
THE MULTIPLE MOLECULAR FORMS OF …
226
THE MULTIPLE MOLECULAR FORMS OF ACETYLCHOLINESTERASE IN SKELETAL MUSCLES: A FUNCTIONAL ANALYSIS Josephine Yuen-Wai Lai A dissertation submitted to Imperial College of Science and Technology in candidature for the Diploma of Imperial College and to the University of London for the degree of Doctor of Philosophy. Department of Biochemistry Imperial College University of London London SW7 2AZ. August, 1985 -1-
Transcript of THE MULTIPLE MOLECULAR FORMS OF …

THE MULTIPLE MOLECULAR FORMS OF ACETYLCHOLINESTERASE IN SKELETALMUSCLES: A FUNCTIONAL ANALYSIS
Josephine Yuen-Wai Lai
A dissertation submitted to Imperial College of Science and Technology in candidature for the Diploma of Imperial College and to
the University of London for the degree of Doctor of Philosophy.
Department of Biochemistry Imperial College University of London London SW7 2AZ.
August, 1985
- 1 -

JOSEPHINE Y. W. LAI
THE MULTIPLE MOLECULAR FORMS OF ACETYLCHOLINESTERASE IN SKELETALMUSCLES: A FUNCTIONAL ANALYSIS
ABSTRACT
A number of molecular forms of acetylcholinesterase (AChE) from
vertebrate skeletal muscles have been identified on sucrose density
gradients. The heaviest form, H2 C, is concentrated at the neuromuscular
junction of twitch muscle fibres, while the light (L^ and L2 ) forms
predominate at the avian tonic fibre endplate. It is thought likely
that the distribution of these forms in muscles may reflect their
functions in different muscle types. However, previous studies on the
distribution of AChE forms in mammalian muscles had not been able to
show any clear relationship between their distribution, functions and
fibre-type-specific localization. Furthermore, the proportion of the H2 C
form relative to other forms is small, which does not reflect its
presumed physiological importance. H2C AChE disappears after chronic
denervation and reappears upon reinnervation, and thus is clearly under
neural control. This study aims to study the relationship between the
distribution of AChE forms and muscle fibre types, as well as the
neurotrophic regulation which may contribute to such distribution.
In marked contrast to previous reports, it was found that H2 C
AChE and a significant H^c component predominate in most mammalian and
avian twitch muscles, and which are the only forms located at the
endplate. L and M forms are confined to the extrajunctional regions, the
variations in their concentrations contribute to the overall variability
in the distribution of AChE forms. Moreover, their contribution may be
overestimated if proteolytic degradation of the H AChE was not
adequately prevented. Slow-twitch fibres contained extrajunctional H2 C
which was also observed in fast-twitch muscle subjected to continuous

low frequency stimulation. On chronic denervation, the endplate H AChE
decreased considerably, while the extrajunctional AChE forms, including
the H AChE, accounted for the overall increase in AChE activity in some
species. It is concluded therefore that endplate H AChE is maintained by
innervation and is by far the major form at the endplate, while the
extrajunctional AChE is probably regulated by muscle activity.
Monoclonal antibodies specific for avian AChE had been raised from
purified chick brain AChE. These antibodies are now used as specific
affinity ligands for the purification of the avian H2 C form.
- i l l -

Preface
The work presented in this dissertation was performed between
October, 1981 and April, 1985 in the Department of Biochemistry,
Imperial College, London. All the work described herein is my own except
where otherwise indicated in the text.
I am deeply grateful to my supervisor, Professor E.A. Barnard,
F.R.S., for his encouragement and advice throughout the course of this
work. I am also indebted to Dr. William R. Randall for his technical
supervision, much inspiring discussion, and for his constructive
criticism of several chapters in this thesis. I wish also to record my
special thanks to Dr. Penelope J. Barnard, Dr. J. Jedrzejczyk, Dr. John
A. Pizzey, Dr. Rudolf Shipolini, Mr. Jonathan Jarvis, Mr. David Green,
Mr. John F.R. Cavanagh, Mr. Anthony Lai and Dr. David Lane for their
invaluable assistance and enjoyable collaboration. I also thank Miss
Alison Bartlett for typing this thesis. Finally, my gratitude goes to
all my colleagues for their friendship and generosity, who have made the
past 3 years a fruitful and memorable experience. I am also grateful to
the Croucher Foundation for generous financial support during the year
83/84.
- 1 v -

To my parents and to Frank

C O N T E N T S
page
Abstract nPreface ivList of abbreviations xList of figures xiList of tables xm
CHAPTER 1
GENERAL INTRODUCTION
1.1 Acetylcholinesterase 21.1.1 Enzyme properties 21.1.2 Inhibition of cholinesterases 41.1.3 Molecular structure of AChE: multiple molecular forms 51.1.4 Localisation of AChE 121.1.5 Purification of AChE 151.1.6 Immunological studies of AChE 16
1.2 Classification of skeletal muscle fibre types 161.2.1 Physiological characterisation 171.2.2 Histochemical and biochemical characterisation 181.2.3 Morphological and uItrastructuraI characterisation 191.2.4 Fibre type composition of muscles 19
1.3 Skeletal muscle AChE in health and disease 211.3.1 Biosynthesis of AChE in skeletal muscles 211.3.2 Neurotrophic regulation of AChE 231.3.3 Avian muscular dystrophy 251.3.4 This thesis 27
•
CHAPTER 2
MULTIPLE MOLECULAR FORMS OF AChE IN MAMMALIAN AND AVIAN SKELETAL MUSCLES
2.1 Introduction 302.2 Materials and Methods 36
2.2.1 Animals 362.2.2 Muscles 362.2.3 Materials 372.2.4 Precautions applied in tissue extraction 372.2.5 Microdissection 402.2.6 Sedimentation analysis 412.2.7 Enzyme assay 41
- V i -

2.3 Results 44
Section I: Effects of extraction conditions on the distribution of AChE forms
2.3.1 Protease activities in muscle extracts and the effects of 44protease inhibitors on AChE extraction
2.3.2 Extraction of AChE by rapid processing In the presence of 45protease Inhibitors
2.3.3 The effect of perfusion 532.3.4 The effect of freezing muscle tissues 53
Section II: Distribution of AChE forms in mammalian and avian skeletal muscles
2.4
2.5
2.3.5 Molecular profiles of AChE In mammalian skeletal muscles2.3.6 Molecular profiles of AChE in avian muscles2.3.7 Molecular profiles of AChE In muscles of mixed fibre type
Discussion
2.4.1 Autolysls of AChE In tissue crude extracts2.4.2 Correlation of the degradation of the H2C form with the
proteolytic activity In tissue crude extracts2.4.3 Inhibition of proteolysis stabilised the distribution of the
molecular forms of AChE2.4.4 Time-course and mechanism of AChE degradation2.4.5 H2c I s a dominant component of mammal Ian skeletal muscle ACHE2.4.6 AChE proflles In fast- and slow-twltch muscles2.4.7 AChE profiles In chicken twitch and tonic muscles2.4.8 Non-endplate AChE In type I and type II fibres2.4.9 Endplate AChE In twitch and tonic fibres
Conclusion
54636666
6670
71
72 75 75777879
80
CHAPTER 3
THE EFFECT OF CHRONIC DENERVATION ON THE ENDPLATE AND NON-ENDPLATE AChE IN HAMALIAN FAST- AND SLOW-TWITCH MUSCLES
3.1 Introduction 83
3.2 Materials and Methods 853.2.1 Animals 853.2.2 Muscles 863.2.3 Surgical procedures 863.2.4 Partial denervation 873.2.5 Muscle extract preparation 873.2.6 Measurement of endplate and non-endplate AChE 87
3.3 Results 88
3.3.1 Denervation-induced changes In AChE activities In different 88species
- V 1 1 -

3.3.2 Effects of denervation on the AChE profiles of fast and 92slow muscles
3.3.3 Differential analysis of endplate and non-endplate AChE in 100normal and denervated muscle fibres
3.4 Discussion 1003.4.1 Changes in AChE activities Induced by chronic denervation 1003.4.2 AChE in the endplates of normal and denervated muscle fibres 1053.4.3 AChE in the non-endplate regions of normal and denervated 106
muscle fibres3.4.4 Effect of partial and total denervation on the two regions of 108
semimembranosus
3.5 Conclusion 110
CHAPTER 4
CHANGES IN MOLECULAR FORMS OF AChE IN CHRONICALLY STIMULATED CHICKENMUSCLES
4.1 Introduction 1124.2 Materials and Methods 115
4.2.1 Nerve stimulation 1154.2.2 Extraction of AChE 1164.2.3 Sedimentation analysis 117
4.3 Results ii7
4.3.1 Changes in AChE activity in stimulated normal chicken PLD 117muscle
4.3.2 Separation of endplate and non-endplate forms of AChE 1224.3.3 The reversal of the effect of stimulation 1224.3.4 Changes in AChE activity in stimulated dystrophic chicken PLD 122
muscle4.3.5 Separation of endplate and non-endplate forms of AChE 1324.3.6 Changes in AChE activities in stimulated leg chicken muscles 132
4.4 Discussion 134
4.4.1 Changes of AChE in stimulated normal chicken PLD 134% 4.4.2 Changes of AChE in stimulated dystrophic chicken PLD 143
4.4.3 Changes of AChE in leg muscles of normal and dystrophic 146chickens
CHAPTER 5
MONOCLONAL ANTIBODIES TO PURIFIED CHICK BRAIN AChE
5.1 Introduction 1505.2 Materials and Methods 151
5.2.1 Materials 1515.2.2 Purification of chick brain AChE 152
- v i i i -

5.2.3 Enzyme assay 1545.2.4 Immunisation and cell culture 1545.2.5 Screening protocol 1565.2.6 Purification of monoclonal antibodies 1565.2.7 Gel electrophoresis and western blotting 1575.2.8 Immunohistochemistry 158
5.3 Results 158
5.3.1 Purification of chick brain AChE 1585.3.2 Characterisation of monoclonal antibodies to AChE 160
5.3 Discussion no
CHAPTER 6
GENERAL DISCUSSION 175
Appendix I: Rapid processing of skeletal muscle crude 181extracts
Appendix II: Molecular size determination of AChE by 183radiation inactivation
References 189
List of publications 209
*
- 1 x -

LIST OF ABBREVIATIONS
Abbreviations
ACB-1 Monoclonal antibody 1
ACB-2 Monoclonal antibody 2
ACh Acetylcholine
AChE Acetylcholinesterase
ALD Anterior latissimus dorsi
AT Anterior tibia1 is
ATPase ATP-phosphohydro1ase
BTEE Benzoyl-tyrosy 1 ethyl ester
Cat Cata1ase
Con A Concanavalin A
DFP Diisopropyl phosphorof1uoridate
DM EM Dulbecco's minimum essential medium
DTNB 5,51—dithiobis—(2-nitrobenzoic acid)
EDL Extensor digitorum longus
FITC Fluorescein isothiocyanate
3-gal 3-Galactosidase
HRP Horse radish peroxidase
ig Immunoglobulin
isoOMPA Tetraisopropy1pyrophosphoramide
LDH Lactate dehydrogenase
MAc N-methy1acr i d i n i urn
NEM N-ethyImaleimide
^ NP-40 Non i det P-40
ONPG O-nitrophenyIgalactosi de
PBS Phosphate buffered saline
PLD Posterior latissimus dorsi
PMSF PhenyImethy1su1fony1f1uoride
(pChE Pseudocholinesterase
RER Rough endoplasmic reticulum
SA Semimembranosus accessorius
SDS-PAGE Sodium dodecyl su1phate-Polyacrylamide gel electrophoresis
SP Semimembranosus proprius
SVI Vipera ammodytes venom trypsin inhibitor
TCA Trichloroacetic acid
- X-

List of figures Rage
Fig 1.1 Schematic representation of the hydrolysis of acetylcholine.
3
Fig 1.2 Hypothetical model of the binding of acetylcholine to the active site of AChE.
3
Fig 1.3 Schematic representation of the collagen tailed form of AChE from Electrophorus.
7
Fig 1.4 Schematic representation of the quaternary structure of the asymmetric and globular forms of AChE.
9
Fig 2.1 Casein hydrolysing activity of tissue crude extracts from various species.
46
Fig 2.2 Dose dependent inhibition of casein hydrolysing activity of rat muscle extracts by bacitracin, SVI and a2 -macroglobulin.
47
Fig 2.3 Dose dependent inhibition of bovine a-chymotrypsin activity by SVI.
48
Fig 2.4 Effect of prolonged storage of guinea pig muscle crude extracts on the distribution of the molecular forms of AChE.
50
Fig 2.5 Protease inhibitors stabilise the distribution of AChE forms in muscle crude extracts.
51
Fig 2.6 Typical sucrose gradient profiles of AChE forms in some mammalian type II muscles.
55
Fig 2.7 Typical sucrose gradient profiles of AChE forms in some mammalian type II muscles.
56
Fig 2.8 Typical sucrose gradient profiles of AChE forms in some mammalian type II muscles.
58
Fig 2.9 Typical sucrose gradient profiles of AChE forms in some mammalian type I muscles.
59
Fig 2.10 Molecular forms of AChE from type I and type II muscles fibres.
61
Fig 2.11 The distribution of molecular forms of AChE in the endplate and non-endplate regions from type I and type II muscles.
62
Fig 2.12 Molecular forms of AChE in the anterior and posterior regions of the chicken adductor profundus.
65
Fig 2.13 Sucrose gradient profile of AChE in the chicken PLD muscle.
67
Fig 2.14 Sucrose gradient profiles of AChE in some muscles of mixed fibre type composition.
68
Fig 2.15 Distribution of molecular forms of AChE in the endplate and non-endplate regions of the rat diaphragm.
69
Fig 3.1 Histochemical characterisation of contralateral normal and denervated rabbit semimembranosus muscle.
89
Fig 3.2 Sucrose gradient profiles of contralateral control and chronically denervated hamster EDL.
93
-XI -

94Fig 3.3 Sucrose gradient profiles of AChE in control and denervated type II muscles.
Fig 3.4 Sucrose gradient profiles of AChE in contralateral 95control and denervated type I muscles.
Fig 3.5 Sucrose gradient profiles of AChE in the endplate 101region of control and denervated guinea pig muscle fibres.
Fig 3.6 Sucrose gradient profiles of extrajunctional AChE in 102 control and denervated guinea pig muscle fibres.
Fig 3.7 Changes in the intensity of endplate AChE staining in 103 denervated guinea pig soleus.
Fig 4.1 Sucrose gradient profiles of AChE in normal PLD after 120different periods of stimulation.
Fig 4.2 Electron micrographs of PLD muscle from a 19-week old 123normal chicken.
Fig 4.3 Sucrose gradient profiles of endplate and non-endplate 125AChE in control and 3-week stimulated normal PLD.
Fig 4.4 Sucrose gradient profiles of AChE in dystrophic PLD 130after different periods of stimulation.
Fig 4.5 Frozen cross-section of PLD muscles from a 14-week 131old dystrophic chicken.
Fig 4.6 Sucrose gradient profiles of endplate and non-endplate 133AChE in control and 5-week stimulated dystrophic PLD.
Fig 4.7 Sucrose gradient profiles of AChE in leg muscles of 137normal chicken after chronic stimulation.
Fig 4.8 Sucrose gradient profiles of AChE in leg muscles of 138dystrophic chicken after chronic stimulation.
Fig 5.1 Sucrose gradient profile of AChE from crude extract 159of one day old chick brain.
Fig 5.2 Analysis of purified chick brain AChE using SDS-PAGE. 162
Fig 5.3 SDS-PAGE of purified ACB-1 and ACB-2. 162 iFig 5.4 Dilution curves of ACB-1 and ACB-2. 165 ;
Fig 5.5 Competition of ACB-1 and ACB-2 for epitopes of AChE. 167
Fig 5.6 Reactivity of ACB-1 to avian AChE by western blotting. 168
Fig 5.7 Immunohistochemical localisation of AChE in unfixed 171chicken PLD muscle using ACB-1 and ACB-2.
- x i i -

List of tables p a g e
Table 1.1 Sedimentation coefficients of the molecular forms of AChE in chicken and rat skeletal muscle.
11
Table 2.1 The fibre type composition of some mammalian and chicken skeletal muscles which had been chosen for AChE analysis.
33
Table 2.2 Combination of protease inhibitors used during AChE extraction from different tissues.
35
Table 2.3 AChE activity in some mammalian muscles. 64
Table 3.1 Relative activities of AChE in denervated rabbit, guinea pig and hamster muscles.
91
Table 3.2 Relative activities of L and H2c form in denervated rabbit, guinea pig and hamster muscles.
97
Table 3.3 Changes in total AChE and its molecular forms in the partially denervated rabbit semimembranosus.
98
Table 3.4 The relative activities (per muscle) of AChE and its 99molecular forms in the denervated guinea pig and rabbit semimembranosus.
Table 4.1 Increases in AChE concentration in the normal PLD after different periods of stimulation.
118
Table 4.2 Reversal of stimulation-induced changes in AChE after cessation of the stimulus.
127
Table 4.3 Decreases in AChE concentration in dystrophic PLD after different periods of stimulation.
128
Table 4.4 Fibre type composition of some normal and stimulated fast-twitch muscles in the lower limb of chicken.
135
Table 4.5 Increases in AChE concentration in leg muscles of normal and dystrophic chickens after different periods of stimulation.
136
Table 5.1 Purification of AChE from day old chick brain. 161
Table 5.2 Reactivity of ACB-1 and ACB-2 with <PChE and AChE from other species.
169
- X I 11 -

CHAPTER 1
GENERAL INTRODUCTION
-1-

1.1 Acetylcholi nesterase
1.1.1 Enzyme properties
The term acetylcholinesterase (AChE; E.C. 3.1.1.7) was first
introduced by Augustinsson and Nachmansohn (1949) to describe the
particular type of esterase shown to be associated with a variety of
conducting tissues (Nachmansohn and Rothenberg, 1944; 1945) and
erythrocytes (Stedman and Stedman, 1935; Hawes, 1940), and is distinct
from the other non-specific cholinesterases or 1 pseudocholinesterases1
( V'ChE; E.C.3.1.1.8) found in serum (Stedman et al., 1932). AChE
catalyses the hydrolysis of the neurotransmitter acetylcholine (ACh)
which is schematically shown in Fig 1.1. AChE has a high affinity for
its main substrate ACh. The optimal substrate concentration has been
well-defined and substrate inhibition at high ACh concentrations has
been shown to occur (Stedman and Stedman, 1935). Structure-substrate
specificity study showed that the rate of hydrolysis decreases with
increasing length of the acyl chain, i.e. in the order: acetyl- >
propionyl- > butyryl-choline (Nachmansohn and Wilson, 1951). In
contrast, ChE exhibits a reverse order of substrate preference.
Based on results from enzyme kinetic studies, a hypothetical
model of enzyme-substrate interaction was first proposed by Nachmansohn
and Wilson (1951). It was suggested that the catalytic site consists of
a negatively charged anionic and an esteratic subsite. The anionic
subsite binds to cationic substrates or inhibitors while the esteratic
subsite, which contains a serine hydroxyl group, undergoes acylation
during a catalytic turnover (Schaffer et al., 1973). Furthermore, the
existence of peripheral anionic sites on AChE has been suggested by the
effect of certain multi-quaternary AChE antagonists on AChE activity at
low ionic strength (Changeux, 1966). Flaxedil, for example, reverses the
-2 -

o? h 3
H3C - N - c h2—c H2- ° - C - C H 3 + 2 H2 0 ;=;
CHg
FIG. 1.1 Schematic representation of the hydrolysis of
acetylcholine.
t
anionicsite
esteraficsite
h 3 c - n - c h 2 - c h 2 - o - c - o h
CH. ICH
FIG. 1.2 Hypothetical model of the binding of acetylcholine to
the active site of AChE. From Nachmansohn and Wilson, 1951.
-3-

enzyme inhibition due to decamethonium although it does not itself bind
to the catalytic anionic site occupied by the inhibitor. It is likely
therefore that allosteric influence is exerted by ligand binding at
peripheral sites (Kitz et al., 1970; Changeux, 1966).
AChE is classified as a serine hydrolase because of its many
similar characteristics to those possessed by classical serine
hydrolases such as a-chymotrypsin. AChE exhibits irreversible active-
site phosphorylation by organophosphorous compounds. Equivalent weight
determination and peptide analysis indicate that phosphorylation only
involves one serine residue. This serine residue is part of a peptide
which shows significant homology with other enzymes of this class
(Schaffer et al., 1973). Both AChE and a-chymotrypsin exhibit bell-
shaped activity versus pH relationship. The apparent pKa of 6-7 for AChE
suggests a general acid-base catalysis by imidazole (Nachmansohn and
Wilson, 1951).
1.1.2 Inhibitors of cholinesterases
'Anticholinesterases' have been commonly used for biochemical
and physiological studies of cholinesterases (for review see Silver,
1974). The inhibition by organophosphorus compounds is characteristic of
cholinesterases. These compounds form a stable phosphorylated complex
with the serine hydroxyl moiety at the esteratic site of the enzyme.
Most organophosphorus compounds, such as diisopropyl phosphorofluoridate
(DFP), react with the enzyme only at the esteratic site. Some
organophosphorus compounds, however, bind simultaneously to both the
anionic and esteratic sites, and yield a stable phosphorylated enzyme
intermediate. The reversibility of these inhibitors depend on the rate
of regeneration by the hydrolysis of the intermediate complex. Esters of
- 4 -

phosphorus, e.g. echothiophate (Tammelin, 1957), substituted carbamic,
such as eserine and neostigmine (Machne and Unna, 1963), and sulfonic
acid derivatives such as phenyl methylsulfonylfluoride (PMSF), also form
stable complexes with the active sites. Another class of
anticholinesterases, which includes the quaternary compounds
decamethonium and edrophonium, interferes with substrate binding by
attaching to the anionic site (Long, 1963). A series of bis-quaternary
compounds, such as BW 284C51, also inhibit substrate binding by blocking
the anionic site (Fulton and Mogey, 1954). BW 284C51 is often used as a
selective inhibitor of AChE, while isoOMPA, an organophosphorus compound
(see earlier), is frequently used as a selective inhibitor of ^ChE.
These two inhibitors operate at different range of concentrations for
optimal selectivity (Holmstedt, 1957). The exact optimal concentrations
may vary between species and method used (Austin and Berry, 1953). For
mammalian and avian tissues, concentrations of 5 X 10"^M and 10~^m for
BW 284C51 and isoOMPA respectively were found to be most effective
(Hall, 1973; Lyles et_a]_., 1979).
1.1.3 Molecular structure of AChE : multiple molecular forms
The existence of multiple stable forms of AChE was first
demonstrated by Massoulie and Rieger (1969) in high ionic strength
extracts of fresh electric organ tissues of Electrophorus. These
molecular forms can be identified on sucrose density gradients, and show
characteristic sedimentation coefficients of 8S, 14S and 18S. Electron
microscopy of the purified enzyme shows that they have an asymmetric
structure consisting of a cluster of globular units attached to a rod
like 'tail' of approx. 50nm long (Dudai _et__al_., 1973). The globular
units, which contain the enzyme catalytic sites, are grouped into
tetramers. Each tail peptide can be joined to one, two or three of these
-5-

t e t r a m e r s ( Bon _ e t _ a l _ . , 1 9 7 6 ) . Each s u b u n i t ha s a m o l e c u l a r m ass o f a b o u t
80 kd (Duda i and S i l m a n , 1 9 7 4 ; R o s e n b e r r y and R i c h a r d s o n , 1 9 7 7 ) and t h e
t a i l p e p t i d e a b o u t 100 kd (Bon e t a l . , 1 9 7 6 ) . T h i s t a i l p e p t i d e i s
i n t e r e s t i n g i n t h a t i t c o n t a i n s a l a r g e number o f h y d r o x y p r o l i n e and
h y d r o x y l y s i n e r e s i d u e s w h ic h i s c h a r a c t e r i s t i c o f c o l l a g e n and i s
s e n s i t i v e t o c o l l a g e n a s e t r e a t m e n t i n a t e m p e r a t u r e d e p e n d e n t manner
(Bon and M a s s o u l i e , 1 9 7 8 ; A n g l i s t e r and S i l m a n , 1 9 7 8 ; R o s e n b e r r y e t a l . ,
1 9 8 0 ) , b u t i s p e p s i n r e s i s t a n t (Mays and R o s e n b e r r y , 1 9 8 1 ) ( F i g 1 . 3 ) .
T h e s e a s y m m e t r i c m o l e c u l e s a g g r e g a t e u n d e r lo w i o n i c s t r e n g t h c o n d i t i o n s
( G r a f i u s and M i l l a r , 1 9 6 5 ; Bon e t a l . , 1 9 7 8 ) . Such a g g r e g a t i o n c a n be
a b o l i s h e d by c o l l a g e n a s e o r l i m i t e d t r y p s i n t r e a t m e n t , a s w e l l a s upon
s t o r a g e , w h er e b y t h e c o l l a g e n - t a i l i s c l e a v e d , y i e l d i n g g l o b u l a r
t e t r a m e r s t h a t do n o t i n t e r a c t a t lo w s a l t c o n d i t i o n (Bon and M a s s o u l i e ,
1 9 7 8 ) . T h e s e t e t r a m e r s a r e l i n k e d t o t h e t a i l by d i s u l p h i d e bo n d s t o two
o f t h e s u b u n i t s , w h ic h form q u a t e r n a r y a s s o c i a t i o n w i t h t h e r e m a i n i n g
d i s u l p h i d e - l i n k e d d i m e r ( R o s e n b e r r y and R i c h a r d s o n , 1 9 7 7 ) , a s shown i n
F i g 1 . 3 . I t i s i m p o r t a n t t o n o t e t h a t t h e p o s i t i o n s o f t h e d i s u l p h i d e
b r i d g e s a r e o n l y h y p o t h e t i c a l . The m u l t i - s u b u n i t m o l e c u l a r f o r m s r e m a in
s t a b l e and a c t i v e e v e n a f t e r f u l l r e d u c t i o n o f t h e d i s u l p h i d e b o n d s ,
i n d i c a t i n g t h a t q u a t e r n a r y a s s o c i a t i o n s b e t w e e n t h e s u b u n i t s , and i n t h e
c a s e o f t h e a s y m m e t r i c f o r m s , b e t w e e n t h e s u b u n i t s and t h e t a i l i s
* i m p o r t a n t i n s t a b i l i s i n g t h e m u l t i m e r i c s t r u c t u r e .
The g l o b u l a r t e t r a m e r s ca n be f u r t h e r d i s s o c i a t e d i n t o d i m e r s
and monomers by l i m i t e d t r y p s i n d i g e s t (Bon and M a s s o u l i e , 1 9 7 6 ) . A
s y s t e m o f n o m e n c l a t u r e f o r t h e a s y m m e t r i c and g l o b u l a r fo rm s h a s b e e n
i n t r o d u c e d by Bon e t a l . ( 1 9 7 9 ) . The g l o b u l a r , t a i l - l e s s f o r m s and
c o l l a g e n - t a i l e d a s y m m e t r i c fo rm s w e r e d e s i g n a t e d a s G and A f o r m s
r e s p e c t i v e l y . Each o f t h e t h r e e m o l e c u l a r f o rm s i n e a c h c a t e g o r y i s
c l a s s i f i e d by a s u b s c r i p t t o i n d i c a t e t h e number o f c a t a l y t i c u n i t s
-6-

oo
FIG- 1-3 S c h e m a t i c r e p r e s e n t a t i o n o f t h e c o l l a g e n - t a i l e d form
o f AChE from E l e c t r o p h o r u s . The c i r c l e s r e p r e s e n t c a t a l y t i c
s u b u n i t s ; - S - S - , d i s u l p h i d e bonds w h ic h l i n k c a t a l y t i c s u b u n i t s
t o g e t h e r o r t o t h e c o l l a g e n t a i l . The t r i p l e h e l i c a l s t r u c t u r e
r e p r e s e n t s t h e c o l l a g e n t a i l w h ose s u b u n i t s a r e a l s o l i n k e d by
i n t e r d i s u l p h i d e b r i d g e s (n*v5). From Mays and R o s e n b e r r y , 1 9 8 1 .
- 7 -

p r e s e n t ( F i g 1 . 4 ) .
In T o r p e d o e l e c t r i c o r g a n e x t r a c t s , a p a r t from t h e c o l l a g e n
t a i l e d f o r m s , a p o p u l a t i o n o f AChE i n s e n s i t i v e t o s a l t c o n c e n t r a t i o n and
i n t e r a c t s w i t h n o n - i o n i c d e t e r g e n t s h a s b e e n i s o l a t e d ( W it z e m a n n , 1 9 8 0 ;
M a s s o u l i e and B o n , 1 9 8 2 ; V i r a t e l l e and B e r n h a r d , 1 9 8 0 ; L e e e t a l . ,
1 9 8 2 ) . The c a t a l y t i c s u b u n i t s i n t h i s po ol o f d i m e r i c and t e t r a m e r i c
f o r m s a r e s e n s i t i v e t o p r o n a s e o r p r o t e i n a s e K d i g e s t i o n w h ic h a b o l i s h e d
t h e i r i n t e r a c t i o n w i t h d e t e r g e n t s (L i and Bon , 1 9 8 3 ) .
A number o f m o l e c u l a r f o r m s o f AChE w ere a l s o f o u n d i n a v a r i e t y
o f mammalian t i s s u e s , i n c l u d i n g human e r y t h r o c y t e membrane ( O t t e t a l . ,
1 9 7 5 ; 1 9 8 2 ) and m u s c l e s ( C a r s o n e t a l . , 1 9 7 9 ) , b o v i n e e r y t h r o c y t e
membrane (B erm an , 1 9 7 3 ) and serum ( R a l s t o n e t a l . , 1 9 8 5 ) , r a t d ia p h r a g m
( H a l l , 1 9 7 3 ; Y ou nk in e t a l . , 1 9 8 2 ) , v e r t e b r a t e h i n d - l i m b m u s c l e s
( G r o s w a l d and D e t t b a r n , 1 9 8 3 a ; G i s i g e r and S t e p h e n s , 1 9 8 3 ; B a r n a r d et_
a l . , 1 9 8 4 a ; L y l e s e t a l . , 1 9 8 2 ; L y l e s and B a r n a r d , 1 9 8 0 ; B a r n a r d e t a l . ,
1 9 8 4 b ; Rotund o and Fam brough, 1 9 7 9 ) , n e u r a l t i s s u e s ( F e r n a n d e z e t a l . ,
1 9 7 9 ; Bon e t a l . , 1 9 7 9 ; Sung and R u f f , 1 9 8 3 ; G r a s s i e t a l . , 1 9 8 2 ) ,
c a r d i a c and smooth m u s c l e s (S k au and B r i m i j o i n , 1 9 8 0 ) , and c h i c k e n
p la s m a ( L y l e s e t a l . , 1 9 8 0 ) . T h e s e AChE fo rm s a r e a l s o s y n t h e s i z e d by
c u l t u r e d m u s c l e c e l l s ( S u g i y a m a , 1 9 7 7 ; R e i g e r e t a l . , 1 9 8 0 ; I n e s t r o s a e t
a l . , 1 9 8 2 ; B u l g e r e t a l . , 1 9 8 2 ) . They a r e t h o u g h t t o be a n a l o g o u s t o
• t h o s e i n E l e c t r o p h o r u s o r T o r p e d o AChE (Bon e t a l . , 1 9 7 9 ; V i g n y e t a l . ,
1 9 7 9 ) , s i n c e a number o f l o w s a l t i n s o l u b l e f o r m s r e s e m b l e t h e
a s y m m e t r i c f o rm s o f E l e c t r o p h o r u s , and t h e low s a l t s o l u b l e fo r m s may
r e p r e s e n t t h e v a r i o u s g l o b u l a r f o rm s w h ic h h a v e b e e n d e f i n e d . On t h e
o t h e r h a n d , i n v i e w o f t h e l i m i t e d i n f o r m a t i o n a v a i l a b l e on t h e
m o l e c u l a r s t r u c t u r e and c o m p o s i t i o n o f t h e s e m o l e c u l a r f o r m s i n
mammalian and a v i a n t i s s u e s , an e m p i r i c a l s y s t e m o f n o m e n c l a t u r e b a s e d
on s e d i m e n t a t i o n c h a r a c t e r i s t i c s o f t h e v a r i o u s m o l e c u l a r fo r m s h a s b e e n
-8-

ooe P n QjO OO
O p
\ /
ooQO oo og4 g2 Gi
FIG- 1.4 S c h e m a t i c r e p r e s e n t a t i o n o f t h e q u a t e r n a r y s t r u c t u r e
o f t h e a s y m m e t r i c (A) and g l o b u l a r (G) fo rm s o f AChE, a s
e s t a b l i s h e d f o r E l e c t r o p h o r u s . The c i r c l e s r e p r e s e n t i n d i v i d u a l
c a t a l y t i c s u b u n i t ; s h o r t l i n e s w h ic h j o i n t h e c i r c l e s r e p r e s e n t
t h e i n t e r s u b u n i t d i s u l p h i d e b r i d g e s . The c o l l a g e n t a i l i s
s y m b o l i s e d by t r i p l e l i n e s . The g l o b u l a r G4 form c o r r e s p o n d s t o
t h e a u t o l y t i c p r o d u c t , and h e n c e r e t a i n s a n o n - c o l l a g e n i c p e p t i d e
from t h e t a i l . T h i s s i m p l i f i e d r e p r e s e n t a t i o n o f t h e m o l e c u l a r
form s d o e s n o t i n c l u d e d i f f e r e n c e s t h a t may e x i s t f o r e a c h
q u a t e r n a r y s t r u c t u r e i n t e r m s o f i n t e r s u b u n i t d i s u l p h i d e b o n d s ,
g l y c o s i d i c r e s i d u e s , i n t r a s u b u n i t c l e a v a g e s , p r e s e n c e o f b i n d i n g
do m ain s o r n o n - c a t a l y t i c s u b u n i t s . A d a p te d from M a s s o u l i e and
Bon, 1 9 8 2 .
- 9 -

p r o p o s e d by B a r n a r d and c o w o r k e r s ( B a r n a r d e t a l . , 1 9 8 2 ; 1 9 8 4 b ) . The
v a r i o u s m o l e c u l a r f o rm s w e r e g i v e n t h e i n i t i a l s L, M o r H d e s i g n a t i n g
l i g h t , medium and h e a v y f o r m s r e s p e c t i v e l y . T h e s e t h r e e g r o u p s w e r e
f u r t h e r s u b d i v i d e d i n t o , 1-2, M, Mc , Hl c and H2 C , w h er e t h e numbers i n
t h e s u f f i x d e n o t e t h e s u b s p e c i e s w i t h i n e a c h g r o u p , and t h e l e t t e r c
d e n o t e s t h e p r e s e n c e o f a c o l l a g e n - l i k e t a i l p e p t i d e . B e c a u s e t h i s
s y s t e m o f n o m e n c l a t u r e h as t h e a d v a n t a g e s o f a d e q u a t e l y d e s c r i b i n g e v e r y
m o l e c u l a r form f o u n d t o - d a t e and d o e s n o t make any p r e s u p p o s i t i o n
c o n c e r n i n g t h e m o l e c u l a r c o m p o s i t i o n s o f t h e v a r i o u s m o l e c u l a r f o r m s , i t
w i l l be u s e d t h r o u g h o u t t h e p r e s e n t w o r k .
The c h a r a c t e r i s a t i o n o f t h e a m p h i p a t h i c f o r m s o f AChE from human
e r y t h r o c y t e membranes ( B r o d b e c k e t a l . , 1 9 8 4 ; R o s e n b e r r y and S c o g g i n ,
1 9 8 4 ; D u t t a - C h o u d h u r y and R o s e n b e r r y , 1 9 8 4 ) i l l u s t r a t e t h e a p p a r e n t
d i v e r s i t y o f t h e m o l e c u l a r s t r u c t u r e s w i t h i n t h e m o l e c u l a r f o r m s .
M o l e c u l a r fo r m s w h ic h h a v e i d e n t i c a l s e d i m e n t a t i o n p a r a m e t e r s c a n be
d i f f e r e n t s t r u c t u r a l l y and p h y s i o c h e m i c a l l y . F o r e x a m p l e , f u r t h e r
d e s i g n a t i o n i s c l e a r l y n e e d e d t o d i s t i n g u i s h h y d r o p h o b i c and h y d r o p h i l i c
f o rm s o f AChE, e . g . l_2 h may be u s e d t o d e s c r i b e t h e h y d r o p h o b i c ( h ) ,
d i m e r i c n a t u r e o f human e r y t h r o c y t e AChE. The v a r i o u s m o l e c u l a r f o rm s
may a l s o d i f f e r i n t h e i r c a r b o h y d r a t e c o n t e n t , w h ic h i s e v i d e n t from
t h e i r d i f f e r e n t l e c t i n b i n d i n g c h a r a c t e r i s t i c s ( R o t u n d o , 1 9 8 4 ) . D e s p i t e
t h i s m o l e c u l a r h e t e r o g e n e i t y , t h e v a r i o u s f o rm s o f AChE w i t h i n e a c h
g r o u p e x h i b i t e s s e n t i a l l y i d e n t i c a l c a t a l y t i c p r o p e r t i e s ( B a r n e t t and
R o s e n b e r r y , 1 9 7 9 ) . The s e d i m e n t a t i o n c o e f f i c i e n t s o f t h e v a r i o u s f o rm s
o f AChE i n mammalian and a v i a n t i s s u e s a r e su m m ar ized i n T a b l e 1 . 1 .
The v a r i o u s AChE fo r m s may be m o d i f i e d o r d i s s o c i a t e d by
p r o t e o l y t i c d e g r a d a t i o n . The c o l l a g e n - t a i l e d f o r m s , i n p a r t i c u l a r , a r e
s u s c e p t i b l e t o a u t o l y s i s upon s t o r a g e ( S i l m a n e t a l . , 1 9 7 8 ) . On t h e
o t h e r h a n d , a s s o c i a t i o n o f t h e l o w e r m o l e c u l a r w e i g h t f o rm s i n t o h i g h
-10-

Table 1.1 Sedimentation coefficients of the molecular fomjs of
AChE in chicken and rat skeletal muscle.
Species ' -Molecular form
L1 L2 M Hlc HZc
Chicken 4 6.5 11.5 14.8 20
Rat 3.5 6 9.9 13 16.7
(Bon, Vigr\y and Massoulie, 1979)

m o l e c u l a r w e i g h t a g g r e g a t e s was o b s e r v e d i n s t o r e d m ous e b r a i n e x t r a c t s
(Adamson e t a l . , 1 9 7 5 ) . S i m i l a r a g g r e g a t i o n o f t h e G f o r m s was o b s e r v e d
i n b o v i n e c a u d a t e n u c l e u s e x t r a c t s ( Hoi 1u n g e r and N i k l a s s o n , 1 9 7 3 ) and
c o u l d b e p r e v e n t e d by d e t e r g e n t ( G r a s s i e t a l . , 1 9 8 2 ) . Such i n v i t r o
i n t e r c o n v e r s i o n o f t h e m o l e c u l a r f o rm s u n d e r m in e s e f f o r t s t o s t u d y t h e
d i s t r i b u t i o n o f t h e s e f o r m s . I t a l s o makes more d i f f i c u l t t h e
c o r r e l a t i o n b e t w e e n c h a n g e s i n t h e d i s t r i b u t i o n o f t h e m o l e c u l a r f o rm s
and v a r i o u s p a t h o l o g i c a l s t a t e s , s u c h a s d e n e r v a t i o n i n d u c e d a t r o p h y
( T h e s l e f f , 1 9 7 4 ; B acou e t a l . , 1 9 8 2 ) and m u s c u l a r d y s t r o p h y ( G i s i g e r and
S t e p h e n , 1 9 8 3 ; L y l e s e t a l . , 1 9 8 2 ) .
1.1.4 Localisation of AChE
The s t r u c t u r a l h e t e r o g e n e i t y o f AChE i n a w i d e v a r i e t y o f
t i s s u e s i s now c o n s i d e r e d t o be r e l a t e d t o t h e i r d i s t r i b u t i o n and
s p e c i f i c l o c a l i z a t i o n i n t h e s e t i s s u e s ( f o r r e v i e w s e e R o s e n b e r r y ,
1 9 8 2 ) . The a m p h i p a t h i c f o r m s o f AChE from human e r y t h r o c y t e s , human
b r a i n and T o r p e d o e l e c t r i c o r g a n i n t e r a c t s t r o n g l y w i t h d e t e r g e n t
m i c e l l e s , and r e q u i r e d e t e r g e n t t o m a i n t a i n t h e i r c a t a l y t i c a c t i v i t y
( R o m e r - L u t h i e t a l . , 1 9 8 0 ; B r o d b ec k and O t t , 1 9 8 4 ) . In human e r y t h r o
c y t e s , a p a p a i n s e n s i t i v e , h y d r o p h o b i c p e p t i d e f r a g m e n t i s t h o u g h t t o be
r e s p o n s i b l e f o r t h e i n t e r a c t i o n o f AChE d i m e r w i t h l i p o s o m e s and i s
l i k e l y t o a s s o c i a t e w i t h t h e e r y t h r o c y t e membrane ( D u t t a - C h o u d h u r y and
R o s e n b e r r y , 1 9 8 4 ) . The a m p h i p a t h i c f o rm s o f AChE i n n e u r a l t i s s u e s , on
t h e o t h e r h a n d , a p p e a r s t o be a t e t r a m e r (M fo rm ) ( R a k o n c z a y e t a l . ,
1 9 8 1 ; S o r e n s e n e t a l . , 1 9 8 2 ; G r a s s i e t a l . , 1 9 8 2 ; R o t u n d o , 1 9 8 4 ) w h ic h
may be c o n v e r t e d t o a l o w s a l t s o l u b l e form by p r o n a s e o r p r o t e i n a s e K
t r e a t m e n t (L i and B o n , 1 9 8 3 ) . A d e t e r g e n t s o l u b l e L^ form h a s b e e n
o b s e r v e d i n a n e u r o b l a s t o m a s y m p a t h e t i c g a n g l i o n h y b r i d c e l l l i n e (T 2 8 )
( L a z a r and V i g n y , 1 9 8 0 ) . I t a p p e a r s t h a t t h e s e d e t e r g e n t s o l u b l e ,
g l o b u l a r AChE f o r m s i n n e u r a l t i s s u e s may h a v e d i f f e r e n t i n t r a c e l l u l a r
- 12-

and e x t r a c e l l u l a r l o c a l i s a t i o n , a s o b s e r v e d by t h e i r d i f f e r e n t i a l
s u s c e p t i b i l i t y t o i n h i b i t i o n by h y d r o p h i l i c i n h i b i t o r s ( T a y l o r e t a l . ,
1 9 8 1 ; L a z a r and V i g n y , 1 9 8 0 ) . I n t r a c e l l u l a r l y , t h e g l o b u l a r f o rm s a r e
a s s o c i a t e d w i t h t h e e n d o p l a s m i c r e t i c u l u m d u r i n g a x o n a l t r a n s p o r t
( M a s s o u l i e and B o n , 1 9 8 2 ; Couraud and Di G i a m b e r a r d i n o , 1 9 8 0 ) .
E x t r a c e l l u l a r l y , t h e y may be a s s o c i a t e d w i t h t h e p la s m a m embrane , a s
d e m o n s t r a t e d h i s t o c h e m i c a l l y i n m o t o r n e u r o n s ( L e w i s and S h u t e , 1 9 6 9 ) .
The lo w s a l t s o l u b l e , d e t e r g e n t i n s e n s i t i v e , g l o b u l a r fo r m s o f
AChE a r e m o st a b u n d a n t i n t i s s u e f l u i d s su c h a s serum ( G i s i g e r and
V i g n y , 1 9 7 7 ; Hodgson and Chubb, 1 9 8 3 ) o r p la sm a ( L y l e s e t a l . , 198 0 and
u n p u b l i s h e d r e s u l t s ) and c e r e b r a l s p i n a l f l u i d (Chubb e t a l . , 1 9 7 6 ) .
They a r i s e p r o b a b l y from t h e s e c r e t i o n o f n e u r a l ( S k a u and B r i m i j o i n ,
1 9 7 8 ; Chubb e t a l . , 1 9 7 6 ; G i s i g e r and V i g n y , 1 9 7 9 ) and m u s c l e c e l l s
( W i l s o n e t a l . , 1 9 7 3 ; B u l g e r e t a l . , 1 9 8 2 ; R ot u n d o and Fambroug h,
1 9 8 0 b ) . T h e s e f o r m s a l s o c o m p r i s e o f a f r a c t i o n o f t h e g l o b u l a r f o rm s i n
t h e r a t d ia p h r a g m ( Y o u n k in e t a l . , 1 9 8 2 ) and n e u r a l t i s s u e s ( L a z a r and
V i g n y , 1 9 8 0 ) . S e l e c t i v e i n h i b i t i o n o f e x t e r n a l l y l o c a t e d AChE f o r m s
sh ow s t h a t t h e s e lo w s a l t s o l u b l e form s a r e l o c a t e d l a r g e l y
i n t r a c e l l u l a r l y and a r e p r o b a b l y c o n f i n e d t o t h e lumen o f t h e
e n d o p l a s m i c r e t i c u l u m ( K r e u t z b e r g e t a l . , 1 9 7 3 ) . In m u s c l e c u l t u r e s ,
t h e y r e p r e s e n t a d i s t i n c t p o o l o f enzyme w i t h h i g h t u r n o v e r r a t e ,
• d e s t i n e d f o r s e c r e t i o n and d o e s n o t fo rm any m e m b r a n e - a s s o c i a t e d
i n t e r m e d i a t e ( R o t u n d o and Fambrough, 1 9 8 0 a ; 1 9 8 0 b ) .
The c o l l a g e n - t a i l e d , a s y m m e t r i c f o r m s o f AChE h a v e a t t r a c t e d
much i n t e r e s t b e c a u s e t h e y ca n be i s o l a t e d i n h i g h c o n c e n t r a t i o n from
e e l e l e c t r i c o r g a n (N ach m an so h n , 1 9 5 9 ) and a r e l o c a l i s e d a t t h e
n e u r o m u s c u l a r j u n c t i o n o f s k e l e t a l m u s c l e s ( H a l l , 1 9 7 3 ; Bon e t a l . ,
1 9 7 9 ; J e d r z e j c z y k e t a l . , 1 9 8 1 ) . I t se em s l i k e l y t h a t t h e s e f o r m s a r e
- 1 3 -

c l o s e l y a s s o c i a t e d w i t h t h e s y n a p t i c f u n c t i o n o f AChE. C o l l a g e n - t a i l e d
f o r m s a r e a l s o f o u n d i n t h e a t r i a o f r a t h e a r t (S k au and B r i m i j o i n ,
1 9 8 0 ) , and i n n e u r a l t i s s u e s ( G i s i g e r e t a l . , 1 9 7 8 ; F e r n a n d e z e t a l . ,
1 9 7 9 ; Skau and B r i m i j o i n , 1 9 8 0 ; G r a s s i e t a l . , 1 9 8 1 ) ( s e e a l s o C h a p t e r
5 ) . The l o c a l i s a t i o n o f t h e a s y m m e t r i c fo rm s i n t i s s u e s o t h e r t h a n
s k e l e t a l m u s c l e s and e l e c t r i c o r g a n h a s n o t b e e n s t u d i e d i n d e t a i l .
H o w e v e r , i t i s known t h a t i n m o to r n e u r o n e s t h e y a r e t r a n s p o r t e d by f a s t
a x o n a l f l o w and m us t t h e r e f o r e be i n t r a c e l l u l a r l y l o c a t e d (Di
G i a m b e r a r d i n o and C o u ra u d , 1 9 7 8 ) . The H fo rm s c a n a l s o b e f o u n d
e x t r a j u n c t i o n a l l y i n human i n t e r c o s t a l m u s c l e s ( C a r s o n e t a l . , 1 9 7 9 ) and
n e o n a t a l r a t s ( S k e l e t j and B r z i n , 1 9 8 0 ) .
T h e r e i s now a g r e a t d e a l o f e v i d e n c e , i n a d d i t i o n t o t h e i r
s o l u b i l i s a t i o n p r o p e r t i e s , t h a t t h e c o l l a g e n - t a i l e d fo rm s a r e a s s o c i a t e d
w i t h t h e b a s a l l a m i n a a t t h e m o t o r e n d p l a t e s ( f o r r e v i e w s s e e M a s s o u l i e
and Bon , 1 9 8 2 ; B r i m i j o i n , 1 9 8 3 ) . The H f o rm s from E l e c t r o p h o r u s
i n t e r a c t s s t r o n g l y w i t h a h e p a r a n - s u l p h a t e p r o t e o g l y c a n - l i k e compound
v i a t h e c o l l a g e n - t a i l u n i t ( V i g n y e t a l . , 1 9 8 2 ; Br andan and I n e s t r o s a ,
1 9 8 4 ) . H o m o g e n i s a t i o n o f r a t m u s c l e e n d p l a t e s w i t h h e p a r i n
p r e f e r e n t i a l l y s o l u b i l i s e s t h e a s y m m e t r i c form s o f AChE ( T o r r e s and
I n e s t r o s a , 1 9 8 3 ) . A r a t p h eoch rom ocy to m a d e r i v e d c e l l l i n e w h ic h l a c k s a
h e p a r a n on t h e c e l l s u r f a c e c o n t a i n s much l e s s e x t r a c e l 1 u l a r l y l o c a t e d
• a s y m m e t r i c form ( I n e s t r o s a e t a l . , 1 9 8 5 ) . H ow ev er , G r a s s i e t a l . ( 1 9 8 3 )
d i d n o t o b s e r v e any s i g n i f i c a n t i n t e r a c t i o n b e t w e e n t h e a s y m m e t r i c form
and v a r i o u s m a t r i x p r o t e i n s . F o c a l AChE s t a i n i n g c a n s t i l l be
d e m o n s t r a t e d h i s t o c h e m i c a l l y (McMahan e t a l . , 1 9 7 8 ) and
i m m u n o c y t o c h e m i c a l l y ( S a n e s and H a l l , 1 9 7 9 ) a f t e r t h e m o t o r axon and t h e
m u s c l e f i b r e h a v e b o t h b e e n r e m o v e d . S e v e r a l o b s e r v a t i o n s s u g g e s t t h e
p r e s e n c e o f AChE o t h e r t h a n t h e a s y m m e t r i c fo rm s a t t h e m o to r e n d p l a t e .
S k e t e l j and B r z i n ( 1 9 7 9 ) f o u n d t h a t c o l l a g e n a s e r e l e a s e d l e s s AChE from
- 1 4 -

t h e e n d p l a t e t h a n p a p a i n , w h ic h a l s o m o d i f i e s membrane bound f o r m s , a s
m e n t i o n e d a b o v e . G l o b u l a r f o r m s w e r e f o u n d t o b e p r e d o m i n a n t a t t h e
e n d p l a t e s i t e i n r a t h i n d - l i m b m u s c l e s ( V i g n y e t a l . , 1 9 7 6 ; F e r n a n d e z e t
a l . , 1 9 7 9 ; Y ou n k in e t a l . , 1 9 8 2 ) and c h i c k e n t o n i c m u s c l e s ( L y l e s and
B a r n a r d , 1 9 8 0 ) .
1 . 1 . 5 P u r i f i c a t i o n o f ACHE
The i s o l a t i o n and p u r i f i c a t i o n o f AChE a r e e s s e n t i a l f o r t h e
s t r u c t u r a l and k i n e t i c d e t e r m i n a t i o n o f t h e e n z y m e . P u r i f i c a t i o n
t e c h n i q u e s h a v e b e e n g r e a t l y im p r o v e d by t h e a p p l i c a t i o n o f a f f i n i t y
c h r o m a t o g r a p h y . A number o f a c r i d i n i u m o r t r i m e t h y l ammonium b a s e d
s y n t h e t i c a f f i n i t y l i g a n d s h a v e b e e n e m p lo y e d ( M a s s o u l i e e t a l . , 1 9 7 6 ;
V a l l e t t e e t a l . , 1 9 8 3 ) . T h e s e l i g a n d s b i n d t o t h e a n i o n i c s i t e o f AChE
v i a t h e ammonium and w h ic h ca n be s u b s e q u e n t l y d i s p l a c e d by AChE
i n h i b i t o r s s u c h a s d e c a m e t h o n i u m . The c h o i c e o f a p a r t i c u l a r a f f i n i t y
l i g a n d g e n e r a l l y d e p e n d s on t h e s o u r c e o f t h e enzyme and t h e c o n d i t i o n s
r e q u i r e d f o r i t s s o l u b i l i s a t i o n . The l e n g t h o f t h e s p a c e r arm and t h e
c o n c e n t r a t i o n o f t h e a f f i n i t y l i g a n d a r e a l s o c r i t i c a l on t h e e f f i c i e n c y
o f t h e l i g a n d ( V a l l e t t e e t a l . , 1 9 8 3 ) . A f f i n i t y c h r o m a t o g r a p h y h a s b e e n
u s e d i n t h e p u r i f i c a t i o n o f AChE from e e l (Dud a i and S i l m a n , 1 9 7 4 ;
R o s e n b e r r y e t a l . , 1 9 8 0 ) and T o r p e d o e l e c t r i c o r g a n ( H o p f f e t a l . , 1 9 7 3 ;
V i r a t e l l e and B e r n h a r d , 1 9 8 0 ; Le e e t a l . , 1 9 8 2 ; Lwebuga-Mukasa e t a l . ,
1 9 7 6 ) , human e r y t h r o c y t e membrane ( O t t e t a l . , 1 9 7 5 ; Berman and Y ou n g ,
1 9 7 1 ; R o s e n b e r r y and S c o g g i n , 1 9 8 4 ) , c h i c k b r a i n ( R o t u n d o , 1 9 8 4 ) ,
mammalian b r a i n ( R a c k o n c z a y e t a l . , 1 9 8 1 ; R e a v i l l and P lumm er, 1 9 7 8 ;
S o r e n s e n e t a l . , 1 9 8 2 ; Chan e t a l . , 1 9 7 2 ; Yamamura e t a l . , 1 9 7 3 ) and
s n a k e venom (Raba e t a l . , 1 9 7 9 ) . P u r i f i c a t i o n o f b o v i n e serum AChE u s i n g
e d r o p h o n i u m - l i n k e d S e p h a r o s e h a s a l s o b e e n a c h i e v e d (H o d g so n and Chubb,
1 9 8 3 ) .
- 1 5 -

