments) and Z line proteins are “stained” by Tl+ [14]; this im-plies that most of the cellular Tl+ ions are bound to proteins and not dissolved in the sur-rounding water: otherwise a very poor contrasting or even a negative staining of proteins
82 LUDWIG EDELMAN

PHYSICAL STATE OF POTASSIUM IN FROG SKELETAL MUSCLE 83
would have to be expected. Of utmost importance is the fact that these results have beenobtained with fully hydrated cryosections. The sections have been kept at about −170 Cand photographed in a Zeiss EM 10CR at a magnification of 5000 (electron exposurebelow 1000 e-/nm2). No mass loss of the area irradiated during photographing could bedetected. From these electron microscopic studies it was concluded that most of cellularcations are bound to cellular proteins. A recent quantitative X-ray microanalysis concern-ing the localization of K+ in rat heart muscle cells led to a similar conclusion [55].Explanation of seemingly incompatible results. The experimental findings presented
in Figure 2 give an explanation for the conflicting results obtained with the described dif-ferent techniques. This explanation is essentially that predicted by Ling et al. [47]: A mi-croelectrode cannot be inserted into a muscle cell without disturbing the cytoplasmicultrastructure in the vicinity of the microelectrode. Furthermore, the trauma does not havea stationary localization around the microelectrode but spreads with time. For instance,after 11 min the disturbed area seen in Figure 2d will exceed the area shown in Figure 6b.A reasonable explanation for the high activity of K+ seen by a K+-sensitive microelectrodeis that K+ ions are released from disturbed cytoplasmic proteins and freely dissolved inthe free water of the disturbed area. The observed local swelling at the place of deterio-
FIGURE 5. Autoradiographs of frozen hydrated frog muscle fibers, (a) Light microscopic 134Cs au-toradiogram of a stretched Cs+-loaded fiber, (b) Electron microscopic 134Cs autoradiogram of astretched Cs+-loaded fiber. The sarcomere length is about 4.4 µm. A line of silver grains betweentwo dark bands (A bands) indicates the Z line (arrow). (c) Electron microscopic 86Rb autoradiogramof a stretched Rb+-loaded fiber. The sarcomere length is about 3.3 µm. Arrows indicate dark linesat the outer edges of an A band. From [7], reprinted by permission.

ration may be caused by the increasing concentration of liberated ions in the disturbedarea with the result of water movement towards this place of reduced water activity.The observed progressive deterioration of the cytoplasm around the microelectrode may
explain why K+-sensitive microelectrodes did not detect large variations in K+-activities;such variations would be expected if a small microelectrode could detect the K+ ions lib-erated only from an A band region (high value) or from an I band region (low value). Onemay speculate that low K+-activities have been detected at the beginning of microelectrodeexperiments (when the damage was still small) and that such findings were neglected be-cause they were attributed to improper sealing of the cell membrane around the micro-electrodes. It could be worthwhile to repeat K+ activity measurements with a simultaneouscontrol of the produced damage to clarify this issue. The possibility however that the highstable K+ activities reported in past microelectrode work were obtained with microelec-trodes which produced no structural artifacts is very unlikely for the following reasons: 1) The microelectrodes used for the present investigations were smaller (tip diameter about0.3 µm) than those used for K+ activity measurements in frog skeletal muscle (e.g., [30],tip diameter up to 1.5 µm). With no exception even these small microelectrodes producedstructural artifacts, which were visible under the light microscope. 2) The center of deteri-oration produced by the microelectrode is near the electrode inside the cell and not at theplace of the membrane where the microelectrode is inserted (see e.g., Figure 2e). Thismeans that the damage is not starting from an improper sealed cell membrane.
