
THE OF BIOLOGICAL No. 25, and Printed U. S. A ... JOURNAL OF BIOLOGICAL CHEMISTRY 0 1988 by The...
7
THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1988 by The American Society for Biochemistry and Molecular Biology, Inc. Vol. 263, No. 21, Issue of July 25, pp. 10408-10414, 1988 Printed in U. S. A. Sequencing of the Gene ant Which Affects the Na+/H+ Antiporter Activity in Escherichia coli * (Received for publication, January 12, 1988) Rachel KarpelS, Yael OlamiS, Daniel TaglichtQ, Shimon Schuldinerg, and Etana PadanS From the $Departmentof Molecular and Microbial Ecology, Life Science Institute and the §Departmentof Molecular Biology, Hadassah Medical School, Hebrew University, Jerusalem 91010, Israel By introduction of deletions and/or insertions, we havedefinedasmall DNA fragment (1.58 kilobase pairs) which contains the ant gene. Thus, transform- ants of all plasmid constructs containing this segment exhibit Antup phenotype, i.e. growth is Li+-resistant and Na+/H+ antiporter activity is increased in isolated everted membrane vesicles. Utilizing the T7 promoter expression system, we also found that this fragment encodes for a single protein of35 kDa. The DNA frag- ment has been sequenced and found to contain an open readingframe of 1085 basepairs.Analysis of the sequence of the predicted protein suggests the presence of 10 putative transmembrane segments in the protein. The sodium proton antiporter plays a central role in the Na’ metabolism of the prokaryotic cell (1-4). In Escherichia coli, as in many other bacteria, this is the only known system for Na+ extrusion and maintenance of a Na+ gradient (Na+in < Na+out) over a wide range of external concentrations (1-5). The antiporter exchanges protons for Na+ in a stoichiometry larger than 1. Thus, it is driven by the electrochemical proton gradient generated by the primary proton pumps. The Na+ gradient directed inward drives Na+/substrate symport sys- tems. In E. coli, four such symporters are known: melibiose, glutamate, proline, and serine (Refs. 6-9, respectively). Na+ symporters are common in halophiles and alkalophiles, and inthelatter also the flagellar motor is driven by aNa+ gradient (10). Since the Na+/H+ antiporter converts A;H+ to A,iiNa+ in a reversible fashion, it may well serve as adevice for energy buffering. Under some conditions, when A;H+ is ex- hausted, the preformed A;N~+ is utilized to replenish it (11- 13). It has also been suggested that the Na+/H+ antiporter is intimately involved in regulation of intracellular pH in bac- terial cells in alkaline environments (1, 14-16). Utilizing intact cells as well as isolated membrane vesicles, the kinetics and bioenergetics of the antiporter have been studied in several bacteria (1, 2, 4, 12). These results show that it is electrogenic and regulated by intracellular pH. A genetic approach has been undertaken in the study of this transport system. In Bacillus alkalophilus, Krulwich and colleagues (1) isolated a mutant which lost both the Na+/H+ *This workwas supported by a grant from the United States- Israel Binational Foundation (to E. P. and S. S.) and a grant from the Niedersachsiches Ministerium fir Wissenschaft und Kunst (to S. S.). The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. The nucleotide sequence(s) reported in this paper has been submitted to the GenBankTM/EMBL Data Bank with accession number(s) 503879. antiporter activity and the capacity to regulate intracellular pH at alkaline pH, implying the role of the Na+/H+ antiporter in pH homeostasis of these bacteria. E. coli mutants that cannot regulate pHi and therefore do not grow at alkaline pHo have been isolated (9, 17). One of these mutations has been mapped in the gene encoding for the subunit of the RNA polymerase and therefore its relationship to Na’/H+ antiport activity and to regulation of pHi is not yet clear (16). Another mutant is impaired in the Na’/H+ antiporter activity, pH homeostasis, and growth at alkaline pH (9). Tsuchiya, Wilson, and colleagues (18) isolated a mutant in E. coli that shows increased rather than decreased antiporter activity. These cells also tolerate much higher Li+ concentra- tions (100 mM) as compared to the wild type cells (10 mM). The Li+ resistance has been related to the excretion of the ion by the enhanced antiporter. This mutation designated antup has recently been mapped in the vicinity of carA at about 0.5 min on the E. coli map (19). The mapping of antup allowed us to survey a series of plasmids, pBR322 derivatives, covering 15 kilobase pairs of wild type DNA spanning the antup region and to identify recombinant plasmids that confer the Antup phenotype. It has been concluded that these plasmids bear the ant wild type gene(s) which, when present in high copy number, produces the Antup phenotype (19). All plasmids producing increased Na+/H+antiporter activ- ity were shown to contain in common at least two fragments in tandem amounting together to 2.77 kilobase pairs (a HindIII-BamHI fragment of 0.92 kb’ and a BamHI-BamHI fragment of 1.85 kb) (19). Since neither of these inserts alone yielded Antup, it has been concluded that either the antgene spans the junction between them or that products encoded by both inserts are required for Antup. The HindIII-BamHI fragment has already been sequenced and shown to contain an open reading frame (or0 of 0.8 kb which encodes for a 28-kDa protein in an in uitro system for coupled transcriptionandtranslation (20, 21). The direct involvement of this protein in the Antup phenotype has been tested and discarded since plasmid constructs in which sub- stantial parts of the putative orf were deleted confer Antup phenotype (19). In order to identify the antgene(s) and product(s), we have subcloned the gene from the 2.77-kb fragment and defined a fragment of 1.58 kb which confers Antup and expresses a membrane-bound protein of apparent molecular weightof 35,000. The DNA fragment was sequenced and shown to contain a putative orf capable of encoding a protein 362 amino acids long. The hydropathic profile obtained from the orf shows the existence of 10 putative transmembrane segments. The abbreviations used are: kb, kilobase pairs; SDS, sodium dodecyl sulfate; Hepes, 4-(2-hydroxyethyl)-l-piperazineethanesul- fonic acid. 10408
Transcript of THE OF BIOLOGICAL No. 25, and Printed U. S. A ... JOURNAL OF BIOLOGICAL CHEMISTRY 0 1988 by The...

THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1988 by The American Society for Biochemistry and Molecular Biology, Inc.
Vol. 263, No. 21, Issue of July 25, pp. 10408-10414, 1988 Printed in U. S. A.
Sequencing of the Gene ant Which Affects the Na+/H+ Antiporter Activity in Escherichia coli *
(Received for publication, January 12, 1988)
Rachel KarpelS, Yael OlamiS, Daniel TaglichtQ, Shimon Schuldinerg, and Etana PadanS From the $Department of Molecular and Microbial Ecology, Life Science Institute and the §Department of Molecular Biology, Hadassah Medical School, Hebrew University, Jerusalem 91010, Israel
By introduction of deletions and/or insertions, we have defined a small DNA fragment (1.58 kilobase pairs) which contains the ant gene. Thus, transform- ants of all plasmid constructs containing this segment exhibit Antup phenotype, i.e. growth is Li+-resistant and Na+/H+ antiporter activity is increased in isolated everted membrane vesicles. Utilizing the T7 promoter expression system, we also found that this fragment encodes for a single protein of 35 kDa. The DNA frag- ment has been sequenced and found to contain an open reading frame of 1085 base pairs. Analysis of the sequence of the predicted protein suggests the presence of 10 putative transmembrane segments in the protein.
The sodium proton antiporter plays a central role in the Na’ metabolism of the prokaryotic cell (1-4). In Escherichia coli, as in many other bacteria, this is the only known system for Na+ extrusion and maintenance of a Na+ gradient (Na+in < Na+out) over a wide range of external concentrations (1-5). The antiporter exchanges protons for Na+ in a stoichiometry larger than 1. Thus, it is driven by the electrochemical proton gradient generated by the primary proton pumps. The Na+ gradient directed inward drives Na+/substrate symport sys- tems. In E. coli, four such symporters are known: melibiose, glutamate, proline, and serine (Refs. 6-9, respectively). Na+ symporters are common in halophiles and alkalophiles, and in the latter also the flagellar motor is driven by a Na+ gradient (10). Since the Na+/H+ antiporter converts A;H+ to A,iiNa+ in a reversible fashion, it may well serve as a device for energy buffering. Under some conditions, when A;H+ is ex- hausted, the preformed A;N~+ is utilized to replenish it (11- 13). It has also been suggested that the Na+/H+ antiporter is intimately involved in regulation of intracellular pH in bac- terial cells in alkaline environments (1, 14-16).
Utilizing intact cells as well as isolated membrane vesicles, the kinetics and bioenergetics of the antiporter have been studied in several bacteria (1, 2, 4, 12). These results show that it is electrogenic and regulated by intracellular pH.
A genetic approach has been undertaken in the study of this transport system. In Bacillus alkalophilus, Krulwich and colleagues (1) isolated a mutant which lost both the Na+/H+
*This work was supported by a grant from the United States- Israel Binational Foundation (to E. P. and S. S.) and a grant from the Niedersachsiches Ministerium fir Wissenschaft und Kunst (to S. S.). The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
The nucleotide sequence(s) reported in this paper has been submitted to the GenBankTM/EMBL Data Bank with accession number(s) 503879.
antiporter activity and the capacity to regulate intracellular pH at alkaline pH, implying the role of the Na+/H+ antiporter in pH homeostasis of these bacteria. E. coli mutants that cannot regulate pHi and therefore do not grow at alkaline pHo have been isolated (9, 17). One of these mutations has been mapped in the gene encoding for the subunit of the RNA polymerase and therefore its relationship to Na’/H+ antiport activity and to regulation of pHi is not yet clear (16). Another mutant is impaired in the Na’/H+ antiporter activity, pH homeostasis, and growth at alkaline pH (9).
Tsuchiya, Wilson, and colleagues (18) isolated a mutant in E. coli that shows increased rather than decreased antiporter activity. These cells also tolerate much higher Li+ concentra- tions (100 mM) as compared to the wild type cells (10 mM). The Li+ resistance has been related to the excretion of the ion by the enhanced antiporter. This mutation designated antup has recently been mapped in the vicinity of carA at about 0.5 min on the E. coli map (19).
The mapping of antup allowed us to survey a series of plasmids, pBR322 derivatives, covering 15 kilobase pairs of wild type DNA spanning the antup region and to identify recombinant plasmids that confer the Antup phenotype. It has been concluded that these plasmids bear the ant wild type gene(s) which, when present in high copy number, produces the Antup phenotype (19).
All plasmids producing increased Na+/H+ antiporter activ- ity were shown to contain in common at least two fragments in tandem amounting together to 2.77 kilobase pairs (a HindIII-BamHI fragment of 0.92 kb’ and a BamHI-BamHI fragment of 1.85 kb) (19). Since neither of these inserts alone yielded Antup, it has been concluded that either the ant gene spans the junction between them or that products encoded by both inserts are required for Antup.
