
The Muscles of the Adult Honey-bee (Apis mellifera L.). Guy D
89
The Muscles of the Adult Honey-bee (Apis mellifera L.). By Guy D. Morison, B.Sc, (Lond,), North of Scotland College of Agriculture, Aberdeen. With 41 Text-figures. PART II. THE HEALTHY MUSCLES OF THE ADULT HONEY-BEE. MUSCLES OF THE ALIMENTARY CANAL, HEART, DIAPHRAGMS, AND THE REPRODUCTIVE ORGANS, AND THE INDIRECT MUSCLES OF THE WINGS. CONTENTS. PAGE SPLANCHNIC MUSCLES OF THE ALIMENTARY CANAL . . . 563 CARDIAC AND DORSAL DIAPHRAGM MUSCLES . . . . 590 MUSCLES OF THE VBNTEAL DIAPHRAGM . . . . . 597 MUSCLES OF THE REPRODUCTIVE ORGANS . . . . . 603 EIBBOUS INDIRECT MUSCLES OF THE WINGS . . . . . 6 1 1 TABLE OE MEASUREMENTS RELATING TO FIBROUS MUSCLE . . 613 ATTACHMENT or MUSCLE . . . . . . . . 639 SUMMARY . . . . . . . . . . 645 BIBLIOGRAPHY . . . . . . . . . . 646 The Muscles of the Alimentary Canal. MUCH has been written about the alimentary canal of the bee, but almost invariably the writings were more concerned with supposed interpretations of the physiology of the parts under review than with the morphology or histology. As examples of the neglect of the histology of the alimentary canal, it may be mentioned that it was in 1918 that White described the third layer of muscles in the ventriculus, in 1923 that Trappmann described the muscles on the Malpighian tubes, and
Transcript of The Muscles of the Adult Honey-bee (Apis mellifera L.). Guy D

The Muscles of the Adult Honey-bee(Apis mellifera L.).
By
Guy D. Morison, B.Sc, (Lond,),North of Scotland College of Agriculture, Aberdeen.
With 41 Text-figures.
PART II .
THE HEALTHY MUSCLES OF THE ADULT HONEY-BEE.
MUSCLES OF THE ALIMENTARY CANAL, HEART, DIAPHRAGMS,
AND THE REPRODUCTIVE ORGANS, AND THE INDIRECT MUSCLES
OF THE WINGS.
CONTENTS.PAGE
S P L A N C H N I C M U S C L E S O F T H E A L I M E N T A R Y C A N A L . . . 5 6 3
C A R D I A C A N D D O R S A L D I A P H R A G M M U S C L E S . . . . 5 9 0
M U S C L E S O F T H E V B N T E A L D I A P H R A G M . . . . . 5 9 7
M U S C L E S O F T H E R E P R O D U C T I V E O R G A N S . . . . . 6 0 3
E I B B O U S I N D I R E C T M U S C L E S O F T H E W I N G S . . . . . 6 1 1
T A B L E O E M E A S U R E M E N T S R E L A T I N G T O F I B R O U S M U S C L E . . 6 1 3
A T T A C H M E N T o r M U S C L E . . . . . . . . 6 3 9
S U M M A R Y . . . . . . . . . . 6 4 5
B I B L I O G R A P H Y . . . . . . . . . . 6 4 6
The Muscles of the Alimentary Canal.MUCH has been written about the alimentary canal of the
bee, but almost invariably the writings were more concernedwith supposed interpretations of the physiology of the partsunder review than with the morphology or histology. Asexamples of the neglect of the histology of the alimentary canal,it may be mentioned that it was in 1918 that White describedthe third layer of muscles in the ventriculus, in 1923 thatTrappmann described the muscles on the Malpighian tubes, and

564 GUY D. MOBISON
1923 when Hertig recorded in detail the appearance of freshcells of the ventriculus. Naturally the alimentary canal of theworker has received more attention than that of the queen anddrone. In fact the comparative anatomy of the enteron in thethree castes is usually completely neglected, and science yetawaits such an account as will deal with the histology duringand after the post-embryonic metamorphic changes of the gut.Especially if such a work includes the tracheation and innerva-tion besides physiological interpretations. The histology of themuscles has been much more neglected than that of the epithelialor glandular parts, and many misstatements about it lie scat-tered in the literature. Zander (1922) gives a short account ofthe comparative structure of the gut in the three castes. Thenomenclature of Snodgrass (1925) for the divisions of thealimentary canal will be adopted in this paper.
A preliminary consideration in the study of the muscles ofthe alimentary canal is that this region is formed in the embryoby two ectodermal invaginations—the stomodeum and theproctodeum—between which there lies the mesenteron derivedfrom the anterior and posterior mesenteron rudiments. All themuscles are derived apparently from the mesoderm—those ofthe mesenteron froni the visceral layer of the mesoderm, thoseof the stomodeum and proctodeum presumably from an anteriorand a posterior mesodermal mass respectively. Nelson hasdescribed the embryology (1915) and the structure of the larva(1925) of the bee in minute detail, but we yet await a likedescription of the changes which take place during the post-embryonic metamorphosis of the bee, especially as all themuscles of the alimentary canal of the larva seem to disappearand then be replaced greatly augmented in, the adult bee.Evenius, J. (1925), and Evenius, C. (1926), have taken steps inthe right direction. Metzer (1910) describes how the stomodealregion passes along the thorax into the post-abdomen in thedeveloping pupa, but a mention of the histology is omitted.In the adult bee of all castes some of the longitudinal muscle-fibres of the honey-stomach or crop pass over the proventri-culus directly on to the walls of the ventriculus, and some of the

MUSCLES OF THE BEE 565
longitudinal fibres of the ventriculus pass directly on to thewalls of the small intestine without showing any histologicaldifference. The crop belongs to the stomodeal (ectodermal)invagin a tion, the ventriculus is the mesenteron which correspondswith an endodermal origin in animals of more typical developmentthan the bee, and the small intestine originates from the proctodeal(ectodermal) invagination, therefore a difference in structure ofthe muscles might have been expected to mark the division ofthese three regions, even though the muscles are of mesodermalorigin. There is a histological difference between the circularmuscles of the three regions. Correlated with the involution ofthe ectoderm to form the stomodeum, many of the muscles ofthis region are placed regularly and conform to the bilateralsymmetry of the insect besides having the histological structureof the typical somatic muscle-fibres. Such muscles do not occuron the proctodeum.
The muscles of every region of the alimentary canal willcontract spontaneously or may be induced to contract on thestimulus of touch, when large parts of the gut are removed froma living bee and are placed in warmed 0-75 per cent, salt solu-tion. The power of contractibility may last for a few minutes,but its duration is variable and may never be manifested. Thislast case is rare when it includes every group of muscles, but itseems to occur sometimes, and then I can only conclude that itmay be analogous to the sudden death which very rarely over-whelms a bee during the operation of having a first thoracicspiracle blocked. In both cases the age of the bee is question-able, but the insect will seem quite as healthy as its fellowswhich suffer exactly the same treatment, and which afterwardsshow very obvious signs of life. Perhaps the handling of thebee injures the nervous system, for death results in a few minutesafter a general paralysis, and yet this is difficult to reconcilewith the degree of independence of the ganglia of the nervoussystem as outlined previously (Morison, 1927).
Clear peristaltic waves passing from front to rear can be seenin the pharynx, oesophagus,-and in the small intestine of aliving bee; but since this means illumination by reflected light

566 GUY D. MORISON
and comparatively low powers of magnification, very little canbe seen of the muscles. The waves may be due to the passageof food, or may be induced by the stimulus of touch. I have notnoted clear peristaltic movement in the crop, ventriculus, orlarge intestine. The very energetic opening and closing move-ments of the proventriculus have been noted by Schonfeld(1886) and many other observers; but the muscles responsiblefor the movements are like the paired, very active muscles of thepharynx, and the two groups are histologically identical withthe paired somatic muscles of the body and more or less unlikethe rest of the muscles of the alimentary canal.
The histological structure of the living muscle-fibres of the .alimentary canal is less clearly seen than in somatic muscle-fibres, because of the opacity of the other layers of the gut.However, the transverse striation is usually seen as well aslongitudinal striae. Nuclei of the fibres are not distinguishablealthough they may be correctly located in the case of thecircular muscles of the ventriculus. The muscles are all practi-cally colourless. Excluding the muscles of the pharynx andproventriculus just mentioned, the characteristics of the musclesof the alimentary canal as opposed to the somatic muscles arethat they are unpaired and that their fibres usually branch andanastomose, whilst some of the fibres form more or less com-plete rings round the gut against which all tend to be flattened.
Much depends upon the staining of the muscles of the alimen-tary canal. Sectioned material mounted in Canada balsamafter embedding in paraffin wax is not very suitable for themuscles because the fibres tend to shrink 20-30 per cent, evenafter most careful treatment; also in most cases they appear asdark structureless rods in a preparation showing a clear pictureof the epithelial and other layers. If sectioned material isrequired, it is best to embed in clove-oil-ether-celloidin followedby hardening of the celloidin block in cedar-wood oil. Theblock can be cleared in the cedar-wood oil, but this is unneces-sary as long as it is firm enough to cut easily. Trimming theblock during the time it lies in the cedar-wood oil accelerates thehardening. The trimmed block should be blotted dry, washed

MUSCLES OF THE BEE 567
in xylol, blotted again, and then oriented on the block-holderof the microtome by slowly building round it a matrix of paraffinwax. The celloidin sections will not remain in the ribbon ofparaffin wax unless the celloidin block is very small compara-tively to the wax surrounding it, besides being of almost thesame hardness as the wax. Sections will cut perfectly at 3/u,thick. Usually there is no trouble in lifting the celloidinsection out of its frame of wax. Mayer's glycerine and albumenfixes the sections to the slide, and then after drying (of minimumduration) they should be placed in xylol to remove any tracesof paraffin wax, and then in a mixture of absolute alcohol andether to dissolve the celloidin. This will leave the sections ofanimal tissue still firmly adherent to the slide, and if they havenot been stained previously they can be stained by any usualmethod including a twenty-four hour mordanting in iron alumprevious to iron haematoxylin. The advantage of the methodis that contraction of the tissues is reduced to a minimum, andthat the whole process takes place at room temperature exceptfor the few minutes that the celloidin block is being encasedin paraffin wax. Even chitinous structures like the wall of thethorax with its attached muscles cut easily and well. Thedisadvantage is that the object embedded should be small (notmore than about 3 mm. cube). Parts completely enclosed intough chitin should be punctured to ensure the entrance of thecelloidin. The chief factor in the process is the very gradualpassage of the fixed tissue through the successive stages. Afterreaching 96 per cent, alcohol the embedding in celloidin isaccomplished very slowly by adding drops of a dilute solutionof celloidin dissolved in absolute alcohol or ether or a mixtureof both. Later some drops of clove oil are added, and then theobject is passed into a stronger mixture of celloidin in cloveoil and ether, and this is allowed to thicken by the evaporationof the ether. The last process is repeated till the object restsin a very thick solution of celloidin in clove oil. A lump of thissubstance containing the tissue is then dropped into chloroformto be hardened superficially, after which it is moulded into asuitable shape and dropped into cedar-wood oil. The process

568 GUY D. MOBISON
lasts about two. weeks for the alimentary canal of a bee, but thetime could be reduced if desired.
Whole mounts of the alimentary canal are the best for show-ing the muscles. The newly dead muscles from the gut of abee which has died apparently naturally, or from a bee killedby narcotics or cold, and the living muscles of the gut pulledout of a living bee and placed in fixative, will have the samehistological appearance if the same fixation and staining isemployed in each case. Formalin (10 per cent.) is the bestgeneral fixative for the alimentary canal when it is required tosee the tracheation and the relationships of the muscle-fibresto one another. It can be followed by staining in aqueous oralmost aqueous solutions of various stains and counterstainsbefore mounting in dilute glycerine which is allowed to becomepure through evaporation and addition. This treatment willleave the size of the muscle-fibres unaltered besides demonstrat-ing the distribution of tracheae and tracheoles. Ranvier's gold-chloride method also retains the air in the tracheae and showsthe arrangement of the muscles, but the prolonged acid treat-ment swells the sarcostyles and the appearance of the fibres isnot quite natural.
I have not been able to consult Brandt's paper on the nervoussystem of the Apidae. All text-books and original papers thatI have consulted on the morphology of the various regions of thealimentary canal completely ignore or are very vague aboutthe innervation of the alimentary canal. In the three castes thestomatogastric nervous system innervates the pharynx, oeso-phagus, honey-stomach, and probably the proventriculus ; thesplanchnic nerves of the last abdominal ganglion of the centralnervous system innervate the ventriculus, small intestine, andlarge intestine ; I do not know if the ventral sympatheticnervous system has any contact with the gut.
The stomatogastric, so-called ' sympathetic ', nervous systemconsists of an anterior f r o n t a l g a n g l i o n lying in front ofthe brain on the top of the pharynx between the muscles, andof two smaller p h a r y n g e a l g a n g l i a placed just behindthe brain on either side of the pharynx. The pharyngeal ganglia

MUSCLES OF THE BEE 569
lie just behind the pair of long, delicate muscles which arise wherethe pharynx merges into oesophagus and which serve to attachthe alimentary canal to the back of the head. Snodgrass (1925)mentions these ganglia and some of the nerves connected withthem, but for a more detailed account of this system in insectsOrlov (1924), Imms (1925), or Snodgrass (1926) should be con-sulted. The c o r p o r a a l l a t a lie between and above thepharyngeal ganglia and apparently in contact with them. Un-fortunately this glandular mass receives no mention in moderntext-books on the bee in spite of Nabert's (1913) detailed accountof it and its morphological relationships to the surroundingtissues. Evidently this is the glandular region found by Pixell-Goodrich (1920). From the anterior frontal ganglion a mediannerve runs forward, giving off many branches to the musclesof the pharynx. Two lateral branches, f r o n t a l c o m m i s -s u r e s , unite the frontal ganglion with the tritocerebrum. Thefrontal commissures for a part of their course, as well as someof the other nerves of the stomatogastric system lying in thehead, are enclosed by ' inversed ' tracheae such as described byJanet (1911). A median r e c u r r e n t n e r v e passes backfrom the frontal ganglion to branch just before uniting with thelateral pharyngeal ganglia. Each pharyngeal ganglion has aconnective passing to the brain. Prom each pharyngeal gan-glion a large nerve runs back along the oesophagus, giving offmany branches which anastomose more or less with one anotherand with those from the other ganglion. The nerves traversethe entire length of the oesophagus and crop, and their size andposition suggests that they may lead to the twigs innervating themuscles of the proventriculus, though I have never seen a con-nexion between the nerves of the crop and the proventriculus.All the larger branches of the stomato-gastric nerves tend to besomewhat zigzag in their course near and on the crop, and to berather wrinkled in appearance. This would allow the crop to ex-pand without undue stretching of the nerves. The nerves of therectum, the only other region of the alimentary canal which suffersconsiderable distension, never show a similar appearance, sincethey are differently arranged and are much more delicate.

570 GUY D. MORISON
The ganglia of the stomatogastric nervous system containmany large, intensely staining, motor cytons with sphericalnuclei (average 9//. diameter). Besides these nuclei there aresmaller, more elliptical nuclei, which are more peripheral and.apparently belong to the neurilemma. All the nuclei in thenerves are like the smaller nuclei, but are more variable in size,5:4:4-12:6:6/x. The nerves supplying muscle-fibres anasto-
TEXT-PIG. 1.
Stomato-gastric nerve plexus on the muscles of the crop. Flemming'sfixative, iron haematoxylin, eosin.
LETTERING FOE TBXT-ITOS. 1-4.
a, nerve axons ; c m, circular muscle-fibres ; gang, ganglion ; I m,longitudinal muscle-fibres; neur, neurilemma; nu, nucleus ;nv, nerve ; nv e, nerve-end.
mose very completely. At the places of anastomosis or branch-ing they often expand into ganglion-like swellings which seemto be similar to the expansions described (Morison, 1927) forthe nerves of somatic muscles as probable trophic, centres.The nerves pass above and below and between the circular andlongitudinal muscle-fibres of the various regions of the stomo-deum (Text-fig. 1), and presumably they also serve the eel-

MUSCLES OF THE BEE 571
lular layers, but I have never seen them uniting therewith.A. Doyere's hillock is not present where nerve and muscle fuse,but the nerve becomes a little expanded in its course, the neuri-lemma passes into the sarcolemma and a strand of nerve sub-stance enters the muscle substance, but the main nerve passeson. I have not detected any sensory nerve-endings on muscle-fibres for this system unless they are similar to those describedabove which are presumed to be motor-endings. The stomato-gastric nervous system presumably receives sensory impulsesfrom the other layers of the stomodeum, but I have not seenany definite signs of contact with the epithelium.
The last abdominal ganglion in all castes sends pairedsplanchnic nerves to innervate the muscles of ventriculus, smallintestine, and rectum. The nerves branch very considerablyand finally anastomose between the muscle-fibres of each of theregions. I cannot distinguish if the anastomoses of one regionpass directly on to another region, but this does not seem tobe the case. The nerve-twigs become thoroughly entangledwith the muscle-fibres and often lie on the upper surface of thebasement membrane, but I have never seen them entering this. •They are often only 3 ̂ wide with ganglion-like swellings wherethey branch or anastomose. They end in a muscle-fibre in asmall Doyere's hillock occupying the sides of only one to twosarcomeres, or, more seldom, may end as in the stomatogastricsystem, i. e. a small lateral branch of nerve substance fuses witha muscle-fibre. Nuclei are usually present at Doyere's hillockand at the ganglionic swellings, and since they resemble thenuclei of nerves supplying somatic muscles, I regard them asbelonging to the neurilemma and the ganglion-like swellingsas probable trophic centres. Only these two types of nerve-endings were seen and both appear motor. If a sensory cellwere present at the end of a nerve, it would be very difficult todetect against the various cells of the alimentary tract. Twotypes of motor nerve-endings were found on the gut of the larvaof O r y c t e s n a s i c o r n i s by Orlov (1924).
The stomatogastric nervous system is well developed in allcastes, and considering its complexity and that it is the only

572 GUY D. MORISON
part of the nervous system serving the stomodeal region, it mustbe of great importance, especially to the worker bee. The cellsof its ganglia suggest that it would be a motor centre for thefore-gut, and certainly the pharynx and proventiculus are veryactive regions. Its sensory function is problematical. Thesystem deserves experimental study and detailed investigation ;likewise the innervation of the rest of the gut. It seems strangethat the muscles of the mesenteron and the proctodeum shouldboth be innervated exclusively by the last abdominal ganglioneven though the complex evolutionary changes of the gangliaand the muscles are borne in mind. The last abdominal gan-glion presumably controls the movements of the ventriculus,small intestine, and rectum, though the muscles of each of theseregions will exhibit slight movement after they are parted fromthe ganglion.
All the regions of the alimentary canal are well supplied withmuscles, but they are not all regions of equal muscular activity,though they would require a supply of O3 and a means of gettingrid of CO2. In this respect the correlation between tracheation,blood-stream, and active muscles is very interesting and is thesame for all three castes. The pharynx is the most muscularand perhaps the most active region in the gut, and its musclesare well supplied with tracheae and lie close to air-sacs, all ofwhich must be bathed by a constant flow of blood directly fromthe aorta. Those muscle-fibres which form bundles like somaticmuscles are provided like them with numerous tracheoles, whilstthe other more typical splanchnic fibres have very few tracheolesbut are better placed for deriving benefit from the blood-stream.The oesophagus is a region of comparatively little activity. Itsmuscles throughout its course in the thorax and propodeum arewithout tracheae, but it rests in a sluggish stream of bloodwhich presumably satisfies its respiratory needs. Soon after itenters the post-abdomen it expands into the crop, and just atthe start of this region it has a few short tracheae directed moreor less backwards. The crop is capable of great distension butis not a region of active muscles. If the normal type of smalltracheae were present on its median region they would be liable

