
The occurrence of NADPH-ferrihemoprotein reductase in Corbula caribea, from a natural oil seep at La...
6
The occurrence of NADPH-ferrihemoprotein reductase in Corbula caribea, from a natural oil seep at La Brea, Trinidad Azad Mohammed * , John Agard Department of Life Sciences, The University of the West Indies, St. Augustine, Trinidad, Trinidad and Tobago Abstract Corbula caribea is the most common non-polychaete macrofaunal organism identified at a large natural oil seep at La Brea in south Trinidad. It is hypothesized that these animals may possess (NADPH-ferrihemoprotein reductase) a component of the Mixed Function Oxygenase system (MFO), which may allow them to ameliorate the potentially deleterious effects resulting from exposure to the high levels of petroleum hydrocarbons within this environment. This study was designed to determine whether organisms from the seep site showed greater enzyme activity when compared to organisms from a non-seep reference site. NADPH-ferrihemoprotein reductase activity was determined by incubating 10 lm cryostat sections with nitro-blue tetrazolium. The reaction product was determined by visual assessment and quantified by measuring the relative mean stain intensity. The intense staining, indicative of enzyme activity was evident in the digestive epithelia of seep animals. Observations indicated that organisms from the seep showed more intense staining, indicating greater enzyme activity, when compared to animals from a non-seep ref- erence site. The relative stain intensity of NADPH-ferrihemoprotein reductase determined for organisms from the seep was 61.30. This was significantly higher than the stain intensity determined for organisms from the non-seep reference site (7.11). This sup- ported visual assessments, which suggested that the seep organisms showed higher enzyme activity than organisms from the non- seep site. The results suggest that NADPH-ferrihemoprotein reductase may be present in Corbula caribea from the seep site and not in those from the non-seep site. It is possible that this enzyme may contribute to these animals ability to tolerate chronic exposure to petroleum hydrocarbons and offer then a selective advantage for survival the seep environment. Ó 2003 Elsevier Ltd. All rights reserved. Keywords: NADPH-ferrihemoprotein reductase; Natural oil seep; Corbula caribea; Mixed function oxidase; Trinidad 1. Introduction Petroleum hydrocarbons pose a pervasive treat to the survival of organisms. However, many organisms pos- sess an innate ability to tolerate or reduce its toxic ef- fects. This ability is reportedly linked to the presence of the Mixed Function Oxygenase (MFO) enzyme system (Bayne et al., 1985). The MFO system consists of at least two protein components: 1. Cytochrome P-450––an iron containing haem protein and 2. NADPH-ferrihemoprotein reductase (NADPH Cytochrome P-450 or neotetrazolium reductase). Various studies have identified MFO activity in dif- ferent marine invertebrates, particularly crustaceans and polychaetes. Such studies include: MFO activity in the stomach of the blue crab, Callinecties sapidus (Singer et al., 1980); pollution effects on MFO activity in the marine polychaete, Nereis virens (Fries and Lee, 1984); Payne (1977) identified MFO activity in teleost, some crustaceans, an elasmobranch, annelids and an echino- derm but was generally absent from gastropods and pelecypod molluscs, coelenterates and macrophytic algae. Lee (1981) reported MFO activity in 18 marine invertebrates belonging to four phyla (Annelida, Arthropoda, Echinodermata and Mollusca) and Lee and Singer (1980) detected activity in the lower portions of the intestine of Capitella capitata after exposure to either petroleum or its components. Cytochrome P-450 activity has been identified in a variety of organisms, including fishes, mammals, birds * Corresponding author. Tel.: +1-868-662-2002x2047; fax: +1-868- 663-5241. E-mail address: [email protected] (A. Mohammed). 0025-326X/$ - see front matter Ó 2003 Elsevier Ltd. All rights reserved. doi:10.1016/j.marpolbul.2003.11.009 www.elsevier.com/locate/marpolbul Marine Pollution Bulletin 48 (2004) 784–789
-
Upload
azad-mohammed -
Category
Documents
-
view
212 -
download
0
Transcript of The occurrence of NADPH-ferrihemoprotein reductase in Corbula caribea, from a natural oil seep at La...