1.1.6 Immunological study of ACHE
The a p p l i c a t i o n o f i m m u n o l o g i c a l t e c h n i q u e s t o t h e s t u d y o f AChE
h a s made p o s s i b l e a number o f a d v a n c e s i n t h e u n d e r s t a n d i n g o f t h e
e n z y m e . F i r s t l y , AChE d o e s n o t s h a r e common a n t i g e n i c d e t e r m i n a n t s w i t h
/ C h E , a s shown by many s t u d i e s , s u g g e s t i n g t h a t t h e tw o e n z y m e s a r e
l i k e l y t o be d i f f e r e n t g e n e p r o d u c t s ( B r i m i j o i n j e t _ a l _ . , 1 9 8 3 ; R o t u n d o ,
1 9 8 4 b ; M i n t z and B r i m i j o i n , 1 9 8 5 b ) . S e c o n d l y , a n t i b o d i e s t o p u r i f i e d r a t
b r a i n AChE b i n d a l l t h e m o l e c u l a r f o rm s w i t h s i m i l a r a f f i n i t i e s (Marsh
e t a l . , 1 9 8 4 ) . In c o n t r a s t , p r e f e r e n t i a l b i n d i n g o f p o l y c l o n a l
a n t i b o d i e s t o d e t e g e n t s o l u b l e , b u t n o t t o i n s o l u b l e AChE, h a s b e e n
o b s e r v e d i n r a t b r a i n ( Z a n e t t a e t a l . , 1 9 8 1 ) and human c a u d a t e n u c l e u s
( S o r e n s e n e t a l . , 1 9 8 2 ) . L i k e w i s e , D o c t o r e t a l . ( 1 9 8 3 ) r e p o r t e d
m o n o c l o n a l a n t i b o d i e s w h ic h a r e a b l e t o d i s t i n g u i s h b e t w e e n a s y m m e t r i c
and g l o b u l a r f o r m s . T h i r d l y , t h e ho m o lo g y b e t w e e n AChE from d i f f e r e n t
s p e c i e s c a n a l s o be s t u d i e d u s i n g m o n o c l o n a l a n t i b o d i e s ( B r i m i j o i n e t
a l . , 1 9 8 3 ; Fambrough e t a l . , 1 9 8 2 ; R o t u n d o , 1 9 8 4 b ; M in t z and B r i m i j o i n ,
1 9 8 5 b ) . T h e s e s t u d i e s show t h a t , i n g e n e r a l , l i t t l e c r o s s - r e a c t i v i t y i s
s e e n b e t w e e n AChE from d i f f e r e n t c l a s s e s o f a n i m a l s , w h i l e more
s u b s t a n t i a l c r o s s - r e a c t i v i t y o c c u r s among d i f f e r e n t o r d e r s o f t h e same
c l a s s . T h i s s u g g e s t s t h e l i k e l i h o o d o f e v o l u t i o n a r y d i s t i n c t i o n s b e t w e e n
t h e s e s p e c i e s o f AChE. F i n a l l y , by e x p l o i t i n g t h e c r o s s - r e a c t i v i t y o f
m o n o c l o n a l a n t i b o d i e s , i m m u n o a f f i n i t y p u r i f i c a t i o n o f r a b b i t b r a i n AChE
h a s b e e n a c h i e v e d u s i n g m o n o c l o n a l a n t i b o d i e s r a i s e d a g a i n s t p u r i f i e d
human e r y t h r o c y t e AChE ( M i n t z and B r i m i j o i n , 1 9 8 5 a ) .
1.2 Classification of skeletal muscle fibre types
S k e l e t a l m u s c l e i s a h e t e r o g e n e o u s t i s s u e due t o a) t h e
e x i s t e n c e o f a s p e c t r u m o f f i b r e s and b) t h e v a r i a b l e p a t t e r n o f t h e i r
a s s e m b l y . S e v e r a l t y p e s o f m u s c l e f i b r e s c a n be d i s t i n g u i s h e d on t h e
-16-

b a s i s o f t h e i r p h y s i o l o g i c a l , h i s t o c h e m i c a l , b i o c h e m i c a l and u l t r a -
s t r u c t u r a l p r o p e r t i e s (V r b o v a e t a l . , 1 9 7 8 ) . T h e s e m u s c l e f i b r e t y p e s
h a v e b e e n c l a s s i f i e d i n t o a number o f c a t e g o r i e s i n mammalian m u s c l e s
( B r o o k e and K a i s e r , 1 9 7 0 ; B a r n a r d e t a l . , 1 9 7 0 ) . A c l a s s i f i c a t i o n s y s t e m
r e l a t e d t o t h a t f o r t h e mammalian s y s t e m h a s a l s o b e e n d e s c r i b e d f o r
a v i a n m u s c l e s (Ashmore and D o e r r , 1 9 7 1 ; B a r n a r d e t a l . , 1 9 8 2 ) .
1.2.1 Physiological characterisation
S k e l e t a l m u s c l e s c a n b e d i v i d e d i n t o two t y p e s , v i z . t w i t c h and
t o n i c m u s c l e s on t h e b a s i s o f t h e i r c o n t r a c t i l e p r o p e r t i e s . T w i t c h
m u s c l e s a r e com p osed o f f a s t - and s l o w - t w i t c h f i b r e s . A c h a r a c t e r i s t i c s
o f t h e s e f i b r e s i s t h e i r a b i l i t y t o p r o p a g a t e a c t i o n p o t e n t i a l s and t o
r e s p o n d w i t h an a l l - o r - n o n e t w i t c h . Both t y p e s o f f i b r e s a r e i n n e r v a t e d
by an en p l a q u e t y p e o f n e r v e e n d i n g s . Th ey c a n be d i s t i n g u i s h e d h o w e v e r
by t h e i r c o n t r a c t i o n - r e l a x a t i o n and f a t i g u a b i 1 i t y c h a r a c t e r i s t i c s ( B u r k e
e t a l . , 1 9 7 1 ; C l o s e , 1 9 7 2 ) .
T o n i c m u s c l e s c o n t a i n f i b r e s w h ic h a r e m u l t i p i e l y i n n e r v a t e d
w i t h en g r a p p e t y p e o f n e r v e e n d i n g s . They r e s p o n d t o s u s t a i n e d s t i m u l i
w i t h s l o w , t o n i c c o n t r a c t i o n and u s u a l l y do n o t e x h i b i t an a c t i o n
p o t e n t i a l . T o n i c f i b r e s a r e r a r e i n mammals, and ca n o n l y b e f o u n d i n
s p e c i a l i s e d m u s c l e s s u c h a s t h e e x t r a o c u l a r and o e s o p h a g e a l m u s c l e s
( Y e l l i n , 1 9 6 9 ; H e s s , 1 9 7 0 ) .
T h e r e a r e a l s o c o n s i d e r a b l e d i f f e r e n c e s i n t h e c h a r a c t e r i s t i c s
o f n e u r o m u s c u l a r t r a n s m i s s i o n b e t w e e n d i f f e r e n t t y p e s o f f i b r e s
( M a g a z a n i k , 1 9 7 9 ; 1 9 8 4 ; McArdle and A l b u q u e r q u e , 1 9 7 3 ) . In mammals ,
f a s t - t w i t c h m u s c l e f i b r e s r e c e i v e a h i g h e r f r e q u e n c y o f m o t o n e u r o n
d i s c h a r g e ( E c c l e s e t a l . , 1 9 5 8 ) , show a h i g h e r f r e q u e n c y o f e n d p l a t e
p o t e n t i a l (M cA rdle and A l b u q u e r q u e , 1 9 7 3 ) and h a v e a h i g h e r q u a n t a l
c o n t e n t p e r e n d p l a t e p o t e n t i a l ( T o n g e , 1 9 7 4 ) t h a n t h o s e o f t h e s l o w -
- 1 7 -

t w i t c h f i b r e s . F u r t h e r m o r e , a c e t y l c h o l i n e (ACh) s e n s i t i v i t y c a n be
d e t e c t e d a l o n g t h e e n t i r e l e n g t h o f t h e s l o w - t w i t c h f i b r e s ( M i l e d i and
Z e l e n a , 1 9 6 6 ) w h i l e s e n s i t i v i t y t o ACh i s r e s t r i c t e d t o t h e e n d p l a t e
r e g i o n i n t h e f a s t - t w i t c h f i b r e s . The s l o w - t w i t c h m u s c l e s may a l s o
c o n t a i n l e s s ACh r e c e p t o r s ( S t e r z e t a l . , 1 9 8 3 ) and AChE ( M a g a z a n ik e t
a l , , 1 9 8 4 ) a t t h e e n d p l a t e r e g i o n .
1.2.2 Histochemical and biochemical characterisation
The h i s t o c h e m i c a l t e c h n i q u e s u s e d t o d i s t i n g u i s h t h e t y p e s o f
m u s c l e f i b r e s a r e b a s e d on t h e i d e n t i f i c a t i o n o f t h e m e t a b o l i c and
c o n t r a c t i l e e l e m e n t s o f m u s c l e f i b r e s ( D u b o w i t z and B r o o k , 1 9 7 3 ) .
A n a e r o b i c f i b r e s a r e c h a r a c t e r i s e d by a h i g h l e v e l o f enzyme m a r k e r s f o r
g l y c o l y t i c m e t a b o l i s m , s u c h a s p h o s p h o r y l a s e , l a c t a t e d e h y d r o g e n a s e and
a - g l y c e r o p h o s p h a t e d e h y d r o g e n a s e . The g l y c o g e n c o n t e n t o f t h e s e f i b r e s
a r e a l s o more a b u n d a n t . T h e i r a e r o b i c c a p a c i t y , on t h e o t h e r h a n d , i s
v e r y l i m i t e d a s i l l u s t r a t e d by lo w s u c c i n a t e d e h y d r o g e n a s e a s w e l l a s
lo w c y t o c h r o m e and m y o g l o b u l i n c o n c e n t r a t i o n s . A e r o b i c f i b r e s , i n
c o n t r a s t , h a v e a h i g h l e v e l o f e n zy m es f o r o x i d a t i v e m e t a b o l i s m and a
l o w l e v e l o f e n zy m es f o r g l y c o l y s i s .
I t was f o u n d t h a t t h e s p e e d o f c o n t r a c t i o n o f s k e l e t a l m u s c l e s
c o r r e l a t e d w i t h t h e a d e n o s i n e t r i p h o s p h a t a s e (A T P a se ) a c t i v i t y ( B a r a n y ,
1 9 6 7 ) and i t s s t a b i l i t y i n d i f f e r e n t pH m e d ia (Baran.y e t a l . , 1 9 6 5 ) . The
s p e e d o f s h o r t e n i n g , t h u s , a p p e a r s t o be an i n h e r e n t p r o p e r t y o f t h e
m y o f i b r i l s , b e i n g c l o s e l y r e l a t e d t o m y o s i n ATPase a c t i v i t y ( B a r a n y and
C l o s e , 1 9 7 1 ) . F a s t - t w i t c h m u s c l e s show h i g h e r m y o s i n ATPase a c t i v i t y
t h a n t h e s l o w - t w i t c h m u s c l e s ( B a r a n y , 1 9 6 7 ) . Th at t h e ATPase a c t i v i t y
may r e f l e c t d i f f e r e n t t y p e s o f c o n t r a c t i l e e l e m e n t i s d e m o n s t r a t e d by
t h e f i n d i n g o f a n t i g e n i c a l l y d i s t i n c t t y p e s o f m y o s i n s i n d i f f e r e n t
t y p e s o f f i b r e s ( A r n d t and P e p e , 1 9 7 5 ; S a r t o r e e t a l . , 1 9 7 8 ) . The
-18-

c u r r e n t c l a s s i f i c a t i o n o f f i b r e t y p e s , w h ic h t a k e s i n t o a c c o u n t t h e
v a r i o u s h i s t o c h e m i c a l and b i o c h e m i c a l e v i d e n c e , d i v i d e s m u s c l e f i b r e s
i n t o 4 b a s i c t y p e s : a) s l o w - t w i t c h - o x i d a t i v e ( t y p e I ) ; b) f a s t - t w i t c h -
o x i d a t i v e - g l y c o l y t i c ( t y p e 1 1 A ) ; c ) f a s t - t w i t c h - g l y c o l y t i c ( t y p e I I B ) ;
d) t o n i c ( o x i d a t i v e - g l y c o l y t i c ; t y p e I I I A and 11 I B ) .
1.2.3 Morphological and ultrastructural characterisation
F a s t - t w i t c h f i b r e s a r e c h a r a c t e r i s e d by a w e l l - d e v e l o p e d
s a r c o t u b u l a r s y s t e m w h i l e t h a t i n s l o w - t w i t c h and t o n i c f i b r e s a r e
r e l a t i v e l y l e s s w e l l d e v e l o p e d ( H e s s , 1 9 6 7 ; P e a c h e y and H u x l e y , 1 9 6 2 ) .
T w i t c h f i b r e s p o s s e s s s t r a i g h t Z d i s k s w h e r e a s t h a t i n s l o w t o n i c f i b r e s
a r e t h i c k and j a g g e d . S l o w - t w i t c h and t o n i c f i b r e s c o n t a i n a l a r g e r
number o f m i t o c h o n d r i a , l i p i d g l o b u l e s and a r e more v a s c u l a r i s e d t h a n
f a s t - t w i t c h f i b r e s .
A n o t h e r d i f f e r e n c e b e t w e e n t h e t w i t c h and t o n i c f i b r e s i s t h e i r
m o t o r e n d p l a t e m o r p h o l o g y . In mammalian m u s c l e s , t h e n e r v e e n d i n g s i n
t h e f a s t - t w i t c h f i b r e s c o v e r a l a r g e r e n d p l a t e r e g i o n and c o n t a i n more
p r e s y n a p t i c v e s i c l e s t h a n t h e s l o w - t w i t c h f i b r e s ( P a d y k u l a and G a u t h i e r ,
1 9 7 0 ; E l l i s m a n _ e t _ a l _ . , 1 9 7 6 ) . Such d i s t i n c t i o n , h o w e v e r , i s n o t a s c l e a r
b e t w e e n t h e t w i t c h and t o n i c f i b r e s i n t h e a v i a n m u s c l e s ( J . C a v a n a g h ,
p e r s o n a l c o m m u n i c a t i o n ) .
1 . 2 . 4 Fibre type composition of muscles
The f o u r t y p e s o f f i b r e s c a n be f o u n d i n v a r i o u s s p e c i e s o f
v e r t e b r a t e s ; h o w e v e r , any g i v e n s p e c i e s o f anim al d o e s n o t n e c e s s a r i l y
p o s s e s s a l l t h e s e t y p e s o f m u s c l e f i b r e s . Q u a n t i t a t i v e d i f f e r e n c e i n t h e
h i s t o c h e m i c a l s t a i n i n g c h a r a c t e r i s t i c s may a l s o o c c u r b e t w e e n f i b r e s o f
d i f f e r e n t s p e c i e s ( G o l d s p i n k and Ward, 1 9 7 9 ; P e t e r e t a l . , 1 9 7 2 ) .
G e n e r a l l y , i n d i v i d u a l m u s c l e c o n t a i n s e i t h e r a p u re p o p u l a t i o n o f o n e
- 1 9 -

t y p e o f f i b r e s o r a m ix e d p o p u l a t i o n o f v a r y i n g p r o p o r t i o n o f s e v e r a l
t y p e s o f f i b r e s ( A r i a n o e t a l . , 1 9 7 3 ) . The f i b r e t y p e c o m p o s i t o n o f a
m u s c l e a p p e a r s t o b e c o n t r o l l e d by t h e p a r t i c u l a r t y p e o f m u s c l e
a c t i v i t y . A good e x a m p l e i s t h e f l i g h t m u s c l e s o f b i r d s . The p e c t o r a l i s
i n n o n - f l y i n g b i r d s s u c h a s t h e c h i c k e n i s w h i t e o r p u r e l y f a s t - t w i t c h -
g l y c o l y t i c , w h i l e t h a t i n t h e f l y i n g b i r d s u c h a s t h e p i g e o n i s r e d o r
f a s t - t w i t c h - o x i d a t i v e (K ha n , 1 9 8 0 ) . The m e t a b o l i c p a t t e r n s a r e r e f l e c t e d
i n t h e number o f m i t o c h o n d r i a and i n t h e c o n t e n t o f g l y c o g e n and l i p i d .
On t h e o t h e r h a n d , t h e c o n t r a c t i l e p r o p e r t i e s a r e c o r r e l a t e d w i t h t h e
s i z e and s h a p e o f t h e m y o f i b r i l s , t h e t h i c k n e s s o f t h e Z d i s k s and t h e
d e v e l o p m e n t o f t h e s a r c o t u b u l a r s y s t e m ( I s h i k a w a , 1 9 8 3 ) .
F i b r e t y p e i s a l s o r e l a t e d t o t h e n a t u r e o f t h e i n n e r v a t i o n . A l l
m u s c l e f i b r e s w i t h i n a m o to r u n i t a r e o f t h e same f i b r e t y p e (B u r k e _et
a l . , 1 9 7 1 ) . C r o s s - i n n e r v a t i o n e x p e r i m e n t s wh er e t h e n e r v e s t o t h e
mammalian f a s t - t w i t c h and s l o w - t w i t c h m u s c l e s a r e i n t e r c h a n g e d , t h e
m u s c l e t y p e s a l s o u n d e r g o an i n t e r c h a n g e ( B u l l e r e t a l . , 1 9 6 0 ; M e l i d i
and S t e f a n i , 1 9 6 9 ; E i s e n b e r g and S a l m o n s , 1 9 8 1 ) . H o w e v e r , i n a v i a n
m u s c l e s , t o n i c and f a s t - t w i t c h m u s c l e s a r e n o t a f f e c t e d by c r o s s
r e i n n e r v a t i o n ( Hni k e t a l . , 1 9 6 7 ) e x c e p t i n n e w ly h a t c h e d c h i c k s ( Z e l e n a
and J i r m a n o v a , 1 9 7 3 ; J i rmanova e t a l . , 1 9 7 1 ) .
The p l a s t i c i t y o f m u s c l e f i b r e s i s a l s o d e m o n s t r a t e d by t h e
e f f e c t o f i n d u c e d a c t i v i t y on m u s c l e f i b r e t y p e c o m p o s i t i o n , w h e t h e r by
c h r o n i c e l e c t r i c a l s t i m u l a t i o n ( S a lm o n s and V r b o v a , 1 9 6 9 ; P e t t e , 1 9 8 0 ;
P e t t e e t a l . , 1 9 7 6 ) o r i n t e n s i v e e x e r c i s e t r a i n i n g (G r e e n e t a l . , 1 9 8 4 ) .
H ence t h e f i b r e t y p e c o m p o s i t i o n o f a m u s c l e i s a d y n a m ic s y s t e m a d a p t e d
t o f u n c t i o n a l d em a n d s . In c o n c l u s i o n , a l t h o u g h t h e t y p i n g o f f i b r e s
p r e s e n t s a somewhat p a r s i m o n i o u s v i e w o f t h e d yn am ic p r o p e r t i e s o f
m u s c l e s , some p r a c t i c a l c l a s s i f i c a t i o n h a v e n e v e r t h e l e s s b e e n p r o v e d
h e l p f u l i n t h e u n d e r s t a n d i n g o f t h e f u n c t i o n a l s i g n i f i c a n c e and t h e
-20-

p a t h o l o g y o f m u s c l e s ( P i z z e y and B a r n a r d , 1 9 8 3 a ; 1 9 8 3 b ; and f o r r e v i e w s
s e e S a lm o n s and H e n r i k s s o n , 1 9 8 1 ; P e t t e , 1 9 8 4 ; B r o o k e and K a i s e r , 1 9 7 4 ) .
1.3 Skeletal muscle AChE in health and disease
1.3.1 Biosynthesis of AChE in skeletal muscles
The m u l t i p l e m o l e c u l a r f o r m s o f AChE i s t h o u g h t t o be
s y n t h e s i z e d i n a s e q u e n t i a l manner by t h e p o s t - t r a n s l a t i o n a l p r o c e s s i n g
and a s s e m b l y o f m u l t i p l e number o f a b a s i c c a t a l y t i c u n i t ( M a s s o u l i e and
B o n , 1 9 8 2 ; B r i m i j o i n , 1 9 8 3 ) . The c o n c l u s i o n t h a t t h e s e c a t a l y t i c
s u b u n i t s a r e s i m i l a r i s b a s e d on k i n e t i c and i m m u n o l o g i c a l o b s e r v a t i o n s .
The k i n e t i c p r o p e r t i e s o f t h e v a r i o u s p u r i f i e d f o r m s o f AChE from
E l e c t r o p h o r u s and T o r p e d o e l e c t r i c o r g a n t i s s u e w e r e f o u n d t o be
s i m i l a r . I m m u n o l o g i c a l i d e n t i f i c a t i o n u s i n g m o n o c l o n a l o r p o l y c l o n a l
a n t i b o d i e s a l s o sh ows t h a t t h e v a r i o u s m o l e c u l a r f o r m s s h a r e
a n t i g e n i c a l l y i d e n t i c a l s u b u n i t s (Marsh e t a l . , 1 9 8 4 ; R o t u n d o , 1 9 8 4 ) .
E l e c t r o p h o r e t i c a n a l y s i s c a r r i e d o u t by W i l s o n and W a lk e r ( 1 9 7 4 )
sh ow ed t h a t t h e s y n t h e s i s o f AChE a f t e r i n h i b i t i o n by o r g a n o p h o s p h a t e
a p p e a r e d f i r s t a s lo w m o l e c u l a r w e i g h t f o r m s , and s u g g e s t e d t h a t t h e s e
l o w m o l e c u l a r w e i g h t fo rm s w e r e p r e c u r s o r s o f t h e h i g h m o l e c u l a r w e i g h t
f o r m s . T h i s was f u r t h e r s u p p o r t e d by t h e r e c o v e r y o f AChE i n
n e u r o b l a s t o m a c e l l c u l t u r e s a f t e r i r r e v e r s i b l e i n h i b i t i o n o f AChE, w h er e
G| ( l_ i ) form a p p e a r s b e f o r e t h e (M) form ( R i e g e r e t a l . , 1 9 7 6 ) . The
r e c o v e r y o f t h e lo w m o l e c u l a r w e i g h t f o r m s p r i o r t o t h e h i g h m o l e c u l a r
w e i g h t f o rm s i s s i m i l a r l y o b s e r v e d i n r a t m u s c l e c u l t u r e ( K o e n i g and
V i g n y , 1 9 7 8 ) , r a t s u p e r i o r c e r v i c a l g a n g l i a ( G i s i g e r and V i g n y , 1 9 7 7 )
and r a t d ia p h r a g m ( S k e l e t j e t a l . , 1 9 8 0 ) .
The m e t a b o l i c r e l a t i o n s h i p b e t w e e n t h e v a r i o u s f o rm s o f AChE h a s
b e e n i n v e s t i g a t e d by m e a s u r i n g t h e t u r n o v e r r a t e o f t h e s e f o r m s . In
-21-

c h i c k m u s c l e c u l t u r e s , i t was f o u n d t h a t t h e AChE m o l e c u l e s w h ic h bound
t o p la s m a membrane ( 7 S and 1 I S ) h a v e h a l f - l i v e s o f a b o u t 50 h o u r s , w h i l e
t h e i n t r a c e l l u l a r po o l o f AChE ( 7 S ) t u r n o v e r more r a p i d l y (R o t u n d o and
Fam broug h, 1 9 8 0 a ) . Such f i n d i n g h a s b e e n c o n f i r m e d by h e a v y i s o t o p e
l a b e l l i n g o f t h e m o l e c u l a r f o r m s i n n e u r a l t i s s u e c e l l l i n e ( L a z a r e t
a l . , 1 9 8 4 ) . The f a c t t h a t t h e membrane bound f o r m s a p p e a r w i t h a 2 - 3
h o u r l a g a f t e r t h e f i r s t a p p e a r a n c e o f t h e i n t r a c e l l u l a r form s u g g e s t s
t h a t t h e membrane bound form i s s y n t h e s i z e d by t h e p r o c e s s i n g o f t h e
i n t r a c e l l u l a r f o rm s ( I n e s t r o s a e t a l . , 1 9 8 5 ) . The s e q u e n c e o f e v e n t s
i n v o l v e d i n t h e p r o c e s s i n g o f t h i s i n t r a c e l l u l a r p o o l o f enzyme h a s b e e n
s t u d i e d i n q u a i l m u s c l e c u l t u r e b a s e d upon t h e c o t r a n s l a t i o n a l
g l y c o s y l a t i o n o f t h e enzym e m o l e c u l e s ( R o t u n d o , 1 9 8 4 a ) . I t was f o u n d
t h a t t h e a s s e m b l y o f t h e m ono m er ic u n i t s i n t o IS and 1 IS fo rm s o c c u r s i n
t h e roug h e n d o p l a s m i c r e t i c u l u m ( RER) , w h i l e t h e c o l l a g e n - t a i l e d f o r m s
a r e p r o b a b l y a s s e m b l e d from t h e 11S f o rm s and t h e c o l l a g e n t a i l i n t h e
G o l g i a p p a r a t u s .
A l t h o u g h t h e p r e c u s o r - p r o d u c t r e l a t i o n s h i p s b e t w e e n t h e g l o b u l a r
and c o l l a g e n - t a i l e d f o rm s o f AChE i s s u p p o r t e d by t h e e v i d e n c e d e s c r i b e d
a b o v e , su c h h y p o t h e s i s d o e s n o t a c c o u n t f o r t h e p o s s i b l e e x i s t e n c e o f
d i f f e r e n t p o o l s o f e n zy m es w h ic h may be r e g u l a t e d s e p a r a t e l y . P r i m a r y
c u l t u r e s o f b o t h m y o tu b e s ( W i l s o n e t a l . , 1 9 7 3 ; B u l g e r e t a l . , 1 9 8 2 ;
R otu n d o and Fam brough, 1 9 7 9 ; W a lk e r and W i l s o n ; C i s s o n and W i l s o n , 1 9 7 7 )
and n e r v e c e l l s ( L a z a r and V i g n y , 1 9 8 0 ; G i s i g e r e t a l . , 1 9 7 7 ) a c t i v e l y
s e c r e t e AChE i n t o t h e medium from a r a p i d l y t u r n o v e r p o o l o f
i n t r a c e l l u l a r enzyme ( R o t u n d o and Fambrough, 1 9 8 0 b ) w h ic h d o e s n o t
i n v o l v e any t r a n s i e n t a s s o c i a t i o n w i t h t h e p la sm a membrane . The s e c r e t e d
and membrance bound AChE, t h e r e f o r e , may r e p r e s e n t tw o d i s t i n c t p o o l s o f
e n z y m e s . F u r t h e r m o r e , l e c t i n b i n d i n g s t u d i e s a l s o s u g g e s t t h a t n e w l y
s y n t h e s i z e d AChE i s n o t n e c e s s a r i l y e n z y m a t i c a l l y a c t i v e ( R o t u n d o ,
1 9 8 4 a ) .
-22-

Some i m m u n o l o g i c a l e v i d e n c e s u g g e s t s t h a t t h e membrane b o u n d ,
d i m e r i c AChE fo rm from T o r p e d o e l e c t r i c o r g a n i s a n t i g e n i c a l l y d i s t i n c t
from t h e s o l u b l e o r a s y m m e t r i c f o rm s ( D o c t o r e t a l . , 1 9 8 3 ) . T o r p e d o AChE
i s a l s o d i f f e r e n t i n t h a t d u r i n g e m b r y o n ic d e v e l o p m e n t , t h e c o l l a g e n
t a i l e d form a p p e a r s e a r l i e r t h a n t h e d i m e r i c form ( B o n , 1 9 8 2 ) .
F u r t h e r m o r e , t h e a s s e m b l y o f t h e c o l l a g e n - t a i l e d form may i n v o l v e non -
c a t a l y t i c s u b u n i t s ( L e e and T a y l o r , 1 9 8 2 ) . T h e s e e v i d e n c e a g a i n s u g g e s t s
t h a t t h e v a r i o u s m o l e c u l a r f o rm s may n o t o r i g i n a t e from a common p o o l o f
' p r e c u r s o r ' u n i t s .
1.3.2 Neurotrophic regulation of AChE
T h e r e i s now a g r e a t d e a l o f e v i d e n c e t o show t h a t AChE o f
s k e l e t a l m u s c l e i s c o n t r o l l e d by t h e m o t o r i n n e r v a t i o n . N e r v e - m u s c l e
c o n t a c t i n v i v o o r i n v i t r o i s a c c o m p a n ie d by a l a r g e i n c r e a s e i n m u s c l e
AChE c o n t e n t i n t h e r a t ( V i g n y e t a l . , 1 9 7 6 ) o r c h i c k ( T o u t a n t e t _ a l _ . ,
1 9 8 3 ) , and s u b s e q u e n t l y f o c a l a c c u m u l a t i o n o f t h e enzyme a p p e a r s
( H a r r i s , 1 9 8 1 ; Z i s k i n d - C o n h a i m e t a l . , 1 9 8 4 ) . A f t e r d e n e r v a t i o n , t h e
e n d p l a t e - a s s o c i a t e d AChE d e c l i n e s i n b o t h mammalian ( H a l l , 1 9 7 3 ; C o l l i n s
and Y o u n k i n , 1 9 8 2 ) and a v i a n m u s c l e s ( S k e t e l j e t a l . , 1 9 7 8 ; S i l m a n e t
a l . , 1 9 7 8 ) . S i m i l a r l y , b l o c k a d e o f n e u r a l t r a n s m i s s i o n by f l a x e d i l
p r e v e n t e d e n d p l a t e AChE a p p e a r i n g i n c h i c k embryo m u s c l e ( B e t z e t a l . ,
1 9 8 0 ) . Such l o s s o f e n d p l a t e AChE c a n b e r e v e r s e d e i t h e r by
r e i n n e r v a t i o n (M al d on ad o e t a l . , 1 9 8 4 ; V i g n y e t a l . , 1 9 7 6 ) o r by d i r e c t
e l e c t r i c a l s t i m u l a t i o n o f t h e m u s c l e c e l l c u l t u r e (L0mo and S l a t e r ,
1 9 8 0 ; W a lk e r and W i l s o n , 1 9 7 5 ; 1 9 7 6 ) . N e r v e - m u s c l e c o - c u l t u r e i n w h ic h
s y n a p t i c t r a n s m i s s i o n i s b l o c k e d by c u r a r e c a n a l s o be r e v e r s e d by
s t i m u l a t i o n ( R u b in e t a l . , 1 9 8 0 ) . M u s c le a c t i v i t y t h u s a p p e a r s t o be
e s s e n t i a l f o r t h e s t a b i l i s a t i o n o f e n d p l a t e AChE. The f o r m a t i o n o f
j u n c t i o n a l ACh r e c e p t o r s and AChE c a n be i n d u c e d by n e u r a l s i g n a l a t t h e
-23-

e a r l i e s t s t a g e s o f j u n c t i o n a l d e v e l o p m e n t , b e f o r e c h a n n e l c o n v e r s i o n
o c c u r s ( B r e n n e r e t a l . , 1 9 8 3 ) . S u b s e q u e n t d e v e l o p m e n t o f t h e j u n c t i o n a l
c h a r a c t e r i s t i c s ca n be m a i n t a i n e d by d i r e c t e l e c t r i c a l s t i m u l a t i o n and
d o e s n o t r e q u i r e t h e p r e s e n c e o f t h e n e r v e . I t i s t h o u g h t t h a t t h e s e
s y n a p t i c s p e c i a l i s a t i o n i s e x e r t e d t h r o u g h t h e m o d i f i c a t i o n o f t h e b a s a l
l a m i n a a t t h e s i t e o f t h e n e r v e - m u s c l e c o n t a c t (W e i n b e r g and H a l l , 1 9 7 9 ;
Lomo and S l a t e r , 1 9 8 0 ; S a n e s e t a l . , 1 9 7 8 ) .
T h e r e i s a l s o e v i d e n c e t o s u g g e s t t h a t some n e u r o t r o p h i c f a c t o r s
may be r e s p o n s i b l e f o r t h e a c t i v i t y - i n d e p e n d e n t r e g u l a t i o n o f t h e n e r v e .
Thus b l o c k a d e o f a x o n a l t r a n s p o r t by c o l c h i c i n e r e s u l t s i n an i n c r e a s e d
r e l e a s e o f e n d p l a t e AChE s i m i l a r t o d e n e r v a t i o n ( I n e s t r o s a e t a l . ,
1 9 7 7 ) , w h e r e a s b o t u l i n u m t o x i n b l o c k a d e o f n e u r o t r a n s m i s s i o n , and t h u s
m u s c l e a c t i v i t y , i s n o t a s e f f e c t i v e a s d e n e r v a t i o n i n d e p l e t i n g
e n d p l a t e AChE (D rachman , 1 9 7 2 ) . F u r t h e r m o r e , n e r v e e x p l a n t s o r t i s s u e
e x t r a c t s h a v e b e e n shown t o i n d u c e ACh r e c e p t o r c l u s t e r i n g i n c l o n e d
m u s c l e c e l l s ( P o d l e s k i e t a l . , 1 9 7 8 ) , i n c r e a s e AChE a c t i v i t y i n newt
o r g a n c u l t u r e ( L e n t z , 1 9 7 4 ) and i n c r e a s e e n d p l a t e AChE i n d e n e r v a t e d r a t
m u s c l e o r g a n c u l t u r e ( D a v e y e t a l . , 1 9 7 9 ; Youn kin e t a l . , 1 9 7 8 ) and
m y o t u b e s ( S a n e s e t a l . , 1 9 8 4 ; L e n t z e t a l . , 1 9 8 1 ) . N e r v e e x t r a c t s a l s o
r e v e r s e t h e d e n e r v a t i o n - i n d u c e d r e d u c t i o n o f AChE i n t h e s u p e r i o r
c e r v i c a l g a n g l i o n o f t h e c a t ( K o e l l e and R u ch , 1 9 8 3 ) . The r e c e n t
• i d e n t i f i c a t i o n o f an ACh r e c e p t o r a g g r e g a t i n g f a c t o r h a s a l s o b e e n shown
t o i n d u c e AChE c l u s t e r i n g ( W a l l a c e e t a l . , 1 9 8 5 ) . T h i s f a c t o r , w h ic h i s
s e c r e t e d by n e u r a l t i s s u e s a s w e l l a s s y n t h e s i z e d by m u s c l e s i s
c o n c e n t r a t e d i n t h e s y n a p t i c c l e f t ( F a l l o n e t a l . , 1 9 8 5 ) and may b e an
i m p o r t a n t f a c t o r i n d i r e c t i n g t h e f o c a l a c c u m u l a t i o n o f AChE d u r i n g
s y n a p t o g e n e s i s and s u b s e q u e n t p r e f e r e n t i a l l o c a l i s a t i o n a t t h e e n d p l a t e .
-24-

1.3.3 Avian muscular dystrophy
M u s c u l a r d y s t r o p h y i n human i s a g e n e t i c a l l y i n h e r i t e d d i s o r d e r .
A number o f ani m al m o d e l s o f m u s c u l a r d y s t r o p h y h a v e b e e n u s e d f o r t h e
s t u d y o f t h i s d i s e a s e . The d y s t r o p h i c c o n d i t i o n i n c h i c k e n i s a
p r o g r e s s i v e a t r o p h y o f t h e f a s t - t w i t c h m u s c l e s , i n p a r t i c u l a r t h e
p e c t o r a l i s , t h e p o s t e r i o r l a t i s s i m u s d o r s i (PLD) and t h e w in g m u s c l e s .
The d i s e a s e i s t h o u g h t t o be o f m y o g e n i c o r i g i n s i n c e no a b n o r m a l i t i e s
c a n be d e t e c t e d i n t h e n e u r a l t r a n s m i s s i o n f u n c t i o n ( H o l l i d a y e t a l . ,
1965; W arnick e t a l . , 1979) a l t h o u g h t h e membrane o f d y s t r o p h i c f i b r e s
e x h i b i t abnormal f i b r i l l a t i o n and h y p e r e x c i t a b i l i t y t o m e c h a n i c a l
s t i m u l i , p o s s i b l y due t o an a l t e r a t i o n i n c a l c i u m c o n d u c t a n c e .
F a s t - t w i t c h f i b r e s s e v e r e l y a f f e c t e d by m u s c u l a r d y s t r o p h y
u n d e r g o r a p i d a t r o p h y . The f i b r e s become l a r g e , r o un de d and i r r e g u l a r .
At t h e same t i m e , s m a l l e r f i b r e s a l s o a p p e a r w h ic h may be a r i s e n from
t h e s p l i t t i n g o f l a r g e f i b r e s . The n u c l e i become more c e n t r a l l y
l o c a l i s e d and more a b u n d a n t , w h ic h i s r e f l e c t e d i n t h e i n c r e a s e o f DNA
c o n t e n t ( Y o s h i k a w a e t a l . , 1984). E x t e n s i v e c o n n e c t i v e and f a t t y t i s s u e
i n f i l t r a t i o n o c c u r s a s t h e d i s e a s e p r o g r e s s e s (Admundson and J u l i a n ,
1956). D e f e c t i v e membrane s t r u c t u r e and p e r m e a b i l i t y i s t h o u g h t t o be
t h e r e s u l t o f i n c r e a s e d l i p i d c o n t e n t i n t h e p la sm a membrane o f
d y s t r o p h i c f i b r e s (Rodan e t a l . , 1974). Such a b n o r m a l i t i e s i n t h e
membrane s t r u c t u r e may l e a d t o t h e l e a k a g e o f m y o f i b r i l l a r p r o t e i n s and
f a u l t y c a l c i u m c o n d u c t a n c e .
A number o f e n z y m e s i n t h e b l o o d h a v e b e e n e m p lo y e d a s m a r k e r s
f o r m o n i t o r i n g t h e p r o g r e s s i o n o f d y s t r o p h y . Abnormal i n c r e a s e i n t h e
l e v e l o f p la sm a a l d o l a s e and g l u t a m i c o x a l i c t r a n s f e r a s e w e r e n o t e d
( C o r n e l i u s e t a l . , 1959). C r e a t i n e k i n a s e and AChE a r e a l s o f o u n d t o be
e l e v a t e d ( Wi l s o n e t a l . , 1973). M u s c le AChE l e v e l was a l s o i n c r e a s e d
w h ic h ca n be s e e n i n m u s c l e a q u e o u s e x t r a c t o f d y s t r o p h i c p e c t o r a l i s a s
-25-

e a r l y a s 5 d a y s e x ov o ( Wi l s o n e t a l . , 1 9 6 9 ) . D e v e l o p m e n t a l s t u d y o f
AChE f o r m s i n normal and d y s t r o p h i c f a s t - t w i t c h m u s c l e s o f c h i c k e n
d y s t r o p h i c l i n e 4 1 3 ( D a v i s , C a l i f o r n i a ) sh ow s t h a t abnormal i n c r e a s e s i n
t h e AChE l e v e l b e g i n a s e a r l y a s 4 d a y s e x ov o and m a i n l y i n v o l v e s t h e
o v e r p r o d u c t i o n o f t h e 7S f o r m . The 20S form i s a l s o i n c r e a s e d m a n y - f o l d
( L y l e s e t a l . , 1 9 7 9 ) when comp ared w i t h normal m u s c l e s o f t h e same a g e
g r o u p . Such d i s t r i b u t i o n r e s e m b l e s t h e e m b r y o n ic d i s t r i b u t i o n o f AChE
f o r m s . In a d d i t i o n , t h e s e d y s t r o p h i c f a s t - t w i t c h f i b r e s r e t a i n a lo w
l e v e l o f l a c t a t e d e h y d r o g e n a s e (LDH) and p h o s p h o r y l a s e , and a r e
c o m p a r a t i v e l y more o x i d a t i v e t h a n t h e normal f i b r e s (A sh m ore and D o e r r ,
1 9 7 1 ) . The l e v e l s o f t h e s e enz ym es a r e s i m i l a r t o t h a t i n im m atu re f a s t -
t w i t c h m u s c l e s , and h e n c e l e a d s t o t h e p r o p o s a l t h a t d y s t r o p h y i s du e t o
a d e r e g u l a t i o n o f f i b r e m a t u r a t i o n ( W i l s o n e t a l . , 1 9 7 9 ; K a p la n and
Ch an , 1 9 6 2 ; Cosmos e t a l . , 1 9 7 9 ) .
I n c r e a s e i n AChE l e v e l s i s a l s o o b s e r v e d i n d e n e r v a t e d c h i c k e n
m u s c l e s . H ow e v e r , i n c r e a s e d AChE l e v e l due t o d y s t r o p h y i s d i f f e r e n t
from d e n e r v a t i o n i n d u c e d i n c r e a s e i n AChE s i n c e t h e H2 C fo rm i s a l s o
e l e v a t e d i n d y s t r o p h i c m u s c l e b u t d i s a p p e a r s i n d e n e r v a t e d m u s c l e .
F u r t h e r m o r e , d y s t r o p h i c m u s c l e s a r e f u n c t i o n a l l y i n n e r v a t e d ; i f a
d y s t r o p h i c m u s c l e i s d e p r i v e d o f i t s n e r v e , t h e AChE l e v e l i n c r e a s e s and
H2 C l e v e l d i s a p p e a r s s i m i l a r t o d e n e r v a t e d normal m u s c l e . H en ce
• d y t r o p h i c m u s c l e s a r e u n d er n e u r a l i n f l u e n c e .
In summary, a number o f s t a b l e , d i s t i n c t i v e m o l e c u l a r f o r m s o f
AChE a r e u b i q u i t o u s i n a v a r i e t y o f v e r t e b r a t e t i s s u e s . The e l u c i d a t i o n
o f t h e s t r u c t u r e s o f t h e s e f o rm s f u r t h e r d e m o n s t r a t e s t h e c o m p l e x
h e t e r o g e n e i t y o f t h e s e m o l e c u l e s . The l o c a l i s a t i o n o f t h e s e m o l e c u l e s ,
i n p a r t i c u l a r t h e d i m e r i c form i n e r t h r o c y t e membranes and t h e c o l l a g e n
t a i l e d f o rm s i n f i s h e l e c t r i c o r g a n , s u g g e s t s t h a t t h e
-26-

c o m p a r t m e n t a l i s a t i o n o f t h e s e f o r m s i s i n h e r e n t i n t h e i r m o l e c u l a r
s t r u c t u r e s . In p u r e s y s t e m s su c h a s t h e two m e n t i o n e d a b o v e , t h i s
r e l a t i o n s h i p se em s i n d i s p u t a b l e . H o w e v e r , t h e m o l e c u l a r form s o f AChE
p r e s e n t i n n e u r a l t i s s u e s and m u s c l e s a r e more c o m p l e x and s u c h
s t r u c t u r e - l o c a l i s a t i o n r e l a t i o n s h i p i s d i f f i c u l t t o d e f i n e .
N e v e r t h e l e s s , t h e l o c a l i s a t i o n o f t h e s e f o rm s h a s g e n e r a l l y b e e n
c o n s i d e r e d t o r e f l e c t , a t l e a s t i n w e l l d e f i n e d s y s t e m s s u c h a s t h e
n e u r o m u s c u l a r j u n c t i o n o f m u s c l e s and n e r v e s y n a p s e s , t h e i r r e s p e c t i v e
f u n c t i o n s . In p a r t i c u l a r , t h e r e l a t i o n s h i p b e t w e e n t h e l o c a l i s a t i o n and
f u n c t i o n o f t h e H2C AChE form i n t h e n e u r o m u s c u l a r j u n c t i o n h as o f t e n
b e e n u s e d a s a m ark er i n d e v e l o p m e n t a l s t u d i e s . H o w e v e r , s i n c e v a r i o u s
e v i d e n c e s u g g e s t s t h a t t h i s form i s n o t e x c l u s i v e t o t h e n e u r o m u s c u l a r
j u n c t i o n no r d o e s i t seem t o be t h e o n l y form i n t h e e n d p l a t e , f u r t h e r
work i s e s s e n t i a l t o c l a r i f y t h e l o c a l i s a t i o n o f t h i s form and i t s r o l e
a s f u n c t i o n a l m a r k e r .
This thesis
I t i s t h e aim o f t h e p r e s e n t work t o e s t a b l i s h more p r e c i s e l y
t h e l o c a l i s a t i o n o f t h e AChE f o rm s and t h e i r r e g u l a t i o n i n s k e l e t a l
m u s c l e s . I t was d e c i d e d t o f o c u s t h e s t u d y on s k e l e t a l m u s c l e s b e c a u s e
o f t h e i r f u n c t i o n a l d i v e r s i t y and t h e i r a m e n a b i l i t y t o f u n c t i o n a l
• m a n i p u l a t i o n by p r o c e d u r e s su c h as d e n e r v a t i o n and a r t i f i c i a l
s t i m u l a t i o n . In a d d i t i o n , a t t e m p t s w e r e made t o s t u d y t h e e f f e c t s o f
m u s c l e a c t i v i t y on t h e abnormal d i s t r i b u t i o n o f AChE f o r m s i n d y s t r o p h i c
a v i a n m u s c l e s i n o r d e r t o g a i n f u r t h e r i n s i g h t i n t o t h e normal
r e g u l a t i o n o f AChE. C h a p t e r 2 d e t a i l s t h e e x p e r i m e n t a l c o n d i t i o n s , n o t
p r e v i o u s l y w e l l - d e f i n e d , w h ich a r e c r i t i c a l f o r t h e r e p r o d u c i b l e i n
v i t r o a n a l y s i s o f AChE m o l e c u l a r f o r m s i n s k e l e t a l m u s c l e s . The
r e l a t i o n s h i p b e t w e e n t h e d i s t r i b u t i o n o f t h e s e m o l e c u l a r f o r m s and
-27-

m u s c l e f u n c t i o n was t h e n d e s c r i b e d . C h a p t e r 3 d e s c r i b e s t h e i n f l u e n c e o f
t h e n e r v e on t h e d i s t r i b u t i o n o f AChE f o rm s i n d i f f e r e n t m u s c l e t y p e s .
C h a p t e r 4 d e s c r i b e s t h e r e s p o n s e o f normal and d y s t r o p h i c m u s c l e s t o
i n d u c e d m u s c l e a c t i v i t y , i n t e r m s o f c h a n g e s i n b o t h t h e m u s c l e
p h y s i o l o g y and t h e d i s t r i b u t i o n o f AChE. The r e l a t i o n s h i p b e t w e e n t h e s e
tw o p a r a m e t e r s was a l s o e x p l o r e d . F i n a l l y , C h a p t e r 5 d e s c i b e d t h e
p u r i f i c a t i o n o f AChE from c h i c k b r a i n and t h e p r o d u c t i o n o f m o n o c l o n a l
a n t i b o d i e s s p e c i f i c a g a i n s t t h i s e n z y m e .
-28-

CHAPTER 2
MULTIPLE MOLECULAR FORMS OF ACHE IN
MMtALIAN AND AVIAN SKELETAL MUSCLES
-29-

2 . 1 INTRODUCTION
In v e r t e b r a t e s k e l e t a l m u s c l e s , t h e m o l e c u l a r form s o f AChE
a r e c o m p r i s e d o f t h e g l o b u l a r L1# l_2 and M fo rm s, and t h e
a s y m m e t r i c c o l l a g e n - t a i l e d H^c and H2c fo rm s ( S e e S e c t i o n
1 . 1 . 3 ) . Some o r a l l o f t h e s e fo rm s c a n o c c u r i n v a r y i n g
p r o p o r t i o n s i n d i f f e r e n t m u s c l e s .
In a v i a n m u s c l e s , i t h a s b e e n p r o p o s e d t h a t t h e d i s t r i b u t i o n
o f t h e m o l e c u l a r for m s o f AChE i s d e p e n d e n t on t h e i r f i b r e t y p e
c o m p o s i t i o n ( B a r n a r d e t a l . , 1 9 8 2 ) . M u s c l e s a r e g e n e r a l l y made
up o f a m i x t u r e o f t y p e I ( s l o w - t w i t c h o x i d a t i v e ) , t y p e IIA
( f a s t - t w i t c h , o x i d a t i v e and g l y c o l y t i c ) , and I IB ( f a s t - t w i t c h
g l y c o l y t i c ) , and t y p e I I I ( t o n i c ) f i b r e s , c l a s s i f i e d by a v a r i e t y
o f h i s t o c h e m i c a l , m o r p h o l o g i c a l , b i o c h e m i c a l and e l e c t r o p h y s i o -
l o g i c a l c h a r a c t e r i s t i c s (V rbova e t a l . , 1 9 7 8 ) . In t h e c h i c k e n
and p i g e o n , t h e H2 c form p r e d o m i n a t e s i n a l l t h e t w i t c h m u s c l e s
s t u d i e d t o - d a t e ( L y l e s e t a l . , 1 9 8 2 ; B a rn a rd e t a l . , 1 9 8 2 ;
B a r n a r d e t a] _ . , 1 9 8 4 ) . S m al l v a r i a t i o n s i n t h e amount o f L and
H^c for m s a p p e a r t o c o r r e l a t e w i t h t h e r e l a t i v e amount o f t y p e
I I and t y p e I f i b r e s . In c o n t r a s t , t o n i c m u s c l e s su c h a s t h e ALD
c o n t a i n l a r g e l y L2 and very l i t t l e H2c fo rm s ( B a r n a r d e t a l . ,
1 9 8 2 ; L y l e s and B a r n a r d , 1 9 8 0 ) . The p r o f i l e o f t h e m o l e c u l a r
f o rm s o f AChE i s t h u s e a s i l y d i s t i n g u i s h a b l e b e t w e e n t h e t w i t c h
and t o n i c m u s c l e s .
I t i s now w e l l e s t a b l i s h e d t h a t H2c i s t h e e n d p l a t e
a s s o c i a t e d form o f AChE i n s k e l e t a l m u s c l e s ( H a l l , 1 9 7 3 ;
J e d r z e j c z y k e t a l _ . , 1 9 8 1 ) . In c h i c k e n s , t h e e n d p l a t e s o f
f a s t - t w i t c h m u s c l e s s u c h a s t h e PLD c o n t a i n l a r g e l y t h e H2c
-30-

form and v e r y l i t t l e g l o b u l a r f o r m s , w h e r e a s t h e form i s
p r e d o m i n a n t i n t h e e n d p l a t e s o f t h e t o n i c ALD ( J e d r z e j c z y k e t
a l . , 1 9 8 4 ) . T h u s , t h e p r e d o m i n a n t m o l e c u l a r f o r m ( s ) o f AChE i n
c r u d e c h i c k e n m u s c l e e x t r a c t s i s l a r g e l y c o n c e n t r a t e d a t t h e
e n d p l a t e .
A l t h o u g h e x t e n s i v e l y s t u d i e d , t h e r e h a s b e e n l i t t l e
c o n s i s t e n c y i n t h e p r o f i l e s o f AChE m o l e c u l a r form s o f mammalian
m u s c l e s c o n t a i n i n g p r e d o m i n a n t l y e i t h e r f a s t o r s l o w f i b r e s .
C o n s i d e r a b l e v a r i a t i o n i n t h e AChE p r o f i l e can o c c u r among
m u s c l e s o f t h e same s p e c i e s , o r b e t w e e n t h e same m u s c l e i n
d i f f e r e n t s p e c i e s ( M a s s o u l i e and B o n , 1 9 8 2 ) . M o r e o v e r , a l t h o u g h
a l l o f t h e s e m u s c l e s ha v e t w i t c h p r o p e r t i e s ( t o n i c m u s c l e s a r e
r a r e i n mammals) , t h e p r o p o r t i o n o f t h e ^ form i s g e n e r a l l y
s m a l l . Thus i n t h e m u s c l e s o f m a tu r e a n i m a l s , t h e amount o f t h e
Hgc f o r m , a s a p r o p o r t i o n o f t h e t o t a l AChE v a r i e s from 5%-7%
i n t h e r a t g a s t r o c n e m i u s o r r e c t u s m u s c l e s ( V i g n y e t a l . . 1 9 7 6 ;
F e r n a n d e z e t a l ., 1 9 7 9 ) , t o 15%-20% i n t h e r a t EDL ( G r o s w a l d and
D e t t b a r n , 1 9 8 3 a ; S e n n i e t a l _ . , 1 9 8 1 ) and t o a b o u t 30% i n t h e
m ous e s o l e u s ( G i s i g e r and S t e p h e n s , 1 9 8 3 ) . As i n a v i a n m u s c l e s ,
t h e H2C form w as a l s o f o u n d t o be a s s o c i a t e d w i t h t h e e n d p l a t e
r e g i o n o f t h e r a t d ia p h r a g m ( H a l l , 1 9 7 3 ; R i e g e r and V i g n y , 1 9 7 6 ;
Y o u n k in e t __ a l . , 1 9 8 2 ) , t h e r a t s t e r n o m a s t o i d (Bon e t a l . , 1 9 7 9 )
and t h e r a b b i t sem im em b ran osu s (B aco u e t a l . , 1 9 8 2 ) . H ow e v e r , a
m a j o r p r o p o r t i o n o f t h e e n d p l a t e - a s s o c i a t e d AChE i n mammalian
m u s c l e s , u n l i k e t h a t i n t h e c h i c k e n , i s p r e s e n t a s g l o b u l a r fo rm s
( M a s s o u l i e and Bon, 1 9 8 2 ) . In o t h e r w o r d s , t h e e n d p l a t e
a s s o c i a t e d H2C AChE c o n s t i t u t e s o n l y a m in o r p a r t o f t h e t o t a l
- 3 1 -