MUSCLES OF THE BEE 573
to rupture. It is without tracheae but numerous large air-sacslie very close and often in contact with it. Presumably itsmuscles get their respiratory needs supplied by the blood, andmay be helped by the adjoining air-sacs. The proventriculusranks very near or equal with the pharynx in muscles andactivity. Its muscles are of the somatic type and have atracheation like them. These tracheae come forward to theproventriculus from the ventriculus. The tracheation of theentire stomodeal region belongs almost exclusively to themuscles, for the non-muscular layers are very delicate andseemingly are aerated by the blood-stream. In the ventriculus,on the contrary, both muscular and cellular parts require aplentiful supply of air, so this region is crowded with tracheae.Here the tracheation has received comment from variousobservers : vide Zander's (1922) photograph or Petersen's (1912)statements, though I have never seen the degree of anasto-mosing tracheoles the latter author describes. The ultimatetracheoles are 0-3/u. wide. They branch considerably and takea very winding course amongst the muscle-fibres on which theyvery rarely end. Their usual course is to pass from the outersurface of the sarcolemma of a muscle-fibre on to the basementmembrane of the ventriculus on which they end blindly, or,very rarely, anastomose with other tracheoles. The broadermuscle-fibres are quite well supplied with tracheoles, but thefinest fibres are often without them and so must depend uponthe blood or adjacent tissues for their respiration. Each Mal-pighian tube is very well supplied with tracheoles coming fromtracheae of various origin depending on the position the tubeoccupies in the body. The tracheoles have a tendency to twistround the tube in long but interrupted spirals. Often twotracheoles will run parallel to one another, and between themthere may lie a muscle-fibre which they demarcate and, at thesame time, obscure (Text-fig. 6). The small intestine is astrongly muscular region which has its muscles very well pro-vided with tracheoles. The tracheoles are often very long, andthough most pass along the outer surface of the muscle-fibressome will run between the fibres and the basement membrane,

574 GUY D. MORISON
therefore they may serve for the respiration of both muscle andepithelium. They usually end on the muscle-fibres, but'mayterminate on the basement membrane usually just above whereepithelial cells meet. The course of the tracheoles on all thesplanchnic muscle-fibres is less regularly longitudinal and showsless bending for the individual sarcomeres than on somaticfibres.
The rectum is capable of enormous and long-sustained disten-sion. It may be divided roughly into three parts dependingupon the degree of distension of which each is normally capable.The first part starts at the end of the small intestine and con-tinues to the end of the rectal glands ; it may be distended con-siderably, but not so much as the second part which is demar-cated from it by a greater bulge. The second part tapers to thethird part, which is a narrow tube leading to the anus. Thedistinction into first and second part may be most clearlymarked between the anterior end of the rectal glands and thestart of the rectum in the queen, but this need not concern usnow. The musculature is thickest at those parts which showthe least distension : most conspicuous on part three, less so onone, and least on two. This seems due to the distension ofthe rectum spacing the individual fibres farther apart, becausethe number of muscle-fibres in a transverse section of any of thethree parts is about the same. Part one has a poor supply oftracheae except at the rectal glands, where they are verynumerous, vide Trappmann (1923) ; part two, like the greaterpart of the crop, is without tracheae ; part three is well sup-plied. All the tracheoles are practically confined to the muscle-fibres except at the rectal glands. Part two, whether it bedistended or not, lies in such a position with regard to theventral diaphragm that it is bathed by the constant stream ofblood passing upwards to the heart. Perhaps this blood is notso well oxygenated as that which leaves the aorta in the head,but it must be sufficient for the respiration of the second partof the rectum.
The histology of the splanchnic muscle-fibres is characterizedby the fibres being more or less flattened against the gut around

MUSCLES OF THE BEE 575
which they anastomose in an irregular meshwork ; they areoften very long but their frequent anastomoses with one anothermake it impossible to specify their length, whilst their width isexceedingly variable ; nuclei are placed usually in the axis ofthe fibre; all the transverse discs described (Morison, 1927) forsomatic fibres may be present, but neither are they so sharplydenned nor are the longitudinal striae so obvious ; neverthelessfibres may sometimes be frayed into fibrils 0-4/x wide. A sarco-lemma apparently is always present, though often impossible todemonstrate in fibres 3/̂ . wide. The sarcolemma is attached tothe contractile substance of the muscle-fibre at the telophrag-mata, and a beaded appearance of the fibre is apparently quitenatural in many cases, especially when the muscle substanceoccupies the whole space of each bead without leaving a spacebetween itself and the sarcolemma. Tracheoles lie against thesarcolemma without piercing it. The attachment of the muscle-fibre to the alimentary canal varies : some of the musclesresemble somatic muscles in their origin and insertion, whilstothers depend upon anastomoses with fibres of their own andother layers for attachment to the gut. By means of theseanastomoses each layer forms a network tube more or less closelyapplied to the basement membrane of the epithelium of the gut,and should one layer lie on top of another, besides the fibresuniting the two layers, there will be places where the outer layerrests upon the basement membrane. The tracheae and nerveswhich twist amongst all the muscle-fibres would also help inbinding the muscles to the gut. I have said that the muscle-fibres rest on the basement membrane of the epithelium of thegut. Perhaps an endothelial membrane covers them and isitself contiguous "with the basement membrane, but I have notseen a definite endothelial membrane for any part of the alimen-tary canal excepting the Malpighian tubes, and on these it seemsto lie chiefly below the muscle-fibres. The presence or absenceof this membrane is ignored in writings on the alimentary canalof the bee. After muscles have been dissected away from therest of the gut, fibres are sometimes seen which have verydelicate threads attached to them at the place of rupture. It
NO. 284 P p

576 GUY D. MORISON
is impossible to determine where these threads belong, but thereis the possibility that they are fragments of the endothelialmembrane.
In modern writings about the internal anatomy of insects,this membrane is often wrongly called the ' peritoneal' mem-brane. As Professor Goodrich pointed out to me (in l i t t . ) ,insects have a haemocoele of which the lining should not beconfused with a peritoneum which is the lining of a portion ofa true coelome. As far as most of the muscles of the bee areconcerned, the only membrane between their contractile sub-stance and the cavity of the haemocoele is the sarcolemm.a ofthe individual muscle-fibres.
The more detailed structure of the pharynx considered as anentity seems to have been ignored in writings since it wasfigured and described by Wolff (1876), though Mclndoo (1916)figures many of the muscles. Wolff's figures show the musclesexcellently except that it seems to me that he has omitted apair of muscles which would be equivalent to another pair ofSchlundkopfofmer, dilators placed between his Schlundkopfoff-nern 5 and 6, as well as a pair of compressors, Schlundkopf-schliessern placed behind the extra pair of dilators. Thepharynx does not vary much in size for the caste—drone andworker are about equal and queen a little larger.
The epipharynx is controlled by a fan-shaped muscle,l e v a t o r ve l i p a l a t i n i , which is probably formed by thefusion of two muscles. It is attached to the anterior dorsal endof the pharynx and in the cavity of the epipharynx. There aresix pairs of m u s c u l i d i l a t o r e s p h a r y n g i s attachedbetween the dorso-lateral wall of the pharynx and the anteriorwall of the head, i. e. clypeus and base of frons. They passcephalad and laterad to attachment on the head, whilst theseventh pair of d i l a t o r s (Wolff's sixth) arise on either sideof the mid-line of the pharynx just posterior to the frontal gan-glion of the stomatogastric nervous system, and pass almostvertically upwards to attachment on the frons. The eighth pairof d i l a t o r s are placed behind the brain just where thepharynx passes into oesophagus and anterior to the stomato-

MUSCLES OF THE BEE 577
gastric pharyngeal ganglia. Each is very long and delicate,and consists of about nine fibres drawn from the longitudinalmuscle-fibres which run forwards to the pharynx or backwardsto the oesophagus. Their innervation from the recurrent nerveof the stomatogastric system is usually easily seen, whilst theopacity of the pharynx makes the innervation of other musclesa matter of difficulty to detect. The anterior dorsal lip of thepharynx is a tough plate on which lie two fan-shaped musclesconstituting the pair of musculi compressores pharyn-gis p r i m i. The muscle-fibres radiate outwards passing overthe edge of the pharynx, and besides these there are other fibreswhich cross the entire pharynx transversely and are attached toits sides. Pharyngeal compressors 2-5 are clearlypaired muscles which start near the mid-dorsal line of thepharynx and run laterad and cephalad to terminate on the sidesof the pharynx. Compressor 2 passes between dilators 2 and 3,compressor 3 between dilators 3 and 4, and so on for the others.Each posterior lateral arm of the chitinous pharyngeal platebears a dorsal, "anteriorly directed hook which gives insertionto two muscles on either side of the pharynx. A musculuspro t rac to r pharyngis originates from a tentorial bar nearits anterior union with the cranium and passes caudad andmesiad to insertion by a tendon on the hook of the arm of thepharyngeal plate. Inserted opposite this muscle and antagoniz-ing its pull is the r e t r ac to r pharyngis superior, whichoriginates from the front of the head. The re t rac to rpharyngis inferior of Wolff is a small pair of muscles.Each muscle adjoins its pair from the opposite side and isattached on one side of the mid-ventral line of the pharynxbehind the pharyngeal plate. It passes caudad to attachmenton either side of the mid-ventral line of the pharynx moreposteriorly. Besides the muscles described above the pharynxpossesses a great number of muscle-fibres which are not groupedtogether in such definite muscular units. On the side of thepharynx a group of strong muscle-fibres attached to theventral edge of the lateral arm of the pharyngeal plate, andpassing cephalad and ventrad to near the mid-ventral line of
p p 2

578 GUY D. MORISON
the pharynx, may be called a l a t e r a l c o m p r e s s o r .Attached to the side of the pharynx just caudad to the lateralcompressor is a similar group of fibres which pass caudad toinsertion along the mid-ventral line of the pharynx, and to theposterior bar of the tentorium on either side. This may becalled the l a t e r a l d i l a t o r . On the dorsal surface of thepharynx a number of muscle-fibres are gathered into cordswhich pass from one side to the other. The arrangement of thecords is not constant and they often overlap the bases of thedorsal compressors. Under all the rest of the muscle-fibreson the anterior dorsal surface of the pharynx there are a fewlongitudinal fibres which do not pass into any of the moredefinite muscles. Towards the posterior part of the pharynxthey become more visible and later they form the (inner) longi-tudinal muscle-fibres of the oesophagus. Behind the insertionof the superior pharyngeal retractors, definite circular (trans-verse) fibres appear. At first they radiate cephalad and ventradcovering the lateral dilators and are attached to the pharynxnear the mid-ventral line, but soon they become more typicallycircular till they are quite indistinguishable from the outer coatof circular muscle-fibres of the oesophagus.
The fibres of muscles of the pharynx which are attached tothe chitinous integument are like ordinary somatic muscle-fibres in histology. The fibres or parts of fibres which lie onthe wall of the pharynx have the general characters of splanchnicfibres. The width of the fibres is fairly constant for the threecastes : 10-15/x in the somatic type of muscle, 10-15/x in thesplanchnic type of fibres. Length of the sarcomeres in restingsomatic fibres average 5/x, in splanchnic fibres 5/x. Nuclei arepresent in the axes of the fibres but difficult to distinguish.
The oesophagus bears a thick covering of muscles which pre-sumably allows a peristaltic movement to pass backwards orforwards as desired at least in the worker bee. Possibly thecontraction of the post-abdominal walls will help in driving thecontents of the crop back to the mouth. An inner, single layerof longitudinal fibres lie close together applied to the basementmembrane of the epithelium of the oesophagus. It consists

MUSCLES OF THE BEE 579
of narrower fibres than the outer circular layer. The fibresare 4-13 \L wide, sarcomeres average 3/x in length, nuclei average7-5 :4-5:4-5/x. Anastomoses of these fibres are very difficultto detect but occur occasionally, and it is not unlikely thatthe circular and longitudinal layers are united by muscle-fibres. Some of the fibres are very long. The fibres of the outerlayer or circular muscles form more or less complete rings roundthe oesophagus. They are flattened fibres 7-22ft wide, sarco-meres average 3/^ in length, nuclei 7*5 : 4*5 :4-5/x. Lateralanastomoses between parallel fibres are quite frequent, and thefibres lie close together. The sizes of the fibres show the samevariation in the three castes.
Crop .—The two muscular layers of the oesophagus are con-tinued over the crop with wider spacing between the individualmuscle-fibres. The distension of the crop does not seem to altertheir histological structure to any marked degree. When thecrop does not contain food its walls tend to fall in folds with themuscle-fibres correspondingly bent and slack. As the cropbecomes distended with food, the fibres are drawn straighterand lose some of their slackness. It is not easy to see themuscle-fibres on a distended crop. The following method isrecommended though not satisfactory : starve some bees forsome hours before letting them feed on colourless, fairly thinsugar syrup. Chloroform them and remove the distended cropwith oesophagus and fore-part of ventriculus, which piece isthen mounted in a drop of the syrup that was fed to the bees,but now coloured with a stain. Exosmosis and opacity may beovercome sufficiently well in some of the specimens for theexamination of the coarser details of the muscles. Text-fig. 2depicts the typical histology of the muscle-fibres. The inner,longitudinal layer of fibres anastomoses more or less. They areflattened 4-14/x wide ; sarcomeres average 4/x in length ;nuclei 8:4:4-10: 6: 6/̂ containing one or more dark-stainingbodies. The nuclei usually lie in the axis of the fibre in acertain amount of undifferentiated sarcoplasm, but they mayoccur near the periphery of the fibres as the latter approach theventriculus. Some of the fibres of this layer pass directly on

580 GUY D. MOBISON
to the ventriculus. The outer, circular muscle-fibres anasto-mose more or less. They are 4-25ju. wide ; sarcomeres average3/x; nuclei 6:4:4-11:5:5/x, containing one or more dark-staining bodies. This layer of muscles stops at the end of thecrop.
For the arrangement and functioning of the muscles of theproventriculus, the work of Trappmann (1923) should be con-sulted. A bundle of inner longitudinal fibres is attached within
TEXT-FIG. 2.
•L m
Portion of muscles of the crop. Flemming's fixative, iron haema-toxylin, eosin. The nuclei are shown in optical section, s, sarco-lemma. Lettering as in Text-fig. 1.
each of the four lobes of the proventriculus. Enclosing thesefour bundles is a ring of circular fibres which are most con-centrated opposite the longitudinal bundles. In the anteriorpart of the proventriculus these fibres tend to. be attachedbetween the sides of each lobe, but caudad they remain morecircular and eventually pass on to the ventriculus as its coatof circular muscle-fibres. Ectad of the circular muscles but stillin the lobes of the proventriculus are a few outer longitudinalfibres which pass backwards on to the ventriculus and minglewith those coming from the crop. The longitudinal muscles
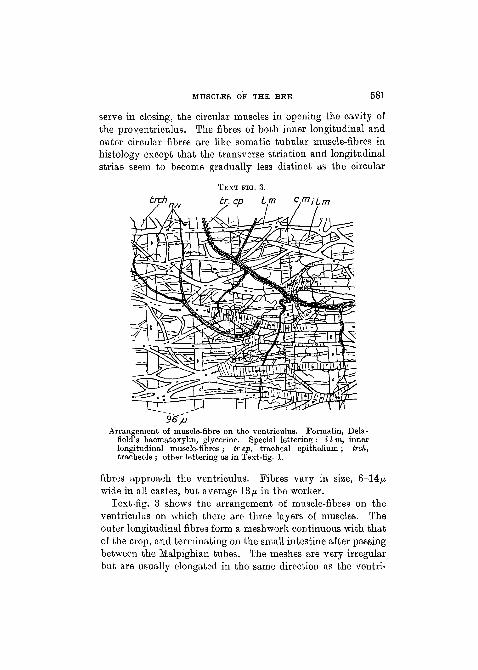
MUSCLES OF THE BEE 581
serve in closing, the circular muscles in opening the cavity ofthe proyentriculus. The fibres of both inner longitudinal andouter circular fibres are like somatic tubular muscle-fibres inhistology except that the transverse striation and longitudinalstriae seem to become gradually less distinct as the circular
trchTEXT-EIG. 3.
br. ep Lm c,miLm
36~/J
Arrangement of muscle-fibre on the ventriculus. Formalin, Dela-field's haematoxylin, glycerine. Special lettering : i I in, innerlongitudinal muscle-fibres ; tr ep, tracheal epithelium ; trch,tracheole ; other lettering as in Text-fig. 1.
fibres approach the ventriculus. Fibres vary in size, 6-14^wide in all castes, but average 13/A in the worker.
Text-fig. 3 shows the arrangement of muscle-fibres on theventriculus on which there are three layers of muscles. Theouter longitudinal fibres form a meshwork continuous with thatof the crop, and terminating on the small intestine after passingbetween the Malpighian tubes. The meshes are very irregularbut are usually elongated in the same direction as the ventri-

582 GUY D. MORISON
culus. Most of the fibres lie in a single layer, but a few fibrestwist above and below the others, so that in a transverse sectionof the ventriculus certain areas would show more than threelayers of muscle-fibres. Also some muscle-fibres pass from theouter longitudinal to the middle circular layer or to the innerlongitudinal layer, so that there is direct muscular continuitybetween all the muscle-fibres of the ventriculus. The largerlongitudinal fibres of the outer layer pass over the annular con-
TEXT-FIG. 4.
Dh
nuHistology of the longitudinal and circular muscle-fibres of the ventri-
culus. Flemming's fixative, iron haematoxylin, eosin. Nuclei ofthe longitudinal fibre are shown in optical section. Lettering asin Text-fig. 1, except D h, Doyere's hillock ; s, sarcolemma;z, telophragma.
strictions of the ventriculus, but some of the delicate fibres ofthe innermost layer dip down into the depressions before linkingup again with other fibres. The longitudinal fibres of the outer-most layer are flattened, very variable in width and varyconstantly during their course 3-35/x in all castes. In theanastomoses the sarcostyles may be seen passing as individualsfrom one fibre to another (Text-fig. 4). Nuclei varying some-what in size 5:4: 3-11 :2-5 : 2-5/u occur singly or in rows in theaxis of the fibres. Sometimes when a fibre is very wide throughthe anastomoses of several smaller fibres, the nuclei lie two

MUSCLES OF THE BEE 583
abreast in small groups. Nuclei were not observed in or on verydelicate fibres, and often they may not occur in a long portionof a wide fibre. They contain one or more dark-staining bodies.
In contrast to both the layers of longitudinal fibres the middlecircular (transverse) fibres are placed verj#regularly on theventriculus. Their width is very constant, average 13/* indrone, 15-2(V in queen and worker. They do not unite withone another directly. Their sarcolemma is often distendedwidely from the contractile sarcostyles in order to accommodateperipheral nuclei and undifferentiated sarcoplasm. The nucleiand the undifferentiated sarcoplasm usually lie on the fibre onthe side away from the gut-wall, and both may stain moreintensely than the rest of the ventriculus if the alimentary canalfrom a living bee is placed directly into dilute Delafi eld's haema-toxylin for some time. The nuclei usually lie peripherally tothe contractile sarcostylar substance, but they may occuraxially completely surrounded by sarcostyles or in a groovebetween the sarcostyles. Pavlovsky and Zarin (1922) havestated that the nuclei only occur peripherally lying in sarco-plasm (= undifferentiated sarcoplasm, mihi) , whilst the fibrilsare disposed in bundles as figured by them. The fibrils formbundles, as seen in transverse sections, where the fibres areabout to anastomose with other muscle-fibres, otherwise thefibrils are fairly evenly distributed in the fibre. The nucleiaverage 8:6:6/^. but may be 19:8:8^, with one nucleolus insmall and more in large nuclei. The peripheral nuclei tend tobe larger than the axial nuclei. Transverse striation and thelongitudinal striae are quite distinct, and the telophragmataoften may be seen to traverse the entire fibre, stretching wellbeyond the contractile elements.
The innermost layer of longitudinal fibres discovered byWhite (1918) in the examination of Nosema infected bees isusually most clearly seen in bees heavily infected with thatdisease, since the epithelial cells may be dissected away toleave a comparatively transparent picture of the muscles.However, the following details are taken from the study ofapparently healthy bees of the three castes. The layer seems

584 GUY D. MORISON
better developed in some bees than in others, but this may bea matter of staining technique. It starts on the fore-part ofthe ventriculus and ends apparently at the Malpighian tubes.Text-fig. 5 depicts the appearance of these fibres. As shownthey branch and a'nastomose very considerably and irregularlywith one another, besides having fibres passing to the middlecircular and outermost longitudinal layers. Fibres measure3-13 JU. in width with some more delicate strands of tissue passingfrom one fibre to another. Nuclei average 6:4'5:4-5/x. They
TEXT-FIG. 5.
nu
cm
Histology of the inner longitudinal muscle-fibres of the ventriculus.Formalin, Delafield's haematoxylin, glycerine. Delicate strandsof protoplasmic material (x) pass between the fibres. Lettering :c m, circular muscle-fibre ; nu, nucleus ; s, ? sarcolemma.
lie singly on the outer surface of the fibres in a small mass ofundifferentiated sarcoplasm which stains more intensely thanthe fibre. Each contains a nucleolus. The striation of the fibresis typical but not easy to distinguish. A sarcolemma seemspresent. The fibres are very much flattened against the base-ment membrane of the ventriculus with which they are con-fused in most sectioned material. Besides a view of sectionedmaterial is not always truthful, since it may show a fibre whichmore properly belongs to the outermost longitudinal layer, as