www.elsevier.com/locate/marpolbul
Marine Pollution Bulletin 48 (2004) 784–789
The occurrence of NADPH-ferrihemoprotein reductasein Corbula caribea, from a natural oil seep at La Brea, Trinidad
Azad Mohammed *, John Agard
Department of Life Sciences, The University of the West Indies, St. Augustine, Trinidad, Trinidad and Tobago
Abstract
Corbula caribea is the most common non-polychaete macrofaunal organism identified at a large natural oil seep at La Brea in
south Trinidad. It is hypothesized that these animals may possess (NADPH-ferrihemoprotein reductase) a component of the Mixed
Function Oxygenase system (MFO), which may allow them to ameliorate the potentially deleterious effects resulting from exposure
to the high levels of petroleum hydrocarbons within this environment. This study was designed to determine whether organisms
from the seep site showed greater enzyme activity when compared to organisms from a non-seep reference site.
NADPH-ferrihemoprotein reductase activity was determined by incubating 10 lm cryostat sections with nitro-blue tetrazolium.
The reaction product was determined by visual assessment and quantified by measuring the relative mean stain intensity. The intense
staining, indicative of enzyme activity was evident in the digestive epithelia of seep animals. Observations indicated that organisms
from the seep showed more intense staining, indicating greater enzyme activity, when compared to animals from a non-seep ref-
erence site. The relative stain intensity of NADPH-ferrihemoprotein reductase determined for organisms from the seep was 61.30.
This was significantly higher than the stain intensity determined for organisms from the non-seep reference site (7.11). This sup-
ported visual assessments, which suggested that the seep organisms showed higher enzyme activity than organisms from the non-
seep site.
The results suggest that NADPH-ferrihemoprotein reductase may be present in Corbula caribea from the seep site and not in
those from the non-seep site. It is possible that this enzyme may contribute to these animals ability to tolerate chronic exposure to
petroleum hydrocarbons and offer then a selective advantage for survival the seep environment.
� 2003 Elsevier Ltd. All rights reserved.
Keywords: NADPH-ferrihemoprotein reductase; Natural oil seep; Corbula caribea; Mixed function oxidase; Trinidad
1. Introduction
Petroleum hydrocarbons pose a pervasive treat to the
survival of organisms. However, many organisms pos-
sess an innate ability to tolerate or reduce its toxic ef-fects. This ability is reportedly linked to the presence of
the Mixed Function Oxygenase (MFO) enzyme system
(Bayne et al., 1985). The MFO system consists of at least
two protein components:
1. Cytochrome P-450––an iron containing haem protein
and
2. NADPH-ferrihemoprotein reductase (NADPHCytochrome P-450 or neotetrazolium reductase).
* Corresponding author. Tel.: +1-868-662-2002x2047; fax: +1-868-
663-5241.
E-mail address: [email protected] (A. Mohammed).
0025-326X/$ - see front matter � 2003 Elsevier Ltd. All rights reserved.
doi:10.1016/j.marpolbul.2003.11.009
Various studies have identified MFO activity in dif-
ferent marine invertebrates, particularly crustaceans and
polychaetes. Such studies include: MFO activity in the
stomach of the blue crab, Callinecties sapidus (Singer
et al., 1980); pollution effects on MFO activity in themarine polychaete, Nereis virens (Fries and Lee, 1984);
Payne (1977) identified MFO activity in teleost, some
crustaceans, an elasmobranch, annelids and an echino-
derm but was generally absent from gastropods and
pelecypod molluscs, coelenterates and macrophytic
algae. Lee (1981) reported MFO activity in 18 marine
invertebrates belonging to four phyla (Annelida,
Arthropoda, Echinodermata and Mollusca) and Leeand Singer (1980) detected activity in the lower portions
of the intestine of Capitella capitata after exposure to
either petroleum or its components.
Cytochrome P-450 activity has been identified in a
variety of organisms, including fishes, mammals, birds

Fig. 1. Map showing the proximity of the seep and non-seep reference
sites, in relation to petroleum hydrocarbon contamination in the Gulf-
of-Paria, Trinidad and Tobago (adapted from Agard et al., 1988).