The HindIII-BamHI fragment has already been sequenced and shown to contain an open reading frame (or0 of 0.8 kb which encodes for a 28-kDa protein in an in uitro system for coupled transcription and translation (20, 21). The direct involvement of this protein in the Antup phenotype has been tested and discarded since plasmid constructs in which sub- stantial parts of the putative orf were deleted confer Antup phenotype (19).
In order to identify the ant gene(s) and product(s), we have subcloned the gene from the 2.77-kb fragment and defined a fragment of 1.58 kb which confers Antup and expresses a membrane-bound protein of apparent molecular weight of 35,000. The DNA fragment was sequenced and shown to contain a putative orf capable of encoding a protein 362 amino acids long. The hydropathic profile obtained from the orf shows the existence of 10 putative transmembrane segments.
The abbreviations used are: kb, kilobase pairs; SDS, sodium dodecyl sulfate; Hepes, 4-(2-hydroxyethyl)-l-piperazineethanesul- fonic acid.
10408

Sequence of Na+/H+ Antiporter Gene 10409
FIG. 1. Subcloning ant. Deletions and/or insertions in the insert of pGM36 were obtained as described under “EX- perimental Procedures.” 1-3, utilizing restriction sites; 4-6, by digestion with Bal31; 5, 7, 8, inserts of pKR322, pKR323, and pKR225, respectively. Each of the constructs was introduced into TA15, and the transformants were tested for growth on 100 mM Li+. In some cases, the Na+/H+ antiporter activity was also determined in isolated everted membrane vesicles. UTT, universal translation terminator sequences; Kan, kanamycin cassette (27). The straight lines describe the inserts in scale. Broken lines depict vector parts, while triangles denote kanamycin cassette or UTT se- quences (not drawn to scale). Restriction sites originated from pGM36 insert are depicted above while those from the vec- tor, cassette, or UTT are indicated below the line describing the DNA fragments.
Deletion Insertion naert of
pGM36
pKR 322
p K R 3 2 3
p K R 2 2 5
EXPERIMENTAL PROCEDURES
Bacterial Strains and Culture Conditions-E. coli K12 strain TA15 is melBLidant+ (19). It is used in this study as a host organism for plasmids, tested for capacity to confer increased Na+/H+ antiporter activity. Cells were grown in L broth or minimal medium A (22) without sodium citrate, supplemented with thiamin (1 pglml), 0.5% glycerol, or melibiose as required. Where indicated, Li+ or Na+ was also added to yield 100 mM. For plates, 1.5% Difco agar was added. For selection of plasmids, 100 pg/ml ampicillin or 50 pg/ml kana- mycin were used.
E. coli MV1190 (kindly provided by Y. Gruenbaum, Life Science Institute, Hebrew University, Jerusalem) is a A(lac-proAB) thisupE44 A(sr1-recA)306::Tn10(tetR)/F’:traD36 proABlacIQlucZ AM15. It was used infected with bacteriophage M13K07 (23) for growth of pUC118 or pUC119 to obtain single-stranded DNA.
Plasmids”pGM36 and pGM42 are pBR322 derivatives (19) and pT7-5 and pGP1-2 were used to express ant. pT7-5 is similar to pT7- 3 except that the p-lactamase gene is reversed and the expression by T7-RNA polymerase is limited exclusively to cloned genes.’ pUC118 and pUC119 are pUC18 and pUC19 (25) derivatives constructed with the intergenic region of M13 by J. Vieira (Dept. of Biochemistry, University of Minnesota, St. Paul, MN) and kindly provided to us by Y. Gruenbaum. They were used for DNA sequencing.
Deletions or Znsertions in pGM36”Since the sequence of the HindIII-BamHI (0.92 kb) segment of the insert of pGM36 was known (20), deletions and/or insertions were introduced into this DNA fragment by utilization of specific restriction sites. The resulting inserts are described in Fig. 1.
1) In pGM36, there are three BanII sites, one at the BamHI- HindIII (0.92 kb) part of the insert and two, very close to the BamHI end of the insert on the pBR322 vector. pGM36 was cut with BanII, and the resulting 2.55-kb fragment (BanII-BanII) was religated to BanII cut pBR322. Restriction enzyme analysis with BamHI and HindIII was used to select plasmids in which the orientation of the insert was identical or reversed with respect to that of pGM36.
2) In the BamHI-Hind111 segment of pGM36 insert, there are two adjacent BstXI sites, unique in pGM36. pGM36 was cut with BstXI. The shortened plasmid was purified, and, after filling its ends with T4 DNA polymerase, religated via a DNA fragment which contained
S. Tabor, unpublished data.
A
irowth on X)mM Li+
N8.l H’ mntiport nctivity
+ + +
- +
+
-
+
+
‘ l k b -
sequences of a universal translation terminator (UTT, the sequence was TTAAAGTTAACTTAA and the oligonucleotide was kindly sup- plied by Dr. H. Sarkar (Roche Institute of Molecular Biology, Nutley, NJ). The terminator was introduced to avoid formation of fused proteins.
3) In pGM36, there are three ClaI sites, two that we found in the insert and one in pBR322 part. The plasmid was cut with ClaI, and the ClaI-ClaI fragment (1.4 kb) was religated to ClaI cut pBR322. Plasmids with the orientation of pGM36 were selected using EcoRI and BamHI.