MUSCLES OF THE BEE 585
apparently belonging to the innermost layer, or it may showmore than three layers of fibres for the ventriculus. Deepstaining in haematoxylin followed by mounting in glycerine isthe best for showing the fibres. The muscles which pass to theMalpighian tubes seem best ascribed to this layer of muscles.
Drone, queen, and worker bees have each about 100 Mal-pighian tubes. Trappmann (1923) has described the appearanceof the muscle-fibres passing round the tubes of the worker.I should like to call attention to the previously describedtracheation of the tubes since it is of some help in localizing the
TEXT-FIG. 6.
27/J
'End of Malpighian tube showing muscle-fibres, trachea (tr), andtracheoles (Irch). Stained Ranvier's gold chloride method;mounted in glycerine.
muscle-fibres, which are usually exceedingly difficult to find.Glycerine mounts of the tubules are most satisfactory afterfixation in formalin followed by staining in aqueous solutions ;or Kanvier's gold chloride method may be used. I have notdetected any innervation of the Malpighian tubes or theirmuscle-fibres, and the only description I have consulted of theinnervation of the tubes in other insects is that of Leydig (1876)and Schindler (1878) for some larvae. The muscle-fibres arepresent on the Malpighian tubes of all castes, but I am not sureif they are present on all the tubes of an individual bee. I findthat the fibres vary from one to four per tube, around whichthey are twisted in a spiral which circles the tube about threetimes.. They measure 3-5 p in transverse diameter and areembedded in the endothelial lining besides being somewhat

586 GUY D. MORISON
flattened against the basement membrane of the Malpighiantubes. The sarcomeres (Text-fig. 7) are 4-5/A long. I have notseen nuclei associated with the fibres, which apparently mayanastomose on the tube (Text-fig. 6). The fibres seem to pass tothe tubes exclusively from the ventriculus and not from the smallintestine, but this may be faulty observation. Under favourableconditions the Malpighian tubes may be seen to perform slightmovements when the living gut of a bee is examined under themicroscope. The movements seem to be the result of contrac-tion of the muscle-fibres of the tubes and not due to the purelymechanical movements of tracheae and other parts. The func-
TEXT-FIG. 7.
Sarcomere of muscle-fibre from Malpighian tube. Formalin, Dela-field's haematoxylin, glycerine.
tion of the muscles would be to help in discharging the contentsof the tubes. Similar muscles have been described lately fora fly. It seems probable that the fibres are present on all theMalpighian tubes of a bee, but owing to technical difficultiesare not brought into view. If they are present only on sometubes, then these have either an advantage or disadvantage overtheir fellows, but all seem alike in structure otherwise.
The small intestine is coated with a single layer of muscle-fibres for the greater part of its length. The fibres are placedcircularly (transversely) except at the start of the intestine,where they are covered by some longitudinal fibres comingfrom the outermost muscle-layer of the ventriculus. Text-fig. 8shows the general aspect of the circular fibres which are charac-terized by being very broad, more or less flattened against theintestinal wall, and with the sarcolemma closely attached to the

MUSCLES OF THE BEE 587
contractile fibrils. They lie very close to one another and some-times they fuse for the distance of a few sarcomeres, or narrowstrands of muscle connect them together so that all the fibreson the small intestine are in muscular continuity with eachother. The circular fibres (Text-fig. 9) are 18-50/u. wide, whilstthe fibres connecting them together are 4-10/u. wide. Sarco-meres average 6/x in length and the muscle does not show very
TEXT-FIG. 8.
trch
Arrangement of muscle-fibres on the small intestine with tracheoles(trch) and nerve (nv) ending at (nv e). At (x) is shown an apparentfusion of the sarcoplasm with the intestinal epithelium. StainedRanvier's gold chloride method, mounted in glycerine.
strong waves of contraction ; telophragmata seem true mem-branes ; individual sarcostyles are seen quite easily at the endsof broken fibres ; each sarcostyle is about 0-3/a wide. Small orlarge nuclei lie scattered irregularly in the axes of the fibres.They are more or less surrounded by undifferentiated sarco-plasm. Small nuclei are irregular spheroids or ellipsoids 3-6/xin longest diameter with usually a single nucleolus. Largenuclei are rarer than the small. They are more or less ellipsoidal,and very variable in size and the number of nucleoli they

588 GUY D. MORIS ON
contain, e.g. 24:10:10/x with two nucleoli and 32:8:8/x witheight nucleoli. Presumably the large nuclei result from the non-separation of numerous small nuclei in the developing pupa.Small nuclei occur in groups, whilst large nuclei occur alone. Thecircular muscles of the small intestine start just behind theMalpighian tubes, and when the small intestine expands intothe rectum they anastomose with one another more frequently,becoming narrower and gradually transforming into the innercircular layer of muscle-fibres of the rectum.
TEXT-FIG. 9.
• 32M; 'Histology of muscle-fibre from small intestine. A portion of nerve
(nv) adjoins the fibre, in which two types of nuclei (nu) are shownin optical section. Flemming's fixative, iron haematoxylin, eosin.
Where the small intestine expands into the rectum a layerof outer longitudinal muscle-fibres appears. There are no freeends to these fibres, for they all anastomose with one anotherwithout leaving a clearly defined trace on the gut to show wherethey started as a meshwork. The meshwork continues to theend of the alimentary canal. Below it lies the meshwork oftransverse or circular fibres. The rectum possesses only thesetwo layers of muscle-fibres (Text-fig. 10). The fibres of bothlayers are more or less flattened against it and resemble oneanother especially towards its middle, which region is subjected
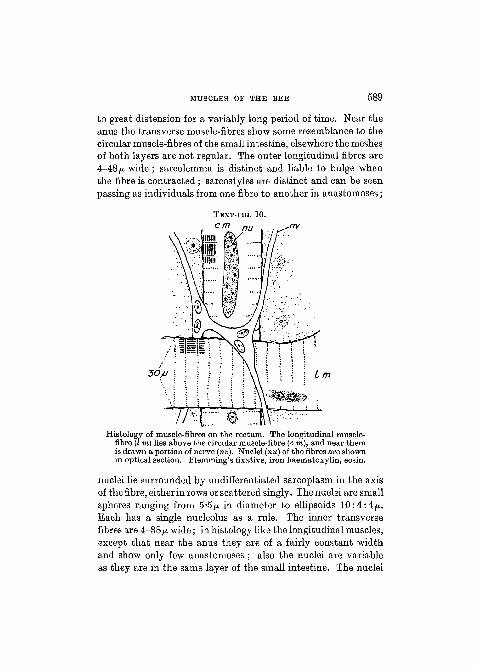
MUSCLES OF THE BEE 589
to great distension for a variably long period of time. Near theanus the transverse muscle-fibres show some resemblance to thecircular muscle-fibres of the small intestine, elsewhere the meshesof both layers are not regular. The outer longitudinal fibres are4-48 fj. wide; sarcolemma is distinct and liable to bulge whenthe fibre is contracted ; sarcostyles are distinct and can be seenpassing as individuals from one fibre to another in anastomoses;
TEXT-FIG. 10.
nu
30/J
Histology of muscle-fibres on the rectum. The longitudinal muscle-fibre (I m) lies above the circular muscle-fibre (c m), and near themis drawn a portion of nerve (nv). Nuclei (nu) of the fibres are shownin optical section. Flemming's fixative, iron haematoxylin, eosin.
nuclei lie surrounded by undifferentiated sarcoplasm in the axisof the fibre, either in rows or scattered singly. The nuclei are smallspheres ranging from 5*5p. in diameter to ellipsoids 10:4:4ft.Each has a single nucleolus as a rule. The inner transversefibres are 4—38/x wide; in histology like the longitudinal muscles,except that near the anus they are of a fairly constant widthand show only few anastomoses ; also the nuclei are variableas they are in the same layer of the small intestine. The nuclei

590 GUY D. MORISON
are arranged as in the longitudinal muscles, but near the rectalglands especially and near the anus, they may be very large withseveral nucleoli, e.g. 18:8:S/x or 48:7:7fi. A few muscle-fibres unite the two meshworks and may be assigned to eitherlayer. The transverse fibres lie directly on the wall of therectum, and the longitudinal fibres after crossing them oftendip down to rest on the rectal wall.
The Ca rd i ac and Dor sa l D i a p h r a g m Musc les .
The heart of the bee is a colourless tube lying in the middleline of the body above the alimentary canal and not far belowthe dorsal surface of the insect. It extends from below post-abdominal tergum 5 to the top of the pharynx in front of thebrain. Towards the posterior end it is expanded into fivechambers, each of which is preceded by a pair of valves—achamber and a pair of valves corresponding to each of the post-abdominal segments 1—5. Each of the five chambers is more orless enlarged behind and constricted in front, and is somewhatflattened dorso-ventrally. For convenience the chambers andvalves will be numbered from behind forwards. Number 1, thelast chamber, is closed at the posterior end. There are five pairsof ostia. A pair of ostia leads into each of chambers 2-5, andthe most anterior pair opens into the heart in front of the fivechambers ; otherwise the pulsatory vessel is a closed tube tillit opens below the brain. The position and structure of the ostiaand associated valves needs some explanation : a typicalchamber is a tube expanded caudad and tapering cephalad.A short length of the sides of this tube fits into the expandedportion of the preceding chamber, but the dorsal and ventralwalls do not fit in, since the dorsal walls of all the chambers areunited in a common plane on the upper surface of the heart,whilst the ventral walls are joined in the same way on the lowersurface. Each side of the expanded portion of the precedingchamber bends inwards and forwards to form a flap lyingparallel to the enclosed sides of the posterior chamber. Eachostium of a pair is a narrow passage which opens vertically intothe posterior region of the chamber it serves. It is the space

MUSCLES OF THE BEE 591
lying on one side of the heart between the infolded ends of thechamber it serves and the sides of the posterior chamber.Blood enters the heart through it and is pushed forwards bythe pulsatory activity of the heart. The two flaps of tissuewhich demarcate an ostium on one side of the heart arrest abackward flow of blood through the ostium, and together withthose from the other side of the heart they form a valve allowingthe blood to flow only forwards through the anterior chambers.The valves 2-4 are best developed, 5 is chiefly membranous, and1 is of such a structure that it would appear to allow bloodentering the ostia to fill chambers 1 and 2.
Muscles are confined to the walls of the chambers and to theposterior part of the tube (aorta) which precedes the chambers.The muscular region constitutes the heart proper, and is theseat of the propulsion of blood. The aorta loses its muscularcovering before it leaves the post-abdomen, and it becomesreduced to a chitinous tube without epithelium and of practicallyuniform calibre till it ends in the head. It takes about eighteensharp-angled bends when near and in the petiole, but it becomesstraight in the proctodeum and passes below the middle phragmaof the thorax and forms a dorsally directed loop between thelongitudinal indirect muscles of the wings. The other end ofthe loop rests on the oesophagus in the prothorax and head. Theheart shows the same structure in all the castes except for theexpected variation in size ; also the chambers of the heart varyin comparative size in individual bees.
The so-called ' alary ' muscles form the muscular part of thed o r s a l d i a p h r a g m which serves indirectly in the suspensionof the heart and is connected with the circulation of blood.There are five pairs of alary muscles corresponding roughly tothe five chambers of the heart, but extending beyond thechambers in front of and behind the heart proper. Each alarymuscle consists of a single layer of fibres radiating like the ribsof a fan from near the antero-lateral margin of post-abdominalsegments 2-6 to near the lower surface of the heart. Here thefibres tend to branch, and finally they all pass into a very delicatenetwork of membranous threads forming a part of a complex
NO. 284 Q q

592 GUY D. MORISON
reticulum with threads from the opposite and all other alary-muscles. The reticulum below the heart is so dense that itprobably functions like a true membrane. Some of the threadsof the meshwork are united to the muscles of the heart, whilstsimilar threads fasten the heart dorsally to the body-wall andare to be seen passing to various glandular cells of the region.The alary muscle-fibres rest upon a delicate membrane (peri-cardial membrane), so that the dorsal diaphragm forms a sheetof tissue enclosing the heart from the rest of the body exceptfor certain passages through which the blood enters. Thesepassages are the spaces lying between the dorsal diaphragmand its points of attachment to the body-wall. They are widestin the posterior segments.
Poletajewa (1886) has described the heart of B o m b u s ,which is very similar to that of the bee ; Pissarew (1898) figuresand describes the heart of the bee, but has overlooked one pairof alary muscles and a chamber of the heart; Arnhart (1906)offers an explanation for the bends in the aorta, and speaksabout systole and diastole without entering into details; Graber(1873) gives a detailed histological account of the entire pulsatoryapparatus of the bee, whilst he mentions the physiology morebriefly ; Girdwoyn (1876) describes and figures five chambersand five pairs of alary muscles, but states that there are fivepairs of ostia in the middle of the chambers, and he was ledastray by Blanchard's idea of the peritracheal circulation of theblood. Zander (1922) describes the heart accurately enoughexcept that he places the ostia in the middle of the chambersand endows the heart with valves for each ostium as well asfive valves marking the anterior boundary of the chambers.Snodgrass (1925) notices the five pairs of alary muscles, but hehas missed the most anterior valve of the heart. He is mistakenin stating that the ostia are in the middle of the chambers andthat there are no special valves between the heart chambers.He deals with the histology briefly.
The innervation of the heart and dorsal diaphragm is com-pletely ignored in all the writings I have consulted, exceptthat of Girdwoyn (1876) in which he says that the heart and

MUSCLES OF THE BEE 593
respiratory apparatus is innervated from ganglia at the side ofthe brain. Unfortunately he does not give any proof for thisstatement. I have not found any nerves supplying the heartitself. A large nerve supplies each alary muscle. The nerve-endings are very easily seen, but I have not traced all the largenerves to their source. Apparently they arise as paired nervesfrom each of the five post-abdominal ganglia of the worker andthe four post-abdominal ganglia of the drone and queen—thelast abdominal ganglion of the drone and queen supplyingthe nerves for the last two pairs of alary muscles. I have seenthe nerves passing from the last abdominal ganglion (of allcastes) to the alary muscles. The other large nerves supplyingthe alary muscles were not traced to their source, but they weretoo large to be branches of the pair coming from the lastabdominal ganglion. The nerves have the same histologicalstructure as those supplying somatic muscles. The endingsform Doyere's hillocks applied to the sides of three to five sarco-meres, and contain one to three nuclei averaging in size7:5:5/x (Text-fig. 11). A distinction into motor and sensorynerves was not observed. In order to find the innervation ofthe heart and alary muscles about sixty hearts of the threecastes were stained by various methods including unsuccessfulattempts at i n t r a - v i t a m staining with methylene blue. Notrace of a nerve was seen on any of the hearts, and it was con-cluded that the very capricious silver-nitrate method mightyield the final solution of the innervation of the heart.
All the muscle-fibres of the heart and alary muscles are wellsupplied with tracheoles (reaching a width 0-3/x) which evenextend on to the aorta until it passes between the indirect musclesof the wings. Though the heart is more or less bathed in blood,which perhaps could supply its respiratory needs without thehelp of the tracheae, it seems better that tracheae should supplythe muscles and even perhaps play the dual role of oxygenatingmuscle and the surrounding blood. Graber attributes to thedorsal diaphragm with its numerous tracheae the function ofgaseous exchange for the blood-stream. The evidence that theblood may transport and surrender oxygen to the tissues
Q q 2

594 GUY D. MORISON
depends on (1) the entire tracheation of the bee; (2) Griffiths(1891, 1892), after chemical analysis of the blood of four speciesof Lepidoptera, stated that in them it appears that oxygen istransported to a considerable extent by means of colourlessproteins in the blood; (3) Keilin (1925, 1926) has found therespiratory protein cytochrome in the tissues of the bee, andprobably it will be found in the blood when this is examined bya suitable technique.
TEXT-FIG. 11.
nv nu
Dh
Histology and innervation of alary muscle-fibres. The nerve ends ina Doyere's hillock (D h) in which lie some nuclei (nv nu). Twotypes of nuclei (nu) are shown in optical section associated with amuscle-fibre. Formalin, methylene blue, glycerine.
According to Graber (1873) the wall of the heart consists ofthree layers : intima, muscularis, and adventitia. My observa-tions on the histology agree with his except for various details.The intima is the non-cellular endocardial membrane which isindistinguishable from the sarcolemma of the circular musclesof the heart. It is best seen in the aorta. The muscular layerencircles the intima almost completely. It consists of semi-circular fibres placed at the sides of the heart, and of some almost

MUSCLES OP THE BEE 595
complete rings on the most posterior chamber. Since the fibresare not continuous over the dorsal and ventral surfaces of theheart, a median dorsal and a median ventral line of intima isobservable. This condition is a relict of the development of theheart. The dorsal and ventral lines are visible throughout thelength of the heart, but they are most conspicuous in front,and pass on to the aorta to be lost where the muscles disappear.The muscle-fibres lie in a single layer, flattened against the sidesof the heart. They are thick and clearly defined in width onall the chambers, but shortly from the start of the aorta theybecome very attenuated and ill-defined in width till they finallydisappear. On the chambers they measure 8-60/* in width,
TEXT-FIG. 12.
it
Histology of a portion of a muscle-fibre of the heart. Flemming'sfixative, iron haematoxylin, eosin. Lettering : z, telophragma.
8-20 JU, in thickness ; on the aorta 20-170 p width, 0-5-6/Athickness. A sarcolemma is present but often very difficult tosee. The transverse striation is very close (1-8-2-3/a betweentelophragmata) and often proceeds in a very zigzag directionacross the fibre. Individual sarcostyles (0-4/x wide) are oftendistinguishable (Text-fig. 12), and sometimes some appear notto lie parallel with the rest but to twist in amongst them—thebend being initiated at a telophragma. The darkly staining sub-stance of the sarcostyles tends to be most conspicuous at thetelophragmata, which are always distinguishable. Nuclei occurin the thicker fibres. They lie singly or in rows in spaces of un-differentiated sarcoplasm between the sarcostyles, either nearerthe centre or periphery of the fibre. The spaces containingnuclei may be scattered or arranged in more or less parallel rows

596 GUY D. MORISON
in the length of the fibre. Each nucleus usually contains a singledarkly stainable body. The nuclei vary slightly in shape andsize but average about 5^ diameter. As Snodgrass remarks, themuscles in longitudinal sections of the heart tend to look likean epithelium owing to the delicacy and the direction of thestriation.
According to Graber (1873) the adventitia or third layer ofthe heart in the Apidae ( sens . 1 a t.) is united with the peri-cardial septum or the membranous layer of the dorsal diaphragmthrough a characteristic network of entangled strands whichunite two similar membranes. I have not been able to assigndefinitely nuclei to either membrane. The adventitia seemsindistinguishable from the sarcolemma of the muscles of theheart, whilst the membranous layer (? two layers) of the dorsaldiaphragm is a definite, slightly fenestrated membrane, andbetween the two a number of exceedingly delicate fibres pass.
Closely associated with the heart in the propulsion of bloodare the dorsal and ventral diaphragms with their muscles.The alary muscles of the dorsal diaphragm have received men-tion above. In them the striation is as described for somaticmuscles, but is usually exceptionally distinct (Text-fig. 11).Tracheation is the same as in somatic muscles. The muscle-fibres appear to lie dorsad to the pericardial septum, which, asGraber said, is never in close union with the ventral surface ofthe heart. They radiate from their point of attachment to thechitin of the body-wall, and pass towards the heart as straight,somewhat dorso-ventrally flattened fibres. When nearing themiddle line of the body under the heart, they may terminatequite abruptly in connective-tissue-like strands, or they branchand anastomose with one another "of the same side of the body.At any rate all the alary muscle-fibres of one side of the bodyfinally branch into exceedingly delicate strands which form acomplex anastomosis under the heart with themselves and withthose of the muscles of the other side of the body. Amongst thisanastomosis of thread-like strands is applied the membrane ofthe pericardial septum. The strands show the xanthoproteicreaction as first noted by Graber.