A. Mohammed, J. Agard / Marine Pollution Bulletin 48 (2004) 784–789 785
and invertebrates. Quattrochi and Lee (1984) reported
cytochrome P-450 activity in several marine crabs,
Libinia emarginata, Callinectes sapidus, Menippe merce-
narie and Uca minax. Lee et al. (1982) identified in-creased activity of Cytochrome P-450 in the crabs,
Callinectes sapidus, Serarina cinerum and Uca pugilator,
and the polychaete (Neries virens) following exposure to
oil. Lee et al. (1982) also identified cytochrome P-450
activity in Uca minax collected from a heavily oiled area,
Uca pugilator collected from an oil spill site, Callinectes
sapidus from a heavy industrial area and in Sesarma
cinerum maintained in marsh sediment. Livingstone(1988) has also reported elevated levels of cytochrome
P-450 and cytochrome b5 in the digestive glands of
mussels exposed to diesel oil. Cytochrome P-450 activity
has also been reported in fish (Platichthys flesus), mol-
luscs (Mytilus edulis and Littorina littorea) and in crabs
(Carcinus maenas), (IOC/UNESCO, 1986). Kurelec
(1985) and Kurelec et al. (1986) suggested that in mol-
lusc, in which levels of P-450 are low, flavin-containingmonooxygenases may be the predominant phase 1
enzyme.
NADPH-ferrihemoprotein reductase is involved in
the electron transfer system of the endoplasmic reticu-
lum. According to Bayne et al. (1985), Moore (1988)
reported that there was no definitive evidence to sug-
gest induction of MFO activity in bivalves, but evidence
of induction of components of the system (NADPH-ferrihemoprotein reductase) has been observed.
However, studies have shown increased activity of
microsomal NADPH-ferrihemoprotein reductase in
Mytilus edulis blood cells following exposure to pheno-
barbital (Moore, 1979). Moore et al. (1987) also re-
ported elevated levels of NADPH-ferrihemoprotein
reductase activity in L. littorea from contaminated sites,
when compared to animals for a clean reference site, andincreased activity following exposure to 400 lg/l of
phenanthrene. Moore (1988) also showed that elevated
levels of NADPH-ferrihemoprotein in the digestive cells
ofMytilus edulis and L. littorea, correlated directly with
total PAH concentration.
The La Brea seep in Trinidad was identified as one of
the three largest natural oil seeps known (Wilson et al.,
1974). Persistent oil intrusion saturates this area withpetroleum hydrocarbons (Agard et al., 1988), making
the environment harsh to the survival of animals.
However, the occurrence of thriving benthic communi-
ties with this environment despite the persistent oil
intrusion, may suggest that organisms here have innate
homeostatic mechanisms to help reduce potentially toxic
effects. In a study conducted in parallel with this work,
Agard et al. suggested that Corbula caribea was the mostabundant non-polychaete organism (46 individuals per
sq. meter) identified at the seep. Surveys conducted
during this research on the western Gulf coast of Trin-
idad, failed to identify any areas with such high densities
of organisms. This present study focused on determining
if NADPH-ferrihemoprotein reductase can be identified
in organisms from the seep site.
2. Methodology
Corbula caribea was collected from a natural oil seep
at La Brea and a non-seep reference site at Carenage,
about 50 km (Fig. 1) north of the seep site. Animals were
collected using as a petit ponar grab and transported to
the laboratory alive.
2.1. Tissue preparation for cytochemistry
Organisms were immediately removed from their
shells and rinsed twice in molluscan saline [0.4 M NaCl,
0.05 M CaCl2 and 0.08 M MgCl2], embedded in a small
amount of Tissue Tek (O.C.T 4583, Sukura Finetechni-
cal Co.) and frozen at )70 �C. Tissue blocks were sub-sequently sectioned at 10 lm using a Re�ıchert Jung
Cryocut 1800 (Eine Gesellsenaft Der Cambridge
instrument) at a cabinet temperature of )15 �C. Sectionswere then flash dried on to clean slides and stored at
)70 �C. Histochemical analysis was done within 24 h
(Pearse, 1980; Kiernan, 1981; Moore, 1988).