AChE a c t i v i t y , t h e m a j o r i t y b e i n g c o n t r i b u t e d by a v a r y i n g
p r o p o r t i o n o f L and M f o r m s , b o t h i n s i d e and o u t s i d e t h e
e n d p l a t e s .
T h i s s e e m i n g l y d i v e r s e c o m p o s i t i o n o f AChE f o rm s i n v a r i o u s
m u s c l e s may be due t o a number o f r e a s o n s . F i r s t l y , mammalian
m u s c l e s a r e u s u a l l y o f m ix e d f i b r e - t y p e c o m p o s i t i o n ( D u b o w i t z and
B r o o k e , 1 9 7 3 ; V r b o v a , e t a l . , 1 9 7 8 ) ; o n l y a f e w ( e . g . g u i n e a p i g
s o l e u s , r a b b i t sem im em br ano su s a c c e s s o r i u s / p r o p r i u s ) a r e
c o m p o se d p u r e l y o f o n e t y p e o f f i b r e ( P e t e r e t a l . , 1 9 7 2 ) .
F u r t h e r m o r e , t h e f i b r e t y p e c o m p o s i t i o n o f t h e same m u s c l e may
a l s o v a r y b e t w e e n s p e c i e s ( G a u t h i e r and P ac ty ku la , 1 9 6 6 ; s e e a l s o
m ous e and g u i n e a p i g s o l e u s , T a b l e 2 . 1 ) . I f , a s i n t h e c h i c k e n ,
t h e AChE c o n t e n t i n mammalian m u s c l e s i s a l s o d e t e r m i n e d by t h e
f i b r e t y p e c o m p o s i t i o n , t h e n v a r i a t i o n s i n t h e AChE p r o f i l e w o u ld
be e x p e c t e d .
S e c o n d l y , i t i s p o s s i b l e t h a t t h e s e a p p a r e n t v a r i a t i o n s a r e
a r t e f a c t s . As m u s c l e c r u d e e x t r a c t s a r e commonly u s e d f o r t h e
a n a l y s i s o f AChE f o r m s , t h e v a r i a b i l i t y i n t h e i r c o m p o s i t i o n and,
i n p a r t i c u l a r , t h e low l e v e l o f W^c form commonly o b s e r v e d i n
mammalian m u s c l e s c o u l d be due t o t h e p r e s e n c e o f e n d o g e n o u s
p r o t e a s e i n t h e m u s c l e e x t r a c t s . I n d e e d , s e v e r a l g r o u p s ha v e
♦ shown t h a t t h e c o l l a g e n - t a i l e d fo rm s o f AChE a r e s e n s i t i v e t o a
number o f p r o t e a s e s w h ic h c o n v e r t them t o g l o b u l a r fo rm s
( M a s s o u l i e and R i e g e r , 1 9 6 9 ; A n g l i s t e r and S i l m a n , 1 9 7 8 ; V ig n y e t
a l . , 1 9 7 9 ; Mays and R o s e n b e r r y , 1 9 8 1 ) . F u r t h e r m o r e , i t w a s shown
t h a t c o n v e r s i o n o f t h e H form i n t o M and L fo rm s o c c u r r e d d u r i n g
s t o r a g e o f t h e t i s s u e p r e p a r a t i o n e i t h e r a s t i s s u e e x t r a c t s
( R o t u n d o and Fambrough, 1 9 7 9 ) o r a s f r o z e n t i s s u e s ( L y l e s e t a l . ,
1 9 8 2 ) . Mammalian s k e l e t a l m u s c l e s a r e known t o c o n t a i n a w i d e
-32-

TABLE 2.1 The fibre type composition of some manual Ian and chicken skeletal muscles which had been chosen for AChE analysis.
*
Muscle type
Fast-tw1 tch
Slow-twltch
Mixed
Tonic
Species MuscleFibreI
type composition IIA 1 IB
(x)I I I
AChEXL
formsSM XH
Anterior t ib ia lis 4 46 50 - 31 6 62EDL 6 37 57 - 30 - 51
Guinea pig Gastrocnemius 12 32 56 - 18 - 61Semimembranosus accessorius <1 22 77 - 32 - 44Vastus latera lis (red) 4 78 18 - 23 6 69Vastus latera lis (white) 0 29 71 - 26 7 67
Anterior tib ia lis* 6 61 33 . 12 63Hamster EDL* 11 31 58 - 21 13 66
Plantarls 17 82 0 - 31 - 55
Anterior t ib ia lis 2 66 32 38 45 17Rat EDL 5 18 77 62 21 12
Sternomastold N.D. 55 23 13
Rabbit Anterior t ib ia lis 3 40 57 _ 41 45EDLa N.D. 44 - 42Semimembranosus accessorius 2 12 86 - 45 - 35
Mouse Anterior t lb la lls a N.D. 26 44 30
Chicken PLD <3 10 90 - 7 11 82
Plantaris* 100 0 0 20 68Guinea pig Semimembranosus proprlus 100 0 0 - 23 - 63
Soleus 100 0 0 - 17 12 71Hamster Soleus 80 20 0 - 18 - 71Rat Soleus 84 16 0 - 15 - 62Rabbit Semimembranosus proprlus* 100 0 0 - 17 21 61
Soleus 96 4 0 - 33 20 47
Ch1ckenb* Adductor profundus (anterior) 31 69 0 _ 24 12 64Rat Diaphragm 40 27 34 33 34 33Mousec* Soleus 42 58 0 - 31 18 51
Chicken* Adductor profundus (posterior) _ - O O 57 14 19Chicken ALD - - - 100 50 30 20
a These muscles were considered to be typical fast-twitch muscles by their electrophyslologlcal and biochemical characteristics. N.D. * fibre type composition not available,
b For some other chicken limb muscles, see also Chapter 4. c The fibre type composition was determined on three C57 black mice.* Fibre type compositions of these muscles were determined In this laboratory, courtesy of J.A . Plzzey and J . Jarv is.
The fibre type compositions of the other muscles were cited from Arlano et a l . . 1973 ; Peter et a l. , 1972 ; Campbell, 1979; Goldsplnk and Ward, 1979; Barnard et a l .. 1982 and Metzger et al.. 1985
- 3 3 -

v a r i e t y o f p r o t e a s e s ( P e n n i n g t o n , 1 9 7 7 ) , i d e n t i f i a b l e by t h e i r
s u b s t r a t e and i n h i b i t o r s p e c i f i c i t i e s , m a i n l y i n m a s t c e l l s
( B e n d i t t , 1 9 5 6 ; Park e t a l . , 1 9 7 3 ; B r a u n - F a l c o a n d S a l f e l d , 1 9 5 9 ;
Woodbury e t a l 1 9 8 1 ; S c h e c h e t e r e t a l 1 9 8 3 ) a n d c o n n e c t i v e
t i s s u e s ( B o i s , 1 9 6 4 ) . The v a r i e t y and s p e c i f i c i t y o f t h e s e
p r o t e a s e s may d i f f e r b e t w e e n d i f f e r e n t t y p e s o f t i s s u e s ( L a g u n o f f
e t __ a l _ . , 1 9 6 2 ) and a c o m p a r a t i v e stucty o f t h e s e p r o t e a s e s i n
s k e l e t a l m u s c l e s o f d i f f e r e n t s p e c i e s i s i n c o m p l e t e . T h e s e
e n z y m e s a r e r e s p o n s i b l e f o r p r o t e i n c a t a b o l i s m , a n d a r e l i k e l y t o
a t t a c k AChE i n v i t r o d u r i n g t h e t i s s u e h o m o g e n i s a t i o n and
e x t r a c t i o n p r o c e d u r e s when c e l l s t r u c t u r e s a r e d i s r u p t e d . H e n c e ,
p r e c a u t i o n a r y m e a s u r e s s h o u l d be t a k e n t o p r e v e n t t h e i n v i t r o
breakdow n o f t h e H form s so t h a t su ch a n a l y s i s a c c u r a t e l y
r e f l e c t s t h e AChE f o rm s p r e s e n t i n v i v o . P r o t e a s e i n h i b i t o r s a r e
c l e a r l y s u i t e d f o r t h i s p u r p o s e . M o s t o f them a r e l o w m o l e c u l a r
w e i g h t , a c t i v e s i t e - d i r e c t e d o r g a n i c compounds o r p e p t i d e s , w h ic h
a r e e i t h e r s y n t h e t i c o r n a t u r a l l y o c c u r r i n g ( m o s t o f t h e
i n h i b i t o r s u s e d i n t h i s s t u d y b e l o n g t o t h e l a t t e r c l a s s ) ( T a b l e 2.
2 ) . P r o t e a s e i n h i b i t o r s e x h i b i t i n g b r o a d s p e c i f i c i t y , s u c h a s
c ^ - m a c r o g l o b u l i n , a r e a l s o u s e d . A s u i t a b l e c o m b i n a t i o n o f
t h e s e i n h i b i t o r s s h o u l d p r o t e c t t h e H AChE from breakd ow n by
s u p p r e s s i n g a l l t h e p r o t e a s e a c t i v i t y p r e s e n t i n t h e m u s c l e .
The f i n a l r e a s o n f o r t h e v a r i a b i l i t y o f AChE p r o f i l e s may b e
due t o t h e d i f f e r e n t a n a t o m i c a l d i s t r i b u t i o n o f AChE form s i n
mammalian m u s c l e f i b r e s . As m e n t i o n e d e a r l i e r , t h e r e i s , i n a l l
c a s e s r e p o r t e d , a l a r g e p r o p o r t i o n o f g l o b u l a r fo rm s p r e s e n t i n
e n d p l a t e - r i c h r e g i o n s o f mammalian m u s c l e s , u n l i k e t h o s e i n t h e
e n d p l a t e s o f c h i c k e n t w i t c h m u s c l e s w h ic h a r e e n r i c h e d i n t h e
- 3 4 -
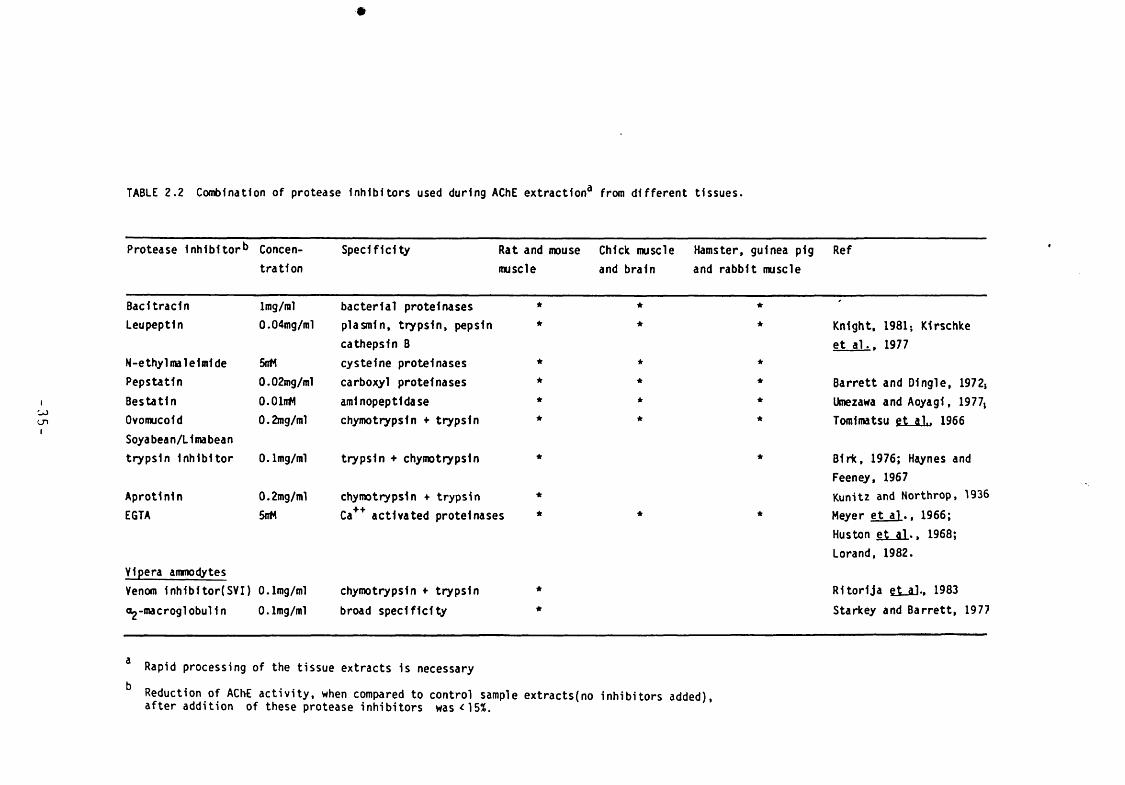
TABLE 2.2 Combination of protease inhib itors used during AChE extraction3 from different tissues.
Protease in h ib ito r^ Concen
tra tio n
Spec ific ity Rat and mouse
muscle
Chick muscle
and brain
Hamster, guinea pig
and rabb it muscle
Ref
Bacitracin lmg/ral bacteria l protelnases * * * *
Leupeptln 0.04mg/ml plasmin, tryps in , pepsin * * * Knight, 1981; Kirschke
cathepsln B e t a l. . 1977
N-ethylnalelmide 5nM cysteine proteinases ★ * *
Pepstatln 0.02mg/ml carboxyl protelnases * * * Ba rre tt and Dingle, 1972,
Bestatln O.OlnM ami nopeptidase ★ * * Umezawa and Aoyagl, 1977,
Ovomucoid 0 .2mg/ml chymotrypsin + trypsin * * * Tomimatsu et al,, 1966
Soyabean/Limabean
tryps in In h ib ito r O.lmg/ml tryps in + chymotrypsin * * B1rk, 1976; Haynes and
Feeney, 1967
Apro tin in 0.2mg/ml chymotrypsin + trypsin * Kunitz and Northrop, 1936
EGTA 5nH Ca++ activated protelnases * * * Meyer e t a l .. 1966:
Huston e t a l .. 1968:
Lorand, 1982.
Ylpera ammodytes
Venom inh ib ito r(S V I) O.lmg/ml chymotrypsin + tryps in * R lto r l ja et al.. 1983
c^-macroglobulin O.lmg/ml broad spec if ic ity * Starkey and Ba rre tt, 1977
Rapid processing of the tissue extracts is necessary
Reduction of AChE a c t iv ity , when compared to control sample extracts(no in h ib ito rs added), a fte r addition of these protease in h ib ito rs was <15%.

H£C f o r m . I t i s c o n c e i v a b l e t h a t , i f t h e g l o b u l a r fo rm s a r e
i n d e e d p a r t o f t h e f u n c t i o n a l AChE i n mammalian m u s c l e s
e n d p l a t e s , t h e y w o u ld a c c o u n t f o r t h e h i g h e r l e v e l o f g l o b u l a r
fo rm s i n mammlian m u s c l e s .
The a im o f t h e p r e s e n t s t u d y i s t o d e t e r m i n e i f t h e r e i s a
c o r r e l a t i o n b e t w e e n t h e f i b r e t y p e c o m p o s i t i o n and t h e
d i s t r i b u t i o n o f AChE f o r m s . To do t h i s , m u s c l e s o f d e f i n e d f i b r e
t y p e c o m p o s i t i o n from s e v e r a l mammalian s p e c i e s w e r e a n a l y s e d . A
d e t a i l e d s t u d y was c a r r i e d o u t a t t h e same t i m e t o i d e n t i f y and
r e l a t e t h e a r t e f a c t u a l e f f e c t s o f e n d o g e n o u s p r o t e a s e s on t h e
d i s t r i b u t i o n o f AChE form s i n v i t r o , and p r e c a u t i o n a r y p r o c e d u r e s
w e r e t h e n d e s i g n e d t o e l i m i n a t e o r m i n i m i z e t h e s e a r t e f a c t s .
2 . 2 MATERIALS AND METHODS
2 . 2 . 1 Antals
G u in ea p i g s ( D u n k i n - H a r t l e y , two m onth s o l d ) , r a b b i t s (New
Z e a l a n d w h i t e , %3kg), c h i c k e n s (London l i n e 4 1 2 , 2 - 4 m on th s o l d )
and r a t s ( S p r a g u e - D a w l e y , %200g) w e r e m a i n t a i n e d i n t h e
B i o c h e m i s t r y d e p a r t m e n t , I m p e r i a l C o l l e g e . H a m s t e r s ( G o l d e n , 3
m o n th s o l d ) , w e r e o b t a i n e d from M i d d l e s e x H o s p i t a l , M i d d l e s e x .
• M ic e ( 1 2 9 / R e J , n o r m a l , 2 m on th s o l d ) w e r e o b t a i n e d from C h a r i n g
C r o s s H o s p i t a l , London.
2 . 2 . 2 Muscles
M u s c l e s w h ic h c o n t a i n p r e d o m i n a n t l y t y p e I o r t y p e I I f i b r e s
w e r e s e l e c t e d from t h e a b o v e a n i m a l s a s l i s t e d i n T a b l e 2 . 1 . The
r a t d ia ph rag m w a s c h o s e n t o r e p r e s e n t mammalian m u s c l e s o f m ix e d
f i b r e t y p e c o m p o s i t i o n .
- 3 6 -

2 . 2 . 3 Materials
B a c i t r a c i n , l e u p e p t i n , p e p s t a t i n , c h i c k e n o v o m u c o id t r y p s i n
i n h i b i t o r ( o v o m u c o i c j ) , a p r o t i n i n , b e s t a t i n , N - e t h y l m a l e i m i d e (NEM),
5 , 5 ' - d i t h i o b i s - ( 2 - n i t r o b e n z o i c a c i d ) (DTNB), t e t r a i s o p r o p y l p y r o -
p h o s p h o r a m i d e ( i soOM PA), BW284C51, b o v i n e l i v e r c a t a l a s e , E . c o l i
B - g a l a c t o s i d a s e , a c e t y l c h o l i n e c h l o r i d e , a - c h y m o t r y p s i n from
b o v i n e p a n c r e a s , a z o c o l l and FITC c o n j u g a t e d c a s e i n w e r e o b t a i n e d
from S ig m a ; a c e t y l t h i o c h o l i n e i o d i d e and T r i t o n X - 1 0 0 from
K o c h - L i g h t Ltd ; c ^ - m a c r o g l o b u l i n from B o e h r i n g e r Mannheim;
[ H] a c e t y l c h o l i n e from Arnersham. The V i p e r a am modytes venom
t r y p s i n i n h i b i t o r ( S V I ) was a g i f t from Dr. R. S h i p o l i n i o f t h i s
d e p a r t m e n t .
2 . 2 . 4 Precautions applied in tissue extraction
The f o l l o w i n g p r e c a u t i o n s w e r e t a k e n d u r i n g t i s s u e
e x t r a c t i o n i n o r d e r t o a c h i e v e o p t i m a l p r e s e r v a t i o n o f t h e
m o l e c u l a r fo rm s o f AChE. The e f f e c t i v e n e s s o f t h e s e p r e c a u t i o n s
w as j u d g e d by t h e r e p r o d u c i b i l i t y o f t h e m o l e c u l a r p r o f i l e s
o b t a i n e d and t h e r e l a t i v e amount o f t h e form p r e s e r v e d .
M u s c l e s from t h e h a m s t e r , g u i n e a p i g and r a t w e r e u s e d i n t h e s e
p r e l i m i n a r y e x p e r i m e n t s .
1 . P e r f u s i o n A n im a ls w e r e d e e p l y a n a e s t h e t i s e d w i t h
b a r b i t u r a t e . The t h o r a c i c c a v i t y was o p e n e d by a m i d l i n e
i n c i s i o n and by c a r e f u l l y c u t t i n g t h e d ia p h r a g m from t h e r i b
c a g e . C a r d i o v a s c u l a r p e r f u s i o n was t h e n p e r f o r m e d u s i n g a
p h o s p h a t e b u f f e r e d s a l i n e (PBS) c o n t a i n i n g lOmM EDTA, pH 7 0 .
A p p r o x i m a t e l y 1 m l / g body w e i g h t o f t h e b u f f e r was a p p l i e d a t
1 0 - 2 0 m l / m i n.
- 3 7 -

Protease inhibitors2 . For tissue homogenisation and
extraction, buffers containing the various protease inhibitors,
their concentrations as listed in Table 2.2, were prepared fresh
before use. Selective combinations of the inhibitors were tested
for their ability to preserve the H2C form in tissue extracts.
None of the inhibitors used had any inhibitory activity on AChE
The activities of endogenous proteases in tissue extracts,
and their inhibition by protease inhibitors, were measured using
azocoll or FITC conjugated casein as substrates.
i) Azocoll hydrolysing activity (Saheki and Holzer, 1974)-
batches of azocoll (2.4%) in 1% Triton X-100/1M NaCl phosphate
buffer, pH 7.0,were mixed (5:1) with the muscle crude extract and
incubated, with vigorous shaking, at 25°C. At different time
intervals, up to 24 hours, 0.5ml of the suspension was mixed 1:1
with 10% TCA and centrifuged at 10,000xg for 3 min. The
absorbance of the supernatant was determined at 520nm.
ii) FITC conjugated casein hydrolysing activity — this assay was
modified from the method by Twining, 1983. Batches of
FITC-casein (0.025%), dissolved in 1% Triton X-100/1M NaCl
phosphate buffer, were mixed (4:1) with the muscle crude extract
and incubated at 37°C; at different time intervals, up to 3
hours, the reaction was terminated by mixing 0.05ml of the
mixture with 0.05ml of 10% TCA. The precipitate was centrifuged
at 10,000xg(3 min). 0.08ml of the supernatant was made up to 0.7ml
with 0.1M phosphate buffer, pH 7 0. The supernatant was excited
at 490nm and its fluorescence recorded at 525nm in a Perkin Elmer
(model no. 203) fluorescence spectrophotometer. The combined
protease inhibitors were prepared in the FITC-casein solution,
or included during homogenisation. Dose-response curves were
constructed for SVI, a^-macroglobulin and bacitracin.
-38-

a-chymotrypsin from bovine pancreas was used as a standard
protease for both azocoll and FITC-casein assays. The enzyme
activity of the standard a-chymotrypsin solution was determined
using BTEE as substrate, and;hydrolysis was monitored at 256 nm
in a Cecil spectrophotometer (Hummel, 1959). Enzyme activity was
calculated as:
Uni ts/ml = Afl256/min X 100__________
964 X vol. of reaction mix
3. Rapid processing of tissue extracts
Muscles were excised immediately upon death and rinsed
briefly in PBS. No more than 6 samples were handled at ar\y one
time in order to minimise the processing time. Muscles were
minced briefly on ice, then either the whole muscle, or an
aliquot of the minced muscle, was homogenised in 10 vol of an
ice-cold phosphate buffer containing 1% Triton X-100, 1M NaCl and
an array of protease inhibitors as described in Table 2.2.
Depending on the sample volume, these muscles were either
homogenised with glass/glass hand homogenisers or with a Polytron
(Kinetica) (3x30s) at 4°C. The homogenates were immediately
centrifuged at 43,500xg for 15 min to obtain the crude extracts,
which were then immediately applied onto sucrose gradients for
sedimentation analysis. The whole process of homogenisation and
extraction was accomplished within 60 min.
Long incubations were also carried out in the presence and
absence of protease inhibitors, up to 4 hours before
sedimentation analysis. The efficiency of extraction of AChE
after long or rapid processing was estimated by comparing the
AChE activity present in the extract with that in the muscle
homogenate.
-39-

4. Rapid freezing of muscles The rat soleus muscle was
excised and immediately dropped into liquid nitrogen. The muscle
was then crushed, in liquid nitrogen, into small pieces and
processed as described above.
2.2.5 Hicrodfssection
This technique enables intact whole fibres, endplates and
non-endplate segments to be dissected from muscle. All
manipulations were carried out under a dissecting microscope
fitted with an ice-cooled platform. The muscle was cleaned of
surface connective tissue. A longitudinal slice was then trimmed
from the muscle so that the whole span of the fibre length was
included. Small bundles of fibres were then teased from the edge
of this slice with microforceps, taking care not to stretch or
damage the fibres. Usually a group of 20-30 fibres could be
isolated and, keeping the fibres moist with PBS, further trimmed
to remove any broken fibres. The intact fibres obtained were
then transferred to a fresh cold surface, gently teased and
chopped.
Endplate and non-endplate segments were also dissected from
muscle fibres. The muscle was cleaned of surface connective
• tissues and a suitable slice of the muscle was stained for
endplate AChE (Karnovsky and Roots, 1964). As soon as the
endplates became visible, they were carefully excised from
individual muscle fibre. Non-endplate segments, each about five
times the length of an endplate,were taken from the same fibres
far removed from the endplate region without entering the
tendonous region. Batches of 20-30 endplates or non-endplate
segments were pooled.
-40-

The dissected whole fibres, endplates and non-endplate
segments were immediately extracted by shaking for 60-90 min in
0.25ml of the above medium at room temperature.
2.2.6 Sedimentation analysis
This procedure was carried out as described previously
(Lyles et__aK, 1982). 5% to 20% linear sucrose gradients were
prepared using a Buchler Auto Densi-Flow IIC apparatus (Haake
Buchler), on a 0.5ml 60% sucrose solution in 12ml polyallomer
centrifuge tubes. The sucrose solutions were made up in 1%
Triton X-100/ 1M NaCI phosphate buffer, pH 7.0. Crude muscle
extracts or suspensions of microdissected samples were layered
onto the gradients and centrifuged in a Beckman SW40Ti rotor atip p
4°C, to a preset total of 1.02 x 10 rad for mammaliani i p
tissues, or 9.6 x 10 rad for avian tissues, in a Beckman
L8-70 ultracentrifuge. Catalase and 0-galactosidase were
included as internal protein standards. After sedimentation, the
gradients were fractionated with the Buchler Auto Densi-Flow IIC
apparatus.
2.2.7 Enzyme assay
The AChE activity of crude extracts were assayed by the
colorimetric method of Ellman et al. (1969), essentially
as described previously (Lyles et al.. 1979; Lyles, 1980), in the
presence of isoOMPA, a ^ChE inhibitor. The assay was carried out
at room temperature in a medium containing InM DTNB/ 0.1M
phosphate/ 10"^M isoOMPA. Extracts (10 pi) were preincubated
in 0.5 ml of this medium for 30 min, after which
- 4 1 -

acetylthiocholine iodide was added to a final concentration of
0.75nM to initiate the enzyme reaction. The rate of hydrolysis
of acetylthiocholine iodide was monitored by the increase in
absorbance at 412 nm on a LKB Ultrolab System 2086 Mark II
Kinetic Analyser. The concentration of AChE was calculated using
Beer's law:
C = AOD where 6= 13.6 t l Jt= 1
The sucrose gradient fractions from whole muscle extracts
were assayed by the same method unless otherwise stated. 0.5 ml
of a double strength DTNB/isoOMPA solution was added to each
fraction and preincubated for 30 min. The enzyme reaction was
initiated by adding a further 0.5 ml of 1.5 mM acetylthiocholine
iodide. The length of incubation varied depending on the rate of
colour development. The absorbance of the gradient fractions was
recorded on a Cecil spectrophotometer after suitable incubation.
AChE from gradient fractions of microdissected sample
extracts, or samples containing NEM which interferes with the
colorimetric assay, were analysed radiometrically using
[ H]-acetylcholine (Johnson and Russell, 1973). A typical
assay mixture contained lCf^M isoGMPA, 0.788mM acetylcholineo
chloride with [ H]- acetylcholine and the gradient fraction or
crude extract, in a final volume of 0.3ml and 0.1ml respectively.
The reaction was terminated by adding 0.1ml of stopping mix (1M
chloroacetic acid/ 2M NaCl/ 0.5M NaOH) to crude extract sample
assays, or 0.15ml of a double strength stopping mix to gradient
fraction assays. The whole fraction was mixed in %4.5 ml of 90%
toluene/10% aniyl alcohol/PP0/P0P0P scintillant. AChE activity
was expressed as the proportion of substrate hydrolysed (in cpm)
- 4 2 -

to the total substrate hydrolysed using excess pure AChE from eel
(approximately lO^cpm). AChE activity was calculated as
fol1ows:
Enzyme units _ Sample cpm blank x nmoles substrate added
(nmoles AChE Eel cpm - blank running time x sample vol
hydrolysed/mi n/ml) (min) (ml)q
The non-specific hydrolysis of [ H] acetylcholine, or
acetylthiocholine iodide, was estimated by running a parallel
assay of buffer blanks, or crude extracts/gradient fractions
inhibited with both isoOMPA and 5 x lCf^M BW284C51 (AChE
inhibitor).
Enzyme marker assays
A) Catalase (11.4S)(EC 1.11.1.6) 1 unit/nl sample was loaded
onto a gradient. The position of the marker was subsequently
detected by assaying for the breakdown of HgOg in the
gradient fractions. 1 ml of reaction mix (0.2 ml of 30% HgOg
in 142 ml of 0.05 M sodium phosphate, pH 7.0) was added to 10 ul
of a gradient fraction in a quartz cuvette. The decrease in
absorbance at 240 nm was immediately measured in a
spectrophotometer preset at O.D. = 0.2 with a reagent blank.
B) B-galactosidase (16 S) (EC. 3.2.1.23) 10 units/100 ul
sample was loaded. The position of this enzyme was determined by
the breakdown of its substrate, O-nitrophenylgalactoside (0NPG).
0.3 ml of 0NPG (0.6 g in 1 lit of 0.1 M phosphate, pH 7.2) was
mixed with 10 ill of a gradient fraction and incubated until a
strong yellow colour developed. The reaction was then stopped by
adding 0.6 ml of 1 M I^COg and the absorbance of the fraction
recorded at 416 nm.
- 4 3 -

2.3 RESULTS
SECTION I. Effects of extraction conditions on the
distribution of ACHE forms.
2.3.1 Protease activities in muscle extracts and the effects of
protease inhibitors on AChE extraction
The possibility that the variable amount of L forms observed
in tissue extracts may be a result of proteolytic degradation of
the higher molecular weight forms was assessed by monitoring the
azocoll or casein hydrolysing activity of the extracts. Both of
these proteins are substrates for a wide spectrum of proteases
which may be present in muscle extracts (Cabib and Ulane, 1973;
Reimerdeo and Klostermeyer, 1976). These non-specific substrates
were employed, instead of the many synthetic peptides used for
quantitative determination of specific proteases, for three
reasons. Firstly, assay systems employing synthetic peptides
could not be used because Triton X-100, which was present in the
medium, interfered with absorbance at 280nm. Secondly, the
non-specific substrates provided an estimate of total proteolytic
activity present in crude extracts so that the combined
inhibitory effect of the protease inhibitors could be assessed.
Finally, an accurate quantitative measurement of individual
protease was not essential.
The measurement of muscle protease activity using azocoll as
substrate was negative. This was probably because the level of
enzyme activity in the extracts was below the sensitivity range
of this assay, since the hydrolysing activity of a standard
- 4 4 -

a-chymotrypsin solution could be measured by this method.
However, the possibility that azocoll was an unsuitable substrate
for muscle proteases could not be ruled out.
The hydrolysis of FITC conjugated casein was a more
sensitive assay system for measuring endogenous protease activity
in muscle extracts. With purified bovine a-chymotrypsin, the
rate of hydrolysis was linear with enzyme concentrations as
previously reported (Twining, 1983). The present results showed
that rat muscles contained a large amount of protease activity
(Fig. 2.1), which was low or negligible in brain tissues and
guinea pig muscles.
The protease activities of three rat muscles, v.t. soleus.
sternomastoid and gastrocnemius were studied. They were found to
have a similar activity, roughly equivalent to 1.2 mil of bovine
pancreatic a-chymotrypsin per ml of extract. The protease
inhibitors, used in combination (Table 2.2), completely inhibited
the protease activity observed in these muscles (Fig. 2.1).
Interestingly, a2-macroglobulin produced an apparent total inhibition at concentrations of O.lmg/ml or above for a period of
60-90 min (Fig. 2.2C). This indicates that ag-macroglobulin
may be one of the most potent protease inhibitors within the
• processing time period. Bacitracin and SVI, on the other hand,
only produced a partial inhibition (Fig. 2.2A,B). The amount of
SVI required to produce 50% inhibition of bovine a-chymotrypsin
was 30 ug/unit of enzyme activity (Fig. 2.3).
2.3.2 Extraction of AChE by rapid processing in the presence of
protease inhibitors
Immediate analysis of freshly prepared crude extracts of
mammalian muscles by sucrose density gradients showed that the
-45-

FIG- 2-1 Casein hyrolysing activity of tissue crude extracts
from various species. 0.02% final concentration of
FITC-conjugated casein in 1M NaCl/1% Triton/O.IM phosphate, pH
7.0, incubated with crude extracts from: O , rat sternomastoid/
rat soleus (16.5 units/hr); # , rat brain/chick brain (1.4
units/hr); ^ , guinea pig hind limb muscles (undetectable);
■ , muscle and brain extracts in the presence of a full array
of protease inhibitors (Table 2.2.). All incubations were
carried out at 37°C. Activity is expressed in arbitrary
fluorescence units per hour. See text for calibration with
bovine a-chymotrypsin. Results were pooled from at least 2
animals.
- 4 6 -

FIG. 2.2 Dose dependent inhibition of casein hydrolysing
activity of rat muscle extracts by bacitracin, SVI and
c^-macroglobulin. (A) 0 , rat sternomastoid; Q , rat
soleus; (B) rat hind limb muscles; (C) rate of hydrolysis of
casein by rat soleus extract in the presence of different
concentrations of c^-macroglobulin; # , O.Olmg/ml; O ,
0 . 0 5 mg/ml; ■ , 0 . 1 mg/ml; □ , 0 . 5 mg/ml. Results were from
a typical experiment; repeat experiments gave similar inhibition
character!' sties.- 4 7 -

i
FIG- 2-3 Dose dependent inhibition of bovine a-chymotrypsin
activity by SVI. 0.5 itM final concentration of BTEE in 0.05 M
CaCl2/0.025 M Tris HC1, pH 7.8 was used as substrate. Reactions
were carried out at room temperature.
-48-

Hg form predominated in most type I and type II muscles, (e.g.
Fig. 2.4A,C). In contrast, if muscle homogenates or crude
extracts were stored (on ice) for a period of time, up to 4 hours
before sedimentation analysis, the H forms were greatly
diminished. At the same time, a proportional increase in the L
forms became apparent. Such changes occurred when the extracts
contained few (e.g. bacitracin, ovomucoid and NEM only) or no
protease inhibitors (Fig. 2.4B,D). The difference observed was
not due to limited extraction of L forms during rapid processing,
as tissue homogenisation and extraction in 1% Triton X-100/1M
NaCl solubilised at least 90% of total AChE (Appendix 1). There
was no difference between the AChE activities extracted slowly or
rapidly from samples of the same muscle.
If protease inhibitors were present in the muscle extracts,
the profile of AChE forms remained stable even after prolonged
incubation at 4°C (Fig. 2.5B). The H2c form in guinea pig
muscles predominated even in the absence of protease inhibitors,
provided that the extracts were processed rapidly (Fig. 2.4 A,C).
In contrast, rat muscles always require protease inhibitors to
suppress the breakdown of the H forms if reproducible results
were to be obtained. Such difference between guinea pig and rat
• muscle extracts correlated with the levels of protease activity
observed in the muscles of the two species. Rapid processing in
the presence of protease inhibitors, therefore, is an essential
procedure for the extraction and analysis of AChE forms.
The particular combination of protease inhibitors used
during AChE extraction differed between species, depending on the
level of protease activities in the particular tissues. Some
effective inhibitor combinations tested in various species are
listed in Table 2.2.
- 4 9 -
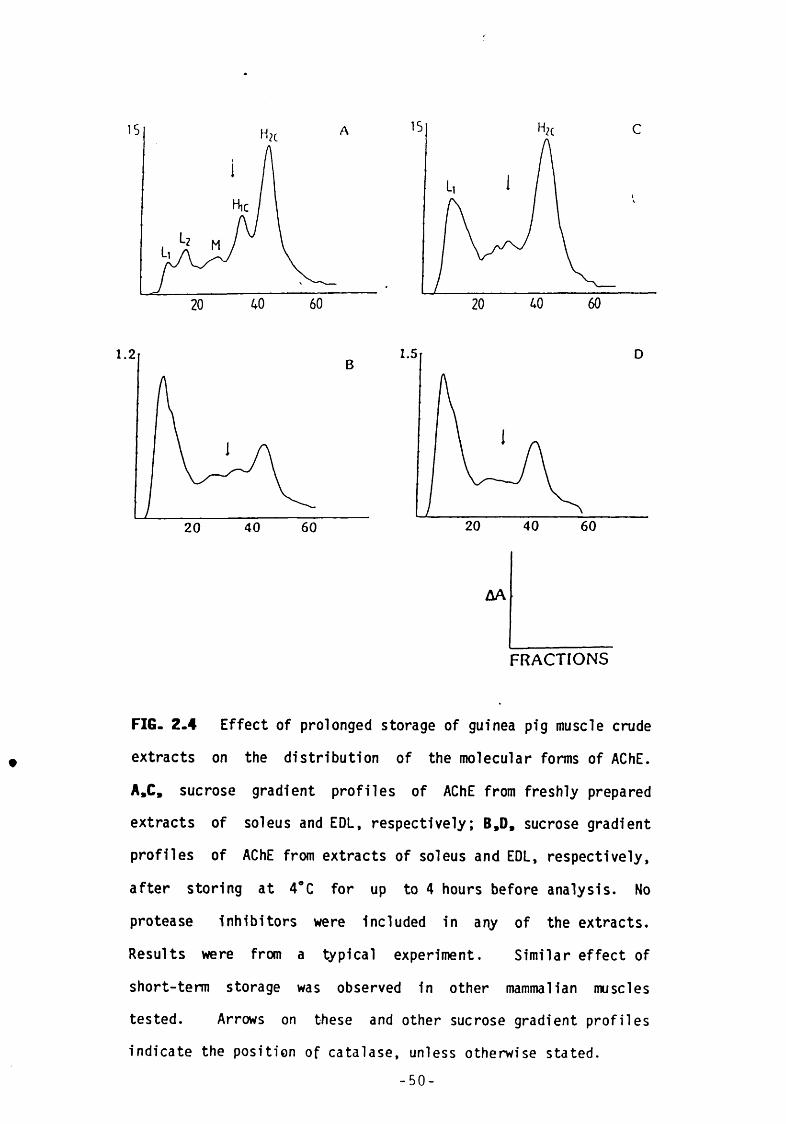
IS A 15 C
AA
FR A CTIO N S
FIG- 2-4 Effect of prolonged storage of guinea pig muscle crude
extracts on the distribution of the molecular forms of AChE.
A,C, sucrose gradient profiles of AChE from freshly prepared
extracts of soleus and EDL, respectively; B,D, sucrose gradient
profiles of AChE from extracts of soleus and EDL, respectively,
after storing at 4°C for up to 4 hours before analysis. No
protease inhibitors were included in any of the extracts.
Results were from a typical experiment. Similar effect of
short-term storage was observed in other mammalian muscles
tested. Arrows on these and other sucrose gradient profiles
indicate the position of catalase, unless otherwise stated.
- 5 0 -

*
-5 1 -

FIG- 2-5 P r o t e a s e i n h i b i t o r s s t a b i l i s e t h e d i s t r i b u t i o n o f AChE
fo rm s in m u s c le c r u d e e x t r a c t s . (A) s u c r o s e g r a d i e n t p r o f i l e o f
AChE i n r a t s o l e u s , p r e p a r e d 1n t h e a b s e n c e o f p r o t e a s e
i n h i b i t o r s and t h e e x t r a c t s t o r e d a t 4°C f o r 1 h o u r b e f o r e
g r a d i e n t a n a l y s i s . D o t t e d l i n e r e p r e s e n t s b a ck g ro u n d a b s o r b a n c e
o f s u c r o s e g r a d i e n t f r a c t i o n s w h ich w e r e i n c u b a t e d i n t h e
p r e s e n c e o f b o th isoOMPA (1 x 1 0 “^M)and BW 284C51 ( 5 x
10-*). (B) s u c r o s e g r a d i e n t p r o f i l e o f AChE i n r a t s o l e u s
p r e p a r e d i n t h e p r e s e n c e o f a f u l l a r r a y o f p r o t e a s e i n h i b i t o r s
( T a b l e 2 . 2 ) , and t h e e x t r a c t a n a l y s e d by g r a d i e n t e i t h e r
im m e d ia t e ly o r a f t e r s t o r i n g a t 4°C f o r 1 h o u r . Same am ount o f
e x t r a c t s was a n a l y s e d i n (A) and ( B ) . U n l e s s i n d i c a t e d ( a s in A ) ,
g r a d i e n t p r o f i l e s shown have b e e n c o r r e c t e d f o r b a c k g r o u n d
a b s o r b a n c e . R e p e a t e x p e r i m e n t s g a v e s i m i l a r r e s u l t s .

FRACTIONS
- 5 2 -

2 . 3 . 3 The effect of perfusion
B lo o d c o n t a i n s L and M fo rm s o f AChE ( s e e g e n e r a l
i n t r o d u c t i o n ) and serum p r o t e a s e s ( f o r r e v i e w , s e e L o r a n d ,1 9 7 6 ) .
P e r f u s i o n w as i n c l u d e d a s a p r e c a u t i o n i n t h e e x t r a c t i o n o f AChE
p r i m a r i l y t o rem ove b l o o d from m u s c le c a p i l l a r i e s . T h is
p r o c e d u r e h a s b e e n em p lo y e d p r e v i o u s l y i n t h e s t u d y o f c h i c k e n
m u s c l e s ( L y l e s e t a l_ . , 1 9 8 2 ) . H ow ever, when u s e d h e r e no
d i f f e r e n c e i n t h e AChE p r o f i l e s w as o b s e r v e d . B e c a u s e o f t h i s ,
and i n v i e w o f t h e f a c t t h a t p e r f u s i o n may in d u c e o s m o t i c damage
t o c a p i l l a r i e s and t h e s u r r o u n d in g t i s s u e , and t h a t f o r m o st
m u s c l e s t h e am ount o f c a p i l l a r y b l o o d * i s s m a l l , p e r f u s i o n was
s u b s e q u e n t l y o m i t t e d from t h e e x t r a c t i o n p r o c e d u r e .
* The re d c o l o u r o f m u s c l e s su ch a s t h e d iap h ragm i s l a r g e l y due
t o m y o g lo b in and m i t o c h o n d r i a l c y t o c h r o m e s i n t h e s lo w f i b r e
( G a u t h i e r , 1 9 6 9 ) and n o t t o b l o o d .
2 . 3 . 4 The effect of freezing nuscle tissues
T h is p r o c e d u r e h a s b een em p lo y e d in s e v e r a l l a b o r a t o r i e s
( G i s i g e r and S t e p h e n s , 1 9 8 3 ; Bacou e t _ a l . , 1 9 8 2 ) f o r t h e s t o r a g e
o f t i s s u e s b e f o r e e x t r a c t i o n . P r e s u m a b ly , a t v e r y lo w
t e m p e r a t u r e , t h e p r o t e a s e a c t i v i t y w o u ld b e i n s t a n t a n e o u s l y
• a r r e s t e d upon f r e e z i n g . T h is was t h e r e f o r e i n c l u d e d a s a
p r e c a u t i o n a r y p r o c e d u r e f o r e x t r a c t i n g AChE from r a t m u s c l e s ,
a l o n g w i t h r a p i d p r o c e s s i n g . H o w ev er , i t was fo u n d t h a t f r e e z i n g
d i d n o t a p p e a r t o make any d i f f e r e n c e in p r e s e r v i n g t h e H£C
AChE, p o s s i b l y b e c a u s e p r o t e o l y s i s was n o t e x t e n s i v e b e f o r e
h o m o g e n i s a t i o n .
In c o n c l u s i o n , i t w as e s t a b l i s h e d t h a t r a p i d p r o c e s s i n g o f
-53-

m u s c le c r u d e e x t r a c t s from f r e s h l y e x c i s e d m u s c l e s , a n d t h e u s e o f
an a p p r o p r i a t e c o m b in a t io n o f p r o t e a s e I n h i b i t o r s , w e r e t h e
s i m p l e s t w ays o f e n s u r i n g r e p r o d u c i b i l i t y In t h e vn v l t r o
a n a l y s i s o f AChE f o r m s .
SECTION II. Distribution of AChE torus in u u n l i a n and
avian skeletal uuscles
2 . 3 . 5 Molecular profiles of AChE in uauualian skeletal uuscles
The r e l a t i v e p r o p o r t i o n s o f AChE fo rm s In t h e v a r i o u s
m u s c l e s a n a l y s e d a r e l i s t e d In T a b le 2 . 1 .
A. F a s t - t w i t c h m u s c l e s
M u s c le s w h ich a r e com p osed p r e d o m in a n t ly o f t y p e I I f i b r e s
w ere a n a l y s e d f o r t h e i r AChE c o n t e n t ( T a b le 2 . 1 ) . T h e s e t y p e I I
m u s c l e s from h a m s t e r , g u i n e a p i g and r a b b i t show ed t h a t t h e r e
w ere two p r i n c i p a l t y p e s o f m o l e c u l a r p r o f i l e s . The f i r s t t y p e
c o n s i s t e d o f a b o u t e q u a l a m o u n ts o f t h e L an d H f o r m s , t h e
s e c o n d c o n s i s t e d p r e d o m in a n t ly o f t h e ^ fo r m . The p r o f i l e s
o f AChE fo rm s i n a l l t h e r a b b i t f a s t - t w i t c h m u s c l e s , a s w e l l a s
i n t h e EDL ( F i g . 2 .4 C ) an d t h e sem im em branosus a c c e s s o r i u s o f
t h e g u in e a p i g , a r e o f t h e fo r m e r t y p e . F i g u r e 2 . 6 i l l u s t r a t e s
• some r e p r e s e n t a t i v e p r o f i l e s o f t h i s t y p e o f d i s t r i b u t i o n .
The s e c o n d t y p e o f d i s t r i b u t i o n o f AChE fo rm s was e x h i b i t e d
by a l l t h e h a m s te r t y p e I I m u s c l e s and some o f t h e g u i n e a p i g
m u s c l e s ( F i g . 2 . 7 ) , w h ere AChE w as d o m in a n t . T h er e w a s no
d i f f e r e n c e i n t h e c o m p o s i t i o n o f AChE form s i n t y p e IIA a n d I IB
f i b r e s , a s shown by t h e r e d and w h i t e r e g i o n s o f t h e g u in e a p i g
v a s t u s m u s c l e . ( T a b le 2 . 1 ) .
- 5 4 -

FIG. 2-6 T y p ic a l s u c r o s e g r a d i e n t p r o f i l e s o f AChE fo rm s in
some mammalian t y p e I I m u s c l e s , A, r a b b i t EDL; B, r a b b i t AT; C,
g u in e a p i g sem i-m em b ra n o su s a c c e s s o r i u s . A l l e x t r a c t s w e r e
p r e p a r e d in t h e p r e s e n c e o f p r o t e a s e i n h i b i t o r s .
-55-

aa
FRACTIONS
FIG- 2.7 T y p ic a l s u c r o s e g r a d i e n t p r o f i l e s o f AChE fo rm s in
some mammalian t y p e I I m u s c l e s . A, h a m s te r AT; B, h a m s te r EDL;
C, g u i n e a p i g v a s t u s l a t e r a l i s ; D, g u i n e a p ig g a s t r o c n e m i u s . A l l
e x t r a c t s w ere p r e p a r e d i n t h e p r e s e n c e o f p r o t e a s e i n h i b i t o r s .
-56-

In c o n t r a s t , t h e t y p e I I m u s c l e s i n r a t and m ouse c o n t a i n e d
a s u b s t a n t i a l am ount o f L and M fo rm s ( F i g . 2 . 8 ) , t h e p r o p o r t i o n s
o f w h ic h r e m a in e d l a r g e e v e n when p r o t e a s e i n h i b i t o r s w e r e u se d
d u r in g e x t r a c t i o n .
B . S l o w - t w i t c h m u s c l e s
Type I m u s c l e s from t h e v a r i o u s mammalian s p e c i e s c l e a r l y
sh ow ed t h e p r e d o m in a n c e o f t h e H2 c form ( F i g . 2 . 9 ) . In
a d d i t i o n , t h e r e w as a l s o an i m p o r t a n t com p onent o f H^c f o r m ,
m o st p r o m in e n t i n t h e r a t s o l e u s ( F i g . 2 . 5 B ) , t o g e t h e r w i t h a
m in o r am ount o f t h e l o w e r m o l e c u l a r w e i g h t fo r m s . The r e l a t i v e
am ount o f t h e L fo rm s v a r i e d b e tw e e n m u s c l e s , r a n g i n g from 15%
( r a t s o l e u s ) t o 33% ( r a b b i t s o l e u s ) o f t o t a l a c t i v i t y . The M
form on t h e o t h e r hand a c c o u n t e d f o r a r e l a t i v e l y m in o r
p r o p o r t i o n o f t o t a l AChE a c t i v i t y . T h e s e AChE p r o f i l e s o f t y p e I
m u s c l e s a r e t h e r e f o r e v e r y s i m i l a r t o some p r o f i l e s o f t y p e I I
m u s c l e s d e s c r i b e d a b o v e ( F i g . 2 . 7 ) .
C. A n a l y s i s o f AChE fo rm s by m i c r o d i s s e c t i o n
S i n c e t h e a b u n d a n c e o f L fo rm s i n r a t m u s c l e s c o i n c i d e d w i t h
t h e e x c e p t i o n a l l y h ig h p r o t e o l y t i c a c t i v i t y fo u n d i n th em , i t was
e s s e n t i a l t o a s c e r t a i n w h e t h e r o r n o t t h e l a r g e p r o p o r t i o n o f L
form s was an a r t e f a c t o f p r o t e o l y s i s . A l t h o u g h p r o t e a s e
i n h i b i t o r s had b e e n shown t o r e d u c e t h e p r o t e o l y t i c a c t i v i t y in
r a t m u s c l e s t o v e r y lo w l e v e l s , i t d id n o t p r e c l u d e t h e
p o s s i b i l i t y o f some r e s i d u a l p r o t e a s e a c t i v i t y . AChE a n a l y s i s
was t h e r e f o r e c a r r i e d o u t u s i n g m i c r o d i s s e c t e d s a m p le s t o
m in i m is e c o n t a m i n a t i o n by e n d o g e n o u s p r o t e a s e s . The AChE c o n t e n t
-57-

FIG. 2.8 T y p ic a l s u c r o s e g r a d i e n t p r o f i l e s o f AChE fo rm s in
some mammalian t y p e I I m u s c l e s . A, r a t s t e r n o m a s t o i d ; B, r a t
EDL; C, m ouse AT. A l l e x t r a c t s w ere p r e p a r e d in t h e p r e s e n c e o f
p r o t e a s e i n h i b i t o r s .
-58-