Ion adsorption in intact and damaged muscle cytoplasm studied by electron microscopy
Ion adsorption in intact skeletal muscle cells has been established by the above discussedelectron microscopic methods. Independent evidence for the view that cytoplasmic pro-teins do weakly bind or adsorb alkali-metal ions has been obtained with the following invitro experiments: Sections of freeze-dried embedded muscle exposed to alkali cation so-lutions show a selective binding of the different ions (e.g., K+. Rb+, Cs+) by the same pro-teins which accumulate the ions in the living cell [8, 9, 11]. Figure 3b shows that freezesubstituted and low temperature embedded muscle can also be used to demonstrate thisphenomenon: The electron-dense Cs+ ions stain the ultrathin section because they are ad-sorbed preferentially to filaments of the A band and to Z line proteins. The ion adsorptionsites are the same as those visualized in frozen hydrated cryosections of Tl+-loaded intactmuscles (Figure 6b). It is noteworthy that proteins of a glutaraldehyde fixed muscle do notbind alkali-metal ions [13]. These results confirm the postulation of the AIH that cellularproteins are able to adsorb alkali-metal ions but only if they are maintained in certain con-formations similar to those which they assume in the living cell. Apparently freeze-sub-stitution and embedding carried out exclusively at rather low temperatures can be used tocapture the capability of proteins to adsorb alkali-metal ions.In the foregoing section we explained the high K+ activity measured by microelectrodes
as being due to ions, which were liberated from proteins of the disturbed cytoplasm. Inother words, we expect that proteins of a disturbed muscle cytoplasm change their con-formation in such a way that alkali-metal ions like K+ or Cs are no longer preferentiallyadsorbed. The best way of testing this possibility would be to freeze very rapidly a musclecell together with an inserted microelectrode, to analyze the K distribution around theelectrode in a frozen hydrated preparation and to determine the ion binding capacity of
84 LUDWIG EDELMAN

proteins around the electrode after freeze substitution and low temperature embedding.Because of technical difficulties we have chosen to start with a simpler method. FromEMOC (effectively membrane-less open-ended cell) studies it is known that after cuttingof a skeletal muscle cell by a razor blade the cytoplasm at the cut end is deteriorating pro-gressively, thereby changing its physicochemical properties [37, 43]. With the electronmicroscopic cryo-methods now available it is possible to investigate the events occurringduring this deterioration under different conditions. First experimental findings obtainedfrom a cut Cs+-loaded muscle are given in Figure 4 and described above. The followingobservations are relevant to the problem of ion adsorption in damaged muscle cytoplasm:
1) The subcellular distribution of electron- dense Cs+ ions is completely different in thedisturbed area near the cut end compared to an area where the muscle appears in a nor-mal resting state (compare Figure 4e with Figure 4c). The disturbed area shows a poorcontrast and some dark precipitates which most likely represent Cs - precipitates. Whetherthe poor contrast is caused by an even distribution of Cs+ ions or whether the Cs+ con-centration is very low in the disturbed area of the freeze substituted and low temperatureembedded preparation remains to be determined by future X-ray microanalytical studies.(It has been speculated that ions, which are freely dissolved in cellular water, cannot be
PHYSICAL STATE OF POTASSIUM IN FROG SKELETAL MUSCLE 85
FIGURE 6. Frozen hydrated cryosections of frog sartorius muscle, (a) Normal K+ containingmuscle. Only very faint ultrastructural details can be seen. A, A band; Z, Z line; (b) Tl+-containingmuscle. Dark myosin filaments (arrows) in the A bands (A) and dark Z lines (Z) indicate sites ofpreferential Tl+ accumulation in the living cell. Bar: 1 µm. From [14], reprinted by permission.

retained as easily as weakly bound ions in the biological specimen during freeze substi-tution and low temperature embedding [16]). In any case it can be concluded that a re-distribution of the electron-dense ions must have occurred. Since the disturbed area showsmuscle contraction (Figure 4a) a finding observed in earlier studies with intact contract-ing muscles is noteworthy [15, 16]. These muscles loaded either with Cs or Tl+ beforefreezing during contraction showed also ion redistribution; this phenomenon confirmed apostulate of a new model for the contraction of living muscle namely that alkali-metalions (e.g., K+ions in a normal K+-containing muscle) are liberated from their original ad-sorption sites during contraction ([38], chap. 16). With these findings and the observationthat muscles impaled with microelectrodes show a local contraction (see Figure 4e) wehave an experimental confirmation for the view that an impaling microelectrode causes alocal liberation of adsorbed ions.2) The disturbed area cannot be stained with the electron-dense Cs+ ions (Figure 4f) as
is the case with intact muscle fibers (Figure 3a) or with intact areas of a cut muscle fiber(Figure 4g). This suggests that proteins of the damaged cytoplasm have lost their abilityto adsorb (or bind) alkali-metal ions. If the ion adsorption capacity of proteins is greatlyreduced during damaging of the cytoplasm an increase of free ions in the cellular wateris inevitable.