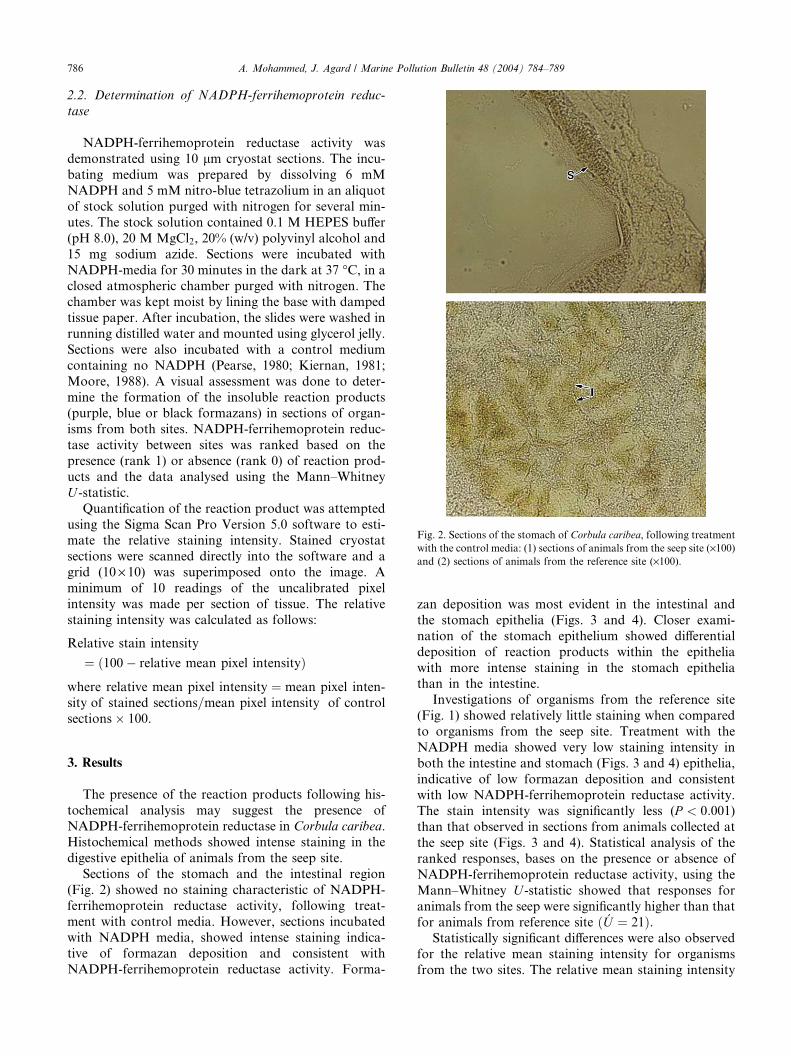
Fig. 2. Sections of the stomach of Corbula caribea, following treatment
with the control media: (1) sections of animals from the seep site (·100)and (2) sections of animals from the reference site (·100).
786 A. Mohammed, J. Agard / Marine Pollution Bulletin 48 (2004) 784–789
2.2. Determination of NADPH-ferrihemoprotein reduc-
tase
NADPH-ferrihemoprotein reductase activity wasdemonstrated using 10 lm cryostat sections. The incu-
bating medium was prepared by dissolving 6 mM
NADPH and 5 mM nitro-blue tetrazolium in an aliquot
of stock solution purged with nitrogen for several min-
utes. The stock solution contained 0.1 M HEPES buffer
(pH 8.0), 20 M MgCl2, 20% (w/v) polyvinyl alcohol and
15 mg sodium azide. Sections were incubated with
NADPH-media for 30 minutes in the dark at 37 �C, in aclosed atmospheric chamber purged with nitrogen. The
chamber was kept moist by lining the base with damped
tissue paper. After incubation, the slides were washed in
running distilled water and mounted using glycerol jelly.