FIG- 2-9 T y p ic a l s u c r o s e g r a d i e n t p r o f i l e s o f AChE form s in
some mammalian t y p e I m u s c l e s . A, h a m s te r s o l e u s ; B, r a b b i t
sem i-m em b ra n o su s p r o p r i u s ; C, g u in e a p i g sem i-m em b ra n o su s
p r o p r i u s . A l l e x t r a c t s w ere p r e p a r e d in t h e p r e s e n c e o f p r o t e a s e
i n h i b i t o r s .
-59-

o f w h o le m u s c le f i b r e s a s w e l l a s o f e n d p l a t e and n o n - e n d p l a t e
r e g i o n s w e r e a n a l y s e d .
U s in g t h e s t e r n o m a s t o i d and s o l e u s o f t h e r a t a s e x a m p le s o f
t y p e I I and t y p e I m u s c l e s r e s p e c t i v e l y , t h e p r o f i l e s o f AChE
fo rm s o b t a i n e d from w h o le m u s c l e f i b r e s w e r e v e r y s i m i l a r t o
t h o s e from w h o le m u s c le e x t r a c t s ( c f . F i g . 2 .1 0 A ,B and F i g . 2 .
8A , F i g . 2 . 5 B ) . The AChE p r o f i l e from t h e s t e r n o m a s t o i d
c o n s i s t e d o f a s u b s t a n t i a l am ount o f t h e L f o r m s , w h i l e t h e
s o l e u s c o n t a i n e d a much s m a l l e r L c o m p o n e n t , w i t h a b o u t eq u a l
a m o u n ts o f t h e H^c an d H2 c f o r m s . D i s s e c t i o n o f t h e e n d p l a t e
and n o n - e n d p l a t e r e g i o n s o f t h e s e m u s c l e s c o n f i r m e d t h e
e n d p l a t e - s p e c i f i c l o c a l i z a t i o n o f t h e H2c form a s o b s e r v e d
p r e v i o u s l y i n t y p e I I m u s c l e s ( J e d r z e j c z y k e t a l . , 1 9 8 1 ; F i g .
2 . 1 1 A , B ) . The l a r g e a m o u n ts o f L and M fo rm s in t h e
s t e r n o m a s t o i d w ere m a in ly l o c a t e d o u t s i d e t h e e n d p l a t e s ( F i g .
2 . 1 1 C ) . The l o w e r m o l e c u l a r w e i g h t fo rm s a t t h e e n d p l a t e s c o u l d
be c o n t a m i n a n t s from t h e n o n - e n d p l a t e r e g i o n s i n c l u d e d in t h e
s a m p le , e s p e c i a l l y s i n c e t h e AChE a c t i v i t y o u t s i d e t h e e n d p l a t e s
i n t h i s c a s e was h i g h . F o r t h e s l o w - t w i t c h s o l e u s ( F i g . 2 . 1 1
B , D , E ) , t h e form w a s fo u n d i n s i d e a s w e l l a s o u t s i d e t h e
e n d p l a t e s , a s i n t h e c a s e o f t h e g u in e a p i g s o l e u s ( s e e C h a p te r
• 3 , F i g . 3 .5 B and 3 .6 B ) and t h e r a b b i t sem im em b ran osu s p r o p r i u s
( J . J e d r z e j c z y k , p e r s o n a l c o m m u n i c a t i o n ) . T h i s e x t r a j u n c t i o n a l
H2C i n t h e r a t s o l e u s a p p e a r e d t o l o c a l i s e c l o s e t o t h e
e n d p l a t e r e g i o n , s i n c e r e g i o n s away from t h e e n d p l a t e s to w a r d t h e
e n d s o f t h e f i b r e s c o n t a i n v e r y much l e s s H2 c form ( F i g . 2 .
1 1 E ) . In t h e g u in e a p i g s o l e u s , h o w e v e r , t h e e x t r a j u n c t i o n a l
H2 c form w as fo u n d a l o n g t h e e n t i r e l e n g t h o f t h e f i b r e . The
- 6 0 -

4 0 A
FRACTIONSFIG- 2-10 M o l e c u l a r form s o f AChE from t y p e I an d t y p e I I m u s c le
f i b r e s . A, r a t s t e r n o m a s t o i d ( t y p e I I ) ; B, r a t s o l e u s ( t y p e I ) .
D o t t e d l i n e in (A) r e p r e s e n t s b a c k g r o u n d a b s o r b a n c e
o f s u c r o s e g r a d i e n t f r a c t i o n s i n c u b a t e d in t h e p r e s e n c e o f b o th
isoOMPA (1 x 1 0 _ 4M) an d BW284C51 ( 5 x 1 0 ’ ^M). A rrow s d e n o t e
t h e c o r r e s p o n d i n g p o s i t i o n s o f c a t a l a s e ( c a t ) and 8- g a l a c t o s i d a s e
( 8- g a l ) . B oth e x t r a c t s w ere p r e p a r e d i n t h e p r e s e n c e o f p r o t e a s e
i n h i b i t o r s . R e p e a t e x p e r im e n t s g a v e s i m i l a r p r o f i l e s .
- 6 1 -

FIG. 2.11 The d i s t r i b u t i o n o f m o l e c u l a r fo rm s o f AChE i n t h e
e n d p l a t e and n o n - e n d p l a t e r e g i o n s from t y p e I and t y p e I I
m u s c l e s . A,B, e n d - p l a t e r e g i o n o f r a t s t e r n o m a s t o i d ( t y p e I I )
and s o l e u s ( t y p e I ) , r e s p e c t i v e l y ; C, n o n - e n d p l a t e r e g i o n o f r a t
s t e r n o m a s t o i d ; D, p e r i j u n c t i o n a l r e g i o n o f r a t s o l e u s ; E,
n o n - e n d p l a t e r e g i o n o f r a t s o l e u s . D o t t e d l i n e d e n o t e s t h e
b a c k g r o u n d a b s o r b a n c e o f s u c r o s e g r a d i e n t f r a c t i o n s . The am ount
o f t i s s u e sa m p le u s e d i n e a c h a n a l y s i s was a r b i t r a r y , . A l l
e x t r a c t s w e r e p r e p a r e d i n t h e p r e s e n c e o f p r o t e a s e i n h i b i t o r s .
R e p e a t e x p e r i m e n t s g a v e s i m i l a r p r o f i l e s .
- 6 2 -

H^c fo r m , a m a jo r form o f t h e AChE i n t h e s o l e u s , w as a l s o
l o c a l i s e d o u t s i d e t h e e n d p l a t e s . T h e s e r e s u l t s s u g g e s t ,
t h e r e f o r e , t h a t t h e a n a l y s i s o f AChE form s by m i c r o d i s s e c t i o n
c o r r e l a t e d w e l l w i t h w h o le m u s c le e x t r a c t s . I t i s a l s o e v i d e n t
from t h e a n a l y s i s o f e n d p l a t e and n o n - e n d p l a t e AChE t h a t t h e h ig h
l e v e l o f L form s i n t h e s t e r n o m a s t o i d m u s c le i s n o t a d e g r a d a t o r y
p r o d u c t o f t h e 1 ^ form s i n c e t h e two a r e s p a t i a l l y s e p a r a t e .
H ow ever , t h e p o s s i b i l i t y t h a t some o f t h e L fo rm s w e r e breakdow n
p r o d u c t s o f t h e M form c o u l d n o t be r u l e d o u t .
A f e a t u r e o f t h e t y p e I m u s c l e s w h ich d i s t i n g u i s h e s them
from t y p e I I m u s c l e s i s t h e r e f o r e t h e p r e s e n c e o f e x t r a s y n a p t i c
p o p u l a t i o n o f and H£C f o r m s . F o r t y p e I a n d t y p e I I
m u s c l e s w h ic h have s i m i l a r HgQ and AChE c o n t e n t p e r u n i t w e i g h t
( T a b l e 2 . 3 ) , t h e am ount o f H£C form i n t h e e n d p l a t e s o f t y p e I
m u s c l e s w o u ld be l e s s th a n t h a t in t h e t y p e I I m u s c l e s s i n c e p a r t
o f t h e H£C AChE i n t h e f o r m e r i s d i s t r i b u t e d o v e r a l a r g e r
vo lu m e o f t h e m u s c l e .
2 . 3 . 6 Molecular profiles of AChE in avian ouseles
P r e v i o u s s t u d i e s show ed t h a t m u s c l e s o f m ix ed t y p e I and
t y p e IIA f i b r e s ( e . g . t h e s a r t o r i u s re d s e c t o r , 40% t y p e I , 60%
• t y p e I I ) c o n t a i n a h i g h e r l e v e l o f t h e L and Hl c AChE th a n a
p u r e t y p e I IB m u s c le su c h a s t h e PLD (B a rn a rd e t a l . . 1 9 8 2 ) . In
t h i s s t u d y t h e a n t e r i o r r e g i o n o f t h e a d d u c t o r p r o f u n d u s (31%
t y p e I , 69% t y p e I IA ) was a l s o fo u n d t o c o n t a i n a s i g n i f i c a n t
am ount o f t h e L form and a d i s t i n c t H^c co m p o n en t ( F i g . 2 . 1 2 ) ,
T h e s e fo rm s w e r e p r o b a b ly c h a r a c t e r i s t i c s o f t h e a v i a n t y p e I an d
IIA f i b r e s , s i n c e a p u r e f a s t - t w i t c h m u s c l e , su c h a s t h e PLD,
- 6 3 -

TABLE 2.3 AChE a ctiv ity in some mammalian muscles
Species Muscle Fibre type [AChE] (U/g) n
guinea pig EDL I I 0.79 ± 0.03 2
SAa 0.37 ± 0.04 7
Vastus la te ra lis (white) 0.45 1
Vastus la te ra lis (red) 0.71 1
Rabbit SA 0.49 ± 0.03 7
Rat EDL 1.03 ± 0.0 2
Sternomastold (red) 1.00 ± 0.02 2
Sternomastoid (white) 1.17 ± 0.12 2
Guinea pig Soleus I 0.49 ± 0.06 5
SPb 0.56 ± 0.04 6
Plantarls 0.41 1
Rabbi t Soleus 0.6 ± 0.07 6
SP 0.55 ± 0.07 7
Rat Soleus 0.31 ± 0.02 2
Rat Diaphragm I + I I 0.69 ± 0.02 2
a Semimembranosus accessorius (see also chapter 3),
b Semimembranosus proprlus (see also chapter 3),
The a c t iv ity of AChE 1s expressed as units per gram tissue wet weight, and
the standard error 1s indicated for n number of determinations.
- 6 4 -

FIG- 2-12 M o l e c u l a r form s o f AChE i n t h e a n t e r i o r and p o s t e r i o r
r e g i o n s o f t h e c h i c k e n a d d u c t o r p r o f u n d u s . Dotted line: s u c r o s e
g r a d i e n t p r o f i l e o f AChE in t h e a n t e r i o r r e g i o n , Solid line:
s u c r o s e g r a d i e n t p r o f i l e o f AChE in t h e p o s t e r i o r r e g i o n .
E x t r a c t s w e r e p r e p a r e d i n t h e p r e s e n c e o f p r o t e a s e i n h i b i t o r s
( T a b l e 2 . 2 ) . R e s u l t s w e r e from a t y p i c a l e x p e r i m e n t .
-65-

d o e s n o t c o n t a i n s i g n i f i c a n t am ou n ts o f t h e s e fo rm s ( F i g . 2 .1 3 ) .
T h i s f i n d i n g i n c h i c k e n m u s c l e s i s c l e a r l y d i f f e r e n t from t h a t in
mammalian m u s c l e s . The t y p e I I I o r t o n i c m u s c l e , su ch a s t h e ALD
( L y l e s e t __ a l . . , 1 9 8 1 ) and t h e p o s t e r i o r r e g i o n o f t h e a d d u c t o r
p r o f u n d u s ( F i g . 2 . 1 2 ) c o n s i s t e d o f L^ and M fo r m s , w i t h v e r y
l i t t l e H£C fo r m .
2 . 3 . 7 Molecular profiles of AChE in anscles of Mixed fibre type
M u s c le s o f m ix ed f i b r e t y p e c o m p o s i t i o n , su ch a s t h e r a t
d ia p h r a g m , w e r e s t u d i e d . In t h i s mammalian m u s c l e , a b o u t eq u a l
am ou n ts o f L, M and H fo rm s w e r e fo u n d ( F i g . 2 . 1 4 ) . The l a r g e L
and M c o m p o n e n ts w e r e p r o b a b ly c o n t r i b u t e d by t h e t y p e I I f i b r e s ,
i f t h e y w ere a ssu m ed t o h a v e a s i m i l a r d i s t r i b u t i o n o f form s a s
t h e s t e r n o m a s t o i d o r t h e EDL. A n a l y s i s o f e n d p l a t e and
n o n - e n d p l a t e AChE a g a i n r e v e a l e d t h e form a s t h e j u n c t i o n a l
co m p o n en t o f AChE, w h i l e t h e L an d M fo rm s w ere r e s t r i c t e d t o t h e
e x t r a j u n c t i o n a l r e g i o n s ( F i g . 2 . 1 5 ) .
2 . 4 DISCUSSION
2 . 4 . 1 Autolysis of AChE in tissue crude extracts
The t e c h n i q u e o f s e d i m e n t a t i o n a n a l y s i s h a s b e e n u s e d
e x t e n s i v e l y f o r t h e s tu d y o f m u l t i p l e m o l e c u l a r form s o f AChE i n
t i s s u e e x t r a c t s . The p r e s e n t r e s u l t s show t h a t t h e r e l a t i v e
p r o p o r t i o n s o f t h e s e form s c o u l d i n a d v e r t e n t l y be g r e a t l y
a l t e r e d i n v i t r o by t h e a c t i v i t i e s o f e n d o g e n o u s p r o t e a s e s , b u t
t h e p ro b lem c o u l d l a r g e l y be a v o i d e d by p r o c e s s i n g c r u d e e x t r a c t s
r a p i d l y in t h e p r e s e n c e o f a s u i t a b l e a r r a y o f p r o t e a s e
i n h i b i t o r s .
-66-

1.0
F IG - 2 - 1 3 S u c r o s e g r a d i e n t p r o f i l e o f AChE in t h e c h i c k e n PLD
m u s c l e .
- 6 7 -

10 3 0 5 0
Fractions
FIG. 2.14 S u c r o s e g r a d i e n t p r o f i l e s o f AChE i n some m u s c l e s o f
m ix e d f i b r e t y p e c o m p o s i t i o n . A, r a t d ia p h ra g m ; B, m ou se s o l e u s .
E x t r a c t s w ere p r e p a r e d i n t h e p r e s e n c e o f p r o t e a s e i n h i b i t o r s .
R e p e a t e x p e r i m e n t s showed s i m i l a r d i s t r i b u t i o n o f m o l e c u l a r
f o r m s .
-68-

FIG- 2-15 D i s t r i b u t i o n o f m o l e c u l a r forms o f AChE i n t h e
e n d p l a t e and n o n - e n d p l a t e r e g i o n s o f t h e r a t d i a p h r a g m . A,
s u c r o s e g r a d i e n t p r o f i l e o f AChE i n t h e e n d p l a t e ; B, s u c r o s e
g r a d i e n t p r o f i l e s o f AChE o u t s i d e t h e e n d p l a t e s . R e s u l t s o f J .
• J e d r z e j c z y k .
-69-

The breakdown o f t h e m u l t i - s u b u n i t AChE fo rm s i n e l e c t r i c
o r g a n t i s s u e e x t r a c t was f i r s t r e p o r t e d by M a s s o u l i e a n d R i e g e r
( 1 9 6 9 ) . L a t e r , R i e g e r a nd V i g n y ( 1 9 7 6 ) n o t e d a t i m e - d e p e n d e n t
r e d u c t i o n o f t h e a n d M fo rm s and a p r o p o r t i o n a l i n c r e a s e
o f L f o r m s i n r a t d ia p h r a g m e x t r a c t s o v e r a p e r i o d o f s e v e r a l
d a y s , a t room t e m p e r a t u r e , and r e g a r d e d t h e s e c h a n g e s t o o s l o w t o
h a v e a n y s i g n i f i c a n t e f f e c t on t h e a n a l y s i s o f AChE f o r m s ,
n o r m a l l y c o m p l e t e d w i t h i n 1 d a y . Rotun do and Fambrough ( 1 9 7 9 )
o b s e r v e d t h a t s t o r i n g c h i c k m u s c l e c r u d e e x t r a c t s l e d t o a r a p i d
l o s s o f H f o rm s i n t h e s e e x t r a c t s , w h i l e i s o l a t e d m o l e c u l a r fo rm s
r e t a i n e d t h e i r s e d i m e n t a t i o n c h a r a c t e r i s t i c s f o r a l o n g t i m e ,
w h e t h e r s t o r e d f r o z e n a t - 2 0 ° C o r i n s o l u t i o n a t 5 ° C . S i l m a n e t
a l . ( 1 9 7 8 ) showed t h a t r e p e a t e d f r e e z i n g and t h a w i n g o f m u s c l e
e x t r a c t s , o r f r e e z e - s t o r i n g t i s s u e s a l w a y s g i v e r i s e t o some
d e g r e e o f breakdown o f t h e f o rm ; t h e y f u r t h e r showed t h a t a
m i x t u r e o f p r o t e a s e i n h i b i t o r s c o u l d p r e v e n t t h e breakdow n o f t h e
h i g h e r m o l e c u l a r w e i g h t f o rm s d u r i n g e x t r a c t i o n (1 h o u r a t 3 8 °C )
from f r e s h c h i c k e n m u s c l e . T h e s e r e s u l t s c l e a r l y show t h a t t h e
c o n v e r s i o n o f t h e AChE f o rm s i n v i t r o i s n o t due t o a s p o n t a n e o u s
i n t e r c o n v e r s i o n o f t h e s e fo rm s, b u t a r e s u l t o f t h e br eakdo w n o f
t h e m u l t i - s u b u n i t c o m p le x i n p a r t by e n d o g e n o u s p r o t e o l y t i c
• d e g r a d a t i o n .
2 . 4 . 2 Correlation of the degradation of the H2c fora with the
proteolytic activity in tissue crude extracts
The t i m e c o u r s e o f t h e i n v i t r o breakdown o f t h e H f o r m s i s
n o t c l e a r b e c a u s e t h e e x p e r i m e n t a l c o n d i t i o n s and t h e t i s s u e
e x t r a c t s u s e d i n v a r i o u s s t u d i e s d i f f e r e d w i d e l y . H o w e v e r , a s
- 7 0 -

t h i s s t u d y h a s shown , t h e r a t e a t w h ic h su ch breakdown t a k e s
p l a c e d u r i n g e x t r a c t i o n h a s b een g r e a t l y u n d e r e s t i m a t e d ,
e s p e c i a l l y i n t h e c a s e o f AChE i n r a t m u s c l e e x t r a c t s .
S u b s t a n t i a l breakdo wn o f H fo rm s o c c u r r e d w i t h i n 1 ho ur a t 4 ° C ,
w h e t h e r i t was k e p t a s c r u d e e x t r a c t ( F i g . 2 . 5 A ) o r a s h o m o g e n a te
( r a t d i a p h r a g m , n o t s h o w n ) . For g u i n e a p i g m u s c l e s , r a p i d
p r o c e s s i n g a l o n e may i n c r e a s e t h e p r o p o r t i o n o f H fo rm s o b s e r v e d ,
b u t t h e m o l e c u l a r p r o f i l e i s h i g h l y v a r i a b l e . The c o n v e r s i o n
from H fo rm s t o L fo rm s may v a r y w i t h t h e p r o t e o l y t i c a c t i v i t y
p r e s e n t i n d i f f e r e n t t i s s u e e x t r a c t s a n d , d e p e n d i n g on s u c h
a c t i v i t y and t h e t i m e t h e H fo rm s a r e e x p o s e d t o t h e p r o t e a s e s ,
w o u ld g i v e r i s e t o a g r e a t v a r i e t y o f m o l e c u l a r p r o f i l e s w h ic h do
n o t r e p r e s e n t t h e d i s t r i b u t i o n i n v i v o ,
2 . 4 . 3 Inhibition of proteolysis stabilised the Molecular
distribution of AChE forus
A v a r i e t y o f p r o t e a s e i n h i b i t o r s w i t h w e l l - d e f i n e d
s p e c i f i c i t y w e r e u s e d i n c o m b i n a t i o n t o i n h i b i t a l l known
p r o t e a s e s w h ic h c o u l d be p r e s e n t i n m u s c l e e x t r a c t s . S e r i n e
p r o t e a s e s a c c o u n t f o r a s i g n i f i c a n t p r o p o r t i o n o f p r o t e a s e
a c t i v i t y i n r a t m u s c l e s (Katnuma and Kominami , 1 9 7 7 ) , p r o b a b l y
• o f m a s t c e l l o r i g i n ( L a g u n o f f and B e n d i t t , 1 9 6 3 ) . A number o f
s e r i n e p r o t e a s e i n h i b i t o r s w h ich do n o t e x h i b i t i n h i b i t o r y
a c t i v i t y on AChE w e r e u s e d i n a d d i t i o n t o e n s u r e t h e i n h i b i t i o n
o f t h e s e p r o t e a s e s , w h ic h may e x h i b i t d i f f e r e n t i n h i b i t o r
s p e c i f i c i t y i n d i f f e r e n t s p e c i e s ( G l e n n e r a nd C oh en , 1 9 6 0 ) . The
c o n c e n t r a t i o n o f t h e s e i n h i b i t o r s r e q u i r e d f o r maximum i n h i b i t i o n
was h i g h , a s e x e m p l i f i e d by t h e d i f f e r e n c e i n t h e amount o f SVI
- 7 1 -

r e q u i r e d f o r 50% i n h i b i t i o n o f b o v i n e a - c h y m o t r y p s i n and o f r a t
m u s c l e p r o t e a s e s ( c . f . F i g . 2 .2 C and F i g . 2 . 3 ) , H o w ev er ,
O g - m a c r o g l o b u l i n a p p e a r e d t o be an e x c e p t i o n : a t a
c o n c e n t r a t i o n o f O . lm g/m l ( 0 . 1 4 p M ) , i t r e d u c e d t h e r a t m u s c l e
p r o t e a s e a c t i v i t y t o m inim al l e v e l . The a p p a r e n t r e c o v e r y o f
p r o t e a s e a c t i v i t y i s p r o b a b l y due t o r e s i d u a l p r o t e a s e and n o t t o
r e v e r s i b l e i n h i b i t i o n , a s p r e v i o u s e x p e r i m e n t s ha ve d e m o n s t r a t e d
t h e i r r e v e r s i b l e n a t u r e o f t h e a g - m a c r o g l o b u l i n - e n z y m e c o m p le x
( B a r r e t t and S t a r k e y , 1 9 7 3 ; S a u n d e r s e t a l . , 1 9 7 1 ) . R e s i d u a l
p r o t e a s e a c t i v i t y was o f t e n a c c o m p a n ie d by a d i s c r e p a n c y i n t h e
o b s e r v e d AChE p r o f i l e s , s u g g e s t i n g t h a t e v e n l o w p r o t e a s e
a c t i v i t y p r e s e n t i n m u s c l e h o m o g e n a t e s c o u l d be s u f f i c i e n t t o
a l t e r t h e r e l a t i v e p r o p o r t i o n o f AChE fo rm s i n v i t r o , and t h a t
t h e d e g r a d a t i o n o f t h e s e fo rm s p r o b a b l y i n v o l v e d a v a r i e t y o f
p r o t e a s e s i n t h e e x t r a c t s . T h i s w o u ld a l s o e x p l a i n t h e h i g h
l e v e l o f L fo rm s o b s e r v e d i n p r e v i o u s i n v e s t i g a t i o n s i n w h ic h
some p r o t e a s e i n h i b i t o r s w ere e m p lo y e d ( G i s i g e r a n d S t e p h e n s ,
1 9 8 3 ; Bacou e t __ a L , 1 9 8 2 ) , s i n c e u n s u i t a b l e o r i n s u f f i c i e n t
i n h i b i t o r s w ou ld be i n e f f e c t i v e . Hence c o m p l e t e i n h i b i t i o n o f
p r o t e a s e a c t i v i t y i s i m p o r t a n t f o r t h e a n a l y s i s o f AChE f o r m s .
# 2 . 4 . 4 . Tiae course and nechanisa of ACHE degradation
The p r o c e s s o f h o m o g e n i s a t i o n f a c i l i t a t e s p r o t e o l y s i s by
e x p o s i n g AChE t o t h e p r o t e a s e s ; t h u s t h e breakdown o f H fo rm s i n
c r u d e e x t r a c t s w o u ld t a k e p l a c e more r e a d i l y t h a n w o u ld o c c u r i n ,
s a y , f r o z e n i n t a c t t i s s u e s . I n i t i a l p e r f u s i o n o r r a p i d f r e e z i n g
b e f o r e h o m o g e n i s a t i o n , t h e r e f o r e , h a d l i t t l e e f f e c t i n p r e v e n t i n g
s u c h br ea k d o w n . The r a p i d i t y w i t h w h ic h t h e t r a n s f o r m a t i o n o f H
- 7 2 -

t o L fo rm s o c c u r r e d d u r i n g t h e f i r s t few h o u r s o f e x t r a c t i o n
c o n t r a s t s w i t h t h e much s l o w e r r a t e o f c o n v e r s i o n o f t h e M f o rm s
t o L f o r m s o b s e r v e d by R i e g e r and V i g n y ( 1 9 7 6 ) . Such d i f f e r e n c e s
c o u l d be due t o t h e i n s t a b i l i t y o r d e g r a d a t i o n o f t h e p r o t e a s e s
t h e m s e l v e s w i t h l o n g i n c u b a t i o n . M o r e o v e r , t h e d i f f e r e n c e i n
t h i s i n i t i a l p r o t e o l y t i c a c t i v i t y o b s e r v e d i n d i f f e r e n t s p e c i e s ,
a p a r t from t h e a c t u a l amount and v a r i e t y o f p r o t e a s e s p r e s e n t ,
may a l s o be i n f l u e n c e d by t h e i r e n z y m i c s t a t e when t h e y a r e
m e c h a n i c a l l y r e l e a s e d : e v i d e n c e s u g g e s t s t h a t r a t m a s t c e l l
p r o t e a s e s a r e s t o r e d i n t h e i r a c t i v e fo rm s i n t h e g r a n u l e s ( C h iu
and L a g u n o f f , 1 9 7 2 ) w h i l e g u i n e a p i g m a s t c e l l p r o t e a s e s a r e
s t o r e d a s z y m o g e n s (U n gar a n d Damgaard, 1 9 5 5 ) .
The q u e s t i o n o f how t h e c o l l a g e n - t a i l e d o r g l o b u l a r fo rm s
a r e b r o k e n down i n v i t r o r e q u i r e s t h e s t r u c t u r a l e l u c i d a t i o n o f
t h e c a t a l y t i c s u b u n i t s and t h e i r a s s o c i a t i o n t o form s t a b l e
q u a t e r n a r y p o l y m e r s . The g l o b u l a r s u b u n i t s a p p e a r t o a s s o c i a t e
w i t h e a c h o t h e r a s w e l l a s w i t h t h e c o l l a g e n - t a i l v i a d i s u l p h i d e
b r i d g e s ( s e e G e n e r a l I n t r o d u c t i o n ) . In a d d i t i o n , e v i d e n c e
s u g g e s t s t h a t s u c h q u a t e r n a r y a s s o c i a t i o n a l s o i n v o l v e s
n o n - c o v a l e n t i n t e r - s u b u n i t i n t e r a c t i o n , s i n c e s u b u n i t
d i s s o c i a t i o n w o u ld n o t o c c u r e v e n a f t e r d i s u l p h i d e r e d u c t i o n
• u n l e s s t h e enzym e i s s u b j e c t e d t o d e n a t u r a t i o n o r l i m i t e d
p r o t e o l y t i c d i g e s t (Bon and M a s s o u l i e , 1 9 7 6 ; Mays and R o s e n b e r r y ,
1 9 8 1 ; G r a s s i e t __ a l . , 1 9 8 2 ; Lee e t __ a l _ . , 1 9 8 2 ) . I t may b e
e n i v s a g e d , t h e r e f o r e , t h a t t h e d i s s o c i a t i o n o f t h e s u b u n i t s from
e a c h o t h e r r e q u i r e s some e n z y m i c c l e a v a g e on t h e p o l y p e p t i d e
b a c k b o n e a t a s i t e w h ic h w o u ld s e p a r a t e t h e b u l k o f t h e c a t a l y t i c
s u b u n i t from t h e r e g i o n w h ic h c o n t a i n s t h e h a l f c y s t e i n e a nd
- 7 3 -

w h i c h i s i n v o l v e d i n t h e i n t e r s u b u n i t i n t e r a c t i o n . Such e n z y m i c
c l e a v a g e i s l i k e l y t o r e l e a s e t h e L fo rm s from t h e M a n d H f o r m s ,
w h ic h i s m o s t commonly a s s o c i a t e d w i t h t h e breakdow n o f
Hg AChE. F u r t h e r m o r e , su ch e n z y m i c c l e a v a g e c a n a l s o o p e r a t e
b e t w e e n t h e g l o b u l a r s u b u n i t s and t h e c o l l a g e n t a i l , g i v i n g r i s e
t o t h e g l o b u l a r M f o r m . L i m i t e d t r y p s i n d i g e s t , f o r e x a m p l e , h a s
b e e n shown t o d e g r a d e t h e ^ form from c h i c k m u s c l e ( R o tu n d o
and Fambrough, 1 9 7 9 ) and e l e c t r i c o r g a n ( M a s s o u l i § a n d R i e g e r ,
1 9 6 9 ) i n a s e q u e n t i a l m an n er , y i e l d i n g L and M f o r m s . The
r e s u l t s p r e s e n t e d i n t h i s c h a p t e r show t h a t i n mammalian m u s c l e
t i s s u e s , t h e d i s s o c i a t i o n o f t h e g l o b u l a r s u b u n i t s from H AChE
o c c u r s r a p i d l y i n v i t r o , s u g g e s t i n g t h a t c e r t a i n r e g i o n s on t h e
p o l y p e p t i d e i n v o l v e d i n q u a t e r n a r y i n t e r a c t i o n s may be
s u s c e p t i b l e t o a t t a c k s by e n d o g e n o u s e n d o p e p t i d a s e . T h a t t h e
s u b u n i t s w h ic h a r i s e from d e g r a d a t i o n a r e i n d i s t i n g u i s h a b l e from
t h e n a t i v e L and M f o rm s by t h e i r s e d i m e n t a t i o n c o e f f i c i e n t s
s u g g e s t s t h a t d e g r a d a t i o n d o e s n o t r e s u l t i n any e x t e n s i v e l o s s
o f t h e s u b u n i t m o l e c u l a r s i z e o r c a t a l y t i c a c t i v i t y . I n d e e d ,
t r y p s i n t r e a t m e n t o f t h e t e t r a m e r i c human serum AChE g e n e r a t e s
a c t i v e monomers w i t h a v e r y s m a l l c h a n g e i n t h e a p p a r e n t
m o l e c u l a r w e i g h t o f t h e f u l l y r e d u c e d c a t a l y t i c s u b u n i t
( L o c k r i d g e and La Du, 1 9 8 2 ) . C t e r m i n a l s e q u e n c i n g o f p u r i f i e d
human serum AChE by L o c k r i d g e ( 1 9 8 4 ) sh ow s t h a t
a c y s t e i n e r e s i d u e p r e s e n t c l o s e t o t h e C t e r m i n u s may b e
i n v o l v e d i n t h e i n t e r s u b u n i t d i s u l p h i d e b o n d i n g . I t i s
t h e r e f o r e , r e a s o n a b l e t o s u g g e s t t h a t t h e p r o x i m i t y o f t h e
d i s u l p h i d e bond t o t h e C t e r m i n u s , and i t s a c c e s s i b i l i t y t o
p r o t e o l y t i c d e g r a d a t i o n c o u l d d e t e r m i n e t h e s u s c e p t i b i l i t y o f t h e
- 7 4 -

m o l e c u l a r fo rm s t o breakdown i n v i t r o . Such s p e c u l a t i o n c a n o n l y
b e v e r i f i e d by t h e e l u c i d a t i o n o f t h e s u b u n i t p e p t i d e s e q u e n c e o f
AChE i n v a r i o u s s p e c i e s .
2 . 4 . 5 is a dominant component of M i l fan skeletal ouscle
AChE
S k e l e t a l m u s c l e s from a number o f mammalian s p e c i e s ha v e
r e v e a l e d t h a t t h e H2 c form c o n s t i t u t e s a s i g n i f i c a n t p r o p o r t i o n
(30%-71%) o f t o t a l AChE a c t i v i t y i n b o t h f a s t - and s l o w - t w i t c h
m u s c l e s ( T a b l e 2 . 1 ) e x c e p t i n r a t f a s t - t w i t c h m u s c l e s w h e r e t h e
am ou nt o f Hg d o e s n o t e x c e e d 20% o f t o t a l AChE. A g r o s s
u n d e r e s t i m a t i o n o f t h e H2 c p o p u l a t i o n i n mammalian m u s c l e s h as
b e e n a r e s u l t o f e x p e r i m e n t s p e f o r m e d i n t h e a b s e n c e o f p r o t e a s e
i n h i b i t o r s l e a d i n g t o a r a p i d breakdown o f H2 c , e s p e c i a l l y i n
r a t m u s c l e s . I t i s a l s o due t o t h e l a c k o f a c o m p r e h e n s i v e s t u d y
o f t h e d i s t r i b u t i o n o f AChE f o rm s i n m u s c l e s from d i f f e r e n t
s p e c i e s , and b a s i n g o b s e r v a t i o n s m o s t l y on r a t f a s t - t w i t c h
m u s c l e s w h o se AChE p r o f i l e s a r e a t y p i c a l i n c o n t a i n i n g a l a r g e
p r o p o r t i o n o f L f o r m s .
2 . 4 . 6 AChE profiles in fast- and slow-twitch wuscles
S l o w - t w i t c h m u s c l e s w h ic h a r e co m p o se d p r e d o m i n a n t l y o f t y p e
I f i b r e s a l l e x h i b i t a d o m in a n t p r o p o r t i o n o f H^AChE and a
l a r g e , d i s t i n c t H^c c o m p o n e n t . No s i g n i f i c a n t d i f f e r e n c e
b e t w e e n t h e s e p r o f i l e s , e x c e p t t h e somewhat v a r i a b l e H^c
c o m p o n e n t , i s a p p a r e n t . On t h e o t h e r h a n d , t h e f a s t - t w i t c h
m u s c l e s w h ic h c o n s i s t l a r g e l y o f t y p e I I f i b r e s r e v e a l a more
v a r i e d and a p p a r e n t l y s p e c i e s - s p e c i f i c d i s t r i b u t i o n o f t h e AChE
- 7 5 -

f o r m s . A g e n e r a l p a t t e r n o f d i s t r i b u t i o n i s p r e v a l e n t i n f a s t
m u s c l e s from t h e same s p e c i e s b u t may d i f f e r b e t w e e n t h e same
m u s c l e from d i f f e r e n t s p e c i e s . Thus a l l r a b b i t f a s t m u s c l e s ha v e
e q u a l a m o u n ts o f L and H f o r m s ; h a m s t e r f a s t m u s c l e s a l w a y s
h a v e a p r e d o m i n a n t H^ c o m p o n e n t w h i l e t h a t i n r a t f a s t m u s c l e s
i s s m a l l . G uin ea p i g f a s t m u s c l e s h a v e a s i m i l a r AChE
d i s t r i b u t i o n a s t h e h a m s t e r f a s t m u s c l e s e x c e p t t h e EDL and t h e
s e m i- m e m b r a n o su s a c c e s s o r i u s w h ic h r e s e m b l e more c l o s e l y t h a t o f
t h e r a b b i t f a s t m u s c l e s . I n t e r - s p e c i e s d i f f e r e n c e s a r e m o s t
s t r i k i n g l y i l l u s t r a t e d by t h e d i f f e r e n t d i s t r i b u t i o n o f AChE
f o rm s i n t h e EDL o f t h e r a t ( F i g . 2 . 8B) a n d g u i n e a p i g ( F i g .
2 . 4 C ) d e s p i t e t h e s i m i l a r i t y b e t w e e n t h e i r f i b r e t y p e
c o m p o s i t i o n . F o r a p a r t i c u l a r s p e c i e s , m u s c l e s w i t h d i f f e r e n t
p r o p o r t i o n s o f t y p e IIA and I IB f i b r e s e x h i b i t s i m i l a r
d i s t r i b u t i o n o f AChE f o r m s , a s shown by t h e two r e g i o n s o f t h e
g u i n e a p i g v a s t u s l a t e r a l i s . T h i s s u g g e s t s t h a t t h e d i s t r i b u t i o n
o f AChE fo rm s i s n o t s i m p l y r e l a t e d t o t h e m e t a b o l i c p r o p e r t i e s
o f m u s c l e f i b r e s . F o r g u i n e a p i g and h a m s t e r , s i m i l a r p r o f i l e s
o f AChE fo rm s a r e f o u n d i n t h e f a s t - and s l o w - t w i t c h m u s c l e s ,
w h e r e a s d i s t i n c t p a t t e r n s o f d i s t r i b u t i o n o f t h e s e fo rm s a r e
a s s o c i a t e d w i t h t h e two f i b r e t y p e s i n r a b b i t ,
mouse and r a t . In m u s c l e s o f m ixe d f i b r e t y p e s s u c h a s t h e r a t
d i a p h r a g m , t h e AChE p r o f i l e i s c h a r a c t e r i s t i c o f a m i x t u r e o f t h e
c o n s t i t u e n t t y p e s , i . e . t y p e I a nd I I f i b r e s . Taken t o g e t h e r ,
t h e s e r e s u l t s s u g g e s t t h a t t h e d i s t r i b u t i o n o f AChE fo rm s
c o r r e l a t e s w i t h t h e f i b r e t y p e c o m p o s i t i o n o f t h e m u s c l e s ;
h o w e v e r , t h e two f i b r e t y p e s do n o t n e c e s s a r i l y h a v e d i f f e r e n t
AChE p r o f i l e s , a s s e e n i n g u i n e a p i g and h a m s t e r s . Where a
- 7 6 -

d i f f e r e n c e i s o b s e r v e d , i t i s m o s t l y due t o a l a r g e r p r o p o r t i o n
o f L f o rm s i n f a s t m u s c l e s r e l a t i v e t o t h a t i n s l o w m u s c l e s . The
p h y s i o l o g i c a l s i g n i f i c a n c e o f t h i s v a r y i n g am ount o f L fo rm s i n
d i f f e r e n t m u s c l e s i s n o t c l e a r ; h o w e v e r , some s p e c u l a t i o n f o r
t h e i r v a r i a b i l i t y i s s u g g e s t e d b e l o w i n s e c t i o n 2 . 4 . 8 . In t h e
c a s e o f r a t f a s t m u s c l e s , a l t h o u g h t h e H2 c form i s a m in o r AChE
c o m p o n e n t , t h e a c t u a l a c t i v i t y i t c o n s t i t u t e s i s s i m i l a r t o t h a t
i n t h e r a t s o l e u s , w h er e H2 c i s m a j o r . T h i s i s due t o t h e
e x c e p t i o n a l l y h i g h l e v e l o f AChE i n t h e f a s t m u s c l e s o f t h e r a t
( T a b l e 2 . 3 ) . Thus t h e H2 c a c t i v i t i e s a r e c o m p a r a b l e i n a l l t h e
m u s c l e s t h a t h a v e b e e n s t u d i e d d e s p i t e t h e p r e s e n c e o f d i f f e r e n t
p r o p o r t i o n s o f t h e l o w e r m o l e c u l a r w e i g h t f o rm s i n d i f f e r e n t
m u s c l e s
2 . 4 . 7 AChE profiles in chicken twitch and tonic auscles
C h ic k e n s k e l e t a l m u s c l e s p r o v i d e f u r t h e r e v i d e n c e f o r t h e
c o r r e l a t i o n b e t w e e n t h e d i s t r i b u t i o n o f AChE fo rm s and t h e f i b r e
t y p e c o m p o s i t i o n o f m u s c l e s . As ha s b e e n m e n t i o n e d a b o v e ,
t y p i c a l f a s t - t w i t c h m u s c l e s suc h a s t h e PLD c o n t a i n l a r g e l y H2c
( F i g . 2 . 1 3 ) , w h i l e t h e t o n i c ALD m u s c l e c o n t a i n s l a r g e l y L and M
f o r m s , w i t h a m in o r H2 c c o m p o n e n t t h a t d i m i n i i i s h e s w i t h
m a t u r a t i o n ( L y l e s and B a r n a r d , 1 9 8 0 ) . T h i s d i f f e r e n t i a l
d i s t r i b u t i o n o f AChE i s a l s o s e e n i n t h e a d d u c t o r p r o f u n d u s
m u s c l e , w h ic h i s d i v i d e d i n t o an a n t e r i o r r e g i o n o f t w i t c h f i b r e s
and a p o s t e r i o r r e g i o n o f t o n i c f i b r e s ( T a b l e 2 . 1 ) . The AChE
fo rm s e x h i b i t e d by t h e two r e g i o n s o f t h i s m u s c l e c o r r e l a t e w e l l
w i t h t h e i r r e s p e c t i v e f i b r e t y p e c h a r a c t e r i s t i c s ( F i g . 2 . 1 2 ) .
- 7 7 -

2 . 4 8 Non-endplate ACHE in type I and type II fibres
The o t h e r f e a t u r e w h ic h d i s t i n g u i s h e s t y p e I f i b r e s from
t y p e I I f i b r e s i s t h e p r e d o m in a n c e o f e x t r a j u n c t i o n a l H^ f o rm s
i n t y p e I f i b r e s , w h e r e a s L and M fo rm s a r e m o s t common o u t s i d e
t h e e n d p l a t e s o f t y p e I I f i b r e s ( F i g . 2 . 1 1 , F i g . 2 . 1 5 and C h a p t e r
3 , F i g s . 3 . 5 a n d 3 . 6 ) . T h e s e d i f f e r e n c e s i n t h e e x t r a j u n c t i o n a l
AChE b e t w e e n t h e two f i b r e t y p e s a r e t h u s r e s p o n s i b l e f o r t h e
v a r i a t i o n s i n t h e o v e r a l l AChE p r o f i l e s . The p h y s i o l o g i c a l and
f u n c t i o n a l s i g n i f i c a n c e f o r s u c h d i f f e r e n c e s i n t h e
e x t r a j u n c t i o n a l AChE p oo l b e t w e e n t h e two f i b r e t y p e s h a s y e t t o
b e d e t e r m i n e d . H ow e v e r , a number o f f a c t o r s may be i m p o r t a n t i n
d e t e r m i n i n g su c h d i s t r i b u t i o n . As h a s b e e n o b s e r v e d i n
s t i m u l a t i o n e x p e r i m e n t s , n o n - e n d p l a t e H fo rm s w e r e i n d u c e d i n
f a s t - t w i t c h m u s c l e s upon c o n t i n u o u s 1 o w - f r e q u e n c y s t i m u l a t i o n
( s e e C h a p t e r 4 ) ; t h e s e e x t r a s y n a p t i c H fo rm s t h u s a p p e a r t o be
a s s o c i a t e d w i t h t h e f u n c t i o n a l o u t p u t o f t h e m u s c l e s . T h i s may
a l s o be t r u e o f t h e mammalian s l o w - t w i t c h m u s c l e s w h ic h r e c e i v e
c o n t i n u o u s t r a i n s o f i m p u l s e s ( V r b o v a , Gordon a n d J o n e s , 1 9 7 8 ) .
The f a s t m u s c l e s , on t h e o t h e r h a n d , a r e r e l a t i v e l y i n a c t i v e i n
a n i m a l s i n c a p t i v i t y . Such i n a c t i v i t y may g i v e r i s e t o a l a r g e
e x t r a j u n c t i o n a l po o l o f g l o b u l a r f o r m s . T h e r e i s no o b v i o u s
• e x p l a n a t i o n f o r t h e s p e c i e s d i f f e r e n c e s o b s e r v e d i n t h e
d i s t r i b u t i o n o f AChE fo rm s i n t h e f a s t t w i t c h m u s c l e s . P r e v i o u s
o b s e r v a t i o n s show t h a t h i g h l e v e l s o f L and M f o rm s w e r e
m a i n t a i n e d i n f a s t m u s c l e s o f r a t t h r o u g h o u t d e v e l o p m e n t and
m a t u r a t i o n ( V i g n y e t a K , 1 9 7 6 ) , w h e r e a s t h e H form
p r o g r e s s i v e l y d i m i n i s h e s d u r i n g t h e same t i m e c o u r s e . The L and
M f o r m s , t h e r e f o r e , a p p e a r t o be r e g u l a t e d d i f f e r e n t l y from t h e
- 7 8 -

H20 AChE. S p e c i e s d i f f e r e n c e s i n t h e l e v e l o f L a n d M form s i n
f a s t m u s c l e s may p o s s i b l y be due t o d i f f e r e n c e s i n t h e
b i o s y n t h e s i s o r p o s t t r a n s l a t i o n a l p r o c e s s i n g o f t h e s e fo rm s.
The p r e s e n c e o f e x t r a j u n c t i o n a l H2 c form i n t y p e I f i b r e s
a l s o a p p e a r s t o c o m p le m e n t t h e l o w c o n c e n t r a t i o n o f
e x t r a j u n c t i o n a l ACh r e c e p t o r s a l o n g t h e e n t i r e l e n g t h o f t h e s e
f i b r e s (McA rdle a nd A l b u q u e r q u e , 1 9 7 3 , M i l e d i a n d Z e l e n a , 1 9 6 6 ) .
The p o s s i b l e p h y s i o l o g i c a l s i g n i f i c a n c e o f t h i s form w i l l be
d i s c u s s e d i n C h a p t e r 3 ( s e e S e c t i o n 3 . 4 . 3 ) .
2 . 4 . 9 Endplate AChE in twitch and tonic fibres
The e n d p l a t e - s p e c i f i c n a t u r e o f t h e H2 c form i s c o n f i r m e d
i n b o t h t y p e I and t y p e I I m u s c l e s . In a d d i t i o n , t h e r e i s a
s i g n i f i c a n t H^c co m p o n e n t a t t h e e n d p l a t e s . T h i s e n d p l a t e
c o m p o n e n t a c c o u n t s f o r t h e common o c c u r r e n c e o f an H^c m o i e t y
i n b o t h t y p e I and t y p e I I f i b r e s o f mammalian m u s c l e s , and
s u g g e s t s t h a t i t may be a f u n c t i o n a l m o l e c u l a r form a t t h e
e n d p l a t e , a l t h o u g h t h e p o s s i b i l i t y t h a t i t i s a d e g r a d a t o r y
p r o d u c t o f t h e HgQAChE c a n n o t be e n t i r e l y r u l e d o u t . The
g l o b u l a r fo rm s o c c u r l a r g e l y a s c o n t a m i n a n t s a t t h e e n d p l a t e s o f
t h e s e t w i t c h f i b r e s , b u t become p r e d o m i n a n t i n t o n i c f i b r e
e n d p l a t e s , s u g g e s t i n g a d i f f e r e n t o r g a n i s a t i o n o f e n d p l a t e AChE
i n t h e t w i t c h and t o n i c f i b r e s ( J e d r z e j c z y k e t a l . . 1 9 8 4 ) .
W h e r e a s ^ i s p r o b a b l y l o c a l i s e d a t t h e b a s a l l a m i n a by
a n c h o r i n g v i a i t s c o l l a g e n - t a i l ( A n g l i s t e r a n d S i l m a n , 1 9 7 8 ;
McMahan e t a l . . 1 9 7 8 ) , t h e L a nd M fo rm s a t t h e e n d p l a t e s may be
a s s o c i a t e d w i t h t h e p la sm a membrane ( L y l e s and B a r n a r d , 1 9 8 0 ) a s
o b s e r v e d f o r t h e b r a i n AChE ( R i e g e r and V i g n y , 1 9 7 6 ; R o t u n d o ,
1 9 8 4 ) .
- 7 9 -

2 . 5 CONCLUSION
In c o n c l u s i o n , b o t h p a s t and p r e s e n t e v i d e n c e s u g g e s t s t h a t
t h e m u l t i p l e m o l e c u l a r f o rm s o f AChE a r e s u s c e p t i b l e t o
p r o t e o l y t i c d e g r a d a t i o n jm v i t r o . Such m o l e c u l a r c o n v e r s i o n
o c c u r s r e a d i l y d u r i n g h o m o g e n i s a t i o n and e x t r a c t i o n i n
c o r r e l a t i o n w i t h t h e l e v e l o f p r o t e o l y t i c a c t i v i t y p r e s e n t i n t h e
t i s s u e s . The d i f f e r e n t l e v e l o f p r o t e o l y t i c a c t i v i t y i n
d i f f e r e n t t i s s u e s , a s w e l l a s t h e d u r a t i o n o f s a m p le p r o c e s s i n g
a r e some o f t h e f a c t o r s w h ic h g i v e r i s e t o v a r i a b i l i t y and l a r g e
r a t i o s o f L form: H form o b s e r v e d f o r d i f f e r e n t m u s c l e s . T h a t
s u c h v a r i a b i l i t y i s a r e s u l t o f p r o t e o l y s i s i s s u p p o r t e d by t h e
f a c t t h a t when p r o t e o l y s i s i s a b o l i s h e d by an e f f e c t i v e
c o m b i n a t i o n o f p r o t e a s e i n h i b i t o r s i n t h e e x t r a c t i o n medium, t h e
m o l e c u l a r p r o f i l e s e x h i b i t a s t a b l e a n d , i n m o s t c a s e s , a
p r e d o m i n a n t form i n a d u l t mammalian m u s c l e s . The breakdown
o f t h e AChE fo rm s p r o b a b l y i n v o l v e s a v a r i e t y o f p r o t e a s e s , a s
i n c o m p l e t e i n h i b i t i o n c o u l d n o t p r e v e n t su c h a r t e f a c t s .
F u r t h e r m o r e , t h e e n d o g e n o u s p r o t e a s e a c t i v i t y p r o b a b l y d e c r e a s e s
w i t h l o n g i n c u b a t i o n , and t h u s i t s i n i t i a l , s h o r t - t e r m e f f e c t h a s
l a r g e l y b e e n o v e r l o o k e d i n t h e p a s t .
The d i s t r i b u t i o n o f AChE form s and t h e f i b r e t y p e
c o m p o s i t i o n o f a m u s c l e a r e i n t e r - r e l a t e d . T h e r e a r e c e r t a i n
c o r r e l a t i o n s b e t w e e n t h e f i b r e t y p e c o m p o s i t i o n and t h e AChE
p r o f i l e s w h i c h , i n t h e c a s e o f f a s t - t w i t c h m u s c l e s a l s o a p p e a r t o
b e s p e c i e s - s p e c i f i c . A t t h e c e l l u l a r l e v e l , t h e d i s t r i b u t i o n o f
AChE a l o n g a m u s c l e f i b r e i s d i c t a t e d by t h e c h a r a c t e r i s t i c s o f
t h e f i b r e t y p e w h e t h e r a t t h e s i t e o f t h e n e u r o m u s c u l a r c o n t a c t ,
- 8 0 -

o r i n t h e r e s t o f t h e f i b r e . E n d p l a t e AChE a p p e a r s t o be
r e g u l a t e d by t h e t y p e o f m o t o n e u r o n e w h ic h s y n a p s e s w i t h t h e
m u s c l e , w h i l e t h e e x t r a j u n c t i o n a l AChE i s p o s s i b l y d e t e r m i n e d by
m u s c l e a c t i v i t y ( W a lk e r and W i l s o n , 1 9 7 5 ; Drachman and W i t z k e ,
1 9 7 2 ) . The f u n c t i o n a l s i g n i f i c a n c e o f t h e e x t r a j u n c t i o n a l AChE
a n d i t s r e l a t i o n s h i p t o t h e e n d p l a t e AChE n e e d s t o be c l a r i f i e d .
I t i s a l s o i m p o r t a n t t o e s t a b l i s h t h e m ec h a n is m s w h ic h d e t e r m i n e
t h e d i f f e r e n t s p a t i a l o r g a n i s a t i o n o f t h e e n d p l a t e AChE i n
d i f f e r e n t f i b r e t y p e s and t h e r o l e t h e s e o r g a n i s a t i o n s p l a y i n
m a i n t a i n i n g t h e c h a r a c t e r i s t i c i n n e r v a t i o n d i f f e r e n t f i b r e t y p e s
r e c e i v e (McArdle and A l b u q u e r q u e , 1 9 7 3 ; Gordon a n d Y r b o v a , 1 9 7 5 ;
C l o s e , 1 9 7 2 ) .
♦
- 8 1 -

CHAPTER 3
THE EFFECT OF CHRONIC DENERVATION ON THE ENDPLATE AND NON-ENDPLATE ACHE
IN MAMMALIAN FAST AND SLOW-TWITCH MUSCLES
♦
- 8 2 -