Conclusion
Theoretical considerations and experimental findings obtained with electron microscopiccryotechniques support the view that the bulk of muscle K+ is adsorbed (weakly bound)to cellular proteins and that this binding can only be detected by methods, which do notdisturb the cytoplasm. Ion sensitive microelectrodes, which impale the cell, produce arti-facts in the cytoplasm and cannot be used to evaluate free and adsorbed ions in intactskeletal muscle.
Appendix
Association-induction hypothesis versus membrane pump theory
According to the classical membrane theory the cell interior is seen as a protein contain-ing solution of free ions in free water separated from the external environment by a verythin cell membrane. In the early 1940’s the membrane theory gained a high degree of pub-lic acceptance when it was able to explain with a few basic postulates the following fourmain physiological properties of living cells [1]: selective solute accumu lation and exclu-sion, selective permeability, volume changes, and cellular electrical potentials. However,a basic assumption of the classical membrane theory was found to be incorrect when thecell membrane was shown to be permeable to Na+ ions [53]. In order to explain the ob-served low cellular Na+ ion concentrations it was necessary to postulate an energy con-suming Na+ pump situated in the cell membrane [3]. Soon it became evident that moreand more pumps had to be postulated in order to understand the observed asymmetric dis-tribution between the inside and outside of living cells of many other substances (for re-view see [47]).
86 LUDWIG EDELMAN

The fundamental correctness of the new membrane pump theory (MPT) was doubtedwhen Ling calculated the energy needed for the Na+ pump to maintain the observed lowcellular Na+ concentration in frog skeletal muscle cells; Ling found the energy require-ments of the Na+ pump under conditions where the energy sources had been blocked weresuch that the Na+ pump alone would consume 15 to 30 times as much energy as the en-tire amount that the cell commands ([31], chap. 8). This finding led Ling to develop a the-ory for a molecular mechanism for the selective accumulation of K+ over Na+ in livingcells, which is not based on hypothetical ion pumps (for review see [31]). Later Ling pre-sented a generalized theory of the living cell called the association-induction hypothesis(AIH) based on the following 3 concepts ([38], p. 375):(Cl) “The bulk of cellular water exists in a state of polarized multilayers; in this state
water tends to exclude solutes and does so to variable degrees depending on the size andcomplexity of the solute. This provides the mechanism for the normal exclusion of Nafrom most cells.”(C2) “Solutes are accumulated by the cell if they are adsorbed onto macromolecules
within the cell; for example, cations are adsorbed onto fixed carboxyl groups and sugarsonto hydrogen bonding qroups of proteins. This provides the mechanism for the normalaccumulation of K+ by most cells.” (C3) “The polypeptide chain is especially well suited for the induction of electron dis-
tribution changes from one side chain to another. This underlies the interaction betweensites that adsorb solutes, permitting them to function in a cooperative manner, and it un-derlies the ability of cardinal adsorbents (e.g., ATP, hormones, drugs) to affect a largenumber of sites in an allosteric manner”. According to the current version of the MPT the above mentioned four basic phenom-
ena of living cells are interpreted as follows: Selective accumulation and exclusion are theresult of energy consuming active and passive transport mechanisms situated in the cellmembrane. Selective permeability is explained as follows: The cell membrane is seen asan envelope made of lipids in which complex charged and uncharged pores are incorpo-rated as well as carriers or other active and passive transport mechanisms that are re-sponsible for the translocation of ions, sugars, amino acids, and other substances acrossthe membrane. Cell volume regulation is determined primarily by the osmotic pressureexerted by solutes that are freely dissolved within free cellular water. K+ - the main cel-lular cation - is thought to be the main solute that helps to balance the osmotic pressureof the cell interior and of the extracellular medium. According to the MPT an intact cellmembrane is essential for the maintenance of cell volume. The electrical potentials of liv-ing cells are membrane diffusion potentials, which can be described by the extra- and in-tracellular concentrations and the membrane permeabilities of certain ions. For instance,the resting potential of nerve and muscle cells is in first approximation a K+-diffusion po-tential as described by the Hodgkin-Katz theory [24]. According to the AIH the four cellular phenomena are quantitatively described by four