MUSCLES OP THE BEE 597
The width of the alary muscle-fibres before they branch isvery constant, 15-20/z. Transverse striation may be seen clearlyin branches only 3ju. wide, and it certainly seems that theindividual sarcostyles pass gradually into the connective-tissue-like strands, but the strands show no trace of transverse stria-tion. The sarcostyles are 0-4-0-5 jn wide ; the strands even finer.
Nuclei are present in the axes of some of the alary muscle-fibres as is normal for tubular muscle-fibres. They lie singly orin rows and are surrounded by a certain amount of undif-ferentiated sarcoplasm. They usually possess a nucleolus andare small 4:2-5 :2-5/x. Besides these nuclei there occur somemuch larger ones at the sides of the fibres (Text-fig. 11). Itappears that the larger nuclei lie under the sarcolemma of themuscle-fibres, but I am not certain on this point after examiningboth sectioned material and whole mounts. Perhaps thesenuclei belong to the pericardial septum? They measure on anaverage 8:4:4/i.
Muscles of t h e V e n t r a l D i a p h r a g m .There does not seem to be a complete and accurate account
of the ventral diaphragm of the bee in any one book. In 1876Wolff figured and described the anterior end of this membranequite accurately, but his work seems to have been overlookedby later authors. The ventral diaphragm of the worker bee isa sheet of muscle-fibres lying horizontally between the nerve-cord and the alimentary canal in the propodeum and post-abdomen. Wolff, Graber (1873, 1876), and others have writtenabout its physiological importance. The cavity of the post-abdomen is divided into three more or less complete horizontalchambers by the dorsal and ventral diaphragms. The dorsaldiaphragm forms the floor of the chamber containing the heart;the ventral diaphragm forms the roof of the chamber containingthe nerve-cord ; and between these two lies the chamber con-taining the alimentary canal and reproductive organs. By thecontraction of the alary muscles the dorsal diaphragm is sup-posed to press down the visceral organs and so force blood intothe uppermost chamber chiefly at its posterior end. Now the

598 GUY D. MORISON
blood is supposed to receive an additional impetus from thepostero-anterior movements of the dorsal diaphragm. Much ofit is sucked or passes into the heart and is pumped out ofthe post-abdomen into the head by means of a closed vessel, theaorta. The aorta opens and ends above the pharynx. In thehead (Janet, 1911) and in the legs and mouth-parts there arechitinous vessels not connected directly with the central pulsa-tory apparatus, but presumably serving in the conveyance ofblood, though their method of functioning does not seem tohave been investigated. At any rate they have no specialmuscles, but probably rely on the anatomy of the bee and theforce exerted by adjoining somatic muscles for a passage ofblood through their tube. Physical and chemical laws and thestructure of the bee coupled with its muscular activity largelydetermine the course of blood, but it becomes very complicatedto explain the probable course of blood to and from the thoracicmuscles, though a thorough knowledge of the entire anatomymakes the problem seem fairly easy to solve. Excepting theheart and aorta there are no blood-vessels in the thorax, pro-podeum, and post-abdomen of the bee, but most of the bloodon its way back to the heart from the head, thorax, and legshas to flow into the lowermost chamber of the post-abdomen,and this chamber starts in the propodeum and is roofed by theventral diaphragm. After blood has entered the lowermostchamber its backward flow is aided by the waves of contractionpassing backwards along the ventral diaphragm. Some bloodcan leave or enter the lowermost chamber from the middlechamber by means of the spaces which occur between the body-wall and the edges of the ventral diaphragm, but most of theblood will be forced towards the posterior end of the lowermostchamber and thence into the middle chamber, and so back to theheart. As Snodgrass states, though there are no true blood-vessels excepting the heart and aorta in the abdomen, yet theblood has to flow along certain quite definite passages. Thetracheation of the muscles of the alimentary canal with regardto the flow of blood has been described already.
The anterior end of the ventral diaphragm is attached to the

MUSCLES OP THE BEE 599
posterior margin of furca 2-3 above the second thoracic nerveganglion. The diaphragm passes back through the propodeumas an ever-widening sheet of muscle-fibres which has only twoother points of attachment—one on either side to the tendonof either musculus propodei retractor dorsalis post-abdominisnear its insertion. The contractions of this pair of muscles isbound to affect the diaphragm, and there is probably some rela-tion to the constriction of the petiole. The diaphragm formsa complete sheet right across the petiole, to the posterior wallsof which it is attached. It lies in the same way in the secondabdominal segment (1st post-abd. seg.), being attached to thelateral ridges of the second urosternum. The attachment is con-tinued backwards for some distance along the third urosternum,and then a space occurs on either side between the diaphragmand the body-wall, and then the next points of attachment arethe anterior processes of the fourth urosternum. It is attachedin the same way to the anterior processes of the fifth, sixth,and seventh urosterna, and there are a corresponding pair ofgaps between it and the body-wall for each of the segments 4-6.Through these gaps pass muscles, the main nerves supplyingthe more dorsal somatic muscles, air-sacs, and tracheae, and apart of the blood. After its attachment to the anterior processesof the seventh urosternum, the diaphragm becomes very muchnarrower and then forked into two very long prongs which areattached to the a n t e r i o r p rocesses of the e i g h t ha b d o m i n a l s p i r a c u l a r p l a t e s . The plate is regarded asa part of the tergum of the eighth abdominal segment, andhitherto, in the post-abdomen, the attachments of the ventraldiaphragm have been to the sterna. The angle of the fork whichis the posterior margin of the diaphragm lies above and a littleanterior to the last abdominal ganglion of the worker and queen.The prongs of the fork are muscular for about half their lengthand then become a delicate but strong fibre which is attached tothe spiracular plate. Worker and queen have the type of ventraldiaphragm described above. The diaphragm in the drone issimilar except that it extends only into the fifth abdominalsegment, and, there being no fork, its most posterior termina-

600 GUY D. MORISON
tions are a little behind the anterior processes of the fifthsternum. However, the posterior margin of the ventral dia-phragm of the drone lies above and in front of the last abdominalganglion as in the other two castes.
The tracheation of the ventral diaphragm is comparativelysparse, consisting largely of tracheoles coming from air-sacs.Perhaps it serves as indirect evidence that the blood may supplythe respiratory needs of some muscles, i.e. that the blood cancarry O2 to the muscle and bear away CO2, since the ventraldiaphragm is washed continuously by blood.
Wolff described the innervation of the ventral diaphragm ascoming from the second thoracic ganglion. Other writers donot mention its innervation, except that Graber (1876) agreeswith Gegenbaur that in insects the pulsating blood-sinus iscontrolled by the central nervous system. Considering thedirection of the movement of the ventral diaphragm, Wolff'sexplanation is very plausible. It seems to me that the ventraldiaphragm is innervated from the second thoracic and the firstabdominal ganglion in all three castes, but I am not decided onthis point. The entire ventral diaphragm can be dissected outof a bee, and in it will be seen very few nerves and nerve-endings.The nerves are like those of the small intestine—nerve-fibresramifying amongst muscle-fibres.
In general the muscle-fibres of the ventral diaphragm forma layer of the thickness of one fibre, but as they anastomosevery considerably the thickness may often consist of two tothree fibres. Most of the fibres lie transversely in the diaphragm,and they are applied very closely to one another except wherethe diaphragm presents a fenestrated appearance. This happensat its start in the propodeum and just before it ends in the post-abdomen, also, more irregularly, near its lateral margins. Thelength of the fibres is very variable owing to their anastomoses,but single fibres do not stretch right across the abdomen. Thedegree of anastomosis is variable for different parts of thediaphragm. A portion of the diaphragm has a very charac-teristic appearance : a tissue of closely applied, anastomosingmuscle-fibres with close and rather obscure striation and few

MUSCLES OF THE BEE 601
or no tracheoles. The fibres (Text-fig. 13) are 5-18//. wide,average 8 /* ; somewhat dorso-ventrally flattened ; with a sarco-lemma ; striation as described for tubular muscles and like thatof the circular muscles of the small intestine. Strong bands ofcontraction were never observed, but a killed and a fixed beeoften has the post-abdominal part of its ventral diaphragmthrown into one or more transverse folds. Nuclei lie in or nearthe axis of the fibre in a greater or less quantity of undifferen-tiated sarcoplasm. They average 10:3:3/x with two to three
TEXT-FIG. 13.
,trch
Histology of muscle-fibres of the ventral diaphragm. Nuclei (nu)of muscle-fibres are shown in optical section. Formalin, Delafield'shaematoxylin, glycerine. Lettering : gang, ganglion ; nv, nerve ;tr, trachea ; trch, tracheole.
dark-staining bodies, but may be 58:3-6 :3-6JU,, containing manydark-staining bodies. No endothelial membrane is found withthe ventral diaphragm.
All the muscle-fibres of the heart, dorsal, and ventral dia-phragms are quite colourless when alive. If a living bee or anewly narcotized bee is dissected in a suitable manner the move-ment of the heart, the dorsal, and ventral diaphragms maybecome visible. Snodgrass has described how a coloured solu-tion when injected into the blood-stream may be seen to com-plete the course of circulation. In spite of many attempts I have

602 GUY D. MOEISON
not been successful in repeating this experiment, nor in isolatinga living active heart on a microscope slide. Graber (1873) sawa heart beating for a quarter of an hour in iodine water after itsremoval from the body.
Nothing is known about the normal rate of heart-beat orthe rate of the antero-posterior pulsatory waves of the ventraldiaphragm, or that of the movements of the dorsal diaphragm.The three regions are capable of independent contractions asmay be seen when they are exposed in a living bee, but since thefibres have inherent irritability and will contract on stimulation,no satisfactory conclusions can be drawn from observations ofthe contractions in an injured bee. Also it should be rememberedthat we have little definite knowledge whence most of thesemuscles derive their stimulus for contraction in the normal bee.It seems probable that many of the actions are 'reflex'. In allthree regions the movement is extremely rapid and apparentlyirregular like the ' breathing ' movements described previously(Morison, 1927). Probably the rate of circulation of the bloodis very closely connected with ' breathing' movements. Cer-tainly the telescoping and bending of the post-abdominal seg-ments must displace quantities of blood, for there does not seemto be much room to spare between the organs for the blood ina bee's post-abdomen, no matter what may be the state ofdistension of the alimentary canal and air-sacs.
The movements of the heart and dorsal and ventral dia-phragms must be co-ordinated in some way. This powerapparently is not localized in the brain or sub-oesophageal gan-glion, for the head can be removed and the three regions willcontinue their activity after a short pause. It seems probablethat the second thoracic ganglion would initiate the contractionof the ventral diaphragm, as well as affecting the respiratorymovements. The dorsal diaphragm is perhaps controlled by allthe abdominal ganglia and especially by the last. The innerva-tion of the heart is unknown. Perhaps the quantity of fluidsurrounding the heart supplies the stimulus for contraction.
The bee of all castes is a very difficult subject for physiologicalexperiments to determine the frequency of heart-beat or move-

MUSCLES OF THE BEE 603
ments of the ventral or dorsal diaphragms, and it is too special-ized an insect for the conclusions derived from the study ofother insects by Fries in 1926 and Carlson in 1906 to be appliedto it rightly. Temperature and light probably influence thefrequency of the movement of the three regions very consider-ably, as noted by Walling (1906) and Demoll (1926) for anumber of insects. During disease and perhaps with age themuscular activity of these organs is probably modified, but wecannot even hazard a guess to what extent.
THE MUSCLES OF THE REPRODUCTIVE ORGANS.
Muscles of the Male Reproductive Organs.
The chitinous parts of the male sexual organs have beendescribed by Zander (1900), whilst Koschewnikoff (1891) hadpreviously described without figuring the histology of theregions, and more recently Bishop (1920) has dealt with thewhole reproductive system very thoroughly and with specialregard to its functioning during the fertilization of the queen.The two latter authors describe the arrangement of the muscle-fibres, but the innervation and tracheation are not recorded, northe detailed histology of the muscle-fibres.
The last abdominal ganglion, the fourth, innervates the entiresexual apparatus, and the extrusion of the drone organs isusually ascribed to a reflex action. The nerves supplying themuscle-fibres end like those described for the stomato-gastricnervous system. Nerve-fibres anastomose amongst the muscle-fibres and their endings on the muscle-fibres are by very shortlateral branches. Doyere's hillocks and terminations of thenerve on other tissues were not seen, nor was a distinction intomotor and sensory nerves observed.
Tracheation follows the course normal to tubular muscles.Tracheae and tracheoles penetrate between the layers of muscle-fibres, and are most numerous where the fibres are most con-centrated.
The peculiarity of the muscle-fibres of the male reproductiveorgans is that a more or less delicate cellular endothelial mem-

604 GUY D. MORISON
brane envelops them, so that they are not freely exposed in thegeneral body-cavity like a l l the other systems of muscles seemto be in the three castes. Another characteristic of the fibres isthat they may be grouped together in fascicles which are notmuscles in themselves, but parts of a layer of fibres. Fasciclesare more distinctive of the longitudinal than of the circularlayers. They are not distinct from one another because fibrescross from one fascicle to another. Muscle-fibres are confinedto the mesodermal portion of the reproductive apparatus. Theyappear as an inner circular closely covered by an outer longi-tudinal layer of fibres on each vas deferens. These layers areat first very inconspicuous, but as the vas deferens expands intothe vesicula seminalis the individual fibres of each layer cometo rest closer together by the addition of new fibres, till eachlayer forms an almost complete envelope one to six fibres thick.The two layers are continued over the whole of the vesiculaseminalis, and similar layers cover the proximal parts of themucous glands with the addition of three innermost bands ofmuscle-fibres which, as Bishop remarks, are only a distortionin the arrangement of the circular layer. The muscle-layersterminate before the distal ends of the mucous glands. Muscle-fibres of various layers are numerous where the vesiculaeseminales are constricted to pass into the mucous glands and atthe junction of the ejaculatory duct with the mucous glands,but they do not pass on to the ejaculatory duct.
The muscle-fibres of the male reproductive organs twist inamongst one another and only rarely anastomose. They aresomewhat flattened against the tissue they cover, and aretypical tubular muscles, containing axial nuclei in rows orsingly embedded in a greater or less quantity of undifferentiatedsarcoplasm. But nuclei are scarce. A sarcolemma is present andthe striation shows the variation described for tubular muscles,and naturally, the genitalia have to be extruded before anyvery marked waves of contraction are discernible on the fibres.After fixation in alcohol the fibres may be broken into discs atthe telophragmata, and after fixation in alcohol followed by longsoaking in glycerine the fibres may be split into individual

MUSCLES OP THE BEE 605
sarcostyles. Sihler's (1895) method of macerating and stainingis also very successful in splitting these and all other tubularmuscle-fibres into sarcostyles.
The longitudinal (outer) muscle-fibres of the vesicula seminalisare 5-1 lju wide, with long sarcomeres 13-17/x, and nuclei12 : 2-4 : 2-4 — 8 : 2: 2 p. On the vas deferens, on the short tubeentering the mucous gland, and on the mucous gland they aresimilar except that on the two former regions the sarcomeresaverage 8/x in length.
The circular (inner) muscle-fibres of the vesiculae seminalesare 6-9 jx wide, with long sarcomeres 13-22/x and nuclei as above.On the vas deferens, on the short tube entering the mucous gland,and on the mucous gland, they are like the longitudinal fibres.
The three innermost bundles of muscle-fibres of the mucousgland are 8-21 //. wide, sarcomeres 6-13^. long, nuclei as above.
For details of the functioning of the muscles and the physio-logy of the reproductive organs the reader is referred to Bishop's(1920, 1920 a) valuable papers.
Muscles of t h e R e p r o d u c t i v e Organs of theQueen .
A very large and scattered literature exists on the reproduc-tive organs of the queen bee, nevertheless there does not seemto be a single account which deals with the entire morphologyof the system in a satisfactory manner. The muscles are almostalways totally neglected, and yet upon them depends theactions of oviposition which are repeated some hundreds ofthousands of times during the life of a prolific queen, and whichmay reoccur nearly 2,000 times a day during a honey-flow(Nolan, 1925). Besides, the muscles which are likely to be con-cerned directly with oviposition are also likely to play an im-portant part during the determination of the sex of the bee.Because, assuming that the sperm-pump regulates the outflowof sperm to the egg, the latter will have to lie in such a positionthat the sperm will be able to enter it, and whatever the positionmay be these muscles will be affected.
Lb'schel (1916) gives an account of the development of the

606 GUY D. MORISON
reproductive organs in the queen ; Bishop (1920 a) describes thesexual apparatus with special regard to fertilization ; Cheshire(1885), Bresslau (1905), Adam (1913) have written about the'sperm-pump'; finally Pruthi (1925) might be consulted withregard to the homologies of the genital ducts in insects.
The oviducts of insects are derived from the mesoderm. Theso-called ' oviducts ' of the queen and worker bee are obviouslyectodermal structures with a chitinous intima bearing back-wardly directed hairs, and they do not homologize in the dronebee with the vas deferens, including the vesiculae seminaleswhich are niesodermal structures. Thanks to Loschel and Meier(1916) we know the development of the parts concerned in thequeen and worker. For the sake of accurate terminology anotherword should be employed for the ' oviducts ' of adult bees,especially as the true oviducts are probably represented byLo'schel's ' Ovarialstrange' in the young pupa. However, theword ' oviduct ' will be retained in this paper.
Innervation is from the last, the fourth abdominal ganglionwhich lies above the anterior end of the vagina. The regionfollowing the ovarioles is surrounded by many diverging nerveswhich are liable to cause confusion in the unravelling of theinnervation. Other writers ignore the innervation, but Cheshire(1885) describes and figures many nerves associated with theparts posterior to the ovarioles, and he states that the muscularcontractions of the sperm-pump are produced by a reflex action.Snodgrass (1925) states that as far as we know the sperm-pumpis controlled by the ' will' of the bee. The numerous gangliathat Cheshire describes as scattered about the organs appear tobe the ganglionic swellings which I have described (Morison,1927) as occurring on the more delicate nerves in most parts ofthe body. The muscle-fibres of the oviducts are innervated byanastomosing nerve-fibres as on the vesiculae seminales of themale. I do not know how the nerves terminate on the musclesof the sperm-pump, though there are numerous very delicatenerves about this region. All other muscles attached to thefemale reproductive organs are innervated like somatic muscles,the nerve-ending in a Doyere's hillock on a fibre.

MUSCLES OF THE BEE 607
The tracheation of the muscles is dense and like that ofsomatic muscle-fibres. On the oviducts the tracheoles seemusually to leave the fibres to end blindly on the epithelium ofthe oviduct, as happens on the ventriculus. Incidentally it maybe remarked that the most complex anastomoses of tracheae inthe bee is exemplified in the covering of the spermatheca, whereneither muscle nor epithelium is served but the mass of sperma-tozoa obtained during fertilization. This case serves as indirectproof of the permeability of chitin to O2 and CO2, for presumablyboth' these gases will be encountered in the metabolism of thespermatozoa during their days or years of vigil in the sperma-theca, and the way of entrance and exit for the gases is via thechitinous walls of tracheae and spermatheca helped only by theblood, the very delicate tracheal epithelium, and the few epithe-lial cells near the opening of the spermatheca.
In contrast to the drone the muscles are confined to the ecto-dermal portions of the reproductive apparatus, and they are notcovered by an endothelial membrane. All the muscle-fibres showthe striation described for somatic fibres, and like them they maybe separated into sarcostyles or broken into discs at the telo-phragmata after suitable treatment. Nuclei are axial in position.
Only longitudinal muscles are present on the oviducts. Theydo not anastomose, but are more or less flattened against theoviduct, on which they form a sparse layer. They are 3-13/nwide, sarcomeres average 3-5/u, axial nuclei occurring singlyor in rows with or without undifferentiated sarcoplasm are9-12:2-4:2-4/u., but are rare. The sarcolemma is distinct. Thefibres start near the apex of each oviduct and continue to thebase, where they are most closely placed to one another. Theirprobable function is to help in the outward passage of the egg.
For details of the anatomy and possible functioning of thesperm-pump the reader is referred to the works of Bresslau andAdam. Three unpaired and three paired muscles are found.The fibres are 3-8^ wide, with axial nuclei rarely present.
A pair of muscles with numerous fibres is attached on eitherside of the mid-dorsal line of the vagina posterior to the basesof the oviducts and anterior to the opening of the spermathecal
NO. 284 R r