Sections were also incubated with a control medium
containing no NADPH (Pearse, 1980; Kiernan, 1981;
Moore, 1988). A visual assessment was done to deter-
mine the formation of the insoluble reaction products(purple, blue or black formazans) in sections of organ-
isms from both sites. NADPH-ferrihemoprotein reduc-
tase activity between sites was ranked based on the
presence (rank 1) or absence (rank 0) of reaction prod-
ucts and the data analysed using the Mann–Whitney
U -statistic.
Quantification of the reaction product was attempted
using the Sigma Scan Pro Version 5.0 software to esti-mate the relative staining intensity. Stained cryostat
sections were scanned directly into the software and a
grid (10 · 10) was superimposed onto the image. A
minimum of 10 readings of the uncalibrated pixel
intensity was made per section of tissue. The relative
staining intensity was calculated as follows:
Relative stain intensity
¼ ð100� relative mean pixel intensityÞ
where relative mean pixel intensity ¼ mean pixel inten-sity of stained sections=mean pixel intensity of control
sections� 100.
3. Results
The presence of the reaction products following his-
tochemical analysis may suggest the presence ofNADPH-ferrihemoprotein reductase in Corbula caribea.
Histochemical methods showed intense staining in the
digestive epithelia of animals from the seep site.
Sections of the stomach and the intestinal region
(Fig. 2) showed no staining characteristic of NADPH-
ferrihemoprotein reductase activity, following treat-
ment with control media. However, sections incubated
with NADPH media, showed intense staining indica-tive of formazan deposition and consistent with
NADPH-ferrihemoprotein reductase activity. Forma-
zan deposition was most evident in the intestinal and
the stomach epithelia (Figs. 3 and 4). Closer exami-
nation of the stomach epithelium showed differential
deposition of reaction products within the epithelia
with more intense staining in the stomach epitheliathan in the intestine.
Investigations of organisms from the reference site
(Fig. 1) showed relatively little staining when compared
to organisms from the seep site. Treatment with the
NADPH media showed very low staining intensity in
both the intestine and stomach (Figs. 3 and 4) epithelia,
indicative of low formazan deposition and consistent
with low NADPH-ferrihemoprotein reductase activity.The stain intensity was significantly less (P < 0:001)than that observed in sections from animals collected at
the seep site (Figs. 3 and 4). Statistical analysis of the
ranked responses, bases on the presence or absence of
NADPH-ferrihemoprotein reductase activity, using the
Mann–Whitney U -statistic showed that responses for
animals from the seep were significantly higher than that
for animals from reference site ð �U ¼ 21Þ.Statistically significant differences were also observed
for the relative mean staining intensity for organisms
from the two sites. The relative mean staining intensity

Fig. 3. Sections of the intestine showing the varying degrees of staining following treatment with the NADPH media: (1) sections of animals from
reference site (·100) and (2) sections of animals from the seep (·100).
Fig. 4. Sections of the stomach showing the varying degrees of staining following treatment with the NADPH media: (1) sections of the animals from
reference site (·100) and (2) sections of animals from the seep (·100).
Table 1
Relative mean staining intensity of NADPH-ferrihemoprotein reductase in Corbula caribea from three sites in Trinidad
Sites Relative mean stain intensity of NADPH-ferrihemoprotein reductase in Corbula caribea, as a
percent of the controls
Intensity of the controls Relative mean stain intensity
Seep site 231.85± 16.46 61.30± 3.46
Non-seep contaminated 200.04± 15.66 7.11± 6.21
A. Mohammed, J. Agard / Marine Pollution Bulletin 48 (2004) 784–789 787
for organisms from the seep was 61.30 ± 3.46 as com-
pared to 7.11 ± 6.21 (Table 1) for the non-seep uncon-
taminated site (ANOVA; Fcal ¼ 57:63, Ftab ¼ 9:55, P <0:001Þ.