sets of equations [39] and interpreted as follows [38, 39]: Solute exclusion is the result ofcellular water properties (see C1) and solute accumulation is due to binding onto cellularmacromolecules (see C2). In most cases this binding is very weak (adsorption) and de-pendent on the very labile state of the cellular protein-ion-water complex (fixed chargesystem). Energy is necessary to maintain the metastable state of the cytoplasm but energyis not required to move solutes into and out of the cell. Selective permeabilities of ions,
PHYSICAL STATE OF POTASSIUM IN FROG SKELETAL MUSCLE 87

sugars, and amino acids are due to cell surface (or membrane) properties. The cell surfaceis seen as a fixed charge system (organized differently than the cytoplasm), which con-tains interstices filled with multilayers of polarized water and proteins with potential ad-sorption sites for ions, sugars, amino acids, and other substances. The movement ofsolutes into and out of the cell may occur via diffusion through the water and/or via ad-sorption and desorption from proteins or other macromolecules (for different views oncell membrane properties according to MPT and AIH see [36]). Cell volume regulation isdetermined by three factors: 1) the tendency of certain macromolecules to build up sev-eral layers of water dipoles (multilayer expansion), 2) the restrictive forces provided bysalt linkages between fixed cationic and anionic groups of cellular proteins (salt linkagerestraint), and 3) the disparity between the particles dissolved in the external solution andthe lower concentration of particles dissolved in the multilayer water of the cytoplasm. (Itis important to realize that according to the AIH the concentration of free K+ and otherions in the cytoplasm is very low and that the decrease in the cell water activity to thevalue that is found in the extracellular fluid is mainly caused by the electrostatic influenceof certain proteins on the cellular water). Cellular electrical potentials are seen as phaseboundary potentials between the cell surface and the extracellular phase and have no di-rect relation to ionic permeabilities. The electrical potential is determined by the densityand nature of the ionic groups on the macromolecules of the cell surface. Taken together the AIH is a general theory based on a few postulations, which is able
to explain the four basic phenomena of living cells in a consistent manner. One may askwhy the theory is not generally accepted or at least generally discussed. The main reasonis probably the strong conviction of most scientists that ion pumps have been proven un-equivocally and that therefore neither the energy argument of Ling nor the logical conse-quence - a model without energy consuming membrane pumps - can be correct.However, are membrane situated pumps and in particular ion pumps responsible for K+
accumulation in and Na+ exclusion from living cells really proven? Many scientists be-lieve that the following observations prove that the Na,K- ATPase - first isolated by Skouin 1957 [52] - is the postulated ion pump (for review see [38], pp. 118–119): 1) Both sys-tems are present in the cell membrane (the ATPase is in the membrane, the postulatedpump must be in the membrane). 2) Both systems utilize ATP but not inosine triphos-phate. 3) Both systems require the presence of Na+ and K+. 4) Both systems require thesame concentration of cations for half-maximal activity; ATPase activity and cation fluxare significantly correlated. 5) Both systems are inhibited by cardiac glycosides.As a consequence of these findings most scientists accepted ouabain inhibition and ATP
induced increase in ion flux as an identification for active transport. In Best and Taylor’stextbook of physiology we read: “An active transport process is defined, not by demon-strating that flux is thermodynamically uphill, but only by demonstrating that flux is cou-pled to metabolism” [51]. This definition, by statement, however is useless for scientificmethods because it is not clear what fundamental assumptions are basic to the definition,hence it cannot be tested [21].Despite this misleading definition of active transport it should have been possible to test
the claim that the Na.K-ATPase is able to transport ions against electrochemical gradients.Indeed, several attempts were undertaken to prove such an active transport with puremembrane preparations. Unfortunately these attempts either failed or the results were atbest equivocal. For instance, the perfused squid axon contains functioning Na,K-ATPaseand is able to increase efflux by addition of ATP, and the efflux is sensitive to ouabain;
88 LUDWIG EDELMAN