3 . 1 INTRODUCTION
I t h a s b e e n shown i n C h a p t e r 2 t h a t t h e d i s t r i b u t i o n o f t h e
m o l e c u l a r fo rm s o f AChE i s o n e o f t h e many d i s t i n c t i v e f e a t u r e s o f f a s t
and s l o w m u s c l e s . Much e f f o r t s h a v e b een made i n t h e l a s t t h r e e d e c a d e s
t o i d e n t i f y f a c t o r s w h ic h c o n t r o l t h e e x p r e s s i o n o f m u s c l e t y p e
c h a r a c t e r i s t i c s ( f o r r e v i e w s s e e G u th , 1 9 6 8 ; J o l e s z and S r e t e r , 1 9 8 1 ) .
The d i f f e r e n t p h y s i o l o g i c a l and m e t a b o l i c c h a r a c t e r i s t i c s e x h i b i t e d by
t h e v a r i o u s m u s c l e f i b r e t y p e s may be r e l a t e d t o d i f f e r e n t p r o p e r t i e s o f
t h e i r r e s p e c t i v e m o t o n e u r o n e s ( C l o s e , 1 9 7 2 ; P a d y k u l a and G a u t h i e r ,
1 9 7 0 ) . One way o f e l u c i d a t i n g t h e r e g u l a t o r y r o l e o f t h e n e r v e i s t o
s t u d y c h a n g e s w h ic h t a k e p l a c e i n a m u s c l e o r m u s c l e f i b r e when t h e
i n n e r v a t i o n h a s b e e n r e m o v e d . The h i s t o c h e m i c a l and m o r p h o l o g i c a l
c h a n g e s w h ic h o c c u r i n m u s c l e s upon d e n e r v a t i o n h a v e b e e n s t u d i e d i n
d e t a i l ( T o w e r s , 1 9 3 5 ; T e n n y so n e t a l . , 1 9 7 7 ) . I t was f o u n d t h a t t h e
rem ova l o f m o t o n e u r o n e s was t h e m a jo r f a c t o r w h ich i n d u c e d d e g e n e r a t i o n
o f t h e m u s c l e . I t was shown t h a t d e v e r v a t i o n i n d u c e d s e v e r e a t r o p h i c
symptoms p a r t l y due t o an i n c r e a s e i n t h e p r o t e o l y t i c c a p a c i t y o f t h e
m u s c l e and t h e s u r r o u n d i n g t i s s u e s ( M a s k r e y e t a l . , 1 9 7 7 ; F e r n a n d e z and
D u e l l , 1 9 8 0 ) . F u r t h e r , t h e r e was a r a p i d r e d u c t i o n o f m y o f i b r i l l a r
p r o t e i n s and a c o n c o m i t a n t i n c r e a s e i n c y t o p l a s m i c p r o t e i n s i n
d e n e r v a t e d r a b b i t m u s c l e s ( S y r o v y e t a l . , 1 9 7 1 ) .
+ D e n e r v a t i o n i n d u c e s s p e c i e s - s p e c i f i c c h a n g e s i n t h e s y n t h e s i s
and d i s t r i b u t i o n o f AChE. D e n e r v a t e d c h i c k e n ( L i n k h a r t and W i l s o n , 1 9 7 5 )
and r a b b i t ( T e n n y s o n e t a l . , 1 9 7 7 ; Bacou e t a l , , 1 9 8 2 ) m u s c l e s e x h i b i t
i n c r e a s e d AChE a c t i v i t y w h e r e a s r e d u c e d AChE a c t i v i t y was o b s e r v e d i n
d e n e r v a t e d m u s c l e s o f r a t ( S e n n i e t a l . , 1 9 8 1 ) and m i c e (McCaman, 1 9 6 6 ) .
D e n e r v a t e d r a t and r a b b i t m u s c l e s b o t h e x h i b i t a r e d u c t i o n o f AChE
c o n c e n t r a t i o n a t t h e e n d p l a t e s i t e s , a s t h e s i z e and i n t e n s i t y o f AChE
s t a i n i n g d i m i n i s h e s w i t h t h e p r o g r e s s i o n o f a t r o p h y . The s t a i n i n g
- 8 3 -

i n t e n s i t y i n c r e a s e s c o n c o m i t a n t l y i n t h e e x t r a - j u n c t i o n a l r e g i o n o f t h e
d e n e r v a t e d r a b b i t g a s t r o c n e m i u s b u t s u c h s t a i n i n g was a b s e n t i n t h e r a t
m u s c l e u n d er t h e same e x p e r i m e n t a l c o n d i t i o n s ( B r z i n and M a j c e n - T r a c e v ,
1 9 8 3 ; T en n y so n e t a l . , 1 9 7 7 ) . Such h i s t o c h e m i c a l o b s e r v a t i o n a g r e e s w i t h
t h e f i n d i n g s o f t h e q u a n t i t a t i v e a n a l y s i s o f j u n c t i o n a l and
e x t r a j u n c t i o n a l AChE a c t i v i t i e s . Thus t h e i n c r e a s e i n t h e AChE a c t i v i t y
i n t h e e x t r a - j u n c t i o n a l r e g i o n i n r a b b i t s e em s t o be r e s p o n s i b l e f o r t h e
o v e r a l l i n c r e a s e i n t o t a l a c t i v i t y ; l a c k o f su c h d e n e r v a t i o n - i n d u c e d
i n c r e a s e s i n t h e e x t r a - j u n c t i o n a l AChE i n t h e r a t m u s c l e t h e r e f o r e
r e s u l t s i n an o v e r a l l d e c r e a s e i n a c t i v i t y . D e n e r v a t i o n a l s o g i v e s r i s e
t o an i n c r e a s e i n t h e r e l e a s e o f e n d p l a t e AChE ( I n e s t r o s a e t a l . , 1 9 7 7 )
and t h e t u r n o v e r o f AChE (Newman e t a l . , 1 9 8 4 ) . F u r t h e r m o r e , C a r t e r and
B r i m i j o i n ( 1 9 8 1 ) f o u n d t h a t t h e 10S fo rm a c c o u n t e d f o r a b o u t 80% o f
AChE r e l e a s e d from t h e d e n e r v a t e d r a t d i a p h r a g m . In c o n t r a s t , e l e c t r i c a l
s t i m u l a t i o n i n t h e a b s e n c e o f t h e n e r v e r e s t o r e d t h e AChE a c t i v i t y t o
normal ( W a lk e r and W i l s o n , 1 9 7 5 ) .
The m ost n o t i c e a b l e c h a n g e i n t h e m o l e c u l a r d i s t r i b u t i o n o f AChE
upon d e n e r v a t i o n i s t h e d e p l e t i o n o f t h e e n d - p l a t e s p e c i f i c , c o l l a g e n
t a i l e d form ( H a l l , 1 9 7 3 ; S e n n i e t a l . , 1 9 8 1 ; C o l l i n s and Y o u n k i n , 1 9 8 2 ;
S k e t e l j e t a l . , 1 9 7 8 ) . F u r t h e r m o r e , AChE c l u s t e r i n g a t t h e o r i g i n a l
e n d p l a t e s i t e o r e c t o p i c n e u r a l c o n t a c t c a n be m a i n t a i n e d by
♦ r e i n n e r v a t i o n ( S e n n i e t a l . , 1 9 8 1 ; M aldo nad o e t a l . , 1 9 8 4 ; V i g n y e t a l . ,
1 9 7 6 ; Lptoo and S l a t e r , 1 9 8 0 ) . M u s c le a c t i v i t y i s t h o u g h t t o b e o n e o f
t h e m o st i m p o r t a n t f a c t o r s i n d e t e r m i n i n g t h e a p p e a r a n c e and t h e
s t a b i l i t y o f e n d p l a t e AChE (R u b in e t a l . , 1 9 8 0 ; K o e n ig e t a l . , 1 9 8 2 ) .
E v i d e n c e a l s o s u g g e s t s t h e r o l e o f m u s c l e a c t i v i t y i n t h e r e g u l a t i o n o f
e x t r a - j u n c t i o n a l AChE ( L i n k h a r t and W i l s o n , 1 9 7 5 ) . The m a i n t e n a n c e o f
AChE by some n e u r o t r o p h i c f a c t o r h a s a l s o been p r o p o s e d ( K o e l l e and
R u ch , 1 9 8 3 ; L e n t z e t a l . , 1 9 8 1 ) .
- 8 4 -

S y r o v y e t a l . ( 1 9 7 1 ) r e p o r t e d t h a t t h e p a t t e r n o f c h a n g e ,
e s p e c i a l l y t h e r a t e o f d e v e l o p m e n t o f a t r o p h y , t h e c o n t e n t o f m y o s i n and
lo w m o l e c u l a r w e i g h t p r o t e i n s , t h e ATPase a c t i v i t y and t h e c o n t r a c t i l e
p r o p e r t i e s o f d e n e r v a t e d r a b b i t f a s t and s l o w m u s c l e s w e r e d i s t i n c t l y
d i f f e r e n t . The m u s c l e t y p e s r e m a i n e d d i s t i n c t e v e n a f t e r d e n e r v a t i o n
( L e w i s e t a l . , 1 9 7 4 ) . T h e s e m u s c l e t y p e - s p e c i f i c c h a n g e s may r e f l e c t
d i f f e r e n t modes o f r e g u l a t i o n by w h ic h t h e f a s t and s l o w n e r v e s n o r m a l l y
e x e r t on t h e i r m u s c l e s (M a g a z a n ik e t a l . , 1 9 7 8 ; McArdle and A l b u q u e r q u e ,
1 9 7 3 ) . R e c e n t l y , Bacou e t a l . ( 1 9 8 2 ) r e p o r t e d an a p p a r e n t i n c r e a s e i n
t h e 16S (H2C) a c t i v i t y i n t h e d e n e r v a t e d se m i- m e m b r a n o su s p r o p r i u s , a
p u r e s l o w - t w i t c h m u s c l e i n t h e r a b b i t , s u g g e s t i n g t h a t t h i s fo rm i s
n o r m a l l y s u p p r e s s e d by t h e n e r v e . Such o b s e r v a t i o n c o n t r a s t s w i t h t h e
n e u r a l i n d u c t i o n o f ^ a c t i v i t y d u r i n g s y n a p t o g e n e s i s i n e m b r o y o n ic r a t
( K o e n i g and V i g n y , 1 9 7 8 ) and c h i c k m u s c l e s ( T o u t a n t e t a l . , 1 9 8 3 ) . The
e x p e r i m e n t s d e s c r i b e d i n t h i s s e c t i o n aim t o c l a r i f y t h e e f f e c t o f
d e n e r v a t i o n on AChE a c t i v i t y and t h e d i s t r i b u t i o n o f t h e m o l e c u l a r f o rm s
i n f a s t and s l o w m u s c l e s i n a number o f mammalian s p e c i e s , and how t h e
v a r i o u s m o l e c u l a r f o rm s c h a n g e i n r e l a t i o n t o t h e o v e r a l l i n c r e a s e o r
d e c r e a s e i n t o t a l AChE a c t i v i t y i n d i f f e r e n t m u s c l e s . The d i f f e r e n t i a l
e f f e c t s o f d e n e r v a t i o n on e n d p l a t e and n o n - e n d p l a t e AChE f o r m s w e r e a l s o
i n v e s t i g a t e d .
3.2 MATERIALS AND METHODS
3.2.1 Animals
New Z e a l a n d W h it e and Old E n g l i s h B l a c k r a b b i t s (^3 k g ) , D u n k in -
H a r t l e y g u i n e a p i g s ( t w o m onth s o l d ) and G olde n h a m s t e r s ( 3 m o n th s o l d )
w e r e u s e d f o r t h i s s t u d y .
- 8 5 -

3.2.2 Muscles
The f o l l o w i n g p e l v i c l im b m u s c l e s o f s i n g l e f i b r e - t y p e
c o m p o s i t i o n w e r e s t u d i e d : t h e f a s t - t w i t c h EDL ( e x t e n s o r d i g i t o r u m
l o n g u s ) ( A r i a n o e t a l . , 1 9 7 3 ) , t h e s l o w - t w i t c h s o l e u s ( P e t e r e t a ! . ,
1 9 7 2 ; C a m p b e l l , 1 9 7 9 ) and t h e se m i- m e m b r a n o su s w h ic h i s made up o f tw o
d i s t i n c t r e g i o n s — t h e sem im em b ran osu s a c c e s s o r i u s (SA) ( f a s t - t w i t c h ) ,
o r t h e o u t e r r e g i o n , and t h e se m im em br ano su s p r o p r i u s (S P ) ( s l o w -
t w i t c h ) , o r t h e i n n e r r e g i o n . The f i b r e - t y p e c o m p o s i t i o n o f t h e s e
m u s c l e s a r e l i s t e d i n T a b l e 1 . 1 . The p u re f i b r e t y p e c o m p o s t i o n o f t h e s e
m u s c l e s f a c i l i t a t e s s e p a r a t e a n a l y s e s o f t h e e f f e c t o f d e n e r v a t i o n on
t h e t y p e I I ( f a s t - t w i t c h ) and t y p e I ( s l o w - t w i t c h ) f i b r e s . The f a c t t h a t
t h e s e m u s c l e s a r e a l l p a r t o f t h e p e l v i c l i m b m u s c u l a t u r e s i m p l i f i e s t h e
s u r g e r y r e q u i r e d f o r d e n e r v a t i o n .
3.2.3 Surgical procedures
A n i m a l s w e r e o p e r a t e d on u n d e r f l u o t h a n e a n a e s t h e s i a . One o f t h e
h i n d l i m b s w e r e d e n e r v a t e d ; t h e m u s c l e s i n t h e u n d e n e r v a t e d l i m b w e r e
u s e d a s c o n t r a l a t e r a l c o n t r o l s . *
The r a b b i t sem im em br ano su s i s i n n e r v a t e d by t h e n . c u t a n e u s
f e m o r i s c a u d a l i s , R r . m u s c u l a r e s n . i s c h i a d i c i and b r a n c h e s o f t h e N.
o b t u r a t o r i u s ( B o r o n e e t a l . , 1 9 7 3 ) . C o m p l e te d e n e r v a t i o n o f t h e s e
m u s c l e s r e q u i r e d s e c t i o n i n g a l l t h e n e r v e b r a n c h e s d i s t a l l y c l o s e t o t h e
m u s c l e . The g u i n e a p i g sem im em br ano su s i s i n n e r v a t e d by t h e N.
i s c h i a d i e u s , t h e n . c u t a n e u s f e m o r i s c a u d a l i s , t h e n . f e m o r i s and t h e n .
s a p h e n u s ( C o o p e r and S c h i l l e r , 1 9 7 5 ) . The EDL and s o l e u s o f r a b b i t ,
g u i n e a p i g and h a m s t e r w e r e r o u t i n e l y d e n e r v a t e d by s e c t i o n i n g t h e N.
i s c h i a d i e u s i n t h e m i d - t h i g h r e g i o n and r e m o v i n g a p p r o x i m a t e l y 6mm o f
t h e d i s t a l n e r v e t r u n k .
* I w i s h t o t h a n k Mr. D a v i d Green f o r k i n d l y p e r f o r m i n g t h e s u r g i c a l
d e n e r v a t i o n p r o c e d u r e s .
-86-

3.2.4 Partial denervation
To s t u d y t h e e f f e c t o f p a r t i a l d e n e r v a t i o n , i t was d e c i d e d t o
u s e t h e r a b b i t se m im em br ano su s w h ic h h a s a c o m p le x n e u r a l i n p u t . P a r t i a l
d e n e r v a t i o n was a c h i e v e d e i t h e r by s e c t i o n i n g t h e n . c u t a n e u s f e m o r i s
c a u d a l i s a l o n e o r o n l y t h e R r . m u s c u l a r i s n . i s c h i a d i c i .
The g u i n e a p i g s w e r e s a c r i f i c e d 2 t o 4 w e e k s p o s t - d e n e r v a t i o n ;
t h e r a b b i t s and h a m s t e r s w e r e a l w a y s s a c r i f i c e d 4 w e e k s p o s t
d e n e r v a t i o n .
3 . 2 . 5 Muscle extract preparation
A n i m a l s w e r e k i l l e d by an o v e r d o s e o f p e n t o b a r b i t o n e . M u s c l e s
w e r e d i s s e c t e d from t h e d e n e r v a t e d and t h e c o n t r a l a t e r a l l i m b s ,
i m m e d i a t e l y w e i g h e d , m in c e d b r i e f l y and h o m o g e n iz e d i n 10 v o l o f an i c e -
c o l d p h o s p h a t e b u f f e r , pH 7 . 0 , c o n t a i n i n g 1M N a C l , 1% T r i t o n , 5 mM EGTA
and t h e f o l l o w i n g p r o t e a s e i n h i b i t o r s : b a c i t r a c i n ( l m g / m l ) , b e n z e m i d i n e
HC1 (2mM), c h i c k e n o v o m u c o id t r y p s i n i n h i b i t o r ( 0 . 2 m g / m l ) , s o y - b e a n
t r y p s i n i n h i b i t o r ( 0 . 1 m g / m l ) , l e u p e p t i n ( 0 . 0 4 m g / m l ) , p e p s t a t i n ( 0 . 2
m g/m l) and NEM (5mM) ( s e e S e c t i o n 2 . 2 . 3 ) . Smal l m u s c l e s w e r e h o m o g e n i s e d
w h o l e , a f t e r m i n c i n g b r i e f l y , w i t h g l a s s / g l a s s hand h o m o g e n i s e r s . L a r g e
m u s c l e s w e r e m in c e d and a b o u t l g o f t h e m i n c e s w e r e h o m o g e n i s e d w i t h a
P o l y t r o n ( K i n e t i c a ) ( 3 x 3 0 s ) a t 4 ° C . The h o m o g e n a t e s w e r e i m m e d i a t e l y
* c e n t r i f u g e d a t 4 3 , 5 0 0 x g f o r 15 min t o y i e l d t h e m u s c l e c r u d e e x t r a c t s .
3.2.6 Measurement of endplate and non-endplate AChE
E n d p l a t e and n o n - e n d p l a t e r e g i o n s w e r e d i s s e c t e d from m u s c l e s
( w h e r e i n d i c a t e d ) and e x t r a c t e d a s d e s c r i b e d p r e v i o u s l y ( s e e S e c t i o n
2 . 2 . 5 ) . T h e s e s a m p le c r u d e e x t r a c t s w e r e f o l l o w e d i m m e d i a t e l y by
s e d i m e n t a t i o n and enzyme a n a l y s i s a s d e s c r i b e d i n S e c t i o n 2 . 2 . 6 and
2 . 2 . 7 .
- 8 7 -

3 . 3 Results
3.3.1 Denervation-induced changes in AChE activities in different
species
C h r o n i c a l l y d e n e r v a t e d m u s c l e s e x h i b i t s e v e r e a t r o p h y and
e x t e n s i v e f a t t y i n f i l t r a t i o n ( F i g . 3 . 1 ) . T h e r e was s u b s t a n t i a l v a r i a t i o n
i n t h e w e i g h t l o s s o f t h e d e n e r v a t e d m u s c l e s b e t w e e n d i f f e r e n t a n i m a l s
and t r e a t m e n t ; on a v e r a g e , t h e t o t a l l y d e n e r v a t e d SA and SP s u f f e r e d
s i m i l a r w e i g h t l o s s , p a r t i a l l y d e n e r v a t e d SA was t h e m o st a f f e c t e d w h i l e
t h e p a r t i a l l y d e n e r v a t e d SP e x h i b i t e d r e l a t i v e l y m in o r w e i g h t c h a n g e s .
In t h e c o n t r a l a t e r a l i n n e r v a t e d f a s t - t w i t c h m u s c l e s , i . e . t h e
EDL and t h e SA, t h e AChE a c t i v i t y r a n g e d b e t w e e n 0 . 3 and 1 . 0 U / g ( w e t
w e i g h t ) ; i n t h e i n n e r v a t e d s l o w - t w i t c h m u s c l e s , t h e a c t i v i t y was b e t w e e n
0 . 5 and 0 . 7 U / g . T h e r e was no a p p a r e n t c o r r e l a t i o n b e t w e e n t h e l e v e l o f
AChE a c t i v i t y and t h e f i b r e t y p e c o m p o s i t i o n o f t h e s e m u s c l e s ( s e e a l s o
T a b l e 2 . 3 ) , c o n t r a r y t o t h a t o b s e r v e d i n c h i c k e n w h er e t y p e I m u s c l e s
o f t e n d i s p l a y e d much h i g h e r a c t i v i t y t h a n t y p e I I m u s c l e s ( L y l e s e t a l . ,
1 9 8 0 ; L y l e s e t a l . , 1 9 8 2 ) .
The AChE a c t i v i t y i n t h e h a m s t e r m u s c l e s d e c r e a s e d upon
d e n e r v a t i o n ( T a b l e 3 . 1 ) , s i m i l a r t o t h a t o b s e r v e d i n r a t m u s c l e s ( H a l l ,
1 9 7 3 ; S e n n i e t a l . , 1 9 8 1 ) . H ow ever , i n c r e a s e s i n AChE a c t i v i t y w e r e
• o b s e r v e d i n t h e d e n e r v a t e d m u s c l e s o f b o t h r a b b i t and g u i n e a p i g when
comp ared t o t h e i r c o n t r a l a t e r a l c o n t r o l m u s c l e s ( T a b l e 3 . 1 ) ( T e n n y s o n e t
a l . , 1 9 7 7 ) . T h e s e c h a n g e s a r e e x p r e s s e d a s t h e r a t i o o f AChE a c t i v i t i e s
o r ' r e l a t i v e l y a c t i v i t y 1 i n t h e d e n e r v a t e d and c o n t r a l a t e r a l c o n t r o l
m u s c l e s . Such r a t i o s v a r i e d b e t w e e n t h e m u s c l e s t h a t w e r e s t u d i e d , b u t
on t h e w h o l e t h e y w e r e somewhat l o w e r i n t h e f a s t m u s c l e s ; f u r t h e r m o r e ,
t h e d e n e r v a t e d sem im em br an osu s from b o t h r a b b i t and g u i n e a p i g d i s p l a y e d
h i g h e r r e l a t i v e a c t i v i t i e s t h a n o t h e r m u s c l e s o f s i m i l a r f i b r e t y p e
-88-


Fig.3.1 Changes in the structure and myosin ATPase staining of the denervated SA (B) and SP (0) rabbit muscles compared to the contralateral control innervated (A, SA; C, SP) muscles. Staining was after preincubation at pH 4.5. Three types of fibres can be distinguished: type 11A stains weakly, type 11B stains intermediately and type I stain intensely. The SP has only type I fibres and the intense ATPase reaction is retained on denervation. The type II fibres atrophy more marked than that seen in the type I fibres. Bar represents 30ym. Results of J. Pizzey.
- 9 0 -

TABLE 3.1. Relative activities of AChE in denervated rabbit,
guinea pig and hamster muscles.
species muscle
fibre
type
relative
activity n
hamster EDL 0.5 ± 0.0 2
guinea pig EDL 1.3 ± 0.2 2
guinea pig SA type II 2.7 ± 0.4 5
rabbit SA(partial)* 4.5 ± 0.6 2
(total) 3.7 ± 0.8 5
hamster soleus 0.8 1
guinea pig soleus 2.5 ± 0.4 2
guinea pig SP 3.5 ± 0.3 5
rabbit soleus type I 2.5 ± 0.4 3
rabbit SP (partial)* 3.3 ± 0.2 2
(total) 4.4 ± 0.7 5
1 Denervation by sectioning the Rr. muscularis N. ischiadici.

composition. The relative activity of the denervated rabbit SP was
similar to that of the denervated guinea pig.
3.3.2 Effects of denervation on the AChE profiles of fast and slow
muscles
In all the type II muscles used in this study, the L and are
equally prominant (Fig. 3.3), except the hamster EDL where the form
predominates (Fig.3.2). Type I muscles contain a large ^ component and
also significant amounts of M and H^c forms (Fig.3.4), contrary to that
previously reported where the H2 C form accounted for a minor proportion
of total activity (Bacou et al., 1982). This discrepancy is likely to be
due to the use of stored, frozen tissue samples, and an inadequate
protection of the AChE from proteolytic degradation, as discussed in
Chapter 2.
It is clear from Fig 3.2, 3.3 and 3.4 that the proportion of
form in these muscles invariably diminished upon denervation. Such
reduction was more pronounced in the fast muscles (averaged 34%) than
that in the slow muscles (averaged 17%). A concomitant increase in the L
forms was observed in denervated guinea pig and rabbit muscles (Fig.3.3
and 3.4), but not in the hamster muscles (Fig.3.2). The fraction of the
area under each peak corresponding to a molecular form was taken to
• represent the proportion of total AChE activity of that molecular form.
The relative change in terms of activity of the different molecular
forms upon denervation is illustrated in Table 3.3.
These figures show that the L forms were increased by many folds
in all denervated muscles except those of the hamster. The activity of
the H2C form, on the other hand, was only marginally reduced in the
denervated fast muscles (Table 3.2), except in the denervated guinea pig
EDL where activity dropped significantly. Almost all the slow
-92-

20 40 60FRACTIONS
FIG- 3-2 Sucrose gradient profile of contralateral control and
chronically denervated hamster EDL. # , contralateral control;
O , denervated (4-week). All extracts in this and subsequent
figures in this chapter were prepared in the presence of protease
inhibitors. Arrows indicate the position of the catalase
marker.- 9 3 -

FIG. 3.3 Sucrose gradient profiles of AChE in control and denervated type II muscles.
Top row: AChE in control muscles. A, guinea pig EDL; B, guinea pig SA; C, rabbit SA. Bottom row:
AChE in denervated muscles. D, guinea pig EDL; E, guinea pig SA; F, rabbit SA (partially
denervated)*, G, rabbit SA (totally denervated). The amount of samples analysed in each case in
this and Figs. 4, 5 and 6 was arbitrary.

*
-95-

1.5 B
#

t FIG. 3.4 Sucrose gradient profiles of AChE in contralateral
control and denervated type I muscles. Top row: AChE in control
muscles. A, guinea pig soleus; B, guinea pig SP; C, rabbit
soleus; D, rabbit SP. Second row: AChE in denervated muscles.
E, guinea pig soleus; F, guinea pig SP; G, rabbit soleus; H,
rabbit SP (partially denervated); I, rabbit SP (totally
denervated).
- 9 5 a -

muscles exhibited an increase in the H^ activity upon denervation, the
increase being most pronounced in the SP muscles. Therefore, although
the relative proportion of W^c form in these slow muscles was reduced
upon denervation, the total activity contributed by this molecular form
actually became greater in most cases (Table 3.2). Hence increases in
the total AChE activity in these denervated muscles were largely due to
the presence of a great excess of L and to a lesser extent the M forms.
The H~ form also contributed to the overall increase in AChE in 2cdenervated slow muscles.
Partial denervation of the rabbit semimembranosus by sectioning
the n. cutaneus femoris caudal is alone had little effect on the muscle.
The muscle underwent only minor weight loss (^5%); neither the AChE
content nor its molecular distribution was altered (Table 3.3). In
contrast, when partial denervation of the muscle was achieved by
sectioning the branches from the sciatic nerve (Rr. muscularis N.
ischiadici), the effect on the semimembranosus was profound (Table 3.1
and 3.2). The AChE activity increased in both the SA and the SP.
Increased activity of the molecular forms was similar in magnitude to
the totally denervated semimembranosus. The differences in the relative
activities, whether of total AChE or of molecular forms, observed
between partial and total denervation were not significant at a 99%
confident limits. There was also no significant difference between the
changes in the proportion of the molecular forms in partially and
totally denervated semimembranosus, as shown in Fig 3.3 and 3.4. If the
relative activities were corrected for the weight differences between
the denervated and control muscles (Table 3.4), the data shows that the
AChE activity per muscle increased slightly; the L forms increased by
similar amount in both type I and type II muscles, while the form
was clearly reduced in the type II but not the type I muscles.
-96-

TABLE 3.2 Relative activities of L and forms in denervated
rabbit, guinea pig and hamster muscle.
species muscle
fibre
type
relative activity
L H2C
hamster EDL 0.4 0.3
guinea pig EDL 2.2 0.4
guinea pig SA II 6.2 1.1
rabbit SA (partial)* 12.1 0.7
(total) 10.3 0.8
hamster soleus 1.0 0.4
guinea pig soleus 4.0 1.4
guinea pig SP I 6.2 2.0
rabbit soleus 5.8 0.7
rabbit SP (partial)* 5.3 2.3
(total) 7.0 3.0
1 Denervation by sectioning the Rr. muscularis N. ischiadici.
- 9 7 -

TABLE 3.3. Changes in total AChE and its molecular forms in the 2
partially denervated rabbit semimembranosus.
relative activity
species muscle total AChE L H2C n
rabbi t SA 1.1 ± 0.2 1.0 1.0 3
rabbit SP 0.9 ± 0.1 0.9 0.95 3
2The semimembranosus was denervated by sectioning the n.
cutaneus femoris caudal is only.
-98-

TABLE 3 . 4 The r e l a t i v e a c t i v i t i e s ( p e r m u s c l e ) o f AChE and i t s
m o l e c u l a r f o rm s i n t h e d e n e r v a t e d g u i n e a p i g and r a b b i t
se m im e m b r a n o s u s .
s p e c i e s m u s c l e
f i b r e
t y p e
r e l a t i v e a c t i v i t y ( p e r m u s c l e )
t o t a l AChE L n
r a b b i t SA ( p a r t i a l ) 1 . 6 ± 0 . 1 4 . 3 0 . 2 2
( t o t a l ) II 1 . 6 ± 0 . 3 4 . 4 0 . 3 5
g u i n e a p i g SA 1 . 2 ± 0 . 1 2 . 8 0 . 5 5
r a b b i t SP ( p a r t i a l ) 2 . 6 ± 0 . 4 4 . 1 2 . 0 2
( t o t a l ) I 1 . 7 ± 0 . 3 2 . 7 1 . 2 5
g u i n e a p i g SP 1 . 7 ± 0 . 2 3 . 0 1 . 0 5
1 D e n e r v a t i o n by s e c t i o n i n g t h e R r . m u s c u l a r i s N . i s c h i a d i c i
*
-99-

3.3.3 Differential analysis of endplate and non-endplate AChE in
normal and denervated muscles
D i f f e r e n t i a l a n a l y s i s o f t h e j u n c t i o n a l and e x t r a - j u n c t i o n a l
AChE by m i c r o d i s s e c t i o n showed t h a t t h e H^c and f o r m s p r e d o m i n a t e a t
t h e e n d p l a t e s o f b o t h t y p e I and t y p e I I f i b r e s ( F i g 3 . 5 A and 3 . 5 B ) .
C h r o n i c d e n e r v a t i o n up t o 4 w e e k s r e d u c e d b u t d i d n o t e l i m i n a t e t h e s e
f o r m s from t h e e n d p l a t e s ( F i g 3 . 5 C and 3 . 5 D ) . The p e r s i s t e n c e o f t h e s e
f o r m s i s c o n s i s t e n t w i t h t h e w ea k en ed b u t v i s i b l e e n d p l a t e AChE s t a i n i n
b o t h t y p e s o f m u s c l e f i b r e s . T h i s i s i l l u s t r a t e d i n F i g 3 . 7 w h i c h s h o w s
t h e s t a i n e d t e a s e d f i b r e s from d e n e r v a t e d and c o n t r a l a t e r a l c o n t r o l
s o l e u s m u s c l e s o f t h e g u i n e a p i g . F u r t h e r m o r e , L f o r m s w h i c h a r e
n o r m a l l y f o u n d o n l y i n t h e n o n - e n d p l a t e r e g i o n s a p p e a r i n t h e e n d p l a t e s
o f t h e d e n e r v a t e d t y p e I f i b r e s ( F i g 3 . 6 A and 3 . 6 B ) . E x t r a j u n c t i o n a l H
f o r m s a r e p r e s e n t i n t h e s o l e u s o f t h e g u i n e a p i g ( F i g 3 . 6 B ) and t h e SP
o f t h e r a b b i t ( J e d r z e j c z y k J . , p e r s o n a l c o m m u n i c a t i o n ) , b u t a r e
n e g l i g i b l e i n t h e t y p e II f i b r e s ( F i g 3 . 6 A ) . T h e s e e x t r a j u n c t i o n a l H
f o r m s r e m a in s i g n i f i c a n t i n t h e d e n e r v a t e d t y p e I f i b r e s ( F i g 3 . 6 C )
w i t h i n t h e 4 week p o s t - d e n e r v a t i o n p e r i o d .
3.4 DISCUSSION
3.4.1 Changes in AChE activities induced by chronic denervation
D e n e r v a t i o n - i n d u c e d c h a n g e s i n AChE a c t i v i t i e s o f s k e l e t a l
m u s c l e s a r e s p e c i e s s p e c i f i c ( B r z i n and M a jc e n - T r a f c e v , 1 9 6 3 ) . In m u s c l e s
w h i c h e x h i b i t an o v e r a l l r e d u c t i o n i n AChE a c t i v i t y upon c h r o n i c
d e n e r v a t i o n , e . g . h a m s t e r EDL, t h e r e i s an e q u a l r e d u c t i o n i n t h e L and
H f o r m s . In c o n t r a s t , d e n e r v a t e d r a b b i t and g u i n e a p i g m u s c l e s e x h i b i t
a l a r g e i n c r e a s e i n AChE a c t i v i t y , w h ic h i s a t t r i b u t e d t o a marked
e l e v a t i o n o f t h e L and M f o r m s . T h e r e i s a l s o an a p p a r e n t d e c r e a s e i n
t h e p r o p o r t i o n o f H fo r m s i n b o t h f a s t and s l o w m u s c l e s , c o n t r a r y t o
-100-

FIG- 3-5 S u c r o s e g r a d i e n t p r o f i l e s o f AChE i n t h e e n d p l a t e
r e g i o n o f c o n t r o l and d e n e r v a t e d g u i n e a p i g m u s c l e f i b r e s . EDL
( t y p e I I ) : A, c o n t r a l a t e r a l c o n t r o l ; C, d e n e r v a t e d . S o l e u s ( t y p e
I): B, c o n t r a l a t e r a l c o n t r o l ; D, d e n e r v a t e d . D o t t e d l i n e
r e p r e s e n t s b a c k g r o u n d a b s o r b a n c e o f g r a d i e n t f r a c t i o n s i n c u b a t e d
i n t h e p r e s e n c e o f b o t h isoOMPA (1 x 1 0 - 4 M) a n d BW284C51 ( 5 x
10"^M). Where b a c k g r o u n d i s n o t i n d i c a t e d , i t h a s b een
s u b t r a c t e d from t h e s a m p le p r o f i l e .
-101-

FRACTIONS FRACTIONS
FIG. 3-6 S u c r o s e g r a d i e n t p r o f i l e s o f e x t r a j u n c t i o n a l AChE i n
c o n t r o l and d e n e r v a t e d g u i n e a p i g m u s c l e f i b r e s . EDL ( t y p e I I ) :
A, c o n t r a l a t e r a l c o n t r o l ; C, d e n e r v a t e d . S o l e u s ( t y p e I): B,
c o n t r a l a t e r a l c o n t r o l ; D, d e n e r v a t e d .
-102-
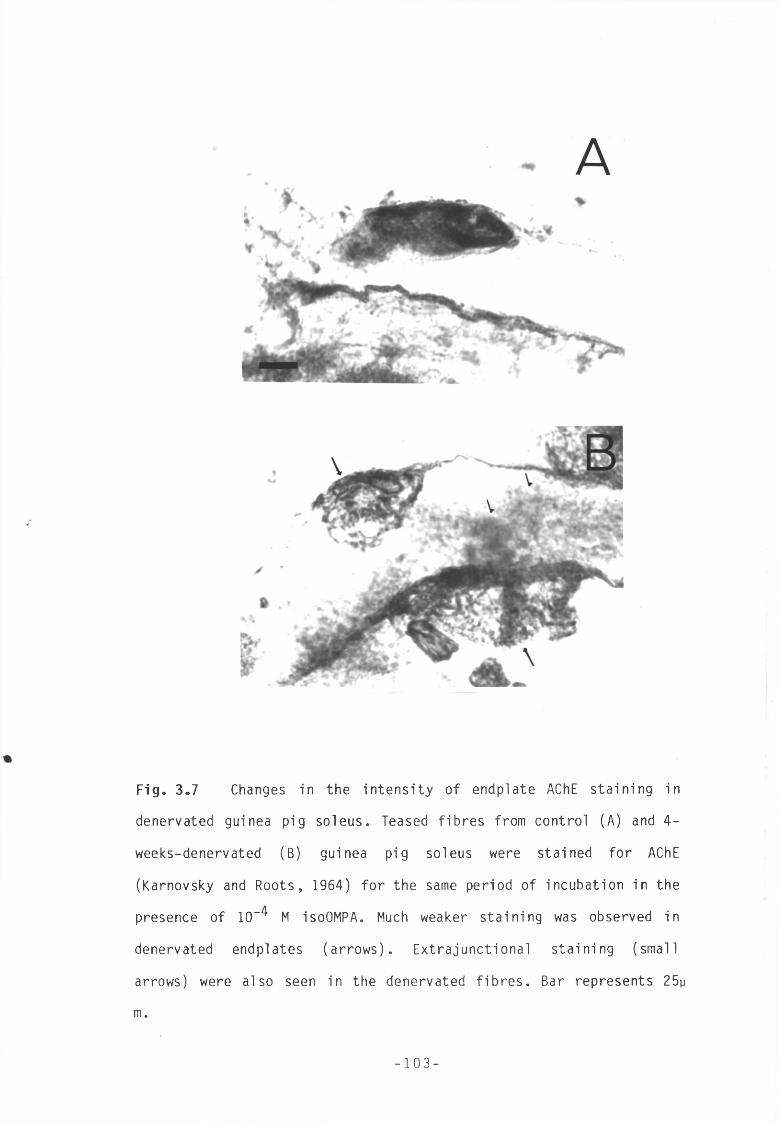
F i g . 3 . 7 Changes in the intensity of endplate AChE staining in
denervated guinea pig soleus. Teased fibres from control (A) and 4-
weeks-denervated (B) guinea pig soleus were stained for AChE
(Karnovsky and Roots, 1964) for the same period of incubation in the
presence of 10"^ M isoOMPA. Much weaker staining was observed in
denervated endplates (arrows). Extrajunctional staining (small
arrows) were also seen in the denervated fibres. Bar represents 25y
m.
- 1 0 3 -

t h a t r e p o r t e d by Ba cou e t a l . ( 1 9 8 2 ) . T h e s e w o r k e r s sh o w ed t h a t w h i l e
t h e p r o p o r t i o n o f ^ f o r m s was r e d u c e d i n t h e d e n e r v a t e d f a s t - t w i t c h SA
( t y p e I I ) , i t was m a r k e d l y i n c r e a s e d i n t h e d e n e r v a t e d s l o w - t w i t c h SP
( t y p e I ) . I t i s c l e a r t h a t t h e d i s c r e p a n c y b e t w e e n t h e p r e s e n t r e s u l t
and t h a t r e p o r t e d p r e v i o u s l y l i e s i n t h e d i f f e r e n t e s t i m a t i o n s o f t h e H
AChE co m p o n e n t i n t h e c o n t r o l S P . A n a l y s i s o f t h e AChE c o m p o s i t i o n i n a
v a r i e t y o f p u r e t y p e I m u s c l e s d e m o n s t r a t e s u n e q u i v o c a l l y t h a t t h e H
AChE i s a m a j o r c o m p o n e n t i n t y p e I m u s c l e s ( s e e C h a p t e r 2 ) , t h u s t h e
a p p a r e n t i n c r e a s e i n t h e p r o p o r t i o n o f t h e H AChE c o m p o n e n t r e p o r t e d by
Bacou e t a l . ( 1 9 8 2 ) i s an a r t e f a c t du e t o t h e u n d e r e s t i m a t i o n o f t h e
amount o f H f o r m s i n t h e c o n t r o l S P . I t may be c o n c l u d e d , t h e r e f o r e ,
t h a t i n b o t h f a s t and s l o w m u s c l e s o f t h e r a b b i t and t h e g u i n e a p i g , t h e
s u b s t a n t i a l i n c r e a s e i n t o t a l AChE a c t i v i t i e s upon d e n e r v a t i o n i s
l a r g e l y due t o an i n c r e a s e i n t h e a c t i v i t y o f t h e L and M form i n t h e
m u s c l e s . The a c c u m u l a t i o n o f L and M f o r m s p r o b a b l y a r i s e s from an
e x c e s s s y n t h e s i s o f t h e s e f o rm s i n t h e d e n e r v a t e d m u s c l e s , s i n c e
d e t a i l e d s t u d i e s by T e n n y s o n e t a l . ( 1 9 7 7 ) showed t h a t l a r g e amount o f
c y t o p l a s m i c AChE a p p e a r e d upon c h r o n i c d e n e r v a t i o n , c l e a r l y
d i s t i n g u i s h a b l e from t h e p a r t i c u l a t e AChE w h ic h a c c o u n t s f o r m o st o f t h e
AChE a c t i v i t y i n normal m u s c l e s . Some r e g e n e r a t i n g f i b r e s , a s w e l l a s
t h e l a r g e number o f AChE c o n t a i n i n g m o n o n u c l e a r c e l l s m u st a l s o
c o n t r i b u t e t o t h e i n c r e a s e i n AChE a c t i v i t y i n t h e a t r o p h i c m u s c l e .
The d e n e r v a t e d t y p e I m u s c l e s o f b o t h r a b b i t and g u i n e a p i g a l s o
e x h i b i t an e l e v a t e d a c t i v i t y o f t h e H2 c f o r m , e v e n t h o u g h s u c h i n c r e a s e
i s m i n o r when comp ared t o t h e i n c r e a s e i n t h e L and M f o r m s ( T a b l e 3 . 2 ) .
T h i s i n c r e a s e d H AChE a c t i v i t y i s p e c u l i a r t o t h e t y p e I m u s c l e s , s i n c e
i n t y p e II m u s c l e s b o t h t h e p r o p o r t i o n and t h e t o t a l a c t i v i t y o f t h e H2 c
a r e r e d u c e d . T h i s i n c r e a s e i n t h e t o t a l H2 c AChE i n t h e t y p e I m u s c l e s
a g r e e s w i t h t h e f i n d i n g s o f Bacou e t a l . ( 1 9 8 2 ) .
-104-

3.4.2 AChE in the endplates of normal and denervated muscle fibres
Bo th t h e anc* a s i g n i f i c a n t amount o f t h e H^c AChE a r e
c o n f i n e d t o t h e e n d p l a t e s o f t y p e I I f i b r e s . T h e s e fo r m s r e m a in
d e t e c t a b l e i n d e n e r v a t e d m u s c l e s 4 w e e k s a f t e r d e n e r v a t i o n and c o n f i n e d
t o t h e e n d p l a t e s i t e . The o v e r a l l r e d u c t i o n i n t h e H f o r m s i n t h e
d e n e r v a t e d t y p e I I f i b r e s t h e r e f o r e r e p r e s e n t s t h e l o s s o f AChE from t h e
e n d p l a t e . A f t e r 4 w e e k s , t h e s e f o rm s a r e r e d u c e d by 40-60%, w h ic h means
t h a t a s i g n i f i c a n t p r o p o r t i o n o f H2 c and Hl c AChE r e m a in a t t h e e n d p l a t e
e v e n when t h e n e r v e i s a b s e n t . T h i s a g r e e s w i t h t h e w e a k e n e d b u t s t i l l
c l e a r l y d i s c e r n a b l e e n d p l a t e AChE s t a i n i n g t y p i c a l o f d e n e r v a t e d m u s c l e s
( B r z i n and M a j c e n - T r a c e v , 1963). AChE s t a i n i n g a l s o sh ow s t h a t t h e
e n d p l a t e s i n t h e d e n e r v a t e d t y p e I f i b r e s a r e now s m a l l e r and o n l y
w e a k l y s t a i n e d ( F i g . 3.7). Hence a l t h o u g h t h e s e f i b r e s a r e c a p a b l e o f
s y n t h e s i z i n g e x c e s s H f o r m s , t h e e n d p l a t e p o p u l a t i o n o f t h e s e form s m us t
h a v e d i m i n i s h e d , a s o b s e r v e d i n t h e d e n e r v a t e d t y p e I I f i b r e s . An
abnormal p o p u l a t i o n o f L AChE i s a l s o a p p a r e n t i n t h e e n d p l a t e s o f
d e n e r v a t e d t y p e I f i b r e s . They may a r i s e from t h e p r o t e o l y t i c breakdown
o f H AChE a t t h e e n d p l a t e , a l t h o u g h i t se em s u n l i k e l y s i n c e v e r y l i t t l e
L f o r m s i s o b s e r v e d i n t h e e n d p l a t e o f d e n e r v a t e d t y p e II f i b r e s .
A l t e r n a t i v e l y , t h e s e fo rm s may r e p r e s e n t n e w l y s y n t h e s i z e d L AChE w h ic h
e i t h e r h a s a s p e c i f i c l o c a l i s a t i o n a t t h e e n d p l a t e s i t e o r a r i s e s from
• t h e g e n e r a l i n c r e a s e i n t h e s y n t h e s i s o f l o w e r m o l e c u l a r w e i g h t f o r m s
a l o n g t h e l e n g t h o f t h e f i b r e s .
The r e d u c t i o n i n e n d p l a t e H AChE s u g g e s t s t h a t t h e c o n c e n t r a t i o n
o f AChE a t t h e n e u r o m u s c u l a r j u n c t i o n i s i n d e p e n d e n t o f t h e s y n t h e t i c
c a p a c i t y o f t h e m u s c l e . I n c r e a s e d s y n t h e s i s o f t h e L, M and i n t h e c a s e
o f t h e d e n e r v a t e d t y p e I f i b r e s , t h e H f o rm s d o e s n o t p r e v e n t t h e
r e d u c t i o n o f AChE a t t h e e n d p l a t e , w h ic h i n v a r i a b l y o c c u r s upon c h r o n i c
d e n e r v a t i o n . I t s u g g e s t s , t h e r e f o r e , t h a t t h e c o n c e n t r a t i o n o f AChE a t
- 105 -

t h e e n d p l a t e may be c o n t r o l l e d by some o t h e r n e u r a l l y r e g u l a t e d f a c t o r s ,
e . g . t h e number o f b i n d i n g s i t e s f o r t h e a t t a c h m e n t o f t h e H f o r m s .
I n d e e d much i n d i r e c t e v i d e n c e ha s s u g g e s t e d t h e a s s o c i a t i o n o f t h e
a s y m m e t r i c fo rm s w i t h t h e b a s a l l a m i n a w h ic h a p p e a r s t o c o n t a i n u n i q u e
e l e m e n t s a t t h e n e u r o m u s c u l a r j u n c t i o n ( S a n e s and H a l l , 1979; S a n e s and
C h e n e y , 1982; S a n e s , 1983). T h e s e e n d p l a t e - s p e c i f i c c h a r a c t e r i s t i c s a r e
a s s o c i a t e d w i t h t h e o r g a n i s a t i o n o f r e c e p t o r s and AChE ( F a l l o n e t a l . ,
1985; W a l l a c e e t a l . , 1985). Cha nge s i n t h e a r c h i t e c t u r e o f t h e b a s a l
l a m i n a upon d e n e r v a t i o n c o u l d t h e r e f o r e be o n e o f t h e c r u c i a l f a c t o r s i n
d e t e r m i n i n g t h e c o n c e n t r a t i o n o f e n d p l a t e AChE. H o w e v e r , t h e f a c t t h a t
t h e H f o r m s re m ai n s p e c i f i c t o t h e e n d p l a t e r e g i o n i n t h e t y p e I I f i b r e s
a l s o s u g g e s t s t h a t e v e n t h o u g h t h e number o f b i n d i n g s i t e s may h a v e b een
r e d u c e d , t h e a t t a c h m e n t o f AChE r e m a i n s s e l e c t i v e . I t h a s b e e n shown
t h a t n e u r a l i n d u c t i o n l e a v e s a p erm a n en t s i g n a l f o r t h e f o c a l
a c c u m u l a t i o n o f t h e H fo r m s (B u rd en e t a l . , 1979). I n c r e a s e d t u r n o v e r o f
t h e en zy m e i n d e n e r v a t e d m u s c l e s (Newman e t a l . , 1984) i s n o t a
s u f f i c i e n t e x p l a n a t i o n f o r t h e r e d u c t i o n o f e n d p l a t e AChE i n c a s e s wh er e
t h e r e a r e c o m p e n s a t o r y i n c r e a s e s i n AChE s y n t h e s i s . H ow ever ,
n e u r o m u s c u l a r i n t e r a c t i o n i s e s s e n t i a l f o r t h e s t a b i l i s a t i o n o f t h e
normal c l u s t e r i n g o f t h e s y n a p t i c AChE. The d e n e r v a t e d t y p e I f i b r e s , on
t h e o t h e r h a n d , c o n t a i n L a s w e l l a s H f o r m s a t t h e e n d p l a t e s i t e s . The
* n a t u r e and l o c a l i s a t i o n o f t h i s L co m p o n en t r e m a i n s t o be c o n f i r m e d ;
h o w e v e r , s i n c e t h e amount o f H fo rm s a t t h e s e e n d p l a t e s i s a l s o r e d u c e d ,
t h e e f f e c t o f d e n e r v a t i o n on t h e e n d p l a t e H fo rm s i s t h e r e f o r e s i m i l a r .
3.4.3 AChE in the non-endplate regions of normal and denervated muscle
fibres
The e x c e s s s y n t h e s i s o f t h e L and M fo rm s i n d e n e r v a t e d t y p e II
m u s c l e i s e n t i r e l y e x t r a s y n a p t i c . The c y t o p l a s m i c AChE s t a i n i n g o b s e r v e d
- 106 -

by T e n n y so n e t a l . ( 1 9 7 7 ) i s l i k e l y t o be c o n t r i b u t e d by t h e s o l u b l e
f r a c t i o n o f t h e s e f o r m s , n o r m a l l y w e l l l o c a l i s e d w i t h i n t h e s a r c o p l a s m i c
r e t i c u l u m . T h e s e l a r g e i n c r e a s e s i n AChE c o u l d a l s o a c c o u n t f o r t h e
i n c r e a s e i n c y t o p l a s m i c p r o t e i n s s e e n i n d e n e r v a t e d r a b b i t m u s c l e s
( S y r o v y e t a l . , 1 9 7 1 ) . In t y p e I m u s c l e s , a p a r t from t h e L and M f o r m s ,
an e x t r a - j u n c t i o n a l p o p u l a t i o n o f F ^ i s n o t e n d p l a t e - s p e c i f i c .
F u r t h e r m o r e , t h i s e x t r a - j u n c t i o n a l H^c AChE m us t b e o v e r - p r o d u c e d
b e c a u s e o f t h e n e t i n c r e a s e i n t h e a c t i v i t y i n d e n e r v a t e d t y p e I
m u s c l e s , d e s p i t e t h e r e d u c t i o n o f t h e e n d p l a t e H AChE c o m p o n e n t . T h i s
s u g g e s t s t h a t t h e e x t r a - j u n c t i o n a l F ^ AChE u n d e r g o e s s i m i l a r c h a n g e s
w h ic h a f f e c t t h e L and M f o rm s o u t s i d e t h e e n d p l a t e , and i s q u i t e
i n d e p e n d e n t o f t h e e n d p l a t e F ^ . In t y p e I I m u s c l e s , d e n e r v a t i o n d o e s
n o t i n d u c e t h e p r o d u c t i o n o f e x t r a - j u n c t i o n a l H d e s p i t e s i m i l a r
e x c e s s i v e s y n t h e s i s o f t h e L and M f o r m s . I t i s c o n c e i v a b l e , t h e r e f o r e ,
t h a t i n c r e a s e d am ounts o f t h e n o n - e n d p l a t e F ^ AChE i s n o t m e r e l y du e t o
an o v e r p r o d u c t i o n o f t h e l o w e r m o l e c u l a r w e i g h t AChE f o r m s . T h i s
a p p a r e n t l a c k o f c o r r e l a t i o n b e t w e e n t h e s y n t h e s i s o f AChE
e x t r a j u n c t i o n a l l y and t h e p r e s e n c e o f j u n c t i o n a l f o rm s i n d i c a t e s t h a t
t h e e x t r a j u n c t i o n a l AChE i s n o t n e c e s s a r i l y t h e p r e c u r s o r s o f t h e
j u n c t i o n a l AChE. D en e jv a t io n h a s b een shown t o i n c r e a s e t h e s e c r e t i o n o f
AChE from m u s c l e t i s s u e ( C a r t e r and B r i m i j o i n , 1 9 8 1 ) , w h ic h r e s e m b l e s
• t h e i n t r a c e l l u l a r s e c r e t o r y po ol o f AChE i n a n e u r a l m u s c l e c u l t u r e
( R o t u n d o and Fam brough, 1 9 8 0 ) .
The p r e s e n c e o f e x t r a j u n c t i o n a l H2C i n t y p e I f i b r e s and i t s
i n c r e a s e d s y n t h e s i s upon d e n e r v a t i o n o f t h e s e f i b r e s may be r e l a t e d t o
some s p e c i f i c l o c a l i s a t i o n o f t h i s form o u t s i d e t h e e n d p l a t e . T h e r e i s
y e t no a v a i l a b l e e v i d e n c e c o n c e r n i n g t h e p o s s i b l e u l t r a - s t r u c t u r a l
l o c a l i s a t i o n o f t h i s e x t r a - j u n c t i o n a l p o p u l a t i o n o f AChE, h o w e v e r ,
t h e p r e s e n c e o f t h i s form n o r m a l l y i n t y p e I f i b r e s and n o t t y p e II
- 107 -