4-6) Since the sequence of the BamHI-BamHI (1.85 kb) part of pGM36 insert was not known, it was shortened from its vector end by applying exonuclease Bal31 (26). pGM36 was cut with SphI which is a unique restriction site of pGM36 located on the vector near the BamHI end of the insert. It is also remote from the plasmid essential genes (beyond the PvuII site) by about 1.5 kb. Therefore, treatment of SphI cut pGM36 with the exonuclease for different time periods allowed marked shortening of the insert before hampering plasmid activity. The shortened plasmids were purified and religated after end filling with a kanamycin casette (27). The latter introduced both a selective marker and restriction sites for further excision of the shortened insert. The plasmid that contains deletion 5 was named pKR322.
7) The short plasmid pKR322 was digested by BamHI, and the 1.15-kb fragment was exchanged with the BamHI-BamHI fragment (1.85 kb) of pGM36. Plasmid with orientation identical with pGM36 was selected using SmaI and HindIII. This plasmid was cut with BstXI and religated via UTT as described for deletion 2 to yield plasmid pKR323.
8 ) While constructing pKR322, the pBR322 part was concomi- tantly digested from the BamHI site up to about the BspMI site so that the BanII sites were deleted. Digestion of pKR322 by SmaI and Hind111 yielded shortened vector (3 kb) which was ligated with the SmaI-Hind111 insert (1.9 kb) of pKR323 containing one BanII site. The construct was then digested with BanII and HindIII, and the 4.3-kb fragment was religated after end filling. The result was plasmid pKR225 with an insert (SmaI-EcoRI) of 1.61 kb.
Expression of ant Gene in Vitro-In order to identify the ant gene product, the T7 promoter expression system was used (24). Different size fragments of pGM36 or pGM42 inserts were subcloned into the polylinker, downstream of the T7 RNA polymerase promoter, of

10410 Sequence of Na+/H+ Antiporter Gene pT7-5 to yield the following recombinant plasmids (Fig. 2).
1) pDT2, pGM42 was cut with BamHI and ClaI, and the 2.3-kb fragment was subcloned between ClaI and BamHI of pT7-5.
2) pDT6, pDT2 was cut by BamHI, and the BamHI-BamHI (1.85 kb) fragment of pGM36 insert was subcloned into it. The plasmid with the orientation of pGM42 was selected using BglII and HindIII.
3) pDT14, pDT6 was cut with BanII, and the 2.41-kb fragment was subcloned into BanII cut pT7-5. ClaI was used to select the plasmid with the depicted orientation.
4) pDT62, pDT6 was cut with BstXI. After end-filling with T4 DNA polymerase, universal terminator sequences (UTT) were ligated into it instead of the BstXI-BstXI fragment (0.13 kb). Insertion was confirmed by digestion with HincII, a restriction site in UTT.
The plasmids were transformed into TA15 carrying pGP1-2 (24). Transformants were grown at 30 "C in minimal medium supple- mented with thiamin and glycerol to a cell density of 0.1 mg of cell proteinlml (0.3-0.4 ODw). The cell suspension was incubated for 15 min at 42 "C to induce the T7-RNA polymerase, rifampicin (200 pg/ ml) was added, and incubation continued for an additional 10 min. Then the culture was shifted back to 30 "C for 30 min. [3SS]Methio- nine (10 pCi) was added to 1-ml aliquots of the cell suspension, and incubation continued for an additional 5 min. The cells were centri- fuged and resuspended in the sampling buffer. Cell proteins were resolved by SDS-polyacrylamide gel electrophoresis (15% polyacryl- amide) and visualized by autoradiography.
DNA Sequencing of the ant Gene-DNA sequencing of both strands of the DNA insert carrying ant of pKR323 was achieved by the dideoxyoligonucleotide technique (28, 29). We used as templates various DNA fragments of pKR323 insert, prepared as described under "Results" and Fig. 3 and subcloned into pUC118 and pUC119. As a primer, we utilized the M13, 17 residue universal primer, pur- chased from Amersham. After obtaining part of the sequence, we synthesized homologous 20-residue oligomers which served as primers for further sequencing (Fig. 3). All of the sequence was determined from both strands.
Oligonucleotide Synthesis-The oligonucleotide described in Fig. 3 was synthesized on an Applied Biosystem model 380B DNA synthe- sizer and purified by polyacrylamide gel electrophoresis as described in the Applied Biosystems Manual.
Everted membrane vesicles were prepared as previously described (19, 30).
Measurement of pH and Estimation of Na+/H+ Antiporter Actiu- ity-Acridine orange fluorescence was monitored to estimate pH maintained across membranes of everted vesicles upon addition of lactate as previously described (19, 31). The reaction mixture con- tained, in 2.5 ml, the following: 60-70 pg of membrane protein, 10 mM Tris-Hepes (pH 8), 140 mM choline chloride, 5 mM MgClz, 1 WM
pGM42
pDT2
pDT6
p DT14
pDT62
ClaI
9 BanII 2 a I
BanII HindIlI
F;L h ClaI
HincII
' 1 k b
FIG. 2. Construction of ant in the T7 promoter expression system. Different size fragments of pGM36 or pGM42 were sub- cloned into the polylinker, downstream of the T7-RNA polymerase promoter (->) of pT7-5 as described under "Experimental Proce- dures." The various inserts are depicted in the figure. Restriction sites originated from pGM42 insert are depicted above and those from the plasmid or cassettes are indicated below the line describing the DNA insert (in scale). UTT, universal translation terminator; broken lines, vector fragment, polylinker; all not drawn to scale.