however, a net Na+ efflux against an electrochemical gradient could not be observed (forreview see [38], p. 127). This “negative” experiment shows that the good correlation be-tween ATPase activity and ion fluxes does not reflect the postulated pumping activity ofthis enzyme but may reflect the hypothesis of Ling that configuration changes of the AT-Pase change its interactions with ions and water with the result of modified ion perme-ability rates. Similar results were obtained with nonleaky “white” ghosts obtained fromred blood cells. These ghosts are not able to accumulate K+ or extrude Na+ despite the factthat they contain normally functioning Na,K- ATPase [45]. The claim that reconstitutedpurified phospholipid-ATPase vesicles pump Na+ has been analyzed in a detailed study byLing and Negendank [44]; they came to the conclusion that ATP did not actually cause anet gain of Na+ by these vesicles and that the results can be better explained by the AIH.The criticism of Ling and Negendank has not been refuted in print.These negative results either require one to refute Ling’s energy argument and to carry
out new test experiments, which unequivocally prove the existence of pumps as postulatedby the MPT, or to adopt an alternative working hypothesis for designing new test experi-ments. In the author’s opinion the AIH is the most advanced alternative model from botha theoretical and experimental standpoint which has proven its usefulness as a workinghypothesis by many correct predictions a few of which should be mentioned here (for acomplete list of successful predictions provided by the AIH see [39]):Since 1965 it has been postulated that the bulk of cellular water is polarized in multi-
layers [32]. Experimental testing [42] shows that 95% of frog muscle water follow theBradley multilayer adsorption isotherm [2]. According to the AIH cell volume regulationis primarily not due to membrane properties but is due to interactions between cellularproteins, ions and water. Experimental testing shows that the maintenance of normalmuscle cell volume, its swelling in hypotonic solutions as well as in concentrated KC1solutions are indifferent to the presence of an intact cell membrane [46]. A logical conse-quence of the multilayer theory of cell water and of water in solutions of certain polymersis that the water molecules suffer motional restriction, in particular rotational motional re-striction. This prediction has been confirmed by quasi-elastic neutron scattering [50, 54](see also [48]). Within the context of the AIH it is postulated that K+ accumulation andNa+ exclusion by living cells is due to cytoplasmic properties. Both phenomena have beenverified by direct exposure of muscle cytoplasm to Ringer’s solution [37]. A prediction ofthe AIH is that K accumulation in muscle follows the distribution of β- and γ-carboxylgroups fixed to cellular proteins. This prediction has been confirmed by electron micro-scopic studies (see Discussion of this paper). The AIH predicts that the electrical poten-tial of cells is dependent on ion adsorption at the cell surface and not on ionpermeabilities. This prediction has been confirmed with guinea pig heart muscle cells byusing K+, Rb+ and Cs+ ions [5].
Acknowledgements. This work was supported by a grant from the city of Homburg/Saar. I wouldlike to thank Mrs. E. Frank for typing the manuscript.
References
1. Boyle, P.J., Conway, E.J. (1941) Potassium accumulation in muscle and associated changes.J. Physiol. 100, 1–63.
PHYSICAL STATE OF POTASSIUM IN FROG SKELETAL MUSCLE 89

2. Bradley RS. (1936) Polymolecular adsorbed films. II. The general theory of the condensa-tion of vapors in finely divided solids. J. Chem. Soc. 1936, 1799–1804.
3. Dean, R.B. (1941) Theories of electrolyte equilibrium in muscle. Biol. Symp. 3, 331–348.4. Dick, DAT, McLaughlin SGA. (1969) The activities and concentrations of sodium and
potassium in toad oocytes. J. Physiol. 205, 61–78.5. Edelmann, L. (1973) The influence of rubidium and cesium ions on the resting potential of
guinea-pig heart muscle cells as predicted by the association-induction hypothesis. Ann. N.Y.Acad. Sci. 204, 534–537.
6. Edelmann, L. (1977) Potassium adsorption sites in frog muscle visualized by cesium andthallium under the transmission electron microscope. Physiol. Chem. Phys. 9, 313–317.
7. Edelmann, L. (1980a) Potassium binding sites in muscle. Electron microscopic visualizationof K, Rb, and Cs in freeze-dried preparations and autoradiography at liquid nitrogen temper-ature using 86Rb and 134Cs. Histochemistry 67, 233–242.
8. Edelmann, L. (1980b) Preferential localized uptake of K+ and Cs+ over Na+ in the A bandof freeze-dried embedded muscle section. Detection by X-ray microanalysis and laser micro-probe analysis. Physiol. Chem. Phys. 12, 509–514.