608 GUY D. MORISON
duct. Each muscle curves over the side of the vagina, and someof its fibres are attached to the anterior margin of the seventhurosternum just posterior to the attachment of each medianintersternal retractor, whilst other fibres become fastened to theventral surface of the vagina. The first lot of fibres would servein anchoring the vagina to the skeleton, also partly as aretractor, the second lot would act as a compressor and inaltering the position of the chitinous fold on the ventral surfaceof the vagina opposite the opening of the spermathecal duct.The attachment of the muscles to the seventh urosternummight be expected from what is known of the development ofthe reproductive organs.
On either side of the mid-dorsal line of the vagina, betweenthe above muscles and the opening of the spermathecal duct,a pair of long, strap-like muscles arises. Each passing ventradto the bursal pouch of its side is attached by a short tendon belowthe anterior margin of the eight abdominal spiracular plate ofits side of the body. The muscles would function in anchoringthe vagina to the skeleton and in pulling it backwards and soantagonizing the pull of the first lots of fibres of the anteriormuscles described above.
Each bursal pouch bears a many-fibred muscle on its anteriorhalf. The muscle curves round the pouch and is attached to itsdorsal and ventral surfaces. It would act as a compressor of thebursal pouch.
The last three pairs of muscles consist of typical somaticfibres 8-15/x wide. Probably all three pairs are utilized in ovi-position, and are helped by many other muscles of the body.They may have a special function during fertilization of the eggand during mating.
Muscles of t he E e p r o d u c t i v e Organs of t heW o r k e r .
The reproductive organs of the worker, like those of thequeen, do not seem to have received a full account of theirmorphology in any one work. Dufour (1834) seems the firstto describe and figure them. Adam (1913) deals with the rudi-
MUSCLES OF THE BEE 609
mentary sperm-pump, Meier (1916) with their developmentfrom pupal to adult stage, and Zander (1922) gives the bestgeneral description of them.
The organs are easily dissected out of a worker fixed andhardened in alcohol. Except for their rudimentary conditionthey are just like the corresponding apparatus of the queen,but the last abdominal ganglion which innervates them is the
TEXT-FIG. 14.
Arrangement of muscle-fibres on oviduct of worker. Formalin,Delafield's haematoxylin, glycerine. Lettering: I m, longitu-dinal muscle-fibres ; nv, nerve.
fifth. The nerves on the oviducts are very delicate (3/J, wide)and they anastomose with one another as well as twistingamongst the muscle-fibres. Their structure is the same as onthe oviducts of the queen. Nerve-endings were not observedon the muscles of the oviducts. Owing to the delicacy of nerve-and muscle-fibres on the oviducts, the two may be confusedquite easily, especially when the cross-striation of the muscleis not easily discernible (Text-fig. 14). Adam has noted that
E r 2

610 GUY D. MOEISON
nerves occur near the rudimentary sperm-pump muscles, butI have not distinguished their terminations on the muscle-fibres of that region. The muscles attached to the vagina areinnervated like the ordinary somatic muscles, and probablythis is the case with the muscle-fibres on the part correspondingto the bursa copulatrix of the queen. Tracheation of the musclesis as described for the queen.
All the muscles found on. the reproductive organs of the queenare represented on the worker, but considerably reduced innumbers and width of fibres.
The sole muscle-fibres of the oviducts take a longitudinalor spiral course with occasional connecting strands betweenindividual fibres (Text-fig. 14). They vary in width 1-9/*.A sarcolemma seems to be present always in fibres down to 3JU.wide. I have not detected it in fibres below this width, butit probably exists as such fibres along parts of their courseoften show slight expansions which seem less stained than themain axis of the fibre. Small nuclei averaging 4: 5: 3/J, Avith oneor two nucleoli occur singly and infrequently along the courseof the fibres. They always seem to lie between the sarcolemmaand the contractile substance. This would be their best positionin order not to impede contraction of the sarcostyles in suchdelicate fibres. The striation is like that of somatic muscle-fibres, but strong contraction waves are rarely seen. The telo-phragmata are distinct membranes, and on various occasions,in fibres which have been subjected to longitudinal strain, theyhave come to lie in an oblique position to the long axis of thefibre (Text-fig. 15). As in the queen the muscle-fibres lie abovea squamous epithelium which covers a chitinous intima bearingbackwardly directed spines 6-1 0/J. long.
The reader is asked to consult Adam's paper for the muscle-fibres at the sperm-pump. All I have seen is some portions ofmuscle-fibres at this part.
The two pairs of muscles attached to the vagina and thepair on the part representing the bursa copulatrix are like thoseof the queen, except that the fibres of the first two pairs areonly 6-9/u. wide, and those of the last pair are about 6/u. wide.

MUSCLES OF THE BEE 611
F ib rous Indi rec t Muscles of the Wings .
The muscle-fibres coming under the histological type offibrous muscle are confined to the paired longitudinal andvertical indirect muscles of the wings. They are characterizedhistologically by being split into sarcostyles (fibrils) veryeasily; by each fibre having many rows of nuclei which arenot axial in position ; by the absence of a sarcolemma ; by the
TEXT-FIG. 15.
lltstitt
Histology of portions of a fibre on an oviduct. Two telophragmata(Z) have been dislocated during preparation of the fibre. Letter-ing : J, light disc ; nu, nucleus ; s, sarcolemma.
presence of bodies (sarcosomes) between the fibrils ; and bytheir colour, which is yellowish in diffused daylight and socontrasts with the almost colourless tubular fibres.
The longitudinal and vertical muscles are similar in the threecastes except for the variation in size, and a tendency for asmaller number of fibres in the worker than in the drone andqueen in which the numbers are about equal. The various racesof bees kept in Great Britain, such as Italian, Carniolan, Dutch,and other dark varieties, seem to have about equal numbers of

612 GUY D. MOEISON
fibres in the individual indirect muscles of the wings. The table(p. 613) of the numbers of muscle fibres was obtained by countingall the fibres for each of the indirect muscles of the wings oftwenty bees of each caste. The bees were obtained from manyparts of Great Britain and include many varieties. Alcohol fixedand hardened material dissected under a Greenough microscopeis the most convenient for obtaining counts of fibres. Only rarelydo equal numbers of fibres occur in both muscles of a pair, butthe difference is usually very slight: one to ten fibres in thethree castes. If a muscle on one side of the thorax has manymore fibres (e. g. thirty) than its fellow of the other side, thenit seems that its antagonist on the same side of the thorax asitself has fewer fibres than its pair of the other side, and so themuscles compensate one another on either side of the thorax.Nevertheless the sums of the fibres of the two muscles on eitherside of the thorax almost always show a slight difference,which is apparently of no importance to the activity of the bee.
A resting fibre is a long, slender, straight, more or lesscylindrical rod which is easily separated from its fellows whenexamined in the fresh state or after fixation in alcohol. Theindividual fibres of both longitudinal and vertical muscles liealmost parallel to those nearest them, but all the fibres in amuscle are not equally parallel to one another because the areasof attachment to the chitin are not the same shape at eitherend of a muscle. Owing to the curvature of the chitin the endsof the fibres vary slightly in shape, being either parallel to oneanother and vertical to the long axis of the fibre or more or lessoblique to one another and to the long axis of the fibre. Thefibres vary in length according to their position in a muscleand according to the caste of the bee, but the length is scarcelyinfluenced by contraction in bees which have been killed withoutthe thorax being opened. The variation in width is the same forboth longitudinal and vertical muscle-fibres. The width isgreatest in the drone, least in the worker. It is constant for thewhole length of fresh muscle-fibres examined on a slide, but infixed material it is not so constant and the latter conditionprobably is maintained during life when the fibre is subject to
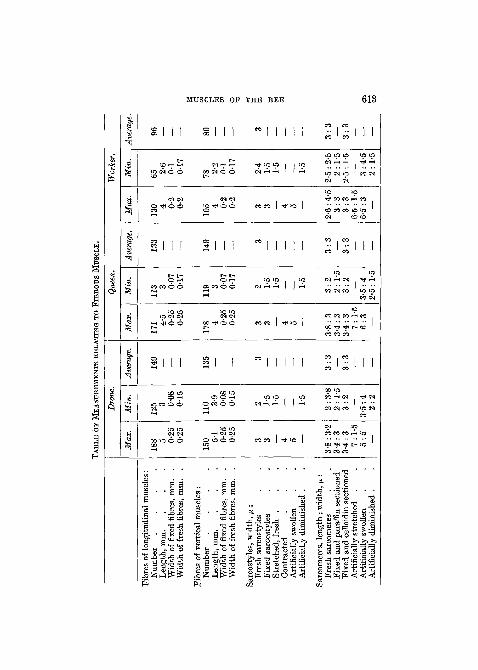
TA
BL
E O
F M
EAST
TBEM
ENTS
RE
LA
TIN
G T
O F
TBEO
TTS
MU
SCL
E.
Fibr
es o
f lo
ngitu
dina
l m
uscl
es:
Num
ber
..
..
Len
gth,
mm
.W
idth
of
fixe
d fi
bres
, mm
. .
Wid
th o
f fr
esh
fibr
es,
mm
. .
Fibr
es o
f ve
rtic
al m
uscl
es :
Num
ber
..
..
Len
gth,
mm
.W
idth
of
fixe
d fi
bres
, mm
. .
Wid
th o
f fr
esh
fibr
es, m
m.
.
Sarc
osty
les,
wid
th, p
:Fr
esh
sarc
osty
les
Fixe
d sa
rcos
tyle
sSt
retc
hed,
fre
sh
.C
ontr
acte
dA
rtif
icia
lly s
wol
len
Art
ific
ially
dim
inis
hed
.
Sarc
omer
es,
leng
th :
wid
th, y
.:F
resh
sar
com
eres
Fixe
d an
d pa
raff
in s
ectio
ned
Fixe
d an
d ce
lloid
in s
ectio
ned
Art
ific
ially
str
etch
ed
.A
rtif
icia
lly s
wol
len
Art
ific
ially
dim
inis
hed
.
Dro
ne.
Max
.
188 5 0-
250-
25
150 5-
10-
250-
25
3 3 4 5 —
3-8
: 3-
23-
4: 3
3-4:
37:
1-5
5:5 —
Min
.
125 3 0-
080-
15
110 3-
90-
080-
15
2 •
1-5
1-5
— —• 1-5
2:3-
82
: 1-
53
:2 —
3-5:
42
:2
Ave
rage
.
149
— — — 135
— — —
3— — — — — 3: 3 — 3:3 — — —
Que
en.
Max
.
171 •4
'5 0-25
0-25
178 4 0-
250-
25
3 3 4 5 —
3-8:
33-
4:3
3-4:
37
: 1-
56
:3 —
Min
.
113 3 0-
070-
17
119 3 0-
070-
17
'2 1-5
1-5
_ — 1-5
3: 2
2:1-
53
:2 —
3-5:
42-
5 :
1-5
Ave
rage
.
- 13
3
. .
— 149
—
3_ . — •
—
— 3: 3 — 3:3 — — —
Wor
ker.
Max
.
130 4 0-
20-
2
105 4 0-
20-
2
3 3 4 5 —
2-6
: 4-
53
: 33
:36-
5 :
1-5
6-5
: 3
—
Min
.
652-
60-
10-
17
782-
20-
10-
17
2-4
1-5
1-5
— — 1-5
2-5
: 2-
52
: 1-
52-
5 :
1-5
— 3:4
-52:
1-5
Ave
rage
96 — 89 —
3— — — — 3: 3 — 3:3 — ——
O

614 GUY D. MOEISON
contraction and unequal pressure along its length from tracheaeand adjoining fibres. The table (p. 613) of the length and widthof fibres is founded on measurements obtained from manydifferent varieties of bees of all castes. The fibres were examinedafter fixation of the undamaged thorax in alcohol or formalin.The fresh fibres were examined in Toison's or Ringer's solutionor in 0-75 salt solution.
Each fibre consists of a number of sarcostyles as long asitself, but extremely narrow in diameter. In both the longi-tudinal and vertical muscle-fibres the number varies between1,000-2,000, depending on the width of the resting fibre andshowing no particular variation for the caste of bee. Thenumbers were obtained as the results of division and multiplica-tion. The area of a thin transverse section of a stained fibre wasmeasured and divided by another measured but smaller areain which the number of sarcostyles had been counted, then thequotient was multiplied by the counted number of sarcostylesto give the approximate number for the whole fibre.
Sarcostyles always seem to be perfect cylinders whenexamined fresh in a suitable solution or after certain fixationand treatment. When ' resting ' in the fresh state in the workerthey measure 2-4-3/J. in diameter. However, they can beartificially stretched with 'ease to a diameter of 1 -5 fx or less.When very strongly ' contracted ' they measure 3-5-4^. Bysubjecting them to dilute acids as in gold-chloride staining, theymay swell to 5/J.. Exosmosis in a hypertonic solution will reducethem to 1-5/x. The table (p. 613) expands what is stated hereabout the dimensions of sarcostyles, but the figures only refer toshort portions of a sarcostyle. In fact figures need wary inter-pretation for ' fresh ' material, since no matter what the treat-ment of the muscle may be, the sarcostyles are subject to moreor less considerable strains before they can be measured, andin a preparation they will vary in width from 1-5 to 4/*. Thesefigures may occur along the length of a single sarcostyle, and itmay be impossible to determine whether they are due to naturalcontraction or artificial stretching, or a combination of the two.Brunnich (1911) gives 0-5-2/x as the width of sarcostyles in the

MUSCLES OP THE BEE 615
bee, but these figures probably were measured from sectionedand shrunken fibres. Kolliker (1888) gives 1-4/n and Biitschli(1891) 3/M as the width of sarcostyles in a number of differentspecies of insects.
Like the tubular muscle-fibre, the fibrous muscle-fibre ismarked by transverse striation. Jordan's (1920) nomenclatureof the striation for the wing-muscles of a wasp will be adoptedin this description. Although striation is very obvious in afibre, it has many of its details obscured, and it is seen in itsgreatest complexity in isolated sarcostyles. This is contrary tothe general case in tubular muscle-fibres, where the extremedelicacy of isolated sarcostyles renders the striation very faint.
The three sarcomeres in each of Text-figs. 20 and 22 show thegreatest complexity of striation that may be seen in a portionof a sarcostyle. They are supposed to be in a state of contrac-tion. A sarcostyle freshly dissected in Toison's solution or egg-albumen and differing only very slightly in refractive indexfrom the surrounding medium, and another sarcostyle elabo-rately and darkly stained, and a third removed from the bodyof a bee kept in a cool room till five days after its death, may allappear as in the figure, though this degree of complexity iscomparatively rare. From their optical and chemical propertiest e l o p h r a g m a t a seem true membranes lying across thecolumn of the fibril at regular intervals. Each telophragmaappears as a narrow, dark, transverse line. Often it is to be seenprojecting slightly on either side of the sarcostyle around whichit would form a frill. Probably a telophragma of a fibril isunited by the frill to a telophragma in each of the other fibrilsin such a way that a membrane is formed right across the fibre."Presumably this happens with all the telophragmata of a fibrilso that the fibril is crossed by a series of membranes parallel toone another and at right angles to the axis. However, the mem-branes cannot be seen in their entirety because of their delicacy,iior are they complete because they are pierced by tracheoles,nuclear spaces, and perhaps other tissue. Thulin (1914), afterexamining a number of animals, voices the arguments for andagainst the existence of a complete membrane, but his methods

616 GUY D. MORISON
of examining insects could be improved. Evidence for the con-clusion that the telophragmata are more or less linked togetherin series may be derived from (1) the appearance of the fibrein which the transverse striations are most marked at thetelophragmata. These crossing at right angles to the long axisof the fibre form a series of uninterrupted planes parallel to oneanother. (2) The way in which distortion of one fibril of anisolated pair of adjoining fibrils may affect the other. (3) Thearrangement of sarcosomes. It has been suggested that thetelophragmata in the muscles of some other animals are doublemembranes. If this is the case in the bee, the most positiveevidence for it is that the sarcostyle when broken usually breaksat its telophragmata. The telophragmata are always visible, butthey will differ in apparent length, depending on the state ofcontraction of the fibrils.
The portion of a sarcostyle enclosed between two succeedingtelophragmata is a s a r c o m e r e . The l e n g t h of all theelements of a sarcostyle is measured longitudinally, whilstthe w i d t h is measured transversely to the long axis ofthe sarcostyle.
The m e s o p h r a g m a seems to be a very delicate, trans-verse membrane dividing the sarcomere into halves. It isusually visible as a thin, dark line across the middle of a coloureddisc (Text-figs. 18, 20) or across a colourless disc (Text-fig. 22).When it is absent its place may be occupied by a more or lesshomogeneous dark disc filling the greater part of the sarcomere(Text-figs. 16, 17, 19), or by a colourless disc (Text-fig. 21)which is bounded at its ends by coloured discs. Jordan (1920)suggests that it may be a membrane which, stretched across thefibril, is so delicate that only under certain conditions of thestretching of fibril is it sufficiently relaxed to become visible.This is a plausible explanation, and after the examination of anumber of fibrils in various stages of contraction, decomposi-tion, and artificial distortion, it seems to me that it is true,only that in many cases the mesophragma remains visible andis marked by a greater or lesser deposition of the denser con-tractile substance on either side of it. It is the most delicate
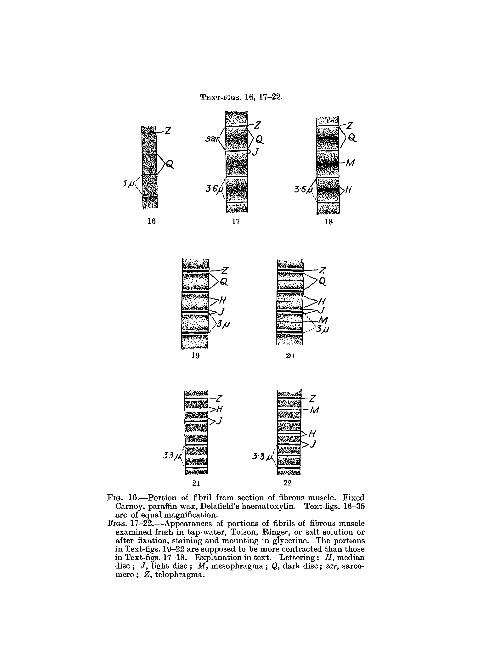
TEXT-ITOS. 16, 17-22.
San
3-6// 36A
-z•>H
>J
21
33fj{
mmmm
mmmmmm
~M
>J
FIG. 16.—Portion of fibril from section of fibrous muscle. FixedCarnoy, paraffin wax, Delafield's haematoxylin. Text-figs. 16-35are of equal magnification.
FIGS. 17-22.—Appearances of portions of fibrils of fibrous muscleexamined fresh in tap-water, Toison, Ringer, or salt solution orafter fixation, staining and mounting in glycerine. The portionsin Text-figs. 19-22 are supposed to be more contracted than thosein Text-figs. 17-18. Explanation in text. Lettering: H, mediandisc ; / , light disc ; M, mesophragma; Q, dark disc; sar, sarco-mere; Z, telophragma.