"""""_ +"P' 4"" p3
+ q----+p,---+
+ + + t t
Insert of pGM 36 "
I I , 0 5001 lOOOl 1270 I
Barn HI Bi I I I d m H I Hindm - ant
1 1 I
2- 3
1100
561 - 4 I I
5 6
561 I
4 2 51
- 1 kb
FIG. 3. Constructions for sequencing of ant. Plasmids used for subcloning were pGM36 and pKR323. Solid lines indicate the extent of fragment from plasmids which have been subcloned for sequencing. Arrows indicate the direction and extent to which the sequence in Fig. 6 was determined from individual fragments serving as templates. Broken arrows show the sequences obtained using synthetic oligonucleotides (Pl-P4) homologous to the following base pairs of the DNA sequence. PI, 848-867; P2, 307-326; P3, 856-875; and P4, 233-249. Each arrow represents results from one to three experiments each involving two to three gels. Broken lines indicate already known sequences (20). The open reading frame of ant (see Fig. 6) is depicted (starts a t 182 and ends at 1267 of the sequence).
acridine orange, and 2 mM K+ lactate. Fluorescence of acridine orange was monitored in an Eppendorf fluorimeter (model 1101M) and recorded by a Goerz recorder (model Re541, Vienna). Exciting light was provided by a mercury lamp and filtered through a 405 nm to 436 nm filter. Emission was measured through a 470 nm to 3000 nm filter.
After the steady state value of ApH was reached, 10 mM NaCl was added and the extent of the Na+-dependent fluorescence enhance- ment was recorded and used to estimate the antiporter activity.
Protein concentration in cells and everted membrane vesicles was determined as described by Lowry et al. (40).
RESULTS AND DISCUSSION
Subcloning of ant-The smallest cloned DNA fragment which produced the Antup phenotype was contained in pGM36 and amounted to 2.77 kb (19). In order to locate the ant gene(s) in this fragment, we either inserted universal translation terminator (UTT) sequences or introduced large deletions at different sites of the insert. The effect of the deletions and/or insertions on the Antup phenotype was then tested by transforming the constructs into TA15 and testing the transformants for growth on high (100 mM) Li'. In addition, everted membrane vesicles were prepared from Li+- resistant isolates and tested for Na+/H+ antiporter activity. Results of such experiments are summarized in Figs. 1 and 4.
Deleting 0.33 kb of DNA from the HindIII end of the insert of pGM36 up to the BanII site (Fig. 1, deletion 1 ) did not affect the Antup phenotype produced by the plasmid. Similar to cells transformed with pGM36, cells transformed with the deleted plasmid showed Li+ resistance and their isolated mem- brane vesicles exhibited markedly enhanced Na+/H+ anti- porter activity. Similarly, deleting the BstXI-BstXI fragment

Sequence of Na+/H+ Antiporter Gene 10411
a mem
:t
nig,KCI
IC
b
d lmln
J NaCl t
TAl5 TAlSlpKR323
FIG. 4. Na+/H+ antiporter activity in everted membranes isolated from TA15/pKR323. Everted membrane vesicles from strains TA15 and TA15/pKR323 were prepared and assayed as de- scribed under “Experimental Procedures.” The reaction mixture con- tained (in 2.5 ml) 10 mM Tris-Hepes (pH 8), 140 mM choline chloride, and 5 mM MgC12. At the onset of the experiment, acridine orange (AO) was added to a 1 p~ concentration, thereafter and, as indicated, membrane vesicles (mem, 60-70 pg of membrane protein) and potas- sium D-laCtate (lact, 2 mM) were added. The detailed protocol is shown in trace A. Where indicated, NaCl (10 mM), nigericin (1 pM), or KC1 (10 mM) was added.
(0.12 kb) of pGM36 insert (Fig. 1, deletion 2) and exchanging it with UTT had no effect on Antup. These results suggest that one side of the ant gene ends before the BstXI site.
Fig. 1 also describes the deletions obtained from the BamHI end of pGM36 insert. It is shown that it is possible to delete up to 0.75 kb of this end without impairing the Antup phe- notype (Fig. 1, deletions 4 and 5). More extensive deletion from this end (Fig. 1 , 3 and 6) hampered the capacity of the insert to produce the Antup phenotype.
Plasmid pKR323 was constructed based on deletions 2 and 5 (Fig. 1). It contains a 1.52-kb fragment up to and a 0.37-kb fragment following the UTT. This plasmid must carry an intact ant gene since pKR323 conferred Li’ resistance and enhanced Na’/H’ antiporter activity of isolated everted mem- brane vesicles as the original plasmid pGM36 (Ref. 19 and Fig. 4). The insert of pKR225 is similar to that of pKR323, but for the Hind111 end which was deleted almost to the UTT insertion. Nevertheless, pKR225 was as active as pKR323 in conferring Li’ resistance (Fig. 1). We therefore conclude that ant is located in the SmaI-UTT (1.52 kb) segment of the SmaI-EcoRI insert (1.61 kb) of pKR225. Formation of fused proteins by the plasmid can be ruled out because of the UTT and because the insert was subsequently transferred to various vectors and found as active in conferring Antup.
Expression of ant Gene-In order to identify the ant gene product, we used the T7 promoter expression system (24). This system allows, in the presence of rifampicin, to express in intact cells, proteins encoded by plasmid genes (inserted downstream of the T7 promoter) rather than chromosomal
genes. Different size fragments of either pGM36 or pGM42 were subcloned into PT7-5 (Fig. 2) and transformed into TA15/pGP1-2. The transformants were pulse-labeled with [”S]methionine in the presence of rifampicin, and the pro- teins resolved by SDS-polyacrylamide gel electrophoresis were visualized by autoradiography (Fig. 5).