9. Edelmann, L. (1981) Selective accumulation of Li+, Na+, K+, Rb+, and Cs+ at protein sitesof freeze-dried embedded muscle detected by LAMMA. Fresenius Z. Anal. Chem. 308,218–220.
10. Edelmann, L. (1983) Electron probe X-ray microanalysis of K, Rb, Cs, and T1 in cryosec-tions of the striated muscle. Physiol. Chem. Phys., 15, 337–344.
11. Edelmann, L. (1984a) Subcellular distri bution of potassium in striated muscles. ScanningElectron Microsc. 1984; 11:875–888.
12. Edelmann, L. (1984b) Frozen hydrated cryosections of thallium loaded muscle reveal sub-cellular potassium binding sites. Physiol. Chem. Phys. and Med. NMR 16, 499–501.
13. Edelmann, L. (1986) Two opposing theories of the cell: experimental testing by cryometh-ods and electron microscopy. Science of Biological Specimen Preparation. M. Muller, R.P.Becker, A. Boyde, J.J. Wolosewick (eds.). Scanning Electron Microscopy, Inc. AMF O’Hare,Chicago, IL 60666, 33–42.
14. Edelmann, L. (1988) The cell water problem posed by electron microscopic studies of ionbinding in muscle. Scanning Microsc. 2, 851–865.
15. Edelmann, L. (1989a) The contracting muscle: a challenge for freeze-substitution and lowtemperature embedding. In: The Science of Biological Specimen Preparation. SEM Inc., AMFO’Hare (Chicago) IL.
16. Edelmann, L. (1989b) Freeze-substitution and low temperature embedding for analyticalelectron microscopy. In: Electron Probe Microanalysis — Applications in Biology and Medi-cine. K. Zierold, HK. Hagler (eds.) Springer Verlag, Berlin, Heidelberg, New York SpringerSeries in Biophysics Vol. 4. PM Bayley (ed.) 33–46.
17. Edzes, H.T., Berendsen, H.J.C. (1975) The physical state of diffusible ions in cells. AnnualReview of Biophysics and Bioengineering 4, 265–284.
18. Eisenmann, 6, Rudin, D.O., Casby, J.U. (1957) Glass electrodes for measuring sodium ion.Science 126, 831–834.
19. Fenn, W.O. (1936) Electrolytes in muscle. Physiol. Rev. 16, 450–487.20. Gersh, I. (1938) Improved histochemical methods for chloride, phosphate-carbonate and
potassium applied to skeletal muscle. Anat. Rec. 70, 311–329.21. Hazlewood, C.F. (1979) A view of the significance and understanding of the physical prop-
erties of cell-associated water. In: Cell Associated Water. W. Drost-Hansen (ed.) AcademicPress, New York, 165–259.
22. Hill, A.V. (1930) The state of water in muscle and blood and the osmotic behavior ofmuscle. Proc. R. Soc. London Ser. B. 106, 477–505.
23. Hinke, J.A.M. (1961) The measurement of sodium and potassium activities in the squidaxon by means of cation-selective glass microelectrodes. J. Physiol. 156, 314–335.
24. Hodgkin, A.L., Katz, B. (1949) The effect of sodium on the electrical activity of the giantaxon of the squid. J.Physiol. 108, 37–77.
90 LUDWIG EDELMAN

25. Huxley, A.F., Niedergerke, R. (1958) Measurement of the striations of isolated muscle fiberswith the interference microscope. J. Physiol. 144, 403–425.
26. Kernan, R.P., MacDermott, M. (1975) Changes in potassium activity within frog sartoriusmuscle fibres during sodium enrichment in potassium-free Ringer fluid. Physiol., 249, 25–26.
27. Khuri, R.N., Hajjar, J.J., Agulian, S.K. (1972) Measurement of intracellular potassium withliquid ion-exchange microelectrodes. J. Appl. Physiol. 32, 419–422.
28. Kostyuk, P.G., Sorokina, Z.A., Kholodova, Yu D. (1969) Measurement of activity of hydro-gen, potassium and sodium ions in striated muscle fibers and nerve cells. In: Glass Micro-electrodes, (eds.) Laval lee M, Schanne OF, Herbert NC. New York: Wiley 322–348.