618 GUY D. MORISON
of the elements of the sarcostyle. I find no signs of it linking upwith other mesophragmata after the style of the telophragmata,except that the sarcosomes which lie outside the fibril aredivided at the levels of the mesophragmata as they are at thetelophragmata. Neither the mesophragma nor any of the otherelements are robust enough to be distinguished as entities intransverse sections of fibrils.
The d a r k d isc is a darker coloured mass of contractilesubstance occupying the greater part of the sarcomere. Instained preparations it may be a homogeneously coloured disc
TEXT-FIGS. 23 AND 24.
WMKMI
—M
24
Fio. 23.—Portion of fibril strongly contracted in white of egg inwhich the living muscle was dissected. Z, telophragma obscuredby more opaque (contractile) substance ; M, position of meso-phragma indicated by more opaque substance ; H, median disc.
FIG. 24.—Portion of a contracted fibril swollen through treatmentwith Ranvier's gold chloride. Lettering as in previous Text-figs.
stretching from telophragma to telophragma (Text-fig. 16), or,in stained and unstained preparations, it may be separated fromeach telophragma by a portion of the l i g h t d i sc (Text-figs. 17,18), or, in stained preparations, it may be with a greater con-centration of darkly stained substance towards the telophrag-mata (Text-figs. 19, 20), or divided into two discs by a light-coloured m e d i a n d i sc (Text-figs. 21, 22), or, finally, concen-trated near or at both the telophragmata (Text-figs. 23, 24, 25,32, 33).
The m e d i a n d i sc is light coloured and it divides the darkdisc transversely into equal portions. Varying in length, it may

MUSCLES OF THE BEE 619
or may not be present, and when present it may or may notbe divided transversely by the mesophragma. It appears instained and unstained preparations and is probably a result ofcontraction of the sarcomere (Text-figs. 21, 22, 23, 24, 25).
The l i g h t d i sc is bisected transversely by the telophragmawhich it separates from the dark discs. Like the median disc, itvaries in length and it may or may not be present. It too isseen in stained and unstained preparations and probably owesits appearance to the state of contraction of the sarcomeres(Text-figs. 17, 19, 21, 22).
TEXT-FIGS. 25 AND 26.
25 26FIG. 25.—Portion of a contracted fresh fibril stained and distorted
by Nile blue sulphate in distilled water. Lettering as in previousText-figs.
FIG. 26.—-Portion of a somewhat contracted fibril obliquely stretchedand examined in water. Lettering as in previous Text-figs.
Physical laws would necessitate an envelope to surround thesarcostyle (cf. Hardy, 1926), but such an envelope would prob-ably be invisible from its delicacy. However, from theaccumulation of evidence, it appears that the sarcostyle hasa membrane which perhaps is seen under certain conditions.Jordan proposed the name s a r c o s t y l i c m e m b r a n e for theenvelope of the sarcostyle in a wasp, and this name is adoptedin this account. The evidence for the existence of a membranemay be presented as follows : (1) A membrane is imposed byphysical laws, but this membrane is independent of a mem-brane of visible structure. (2) The beaded appearance of a sarco-

620 GUY D. MORISON
style in a hypotonic solution, when the side walls of each sarco-mere bulge outwards but are constricted at the telophragmata(Text-fig. 27), and the opposite appearance when the side wallsare concave and the telophragmata project (Text-fig. 25), bothsuggest that the sarcostyle possesses an osmotic membrane.(3) The appearance of the telophragmata described under (2)also suggests that they are attached to a peripheral membrane.(4) Jordan (1920) finds that destaining transverse sections ofsarcostyles of the wasp leaves a central dot and a marginal ring,but I have not observed this for the bee. (5) Stiibel (1920), after
TEXT-FIGS. 27 AND 28.
Ubf
27 28
F I G . 27.—Portion of a fibril distorted by t reatment with dilute aceticacid. Lettering as in previous Text-figs.
F I G . 28.—Terminal portion of a fibril broken from its a t tachmentto the integument. Carnoy's fixative, paraffin wax, Delafield'shaematoxylin. Let ter ing: 'tf, tonofibrillae; Q, dark disc ; Z, telo-phragma.
examining various insects, including Apis, claims that by dark-ground illumination a membrane can be seen to envelop thefibril. My apparatus for dark-ground illumination was notquite so optically perfect as his, but my conclusion was thatreliance could not be placed on the appearance of a membraneunder dark-ground or oblique illumination or very brightreflected light. (6) Van Gehuchten (1886), after examiningmany insect muscles, described a membrane for fibrous musclefibrils. Marcus (1922), from the examination of B o m b u s ,favours the presence of an envelope, but he considers that thefibril is composed of still smaller units, and some of his methods

MUSCLES OF THE BEE 621
are open to the objection that they dissolve out substancesessential to the fibril and are liable to give rise to artifacts.Janisch (1924), who worked on B o m b u s t e r r e s t r i s , isopposed to the idea of Marcus of an enveloping sarcostylicsheath. Ebner (1918), who examined B o m b u s and V e s p a ,finds a membrane.
Whilst the existence of a sarcostylic membrane for theindividual fibrils seems probable, I do not find a sign of a sarco-lemma for the individual fibres. This agrees with the observa-tions of most histologists who worked on fibrous muscles ofvarious Hymenoptera. Numerous air-sacs or tracheae areclosely applied to the periphery of the fibres, and in transversesections they often appear exactly like an encircling niernbrane.They are arranged in such a way that they would bind thesarcostyles together.
Briinnich (1911) and Kielich (1918) observed that nucleiwere present in fibrous muscle-fibres of the bee ; Snodgrass wasnot able to find them, whilst most other authors ignore theirexistence. Nuclei occur in rows between the sarcostyles. Thisis their usual position in muscles of this type in other insects.With the hope of seeing nuclei in the fresh state or more or lessisolated and stained, I have examined, without success, manythousands of fresh fibres dissected in many different media.Yet nuclei are present. In fact hundreds may lie in a fibre andyet be undiscernible owing to their resemblance to the sarco-somes which lie in the same position between the sarcostyles.Tissue stained in iron haematoxylin after fixation in osmiumbichromate shows the most elaborate pictures of the sarco-styles and sarcosomes, but it becomes very difficult or im-possible to distinguish the nuclei from the sarcosomes. The bestmethod of demonstrating nuclei is to fix in an alcoholic fixative(like Carnoy I or II) which dissolves certain substances in thesarcosomes and presumably in the fibrils, stain deeply in Dela-field's haematoxylin, differentiate very considerably in acid-alcohol, and counterstain with eosin. This method shows verylittle detail for the sarcostyles, whilst the sarcosomes are barelydiscernible, but the nuclei appear brightly contrasted against

622 GUY D. MORISON
an eosin-stained background, especially if the tissue has beenexposed to ammonia vapour or a dilute alkaline bath after theacid alcohol. The objection to this method is that it probablyshrinks the nuclei, though they do not show any obvious signsof shrinkage. It certainly reduces the diameter of the sarcostylesand the size of the sarcosomes, especially if paraffin embeddingis adopted instead of celloidin. Romanovsky stains, Light-greenand Benda-Ehrlich-Bondi amongst other stains, did not provesatisfactory for demonstrating nuclei.
TEXT-FIG. 29.
Nuclei of fibrous muscle. Carnoy's fixative, paraffin wax,Delafield's haematoxylin.
Nuclei measure 7-6:4:4-15:2:2/u, in tissue which shows theminimum or no shrinkage for the sarcostyles. They are veryconstant in size, about 10:2-5 :2-5/u for all castes. When stainedin iron haematoxylin they are seen to contain one or moredarkly staining bodies, whilst Delafield's haematoxylin revealsthese and other smaller bodies equally stained and placedseemingly towards the periphery of the nucleus (Text-fig. 29).Seven or eight rows of nuclei may be seen in a 7/n thick sectionof a fibre 110/x wide, and forty to fifty nuclei will appear in atransverse section of a similar fibre. The width of two to threesarcostyles separates the most peripheral nuclei from the surfaceof the fibre. Each of the intervals between the individual nuclei

MUSCLES OP THE BEE 623
of a row is often not much longer than the length of a nucleus.A row may proceed for a long way through a fibre, but I do notknow if any row traverses the whole length, nor do I know if itis in contact with the same sarcostyles for all the distance ofits course. The nuclei lie cushioned in more or less undifferen-tiated sarcoplasm, i.e. protoplasm which is included in themuscle-fibre but which is not sarcostylar nor apparently belong-ing to sarcosomes or nucleus. The positions of the rows ofnuclei are not determined either by the hairs above the inser-tion of the muscle-fibre or by the reticulate sculpturing of theexternal surface of the cuticle, though the former may suggestat times a controlling influence in some longitudinal sections ofthe fibres.
Owing to the difficulties of distinguishing the nuclei fromsarcosomes after osmium bichromate fixation and iron haema-toxylin staining, no interpretation is offered of the relationshipbetween nuclei, sarcosomes, sarcostyles, and tracheoles (cf.Holmgren, 1908, 1910), but from the numbers of nuclei it issupposed that they would have an important role in the physio-logy of the fibre.
Judging from the literature contraction of living fibrousmuscles of insects has been observed but seldom. Merkel (1872)claimed to have seen it happening in the muscles of the beedissected in white of egg. I have not seen fibrous muscledefinitely contracting either as whole muscles or as smallerunits such as fibres or fibrils, in spite of using many differentmedia at varying temperatures for the examination of freshtissue. Perhaps after a quantitative chemical analysis of themuscle a suitable fluid will be found which will resemble thesarcosomes in composition. A continuously high temperature(30°-40° C.) may be necessary also for the contractions to becontinued.
The isolated fresh fibre is yellowish by diffused daylighteither reflected or transmitted, but the fresh fibril, isolated ina colourless solution like Toison, Einger, or 0-75 per cent, salt,is colourless and marked only by certain less transparent discs.These transparent and less transparent discs are arranged
NO. 234 S S

624 GUY D. MORISON
amongst the sarcomeres of a fibril in the ways described above.There is a gradual transition in the ways of arrangement of thediscs which corresponds with a difference in the comparativelengths and widths of the sarcomeres. Since a certain arrange-ment corresponds with a maximum increase in the width coupledwith a minimum length of the sarcomeres, this arrangement(Text-figs. 19, 20) is assumed to mark the greatest degree ofcontraction, as seen in fibrils examined in most liquid mediabefore or after fixation. However, a portion of fresh fibril mayhave the appearance shown in Text-fig. 23 when stronglycontracted after dissection in white of egg. In these cases eachtelophragma is more or less obscured on both its sides by thesubstance of the dark discs, whilst the median disc is visiblemore or less as a light area. The opposite case (Text-fig. 17),when each telophragma appears as a thin dark line traversinga light disc, and the dark disc occupies the middle of the sarco-mere to the exclusion of the median disc, is assumed to be theresting state of the fibril, since at this stage the sarcomeres arelonger than wide. The mesophragma may or may not be seenin either state. Between these two extreme stages there comethe intermediate stages when the dark disc is being divided intohalves and is more concentrated either at the telophragmata orthe mesophragma. Therefore contraction seems to consist essen-tially of a more opaque, motile substance concentrating at thetelophragmata. The stages described above can be seen infresh tissue; however, the whole fibril will absorb most stainsafter a time, and then it is seen that the less transparent discsbecome more deeply coloured than the transparent discs. Sincethe details of striation are more easily seen hi stained than inunstained tissues, it is usual to adopt methods of fixation andstaining for the observation of fibrils, especially as stainingbrings into view the intimate relationship between the degreeof contraction of the sarcomeres and the state of thesarcosornes.
Sarcosomes are complex organic substances which are aggre-gated in small semi-fluid bodies placed between the fibrils inperfect regularity with the striations. Their chemistry will be

MUSCLES OF THE BEE 625
discussed in greater detail later and it will suffice now to saythat they seem to consist of albumin, glucose, cytochrome,lecithin, and probably other substances. Their function seemsto be that of reserve ' food ' material which can be utilizedduring very rapid and rhythmically continued museular con-traction. Similar bodies have been found widely in the animalkingdom in muscle which contracts under the three associatedconditions of rhythm, rapidity, and more or less long duration.They appear in the tongue (Svartz, 1914) and heart (Holmgren,1907) of man and other vertebrates, in the pectoral muscles ofbirds and bats, and apparently in all insects which have fibrousmuscles in the thorax. They may be present in tubular muscle,but I never found a trace of them in the tubular muscles of thebee. Their reported occurrence in tubular muscles of someinsects has been doubted, and it seems advisable that such casesshould be re-examined chiefly with regard to the size of thesarcostyles. Modern investigators consider the sarcostyles oftubular muscle very delicate, and if they are of the delicacystated, 0-2-0-4/A diameter, it is scarcely to be expected thatsarcosomes would be large enough to be visible in the samearrangement as in the fibrous muscle of the bee.
The literature on sarcosomes in the muscles of the animalkingdom is very extensive and scattered, and it suffers chieflyfrom its authors' lack of knowledge of the chemistry of thebodies concerned. Many detailed histological accounts havebeen published and techniques have been devised to show bodiesin the place normally occupied by semi-fluid substance in theliving animal, but what is needed is a detailed chemical analysisof the substance and the adjacent muscle-tissue. It seems thatsarcosomes, at least in the fibrous muscle of the bee, are intimatelybound with the contraction of the sarcomeres, and if this be thecase a chemical investigation of them may supply the long-sought steps in the chemical changes to which the contractionof muscle is due. Largely owing to the lack of knowledge ofthe chemistry of sarcosomes, various authors have stated quitedefinitely that the sarcosomes are either mitochondria or Golgibodies, whilst others as definitely deny this. The problem of the
S S 2

626 GUY D. MORISON
relationship of these various bodies needs detailed investigation,as some people have remarked.
After reading Nath's paper (1926) it seems to me that thesarcosomes are probably very closely allied to mitochondria orthe Golgi apparatus or perhaps to both, though Nath does notinclude a reference to sarcosomes in his work.
Many authors (e. g. Holmgren) have been so struck by theregular arrangement of sarcosomes between the muscle fibrilsthat they have stated that the sarcosomes are as constant intheir position and appearance as the striation of the fibril towhich they are intimately related ; other authors have noticedthe rows of sarcosomes between the muscle-fibres, but they havenot been so decided about the regularity nor the intimaterelationships between the sarcosomes and the striation of thefibrils. However, it seems to me that the former school willprove to be right, whenever techniques are adopted which willpresent on the slide perfectly longitudinal or transverse sectionsof sarcostyles cut at the thickness (2-4 JX) of the sarcostyleswhich must not have altered in size from their living state at anytime during the process of preparing the mount. The mostsatisfactory method for demonstrating sarcosomes and fibrousmuscle-fibrils in the bee is that of fixation in osmium bichromateor Flemming's fluid without acetic acid followed by embeddingclove-oil celloidin and staining in iron haematoxylin as describedfor the alimentary canal muscles.
It should be pointed out that the sarcosomes or microscopicbodies described by various authors as lying between the sarco-styles of several animals are not likely to be homologousthroughout the animal kingdom. Holmgren (1908) divided thesarcosomes found in the muscles of insects and other animalsinto two categories : Q-sarcosomes, which are large and asso-ciated with the dark disc, and J-sarcosomes, winch are smallerand intimately associated with the accessory disc. The J-sarco-somes are still recognized by histologists, but not in Holmgren'soriginal meaning. I have not seen them in the tubular or fibrousmuscles of the bee, in which the accessory disc is also absent.Only the Q-sarcosomes are present in the fibrous muscles of the

MUSCLES OF THE BEE 627
bee. In the bee the proportions of the chemical constituents ofthe sarcosomes would probably vary with the degree of con-traction of the fibrils, and no microscopical technique yetdevised can show a 11 the constituents as they would lie in thesarcosomes and fibrils at any stage of contraction during life.The appearances about to be described are what seems the mostapproximate representation of the passage of substance betweensarcosomes and fibrils during the activity of the latter, but itshould be remembered that the essential sugar and variousother constituents have been dissolved out and replaced byother compounds.
TEXT-FIG. 30.
The shapes assumed by the sarcosomes when examined in differentmedia.
Portions of longitudinal sections of fibres depicted in Text-figs. 81, 32, 33 show the relation of sarcosomes to fibrils intissue which has been deeply stained in iron haematoxylin afterthe treatment described above. In the resting stage (Text-fig. 31) the fibrils are stained only slightly, the sarcomeres arelonger than broad, and the telophragmata visible as fine trans-verse lines, whilst the sarcosomes are very darkly stained andlarge and conspicuous. In the transitional stage (Text-fig. 32)the fibrils are more darkly stained, the sarcomeres are broader,the telophragmata are dark transverse bands, and the sarco-somes are paler in colour and smaller. In the last stage of con-traction (Text-fig. 33) the sarcostyles are quite pale in colour,but the sarcomeres are short and broad and the telophragmataare marked by long very deeply stained discs, whilst the sarco-somes are barely visible. As the fibrils contract, the space
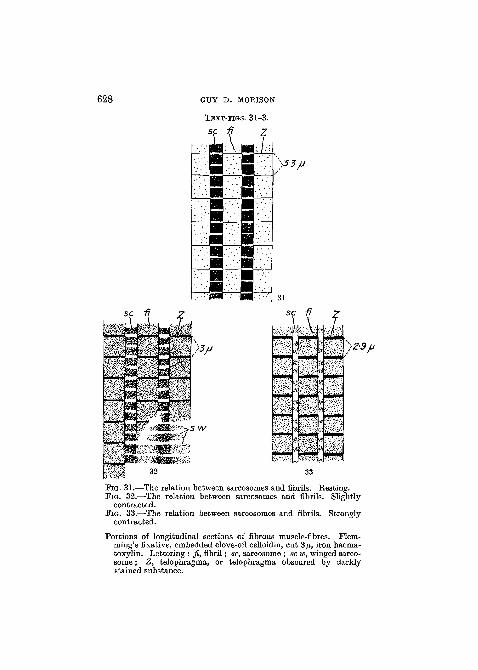
628 GUY D. MORISON
TEXT-FIGS. 31-3.
')3-3/J
yz-9/j
FIG. 31.—The relation between sarcosomes and fibrils. Resting.FIG. 32.—The relation between sarcosomes and fibrils. Slightly
contracted.FIG. 33.—The relation between sarcosomes and fibrils. Strongly
contracted.
Portions of longitudinal sections of fibrous muscle-fibres. Flem-ming's fixative, embedded clove-oil celloidin, cut 3n, iron haema-toxylin. Lettering : fi, fibril; sc, sarcosome ; sc w, winged sarco-some; Z, telophragma, or telophragma obscured by darklystained substance.

MUSCLES OT? THE BEE 629
between them is reduced to the exclusion of the sarcosomes.The sarcosomes in longitudinal sections of a fibre may rest likea row of beads between two adjacent fibrils—two sarcosomesalways touching each lateral margin of a sarcomere, or theymay be connected to one another transversely as depicted(Text-fig. 32). According to the way in which the sarcosomesubstance has been sectioned and its degree of condensationbetween the sarcostyles, it may have a dark central body anda pair of lateral expansions when it is generally called ' winged'(Text-fig. 32). The sarcosome substance in transverse sectionsof fibres is seen more or less to fill the spaces between fibrils(Text-fig. 34).
TEXT-FIG. 34.
Transverse section of fibrils (fi) and sarcosome substance (sc). Flemming'sfixative, embedded clove-oil celloidin, cut 3 ix, iron haematoxylin.
The so-called ' sarcosomes ' in a section of a fibre are afixation-product of a semi-fluid substance. Depending on thestate of contraction of the fibre, they may be entities or onlythe more condensed portions of a semi-fluid layer which extendsright across the fibre. Bach layer of sarcosome substance,whether it is composed of isolated sarcosomes or of sarcosomesunited to one another by wing-like expansions, seems boundedbetween successive telophragmata and mesophragmata of thefibre. Therefore the number of layers of sarcosome substance ina fibre would be double the number of sarcomeres of the fibrilwith the most sarcomeres in the same fibre. Only it is probablyless than this, because the terminal two or three sarcomeres ateither end of a fibril are connected with the insertion of themuscle on the chitin, and they do not seem to have sarcosomes.

630 GUY D. MORISON
Since the sarcosome substance is more or less fluid it must bekept in place by some method. The telophragmata show fairevidence of being limiting membranes beyond the sarcostyles,but the only sign of the extension of the mesophragmata is thatthe sarcosomes are separated from one another at the levels ofsuccessive mesophragmata as well as at the telophragmata.There is no sarcolemma to prevent the substance oozing fromthe sides of the fibre, but the substance is never present in greatquantity, and its intimacy with sarcostyles and tracheoles shouldbe ample in stopping its outward flow by the phenomenon ofcapillarity alone. The fibrils at the periphery of a fibre havenot got sarcosomes on their exterior surface.
As far as I can determine, the sarcosome substance enters thesarcomeres over the general surface of each half of a dark disc,and it moves concurrently towards the telophragmata for thecontraction of the sarcomeres. When leaving it takes the samepassage only now it is more concentrated at the mesophragma.A large portion of the dark-staining substance may enter eachhalf-sarcomere, but there always seems to be a residue left out-side. In the relaxed fibril when the substance is very conspicuousoutside, the fibril is yet stained faintly the same colour as thesubstance, but it is impossible to say if the staining indicatesthat some of the substance still remains in the fibril.
The layers of sarcosome substance must be in contact withrows of nuclei and numerous tracheoles as well as the sarco-styles. It seems quite probable that the nuclei are intimatelyconnected with a chemical change in the substance as Holmgrenhas described for various insects, but my methods of staininghave revealed nothing. The tracheoles must pierce the substancein all directions and probably they are very essentially connectedwith it in the exchange of gases, but again my staining techniquefails in answering. Finally, it must be concluded that the bloodpercolates through the fibres to this nutritive substance todeposit and remove most of the products of metabolism.
From the examination of many sections of whole thoracesof bees, it seems to me that when a fibrous muscle contractsa definite wave of contraction of the fibres takes place in one

MUSCLES OF THE BEE 631
direction and results in a greater diameter for the who lemuscle, and not in a greater diameter passing as a wave throughthe muscle. This is achieved by some of the fibres being mostcontracted near one end of attachment, others most, contractedin the middle, and others most contracted near the other endof attachment. The benefit of this method is that a strong waveof contraction does not surge along the muscle to compressadjoining organs such as the aorta and the antagonistic pair ofmuscles. I do not know the working of the regulatory apparatus,but the reader is asked to refer to what was said about thesemuscles when they were named as somatic muscles (Morison,1927), and Kahn (1916) may also be consulted for the rate oftransmission of impulse in muscle and nerve, though his descrip-tion applies to tubular muscles of some Orthoptera.
The length of the fibres permits a number of rhythmic con-tractions to be present in a-fibre at the same time. In fact itseems that during the beating of the wings all the muscle fibresare in a state of constant tension, only the tension becomesalternately stronger in each pair of muscles and so the wingsare raised and lowered. Though all the muscle-fibres, when thewings are beating, are in a state of tension which is most markedwhere the waves are broadest, yet these waves never consistof very intensely contracted fibres (judging by the comparativelengths and widths of their sarcomeres), and the spaces betweenthe waves are places of rest, and the sarcomeres are alwaysabsorbing or passing out sarcosome substance.
Essentially this idea of the contraction of a fibrous musclemeans that a fibre has a certain independence in contractionwhen working in harmony with the others of its muscle. Theidea was propounded also for tubular somatic muscles (Morison,1927).
It seems desirable to mention here that healthy fibrousmuscle may have the various appearances described above andyet be distorted artificially. After improper sectioning a fibremay have its sides showing a parallel wavy outline ; undercertain conditions a fibril at any stage of contraction may beartificially stretched so that its elements are arranged obliquely