f i b r e s s u g g e s t s t h a t i t may be r e l a t e d t o t h e many p h y s i o l o g i c a l
d i s t i n c t i o n s b e t w e e n t h e two f i b r e t y p e s . One s u c h p e r t i n e n t d i s t i n c t i o n
i s t h e e x t r a j u n c t i o n a l ACh s e n s i t i v i t y due t o t h e p r e s e n c e o f
e x t r a s y n a p t i c ACh r e c e p t o r s i n normal t y p e I f i b r e s . The p r e s e n c e o f
e x t r a - j u n c t i o n a l H AChE may b e r e l a t e d t o t h e f u n c t i o n o f t h i s
p o p u l a t i o n o f r e c e p t o r s . E x t r a j u n c t i o n a l ^ AChE h a s a l s o b e e n o b s e r v e d
i n n e o n a t a l r a t m u s c l e s ( S k e t e l j and B r z i n , 1 9 8 0 ) and human i n t e r c o s t a l
m u s c l e s ( C a r s o n e t a l . , 1 9 7 9 ) . F u r t h e r m o r e , imm ature f i b r e s e x h i b i t t y p e
I c h a r a c t e r s i t i c s ( E c c l e s e t a l . , 1 9 6 0 a ) and c o n t a i n e x t r a j u n c t i o n a l ACh
r e c e p t o r s ( H a r r i s , 1 9 8 1 ) . B a se d on t h e s e o b s e r v a t i o n s , t h e r e f o r e , t y p e I
f i b r e s may r e p r e s e n t a c l a s s o f p a r t i a l l y d i f f e r e n t i a t e d f i b r e s w h ic h
r e t a i n c e r t a i n e m b r y o n ic c h a r a c t e r i s t i c s .
3.4.4 Effect of partial and total denervation on the two regions of
semi membranosus
The e f f e c t o f d e n e r v a t i n g t h e s c i a t i c b r a n c h t o t h e
se m im em b ra n o su s was a s e f f e c t i v e a s when a l l 3 n e r v e s w e r e r e m o v e d ,
s u g g e s t i n g t h a t t h e s c i a t i c t r u n k i s t h e main m o t o n e u r o n e i n p u t t o t h e
tw o r e g i o n s o f t h e m u s c l e . The o b t u r a t o r n e r v e c a r r i e s a m ix e d s e n s o r y
and m o t o r i n p u t w h ic h i s i m p o r t a n t i n t h e r e f l e x m ec h a n ism f o r
c o o r d i n a t e d l o c o m o t i o n . The o b s e r v a t i o n t h a t t h i s n e r v e and t h e c a u d a l
f e m o r a l c u t a n e o u s n e r v e i n p u t do n o t r e d u c e t h e e x t e n t o f d e n e r v a t i o n -
i n d u c e d e l e v a t i o n o f AChE s u g g e s t s t h a t t h e s e n e r v e s p r o b a b l y r e g u l a t e a
r e l a t i v e l y s m a l l number o f m o t o r u n i t s , and may n o t r e i n n e r v a t e t h e
d e n e r v a t e d f i b r e s n o r m a l l y i n n e r v a t e d by t h e s c i a t i c n e r v e .
The AChE c o n t e n t p e r m u s c l e i n d e n e r v a t e d r a b b i t and g u i n e a p i g
m u s c l e s i s f r a c t i o n a l l y h i g h e r t h a n t h a t i n t h e c o n t r o l a n i m a l s d e s p i t e
t h e s e v e r e a t r o p h y o f t h e s e m u s c l e s . T h i s i n d i c a t e s t h a t t h e i n c r e a s e i s
a r e s u l t o f i n c r e a s e d s y n t h e s i s and n o t an a p p a r e n t i n c r e a s e due t o
- 108 -

w e i g h t l o s s . The n o t i c e a b l y h i g h e r AChE a c t i v i t y i n t h e p a r t i a l l y
d e n e r v a t e d SP m u s c l e when compared t o i t s t o t a l l y d e n e r v a t e d c o u n t e r
p a r t i s a r e s u l t o f i t s r e l a t i v e l y m in o r a v e r a g e w e i g h t l o s s . H o w e v e r ,
t h e s e m u s c l e s e x h i b i t e d s i m i l a r d e n e r v a t i o n - i n d u c e d c h a n g e s i n t h e AChE
a c t i v i t i e s a s t h e t o t a l l y d e n e r v a t e d m u s c l e s . Type I f i b r e s a r e known t o
a t r o p h y more s l o w l y t h a n t h e t y p e I I f i b r e s ( N i e d e r l e and Mayr, 1 9 7 8 ) ,
and p e r h a p s some r e s i d u a l a c t i v i t y i n t h e m u s c l e due t o t h e o b t u r a t o r
i n p u t may s l o w down t h e d e g r a d a t i o n o f t h e m y o f i b r i l l a r p r o t e i n s i n t h e
t y p e I m u s c l e . T h i s o b s e r v a t i o n , h o w e v e r , r e q u i r e s c o n f i r m a t i o n s i n c e
t h e number o f s a m p l e s i n v e s t i g a t e d i s s m a l l and e x h i b i t l a r g e
d i f f e r e n c e s . The p a r t i a l l y d e n e r v a t e d SA was s i m i l a r t o t h e t o t a l l y
d e n e r v a t e d SA i n t e r m s o f t h e c h a n g e i n AChE a c t i v i t y a s w e l l a s t h e
d e g r e e o f a t r o p h y , s u g g e s t i n g t h i s r e s i d u a l i n n e r v a t i o n i s i n e f f e c t i v e
i n m a i n t a i n i n g normal r e g u l a t i o n o f t h e t y p e I I f i b r e s . The e f f e c t o f
p a r t i a l d e n e r v a t i o n t h e r e f o r e d i f f e r s l i t t l e from t h a t o f t o t a l
d e n e r v a t i o n on t h e s y n t h e s i s o f AChE i n t y p e I and t y p e I I f i b r e s , b u t
may h a v e d i f f e r e n t i n f l u e n c e s on t h e s t r u c t u r a l e l e m e n t s b e t w e e n t h e two
f i b r e t y p e s . T h e r e a r e c e r t a i n p e c u l i a r i t i e s a s s o c i a t e d w i t h t h e r a b b i t
SP: t h e m u s c l e i s t o t a l l y e n v e l o p e d by t h e SA; h i s t o c h e m i c a l l y , t h e t y p e
I f i b r e s i n t h i s m u s c l e , l i k e t h o s e i n t h e s o l e u s , d i s p l a y much h i g h e r
r e s i s t a n c e t o a l k a l i n e p r e t r e a t m e n t t h a n t h e t y p e I f i b r e s f o u n d i n
• m ix e d m u s c l e s ( J . P i z z e y , p e r s o n a l c o m m u n i c a t i o n ) . T h e r e f o r e t h e s e
f i b r e s i n a p u r e s l o w - t w i t c h m u s c l e may be c o n s i d e r e d t e n t a t i v e l y ,
a c c o r d i n g t o t h e i r h i s t o c h e m i c a l c h a r a c t e r i s t i c s , a s a s u b - t y p e o f
mammalian t y p e I f i b r e s . I t i s n o t w i t h i n t h e s c o p e o f t h i s s t u d y t o
c l a r i f y t h e p r o p e r t i e s o f t h e s e t y p e I f i b r e s . The q u e s t i o n s a s t o
w h e t h e r t h e r e s p o n s e o f t h e SP t o p a r t i a l d e n e r v a t i o n i s p e c u l i a r t o t h e
m u s c l e i t s e l f , o r t y p i c a l o f t h i s s u b - t y p e o f t y p e I f i b r e s , o r w h e t h e r
i t d e m o n s t r a t e s t h e d i f f e r e n c e s b e t w e e n t y p e I and t y p e II f i b r e s a r e
- 109 -

s t i l l t o be s e t t l e d .
3.5 Conclusion
In c o n c l u s i o n , i t was f o u n d t h a t b o t h t h e t y p e I and t y p e I I
f i b r e s e x p r e s s s i m i l a r AChE a c t i v i t i e s and c o n t a i n a s i g n i f i c a n t
p r o p o r t i o n o f H fo r m s w h ic h i s e n d p l a t e s p e c i f i c . In a d d i t i o n , t y p e I
f i b r e s c o n t a i n a s i g n i f i c a n t amount o f e x t r a - j u n c t i o n a l H f o r m s . Bo th
p a r t i a l and t o t a l d e n e r v a t i o n o f m u s c l e s i n g u i n e a p i g and r a b b i t
i n d u c e d an i n c r e a s e i n t o t a l a c t i v i t y , a t t r i b u t e d t o a l a r g e i n c r e a s e i n
t h e s y n t h e s i s o f e x t r a - j u n c t i o n a l AChE f o r m s , i n p a r t i c u l a r t h e L f o r m s .
The i n c r e a s e i n e x t r a - j u n c t i o n a l H2 c form i n t h e t y p e I f i b r e s was
c o n c o m i t a n t w i t h t h e i n c r e a s e i n t h e l o w e r m o l e c u l a r w e i g h t f o r m s , and
a c c o u n t e d f o r t h e i n c r e a s e i n t h e t o t a l H2 c a c t i v t i y . The e n d p l a t e AChE
f o r m s w e r e r e d u c e d upon d e n e r v a t i o n , b u t a s i g n i f i c a n t f r a c t i o n r e m a i n e d
a t t h e e n d p l a t e s i t e . D e n e r v a t e d t y p e I and t y p e I I f i b r e e x h i b i t
s i m i l a r i n c r e a s e i n e x t r a - j u n c t i o n a l AChE and s i m i l a r r e d u c t i o n o f
e n d p l a t e AChE; i n t h e c a s e o f t y p e I f i b r e s , t h e e n d p l a t e and n o n -
e n d p l a t e H2 c AChE w e r e t h u s r e g u l a t e d s e p a r a t e l y . The p r e s e n c e o f no n -
e n d p l a t e H2 c form i n t y p e I m u s c l e s and i t s i n c r e a s e upon d e n e r v a t i o n i s
i n marked d i s t i n c t i o n from t y p e I I m u s c l e s . T h e s e d i f f e r e n c e s may be
r e l a t e d t o t h e e x t r a - j u n c t i o n a l s e n s i t i v i t y o f t y p e I f i b r e s t o ACh.
-110-

CHAPTER 4
CHANGES IN MOLECULAR FORMS OF AChE IN CHRONICALLY STIMULATED
CHICKEN MUSCLE.
- 1 1 1 -

4 . 1 INTRODUCTION
The n e u r o n a l c o n t r o l o f a m u s c l e c o u l d be e x e r t e d by t h e
a c t i o n o f ( a ) some c h e m i c a l m o d u l a t o r ( s ) o r i g i n a t i n g i n t h e m o to r
n e r v e ; ( b ) t h e r e p e t i t i v e d e p o l a r i s a t i o n o f t h e m u s c l e membrane
by t h e c o n d u c t e d a c t i o n p o t e n t i a l s - ( c ) c o m b in e d e f f e c t s o f ( a )
and ( b ) . I t h a s b e e n s u g g e s t e d t h a t t h e s e a c t i o n s may have
d i f f e r e n t i a l e f f e c t s i n d e t e r m i n i n g t h e v a r i o u s p r o p e r t i e s o f t h e
m u s c l e ( W a lk e r and W i l s o n , 1 9 7 5 ) .
M u s c l e a c t i v i t y i s e s s e n t i a l f o r AChE c l u s t e r i n g a t t h e
n e u r o m u s c u l a r c o n t a c t . The b l o c k a d e o f c o n d u c t i o n i n t h e s c i a t i c
n e r v e t r u n k by t h e s u b - p e r i n e u r a l a p p l i c a t i o n o f t e t r o d o t o x i n
( B u t l e r e t a K , 1 9 7 8 ) o r by c o m p r e s s i o n ( C a n g i a n o e t a K , 1 9 8 0 )
i n d u c e d a 50% d e c r e a s e i n r a t h i n d - l i m b m u s c l e AChE and i t s l o s s
from t h e e n d p l a t e . L i k e w i s e , t h e b l o c k a d e o f m yo tub e c o n t r a c t i o n
i n m u s c l e c e l l c u l t u r e by t e t r o d o t o x i n ( W a lk e r and S t r o h m a n ,
1 9 7 6 ; W a lk e r and W i l s o n , 1 9 7 6 ; R i e g e r e t a l . . 1 9 8 0 ) , and i n
i n n e r v a t e d c u l t u r e s by d - t u b o c u r a r i n e ( R u b in e t a l .. 1 9 8 0 ) p r e v e n t
t h e a p p e a r a n c e o f j u n c t i o n a l AChE. C h r o n ic s t i m u l a t i o n ( a t 0 . 5
Hz) t h r o u g h e l e c t r o d e s i n s e r t e d a t t h e s p i n a l c o r d o f t h e c h i c k
embryo i n c r e a s e d t h e number o f n e u r a l c o n t a c t s on t h e PLD m u s c l e
f i b r e w i t h a p l a q u e o f AChE a c t i v i t y p r o d u c e d a t e a c h c o n t a c t
(Renaud e t a ^ . , 1 9 7 8 ) . D i r e c t e l e c t r i c a l s t i m u l a t i o n r e s t o r e s
t h e normal AChE l e v e l i n d e n e r v a t e d m u s c l e i n v i v o ( L i n k h a r t and
W i l s o n , 1 9 7 5 ) o r i n v i t r o ( W a lk e r and W i l s o n , 1 9 7 5 ) , a s w e l l a s
AChE c l u s t e r i n g (F ra nk e t aiK, 1 9 7 5 ; L0mo and S l a t e r , 1 9 8 0 ;
B r e n n e r e t a l . , 1 9 8 3 ) .
-112-

E l e c t r i c a l s t i m u l a t i o n o f a mammalian f a s t - t w i t c h m u s c l e a t
a c o n t i n u o u s l o w f r e q u e n c y (10 Hz) w h ic h i s t y p i c a l o f a s l o w -
t w i t c h f i b r e i n d u c e s t h e m u s c l e t o e x p r e s s s l o w - t w i t c h f i b r e t y p e
c h a r a c t e r i s t i c s ( S a lm o n s and V r b o v a , 1 9 6 9 ; C o t t e r e t a l . , 1 9 7 3 ;
P e t t e e t a \_. , 1 9 7 3 ; Romanul e t a K , 1 9 7 4 ; S a l m o n s and S r e t e r ,
1 9 7 6 ) . T h i s t r a n s f o r m a t i o n i s a p r o g r e s s i v e p r o c e s s w h ic h
i n v o l v e s an i n i t i a l c h a n g e i n t h e e n d o p l a s m i c r e t i c u l u m and
c a l c i u m s e q u e s t r a t i o n (H e i lm a n n e t a K , 1 9 8 1 ; H ei lm an n and P e t t e ,
1 9 7 9 ; Mabuchi e t a K , 1 9 8 2 ) . A r a p i d i n c r e a s e i n t h e e n z y m e s f o r
o x i d a t i v e m e t a b o l i s m o c c u r s , a c c o m p a n i e d by a r e c i p r o c a l d e c r e a s e
i n t h e e n z y m e s f o r g l y c o l y t i c m e t a b o l i s m (K lu g e t a K , 1 9 8 3 ) .
A f t e r p r o l o n g e d s t i m u l a t i o n , t h e l i g h t c h a i n m y o s i n f e a t u r e s o f
f a s t - t w i t c h m u s c l e a r e r e p l a c e d by t h o s e o f s l o w - t w i t c h m u s c l e
( R u b i n s t e i n e t a K , 1 9 7 8 ) r e s u l t i n g i n a c o n t r a c t u r e t y p i c a l o f a
s l o w - t w i t c h m u s c l e ( S a lm o n s and V rb o v a , 1 9 6 9 ) . S t u d i e s u s i n g
e l e c t r o n m i c r o s c o p y r e v e a l a l a r g e i n c r e a s e i n t h e number o f
m i t o c h o n d r i a and c a p i l l a r i s a t i o n i n s t i m u l a t e d m u s c l e s ( S a l m o n s
and V rb ova , 1 9 6 9 ; S a lm o n s e t a K , 1 9 7 8 ) . The Z d i s k s , w h ic h a r e
n o r m a l l y t h i c k e r i n t h e s l o w - t w i t c h t h a n t h e f a s t - t w i t c h f i b r e s
( G a u t h i e r and P a d y k u l a , 1 9 6 6 ) a l s o become n o t i c e a b l y t h i c k e r i n
t h e s t i m u l a t e d f a s t - t w i t c h m u s c l e ( E i s e n b e r g and S a l m o n s , 1 9 8 1 ;
• S a lm o n s e t a K , 1 9 7 8 ) . Such a d a p t i v e c h a n g e s a r e a t t r i b u t e d t o
t h e dyna mic and p l a s t i c n a t u r e o f m u s c l e ( P e t t e , 1 9 8 0 ; Sa lm o n s
and H e n r i k s s o n , 1 9 8 1 ) . The t r a n s f o r m a t i o n o f f i b r e t y p e s i s
r e v e r s i b l e ( S a l m o n s 1 9 8 0 ) and r e c e n t e v i d e n c e s u g g e s t s t h a t t h e
c h a n g e s a r e due t o t h e i n c r e a s e d t o t a l a c t i v i t y r a t h e r t h a n t o
t h e p a r t i c u l a r f i r i n g p a t t e r n i m p o s e d on t h e m u s c l e , s i n c e
s t i m u l a t i n g t h e m u s c l e c h r o n i c a l l y a t 2 0 o r 40 Hz p r o d u c e d
s i m i l a r c h a n g e s a s t h a t a t 5 o r 1 0 Hz ( E e r b e e k e t a l . , 1 9 8 4 ) .
-113-

The v a r i o u s e v i d e n c e p r e s e n t e d i n C h a p t e r s 2 a n d 3 c l e a r l y
i n d i c a t e s a s p e c i f i c r e l a t i o n s h i p b e t w e e n t h e d i s t r i b u t i o n o f t h e
v a r i o u s f o rm s o f AChE and t h e f i b r e t y p e c o m p o s i t i o n o f m u s c l e s .
The n e x t s t a g e o f t h i s s t u d y i s t o i n v e s t i g a t e t h e e f f e c t s o f
c h r o n i c s t i m u l a t i o n on t h e d i s t r i b u t i o n o f t h e s e f o r m s , w h ic h may
s h e d f u r t h e r l i g h t on t h e way AChE i s r e g u l a t e d by t h e
c o n t r a c t i l e a c t i v i t y o f t h e m u s c l e .
A f u r t h e r d i m e n s i o n t o t h e s t u d y o f t h e n e u r a l r e g u l a t i o n o f
m u s c l e AChE i s p r o v i d e d by t h e l a r g e c h a n g e s i n t h e d i s t r i b u t i o n
o f t h i s enzyme i n t h e m u s c l e o f a m u t a n t , g e n e t i c a l l y d y s t r o p h i c ,
c h i c k e n . I n t h e s e h o m o z y g o t e s , a l a r g e e x c e s s o f AChE, i n
p a r t i c u l a r t h e L f o r m s , i s s y n t h e s i z e d by t h e d y s t r o p h i c
f a s t - t w i t c h m u s c l e s s u c h a s t h e PLD ( L y l e s e t a K , 1 9 7 9 ) . The
s y n t h e s i s and d i s t r i b u t i o n o f AChE fo rm s i n t h e a d u l t d y s t r o p h i c
m u s c l e s r e t a i n an e m b r y o n ic p r o f i l e . N e v e r t h e l e s s , n e u r o n a l
c o n t r o l i s e x e r t e d s i n c e d e n e r v a t i o n a g a i n r e s u l t s i n a r e d u c t i o n
o f t h e H f o rm s ( L y l e s e t a l^ . , 1 9 8 2 ) . I n t h i s s t u d y e x p e r i m e n t s
w e r e p e r f o r m e d w h er e b y t h e normal and d y s t r o p h i c PLD m u s c l e o f
t h e c h i c k e n was c h r o n i c a l l y s t i m u l a t e d i n v i v o . The a im i s t o
s e e how t h e m u s c l e AChE i s m o d i f i e d u n d e r t h i s c h a n g e o f
e l e c t r i c a l a c t i v i t y . I t h a s b e e n p r e v i o u s l y shown t h a t t h e
m o rp h o lo g y o f t h e d y s t r o p h i c p a t i g i a l i s m u s c l e i n y o u n g c h i c k e n
a p p e a r e d t o b e n o r m a l i s e d by i n c r e a s e d m u s c l e a c t i v i t y (A sh m o re ,
1 9 8 2 ) . F u r t h e r m o r e , c h r o n i c lo w f r e q u e n c y s t i m u l a t i o n o f
d y s t r o p h i c mouse m u s c l e s a l s o s t r e n g t h e n e d t h e t w i t c h t e n s i o n and
p r e v e n t e d t h e d e g e n e r a t i o n o f t h e m u s c l e s ( L u t h e r t e t _ a T , 1 9 8 0 ;
Vrbova and Ward, 1 9 8 1 ) . I t i s t h e r e f o r e i m p o r t a n t t o s e e w h e t h e r
o r n o t t h e d y s t r o p h i c p a t t e r n o f AChE c o n t e n t i n d y s t r o p h i c
c h i c k e n ca n a l s o be n o r m a l i z e d by i n d u c e d a c t i v i t y .
-114-

In a d d i t i o n , a p r e l i m i n a r y stu<fy o f AChE i n a f e w
f a s t - t w i t c h m u s c l e s i n t h e l o w e r l im b o f b o t h normal and
c | y s t r o p h i c c h i c k e n , and t h e e f f e c t o f c h r o n i c s t i m u l a t i o n on t h e
AChE a c t i v i t y and d i s t r i b u t i o n o f fo rm s i n t h e s e m u s c l e s i s a l s o
p r e s e n t e d . Such s t u d y a i m s t o compare b e t w e e n t h e e f f e c t o f
c h r o n i c s t i m u l a t i o n on f u n c t i o n a l l y more a c t i v e m u s c l e s , su ch a s
t h e l e g m u s c l e s , and t h e i n a c t i v e , p u r e l y t y p e I IB , PLD.
4 . 2 MATERIALS AMD METHODS
4 . 2 . 1 N e r v e s t i m u l a t i o n
S i l v e r e l e c t r o d e s w e r e i m p l a n t e d i n c o n t a c t w i t h t h e PLD
n e r v e , and i m p u l s e s a t 10 Hz w e r e d e l i v e r e d t o them from a
t o t a l l y i m p l a n t e d , e n c a p s u l a t e d p r i n t e d - c i r c u i t s t i m u l a t o r . The
c o n s t r u c t i o n o f t h e s t i m u l a t o r s and t h e s u r g i c a l i m p l a n t a t i o n
p r o c e d u r e s w e r e c a r r i e d o u t by Mr. J . J a r v i s ( B a r n a r d e t a l . ,
1 9 8 5 ) . The c h i c k e n s u s e d w e r e l i n e 4 1 2 ( n o r m a l ) and l i n e 4 1 3
( h o m o z y g o u s f o r m u s c u l a r d y s t r o p h y ; d y / d y ) , b o t h m a i n t a i n e d a t
t h e I m p e r i a l C o l l e g e b r e e d i n g c o l o n y ( W i l s o n e t a K , 1 9 7 9 ) .
The s u c c e s s o f su ch s t i m u l a t i o n r e l i e s upon u n i n t e r r u p t e d
i m p u l s e t r a n s m i s s i o n t o t h e m u s c l e v i a t h e c o n t a c t b e t w e e n t h e
* e l e c t r o d e s and PLD n e r v e . O c c a s i o n a l l y i n t e r r u p t i o n s have a r i s e n
b e c a u s e o f e x c e s s i v e f i b r o u s i n f i l t r a t i o n b e t w e e n t h e e l e c t r o d e s
and t h e n e r v e , w h i c h was d e t e c t a b l e a t a u t o p s y . T h u s , a l t h o u g h
h i s t o c h e m i c a l l y some m u s c l e s m i g h t show a m in o r s h i f t from t y p e
IIB t o IIA f i b r e s , v e r y l i t t l e c h a n g e i n f a c t o c c u r r e d i n t h e
AChE a c t i v i t y . T h e s e c a s e s w e r e d i s c a r d e d , i . e . , o n l y t h o s e
s t i m u l a t e d l i m b s w h ic h showed normal w h o l e - m u s c l e t w i t c h
- 115 -

r e s p o n s e s a t t h e t i m e o f t e r m i n a t i o n ( 2 - 8 w e e k s a f t e r
i m p l a n t a t i o n ) w e r e u s e d . I m p l a n t a t i o n o f e l e c t r o d e s t o t h e PLD
f o r t h e same d u r a t i o n , b u t w i t h o u t a c t u a l s t i m u l a t i o n , was a l s o
i n c l u d e d a s c o n t r o l .
C h r o n i c s t i m u l a t i o n ( 1 0 Hz) o f t h e l o w e r l im b m u s c l e s was
a c h i e v e d by c o n n e c t i n g t h e e l e c t r o d e s t o t h e l o w e r s e c t i o n o f t h e
s c i a t i c n e r v e , a b o u t 2cm a b o v e t h e p a t e l l a . By l e a v i n g t h e u p p e r
l im b m u s c l e s u n i n t e r r u p t e d , su c h s t i m u l a t i o n d i d n o t c a u s e a n y
n o t i c e a b l e d i s c o m f o r t o r r e s t r i c t i v e movement t o t h e c h i c k e n
d u r i n g t h e e x p e r i m e n t a l p e r i o d .
6 - w e e k - o l d normal c h i c k e n s w e r e s t i m u l a t e d f o r 1 and 3 w e e k s
and a 7 - w e e k - o l d d y s t r o p h i c c h i c k e n was s t i m u l a t e d f o r 3 w e e k s
b e f o r e s a c r i f i c e . The EDL, a n t e r i o r t i b i a l i s (AT) and l a t e r a l
g a s t r o c n e m i u s w e r e s e l e c t e d f o r AChE a n a l y s i s .
4 . 2 . 2 E x t r a c t i o n o f AChE
The s t i m u l a t e d PLD m u s c l e and t h e c o n t r a l a t e r a l u n s t i m u l a t e d
m u s c l e w ere e x c i s e d i m m e d i a t e l y a f t e r d e a t h and c o o l e d on i c e . A
s l i c e from t h e c e n t r e o f e a c h m u s c l e was h o m o g e n is e d ( g l a s s /
g l a s s ) i n 10 v o l s o f i c e - c o l d p h o s p h a t e b u f f e r c o n t a i n i n g 1%
T r i t o n X - 1 0 0 , l M - N a C l , 5 mM EGTA and t h e f o l l o w i n g p r o t e a s e
i n h i b i t o r s : b a c i t r a c i n ( l m g / m l ) , c h i c k e n o v o m u c o id t r y p s i n
i n h i b i t o r ( 0 . 2 m g / m l ) , l e u p e p t i n ( 0 . 0 4 m g / m l ) , p e p s t a t i n
( 0 . 0 2 m g / m l ) , and NEM (5nM ). The h o m o g e n a t e s w ere i m m e d i a t e l y
c e n t r i f u g e d a t 4 3 , 5 0 0 x g f o r 15 min t o y i e l d t h e m u s c l e c r u d e
e x t r a c t s . I n some c a s e s , a s a m p le from t h e same m u s c l e s l i c e was
( w h e r e s t a t e d ) m i c r o d i s s e c t e d on an i c e - c o l d p l a t f o r m a s
d e s c r i b e d i n S e c t i o n 2 . 2 . 5 .
- 1 1 6 -

4 . 2 . 3 S e d im e n t a t io n a n a l y s e s
A l i q u o t s o f t h e c r u d e e x t r a c t , o r t h e e x t r a c t o f t h e m i c r o -
d i s s e c t e d s p e c i m e n s w e r e a n a l y s e d on a l i n e a r s u c r o s e d e n s i t y
(5%-20%) g r a d i e n t c o n t a i n i n g b u f f e r e d 1% T r i t o n X - 1 0 0 / lM - N a C l
( s e e s e c t i o n 2 . 2 . 6 ) . The g r a d i e n t f r a c t i o n s w e r e a n a l y s e d f o r
AChE a c t i v i t y a s d e s c r i b e d i n s e c t i o n 2 . 2 . 7 . The a c t i v i t y o f
V'ChE i n t h e m u s c l e c r u d e e x t r a c t s was a l s o m e a s u r e d by t h e same
m e t h o d s , u s i n g 5 X 1 0 “ ^1 BW284C51 t o i n h i b i t AChE. The
r e l a t i v e a c t i v i t y was d e f i n e d a s t h e r a t i o o f t h e AChE a c t i v i t y
o f t h e s t i m u l a t e d m u s c l e and t h e AChE a c t i v i t y o f i t s
c o n t r a l a t e r a l c o n t r o l .
4 . 3 . RESULTS
4 . 3 . 1 . C h a n g es i n AChE a c t i v i t y i n s t i m u l a t e d norm al c h ic k e n PLD
m u s c le
C h r o n i c lo w f r e q u e n c y s t i m u l a t i o n o f t h e normal c h i c k e n PLD
m u s c l e i n d u c e d a r e m a r k a b l e i n c r e a s e i n i t s AChE a c t i v i t y when
com p are d w i t h t h e c o n t r a l a t e r a l , u n s t i m u l a t e d c o n t r o l m u s c l e
( T a b l e 4 . 1 ) . A s i m i l a r i n c r e a s e was o b s e r v e d i n ^ChE a c t i v i t y ,
t o a b o u t 2 - 3 t i m e s t h e u n s t i m u l a t e d m u s c l e . The c o n t r a - l a t e r a l
c o n t r o l PLD u s e d i n t h i s s t u d y c o n t a i n e d i d e n t i c a l AChE a c t i v i t y
and m o l e c u l a r c o m p o s i t i o n a s t h e PLD i n u n t r e a t e d c h i c k e n s .
I m p l a n t e d durnny e l e c t r o d e s had no e f f e c t on t h e h i s t o l o g y , t h e
t o t a l AChE a c t i v i t y o r t h e d i s t r i b u t i o n o f AChE fo rm s o f t h e
m u s c l e .
PLD i n c h i c k e n s i m p l a n t e d w i t h s t i m u l a t o r s b e t w e e n 4 - 6 w e e k s
o f a g e The g r e a t e s t c h a n g e o c c u r r e d a f t e r 2 w e e k s , when
- 117 -

T a b l e 4 . 1 I n c r e a s e s i n AChE c o n c e n t r a t i o n i n t h e normal PLD
a f t e r d i f f e r e n t p e r i o d s o f s t i m u l a t i o n .
Sa m ple
F i n a l
a g e ( w k s )
S t i m u l a t i o n
( w k s )
C o n tr o l [ A C h E ]
( U / g )
R e l a t i v e
a c t i v i t y
A 6 2 0 . 2 5 5 . 5
B 8 3 0 . 3 3 . 2
C 9 5 0 . 4 3 . 1
D 12 3 0 . 2 2 . 8
E 19 8 0 . 3 2 . 6
In T a b l e s 4 . 1 , 4 . 2 a nd 4 . 3 , t h e c o n t r o l o f AChE c o n c e n t r a t i o n i s
shown f o r t h e c o n t r a l a t e r a l m u s c l e i n t h e s t i m u l a t e d b i r d , and
( l a s t c o lu m n ) t h e AChE c o n c e n t r a t i o n i n t h e s t i m u l a t e d m u s c l e
r e l a t i v e t o t h a t i n t h e c o n t r a l a t e r a l m u s c l e i s g i v e n a s t h e
" r e l a t i v e a c t i v i t y " .
*
- 118 -

s t i m u l a t e d PLD m u s c l e e x h i b i t e d a p p r o x i m a t e l y 5 t i m e s t h e normal
AChE a c t i v i t y . T h i s i n c r e a s e i n t o t a l a c t i v i t y was t h e r e s u l t o f
a l a r g e i n c r e a s e (27 f o l d ) i n t h e a c t i v i t y o f t h e L2 f o r m ,
a c c o m p a n i e d by p a r a l l e l b u t s m a l l e r i n c r e a s e s i n t h e o t h e r
m o l e c u l a r fo rm s (M, 7 f o l d ; H, 2 f o l d ) ( c f . F i g 4 . 1A and 4 . I B ) .
F o r s t i m u l a t i o n p e r i o d o f 3 - 5 w e e k s , t h e i n c r e a s e i n t o t a l AChE
a c t i v i t y was r e d u c e d t o o n l y 3 f o l d . T h i s was a c c o m p a n i e d by a
p r o g r e s s i v e r e d u c t i o n o f t h e l_2 a c t i v i t y w h ich was 16 t i m e s
t h e c o n t r o l v a l u e a f t e r t h r e e w e e k s ( F i g . 4 . 1 C ) , and n i n e t i m e s
h i g h e r a f t e r f i v e w e e k s ( F i g . 4 . I D ) . The r e l a t i v e a c t i v i t y o f
t h e M and H fo rm s r e m a i n e d f a i r l y c o n s t a n t o v e r t h e 2 - 5 week
p e r i o d o f s t i m u l a t i o n , a t t h e end o f w h ic h t h e y d i s p l a y e d
r e l a t i v e a c t i v i t i e s o f 6 and 2 . 5 , r e s p e c t i v e l y . A f t e r f i v e w e e k s
s t i m u l a t i o n , t h e r e was a n o t i c e a b l e i n c r e a s e i n t h e H^c form
( F i g . 4 . I D ) , a form w h ic h was a l m o s t a b s e n t i n u n t r e a t e d b i r d s .
PLD i n c h i c k e n s i m p l a n t e d a t 9 - 1 1 w e e k s o f a g e The AChE
i n t h e s e s t i m u l a t e d m u s c l e s was a b o u t 3 t i m e s t h e normal AChE
a c t i v i t y a t , o r a f t e r 3 w e e k s s t i m u l a t i o n ( T a b l e 4 . 1 ) . I t was
c l e a r from t h e m o l e c u l a r p r o f i l e s o b t a i n e d from t h e s e m u s c l e s
( F i g 4 . I F , 4 . 1 G ) t h a t t h e H form s w e r e a l w a y s p r e d o m i n a n t , a s i n
t h e c o n t r o l m u s c l e ( F i g . 4 . I E ) , b u t t h e H^c form a g a i n
* i n c r e a s e d g r e a t l y . T h e s e o l d e r b i r d s , h o w e v e r , showed a more
r a p i d p r o g r e s s t o t h e f i n a l p a t t e r n , w h ic h was n o t c o m p l e t e a f t e r
5 w e e k s s t i m u l a t i o n i n t h e 4 - w e e k b i r d . The f i n a l i n c r e a s e s
i n a c t i v i t y f o r e a c h o f t h e m o l e c u l a r form s w e r e q u i t e s i m i l a r
( L , 3 . 7 ; M, 2 . 0 ; H, 2 . 7 t i m e s t h e i r c o n t r o l v a l u e s ) .
E l e c t r o n m i c r o s c o p y r e v e a l e d a r e m a r k a b l e i n c r e a s e i n t h e
number o f m i t o c h o n d r i a and l i p i d g l o b u l e s i n t h e PLD w h ic h had
- 119 -

*
-120-

FIG- 4.1 Sucrose gradient profiles of AChE in normal PLD after
different periods of stimulation. In 6-9 weeks old chickens: A,
contralateral unstimulated control; B-D, after 2, 3 and 5 weeks
stimulation, respectively. In 12-19 weeks old chickens: E,
contralateral unstimulated control; F-G, after 3 and 8 weeks
stimulation, respectively. E and F were from muscles used for
endplate and non-endplate analysis (Fig. 4.3). The amount of
enzyme analysed in each case in this and other figures in this
chapter was arbitrary. Arrow indicates the position of the
catalase (11.4S) marker.

FRACTIONS
-121-

been stimulated for 8 weeks (Fig. 4.2). These ultrastructural
changes correlate with the progressive change of the fibre type
composition from a dominant type I IB (>80%) to a dominant type
IIA (>70%) (Barnard et al., 1985).
4.3.2 S e p a r a t io n o f e n d p l a t e a n d n o n - e n d p la t e fo r m s o f ACME
The non-endplate region of the stimulated fibres in normal
chickens (Fig. 4.3C) contains significant amounts of
extra-junctional H^c and forms which were negligible in
the control samples. In contrast, in the endplates of the
stimulated fibres, there was little change from the normal
profile; the H^c form did, however, become significant,
although still very minor (Fig.4.3A, B).
4.3.3 T h e r e v e r s a l o f t h e e f f e c t o f s t i m u l a t i o n
Mature chickens whose PLD muscles had been stimulated for
4-5 weeks were examined at intervals after the stimulation, had
ceased (Table4.2). After a 1-week rest, the elevation of AChE
content was halved to a value near or (after a two-week rest)
even below that for control muscle. The profile of the
molecular forms in such a muscle in the latter state was
restored to that seen in a control muscle.
4.3.4. C h a n g e s i n AChE a c t i v i t y i n s t i m u l a t e d d y s t r o p h ic c h ic k e n
PLD m u s c le
Stimulators were implanted into dystrophic chickens of 4-6
weeks or 9-10 weeks of age. Chronic stimulation of the PLD nerve
led to a clear reduction in the AChE activity of these muscle by
-122-

- 123 -

FIG- 4.2 Electron micrographs of PLD muscle from a 19 week old
normal chicken. (A) contralateral unstimulated PLD; CB) 8-week-
stimulated PLD. 1, lipid globules; m, mitochondria. Magnifica
tion: 14,600x. Results of J. Jarvis and J. Cavanagh.

W * -- !% x *
' W _i ’ Jk !B ”*« ££ •?»% ..**;.-a--Si
|g&|M v. $»- f l
t xi ,=3b
| | 1 1- -
„., ir >/•-
♦% * -
IH| *#• -*• • - ■
:.v. 1 % *;."- *"''■ w
•(*V •_ ’’ "
■“•4 -
A : . I ; ! :; -':iiPjfe %!ff V ^ w r ^ n...i-r "&• ?KJL ^
i
* T \ : • 4 !
1 i ^2JT " -C ..1 i. r
124
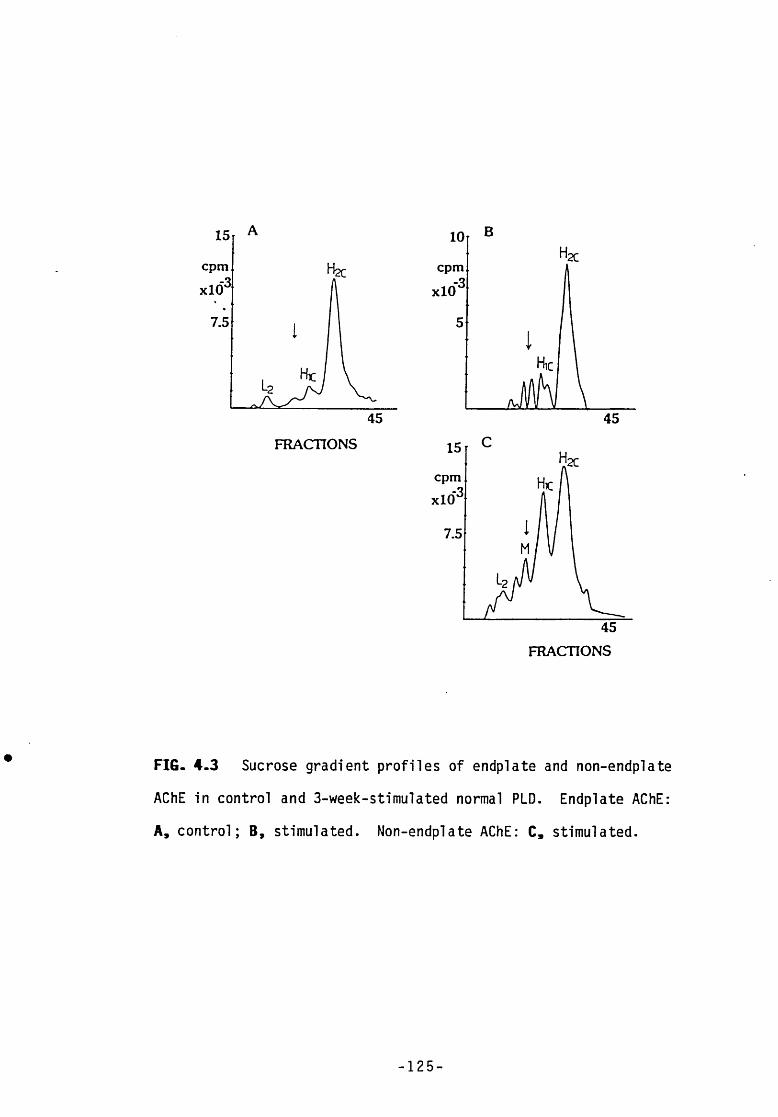
FIG. 4.3 Sucrose gradient profiles of endplate and non-endplate
AChE in control and 3-week-stimulated normal PLD. Endplate AChE:
A, control; B, stimulated. Non-endplate AChE: C, stimulated.
- 125 -

-127-
Table 4.2 Reversal of stimulation-induced changes in AChE after cessation of
the stimulus.
Sample Age(wks)Stimulation
(wks)Control
[AChE] (U/g)Interruption (days)
Relative al1 forms
activity E IT H
F 15 4 0.6 oa 3.916 0.7 7 2.0
G 15 4 0.35 oa 3.017 0.5 14 0.5
H 14 5 0.3 7-14b 0.5 0.5 0.5 0.5
Stimulation was given continuously as shown and was then (where indicated)
interrputed for the period shown (with the electrodes remaining implanted), with
total loss of the induced twitching of the muscle. At the end of that period the PLD
was removed for the measurements. Note that within 2 weeks of cessation of impulses,
the AChE levels in the stimulated muscle regress to within the normal (see Table 4.1)
range, and that all the forms are equally changed.
a. A biopsy sample was taken for the first analysis, for both the PLDs, at the
time of cessation.
b. In this case, the cessation of stimulation and twitching was adventitious
due to fibrous growth over the electrodes contact surface , so that the period of
cessation was not precisely known.

Table 4.3 Decreases in AChE concentration in dystrophic PLD
after different periods of stimulation.
Sample
Final
age(wks)
Stimulation
(wks)
Control [AChE]
(U/g)
Relative
activi ty
A 6 2 2.8 0.3
B 8 3 3.4 0.6
C 9 4 3.2 0.6
D 14 5 2.2 0.4
E 17 8 0.7 1.0
Each value (except A and E) is the mean for 2 or 3 birds.
♦
- 128 -

40% to 70% (Table 4.3). The AChE activity of some of the
stimulated dystrophic muscles however was still about twice that
of the stimulated normal muscles. 0ChE activity, which was
greatly elevated in the dystrophic birds (Lyles et_a]_., 1979),
also declined with stimulation but remained on average 1.5 times
higher than that in the stimulated normal PLD (not shown).
All the molecular forms present in the dystrophic muscle
were reduced upon stimulation, but the reduction of the L forms
was especially noticeable (Fig. 4.4). This preferential
reduction of the L forms was much greater in the more mature
chickens (Fig. 4.4F). After two weeks chronic stimulation, the
dystrophic PLD showed an AChE molecular form profile similar to
that in the stimulated normal muscle (cf. Fig. 4.4B and 4.IB).
This change in activity involved a 70% reduction in the total
amount of the L and M forms,and a 80% reduction in the
form. Thereafter, there was no futher significant change in the
total AChE activity (Table 4. 3) although the relative amount of
the different forms did change, so that eventually both the
activity and distribution of AChE forms resemble that of a
stimulated normal PLD (Fig. 4.4F). This state was reached
earlier in the 14-week birds than in the younger ones. The
profile of a 17 -week old chicken which had been stimulated for 8
weeks was identical to that of the stimulated 14-week birds- The
reduction in the contribution of the L forms averaged 80%. The M
and H forms in contrast were much less affected (0 and 20%
respectively).
The stimulated dystrophic PLD underwent a remarkable
improvement in the structure of the muscle fibres (Fig. 4.5),
- 129 -

F IG - 4 . 4 S u c r o s e g r a d i e n t p r o f i l e s o f AChE i n d y s t r o p h i c PLD
a f t e r d i f f e r e n t p e r i o d s o f s t i m u l a t i o n . In 6 - 9 week o l d
c h i c k e n s : B , 2 w e e k s ; D , 3 - 4 w e e k s . In 14 week o l d c h i c k e n s : F ,
5 w e e k s , A,C a n d E w e r e c o n t r o l p r o f i l e s c o r r e s p o n d i n g t o B , D
a n d F r e s p e c t i v e l y . E an d F w ere from m u s c l e s u s e d f o r e n d p l a t e
and n o n - e n d p l a t e a n a l y s i s ( F i g . 4 . 6 ) .
- 130 -

F IG . 4 . 5 Frozen cross section of PLD muscles from a 14 week old
dystrophic chicken. (A) contralateral unstimulated PLD; (B)
5-week-stimulated PLD. Sections were stained with haematoxylin
and Sirius red. Magnification: 350x. bar represents 20 pm.
Results of J . Jarvi s.
- 1 3 1 -

w h ich w ere now c l o s e l y p ack ed and p o ly g o n a l i n sh a p e in
c r o s s - s e c t i o n . T h ere was a g e n e r a l r e d u c t i o n in f i b r e
h y p e r t r o p h y and t h e n u c l e i became l a r g e l y p e r i p h e r a l . The number
o f t y p e IIA f i b r e s , a s in s t i m u l a t e d normal PLD, i n c r e a s e d
s u b s t a n t i a l l y (B a r n a r d e t a t , 1 9 8 5 ) .
4 . 3 . 5 . S e p a r a t io n o f e n d p l a t e a n d n o n - e n d p la t e fo r m s o f ACME
F i g . 4 .6 A sh o w s t h e m o l e c u l a r form s p r e s e n t a t t h e e n d p l a t e s
o f a d y s t r o p h i c PLD. T h ere was c l e a r l y an abnorm al p r e s e n c e o f
t h e form a t t h e e n d p l a t e , a s fo u n d p r e v i o u s l y i n d y s t r o p h i c
c h i c k e n s o f a d i f f e r e n t a g e ( J e d r z e j c z y k e t a l . , 1 9 8 1 ) . Upon
s t i m u l a t i o n t h i s c o n t r i b u t i o n o f t h e form w as l a r g e l y
e l i m i n a t e d ( F i g . 4 . 6 B ) . O u t s i d e t h e e n d p l a t e s ( F i g . 4 . 6 C ) , a
l a r g e am ount o f and ^ AChE w ere fo u n d in t h e d y s t r o p h i c
PLD f i b r e s , w h ic h w ere a b s e n t in t h e h e a l t h y PLD m u s c l e . The
s t i m u l a t e d d y s t r o p h i c PLD m u s c le f i b r e s c o n t a i n e d , i n c o n t r a s t ,
v e r y l i t t l e L fo rm s e x t r a j u n c t i o n a l l y ( F i g . 4 . 6 D ) . T h i s r e g i o n
w as i n s t e a d d o m in a te d by t h e Hjc and f o r m s . H ence t h e
c e l l u l a r d i s t r i b u t i o n o f t h e s e m o l e c u l a r form s in a s t i m u l a t e d
d y s t r o p h i c PLD f i b r e was v e r y s i m i l a r t o t h a t in a s t i m u l a t e d
norm al PLD f i b r e ( c f . F i g . 4 .6 B ,D and 4 . 3 B , C ) .
4 . 3 . 6 C h a n g es i n AChE a c t i v i t y i n s t i m u l a t e d c h ic k e n l e g m u s c le s
The p r e d o m in a n t ly f a s t - t w i t c h m u s c l e s su ch a s t h e EDL and
t h e AT e x h i b i t e d s i m i l a r AChE a c t i v i t y a s t h e PLD m u s c le ( T a b l e
4 . 5 ) . C h r o n ic s t i m u l a t i o n i n c r e a s e d t h e t o t a l AChE a c t i v i t y i n
t h e s e m u s c l e s , a l t h o u g h t h e m a g n itu d e o f such i n c r e a s e wasa
v a r i a b l e . Thus t h e AChE a c t i v i t y i n t h e s t im u la te d AT and EDL was
- 132 -

F IG . 4 . 6 S u c r o s e g r a d i e n t p r o f i l e s o f e n d p l a t e and n o n - e n d p l a t e
AChE in c o n t r o l and 5 - w e e k - s t i m u l a t e d d y s t r o p h i c PLD. E n d p la te*
AChE: A , c o n t r o l ; B , s t i m u l a t e d . N o n - e n d p la t e AChE: C , c o n t r o l ;
D , s t i m u l a t e d .
- 1 3 3 -

a b o u t t w i c e t h e c o n t r a l a t e r a l c o n t r o l m u s c l e s a f t e r 1 and 3 w e e k s
o f s t i m u l a t i o n r e s p e c t i v e l y , w h i l e t h a t in t h e g a s t r o c n e m i u s was
a n e a r 5 f o l d i n c r e a s e . T h e s e c h a n g e s w e r e a c c o m p a n ie d by an
i n c r e a s e i n t h e t y p e IIA and t y p e I f i b r e s w h ic h w e r e
p a r t i c u l a r l y d i s t i n c t i n t h e g a s t r o c n e m i u s ( T a b l e 4 . 4 ) .
T h e s e f a s t - t w i t c h m u s c l e s c o n t a i n e d p r e d o m in a n t ly
AChE. T h er e w as l i t t l e d i f f e r e n c e i n t h e p r o p o r t i o n o f t h e
form b e tw e e n t h e t h r e e m u s c l e s s t u d i e d e v e n th o u g h t h e EDL
c o n t a i n e d a l a r g e p r o p o r t i o n o f t y p e I f i b r e s w h ich w e r e t h o u g h t
t o c o n t a i n g r e a t e r a m o u n ts o f L^ AChE ( L y l e s e t a K , 1 9 8 2 ;
B arn ard e t a K , 1 9 8 2 ) . The i n c r e a s e in t o t a l AChE a c t i v i t y i n
b o th t h e AT (1 w eek ) and EDL ( 3 w e e k s ) w as due t o an i n c r e a s e i n
t h e am ount o f fo rm . On t h e o t h e r h an d , t h e l a r g e i n c r e a s e
i n AChE in t h e g a s t r o c n e m iu s was c o n t r i b u t e d by an i n c r e a s e i n
a l l t h e fo rm s ( F i g . 4 . 7 and T a b le 4 . 5 ) .
The AChE a c t i v i t i e s i n t h e s e m u s c l e s in d y s t r o p h i c c h i c k e n s
w e r e o n l y m o d e r a t e l y e l e v a t e d ( T a b le 4 . 5 ) . A n a l y s i s o f t h e AChE
form s show ed t h a t in t h e c o n t r o l a l l t h e m o l e c u l a r fo rm s w e r e
o v e r p r o d u c e d : in t h e AT, L ,8 ; M ,3 ; 3 t i m e s t h e normal
AT ( F i g - 4 . 8 A ) ; i n t h e g a s t r o c n e m iu s , L, 13 • H2 c , 2 t i m e s
t h e normal g a s t r o c n e m iu s ( F i g . 4 . 8 C ) . C h r o n ic s t i m u l a t i o n o f
t h e s e m u s c l e s r e s u l t e d in an i n c r e a s e i n AChE c o n t e n t , c o n t r a r y
t o t h a t o b s e r v e d i n t h e d y s t r o p h i c PLD. I t l e d t o a p r e f e r e n t i a l
i n c r e a s e in t h e H^ AChE i n t h e AT ( F i g . 4 . 8 B ) , b u t i n t h e
g a s t r o c n e m i u s , o n l y t h e L^ AChE was s i g n i f i c a n t l y e l e v a t e d .
4 . 4 DISCUSSION
4 . 4 . 1 C h a n g es o f AChE i n s t i m u l a t e d norm al c h ic k e n PLD
The r e s u l t s o f t h i s c h a p t e r show t h a t c h r o n i c lo w f r e q u e n c y
- 134 -
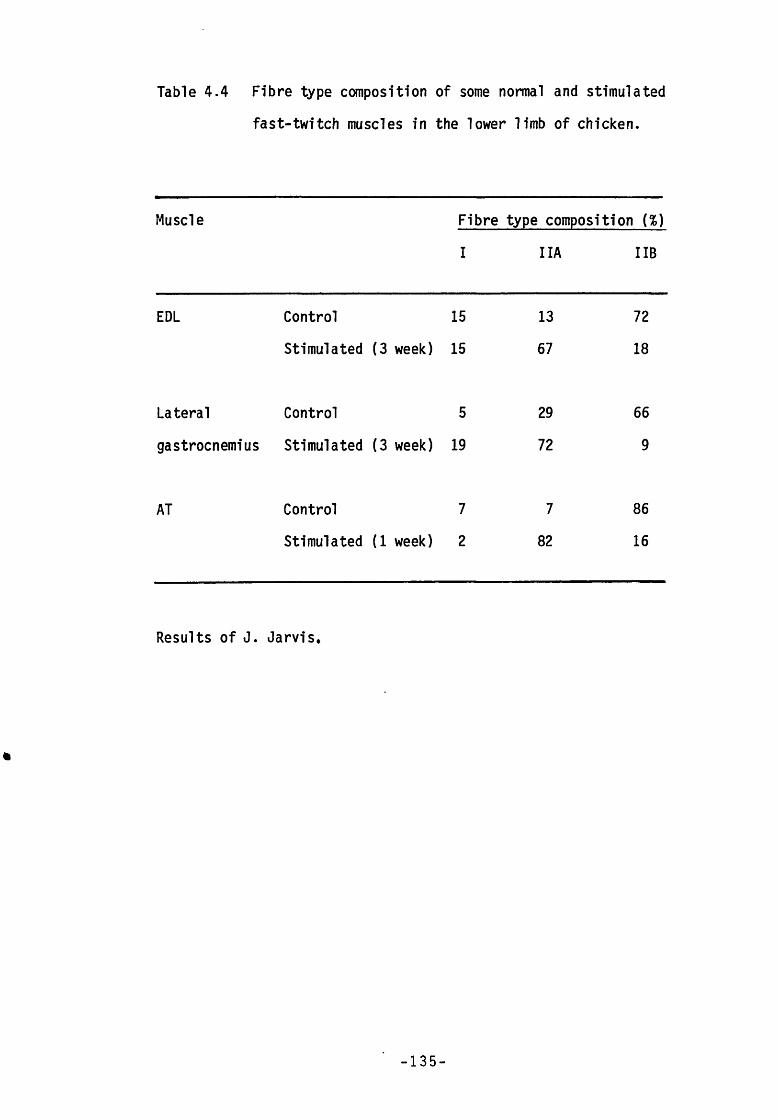
Table 4.4 Fibre type composition of some normal and stimulated
f a s t - t w i t c h m u s c l e s i n t h e lo w e r l im b o f c h i c k e n .
M u s c le F i b r e t y p e c o m p o s i t i o n [%)
I IIA I IB
EDL C o n tr o l 15 13 72
S t i m u l a t e d ( 3 w eek ) 15 67 18
L a t e r a l C o n tr o l 5 29 66
g a s t r o c n e m i u s S t i m u l a t e d ( 3 w eek ) 19 72 9
AT C o n tr o l 7 7 8 6
S t i m u l a t e d (1 w eek ) 2 82 16
R e s u l t s o f J . J a r v i s