29. Lee, C.O., Armstrong, W.M. (1974) State and distribution of potassium and sodium ions infrog skeletal muscle. J. Membrane Biol. 15, 331–362.
30. Lev, A.A. (1964) Determination of activity and activity coefficients of potassium andsodium ions in frog muscle fibres. Nature 201, 1132–1134.
31. Ling, G.N. (1962) A Physical Theory of the Living State. The association-induction hy-pothesis. Blaisdell, Waltham, MA 1-680.
32. Ling, G.N. (1965) The physical state of water in living cells and model systems. Ann. NY.Acad. Sci. 125, 401–417.
33. Ling, G.N. (1969) Measurement of potassium ion activity in the cytoplasm of living cells.Nature 221, 386–387.
34. Ling, G.N. (1977a) Thallium and cesium in the muscle cells compete for the adsorption sitesnormally occupied by K+. Physiol. Chem. Phys. 9, 217–222.
35. Ling, G.N. (1977b) K+ localization in muscle cells by autoradiography, and identification ofK+ adsorbing sites in living muscle with uranium binding sites in electron micrographs offixed preparations. Physiol. Chem. Phys. 9, 319–327.
36. Ling, G.N. (1977c) The functions of polarized water and membrane lipids: a rebuttal. Phys-iol. Chem. Phys. 9, 301–311.
37. Ling, G.N. (1978) Maintenance of low sodium and high sodium levels in resting musclecells. J. Physiol. 280, 105–123.
38. Ling, G.N, (1984) In Search of the Physical Basis of Life. Plenum Publishing Corp. NewYork, 1–791.
39. Ling, G.N. (1992) A Revolution in the Physiology of the Living Cell. R. Krieger Publ. Co.,Melbourne, FL.
40. Ling, G.N., Bohr, G. (1969) Studies on ionic distribution in living cells: I. Long-term preser-vation of isolated frog muscles. Physiol. Chem. Phys. 1, 591–599.
41. Ling, G.N., Bohr, G. (1971) Studies on ion distribution in living cells: IV. Effect of oubainon the equilibrium concentrations of Cs+, Rb+, K+, Na+ and Li+ ions in frog muscle cells.Physiol. Chem. Phys. 3, 573–583.
42. Ling, G.N., Negendank, W.G. (1970) The physical state of water in frog muscle. Physiol.Chem. Phys. 2, 15–33.
43. Ling, G.N., Ochsenfeld, M.M. (1973) Mobility of potassium ion in frog muscle cells, bothliving and dead. Science 181, 78–81.
44. Ling, G.N., Negendank, W. (1980) Do isolated membranes and purified vesicles pumpsodium? A critical review and reinterpretation. Persp. Biol. Med. 23, 215–239.
45. Ling, G.N., Tucker, M. (1983) Only solid red blood cell ghosts transport K+, and Na+ againstconcentration gradients: hollow intact ghosts with K+-Na+ activated ATPase do not. Physiol.Chem. Phys. and Med. NMR 15, 311–317.
46. Ling, G.N., Walton, C.L. (1976) What retains water in living cells? Science 191, 293–295.47. Ling, G.N., Miller, C., Ochsenfeld, M.M. (1973) The physical state of solutes and water in
living cells according to the association-induction hypothesis. Ann. N.Y. Acad. Sci. 204, 6–50.48. Negendank, W. (1986) The state of water in the cell. Science of Biological Specimen Prepa-
ration. M. Muller, R.P. Becker, A. Boyde, J.J. Wolosewick (eds.). Scanning Electron Mi-croscopy Inc., AMF O’Hare Chicago, IL, 60666, 21–32.
49. Plattner, H. (1988) Buchbesprechungen. Optik 80, 136.
PHYSICAL STATE OF POTASSIUM IN FROG SKELETAL MUSCLE 91

50. Rorschach, H.E., Bearden, D.W., Hazlewood, C.F., Heidorn, D.B., Nicklow, R.M. (1987)Quasi-elastic scattering studies of water diffusion. Scanning Microsc.1, 2043–2049.