788 A. Mohammed, J. Agard / Marine Pollution Bulletin 48 (2004) 784–789
4. Discussion
Cytochemical analyses suggest that NADPH-ferrihe-
moprotein may be present in the digestive epithelia ofthe marine bivalve Corbula caribea found at the seep
site. According to Bayne et al. (1985), Moore (1988)
suggested that there was no definitive evidence of induc-
tion of MFO activity in bivalves, though evidence of
NADPH-neotetrazolium reductase and NADPH pro-
ducing enzymes was present. Studies conducted in par-
allel with this work, Agard et al. suggested that Corbula
caribeawas the most abundant non-polychaete organism(46 individuals per sq. meter) identified at the seep.
Additional surveys conducted during the duration of this
project along the Gulf coast of Trinidad, failed to iden-
tify such densities at any other location even at the ref-
erence site. This enzyme is involved in the detoxification
pathway for petroleum hydrocarbons, in particular
PAHs. It is probable that this enzyme may contribute to
the ability of Corbula caribea to adapt survive within theseep environment. Agard et al. (1988) reported varying
concentration of petroleum hydrocarbons (3.0–1824.4
lg/g dry wt. Crysene equivalent) along the West Coast of
Trinidad. The area, which they identified as showing the
highest contamination (Fig. 1), coincided with the seep
site under investigation. The reference site showed only
low levels of petroleum hydrocarbons.
Analysis of Corbula caribea, obtained from two sitesshowed significant differences in stain intensity, which
suggest that there was a difference in enzyme activity
between the seep and non-seep organisms. Animals from
the seep showed higher staining intensity when com-
pared to animals from an uncontaminated reference site.
It is therefore possible that the presence of NADPH-
ferrihemoprotein reductase may be an adaptive response
to exposure to high levels of petroleum hydrocarbons.The differential NADPH-ferrihemoprotein reductase
activity observed in animals from both sites may not be
due to geographical separation of populations, but may
be more closely related to the levels of hydrocarbons
within the environment. It is therefore probable that the
presence of elevated levels of NADPH-ferrihemoprotein
reductase in Corbula caribea at the seep may be an
adaptive response to the high levels of hydrocarbonsin the environment. Further, it is evident that there
is a higher concentration of NADPH-ferrihemoprotein
reductase in the stomach than in the intestine. Moore
(1988), also observed higher NADPH-ferrihemoprotein
reductase activity in Mytilus edulis and L. littorea col-
lected from contaminated sites, than in animals collected
from an uncontaminated site. Similar results were also
reported for L. littorea from contaminated sites (Mooreet al., 1987). The stomach would represent the region
where the most direct exposure would take place. Food
material and sediment particles that would be ingested
may undoubtedly contain high levels of hydrocarbons,
which would have to be detoxified before toxic effect
can manifest. This detoxification can be facilitated by
the NADPH-ferrohemoprotein reductase. It may also
be possible that these organisms associated miofaunalcommunities in areas such as gills which can also serve
to reduce the toxic effect of the hydrocarbons through
absorption and biotransformation, however, this has yet
to be investigated.
The hydrocarbons within the area may also serve as a
carbon source for organisms within the seep ecosystem.
Dhanraj (1999) was able to identify significantly high
levels of oil degrading bacteria at the seep site. Thesebacteria may be able to succeed in transforming the
carbon from the petroleum hydrocarbons and incorpo-
rating it into the food chain. This may be one of the
contributing factors for the high numbers of organisms
identified at the seep.
References
Agard, J.B.R., Gobin, J., Boodosingh, M., 1988. Petroleum residues in
surficial sediments from the Gulf-of-Paria, Trinidad. Mar. Pollut.
Bull. 19, 231–233.
Bayne, B.L., Brown, D.A., Burns, K., Dixon, D.R., Ivanovici, A.,
Livingstone, D.R., Lowe, D.M., Moore, M.N., Stebbing, A.R.D.,
Widdows, J., 1985. The Effects of Stress and Pollution on Marine
Animals. Praeger Publishers, New York.
Dhanraj, D.K., 1999. The ecology of hydrocarbon degrading bacteria
at a natural oil seep. M.Phil Thesis. Department of Life Sciences,
University of the West Indies, St Augustine, Trinidad and
Tobago.