-136
-
Table 4.5 Increases in AChE concentration in leg muscles of normal and dystrophic chickens after
d i f f e r e n t p e r i o d s o f s t i m u l a t i o n .
M u s c le F i n a l
a g e ( w k s )
L e n g th o f
S t i m u l a t i o n ( w k s )
[AChE] ( U / g ) R e l a t i v e a c t i v i t y
C o n tr o l S t im a l l form s L MH1C H2C
AT( M*) 7 1 0 . 3 5 0 . 6 6 1 . 9 2 . 3 - - 1 . 1
EDL(N) 9 3 0 . 4 6 0 . 7 2 1 . 6 2 . 7 2 . 0 - 1 . 1
g a s t r o c n e m i u s ( N ) 9 3 0 . 5 3 2 . 4 2 4 . 6 1 3 . 3 5 . 4 6 . 5 3 . 5
AT(D*) 10 3 1 . 3 3 . 9 3 . 0 1 . 9 2 . 9 - 2 . 4
g a s t r o c n e m i u s ( D ) 10 3 1 . 5 2 . 6 1 . 7 2 . 2 - - 1 . 0
* N = norm al c h i c k e n
D = d y s t r o p h i c c h i c k e n

ConN StimN
AA
FRACTIONS
FIG- 4.7 S u c r o s e g r a d i e n t p r o f i l e s o f AChE i n l e g m u s c l e s o f
norm al c h i c k e n a f t e r c h r o n i c s t i m u l a t i o n . Con N ( c o n t r a l a t e r a l
u n s t i m u l a t e d normal m u s c l e ) ; S t im N ( s t i m u l a t e d normal m u s c l e ) .
A,B: a n t e r i o r t i b i a l i s ; C,D: EDL E,F: g a s t r o c n e m i u s . EDL and
g a s t r o c n e m i u s w ere s t i m u l a t e d f o r 3 w e e k s ; AT was s t i m u l a t e d f o r
1 w e e k .
- 137 -

ConD Sti mD
AA
FR A CTIO N S
F IG - 4 - 8 S u c r o s e g r a d i e n t p r o f i l e s o f AChE i n l e g m u s c l e s o f
d y s t r o p h i c c h i c k e n a f t e r c h r o n i c s t i m u l a t i o n . Con D
( c o n t r a l a t e r a l u n s t i m u l a t e d d y s t r o p h i c m u s c l e ) ; S t im 0
( s t i m u l a t e d d y s t r o p h i c m u s c l e ) . A ,B : a n t e r i o r t i b i a l i s ; C ,D :
g a s t r o c n e m i u s . B oth m u s c l e s w ere s t i m u l a t e d f o r 3 w e e k s .
- 138 -

s t i m u l a t i o n b r i n g s a b o u t a l a r g e i n c r e a s e i n t h e a c t i v i t y o f AChE
i n normal PLD m u s c l e . T h is may b e a t t r i b u t e d t o i n c r e a s e d
b i o s y n t h e s i s i n t h e s t i m u l a t e d m u s c le i n r e s p o n s e t o i n c r e a s e d
m u s c le a c t i v i t y . T h i s d e d u c t io n i s b a s e d on o t h e r o b s e r v a t i o n s
on e l e c t r i c a l l y - s t i m u l a t e d mammalian m u s c l e s . T h u s , H e i l i g and
P e t t e ( 1 9 8 3 ) show ed t h a t s t i m u l a t i o n a t 10 Hz f o r 4 w e e k s o f
r a b b i t f a s t - t w i t c h m u s c l e s i n c r e a s e d t h e t o t a l RNA i n t h e m u s c le
an d t h e s lo w m y o s in l i g h t c h a i n mRNAs. S i m i l a r s t u d i e s by
M a s t r i , S a lm on s and Thomas ( 1 9 8 2 ) showed t h a t t h e c o n t e n t o f
p r o t e i n s y n t h e s i s - a s s o c i a t e d p o ly a m in e s and o f t h e en zy m es
p r o d u c in g them w as a l s o m a rk ed ly e l e v a t e d by c h r o n i c s t i m u l a t i o n .
H o w ev er , t h e e f f e c t o f s t i m u l a t i o n on p r o t e i n s y n t h e s i s i s
d e p e n d e n t upon t h e t y p e s o f m u s c le p r o t e i n : t h e l e v e l s o f enzym es
o f o x i d a t i v e m e t a b o l i s m a r e c o n s i d e r a b l y i n c r e a s e d i n s t i m u l a t e d
mammalian m u s c l e s , b u t a r e r e d u c e d f o r some o t h e r p r o t e i n s , e . g .
t h o s e r e s p o n s i b l e f o r c a l c i u m s e q u e s t r a t i o n and pumping a s w e l l
a s en zy m es o f a n a e r o b i c m e t a b o l i s m (K lug e t al^., 1 9 8 3 ) . In
a c c o r d a n c e w i t h t h i s , an abnorm al i n c r e a s e in t h e s y n t h e s i s o f
t h e "slow " i s o f o r m s o f v a r i o u s c o n t r a c t i l e p r o t e i n c h a i n s o c c u r s
w h i l e t h e p r o d u c t io n o f t h e c o r r e s p o n d i n g " f a s t " i s o f o r m s
d e c l i n e s , in r e s p o n s e t o l o n g p e r i o d s o f s t i m u l a t i o n o f f a s t -
t w i t c h mammalian m u s c l e s a t f r e q u e n c i e s (1 0 Hz) c h a r a c t e r i s t i c o f
t h e n a t u r a l a c t i v i t y o f s l o w - t w i t c h f i b r e s ( P e t t e e t a K , 1 9 7 6 ;
R u b i n s t e i n e t a K , 1 9 7 8 ) . In g e n e r a l , t h e r e s p o n s e t o su ch
a p p l i e d s t i m u l a t i o n , o r t o n a t u r a l s t i m u l a t i o n i n s u s t a i n e d
e x e r c i s e (S a lm o n s and H e n r i k s s o n , 1 9 8 1 ; G reen e t , 1 9 8 3 ) , i s a
' s w i t c h - o n ' o f t h e p r o t e i n s r e q u i r e d f o r s l o w - f i b r e a c t i v i t y and
a ' s w i t c h - o f f ' o f t h o s e f o r f a s t - f i b r e a c t i v i t y .
- 139 -

A v ia n m u s c l e s have n o t p r e v i o u s l y b e e n c h r o n i c a l l y
s t i m u l a t e d and a n a l y s e d w i t h r e s p e c t t o p r o t e i n s y n t h e s i s o r mRNA
c h a n g e s , b u t t h e o v e r a l l e f f e c t s o f in d u c e d a c t i v i t y on c h i c k e n
m u s c l e s a r e l i k e l y t o be s i m i l a r . We h a v e o b s e r v e d i n a p a r a l l e l
s t u d y (B a r n a r d e t a ^ . , 1 9 8 5 ) u s i n g f i b r e - t y p i n g t e c h n i q u e t h a t in
t h e s t i m u l a t e d c h i c k e n PLD m u s c l e s u s e d h e r e , a s i m i l a r
t r a n s f o r m a t i o n o f g l y c o l y t i c t y p e IIB f i b r e s t o h i g h l y o x i d a t i v e
t y p e IIA f i b r e s h a s o c c u r r e d . The i n c r e a s e o b s e r v e d i n t h e
m it o c h o n d r i a and l i p i d c o n t e n t r e s e m b l e s c l o s e l y t h e
u l t r a s t r u c t u r a l m o rp h o lo g y o f a p r e d o m in a n t ly t y p e I IA m u s c l e ,
nam ely t h e a d d u c t o r s u p e r f i c i a l i s ( J . C avan agh , p e r s o n a l
c o m m u n ic a t io n ) . T h er e was no s u b s t a n t i a l c h a n g e , h o w e v e r , i n t h e
t i m e - t o - p e a k o f i s o m e t r i c t w i t c h c o n t r a c t i o n o r t h e h a l f -
r e l a x a t i o n t im e o f t h e m u s c l e , up t o 8 w e e k s o f c o n t i n u o u s
s t i m u l a t i o n . T h i s i s i n c o n t r a s t t o t h e b e h a v i o u r o f s i m i l a r l y
s t i m u l a t e d mammalian f a s t - t w i t c h m u s c l e s , w h ose c o n t r a c t i l e
c h a r a c t e r i s t i c s and c y t o c h e m i s t r y a r e s i m i l a r t o t h o s e o f
s l o w - t w i t c h - o x i d a t i v e t y p e I f i b r e s ( R u b i n s t e i n e t a K , 1 9 7 8 ;
H eilm an n and P e t t e , 1 9 7 9 ) . In t h i s t r a n s f o r m a t i o n from t y p e I IB
t o IIA i n t h e c h i c k e n m u s c l e , i t i s c l e a r t h a t t h e o x i d a t i v e
enzym e s y n t h e s i s i s a c t i v a t e d and i t i s now shown h e r e t h a t
* m u s c le AChE p r o d u c t io n a l s o g r e a t l y i n c r e a s e s . T h is i s
i n t e r e s t i n g , s i n c e normal a v i a n t y p e IIA f i b r e s do n o t c o n t a i n , o n
a v e r a g e , more AChE th a n do t h e t y p e I IB f i b r e s , w h e r e a s t h e
c o n t e n t i n a v i a n m u s c l e s r i c h i n t y p e I f i b r e s i s a b o u t 1 0 - f o l d
g r e a t e r (B a r n a r d e t a K , 1 9 8 2 , 1 9 8 4 ) . H en ce , i t a p p e a r s t h a t t h e
t r a n s f o r m e d f i b r e s have a t t a i n e d some b u t n o t a l l o f t h e v a r i o u s
s p e c i f i c p r o p e r t i e s o f t y p e I f i b r e s .
- 140 -

M a s t r i e t a]_. ( 1 9 8 2 ) show ed t h a t t h e b i o s y n t h e t i c a c t i v i t i e s
o f a s t i m u l a t e d f a s t - t w i t c h mammalian m u s c le a r e t r a n s i e n t l y
e l e v a t e d i n a p h a s i c m an n er , many t im e s a b o v e n o r m a l , b e f o r e
l e v e l l i n g o f f t o a new s t e a d y s t a t e t y p i c a l o f a s l o w - t w i t c h
m u s c l e . A s i m i l a r t i m e - d e p e n d e n t c h a n g e i n t o t a l mRNA c o n t e n t o f
su ch m u s c le h a s a l s o b een o b s e r v e d ( P e t t e , 1 9 8 4 ) . In t h i s s t u d y ,
i t w as fo u n d t h a t AChE f o l l o w e d a s i m i l a r p h a s i c i n c r e a s e d u r in g
t h e c o u r s e o f s t i m u l a t i o n . A f t e r two w eek s o f c h r o n i c
s t i m u l a t i o n , t h e PLD m u s c le e x h i b i t e d t h e h i g h e s t l e v e l o f AChE
a c t i v i t y , a b o u t 5 t i m e s t h a t o f t h e c o n t r o l , w h ic h t h e r e a f t e r
m a in t a i n e d a t a l o w e r l e v e l o f 3 t i m e s t h e c o n t r o l v a l u e . T h is
p h a s i c p a t t e r n was b e t t e r i l l u s t r a t e d by t h e r e m a r k a b le i n c r e a s e
i n t h e L fo rm s o b s e r v e d o v e r t h e 8 w e e k s o f s t i m u l a t i o n . As t h e
e l e v a t e d l e v e l o f t h e H AChE w a s , o n c e a t t a i n e d , c o n s t a n t
t h r o u g h o u t t h e s t i m u l a t i o n p e r i o d , t h e l a t e r p r o g r e s s i v e
r e d u c t i o n in t h e L form s f o l l o w i n g t h e i n i t i a l i n c r e a s e was n o t a
r e s u l t o f a p r o g r e s s i v e i n c r e a s e in t h e a s s e m b ly o f t h e H f o r m s ,
b u t i s l i k e l y t o r e f l e c t a r e a l r e d u c t i o n i n t h e s y n t h e s i s o f t h e
L f o r m s . M i c r o d i s s e c t e d e n d p l a t e s and n o n - e n d p l a t e r e g i o n s o f
t h e 3 -w eek s t i m u l a t e d PLD i n d i c a t e d t h a t a t l e a s t p a r t o f t h e
e x c e s s H AChE i s l o c a t e d e x t r a - j u n c t i o n a l l y ( F i g . 4 . 3 ) . T h e s e
* o b s e r v a t i o n s s u g g e s t a b u i l d - u p o f c o l l a g e n - t a i l e d AChE i n an
e x t r a j u n c t i o n a l p o o l w h ic h i s n o t im m e d ia t e ly t r a n s p o r t e d t o t h e
e n d p l a t e . The H2 c form r e m a in e d t h e o n l y e n d p l a t e - s p e c i f i c
fo r m , w h i l e t h e H^c , l a r g e l y e x t r a j u n c t i o n a l , c o u l d be t h e
im m e d ia te p r e c u r s o r o f t h e H2 c fo r m , and n o t i t s breakdow n
p r o d u c t a t t h e e n d p l a t e s . The am ount o f H AChE a t e a c h e n d p l a t e
- 141 -

w as n o t s i g n i f i c a n t l y g r e a t e r th a n t h a t i n t h e u n s t i m u l a t e d
m u s c l e , s o we d e d u c e t h a t t h e r e a r e a f i x e d number o f s i t e s f o r
i t s a t t a c h m e n t i n t h e b a s a l la m in a t h e r e , l e a d i n g t o t h e e x c e s s H
AChE a c c u m u la t in g in i n t e r n a l m em branes. On t h e o t h e r h a n d ,
t h e s e e x t r a j u n c t i o n a l H AChE may r e s e m b le t h e e x t r a j u n c t i o n a l H
AChE fo u n d in mammalian t y p e I f i b r e s , w h ic h s u g g e s t s some
p o s s i b l e l o c a l i s a t i o n o r f u n c t i o n o f t h e s e form s o u t s i d e t h e
e n d p l a t e i n r e s p o n s e t o i n c r e a s e d m u s c le a c t i v i t y .
T h e r e i s now c o n s i d e r a b l e e v i d e n c e t o i n d i c a t e t h a t t h e
b i o s y n t h e s i s and a s s e m b ly o f AChE fo rm s i s p r o g r e s s i v e ,
com m encing w i t h t h e s y n t h e s i s o f L form s in t h e s a r c o p l a s m i c
r e t i c u l u m ( s e e G en era l I n t r o d u c t i o n ) . The L form s a r e
s u b s e q u e n t l y a s s e m b le d i n t o M and H f o r m s . By f o l l o w i n g t h e
g l y c o s y l a t i o n o f t h e a c t i v a t e d AChE m o l e c u l e s , R otundo ( 1 9 8 4 a )
h a s shown t h a t t h e M form i s a s s e m b le d in t h e rough e n d o p la s m ic
r e t i c u l u m (RER), t r a n s p o r t e d t o t h e G o lg i a p p a r a t u s w h ere i t i s
a s s e m b le d i n t o t h e c o l l a g e n - t a i l e d H f o r m s , and f i n a l l y e x p o r t e d
t o t h e e x t r a c e l l u l a r m a t r i x a t t h e n e u r o m u s c u la r j u n c t i o n . In
t h e u n s t i m u l a t e d s t a t e , t h e r e t h u s a p p e a r s t o be a b a l a n c e
b e tw e e n t h e s y n t h e s i s o f t h e L form s and t h e i r a s s e m b ly i n t o H
fo rm s su ch t h a t o v e r 80% o f t h e AChE i s p r e s e n t a s t h e j u n c t i o n a l
H2C form i n t h e PLD m u s c l e . A b u i l d - up o f H fo rm s was
o b s e r v e d when a new s t e a d y d i s t r i b u t i o n o f AChE fo rm s had b een
r e a c h e d in t h e s t i m u l a t e d PLD. The t u r n o v e r o f e n d p l a t e H AChE
i s in o t h e r s i t u a t i o n s known t o be v e r y s lo w com p ared t o t h a t o f
i n t e r n a l L AChE ( R o tu n d o , 1 9 8 4 ; B r i m i j o i n , 1 9 8 3 ; Newman e t a l . ,
1 9 8 4 ) , so t h i s new s t e a d y s t a t e o f H2c AChE ( a t 3 t i m e s t h e
normal l e v e l , o v e r a l l ) can be u n d e r s t o o d in t e r m s o f t h e s e
d i f f e r e n t i a l l i f e t i m e s and t h e i n i t i a l f a s t and l a t e r s lo w
s y n t h e s i s o f t h e L p r e c u r s o r .
- 1 4 2 -

T h a t t h e e f f e c t s o f s t i m u l a t i o n a r e n o t p e r m a n en t and
r e q u i r e t h e c o n t i n u o u s a c t i v i t i e s o f t h e n e r v e i s d e m o n s t r a t e d by
t h e r e v e r s a l i n AChE a c t i v i t y and t h e r e t u r n o f i t s m o l e c u l a r
p r o f i l e , upon c e s s a t i o n o f t h e e l e c t r i c a l i n p u t ( T a b l e 4 . 2 ) ,
P r o d u c t i o n o f t h e L AChE r a p i d l y d e c l i n e s and t h e a s s e m b ly i n t o
H£C v i a H^c i s d i m i n i s h e d t o i t s normal lo w l e v e l . The
h i s t o c h e m i c a l c h a n g e s a l s o u n d er w en t a s i m i l a r r e v e r s a l , a l t h o u g h
o v e r a som ew hat s l o w e r t im e c o u r s e ( J . J a r v i s , p e r s o n a l
c o m m u n i c a t i o n ) . The r e v e r s a l o f t h e e f f e c t o f
s t i m u l a t i o n - i n d u c e d c h a n g e s i n c o n t r a c t i l e a c t i v i t i e s o f
mammalian t y p e I I m u s c le h a s a l s o b e e n r e p o r t e d ( S a lm o n s , 1 9 8 0 ) .
The PLD AChE a c t i v i t y g r a d u a l l y a p p r o a c h e d a l e v e l s i m i l a r t o
t h a t o f t h e c o n t r o l PLD a f t e r 14 d a y s ( T a b l e 4 . 2 ) .
O v e r a l l , t h e e f f e c t s s e e n i n normal a v i a n m u s c le show t h a t
t h e p r o d u c t i o n o f m u s c le AChE i s r e g u l a t e d by t h e a c t i v i t y o f t h e
m o to r n e r v e . The e f f e c t s on m u s c le AChE o f d e n e r v a t i o n and o f
c o n t r a c t i o n b l o c k a d e , a s r e v ie w e d b r i e f l y i n t h e I n t r o d u c t i o n ,
co m b in ed w i t h t h i s e v i d e n c e on s t i m u l a t i o n e f f e c t s , s e r v e t o show
t h a t e l e c t r i c a l a c t i v i t y p e r S £ i s an i m p o r t a n t c o n t r o l l i n g
f a c t o r f o r t h i s e n z y m e . On t h e o t h e r h a n d , a s n o t e d e a r l i e r
c e r t a i n e f f e c t s o f n e r v e e x t r a c t s on m u s c le AChE show t h a t
* t r o p h i c f a c t o r s can a l s o c o n t r i b u t e t o i t s r e g u l a t i o n .
4 . 4 . 2 C h a n g es o f AChE i n s t i m u l a t e d d y s t r o p h ic c h ic k e n PLD
In c o n t r a s t t o t h e i n c r e a s e i n AChE a c t i v i t y i n s t i m u l a t e d
normal m u s c l e s , c h r o n i c low f r e q u e n c y s t i m u l a t i o n o f d y s t r o p h i c
PLD m u s c l e s u n e q u i v o c a l l y l e d t o a r e d u c t i o n o f AChE a c t i v i t y in
t h e s e m u s c l e s . One o f t h e s a l i e n t f e a t u r e s o f a v i a n d y s t r o p h y i s
- 143 -

t h e l a r g e i n c r e a s e in AChE a c t i v i t y i n a f f e c t e d m u s c l e s ( W i l s o n
e t a K , 1 9 7 9 ) ; t h i s i n c r e a s e i s due t o t h e e x c e s s i v e s y n t h e s e s o f
a l l m o l e c u l a r f o r m s , b u t i n p a r t i c u l a r t h e L fo rm s ( c f . F i g .
4 . 1 A, 4 . 4 A ) . C h r o n ic s t i m u l a t i o n in t u r n p r e f e r e n t i a l l y
s u p p r e s s e d t h e s y n t h e s i s o f L f o r m s ; t h i s e f f e c t w as m o s t c l e a r l y
i l l u s t r a t e d i n t h e PLD o f t h e 1 4 - w e e k - o l d c h i c k e n w h ic h had b e e n
s t i m u l a t e d f o r 5 w e e k s ( F i g . 4 . 4 3 , F ) . The AChE a c t i v i t y a s w e l l
a s i t s m o l e c u l a r p r o f i l e i n t h e s t i m u l a t e d d y s t r o p h i c m u s c le
r e s e m b le d c l o s e l y t h a t o f a s i m i l a r l y s t i m u l a t e d normal PLD
( F i g . 4 . 1 F ) . The same i s t r u e f o r t h e norm al and d y s t r o p h i c
m u s c l e s w h ic h had b e e n s t i m u l a t e d f o r 2 w e e k s ( c f . F i g . 4 . 1 B and
4 . 4 B ) . Over t h e 5 -w eek s t i m u l a t i o n a t 1 0 Hz, t h e r e was c l e a r l y a
p r o g r e s s i v e r e d u c t i o n o f t h e p r o p o r t i o n o f L fo rm s i n t h e
d y s t r o p h i c m u s c l e . T h e s e o b s e r v a t i o n s a r e i n t e r e s t i n g , a s t h e y
show t h a t c h r o n i c s t i m u l a t i o n b r i n g s a b o u t c o m p a r a b le
d i s t r i b u t i o n o f AChE fo rm s in normal and d y s t r o p h i c PLD d e s p i t e
t h e p a t h o l o g i c a l c h a n g e s t h a t had t a k e n p l a c e i n t h e d y s t r o p h i c
m u s c le p r i o r t o s t i m u l a t i o n . The d i f f e r e n t i a l l o c a l i s a t i o n o f
AChE i n and o u t s i d e t h e n e u r o m u s c u la r j u n c t i o n s u b s t a n t i a t e s
t h e s e r e s u l t s . H ence i t sh ow s t h a t a l t h o u g h t h e d y s t r o p h i c
m u s c le c o n s i s t e d o f abnorm al am ou n ts o f L an d H2 C form o u t s i d e
t h e e n d p l a t e s , s t i m u l a t i o n t r a n s f o r m e d t h i s d i s t r i b u t i o n i n t o a
p r o f i l e w i t h p r e d o m in a n t ly H^c and H2 C fo r m s , a s i n t h e
s t i m u l a t e d normal m u s c le ( c f . 4 .6 D and 4 . 3 0 . H i s t o c h e m i c a l and
m o r p h o lo g i c a l f i n d i n g s (B a r n a r d e t a l^ . , 1 9 8 5 ) p r o v i d e f u r t h e r
i n f o r m a t i o n f o r t h e s i m i l a r i t i e s b e tw e e n t h e r e s p o n s e s o f t h e
normal and d y s t r o p h i c PLD t o s t i m u l a t i o n ; t h e y i n c l u d e i n c r e a s e d
f a t i g u e r e s i s t a n c e and t y p e I IB t o 11A t r a n s f o r m a t i o n o f f i b r e
- 144 -

t y p e . I n d e e d , t h e r e m a r k a b le im p ro v em en t in t h e s t r u c t u r e and
i n t e g r i t y o f t h e d y s t r o p h i c m u s c le f i b r e s upon s t i m u l a t i o n
d e m o n s t r a t e s t h e e f f e c t o f c h r o n i c s t i m u l a t i o n n o t s e e n w i t h
norm al m u s c l e s .%
T h e r e a r e , t h e r e f o r e , two d i s t i n c t i v e a s p e c t s w i t h r e g a r d t o
t h e c h a n g e s i n AChE i n t h e s t i m u l a t e d d y s t r o p h i c PLD. F i r s t l y ,
t h e s y n t h e t i c m a c h in e r y f n t h e d y s t r o p h i c m u s c le r e s p o n d e d t o
I n c r e a s e s 1n c o n t r a c t i l e a c t i v i t y In much t h e same way a s a
norm al m u s c l e d i d . S t i m u l a t i o n m a i n t a i n e d l a r g e am ou n ts o f t h e
e x t r a - j u n c t i o n a l H AChE a f t e r an I n i t i a l a c c u m u l a t i o n o f t h e L
f o r m s . S e c o n d l y , t h e e x c e s s i v e s y n t h e s i s o f t h e L fo r m s w as
r a p i d l y s u p p r e s s e d . T h e s e s i m u l t a n e o u s b u t o p p o s i n g e f f e c t s a r e
c o n s i s t e n t w i t h t h e Id e a t h a t t h e d y s t r o p h i c a v i a n m u s c l e I s
a r r e s t e d a t an e a r l y d e v e lo p m e n t a l s t a g e (L AChE b e i n g v e r y h ig h
In norm al e m b r y o n ic m u s c l e : L y l e s e t a K , 1 9 7 9 ) b u t I s s t i l l
c a p a b l e o f r e s p o n d i n g t o Im posed r e g u l a t o r y f a c t o r s .
An abnorm al e x c e s s o f AChE o c c u r s 1n b o t h t h e e n d p l a t e
and n o n - e n d p l a t e r e g i o n s o f d y s t r o p h i c m u s c l e ( F 1 g . 4 . 6 A ,C ) , an d
I s e l i m i n a t e d by Im posed m u s c le a c t i v i t y ( F 1 g . 4 . 6 B , D ) . T h i s
sh ow s t h a t t h e e l e c t r i c a l e f f e c t s a r e n o t e x e r t e d o n l y on t h e
e x t r a - j u n c t i o n a l membrane a s s e e n 1n s t i m u l a t e d normal PLD. The
e l e c t r i c a l l y s t i m u l a t e d d y s t r o p h i c PLD m a i n t a i n s m o s t o f I t s
e x c e s s AChE a s c o l l a g e n - t a i l e d m o l e c u l e s w h ic h a r e l o c a t e d
e x t r a - j u n c t l o n a l l y ( F 1 g . 4 . 6 D ) , w h ic h 1 s t h e same s t a t e a s 1 s
fo u n d 1n t h e e l e c t r i c a l l y s t i m u l a t e d normal PLD.
T h e s e p a r a l l e l s show t h a t a l t h o u g h t h e o v e r p r o d u c t i o n o f
AChE 1 s g e n e t i c a l l y d e t e r m in e d In t h e d y s t r o p h i c a v i a n f a s t
m u s c l e s , I t i s n e v e r t h e l e s s n o t e n t i r e l y rem oved from r e g u l a t o r y
- 145 -

i n f l u e n c e s . I t i s b r o u g h t a b o u t by s u s t a i n e d e l e c t r i c a l a c t i v i t y
t o t h e same s t a t e , i n a m ou n t , fo rm s and l o c a t i o n s , a s i n t h e
s i m i l a r l y s t i m u l a t e d normal m u s c l e . The n a t u r e o f t h e p a r t i c u l a r
c o n t r o l o f AChE w h ic h i s abnorm al i n t h e d y s t r o p h i c p h e n o t y p e ,
an d w h ich may be a r e f l e c t i o n o f an e x t e n d e d im m atu re s t a t e o f
t h e s e a n i m a l s (Bandm an, 1 9 8 4 ) , a w a i t s f u r t h e r s t u d y , s i n c e i t i s
l i k e l y t o th ro w new l i g h t on r e g u l a t o r y c o n t r o l s i n m u s c le
d e v e lo p m e n t .
4 . 4 . 3 C h a n g es o f AChE i n l e g m u s c le o f norm al a n d c jy s t r o p h ic
c h i c k e n s
AChE i n t h e f a s t t w i t c h l e g m u s c l e s p r e s e n t e d i n t h i s s t u d y ,
l i k e t h e f a s t - t w i t c h PLD m u s c l e , c o n s i s t s l a r g e l y o f HgQ*
C h r o n ic s t i m u l a t i n g a v e r i s e t o a s u b s t a n t i a l e l e v a t i o n o f t h e
L2 f o r m , when t h e m u s c le f i b r e s becom e p r e d o m in a n t ly t y p e I I A .
U n l i k e t h e s t i m u l a t e d norm al PLD, t h e s e m u s c l e s (EDL an d AT) do
n o t a p p e a r t o u n d er g o a p h a s i c i n c r e a s e in AChE a s t h e r e was
no d i f f e r e n c e b e tw e e n 1 week o r 3 w eek s s t i m u l a t i o n . T h is
p r e f e r e n t i a l i n c r e a s e i n t h e Lg AChE i s t h u s s i m i l a r b u t
som ew hat l e s s m arked a s t h a t o b s e r v e d i n t h e s t i m u l a t e d PLD. On
t h e o t h e r h a n d , t h e g a s t r o c n e m i u s r e s p o n d e d t o c h r o n i c
s t i m u l a t i o n w i t h a r e m a r k a b le i n c r e a s e i n t o t a l AChE a c t i v i t y .
A f t e r 3 w e e k s c o n t i n u o u s s t i m u l a t i o n , t h e AChE a c t i v i t y
a p p r o a c h e d t h a t o b s e r v e d i n a h i g h l y o x i d a t i v e m u s c le su c h a s t h e
r e d s a r t o r i u s o r t h e m ed ia l g a s t r o c n e m i u s (B a r n a r d e t al^., 1 9 8 2 ) .
The i n c r e a s e in t h e number o f t y p e I f i b r e s i n t h e
s t i m u l a t e d g a s t r o c n e m i u s s u g g e s t s t h a t c h r o n i c s t i m u l a t i o n a t 10
Hz ca n in d u c e a f a s t t o s lo w t r a n s f o r m a t i o n in c h i c k e n m u s c l e s .
- 146 -

The f a c t t h a t m u s c l e s r e s p o n d d i f f e r e n t l y t o t h e same s t i m u l a t i o n
s c h e d u l e s u g g e s t s f a c t o r s o t h e r th a n t h e i m p u ls e f i r i n g p a t t e r n
d e t e r m in e t h e f i b r e t y p e c o m p o s i t i o n o f a m u s c l e . I t a l s o
s u g g e s t s t h a t p r e f e r e n t i a l u t i l i s a t i o n o f t h e g a s t r o c n e m i u s may
h a v e o c c u r r e d d u r in g s t i m u l a t i o n , s i n c e t h e p r o p o r t i o n o f t y p e I
f i b r e s in t h e EDL o r AT was n o t a f f e c t e d .
In t e r m s o f i n c r e a s e d l e v e l s o f AChE, l e g m u s c l e s in
d y s t r o p h i c c h i c k e n s a r e l e s s a f f e c t e d when com p ared t o t h e
d y s t r o p h i c PLD, w h ich c o r r e l a t e s w i t h t h e i n e x t e n s i v e e f f e c t o f
d y s t r o p h y on t h e s e m u s c l e s . The t o t a l AChE a c t i v i t y i s i n c r e a s e d
m o d e r a t e l y , b u t i s h i g h e r th a n p r e v i o u s l y o b s e r v e d ( L y l e s e t a l . ,
1 9 8 2 ; B arn ard e t a l^ . , 1 9 8 2 ) . AChE a c t i v i t i e s i n c r e a s e d i n t h e s e
l e g m u s c l e s in d y s t r o p h i c c h i c k e n s upon s t i m u l a t i o n . In t h e
s t i m u l a t e d AT, H2c and a s i g n i f i c a n t H^c c o m p o n e n t now
p r e d o m in a te ( F ig 4 . 8 B ) . The i n c r e a s e in t h e s e fo rm s and t o t a l
AChE a r e a b o u t 5 t i m e s t h a t e x h i b i t e d by t h e normal AT,
s u g g e s t i n g t h a t t h e m u s c le b e h a v e s more l i k e a normal m u s c le th a n
a d y s t r o p h i c m u s c le a s i s e x p e c t e d from i t s m o r p h o l o g i c a l l y
normal c h a r a c t e r i s t i c s (B a r n a r d e t a l . , 1 9 8 2 ) . H ow ever , s i n c e
b o th t h e AChE a c t i v i t y and i t s m o l e c u l a r d i s t r i b u t i o n i n d i c a t e s
s i g n s o f a b n o r m a l i t y , i t i s l i k e l y t h a t t h e m u s c le d o e s c o n t a i n
p a t h o l o g i c a l IIB f i b r e s w h ich a c c o u n t f o r t h e i n c r e a s e d L an d M
s y n t h e s i s , b u t t h e c h a n g e s t h a t o c c u r i n t h e s e f i b r e s a r e
o b s c u r e d by t h e c h a n g e s t h a t t a k e s p l a c e i n t h e m a j o r i t y o f
normal f i b r e s . A v e r y d i f f e r e n t p a t t e r n o f c h a n g e , h o w e v e r ,
t a k e s p l a c e i n t h e g a s t r o c n e m i u s o f t h e d y s t r o p h i c c h i c k e n . T h is
m u s c l e , l i k e t h e d y s t r o p h i c PLD, c o n t a i n s a l a r g e L c o m p o n e n t ,
e v e n th o u g h i t d o e s n o t e x h i b i t s e v e r e d y s t r o p h i c sym p tom s. When
- 147 -

i t was s t i m u l a t e d , t h e s y n t h e s i s o f t h e L form was f u r t h e r
i n c r e a s e d , w h i l e l i t t l e i n c r e a s e in t h e H c o m p o n e n t i s o b s e r v e d .
Such c h a n g e s in t h e g a s t r o c n e m iu s i s t h e r e f o r e i n c o n s i s t e n t w i t h
t h e o b s e r v a t i o n s i n e i t h e r s t i m u l a t e d normal o r d y s t r o p h i c
m u s c l e s ; i n v ie w o f t h e l i m i t e d number o f e x p e r i m e n t s c a r r i e d o u t
f o r t h i s s t u d y , f u r t h e r e x p e r i m e n t a t i o n i s n e c e s s a r y t o v e r i f y
t h e p r e s e n t r e s u l t s .
T h e s e p r e l i m i n a r y f i n d i n g s w i t h c h i c k e n l e g m u s c l e s have
p r o v i d e d more s u b s t a n t i a l i n f o r m a t i o n on t h e e f f e c t s o f
s t i m u l a t i o n on AChE a c t i v i t y and d i s t r i b u t i o n i n f a s t m u s c l e s .
I t i s c l e a r t h a t c h r o n i c lo w f r e q u e n c y s t i m u l a t i o n i n d u c e s an
i n c r e a s e i n t h e o x i d a t i v e c a p a c i t y o f t h e m u s c l e s , a c c o m p a n ie d by
an i n c r e a s e i n AChE s y n t h e s i s . H ow ever, t h e e x t e n t o f i n c r e a s e
i n AChE and t h e p a t t e r n o f c h a n g e i n ter m s o f t h e m o l e c u l a r
c o m p o s i t i o n a p p e a r s t o v a r y b e tw e e n m u s c l e s . S t i m u l a t i o n c a n
a l s o i n d u c e a f a s t t o s lo w t r a n s f o r m a t i o n in c h i c k e n m u s c l e s , !I
w h ic h i s i n d i c a t e d by t h e i n c r e a s e d number o f t y p e I f i b r e s and
t h e l e v e l o f AChE a c t i v i t y c h a r a c t e r i s t i c o f t h e s e f i b r e s . I t
seem s t h a t t h e c o n t r a c t i l e a c t i v i t y a s s o c i a t e d w i t h l e g m u s c l e s
d o e s f a c i l i t a t e t h e f a s t t o s lo w t r a n s f o r m a t i o n . H o w ev er , t h e r e
i s y e t no c l e a r e v i d e n c e w h ich s u g g e s t s t h e f i b r e t y p e
c o m p o s i t i o n o f a m u s c le c o r r e l a t e s w i t h i t s t e n d e n c y o f f a s t t o
s lo w t r a n s f o r m a t i o n . The s e e m i n g ly c o m p le x r e s p o n s e o f l e g
m u s c l e s t o s t i m u l a t i o n may in p a r t be due t o t h e t e c h n i c a l
l i m i t a t i o n o f t h e p r e s e n t s t i m u l a t i o n p r o t o c o l when d e a l i n g w i t h
t h e a n a t o m i c a l l y much m ore co m p le x m u s c u la t u r e i n t h e l e g .
- 148 -

CHAPTER 5
MONOCLONAL ANTIBODIES TO PURIFIED CHICK BRAIN AChE
*
- 149 -

5 . 1 INTRODUCTION
The p h y s i o l o g i c a l e v i d e n c e p r e s e n t e d i n t h e p r e v i o u s
c h a p t e r s s u g g e s t s t h a t t h e s y n t h e s i s and d i s p o s i t i o n o f t h e
m u l t i p l e m o l e c u l a r fo rm s o f AChE i s r e l a t e d t o t h e f u n c t i o n o f
s k e l e t a l m u s c l e s . The l o c a l i s a t i o n o f t h e s e fo rm s i s d e t e r m in e d
by t h e i r c o m p a r t m e n t a l i s a t i o n d u r in g b i o s y n t h e s i s ( R o t u n d o , 1 9 8 4 )
and by t h e i r f i n a l d e s t i n a t i o n w i t h i n t h e m u s c le f i b r e s . The
m o l e c u l a r s t r u c t u r e s o f t h e s e form s a p p e a r t o be a d a p t e d t o t h e i r
s p e c i f i c l o c a l i s a t i o n , w e l l i l l u s t r a t e d by t h e membrane bound
d i m e r i c form o f AChE from human e r y t h r o c y t e s , T o rp ed o e l e c t r i c
o r g a n and human b r a i n , and t h e c o l l a g e n - t a i l e d form s i n m u s c l e s
( s e e g e n e r a l i n t r o d u c t i o n ) . I f i n d e e d t h e l o c a l i s a t i o n o f AChE
fo rm s i s f u n c t i o n a l l y r e l a t e d , and su ch l o c a l i s a t i o n d e p e n d s on
t h e s t r u c t u r a l h e t e r o g e n e i t y o f t h e AChE f o r m s , t h e e l u c i d a t i o n
o f t h e m o l e c u l a r s t r u c t u r e s o f t h e v a r i o u s e n z y m ic fo rm s s h o u l d
p r o v i d e im p o r t a n t i n f o r m a t i o n on t h e r e g u l a t i o n o f s y n t h e s i s ,
l o c a l i s a t i o n , and f u n c t i o n o f t h e s e fo r m s .
The p u r i f i c a t i o n o f AChE fo rm s from s k e l e t a l m u s c le h a s b een
ham pered by t h e i r lo w c o n c e n t r a t i o n . H o w ev er , m o n o c lo n a l
a n t i b o d i e s r a i s e d a g a i n s t p u r i f i e d mammalian AChE from an
a b u n d a n t s o u r c e su ch a s b r a i n t i s s u e s and e r y t h r o c y t e s h a s shown
t h a t t h e s e a n t i b o d i e s c r o s s - r e a c t e d w i t h AChE from o t h e r t i s s u e s
o f t h e same s p e c i e s (Fam brough e t , 1 9 8 2 ) , o r AChE from o t h e r
mammalian s p e c i e s ( B r i m i j o i n e t a l 1 9 8 3 ) The e x p l o i t a t i o n o f
t h i s r e a c t i v i t y f o r a n t i g e n p u r i f i c a t i o n from a d i f f e r e n t t i s s u e
s o u r c e h a s r e c e n t l y b e e n d e m o n s t r a t e d by M in tz and B r i m i j o i n
- 150 -

( 1 9 8 5 ) . Such im m u n o a f f i n i t y c h r o m a t o g r a p h ic t e c h n i q u e s may t h u s
b e u s e d t o p u r i f y m u s c l e AChE. F u r t h e r m o r e , m o n o c lo n a l
a n t i b o d i e s may a l s o be u s e d i n t h e im m u n o h is to c h e m ic a l
l o c a l i s a t i o n o f AChE, p a r t i c u l a r l y a t t h e e l e c t r o n m i c r o s c o p i c
l e v e l , u s i n g c o l l o i d a l g o l d - p r o t e i n A (R o th e t a l . , 1 9 7 8 ) o r
g o l d - l a b e l l e d s e c o n d a n t i b o d y ( S t e r n b e r g e r , 1 9 7 9 ) t e c h n i q u e .
S i m i l a r a p p r o a c h h a s r e c e n t l y b een a p p l i e d t o t h e l o c a l i s a t i o n o f
T o rp ed o a c e t y l c h o l i n e r e c e p t o r and t h e Mr 4 3 , 0 0 0 p r o t e i n
( S e a l o c k e t a l . , 1 9 8 4 . ) T h i s c h a p t e r d e s c r i b e s t h e p u r i f i c a t i o n
o f c h i c k b r a i n AChE and t h e c h a r a c t e r i s a t i o n o f t h e m o n o c lo n a l
a n t i b o d i e s w h ic h w e r e r a i s e d a g a i n s t t h e p u r i f i e d e n z y m e . The
c u r r e n t a p p l i c a t i o n o f t h e s e m o n o c lo n a l a n t i b o d i e s i s a l s o
a s s e s s e d .
5 . 2 MATERIALS AND METHODS
5 . 2 . 1 M a t e r i a l s
C h e m ic a l s f o r p u r i f i c a t i o n ; C o n c a n a v a l in A - A g a r o s e
(C o n A -A g a r o se ) was o b t a i n e d from BRL; a - m e t h y lm a n n o s id e from
F lu k a ; D E A E -Sephacel from S ig m a ; d eca m eth o n iu m from K o c h - L ig h t .
The a f f i n i t y r e s i n , N - m e t h y l a c r i d i n i u m - S e p h a r o s e ( 0 . 3 5 p m o le s
l i g a n d / m l S e p h a r o s e ) w as a g i f t from P r o f e s s o r J . M a s s o u l i e .
C h e m ic a l s f o r SDS PAGE: H igh m o l e c u l a r w e i g h t p r o t e i n s t a n d a r d s
w e r e o b t a i n e d from P h a r m a c ia ; NaDoDSO^ ( S D S ) , d i t h i o t h r e i t o l
from S igm a; p o l y a c r y l a m i d e , t e t r a m e t h y l e t h y l e n e d i a m i n e and
ammonium p e r s u l p h a t e from BDH.
- 151 -

C h e m ic a l s f o r i m m u n i s a t i o n , c u l t u r e and c l o n i n g : F r e u n d s
a d j u v a n t was o b t a i n e d from D i f c o l a b s ( D e t r o i t M i c h . ) ; DMEM,
f e t a l c a l f serum and p e n i c i l l i n / s t r e p t o m y c i n from G ib c o ; s o f t
a g a r o s e from M i l e s S c i e n t i f i c ; a z a s e r i n e , h y p o x a n t h in e from
S ig m a .
C h e m ic a l s f o r s c r e e n i n g and c h a r a c t e r i s a t i o n o f m o n o c lo n a l
a n t i b o d i e s :
R a b b i t a n t i - m o u s e Ig w as o b t a i n e d from M i l e s S c i e n t i f i c ;
FITC and h o r s e r a d i s h p e r o x i d a s e (HRP) c o n j u g a t e d r a b b i t
a n t i - m o u s e I g from DAKO PATTS; p r o t e i n A - S e p h a r o s e , b o v i n e serum
a lb u m in (B S A ), d ia m i n o b e n z i d i n e t e t r a h y d r o c h l o r i d e , N o n id e t
P - 4 0 ( N P - 4 0 ) w e r e from S ig m a ; h y d r o g e n p e r o x i d e (HgOg) and
n i c k e l s u l p h a t e (N iSO ^) w e r e from BRL.
E q u ip m en t
Amicon u l t r a f i l t r a t i o n u n i t f i t t e d w i t h D i a f l o YM30 f i l t e r s
was o b t a i n e d from A m icon ; m i c r o - t i t r e t r a y s and t i s s u e c u l t u r e
t u b e s from G ib c o (N u n c lo n D e l t a ) ; p o l y e t h y l e n e m i c r o t i t r e d i s h
from F a lc o n ( m i c r o t e s t I I I ) ; e l e c t r o b l o t t i n g u n i t from B io r a d and
n i t r o c e l l u l o s e p a p e r from S c h i n d l e r and S c h u e l l . O th e r c h e m i c a l s
a r e a s l i s t e d i n s e c t i o n 2 . 2 . 3 .
5 . 2 . 2 P u r i f i c a t i o n o f c h i c k b r a in ACHE
One day o l d w h i t e l e g h o r n c h i c k s w ere s a c r i f i c e d by
d e c a p i t a t i o n , and t h e b r a i n s rem oved and k e p t on i c e . A b o u t 60g
o f t i s s u e was p o o l e d and h o m o g e n is e d , u s i n g a p o l y t r o n , i n 10 v o l
o f 0.4M N a C l / 1 2 .5 mM p h o s p h a te b u f f e r , pH 7 . 5 c o n t a i n i n g ( a s
- 152 -

p r o t e a s e i n h i b i t o r s ) 5 nM EGTA and b a c i t r a c i n ( l m g / m l ) . The
h o m o g e n a te w as t h e n c e n t r i f u g e d a t 3 0 ,0 0 0 * g f o r 3 0 m in . The
s u p e r n a t a n t w as d e c a n t e d and t h e p e l l e t s w e r e p o o l e d and
h o m o g e n is e d i n 8 v o l o f 0.5M N aC l/0 .5% T r i t o n X - 1 0 0 / 5 0 mM
p h o s p h a t e b u f f e r , pH 7 . 5 c o n t a i n i n g b a c i t r a c i n ( l m g / m l ) , th e n
c e n t r i f u g e d a t 1 0 0 OOOxg f o r 6 0 m in . S u p e r n a t a n t s w e r e th e n
p o o l e d and p a s s e d t h r o u g h a p r e - e q u i l i b r a t e d 5 0 ml Con A -A g a r o s e
colum n o v e r n i g h t a t 4 ° C . The co lum n w as t h e n w ash ed w i t h 8
co lu m n v o l o f 0.5M N a C l /0 .5 % T r i to n X - 1 0 0 / 1 0 nM p h o s p h a t e b u f f e r ,
pH 7 . 5 , f o l l o w e d by 5 co lu m n v o l o f 0 . 5 M a - m e t h y lm a n n o s i d e ( i n
wash b u f f e r ) t o e l u t e t h e bound enzym e o v e r n i g h t a t 4 °C . The
e l u a t e was t h e n d i a l y s e d a g a i n s t 2 x 4 l i t o f 0.5% T r i t o n X - 1 0 0 / 1 0
mM p h o s p h a t e b u f f e r , pH 7 . 5 f o r a t o t a l o f 16 h o u r s , b e f o r e
p a s s i n g t h e d i a l y s a t e th r o u g h a 4 0 ml D E A E -Sephacel c o lu m n ,
p r e - e q u i l i b r a t e d w i t h d i a l y s i s b u f f e r , a t a b o u t 6 0 m l / h o u r . The
enzym e w as e l u t e d w i t h a 0 - 2 5 0 mM NaCI g r a d i e n t i n 18 0 ml o f
d i a l y s i s b u f f e r , and 2 ml f r a c t i o n s w ere c o l l e c t e d . F r a c t i o n s
c o n t a i n i n g t h e peak AChE a c t i v i t y w e r e p o o le d and d i a l y s e d w i t h
t h e T r i t o n / p h o s p h a t e b u f f e r a s a b o v e . The d i a l y s a t e was t h e n
s u b j e c t e d t o a f f i n i t y c h r o m a to g r a p h y u s i n g a 20 ml
N - m e t h y l a c r i d i n i u m - S e p h a r o s e co lu m n ( V a l l e t t e e t a l . , 1 9 8 3 ) ,
w a sh e d w i t h 10 mM N aC l/0 .05% T r i t o n X - 1 0 0 / 1 0 mM p h o s p h a t e , pH 7 . 5 ,
and e l u t e d w i t h t h e same b u f f e r c o n t a i n i n g 10 mM d e c a m e th o n iu m .
F r a c t i o n s w h ic h c o n t a i n e d t h e peak AChE a c t i v i t y w e r e p o o l e d and
d i a l y s e d a g a i n s t 2 x 4 l i t o f wash b u f f e r a t 4°C f o r 2 4 h o u r s . The
d i a l y s a t e o b t a i n e d w as c o n c e n t r a t e d , u s i n g an Ami con
u l t r a f i l t r a t i o n c e l l , t o a b o u t 0 . 5 m l. F r e s h p r e p a r a t i o n
o b t a i n e d t h i s way w as e i t h e r u s e d f r e s h f o r im m u n is a t io n o r
s t o r e d f r o z e n a t -2 0 °C f o r f u r t h e r a n a l y s i s o f p u r i t y .
- 153 -