51. Schwartz, I.L., Walter, R., Fein, M., Wyssbrod, H.R. (1978) or (1985) In: Best and Taylor’sPhysiological Basis of Medical Practice. JR. Brobeck, (ed.) Williams and Wilkins, Baltimore,MP. 1–23 (10th edition: 1978, 11th edition: 1985).
52. Skou, J.C. (1957) The influence of some actions on an adenosine triphosphatase from pe-ripheral nerves. Biochim. Biophys. Acta 23, 394– 401.
53. Steinbach, H.B. (1940) Sodium and potassium in frog muscles. J. Biol. Chem. 133,695–705.
54. Trantham, E.C., Rorschach, H.E., Clegg, J.S., Hazlewood, C.F., Nicklow, R.M., Wakabayashi,N. (1984) Diffusive properties of water in artemia cysts as determined from quasi-elasticneutron scattering spectra. Biophys. J. 45, 927–938.
55. von Zglinicki, T. (1988) Monovalent ions are spatially bound within the sarcomere. Gen.Physiol. Biophys. 7, 495–504.
56. Walker, J.L., Brown, H.M. (1977) Intracellular ionic activity measurements in nerve andmuscle. Physiol. Rev. 57, 729–778.
Discussion with Reviewers
C.F. Hazlewood: In studies of many non-muscle cells, the activity of potassium (mea-sured by the potassium sensitive electrode) has been shown to be significantly reduced.Can you explain how the ion sensitive electrodes can give these opposite results?
Author: This problem has been discussed at length by Ling (text reference [38], pp. 252-257). He came to the following conclusion (p. 253): “There are three possible sources ofartifacts in the intracellular microelectrode recording of intracellular K+-activity coeffi-cient: (1) liberation of adsorbed K+, (2) localized depolarization of water with rise in itssolubility for K+, and (3) interference by charged amino groups on proteins. Since allthree of these sources of artifacts favor the recording of a spuriously high in activity coef-ficient, it is remarkable that one finds so many reports of a low K+-activity coefficient (inepithelial cells, see Table on p. 255). A likely cause for this is a greater stability of the cy-toplasm of these epithelial cells when compared to that of, for example, nerve andmuscle.”
C.F.Hazlewood: It has been reported that cell nuclei swell (in situ and in vitro) when ex-posed to monovalent ionic concentrations of the order of 150 mM/l (see Kellermayer M.(1981). Soluble and “loosely bound” nuclear proteins in regula tion of the ionic environ-ment in living cell nuclei. In: Intl. Cell Biology. H.G. Schweiger (ed.) Springer Verlag,Heidelberg. 915-924; Hazlewood C.F., Kellermayer M. (1988). Ion and water retention bypermeabilized cells. Scanning Microsc. 2, 267-273). These time dependent changes in nu-clei came to mind when I saw your slide of the changes in the sarcoplasm at the tip of themicroelectode. Do you think these observations are related in any way to what is goingon at the tip of your microelectrode?
Author: It is possible that the local swelling of the muscle observed at the place of theelectrode impalement is not only due to movement of free water towards the place whereliberated K+ ions accumulate but also to an additional mechanism; cytoplasm exposed tohigh concentrations of alkali-metal ions may swell because specific salt linkages between
92 LUDWIG EDELMAN

neighbored proteins may be dissociated leading to an expanded but still coherent cyto-plasm with water polarized in multilayers [38, p. 445; 46]. A similar mechanism may beresponsible for the swelling of nuclei exposed to high concentrations of monovalent ions.
Th. von Zglinicki:Micrographs like your Figure 6 contain an enormous amount of quan-titative information which could easily be obtained by microdensitometry. I would like toencourage you to do so and to compare the results with the expected distribution of puta-tive binding sites.
Author: Such studies are planned. In order to obtain reproducible quantitative informationit is, however, necessary to produce first reproducible cryosections with the lowest possiblecompression during sectioning. We are currently trying to achieve this precondition.
Like the preceding article by Dr. Gilbert N. Ling, this article was also once published inScanning Microscopy, which is now defunct. Because the paper contains important infor-mation that must be preserved, like Dr. Ling I also am grateful for the opportunity toreprint the article in Physiol. Chem. Phys, and Med. NMR.
Dr. Ludwig Edelmann
PHYSICAL STATE OF POTASSIUM IN FROG SKELETAL MUSCLE 93


