632 GUY D. M0R1S0N
to its long axis (Text-fig. 26), or till its sarcomeres lose theirproportions ; dilute organic acids swell the fibrils till they pre-sent a beaded appearance constricted at the telophragmata(Text-fig. 27); hypertonic solutions will cause the telophrag-mata to project and the sides of the sarcomeres to be concave(Text-fig. 25). A fibril with a brush-like end is one that has beenbroken near its insertion on the chitin (Text-fig. 28).
When fresh or fixed tissue is dissected in a liquid medium thesarcosomes become disassociated from the fibrils, and their shapewill be determined by their chemical composition and by thatof the medium of examination and by the previous treatmentof the tissue. In Text-fig. 30 are depicted some of the shapesthey may assume.
Tracheation of the fibrous muscles of the bee is exceedinglycomplex, but it is obvious in many of its details in fresh muscle,and it has attracted the attention of older observers, yet inmodern text-books its description is hopelessly inadequate.Leydig (1859) noticed that round the fibrils the tracheolesformed a network like the anastomosing blood capillaries ofvertebrates ; Cajal (1890) has described this tracheolar systemfor various insects including V e s p a ; Holmgren (1908) de-scribes the system for B o m b u s t e r r e s t r i s , and mentionsthat Apis is similar; Kielich (1918) mentions the tracheolesin fibres ; Snodgrass figures the air-sacs and the larger branchesof the tracheae.
In order to see all the air-sacs and tracheae filled with airit is usually necessary to dissect a freshly killed bee in a strongsolution of salt or glycerine in water. However, the tracheolesappear quite clearly in tap-water or Toison's or Einger's solu-tion when examined by transmitted light under high-powerobjectives. Dark-ground illumination or very strong reflectedlight shows the tracheoles running like white threads betweenthe fibrils. After a variable period of time the tracheoles willnot be seen because of the opacity produced by the coagulation,of the proteins of the fibre. Therefore any reagent which coagu-lates the proteins renders the tracheoles invisible, and if it hasconsiderable wetting power like alcohol it will obscure the

MUSCLES OP THE BEE 633
tracheal system almost completely by displacing the air in thevessels. This, combined with the similarity of the refractiveindices of tracheoles and most mounting media, leaves thetracheoles invisible in the finished mount, and this is the reason•why most histologists ignore the tracheation of insect muscles.To observe the minute tracheation in permanent mounts it isnecessary to adopt the method of silver impregnation advocatedby Cajal and Holmgren, or the method of developing a latentsilver image after immersion of the tissue in a dilute solution ofsilver nitrate.
The tracheae passing to the muscles are delicate walled andthey usually branch from air-sacs. They are particularly abun-dant on the exterior of the muscles and are still very numerousbetween the innermost fibres. When penetrating the musclethey lie against the fibres in rows which are parallel to oneanother and at right angles to the long axis of the fibres. Thetracheae branch as usual giving off smaller branches whichsupply individual fibres. The final tracheae terminate ratherabruptly in a number of wide tracheoles on the fibres. After ashort course and a little branching the wide tracheoles end ina meshwork of minute tracheoles (0-2^ diameter). It can be seenthat every sarcomere on the exterior of a fresh fibre has at leastone tracheole passing over it, and many of the tracheoles canbe seen to pass into the fibre between the fibrils. There are nofree ends to the tracheoles ; they form a complete anastomosisthroughout a fibre. Employing the silver-nitrate method ofimpregnation the older histologists demonstrated the regularityof the position of the tracheoles, and its relation to the sarco-meres in fibrous muscles of insects, but I have not read a detailedaccount of the application of this method to the honey-bee.Text-figs. 35 and 36 show portions of sections impregnated withsilver. Every sarcomere is ringed almost completely by atracheole or portions of adjoining tracheoles. It will be seenthat tracheoles extend across the fibre in more or less parallelseries corresponding to the sarcomeres. The tracheoles of theseseries are liable to twist about amongst the fibrils and to takean oblique or vertical course in order to link up with other
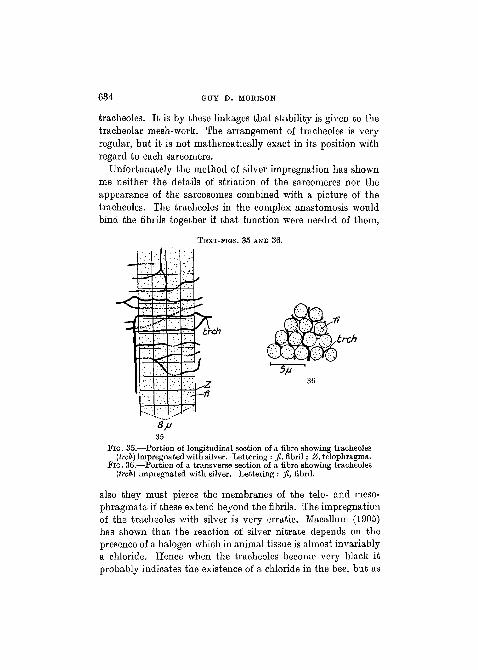
634 GUY D. MORISON
tracheoles. It is by these linkages that stability is given to thetracheolar mesh-work. The arrangement of tracheoles is veryregular, but it is not mathematically exact in its position withregard to each sarcomere.
Unfortunately the method of silver impregnation has shownme neither the details of striation of the sarcomeres nor theappearance of the sarcosomes combined with a picture of thetracheoles. The tracheoles in the complex anastomosis wouldbind the fibrils together if that function were needed of them,
TEXT-PIGS. 35 AND 36.
FIG. 35.—Portion of longitudinal section of a fibre showing tracheoles(trch) impregnated with silver. Lettering : fi, fibril; Z, telophragma.
FIG. 36.—Portion of a transverse section of a fibre showing tracheoles(trch) impregnated with silver. Lettering : fi, fibril.
also they must pierce the membranes of the telo- and meso-phragmata if these extend beyond the fibrils. The impregnationof the tracheoles with silver is very erratic. Macallum (1905)has shown that the reaction of silver nitrate depends on thepresence of a halogen which in animal tissue is almost invariablya chloride. Hence when the tracheoles become very black itprobably indicates the existence of a chloride in the bee, but as

MUSCLES OF THE BEE ' 635
the majority of attempts at impregnation are failures the halogencompound must have been washed out during fixation often todeposit the silver elsewhere on the fibre. Owing to the depositionof silver compound the tracheoles measure 0-3/x diameter.
No nuclei or epithelium were detected for the tracheoles.The tracheoles within the fibre apparently lie as intra-cellularstructures in the semi-fluid sarcosome substance which wouldbe very similar to an epithelium. However, the demarcation oftracheoles with silver seems to show that they are occasionallyenveloped by a chemical which holds the metal, whilst thesarcosome substance never does so. This may be used as anargument for the presence of a tracheolar epithelium. Thetracheae have the usual epithelium as far as I can determine,but sometimes against their sides on a fresh muscle-fibre theseare very minute (1-6 fx diameter) globules of neutral fat. Promthe arrangement of the globules and from what can be seenabout them, a very delicate substance seems to hold them in itsmeshes, and it seems that this substance is the tracheal epithe-lium more or less dislocated during the examination of the fibre.Kolliker (1888) and Jordan (1920 a) mention that fat is foundin the thoracic muscles of some insects, but their accounts arenot detailed, and they do not suffice to explain how the fatcomes to lie in this position in an adult bee. The fat does notreach its position through escaping from other tissues, for it isnot present in muscles, sarcosomes, nerves, or in such quantityin blood, and these with the tracheae are all the tissue foundbetween fibrous muscle. It shows its fatty nature and neutralityin Sudan III, scarlet red, Nile blue sulphate and chloride, andosmic-acid vapour.
The reader is asked to consult what was written about theinnervation of somatic muscles in the earlier part of this paper(Morison, 1927). The innervation of fibrous muscle is very diffi-cult to study. Snodgrass figures the bases of some of the largernerves which send branches to fibrous muscles, but nobodyseems to have attempted more than this for the bee. In factthere does not seem to be a published description of the com-plete innervation of fibrous muscles of the thorax in any insect.

636 GUY D. MOBISON
It will be seen from the account below that our knowledge ofthe innervation of the chief muscles of flight in the bee is stillvery incomplete.
Using alcohol-fixed material I managed to follow the branchesof apparently all the larger nerves supplying the two pairs offibrous muscles. The branches are very long and delicate, andthey seem to be perfectly constant in position for the threecastes, though the point at which they start branching from themain ganglion is not so fixed. All the branches arise from thesecond thoracic ganglion or the nerves just anterior to it. Bachlongitudinal muscle has on its inner face near the middle anerve passing dorsad and across the axes of its fibres. This nervepasses just laterad of the aorta, and it arises as a branch froma nerve of the anterior part of the ganglion. It gives off threepairs of branches which run cephalad or caudad along the fibresbetween which they enter. On the outer face there are threenerves : one near the posterior end curves upwards and branchesamong the fibres of the posterior dorsal region ; the secondpasses cephalad and dorsad and is the largest nerve. It gives offsmaller branches at intervals. The third is a small nerve passingdorsad at the interior. The first and second seem to have acommon origin from the mid-lateral region of the ganglion, andthe foremost nerve is a branch of a large anterior nerve, W2Nv(Snodgrass). Each vertical muscle has on its internal face manylong, delicate branches passing more or less dorsad from a smallnerve given off from, near the base of WzNv (Snodgrass).Another small nerve arising from the same place runs ventradand caudad and almost encircles the muscle whilst supplyingbranches to its ventral parts. The innervation so far describedseems an adequate supply for the muscles except that the upperportion of a vertical muscle has no nerves on its exterior lateralsurface. Now the nerves W2Nv (Snodgrass) are conspicuousand curve up towards this region from the anterior and theposterior sides of the muscle, but they supply the tubularmuscles of the wings. Yet it seems possible that from theirupper portions delicate branches might enter the fibrous muscle,though I have not seen such branches.

MUSCLES OF THE BEE 637
Oajal (1890) concluded after examining a number of insectsthat as he did not see Doyere's hillocks on fibrous muscle-fibres,such nerve-endings were probably not present, especially asGolgi's method of colouring revealed that there was a plexusof nerve-fibres and multipolar nerve-cells round each fibre.However, a union between the plexus and the central nervous
TEXT-FIG. 37.
Nerve-ending on fibrous muscle-fibre. Stained blue with Giemsa'sstain. Lettering : fi, fibrils ; nv, nerve ; tr, trachea.
system was not seen definitely. All my efforts at demonstrationof the nerves by this method were failures, but success may awaitfurther trials. I have examined very carefully many thousandsof fibres after they were treated in many different ways to shownerve-tissue, but out of the total number I saw only six fibreswith exceedingly delicate nerve-tissue attached to them. Fromthe little that I have seen, it appears that the long, delicatenerves described above divide into more delicate branches which

638 GUY D. MORISON
finally reach the width of 3/x. These branches anastomose, butI do not know to what extent. They rest at any point alongthe length of a fibre which they do not penetrate. At the pointof branching or anastomosis there is usually a small nucleus(5-3:3:3ft) lying in an expansion of the nerve. The nucleusseems to belong to the neurilemma. Only in one case (Text-fig. 37) did I see a cell which looked like a multipolar nerve-cell.
TEXT-FIG. 38.
sc nu
trch.
Reconstruction of a portion of a fibre showing the probable relation-ship between fibrils, sarcosomes, and traoheoles during a periodof slight contraction. Lettering : fi, fibril; M, mesophragma ;nu, nucleus ; sc, sarcosome ; trch, tracheole; Z, telophragma;
This cell was in contact with the nerves and seemed to representthe type of innervation described by Cajal for various insects.
Text-fig. 38 is drawn to show the structure of a portion ofa fibrous muscle-fibre as it would probably exist when in a stateof slight contraction during life. It is a reconstruction based onthe different methods of staining required to show each of theconstituents of the fibre, and it is never seen in all its detail inany one microscopical preparation.

MUSCLES OF THE BEE ' 639
On t h e A t t a c h m e n t of Musc le .
Only after a muscle-fibre is attached between at least twopoints is its essential power of contractility of any use to the bee.The different morphological types of muscle will have theirmethods of attachment described in turn, but that of thesomatic fibrous muscles will be mentioned at the end.
As stated previously the fibres of a somatic tubular muscleusually have a firm origin from the cuticle and an insertion ona chitinous tendon. Essentially a fibre consists of fibrils as longas itself, and since its contractile power lies in the fibrils thequestion to answer is how a fibril is attached to the chitin.
The lumen of tubular fibres is almost invariably closed at theend when the fibre is attached to a delicate tendon. It is openless rarely when the fibre is attached to strong cuticle (Text-fig. 40). The closure is affected by the gradual convergence ofthe fibrils towards the axis of the fibre. The diameter of thelumen near the attachment of the fibre will vary with the degreeof contraction of the fibre at the same region, though its diameterat the actual point of attachment is fixed.
In sections of fibres prepared by the usual methods of fixationand staining, the fibrils will be seen clearly to pierce the hypo-dermis and to be attached directly to the cuticle (Text-figs. 39,40). However, this is not the case. Each small portion of thebee differs from the rest in its chemistry, but a special treat-ment is often needed to render the difference visible. Each fibrilis attached to the cuticle by a very short zone (confined to theterminal sarcomere) which is apparently transitional betweencontractile sarcoplasm and chitin. I do not know of a reallysatisfactory method of demonstrating the transitional zone.After the tissue has been stained black in iron haematoxylin, analcoholic solution of bleu de Lyon will colour the zone bluewhilst it stains delicate chitin bright blue. The method of silverimpregnation which shows the boundaries of cells is also useful.The other methods of testing and staining pure chitin are diffi-cult to interpret or they injure or destroy the muscle elements.
The transitional zone does not seem greatly concerned withNO. 284 T t
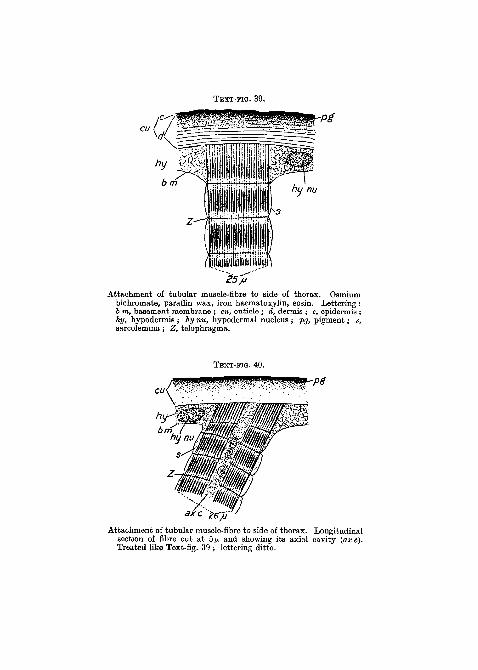
Attachment of tubular muscle-fibre to side of thorax. Osmiumbichromate, paraffin wax, iron haematoxylin, eosin. Lettering:6 m, basement membrane ; cu, cuticle ; d, dermis ; e, epidermis ;hy, hypodermis ; hy nu, hypodermal nucleus ; pg, pigment; s,sarcolemma; Z, telophragma.
TEXT-FIG. 40.
Attachment of tubular muscle-fibre to side of thorax. Longitudinalsection of fibre cut at 5/x and showing its axial cavity (axe).Treated like Text-fig. 39 ; lettering ditto.

MUSCLES OF THE BEE 641
the contraction of the fibre, but it may be obscured when thecontraction occurs near it. The fibre seems more deeply buriedin the hypodermis than the length of the zone (Text-fig. 40).The hypodermal nuclei are large and easily seen. Among themoccur some larger solitary nuclei which are concerned with thehairs of the body. On the tendons and underlying the cuticlethe hypodermis consists of a distinct single layer of squamousepithelial cells which have the long axes of their ellipsoidal orcircular nuclei parallel to the chitin they envelop. When anumber of muscle-fibres are attached to the tendon or cuticlethe hypodermal nuclei are scattered between the bases of thefibres, but it is impossible to determine the lateral boundaries ofthe cells, so the hypodermis may be a syncytium in these parts.Lying above the hypodermis is a very delicate membrane whichis the basement (? endothelial) membrane. The sarcolemma ofthe fibres seems to pass into this membrane (Text-figs. 39, 40)when the fibres are attached to tough cuticle.
A complication now arises. The sarcomeres when in theirresting position are about equal in length and are at right anglesto the long axis of the fibres, so that when both ends of attach-ment of a fibre are parallel to one another all the sarcomeres arecomplete units. But a fibre may have one or both its ends moreor less oblique to its long axis, and in these cases the end sarco-meres perhaps are not complete units ? I cannot decide on thispoint. In many cases the terminal telophragmata are parallelto the cuticle, and the middle sarcomeres are not all at rightangles to the long axis of the fibre.
The attachment of a muscle-fibre to a delicate tendon isessentially similar to its attachment to tough chitin. Thedifference is that the axial fibrils may be a little longer thanthe peripheral fibrils, but they are not long enough to lie ina distinct cup of the tendon. The sarcolemma passes graduallyinto the tendon, and it probably unites with the basementmembrane. At any rate the sarcolemma, the transitional zone,the delicate chitin of the tendon, and the basement membrane(when visible) become indistinguishable from one another.
Those muscles of the alimentary canal which are like somaticT t2

642 GUY D. MORISON
tubular muscles are affixed like them when attached to chitin.When they resemble splanchnic tubular muscles they areattached like the latter. The splanchnic tubular muscles formrings or anastomosing networks over the gut, and are held inplace by their inter-relation with one another, and by theclosely applied tracheae and nerves. They do not seem to beactually affixed to the gut, and, as stated previously, a definiteendothelial membrane cannot be seen covering them.
The cardiac muscle-fibres form semicircles on either side ofthe heart. Their sarcolemma seems to be bound totally withthe intima and adventitia of the heart. At the ends the fibrilsseem to terminate like those of somatic tubular muscle ontendon, but it is not possible to state the facts definitely. The alarymuscle-fibres just beneath the heart pass almost imperceptiblyinto a non-contractile membrane. At the sides of the body theyare attached to the cuticle partly by the membrane of the peri-cardial septum and partly after the manner of somatic tubularmuscles. They are attached along their course to the pericardialseptum by their sarcolemma. The ventral diaphragm is attachedto the cuticle either like somatic tubular muscle-fibres, or bymeans of delicate, tough fibres which seem to be the sarco-lemmata not yet sufficiently strengthened to be called a tendon.The striation disappears gradually as the fibres get narrowerand pass into the membranous fibre. In the heart and dorsaldiaphragm, and to a less extent on the ventral diaphragm,the tracheal system probably helps in supporting the muscle-fibres.
When muscles of the type of somatic tubular muscles areattached to the reproductive organs they are fixed to chitinin their typical way. The other muscles on the reproductiveorgans depend on their anastomoses with one another, theirtwisting, the support given by the tracheae and the nerves and,in the drone, the endothelial membrane.
Fibrous' muscle-fibres are attached to chitin essentially in thesame way as tubular muscle-fibre. The chief difference lies inthe much larger size of the fibrous muscle-fibre, which necessi-tates a correspondingly larger area of attachment on the chitin

MUSCLES OF THE BEE 643
and so introduces complications. One complication lies in theposition of the hairs, and another in the accommodation forhypodermal nuclei.
The vertical indirect muscles of the wings are attached tochitin which bears numerous hairs of two kinds, and the longi-tudinal muscles are attached cephalad to a large area of hair-bearing chitin and a small area of hairless chitin, whilst caudadthey are united to hairless chitin. The attachment to hair-bearing and hairless chitin is similar, but when hairs are present
TEXT-FIG.
•jsa
Bg
41.
Attachment of fibrous muscle to the dorsal wall of the thorax.Osmium bichromate, paraffin wax, cut 3^, iron haematoxylin,bleu de Lyon. Lettering : av, alveolus of hair ; c«, cuticle ;d, derrais ; e, epidermis ; fi, fibril; hr, hair; hy, hypodermis,with nucleus hy nu ; pg, pigment; sc, sarcosome ; //, tonofibrils ;tri, trichogenous cell.
over the part of attachment of a fibre the muscle fibrils have tobe grouped together so as to avoid the cavity in the cuticle atthe base of the hair. There is no very obvious grouping of thefibrils to accommodate the hypodermal nuclei or the nuclei ofthe fibre itself.
Text-fig. 41 shows the insertion of fibrous muscle on thecuticle. The fibrils lose their striation as they enter the hypo-dermis, and each fibril splits into about eight delicate rods,t o n o f i b r i l l a e (mihi nee Snodgrass), which from their