Fries, C.R., Lee, R.F., 1984. Pollutant effects on the mixed function
oxygenase (MFO) and reproductive system of marine polychaete
(Nieris virens). Mar. Biol. 79, 187–193.
Intergovernmental Oceanographic Commission (IOC), 1986. IOC
workshop on the biological effects of pollutants. No. 53, UNESCO,
48p.
Kiernan, J.A., 1981. Histological and Histochemical Methods, Theory
and Practices. Pergamon Press, Oxford.
Kurelec, B., 1985. Exclusion activation of aromatic amines in the
marine mussel (Mytilus edulis) by FAD containing monooxygena-
ses. Biochem. Biophys. Res. Commun. 127, 773–788.
Kurelec, B., Brituik, S., Krea, S., Zahn, R.K., 1986. Metabolic fate of
aromatic amines in the mussel (Mytilus galoprovincialis). Mar. Biol.
91, 523–527.
Lee, R.F., 1981. Mixed Function Oxygenase (MFO) in marine
invertebrates. Mar. Biol. Lett. 2, 87–105.
Lee, R.F., Singer, S.C., 1980. Detoxifying enzyme systems in marine
polychaetes: increases in activity after exposure to aromatic
hydrocarbons. Rap. P.-V. R�eun. Cons. Int. Explor. Mer. 179,
29–32.
Lee, R.F., Conner, J.W., Page, D., Ray, L.E., Giam, C.S., 1982.
Cytochrome P-450 dependent Mixed Function Oxygenase system
in marsh crabs. Physiol. Mech. Mar. Pollut. Toxic.
Livingstone, D.R., 1988. Responses of microsomal NADPH-cyto-
chrome c reductase activity and cytochrome P-450 in the digestive
glands inMytilus edulis and Littorina littorea to environmental and
experimental exposure to pollutants. Mar. Ecol. Prog. Ser. 46 (1–
3), 37–43 (MEPS special. Biological Effects of Pollutants: Results
of a Practical Workshop).
Moore, M.N., 1988. Cytochemical responses of the lysosomal
system and NADPH-ferrihemoprotein reductase in the molluscan

A. Mohammed, J. Agard / Marine Pollution Bulletin 48 (2004) 784–789 789
digestive cells to environmental and experimental exposure to
xenobiotics. Mar. Ecol. Prog. Ser. 46, 81–89.
Moore, M.N., 1979. Cellular responses to polyaromatic hydrocarbons
and phenobarbital inMytilus edulis. Mar. Environ. Res. 2, 255–263.
Moore, M.N., Pipe, R.K., Farrar, S.V., Thomson, S., Donkin, P.,
1987. Lysosomal and microsomal responses to oil-derived hydro-
carbons in Littorina littorea. In: Capuzzo, J.M., Kester, D.R.
(Eds.), Oceanic Processes in Marine Pollution; Biological Processes
and Waste in the Oceans, 1. Robert E. Kriege Publishing
Company, Florida, pp. 89–97.
Pearse, A.G.E., 1980. Histochemistry: Theoretical and Applied, Vol. 1:
Preparation and optical technique. In: Preparation and Optical
Technique, vol. 1. J&A Churchill Publications, London.
Payne, J.F., 1977. Mixed Function Oxygenase in marine organisms in
relation to petroleum hydrocarbon metabolism and detection. Mar.
Pollut. Bull. 8 (5), 112–116.
Quattrochi, L.C., Lee, R.F., 1984. Microsomal cytochrome P-450
from marine crabs. Comp. Biochem. Physiol. 79C (1), 171–
176.
Singer, S.C., Paul Jr., E.M., Gonsoulin, F., Lee, R.F., 1980. Mixed
Function Oxygenase activity in the Blue crab, Callinectes sapidus,
characterization of enzyme activity from stomach tissue. Comp.
Biochem. Physiol. 65C, 129–134.
Wilson, R.D., Monaghan, P.H., Osanik, A., Price, L.C., Rogers,
M.A., 1974. Natural marine oil seeps. Science 184, 857–
865.


