In accordance with previous results (19) when the BamHI- BarnHI and the BamHI-Hind111 fragments of pGM36 insert were subcloned, in tandem, downstream of the T7 promoter (Fig. 2, pDT6), two proteins were expressed with apparent M , of 35,000 and 28,000, respectively (Fig. 5, lane 2). This con- struct also produced the Antup phenotype in TA15 cells. It is interesting to note that even though both proteins are trans- lated from the same mRNA molecule, the expression of the 35-kDa one is much weaker than that of the 28-kDA.
As predicted by the subcloning data described above, the inserts containing a t least the fragment BamHI-UTT of pKR323 insert also produced the Antup phenotype when constructed downstream of the T7 promoter. These constructs pDT14 and pDT62 (compare Fig. 2 to Fig. 1) both expressed the 35-kDa protein (Fig. 5, lanes 3 and 4, respectively). How- ever, pDT14 expressed in addition a truncated part of the 28- kDa protein (14 kDa) (Fig. 5, lane 3), whereas pDT62 did not express the latter protein at all (Fig. 5, lane 4 ) . Another construct pDT2 (Fig. 2) which does not contain the minimal DNA fragment needed for Antup activity, but bears the entire orf of the 28-kDa protein, expressed this protein (not shown) but did not confer the Antup phenotype. Taken together, these results imply that the 35-kDa protein is the one involved in conferring the Antup phenotype.
DNA Sequencing of the ant Gene-A part of the DNA sequence of pKR323 insert was already known. This is its BarnHI-Hind111 (0.92-kb) segment (Ref. 20 and Fig. 6). In order to sequence the remaining BamHI-BamHI fragment, we used pGM36 and pKR323 as sources of DNA to get the fragments as described and numbered in Fig. 3.
BglII-BamHI (0.54 kb), and 4) BglII-Hind111 (1.44 kb) frag- ments were restricted from pGM36, 5) BamHI-BamHI (0.89 kb) fragment from deletion 6 (see Fig. 1) and BamHI-BamHI fragment (1.1 kb) from pKR323. The fragments were sub- cloned into pUC118 or pUC119. Fig. 3 diagrams the subclones
1) BamHI-BamHI (1.85 kb), 2) BamHI-BglII (1.3 kb), 3)
1 2 3 4 5 k Do
45 -ont 30 ORF-
bPI 7 - -- 20
a FIG. 5. Expression of ant. pT7-5 derivatives constructed as de-
scribed under “Experimental Procedures” and Fig. 2 were trans- formed into TA15/pGP1-2. Thirty min after heat induction, trans- formants were pulse-labeled with [“S]methionine in the absence (lane I ) and in the presence of (lanes 2-5) of rifampicin as described under “Experimental Procedures.” The proteins resolved on SDS-polyacryl- amide gel electrophoresis were identified by autoradiography. ORF, previously described open reading frame (19, 20). Plasmids tested (see Fig. 2) were pT7-5 ( l a n e 1, before addition of rifampicin; lane 5, after addition of rifampicin), pDT6 (lane 2 ) , PDT 14 (lune 3 ) , pDT62 (lane 4 shown in duplicate).
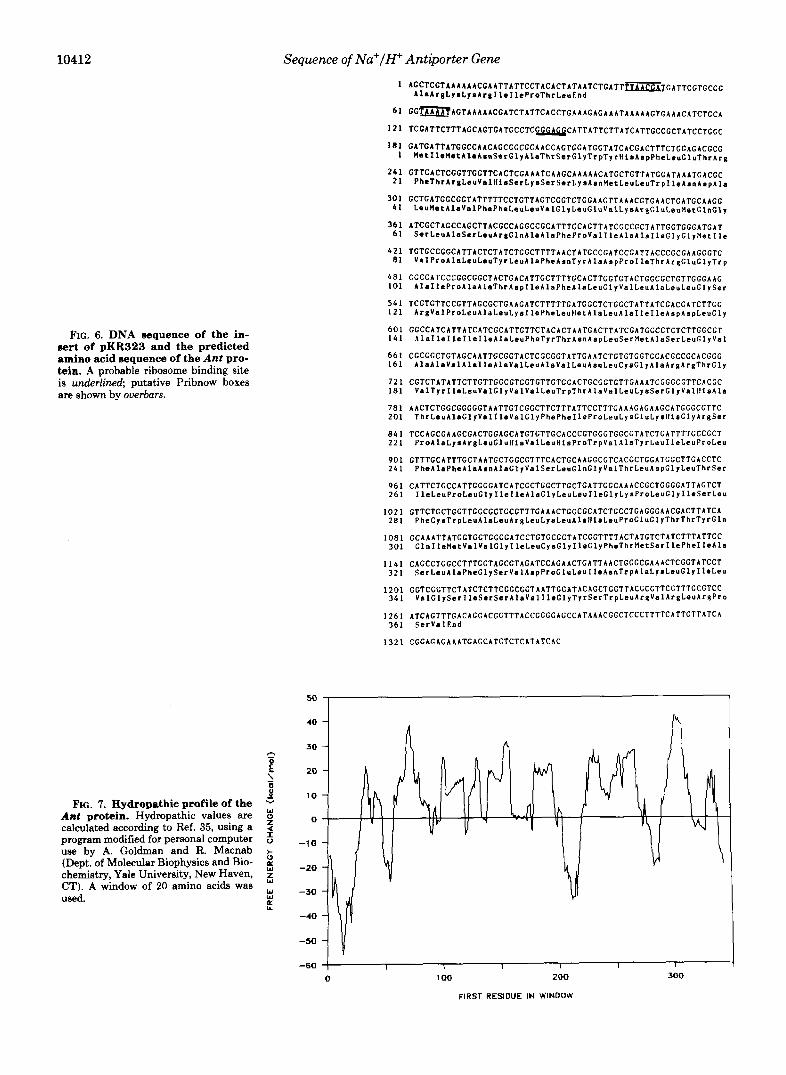
10412 Sequence of Na+/H+ Antiporter Gene
FIG, 6. DNA sequence of the in- sert of pKR323 and the predicted amino acid sequence of the Ant pro- tein. A probable ribosome binding site is underlined; putative Pribnow boxes are shown by ouerbars.