644 GUY D. MOBISON
staining reactions seem like the transitional zone of tubularmuscle fibrils. The tonofibrillae are attached to the stratifiedcuticle, which they do not seem to enter. The hypodermal nucleiare scattered amongst the bases of the fibrils. By most methodsof staining they stain more sharply than the nuclei of the largeand small hairs. The latter generally stain a diffused darkcolour, and both these types stain differently from the musclenuclei. Sections to show tonofibrillae should be cut at 3 /x (or less)and stained black in iron haematoxylin and then stained inbleu de Lyon.
Both tubular and fibrous muscle-fibres are attached to thechitin by a region apparently transitional between sarcoplasmand chitin. Each fibril of tubular muscle seems to undergo thischange in composition as it approaches the cuticle. Each so-called ' fibril' of fibrous muscles splits into a number (abouteight) of tonofibrillae which seem identical with the transitionalregion of tubular fibrils. A fibrous fibril can sometimes beisolated (Text-fig. 28) showing the free tonofibrillae at its end.The question now arises whether the tonofibrillae of fibrousmuscle ' fibrils ' are homologous with the transition zone oftubular muscle-fibrils, and if so whether the fibrous muscle-' fibrils ' should be regarded as a f ibre in which the fibrils arevery completely fused except at the ends. Some observersconsider this to be the case after examining fibrous musclesof other insects. The diameter of the tonofibrillae and thetubular muscle-fibrils is about the same (0-4/x). However,I have not seen any other signs of a longitudinal splittingof the fibrous fibril, so I shall leave the problem as it standsat present.
In the bee it seems probable that the transitional zonebetween muscle and cuticle for both tubular and fibrous muscleswill prove to be derived from the hypodermis. Berlese (1909)figures and describes this region as derived from the elongatedcells of the hypodermis in the developing pupa of a Hymano-pteron. Janet (1895,1895 a) writes that the tendons of attach-ment for tubular muscle-fibres in the bee are hypodermal inorigin, and that they tend to form cornet-shaped cups into

MUSCLES OF THE BEE 645
which the muscular portion of the fibre fits. He does not explainwhat happens to the sarcolemma at the place of insertion, themethod of attachment of the ultimate contractile elements, howthe nuclear axis terminates or the details of the relation of themuscle-fibre to the hypodermis, and he omits to describe theattachment of fibrous muscle. What he describes and figures asterminal ' cornets ' in the bee really pass imperceptibly from
, chitin into the sarcolemmata of the fibres.Different views have been or are held on the attachment of
muscle to chitin in insects. One view is that muscle is attachedby means of the hypodermis. Beside Berlese and Janet, whomay be considered as holding this opinion in the instancesquoted above, Stamm (1909), after working on Dipterouslarvae and adults, considers that insertion is always indirect,and Biley (1908) from the examination of an Orthopteron, adragon-fly, and some other insects, takes the view that fibresare attached through the intermediary of hypodermal cells, buthe does not accept Snethlage's (1905) view that the muscle-fibres of Arthropods are structurally prolongations of the chitino-genous cells. The view that muscles are inserted directly on thecuticle finds adherents in Snethlage, and in Jordan (1919), whoconsidered that muscle-fibres in a mantis were inserted directlyon the cuticle. Holmgren, N. (1910), from observations onDipterous larvae, considers that muscle may be inserted directlyon the cuticle or by the intervention of a hypodermis. Snodgrass(1925) does not describe in detail the attachment of the fibresin the bee. My findings do not agree completely with his. •
SUMMARY.
1. The entire musculature of the alimentary canal is describedin gross and in histological detail. The development of themuscle is considered. The innervation is described, likewise thetracheation and its relation to muscular activity and the blood-stream.
2. The heart is described with a detailed histological accountof its muscle-fibres. Its tracheation is described and its apparentlack of innervation is discussed.

646 GUY D. MOEISON
3. The ' alary ' muscles of the dorsal diaphragm are describedwith a detailed account of their histology, innervation, andtracheation.
4. The ventral diaphragm is described as well as the histology,innervation, and tracheation of its muscle-fibres. The course ofblood and physiological questions connected with it receivediscussion.
5. The muscles of the reproductive organs of drone, queen,and worker are described with particular reference to thehistology, innervation, tracheation, and physiology of theirfibres.
6. The indirect muscles of the wings (fibrous muscle) havetheir histology, innervation, and tracheation described in detail.The method of contraction of the entire muscles and of theindividual fibres and fibrils is discussed. The sarcosomes aredescribed with their physiological significance to contraction.
7. The attachment of all the types of muscle found in the beeis described in histological detail. Different opinions of muscleattachment to chitin are summarized.
8. Throughout the paper, histological measurements are givenfor the various types of muscle-fibres and their nuclei in the threecastes of bee. Since in the three castes the histological appear-ance is so similar for each type of muscle, the illustrations havebeen limited to portions of the muscles of worker bees.
BlBLIOGBAPHY.
Adam, A. (1913).—" Bau und Mechanismus des Receptaculum seminis beiden Bienen, Wespen und Ameisen ", ' Zool. Jahrb. Anat. u. Ontog.', 35,S. 1-74, Taf. 1-3.
Arnhart, L. (1906).—" Die Bedeutung der Aortensohlangenwindungen desBienenherzens ", ' Zool. Anz.', 30, S. 721-2.
Berlese, A. (1909).—' Gli Insetti', vol. i. Milan.Bishop, G. H. (1920).—" Fertilization in the Honey-bee. I. The male
sexual organs : their histological structure and physiological function-ing ", ' Joum. Exper. Zool.', 31, pp. 225-66, Pis. 1-3.
(1920 a).—" II. Disposal of the sexual fluids in the organs of thefemale ", ibid., pp. 267-86, 2 figs.
Bresslau, E. (1905).—" Der Samenblasengang der Bicnenkonigin ", ' Zool.Anz.', 29, S. 299-323, 7 Abb.

MUSCLES OF THE BEE 647
Brunnifih, K. (1911).—"Die Muskeln bei der Biene ", ' SchweizerischeBienenztg.', 7, S. 277-83, figs. 52-60.
Biitschli, 0., und Schewiakoff, W. (1891).—" Ueber den feineren Bau derquergestreiften Muskeln von Arthropoden", ' Biol. Zentralbl.', 11,S. 33-9, figs. 1-7.
Cajal, S. Ramon y (1890).—" Coloration par la methode de Golgi des ter-minaisons des trachees et des nerfs dans les muscles des ailes des in-sectes ", ' Zeits. wiss. Mikr.', 7, pp. 332-42, PL ii, 3 figs.
Cheshire, F. R. (1885).—" The Apparatus for differentiating the sexes inBees and Wasps. An anatomical Investigation into the Structure of theReceptaculum Seminis and adjacent parts ", ' Journ. Roy. Micr. Soc.',Ser. ii, v, pp. 1-15, Pis. i-ii.
Demoll, R. (1926).—"Die Atmung der luftatmenden Insekten ", ' Zool.Anz.', 69, S. 8-16.
Dufour, L. (1834).—" Recherches anatomiques et physiologiques sur leaOrthopteres, les Hymenopteres et les Neuroptdres ", 'Mem. presentedpar divers savants a l'Acad. Roy. des Sci. de l'Inst. de France', vii,pp. 265-647, 13 pis., 1841.
Ebner, V. E. v. (1918).—" Uber den feineren Bau der Flugelmuskelfasernder Insekten ", ' Sitz.-Ber. Ak. Wiss. Wien, Math.-nat. Kl.', Abt. iii,Bd. 127, S. 3-32, Taf. 1.
Evenius, J. (1925).—" Die Entwicklung des Zwischendarms der Honig-biene ", ' Zool. Anz.', 63, S. 49-64.
Evenius, Christa (1926).—"Der Verschluss zwischen Vorder- und Mittel-darm bei der postembryonalen Entwicklung von Apis mellifica L.",ibid., 68, S. 249-62, 10 figs.
Gehuchten, A. van (1886).—" Sur la structure intime de la cellule muscu-laire striee ", ' La Cellule ', 2, pp. 289-453, Pis. 1-6.
Girdwoyn, M. (1876).—' Anatomie et physiologie de l'Abeille ', 39 pp.,12 pis. French translation by Pillain, A.
Graber, V. v. (1873).—" Uber den propulsatorischen Apparat der In-sekten ", ' Archiv f. Mikr. Anat.', 9, S. 129-96, Taf. viii-x.
(1876).—" tlber den pulsirenden Bauchsinus der Insekten", ibid.,Bd. 12, S. 575-82, Taf. xxiv.
Griffiths, A. B. (1891).—" On the blood of Invertebrata ", ' Proc. Roy.Soc. Edinburgh', 18, pp. 288-94.
(1892).—Ibid., 19, pp. 116-30.Hardy, W. B. (1926).—" Properties of thin films ", ' Nature ', London, 118,
pp. 700-1, fig. 1.Hertig, M. (1923).—" The normal and pathological Histology of the Ven-
triculus of the Honey-bee, with special reference to infection with Nosemaapis ", ' Joum. Parasitology ', 9, pp. 109-40, Pis. ix-xi.
Holmgren, E. (1907).—"tlber die Sarkoplasmakorner quergestreifterMuskelfasern ", ' Anat. Anz.', 31, S. 609-21, Taf. 2.

648 GUY D. MOEISON
Holmgren, E. (1908).—" Uber die Trophospongien der quergestreiftenMuskelfasern, nebst Bemerkungen iiber den allgemeinen Bau dieserFasern ", ' Archiv f. Mikr. Anat.', 71, S. 165-247, Taf. xiii-xx.
(1910).—" Untersuchungen iiber die morphologisch nachweisbarenstofflichen Umsetzungen der quergestreiften Muskelfasern ", ibid., Bd. 75,S. 240-336, Taf. viii-xiii.
Holmgren, N. (1910).—" Uber die Muskelinsertionen an das Chitin bei denArthropoden ", ' Anat. Anz.', 36, S. 116-22.
Imms, A. D. (1925).—'A General Text-book of Entomology'. Methuen& Co., London.
Janet, C. (1895).—" Sur les muscles des Fourmis, des Guepes et desAbeilles ", ' C. R. Acad. Sci. Paris ', 121, pp. 610-13, figs.
(1895 a).—" Structure des membranes articulaires des tendons et desmuscles ", ' Etudes sur les Fourmis, les Guepes et les Abeilles ', 12me
Note, 25 pp., 11 figs., Limoges.(1911)..—"Sur l'existence d'un organe chorodontal et d'une vesieule
pulsative antennaire chez l'Abeille et sur la morphologie de la tete decette espece ", ' C. R. Acad. Sci. Paris ', 128, pp. 249-53, 2 figs.
Janisch, E. (1924).—" Zum Bau der quergestreiften und glatten Muskel-siiulchennach Untersuchungen an Bombus terrestris und Helix pomatia ",' Anat. Anz.', 57, S. 246-55.
Jordan, H. E. (1919).—" Studies on striped Muscle Structure. V. Thecomparative Histology of the leg and wing muscles of the Mantis, withspecial reference to the N-discs and the sarcosomes ", ' Anat. Record ',16, pp. 217-46, 3 pis.
(1920).—" Studies, &c. VI. The comparative Histology of the legand wing muscles of the Wasp, with special reference to the phenomenonof stripe reversal during contraction and to the genetic relation betweencontraction bands and intercalated discs ", ' Amer. Journ. Anat.', 27,pp. 1-67, 48 figs.
(1920a).—"Studies, &c. VII. The development of the sarcostyleof the wing muscle of the Wasp, with a consideration of the physico-chemical basis of contraction ", ' Anat. Record', 19, pp. 97-123, 2 pis.
Kahn, R. H. (1916).—" Zur Physiologie der Insektenmuskeln ", Pfliiger'sArchiv f. Physiol., 165, S. 285-336, 31 figs.
Keilin, D. (1925).—" On Cytochrome, a Respiratory Pigment, Common toAnimals, Yeast, and Higher Plants ", ' Proc. Roy. Soc. London ', Ser. B,98, pp. 312-39.
(1926).—" A Comparative Study of Turacin and Haematin and itsbearing on Cytochrome ", ibid., vol. 100, pp. 129-51.
Kielich, J. (1918).—" Beitrage zur Kenntnis der Insektenmuskeln ", ' Zool.Jahrb., Abt. Nat. Ont.', 50, S. 515-36, Taf. 25-6.
Kolliker, A. (1888).—" Zur Kenntnis der quergestreiften Muskelfasern ",' Zeits. wiss. Zool.', 47, S. 689-708, Taf. 14-15.

MUSCLES OF THE BEE 649
Koschewnikoff, G. (1891).—" Zur Anatomie der mannlichen Geschlechts-organe der Honigbiene ", ' Zool. Anz.', 14, S. 393-6.
Leydig, F. (1859).—" Zur Anatomie der Insekten ", Miiller's Archiv f.Anat. u. Physiol. u. wiss. Med., S. 33-89 u. 149-83, Taf. 2-4.
(1876).—" Bemerkungen iiber die Hautdecke und Nerven der Driisenbei Insecten ", ' Archiv f. Mikr. Anat.', 12, S. 536-50, Taf. xxiii.
Loschel, F. (1916).—" III. Die postembryonale Entwicklung des Ge-schleohtsapparates der Bienenkonigin (Apis mellifica L.) ", ' Zeits.angew. Entom.', 3, S. 21-44, Taf. iii-iv u. 5 Abb.
MaGallum, A. B. (1905).—" On the Nature of the Silver Reaction in Animaland Vegetable Tissues ", ' Proc. Roy. Soc. London ', Ser. B, 76, pp. 217-29.
Marcus, H. v. .(1922).—" Weitere Untersuchungen iiber den Bau quer-gestreifter Muskeln ", ' Anat. Anz.', 55, S. 475-97, 8 Abb.
Mclndoo, N. E. (1916).—' The sense organs of the mouth-parts of the Honey-bee ', Smithsonian Misc. Coll. 65, no. 14, pp< 1-55, 10 figs.
Meier, K. (1916).—" IV. Die postembryonale Entwicklung des Geschlechts-apparates der Arbeitsbiene (Apis mellifica L.) ", ' Zeits. angew. Entom.',3, S. 45-65, Taf. v-vi u. 3 Abb.
Merkel, F. (1872).—" Die quergestreifte Muskel. Das primitive Muskel-element der Arthropoden", ' Archiv f. Mikr. Anat.', 8, S. 244-68,Taf. xiii.
Metzer, C. (1910).—" Die Verbindung zwischen Vorder-und Mitteldarm beider Biene ", ' Zeits. wiss. Zool.', 96, S. 539-71, Taf. xxiv-xxv.
Morison, G. D. (1927).—" The Muscles of the Adult Honey-bee (Apismellifera L.). Part I. The Healthy Muscles of the Adult Honey-bee.Somatic Musculature", 'Quart. Journ. Micr. Sci.', 71, pp. 395-463,12 figs.
Nabert, A. (1913).—" Die Corpora allata der Insekten ", ' Zeits. wiss. Zool.',104, S. 181-358, 8 Abb., Taf. viii-xii.
Nath, Vishwa (1926).—" On the present position of the Mitochondria andthe Golgi Apparatus ", ' Biol. Rev. Cambridge Phil. Soc.', ii, pp. 52-79.
Nelson, J. A. (1915).—' The Embryology of the Honey-bee ', PrincetonUniversity Press, 282 pp., 95 figs., 6 pis.
(1924).—"Morphology of the Honey-bee Larva", 'Journ. Agric.Res.', 28, no. 12, pp. 1167-213, 8 pis., 5 text-figs. Washington, D.C.
Nolan, W. J. (1924).—" The Brood-rearing Cycle of the Honey-bee ",' U.S. Dept. Agric, Dept. Bull.', 1349, pp. 1-56.
Orlov, J. (1924).—" Die Innervation des Darmes der Insekten (Larven vonLamellicorniern) ", ' Zeits. wiss. Zool.', 122, S. 425-502, 15 Text-figs.,Taf. xi-xiii.
Pavlovsky, E. N., and Zarin, E. J. (1922).—" On the Structure of theAlimentary Canal and its Ferments in the Bee (Apis mellifera L.) ",' Quart. Journ. Micr. Sci.', 66, pp. 509-56, Pis. 15-17.

650 GUY D. MOEISON
Petersen, H. (1912).—" Beitrage zur vergleichenden Physiologie der Ver-dauung. V. Die Verdauung der Honigbiene", Pfliiger's Archiv f.Physiol., 145, S. 121-51, Taf. 1-2.
Pissarew, W. J. (1898).—" Das Herz der Biene " , ' Zool. Anz.', 21, S. 282-3,1 Abb.
Pixell-Goodrich, Helen L. M. (1920).—" Determination of Age in Honey-bees ", ' Quart. Journ. Micr. Sci.', 64, pp. 191-206, PI. 2.
Poletajewa, Olga (1886).—" Du ccaur des Insectes ", ' Zool. Anz.', 9,pp. 13-15.
Pruthi, Hem Sing (1925).—" Homologies of the Genital Ducts of Insects ",' Nature ', London, 115, p. 763.
Riley, W. A. (1908).—"Muscle attachments in insects", 'Ann. Entom.Soc. Amer.', 1, pp. 265-72, PI. 24.
Sohindler, E. (1878).—" Beitrage zur Kenntnis der Malpighi'schen Gefasseder Insecten ", ' Zeits. wiss. Zool.', 30, S. 587-660, Taf. 38-40.
Schbnfeld, P. (1886).—" Die physiologische Bedeutung des Magenmundesder Honigbiene ", ' Archiv f. Anat. u. Physiol. (Physiol. Abt.)', S. 451-8.
Sihler, C. (1895).—" tJber eine leichte und sichere Methode, die Nervenendi-gung an Muskelfasern und Gefassen nachzuweisen", ibid., S. 202-8,3 Abb.
Snethlage, E. (1905).—" Ueber die Frage vom Muskelansatz und die Her-kunft der Muskulatur bei den Arthropoden ", ' Zool. Jahrb., Abt. f.Morphol.', 21, S. 495-514, Taf. 29-30.
Snodgrass, R. E. (1925).—' Anatomy and Physiology of the Honey-bee', pp. xvi + 328, 108 figs. McGraw-Hill Book Co., New York andLondon.
(1926).—" The Morphology of Insect Sense Organs and the SensoryNervous System ", Smithsonian Misc. Coll. 77, No. 8, Publication 2831,80 pp., 32 figs.
Stamm, R. H. (1909).—" Ueber die Muskelinsertionen an das Chitin beiden Arthropoden ", ' Anat. Ariz.', 34, S. 337^t9, 7 Abb.
Stiibel, H. (1920).—" Mikroskopisch wahrnehmbare Veranderungen derQuerstreifung des Muskels nach Versuchen am Frosch- und Insekten-muskal ", Pfliiger's Archiv f. Physiol., 180, S. 209-49, 4 Abb.
Svartz, N. (1914).—" Studien iiber quergestreifte Muskulatur beim Men-schen mit besonderem Bezug auf die Nahrungsaufnahme der Muskel-fasern ", ' Anat. Anz.', 45, S. 538-48, 5 Abb.
Thulin, I. (1914).—" 1st die Grundmembrane eine konstant vorkommendeBildung in den quergestreiften Muskelfasern ? " ' Archiv f. Mikr. Anat.',86, S. 318-37, Taf. xi. 4 Abb.
Trappmann, W. (1923).—"Die Malpighischen Gefasse von Apis mellificaL.", 'Archiv f. Bienenkunde ', 5. Jahrg., S. 177-89, 9 Abb.
" Anatomie und Physiologie des Zwischendarmes von Apia mellificaL.", ibid., S. 190-203, 3 Abb.

MUSCLES OF THE BEE 651
Walling, Eulalia, V. (1906).—" The influence of gases and temperature onthe cardiac and respiratory movements in the Grasshopper ", ' Journ.Exper. Zool.', 3, pp. 621-9.
White, G. F. (1918).—" A note on the muscular coat of the ventriculus ofthe Honey-bee (Apis mellifica)", ' Proc. Entom. Soc. Washington', 20,pp. 152^, fig. 1.
Wolff, 0. J. B. (1876).—" Das Biechorgan der Biene nebst einer Beschrei-bung des Bespirationswerkes der Hymenopteren, des Saugriissels undGeschmackorganes der Blumenwespen ", ' Nova Acta Acad. C. Leop.-Carol. Nat. Cur.', 38, S. 1-254, Taf. i-viii. Dresden.
Zander, E. (1900).—" Beitrage zur Morphologie der mannliohen Ge-schlechtsanhange der Hymenopteren " , ' Zeits. wiss. Zool.', 67, S. 461-89,9 Abb., Taf. xxvii.
(1922).—" Bau der Biene ", 2. Aufl. (' Handb. d. Bienenkunde ', iii,Ulmer, Stuttgart).


