so
h
E 1 ro
FIG. 7. Hydropathic profile of the Ant protein. Hydropathic values are $ calculated according to Ref. 35, using a program modified for personal computer 5 use by A. Goldman and R. Macnab + (Dept. of Molecular Biophysics and Bio- chemistry, Yale University, New Haven, 5 CT). A window of 20 amino acids was U S d . W E
-60 I I 1 I I I
0 1 00 ZOO 300
FIRST RESIDUE IN WINDOW

Sequence of Na+/H+ Antiporter Gene 10413
In E
I n FIG. 8. Model of the secondary
structure of the Ant protein. The model shown is based on the predictions D of the hydropathic profile. Putative I L transbilayer segments are shown in boxes n connected by hydrophilic segments.
L
'n I
a
iu a
. . 8
FIG. 9. Hypothetical secondary structure of ant mRNA. The most stable RNA structure in the segment 75-275 was chosen using the FOLD program of the University of Wisconsin Sequence Analysis Software package (UWGCG) (39). The free energy of this structure is -45.2 kcal/mol. When the segment used was shortened (125- 235), the stem loops obtained were identical (now shown), and the free energy of the structure was -31.6 kcal/ mol.
so obtained and the extent to which the sequence was deter- mined from each.
The nucleotide sequence, 1300 bp from the SmaI end of the DNA insert of pKR322, is shown in Fig. 6. This insert, which includes a previously sequenced segment (starting at nucleo- tide logo), encodes a long open reading frame.
There is some ambiguity about the exact location of the 5' end of the protein. The first methionine in this sequence is found in the triplet starting at nucleotide 182. This methio-
nine is an obvious potential candidate for initiation of trans- lation. However, the only classical ribosome binding site (32) in this area is found 35 nucleotides upstream of the methio- nine. Very close and downstream of this Shine-Dalgarno sequence, there are three ATT triplets which have been reported in other systems as sites for initiation of translation (33).
The promoter has not yet been identified. However, up- stream of the open reading frame there are sequences (under-

10414 Sequence of Na+/H+ Antiporter Gene
lined in Fig. 6) that might represent promoter elements (34). The open reading frame starting at the methionine is ca-
pable of encoding a polypeptide which contains 362 amino acids (relative molecular mass = 38,683). The calculated M, is slightly higher than the value of -35,000 measured by SDS- polyacrylamide gel electrophoresis, a property reported for many integral membrane proteins.
When the hydropathy of the protein along the amino acid sequence is evaluated according to the method of Engelman et al. (35), it becomes apparent that the product of the ant gene contains 10 putative transmembrane spanning segments linked by hydrophilic segments of variable length (Fig. 7). According to this analysis, the first amino acid of each of the hydrophobic segments would be: 32, 70,99, 129, 153, 179,231, 253, 302, and 335 (Fig. 8). The largest hydrophilic areas can be detected at the amino terminus of the chain (amino acids 1-31), and at the segments that span from amino acid 53-69, 199-230, 273-301. Almost identical results were obtained when the hydropathy was evaluated by the method of Kyte and Doolittle (36).
A striking feature in the 5"region of the gene is the presence of a quite extensive putative secondary structure in the RNA. The minimal energy foldings in the segment between the bases 75 and 275 were calculated (371, and a stable structure (free energy-45.2 kcal/mol) can be predicted on this basis (Fig. 9). It has been claimed that this type of structure overlapping with the area of initiation of translation reduces translation efficiency (38). Several other factors could, in theory, explain either by themselves or in a concerted manner the poor translation efficiency observed in the T7 polymerase expression system (Fig. 5). Among them it is worth noticing that there is no classical ribosome binding site upstream of the first methionine or that the initiation of translation may be located in a nonclassical codon.
CONCLUDING REMARKS
We have subcloned and sequenced the DNA insert bearing the ant gene of E. coli. When cloned in a multicopy plasmid, this gene causes an increased rate of Na+/H+ antiporter activity.
A membrane-bound protein with an apparent M, in SDS- polyacrylamide gel electrophoresis of 35,000 was detected using the in vivo T7 polymerase expression system.
In the DNA insert we found an orf which encodes for a 362-amino-acid-long protein (Mr = 38,683). Analysis of the predicted sequence of the protein using either the Engelman or the Kyte-Doolittle paradigms suggests the presence of 10 putative transmembrane segments in the protein. A stem- loop structure in the upstream region of the gene was also predicted which may affect transcription and/or translation.
We suggest that the ant gene codes for the Na+/H+ anti- porter or one of its subunits. A proof for this suggestion is being searched now by purification of the active antiporter and comparison of part of the protein sequence to the gene sequence.
Acknowledgments-We thank H. Sorek, C. Prody, and A. Ginatt for help in the sequencing; Amos Oppenheim and Gad Glaser for helpful discussion; Y. Deutsh for the preparation of the oligonucleo- tides; R. Macnab for helpful discussions and for running the FOAM V8.4 program (to analyze protein hydropathy), and D. Kornitzer for running the FOLD program (to analyze RNA secondary structure). The use of UWGCG Software is funded by the David and Irene Sala Molecular Genetics Laboratory.
REFERENCES 1. Krulwich, T. A. (1986) J. Membr. Biol. 89, 113-125 2. Leblanc, G., Bassilana, M., and Damiano, E. (1987) Comp. Reu.
3. West, I. C., and Mitchell, P. (1974) Biochem. J. 144,87-90 4. Rosen, B. P. (1986) Annu. Reu. Microbiol. 40, 263-286 5. Castle, A. M., Macnab, R. M., and Schulman, R. G . (1986) J.
6. Tsuchiya, T., Raven, J., and Wilson, T. H. (1977) Biochem.
7. Stewart, L. M. D., and Booth, I. R. (1983) FEMS Microbiol. Lett.
8. MacDonald, R., Lanyi, S. K., and Green, R. V. (1977) Proc. Natl.
9. Ishikawa, T., Hama, H., Tsuda, M., and Tsuchiya, T. (1987) J .
10. Hirota, N., Kitada, M., and Imae, Y. (1981) FEBS Lett. 132 ,
11. Osterhelt, D., Hartmann, R., Michel, H., and Wagner, G. (1978) in Energy Conservation in Biological Membranes (Schager, G.,
12. Schuldiner, S., and Fishkes, H. (1978) Biochemistry 1 7 , 706-711 and Klingenberg, M., eds) Springer Verlag, Heidelberg
13. Brown, I. I., Galperin, M. Y., Glagolev, A. N., and Skulachev, V.
14. Padan, E., Zilberstein, D., and Rottenberg, H. (1976) Eur. J.
15. Padan, E., Zilberstein, D., and Schuldiner, S. (1981) Biochim.
16. Booth, I. R. (1985) Microbiol. Reu. 49, 354-378 17. Zilberstein, D., Padan, E., and Schuldiner, S. (1980) FEBS Lett.
116,177-180 18. Niiya, S., Yamasaki, K., Wilson, T. H., and Tsuchiya, T. (1982)
J. Biol. Chem. 257,8902-8906 19. Goldberg, E. B., Arbel, T., Chen, J., Karpel, R., Mackie, G. A.,
Schuldiner, S., and Padan, E. (1987) Proc. Natl. Acad. Sci. U.
Chem., in press
Biol. Chem. 261,3288-3294
Biophys. Res. Commun. 7 6 , 26-31
19,161-164
Acad. Sci. U. S. A. 7 4 , 3167-3170
Biol. Chem. 2 6 2 , 7443-7446
278-280
P. (1983) Eur. J. Biochem. 134,345-349
Biochem. 63,533-541
Biophys. Acta 6 5 0 , 151-166
S. A. 84,2615-2619 20. Mackie, G. A. (1986) Nucleic Acids Res. 14 , 6965-6981 21. Mackie, G. A., and Parsons, G. D. (1982) Can. J. Biochem. 6 0 ,
22. Davis, B. D., and Mingioli, E. S. (1950) J. Bacteriol. 6 0 , 17-28 23. Dotto, P. D., and Zinder, N. D. (1984) Nature 3 1 1 , 279-280 24. Tabor, S., and Richardson, C. C. (1985) Proc. Natl. Acad. Sci. U.
25. Yanisch-Perron, C., Vieira, J., and Messing, J. (1985) Gene
26. (Davies, L. G., Dibner, M. D., and Batley, J. F., eds) (1986) Methods in Molecular Biology, Elsevier Scientific Publishing Co., New York
338-346
S. A. 82,1074-1078
(Amst.) 33,103-119
27. Ruther (1980) Mol. & Gen. Genet. 178,475-477 28. Sanger, F., Nicklen, S., and Coulso, A. R. (1977) Proc. Natl. Acad.
29. Sanger, F., and Couison, A. R. (1978) FEBS Lett. 87, 107-110 30. Rosen, B. P. (1986) Methods Enzymol. 125,328-336 31. Ambudkar, V., Zlotnick, G. W., and Rosen, B. P. (1984) J. Biol.
32. Shine, J., and Dalgarno, L. (1974) Proc. Natl. Acad. Sci. U. S. A.
33. Sacerdot, C., Fayat, G., Dessen, P., Springer, M., Plumbridge, J. A,, Grunberg-Manago, M., and Blanquet, S. (1982) EMBO J.
Sci. U. S. A. 74 , 5463-5467
Chem. 259,6142-6146
71,1342-1346
1,311-315 34. Pribnow, D. (1975) Proc. Natl. Acad. Sei. U. S. A. 72,784-789 35. Engelman, D. M., Steitz, T. A,, and Goldman, A. (1986) Annu.
36. Kyte, J., and Doolittle, R. F. (1982) J. Mol. Biol. 157, 105-132 37. Freier, S. M., Kierzek, R., Jaeger, J. A., Sugimoto, N., Caruthers,
M. H., Nielson, T., and Turner, D. H. (1986) Proc. Natl. Acad. Sci. U. S. A . 83,9373-9377
38. Steitz, A. J. (1979) Biological Regulation and Development (Gold- berger, R. F., ed) Vol. 1, pp. 349-399, Plenum Publishing Corp, New York
39. Devereux. J.. Haeberli. P.. and Smithies, 0. (1984) Nucleic Acids
Rev. Biophys. Biophys. Chern. 15,321-353
~~ ~~
Res. 12,387-395
(1951) J. Biol. Chem. 193,265-275
, ,
40. Lowry, 0. H., Rosebrough, N. J., Farr, A. L., and Randall, R. J.


