5 . 2 3 E n zy u e a s s a y
AChE a c t i v i t y w as a s s a y e d by t h e r a d i o m e t r i c m ethod a s
d e s c r i b e d in s e c t i o n 2 . 2 . 7 . A l l a s s a y s w e r e p e r fo r m e d i n t h e
p r e s e n c e o f 1 0 “ S l isoOMPA. P r o t e i n w as m ea su red by t h e m ethod
o f Lowry e t a l . ( 1 9 5 1 ) . BSA w as u s e d a s t h e p r o t e i n s t a n d a r d .
5 . 2 . 4 i H u n i s a t i o n a n d c e l l c u l t u r e
6 - 8 w e e k s o l d f e m a le BALB/c m ic e w e r e im m unised i n t r a -
p e r i t o n e a l l y ( i p . ) , w i t h a p p r o x i m a t e ly 15 pg o f p u r i f i e d AChE i n
c o m p le t e F r e u n d s a d j u v a n t . T h e s e m ic e w ere b o o s t e d w i t h s i m i l a r
a m o u n ts o f p u r i f i e d enzym e i n c o m p le t e F r e u n d s a d j u v a n t e v e r y 2
w eek s u n t i l a p o s i t i v e t i t r e was d e t e c t e d i n t h e serum ( b l o o d
sa m p le w as drawn from t h e t a i l v e i n and t h e serum t e s t e d f o r
a n t i b o d y t i t r e by t h e s o l i d p h a s e m eth od , s e e b e l o w ) . A f i n a l
i n j e c t i o n ( 1 5 pg p u r i f i e d en zy m e) i p . w as g i v e n 4 d a y s b e f o r e
h y b r i d i s a t i o n w as p e r fo r m e d .
S p l e e n c e l l s w e r e h y b r i d i s e d by a m o d i f i c a t i o n ( K e n n e t t e t
a ] . , 1 9 7 8 ) o f t h e K o h l e r - M i l s t e i n p r o c e d u r e ( 1 9 7 5 ) . The s p l e e n
w as g e n t l y d i s s o c i a t e d m e c h a n i c a l l y in 2 ml o f DMEM and t h e c e l l
s u s p e n s i o n was rem o v ed , w a sh ed i n 18 ml o f DMEM and p e l l e t e d .
E x p o n e n t i a l l y g r o w in g inyeloma c e l l s (SP 2/0A G 14 n o n - s e c r e t i n g c e l l
l i n e ) w e r e s i m i l a r l y w a sh ed and 10^ myeloma c e l l s w e r e
c o m b in e d , m ixed w i t h t h e s p l e e n c e l l s and p e l l e t e d . F u s io n was
i n i t i a t e d by a d d in g t o t h e p e l l e t 0 .3 m l o f a 3 3 $ s o l u t i o n o f
p o l y e t h y l e n e g l y c o l 150 0 (BDH) in DMEM. The s u s p e n s i o n was
m ix e d , c e n t r i f u g e d f o r 5 min a t 7 0 0 x g , and t h e p e l l e t w as
a s p i r a t e d d r y and g e n t l y r e s u s p e n d e d i n 1 0 0 ml o f s e l e c t i v e
medium (DMEM, 15$ f e t a l c a l f s e ru m , lp g /m l a z a s e r i n e , 1 0 0 pM
- 154 -

h y p o x a n t h i n e , 1 0 0 u n i t s / m l p e n i c i l l i n / s t r e p t o m y c i n ) . C e l l s w e r e
d i s t r i b u t e d i n t o t e n 96 w e l l m i c r o t i t r e t r a y s and f e d w i t h 1 0 0 pi
o f g r o w th medium ( w i t h o u t a z a s e r i n e and h y p o x a n t h i n e ) a f t e r 7
d a y s o f i n c u b a t i o n a t 3 7 ° C in a h u m i d i f i e d 5 % CO^/95% O2
a t m o s p h e r e . Hybridoma s u p e r n a t a n t s w e r e a s s a y e d f o r
i m m u n o r e a c t i v i t y a s so o n a s t h e c e l l s r e a c h e d c o n f l u e n c e .
H ybridom as w h o se m ed ia w ere p o s i t i v e f o r AChE b i n d i n g ( s e e
b e lo w ) w e r e ex p a n d e d and c l o n e d in s o f t a g a r o s e by t h e m eth od o f
C i v i n an d B a n q u e r in g o ( 1 9 8 3 ) . B r i e f l y , h ybridom a c e l l s t o b e
c l o n e d w e r e s u s p e n d e d in DMEM s u p p le m e n te d medium and f i l t e r e d
t h r o u g h n y lo n mesh t o rem ove c lu m p s o f c e l l s . V i a b l e c e l l c o u n t s
w e r e p e r fo r m e d w i t h a h a e m o c y to m e te r and s e r i a l 1 0 f o l d d i l u t i o n s
5w e r e made t o o b t a i n 2 s e t s o f c u l t u r e t u b e s c o n t a i n i n g 10 ,
1 0 ^ , 10^ and 10^ hybridom a c e l l s i n 150 pi o f medium p e r
c l o n i n g t u b e . F or e a c h s e t o f 8 hybridom a c l o n i n g t u b e s , 9 m is
o f 2 x DMEM s u p p le m e n t e d medium w e r e m ix ed w i t h 9 m is o f 3%
a g a r o s e a t 3 7 ° C. Two m is o f t h i s m ix t u r e w e r e a d d ed t o e a c h
c l o n i n g t u b e , and t h e c o n t e n t m ixed w e l l by t r i t u r a t i o n . T h e s e
t u b e s w e r e t h e n h e l d a t 4°C f o r 45 min t o a l l o w t h e a g a r o s e t o
g e l , a f t e r w h ic h t h e y w e r e i n c u b a t e d a t 37°C i n a h u m i d i f i e d
5% C02/95$ 0^ a t m o s p h e r e u n t i l m a c r o s c o p ic g r o w th o f c l o n e s
a p p e a r e d .
I n d i v i d u a l c l o n e s w as c a r e f u l l y rem oved from t h e a g a r o s e
medium w i t h a p a s t e u r p i p e t t e and t r a n s f e r r e d t o 24 w e l l c u l t u r e
d i s h e s c o n t a i n i n g 1 ml o f DMEM s u p p le m e n t e d m edium , w i t h
t r i t u r a t i o n t o d i s p e r s e t h e c e l l s . The c e l l s w e r e a l l o w e d t o
e x p a n d t o s u b c o n f l u e n c y a t w h ich c u l t u r e medium w as rem oved and
- 155 -

t e s t e d f o r t h e p r e s e n c e o f AChE b i n d i n g a c t i v i t y by t h e s o l i d
p h a s e m ethod ( s e e b e l o w ) .
5 . 2 . 5 S c r e e n in g p r o t o c o l
H ybridoma medium w a s s c r e e n e d f o r t h e p r e s e n c e o f AChE
b i n d i n g a n t i b o d i e s by a s o l i d p h a s e im m u n oad sorb an ce a s s a y . 100
pi o f hybridom a medium w as i n c u b a t e d f o r a t l e a s t 3 h o u r s a t room
t e m p e r a t u r e in a 9 6 w e l l p o l y e t h y l e n e m i c r o t i t r e d i s h w h ic h had
b e e n p r e c o a t e d w i t h r a b b i t a n t im o u s e I g (3 0 p g /m l ) and
p r e - a d s o r b e d w i t h 0.5% BSA i n p h o s p h a t e b u f f e r e d s a l i n e (PBS:
150 mM N aC l, 5 0 irM p h o s p h a t e , pH 7 . 4 ) . The w e l l s w e r e th e n
w a sh ed 3 t i m e s w i t h 2 0 0 pi o f PBS and 100 pi o f an AChE s o l u t i o n
from an e x t r a c t o f o n e day o l d c h i c k b r a i n ( h o m o g e n is e d 1 : 1 0 0 i n
PBS + 0.5% T r i t o n X -1 0 0 ) was a d d ed and sh a k e n f o r a t l e a s t 4
h o u r s a t 4 ° C . The w e l l s w e r e w ash ed 3 t i m e s w i t h 2 0 0 pi o f PBS +
0.05% N P-40 and AChE i m m o b i l i z e d on t h e s u r f a c e was a s s a y e d
d i r e c t l y in t h e w e l l i n a vo lum e o f 1 0 0 p i . T y p i c a l l y t h e
n o n - s p e c i f i c b i n d i n g was l e s s th a n 2% and t h e s i g n a l t o n o i s e
r a t i o o f medium from p o s i t i v e c l o n e s w as a b o u t 7 0 : 1 . T h i s s o l i d
p h a s e a s s a y was a d d i t i o n a l l y u s e d f o r a f f i n i t y and c o m p e t i t i o n
m ea su r e m e n t w i t h m o d i f i c a t i o n a s d e s c r i b e d i n t h e R e s u l t s
S e c t i o n .
5 . 2 . 6 P u r i f i c a t i o n o f M o n o c lo n a l a n t i b o d i e s
A n t i b o d i e s w ere p u r i f i e d from h yb r id om a medium u s i n g a 1 . 5
ml p r o t e i n A - S e p h a r o s e CL4B c o lu m n . The medium w as r e c y c l e d
t h r o u g h t h e p r o t e i n A c o lu m n , e q u i l i b r a t e d w i t h 0.1M T r i s b u f f e r ,
- 156 -

pH 8 . 0 o v e r n i g h t a t 4 ° C . The colum n w a s t h e n w a sh ed e x t e n s i v e l y
w i t h t h e same b u f f e r , and e l u t e d w i t h 0.1M c i t r a t e b u f f e r a t
l o w e r pH a c c o r d i n g t o (Ey e t a l . , 1 9 7 8 ) . 0 . 9 ml f r a c t i o n s w ere
c o l l e c t e d and t h e p r o t e i n c o n t e n t i n e a c h f r a c t i o n was d e t e r m in e d
by a b s o r b a n c e a t 2 8 0 n m /260 nm. The f r a c t i o n s w h ich c o n t a i n e d
peak p r o t e i n c o n t e n t w e r e p o o l e d and d i a l y s e d a g a i n s t 10 nM T r i s ,
pH 8 . 0 and c o n c e n t r a t e d w i t h an Am icon u l t r a f i l t r a t i o n c e l l . The
p r o t e i n c o n c e n t r a t i o n and t h e p u r i t y o f t h e f i n a l im m u n o g lo b u l in
s o l u t i o n was d e t e r m in e d by t h e m ethod o f Lowry e t al^. ( 1 9 5 1 ) and
SDS PAGE ( s e e b e l o w ) , r e s p e c t i v e l y . The p r o t e i n A co lu m n was
c l e a n e d w i t h 10 0 co lu m n v o l o f 0 . 5 M N a C l/ 0.5% N P - 4 0 / 0 . 1 M
c i t r a t e b u f f e r , pH 3 . 0 , and s u b s e q u e n t l y e q u i l i b r a t e d w i t h 0 . 1 M
T r i s , pH 8 . 0 .
5 . 2 . 7 G el e l e c t r o p h o r e s i s a n d w e s t e r n b l o t t i n g
SDS-PAGE u s i n g 10% S D S - p o ly a c r y l am ide g e l s was c a r r i e d o u t
by t h e m eth od o f Laemmli ( 1 9 7 0 ) . S a m p le s w ere d e n a t u r e d w i t h 1%
SDS and 1% d i t h i o t h r e i t o l a t 96°C p r i o r t o e l e c t r o p h o r e s i s .
P h o s p h o r y l a s e b (M^ 9 7 , 0 0 0 ) , BSA (M^ 6 8 , 0 0 0 ) , c a t a l a s e (M^
6 0 , 0 0 0 ) and o v a lb u m in (M^ 4 5 , 0 0 0 ) w e r e u s e d a s p r o t e i n
s t a n d a r d s , P r o t e i n b a n d s w ere s t a i n e d w i t h C o o m a ss ie B l u e .
P r o t e i n s from t h e p o l y a c r y l a m i d e g e l w e r e e l e c t r o - b l o t t e d t o
n i t r o c e l l u l o s e p a p e r f o r 16 h o u r s by t h e m ethod o f Tow bin e t a l .
( 1 9 7 9 ) . The b l o t w as a d s o r b e d w i t h hybridom a medium a t room
t e m p e r a t u r e f o r 3 h o u r s , w ash ed and i n c u b a t e d w i t h 2 0 ml o f HRP
c o n j u g a t e d r a b b i t a n t i - m o u s e Ig ( d i l u t e d 1 : 3 0 0 ) w h ic h was
d e v e l o p e d u s i n g d i a m i n o b e n z i d i n e t e t r a h y d r o c h l o r i d e ( 0 . 2 5 m g / m l )
+ H2 0 2 (0 .05% v / v ) and N iS 0 4 ( 0 .3 % ) .
- 157 -

5 . 2 . 8 i H u n o h l s t o c h a r i s t r y
F r o z e n s e c t i o n s ( 1 0 urn) o r t e a s e d f i b r e s from a d u l t c h i c k e n
PLD w e r e i n c u b a t e d w i t h hybridom a c o n d i t i o n e d medium f o r 3 h o u r s ,
w a sh e d 3 t i m e s i n PBS and i n c u b a t e d w i t h e i t h e r FITC o r
p e r o x i d a s e c o n j u g a t e d r a b b i t a n t i - m o u s e Ig f o r 1 - 2 h o u r s a t room
t e m p e r a t u r e . P r e p a r a t i o n s w e r e t h e n r i n s e d 5 t i m e s i n PBS.
F I T C - l a b e l l e d s e c t i o n s w ere m ounted i n g l y c e r o l - P B S - j e l l y and
o b s e r v e d u n d er a f l u o r e s c e n c e m i c r o s c o p e . P e r o x i d a s e l a b e l l e d
m u s c le f i b r e s w e r e d e v e lo p e d a s d e s c r i b e d a b o v e and m ounted i n
Aquamount m o u n ta n t (BDH) and o b s e r v e d u n d e r a p h a s e c o n t r a s t
m i c r o s c o p e .
5 . 3 RESULTS
5 . 3 . 1 P u r i f i c a t i o n o f c h i c k b r a in AChE
A b ou t 90% o f t h e c h i c k b r a i n AChE e x t r a c t e d i n t h e h ig h
s a l t / T r i t o n b u f f e r e x i s t s a s an I I S form ( F ig 5 . 1 ) . T h e r e i s
a l s o a m in or f r a c t i o n o f 7S form an d 20S form . S i n c e t h e enzym e
i s a g l y c o p r o t e i n and e x h i b i t s b i n d i n g t o Con-A (R o tu n d o and
Fam brough, 1 9 7 9 ) , t h i s p r o p e r t y i s e x p l o i t e d i n t h e f i r s t s t e p o f
i t s p u r i f i c a t i o n , by a p p l y i n g t h e e x t r a c t t o c h r o m a to g r a p h y on a
Con-A A g a r o s e c o lu m n , a f t e r w h ich t h e bound p r o t e i n s a r e
d i s p l a c e d w i t h 0 . 1 M a - m e t h y lm a n n o s i d e . F o l l o w i n g d i a l y s i s , t h e
enzym e i s s u b j e c t e d t o i o n e x c h a n g e c h r o m a to g r a p h y on a DEAE-
S e p h a c e l colum n w h ic h i s n e c e s s a r y f o r t h e e f f i c i e n t b i n d i n g o f
t h e enzym e t o t h e N - m e t h y l a c r i d i n i u m - S e p h a r o s e (MAc) a f f i n i t y
c o lu m n . S i n c e t h e a f f i n i t y r e s i n e x h i b i t s i o n e x c h a n g e
p r o p e r t i e s ( M a s s o u l i e and B on , 1 9 7 6 ) , t h e i o n i c s t r e n g t h o f t h e
- 158 -

FRACTIONS
FIG. 5.1 S u c r o s e g r a d i e n t p r o f i l e o f AChE from c r u d e e x t r a c t o f
o n e day o l d c h i c k b r a i n . AChE was e x t r a c t e d w i t h 8 v o l o f
0 . 5 M N aC l/0 .5% T r i t o n X - 1 0 0 / 5 0 nfll p h o s p h a t e b u f f e r , pH 7 . 5 ,
a f t e r b r a i n t i s s u e had b e e n p r e - h o m o g e n i s e d and p e l l e t e d w i t h 10
v o l o f 0 . 4 M N a C l / 1 2 .5 mM p h o s p h a t e b u f f e r , pH 7 . 5 ( s e e t e x t ) .
B a c i t r a c i n ( lm g /m l ) and 5 nM EGTA w e r e i n c l u d e d a s p r o t e a s e
i n h i b i t o r s . Arrow i n d i c a t e s t h e p o s i t i o n o f t h e c a t a l a s e
m a rk er .
- 159 -

DEAE colum n e l u a t e m u st be r e d u c e d by d i a l y s i s p r i o r t o i t s
a p p l i c a t i o n t o t h e MAc c o lu m n . Under t h e s e c o n d i t i o n s , m ore th a n
95% o f t h e enzym e i s bound t o t h e a f f i n i t y r e s i n . N ear c o m p le t e
e l u t i o n i s a c h i e v e d by d i s p l a c i n g t h e enzym e w i t h d e c a m e th o n iu m .
U s in g t h i s p u r i f i c a t i o n p r o c e d u r e , an e n r ic h m e n t o f o v e r 2 , 0 0 0
f o l d o f AChE was o b t a i n e d ( T a b l e 5 . 1 ) .
The p u r i f i e d enzym e m i g r a t e s a s a m a jo r band on SDS g e l w i t h
a n a p p r o x im a t e o f 1 1 0 , 0 0 0 ( F i g . 5 . 2 b ) ( c . f . A p p e n d ix 2 ) .
The band i s c h a r a c t e r i s t i c a l l y d i f f u s e due m o s t l i k e l y t o t h e
e x t e n s i v e g l y c o s y l a t i o n o f t h e p r o t e i n . The p u r i f i e d enzym e i so
r e a d i l y l a b e l l e d w i t h [ H]DFP ( F i g . 5 . 2 c ) w h o se b i n d i n g i s
- 5b l o c k e d by p r e - t r e a t i n g and c o - i n c u b a t i n g w i t h 2 x 10 e s e r i n e
( F i g . 5 . 2 d ) .
5 . 3 . 2 C h a r a c t e r i s a t i o n o f M o n o c lo n a l a n t i b o d i e s t o AChE
A. S u b c l a s s i f i c a t i o n and p u r i f i c a t i o n
T h r e e h ybridom a s e c r e t i n g a n t i b o d i e s t h a t b in d c h i c k b r a i n
AChE w ere i s o l a t e d and c l o n e d . Two o f th em , d e s i g n a t e d ACB-1 and
ACB-3 a r e o f s u b c l a s s IgG£a w h e r e a s ACB-2 i s o f s u b c l a s s
IgG ^. ACB-3 b i n d s AChE from b o t h c h i c k b r a i n and m u s c l e , and
r e c o g n i s e s a 1 1 0 , 0 0 0 d p e p t i d e on n i t r o c e l l u l o s e b l o t . H ow ever ,
s i n c e i t i s s e c r e t e d from hybridom a c e l l s a t v e r y lo w q u a n t i t i e s
( < 0 .1 ug/ml o f c o n f l u e n t c e l l s ) , o n l y ACB-1 an d ACB-2 w e r e
f u r t h e r c h a r a c t e r i s e d . A f t e r b i n d i n g a n t i b o d i e s t o t h e p r o t e i n
A - S e p h a r o s e c o lu m n , ACB-1 was e l u t e d from t h e co lu m n a t pH 4 . 5 ,
and ACB-2 a t pH 6 . SDS-PAGE o f t h e p u r i f i e d a n t i b o d i e s showed
t h e c h a r a c t e r i s t i c l i g h t (Nlr 2 2 , 0 0 0 ) and h ea v y (Mr 5 5 , 0 0 0 )
c h a i n s ( F i g . 5 . 3 b ) (C o h e n , 1 9 6 7 ) .
- 160 -

T a b le 5 . 1 P u r i f i c a t i o n o f AChE from day o l d c h i c k b r a i n .
Sam p le V o l .
(m l)
T o t a l
AChE
( p m o l/m in )
P r o t e i n 3
( m g /m l )
S p e c i f i c
a c t i v i t y
(p m o l /m in /m g p r o t e i n )
P u r i f i c a t i o n
f a c t o r
AChE
r e c o v e r y
mc r u d e e x t r a c t 4 3 0 40 2 2 1 0 7 0 . 1 9 10 0
Con A e l u a t e 3 9 5 165 130 1 . 2 7 6 7 41
DEAE e l u a t e 61 91 32 2 . 8 4 1 4 . 9 23
MAc c o n c e n t r a t e 0 . 3 7 1 . 2 0 . 1 6 44 5 2 3 4 2 18
a P r o t e i n c o n t e n t o f s a m p le s w as d e t e r m in e d by t h e m eth od o f Lowry e t a l . ( 1 9 5 1 ) , u s i n g BSA a s
p r o t e i n s t a n d a r d .

a b e d
FIG- 5.2 Analysis of purified chick brain AChE using SDS-PAGE.
a, chick brain crude extract; b, AChE preparation stained with
Coomassie blue; c, [3H]DFP (10_4M DFP) labelling of purified
AChE; d, [3H]DFP C10-4M DFP) labelling of purified AChE
blocked by pre-treatment with 2 x 10”t I eserine. Numbers on
the right indicate the positions and the size (xlOOO) of the high
molecular weight protein markers.
-162-

ab
68k-
A6K~*
FIG. 5.3 SDS-PAGE of purified ACB-1 and ACB-2. a, protein
markers; b, purified ACB-2. Purified ACB-1 showed identical
banding pattern.
- 1 6 3 -

B . A f f i n i t y o f m o n o c l o n a l a n t i b o d i e s t o c h i c k b r a i n AChE
The b i n d i n g a f f i n i t y o f ACB-1 a nd ACB-2 t o c h i c k b r a i n AChE
w e r e t e s t e d i n v a r y i n g d i l u t i o n s o f p u r i f i e d a n t i b o d i e s ( F i g .
5 . 4 ) . A ss u m in g t h a t b i n d i n g i n t h e s o l i d p h a s e a s s a y f o l l o w s
m a s s - a c t i o n l a w s ; w i t h o n e e p i t o p e p e r enzyme monomer and two
r e c o g n i t i o n s i t e s p e r IgG m o l e c u l e , an a p p a r e n t d i s s o c i a t i o n
c o n s t a n t f o r t h e a n t i g e n - I g G c o m p l e x c o u l d be e v a l u a t e d . S i n c e
t h e c o n c e n t r a t i o n o f a n t i g e n was l o w ( 0 . 0 2 nM), b i n d i n g a s s a y s
o b e y s t h e e q u a t i o n
b = 2 [ A b ]
2 [ A b ] + Kd
w h e r e b i s t h e f r a c t i o n o f e p i t o p e bo un d, [A b ] i s t h e m o l a r
c o n c e n t r a t i o n o f a n t i b o d y , and i s t h e d i s s o c i a t i o n c o n s t a n t .
F u r t h e r m o r e , a s i t i s l i k e l y t h a t b i n d i n g o f a m u l t i - s u b u n i t AChE
m o l e c u l e t o an IgG m o l e c u l e r e q u i r e s a minimum o f 1 e p i t o p e i n
t h e s o l i d p h a s e a s s a y , b i n d i n g o f AChE c o u l d t h e n be e x p r e s s e d by
c o r r e c t i n g b f o r t h e number o f e p i t o p e s ( n ) p e r m o l e c u l e o f AChE,
h e n c e :
B = 1 - ( l - b ) n
w h e r e B i s t h e f r a c t i o n o f AChE bo un d, and n = 4 f o r t h e
t e t r a m e r i c c h i c k e n b r a i n form o f AChE. The a p p a r e n t
c a l c u l a t e d from 50% b i n d i n g f o r IgG i s 0 . 8 5 rM f o r ACB-1 a n d 0 . 7 7
nM f o r ACB-2.
C. M o n o c lo n a l a n t i b o d i e s r e c o g n i s e s e p a r a t e a n t i g e n i c s i t e s
I t was f o u n d t h a t t h e b i n d i n g o f s a t u r a t i n g am oun ts o f ACB-2
t o AChE d i d n o t i n h i b i t t h e s u b s e q u e n t b i n d i n g o f ACB-1 t o AChE.
ACB-2-AChE c o m p l e x b i n d s t o i m m o b i l i s e d ACB-1 t h r o u g h o u t a r a n g e
- 164 -

FIG- 5-4 D i l u t i o n c u r v e s o f ACB-1 and ACB-2. 0 . 2 m i l l i u n i t s o f
AChE from on e day o l d c h i c k b r a i n was i n c u b a t e d w i t h a
number o f s e r i a l d i l u t i o n s o f e i t h e r ACB-1 o r ACB-2 i n a t o t a l
v o lu m e o f 100 ul f o r a minimum o f 4 h o u r s a t 4 °C . The s o l u t i o n s
w e r e t h e n t r a n s f e r r e d t o t h e 9 6 w e l l p o l y e t h y l e n e m i c r o - t i t r e
d i s h and i n c u b a t e d o v e r n i g h t a t 4 ° C . The w e l l s w e r e t h e n w a sh ed
3 t i m e s w i t h 2 0 0 ill o f PBS + 0.05% N P -4 0 . I m m o b i l i s e d
e n z y m e - a n t i b o d y c o m p le x was m e a s u r e d by t h e r a d i o m e t r i c a s s a y .
The am ou nt o f enzyme bound i s e x p r e s s e d a s a p e r c e n t a g e o f enzyme
a d d e d . The c u r v e s w ere f i t t e d by e y e and c a l c u l a t i o n s o f
a p p a r e n t d i s s o c i a t i o n c o n s t a n t s f o r t h e a n t i b o d i e s ( s e e t e x t )
w e r e p e r f o r m e d u s i n g 50% b i n d i n g v a l u e .
- 165 -

o f c o n c e n t r a t i o n s ( F i g . 5 . 5 ) . B i n d i n g o f t h i s c o m p le x i s s i m i l a r
t o t h e b i n d i n g o f e q u i v a l e n t am oun ts o f AChE s u g g e s t i n g t h a t t h e
two a n t i b o d i e s r e c o g n i s e d i f f e r e n t e p i t o p e s . F u r t h e r m o r e , ACB-1
r e c o g n i s e s S D S - d e n a t u r e d AChE i n n i t r o c e l l u l o s e b l o t s ( s e e F i g .
5 . 6 a ) , w h i l e ACB-2 o n l y r e c o g n i s e s t h e n a t i v e form o f t h e
e n z y m e .
D. C r o s s - r e a c t i v i t y w i t h AChE from o t h e r t i s s u e s and s p e c i e s
ACB-1 was t e s t e d by t h e s o l i d p h a s e a s s a y a s w e l l a s
b l o t t i n g m et h od f o r i t s b i n d i n g t o AChE from o t h e r t i s s u e s and
s p e c i e s . T h i s a n t i b o d y r e c o g n i s e s AChE from c h i c k b r a i n , p l asm a
and m u s c l e a s w e l l a s AChE from q u a i l b r a i n , b u t d o e s n o t
r e c o g n i s e AChE from any o t h e r a n i m a l s t e s t e d ( T a b l e 5 . 2 ) . ACB-2
w as t e s t e d o n l y by t h e s o l i d p h a s e a s s a y s i n c e i t d o e s n o t b i n d
t o S D S - d e n a t u r e d AChE. The r e s u l t show s t h a t t h i s a n t ib o c t y h a s
s i m i l a r r e c o g n i t i o n f o r c h i c k AChE, b u t d o e s n o t r e c o g n i s e q u a i l
b r a i n AChE, o r AChE from o t h e r s p e c i e s ( T a b l e 5 . 2 ) . N e i t h e r
a n t i b o d y b i n d s t o V>ChE from c h i c k p las ma o r b r a i n . A l l t h e AChE
fo rm s i n c h i c k m u s c l e a r e r e c o g n i s e d by b o t h ACB-1 a nd ACB-2.
ACB-1 r e c o g n i s e s a p o l y p e p t i d e (Mr 1 1 0 , 0 0 0 ) from p u r i f i e d
c h i c k b r a i n AChE ( F i g . 5 . 6 a ) , from b r a i n and m u s c l e c r u d e
e x t r a c t s o f c h i c k ( F i g . 5 . 6 b and c ) and q u a i l ( F i g . 5 . 6 d and e) ,
b u t d o e s n o t b i n d d e t e c t a b l y t o any p e p t i d e i n r a b b i t , r a t o r
g u i n e a p i g b r a i n , T o r p e d o o r E l e c t r o p h o r u s e l e c t r i c o r g a n
e x t r a c t s .
E. I m m u n o h is t o c h e m i c a l a p p l i c a t i o n o f ACB-1 and ACB-2
Bo th ACB-1 and ACB-2 a r e h i g h l y c o n c e n t r a t e d i n t h e e n d p l a t e
- 1 6 6 -

ANTIBODY IN SOLID PHASE (ng)
FIG- 5-5 C o m p e t i t i o n o f ACB-1 and ACB-2 f o r e p i t o p e s o f AChE.
AChE was f i r s t e x p o s e d t o a s a t u r a t i n g c o n c e n t r a t i o n o f ACB-2 and
t h e n AChE-ACB-2 c o m p le x e x p o s e d t o ACB-1 a t t a c h e d t o s o l i d p h a s e .
The d i l u t i o n c u r v e o f AChE-ACB-2 ( # ) c l o s e l y r e s e m b l e s t h e
c o n t r o l c u r v e ( O ) i n w h ic h AChE i s n o t i n i t i a l l y c o m p l e x e d
w i t h ACB-2. The c u r v e i s f i t t e d by e y e . P e r c e n t AChE b i n d i n g i s
e x p r e s s e d a s i n F i g . 5 . 4 .
- 167 -
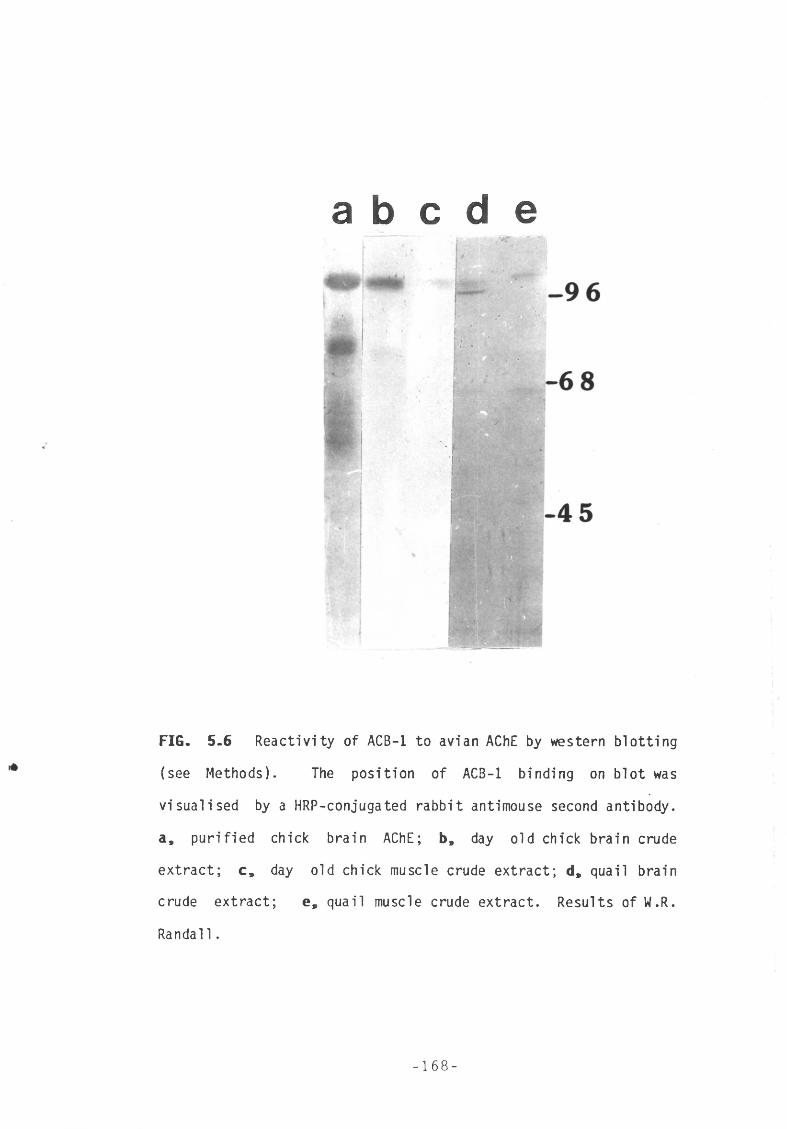
a b c d e
FIG- 5-6 Reactivity of ACB-1 to avian AChE by western blotting
(see Methods). The position of ACB-1 binding on blot was
visualised by a HRP-conjugated rabbit antimouse second antibody,
a, purified chick brain AChE; b, day old chick brain crude
extract; c, day old chick muscle crude extract; d, quail brain
crude extract; e, quail muscle crude extract. Results of W.R.
Randall.
- 1 6 8 -

TABLE 5 . 2 R e a c t i v i t y ® o f ACB-1 and ACB-2 w i t h (pChE and AChE
from o t h e r s p e c i e s .
AChE S o u r c e 15 ACB-1 ACB-2
C h i c k e n b r a i n + +
m u s c l e + +
Q u a i l b r a i n + -
m u s c l e + -
T o r p e d o e l e c t r i c o r g a n - -
E l e c t r o p h o r u s e l e c t r i c o r g a n - -
R a b b i t b r a i n - -
R a t b r a i n - -
G u in ea p i g b r a i n - -
C h ic k e n <PChE ( b r a i n ) - -
( p l a s m a ) - -
a B i n d i n g o f AChE and ^ChE was m e a s u r e d by t h e s o l i d p h a s e
a s s a y .
k AChE a c t i v i t y was m e a s u r e d i n t h e p r e s e n c e o f 1 X 10"^M
isoOMPA f o r a l l b u t t h e l a s t two s a m p l e s . <pChE was m e a s u r e d i n
t h e p r e s e n c e o f 5 X 10"^M BW284C51 f o r t h e s e s a m p l e s .
- 1 6 9 -

r e g i o n s o f a d u l t PLD m u s c l e , a s HRP o r FITC c o n j u g a t e d r a b b i t
a n t i - m o u s e Ig l a b e l l i n g c l e a r l y sh ow s on t e a s e d f i b r e s ( F i g .
5 . 7 A ) o r f r o z e n t r a n s v e r s e s e c t i o n s ( F i g . 5 . 7 B ) t r e a t e d w i t h
e i t h e r o f t h e a n t i b o d i e s . The d e p o s i t i o n o f a n t i b o d y l a b e l l i n g
r e v e a l s s t r u c t u r e s c h a r a c t e r i s t i c o f t h e en p l a q u e m o to r
e n d p l a t e s a s o b s e r v e d by t h e c o n v e n t i o n a l h i s t o c h e m i c a l s t a i n i n g
f o r e n d p l a t e AChE ( H e s s , 1 9 6 1 ) .
5 . 4 DISCUSSION
AChE from c h i c k b r a i n h a s b een p u r i f i e d , u s i n g a c r i d i n i u m
a f f i n i t y c h r o m a t o g r a p h y , by a b o u t 2 , 0 0 0 f o l d . The e f f i c i e n c y o f
t h i s p r o c e d u r e i s g r e a t l y im p r o v e d by c h r o m a t o g r a p h y on t h e
DEAE-Sephacel co lu mn p r i o r t o a f f i n i t y c h r o m a t o g r a p h y s u g g e s t i n g
t h a t t h e l o w r e t e n t i o n and r e c o v e r y o f t h e c h i c k b r a i n enzyme on
t h e MAc colum n r e p o r t e d by V a l l e t t e e t a l . ( 1 9 8 3 ) may be due t o
i n t e r f e r e n c e by c o n t a m i n a t i n g p r o t e i n s i n t h e e x t r a c t .
I d e n t i f i c a t i o n o f t h e p u r i f i e d p r o t e i n a s AChE i s b a s e d on i t s
s u b u n i t m o l e c u l a r s i z e , w h ich a g r e e s w e l l w i t h t h a t p r e v i o u s l y
o b s e r v e d ( A p p e n d ix 2 ) ( M a s s o u l i e and Bon , 1 9 8 2 ; R o t u n d o , 1 9 8 4 ) ,
i t s s p e c i f i c i n h i b i t i o n by BW284C51 ( R a n d a l l e t a l . , 1 9 8 5 ) ,
[ H]DFP b i n d i n g w h ic h i s i n h i b i t e d by e s e r i n e , and
p r e c i p i t a t i o n o f AChE, b u t n o t ^ChE, by m o n o c lo n a l a n t i b o d i e s
d i r e c t e d a g a i n s t t h e p u r i f i e d p r o t e i n . I t i s c o n c l u d e d
t h e r e f o r e , t h a t t h e p u r i f i e d p r o t e i n i s AChE. T h a t t h e
a n t i b o d i e s r e c o g n i s e d e g l y c o s y l a t e d AChE (M^ 1 0 0 , 0 0 0 ) show s
t h a t t h e a n t i g e n i c d e t e r m i n a n t s a r e a s s o c i a t e d w i t h t h e
p o l y p e p t i d e and n o t t h e c a r b o h y d r a t e m o i e t y ( R a n d a l l e t a l
- 170 -
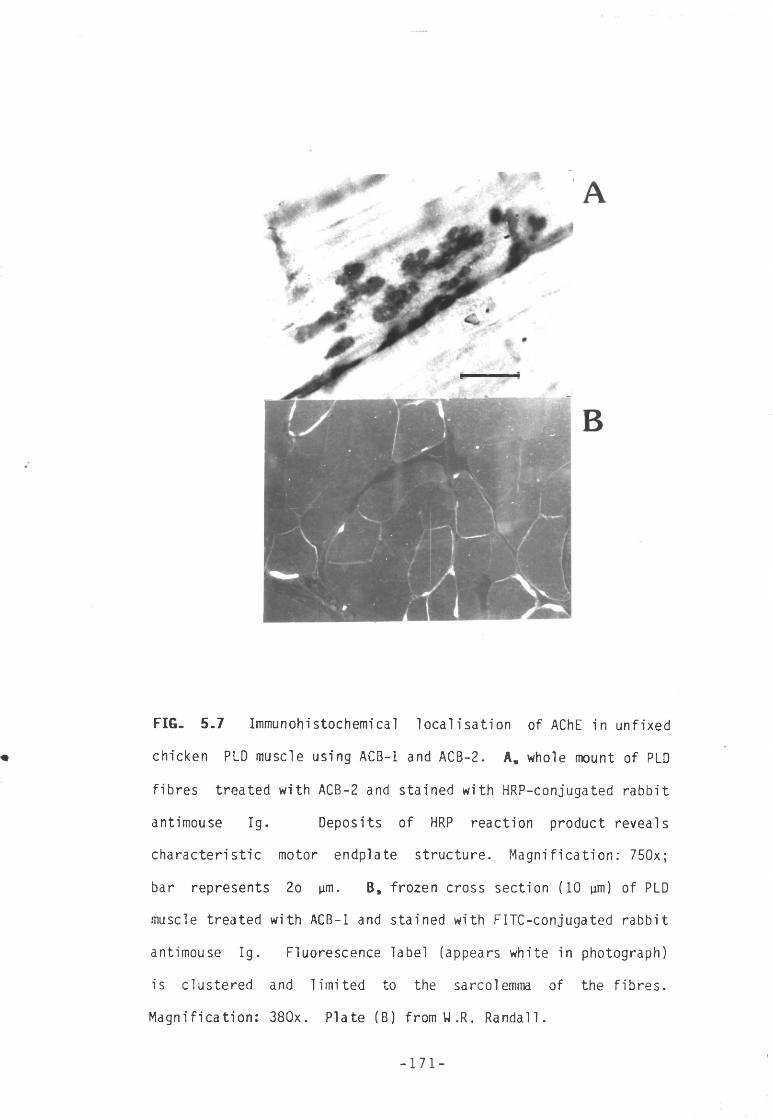
FIG- 5.7 Immunohistochemical localisation of AChE in unfixed
chicken PLD muscle using ACB-1 and ACB-2. A, whole mount of PLD
fibres treated with ACB-2 and stained with HRP-conjugated rabbit
antimouse Ig. Deposits of HRP reaction product reveals
characteristic motor endplate structure. Magnification: 750x;
bar represents 2o urn. B, frozen cross section (10 urn) of PLD
muscle treated with ACB-1 and stained with FITC-conjugated rabbit
antimouse Ig. Fluorescence label (appears white in photograph)
is clustered and limited to the sarcolemma of the fibres.
Magnification: 380x. Plate (B) fromW.R. Randall.
- 171 -

1 9 8 5 ) . The a n t i b o d i e s show h i g h a f f i n i t y f o r t h e a v i a n form o f
AChE. T h e r e i s no c r o s s - r e a c t i v i t y w i t h $ChE o r AChE from
mammals o r f i s h . T h i s l a c k o f c r o s s - r e a c t i v i t y h a s b e e n
s i m i l a r l y o b s e r v e d i n m o n o c lo n a l a n t i b o d i e s r a i s e d a g a i n s t human
e r y t h r o c y t e AChE (Fambrough e t a l . , 1 9 8 2 ) , w h er e t h e a n t i b o d i e s d i d
n o t r e c o g n i s e a v i a n AChE. T h i s s u g g e s t s l i m i t e d ho m o lo g y b e t w e e n
t h e a n t i g e n i c r e g i o n s o f a v i a n and mammalian o r f i s h AChE. The
f a c t t h a t ACB-2 o n l y b i n d s t o t h e n a t i v e form o f t h e enzyme
s u g g e s t s t h a t i t r e c o g n i s e s a c o n f o r m a t i o n a l d e p e n d e n t a n t i g e n i c
s i t e . S i n c e t h e y b i n d a l l t h e fo rm s e q u a l l y w e l l , t h e y m u s t
r e c o g n i s e a common e p i t o p e on t h e c a t a l y t i c s u b u n i t s . I t d o e s
n o t , h o w e v e r , p r e c l u d e t h e p o s s i b l e d i f f e r e n c e s i n t h e m o l e c u l a r
s t r u c t u r e o f t h e c a t a l y t i c s u b u n i t s b e t w e e n t h e v a r i o u s m o l e c u l a r
f o r m s , a s p r e v i o u s s t u d i e s have s u g g e s t e d ( D o c t o r e t a l ^ . , 1 9 8 3 ;
S o r e n s e n e t a l . , 1 9 8 2 ) .
Two p o l y p e p t i d e c h a i n s , t er m e d a and B, w e r e r e c o g n i s e d by
SDS-PAGE from p u r i f i e d c h i c k e n b r a i n AChE ( R o t u n d o , 1 9 8 4 ) . T h i s
s u b u n i t p a t t e r n w a s , h o w e v e r , n o t o b s e r v e d i n o u r p u r i f i e d
p r e p a r a t i o n o f c h i c k b r a i n AChE ( F i g . 5 . 6 a ) . The d i f f e r e n c e i n
t h i s s u b u n i t b a n d in g p a t t e r n may be a t t r i b u t e d t o t h e g e n o t y p e o f
t h e c h i c k e n s t r a i n s u s e d i n t h e two s t u d i e s ( R a n d a l l e t a l . ,
1 9 8 5 ) . T h i s i s e v i d e n t from t h e s t u d y o f a n o t h e r l i n e o f
c h i c k e n , l i n e 4 1 2 ( D a v i s , C a l i f o r n i a ) , w h er e i n d i v i d u a l s e x h i b i t
v a r y i n g b a n d i n g p a t t e r n s o f AChE. Some c o n t a i n o n l y t h e a c h a i n ,
some c o n t a i n o n l y t h e B c h a i n and some c o n t a i n b o t h . T h i s
v a r i a t i o n i s a l s o i n d e p e n d e n t o f t h e a g e o f t h e b i r d o r t h e
t i s s u e s o u r c e o f t h e e n z y m e . I t i s l i k e l y , t h e r e f o r e , t h a t t h e
h e t e r o g e n e i t y i n t h e s u b u n i t s i z e o b s e r v e d i n b o t h c a s e s i s due
t o t h e a l l e l i c d i f f e r e n c e o f t h e AChE g e n e i n t h e s e s t r a i n s o f
b i r d s .
- 1 7 2 -

The p u r i f i c a t i o n o f t h e AChE form m u s c l e u s i n g t h e s e
m o n o c l o n a l a n t i b o d i e s a s i m m u n o a f f i n i t y l i g a n d s i s c u r r e n t l y
b e i n g u n d e r t a k e n a t t h i s l a b o r a t o r y . I m m o b i l i s d ACB-1 a l s o b i n d s
t h e enzyme w i t h h i g h a f f i n i t y , h o w e v e r , t h e r e c o v e r y o f t h e
enzyme i s much l o w e r t h a n t h a t w i t h ACB-2. H e n c e , ACB-2 w h i c h
r e c o g n i s e s a c o n f o r m a t i o n a l d e p e n d e n t e p i t o p e i s s e l e c t e d f o r t h e
i m m u n o a f f i n i t y p u r i f i c a t i o n o f t h e e n z y m e , w h i l e ACB-1 i s
s u i t a b l e f o r a n a l y t i c a l s t u d i e s o f t h e p u r i f i e d e n z y m e .
I m m u n o a f f i n i t y c h r o m a t o g r a p h y u s i n g i m m o b i l i s e d ACB-2 on
S e p h a r o s e co lum n b i n d s AChE from one day o l d c h i c k m u s c l e
e x t r a c t s u n d e r h i g h i o n i c s t r e n g t h c o n d i t i o n s ( 0 . 5 M N aC l /0 .5%
T r i t o n X - 1 0 0 / 1 0 nM HEPES), w h ic h i s n e c e s s a r y f o r t h e
s o l u b i l i s a t i o n o f t h e a s y m m e t r i c f o r m s . T h e s e c o n d i t i o n s a l s o
a l l o w t h e e l i m i n a t i o n o f t h e l a r g e amount o f c o n t a m i n a n t s , i n
p a r t i c u l a r t h e m y o f i b r i l l a r p r o t e i n s , w h ic h c a n n o t be remove d by
c o n v e n t i o n a l p r o t e i n p u r i f i c a t i o n t e c h n i q u e s ( u n p u b l i s h e d d a t a ) .
The bound AChE i s a l s o e a s i l y e l u t e d w i t h h i g h pH b u f f e r (pH 11)
w i t h a b o u t 80% r e c o v e r y .
In summary, c h i c k b r a i n AChE h a s b e e n e n r i c h e d by o v e r 2 , 0 0 0
f o l d u s i n g a f f i n i t y c h r o m a t o g r a p h y . T h re e m o n o c lo n a l a n t i b o d i e s
w h i c h r e c o g n i s e t h i s enzyme ha v e b e e n o b t a i n e d and two o f th em ,
ACB-1 and ACB-2, h a v e b e e n c h a r a c t e r i s e d . Both a n t i b o d i e s
e x h i b i t s i m i l a r b i n d i n g a f f i n i t y t o t h e enzyme and r e c o g n i s e
s e p a r a t e a n t i g e n i c s i t e s on t h e p o l y p e p t i d e b a c k b o n e . T h e s e
a n t i b o d i e s do n o t c r o s s - r e a c t w i t h c h i c k $C hE ,m am m al ian o r f i s h
AChE, b u t ACB-1 d o e s r e c o g n i s e q u a i l AChE. T h e s e a n t i b o d i e s b i n d
a t h i g h c o n c e n t r a t i o n t o t h e e n d p l a t e s o f a d u l t c h i c k e n m u s c l e .
I m m u n o a f f i n i t y p u r i f i c a t i o n o f t h e AChE u s i n g t h e s e
a n t i b o d i e s i s now i n p r o g r e s s .
- 173 -

C h a p t e r 6
GENERAL DISCUSSION
- 1 7 4 -

AChE a t t h e n e u r o m u s c u l a r j u n c t i o n : m o l e c u l a r f o r m s and r e g u l a t i o n
It is widely thought that the distribution of AChE forms has
important functional significance. The present study demonstrates
unequivocally that the heavy AChE predominates at the neuromuscular
junction of vertebrate twitch muscle where very little L and M forms are
present. It was found that the technique of isolating the neuromuscular
endplates and the precautionary measures employed to protect the heavy
AChE forms from proteolytic degradation are both crucial factors in the
identification of heavy AChE as the endplate specific forms. Although
H2 C is with no exception the predominant form in all endplates, in some
muscles a significant H^c component is also found. The evidence is as
yet insufficient to determine whether this H^c component is a stable
form at the endplate, or a degradatory product of H2 C . The latter
possibility cannot be ruled out from the present result since some lower
molecular weight forms are always seen in the endplate despite the use
of protease inhibitors. Further studies are therefore required to
establish the nature of this component and whether it has a role in the
neuromuscular endplates.
Reduction of the amount of endplate AChE upon chronic
denervation may be brought about by 3 ways: (a) an impaired synthesis of
the collagen-tail, (b) an increased degradation of the enzyme, and (c) a
reduced capacity for the attachment of these forms at the endplate. (a)
appears unlikely, at least in denervated slow-twitch muscles where the
content of the asymmetric forms actually increases, (b) has been shown
in rat to be involved in the reduction of endplate AChE following
denervation, as a result of increased turnover rate. In denervated type
I fibres, L forms become predominant at the endplate, which may be
readily contributed by the increased degradation of the H forms.
However, this process may not be the only cause of these changes in
- 175 -

denervated muscles which over-produce AChE. On the other hand, there is
a good deal of evidence that AChE is associated with the basal lamina
where AChE clustering occurs and which is known to have a highly
specific organization at the synapse. An impairment of this specialized
structure in disease (Inestrosa, 1985) or absence of nerve may reduce
the attachment of the H AChE to the endplate region.
Neuromuscular contact is important in maintaining the normal
concentration of H AChE at the endplate. However, the enzyme does not
rapidly disappear upon denervation and the clustering of these forms is
largely maintained. Such persistence of endplate AChE may be attributed
to the increase in AChE content in denervated rabbit and guinea pig
muscles. In rat muscles where AChE content is drastically reduced upon
denervation, the level of endplate AChE disappears more rapidly. The
persistence of AChE at denervated endplates therefore appears to depend
largely on the capacity of the denervated muscle in producing H AChE.
The fact that avian tonic fibres contain a majority of L forms
at the endplate, while twitch fibres contain a majority of H forms
suggests a fundamental difference in the organisation of endplate AChE
in the two fibre types. Furthermore, since embryonic and neonatal tonic
fibres are capable of synthesizing significant amount of H AChE, the
organization of endplate AChE is therefore a highly selective process
which evolves according to the function of the muscle. The failure to
obtain cross-reinnervation between type II and tonic fibres in mature
chickens (Hnik et al., 1967), suggests that the endplate forms of AChE
in mature muscles are not inducible, and that the quaternary association
of the AChE catalytic units within the synapse may be specific to the
type of neural transmission of the muscle (Barnard et al., 1984b). In
this respect, the distribution of AChE forms clearly correlates with
the fibre type composition/function of a muscle. To understand the
- 176 -

p r e c i s e n a t u r e o f t h e c o n t r o l m ec h a n ism r e q u i r e s s t u d i e s a t t h e
m o l e c u l a r l e v e l , w h ic h w i l l l e a d n o t o n l y t o t h e u n d e r s t a n d i n g o f t h e
r e g u l a t o r y m ec h a n is m u n d e r l y i n g t h e p o ly m o r p h is m o f AChE, b u t may a l s o
p r o v i d e a u s e f u l model f o r t h e s t u d y o f t h e c o n t r o l o f g e n e e x p r e s s i o n
i n m u s c l e s .
Extrajunctional AChE forms: function and regulation
C o n t r a r y t o t h e e n d p l a t e AChE f o r m s w h ic h e x h i b i t an a p p a r e n t
a s s o c i a t i o n w i t h t h e f u n c t i o n a l a c t i v i t y o f t h e m u s c l e , t h e d i s t r i b u t i o n
o f e x t r a j u n c t i o n a l AChE i s much more v a r i a b l e . The l o w e r m o l e c u l a r
w e i g h t fo r m s a r e g e n e r a l l y c o n f i n e d t o t h e e x t r a j u n c t i o n a l r e g i o n i n
t w i t c h m u s c l e s ; t h e a c t i v i t y t h e s e f o rm s c o n t r i b u t e s t o t h e t o t a l AChE
i s sm a l l i n t y p e I m u s c l e s , b u t v a r i e s c o n s i d e r a b l y i n t y p e I I m u s c l e s
o f d i f f e r e n t s p e c i e s . The m o s t s t r i k i n g d i f f e r e n c e was s e e n b e t w e e n t h e
h a m s t e r and r a t f a s t m u s c l e s , w h e r e t h e L and M c o m p o n e n t s a r e s m a l l i n
t h e f o r m e r , b u t a r e d o m in a n t i n t h e l a t t e r . The p h y s i o l o g i c a l
s i g n i f i c a n c e o f t h e s e e x t r a j u n c t i o n a l f o rm s i s n o t w e l l e s t a b l i s h e d . The
e v i d e n c e o f d i s t i n c t m e t a b o l i c p o o l s o f AChE i n c u l t u r e d m u s c l e c e l l s
and t r a n s f o r m e d n e r v e c e l l l i n e r a i s e s s e v e r a l i m p o r t a n t q u e s t i o n s : ( 1 )
Do t h e s e m e t a b o l i c p o o l s o f AChE a l s o e x i s t i n m a t u r e m u s c l e s i n v i v o ?
( 2 ) What i s t h e i r r e l a t i o n s h i p w i t h t h e f u n c t i o n a l a s y m m e t r i c f o rm s a t
t h e e n d p l a t e ? And ( 3 ) What i s t h e p h y s i o l o g i c a l s i g n i f i c a n c e o f t h e s e
m e t a b o l i c p o o l s . D i f f e r e n t am oun ts o f e x t r a j u n c t i o n a l AChE i n d i f f e r e n t
m u s c l e s may i n d i c a t e v a r i a t i o n s i n t h e m e t a b o l i c r e g u l a t i o n o f t h e
enzyme i n d i f f e r e n t s p e c i e s . I t i s g e n e r a l l y a g r e e d t h a t a c t i v e ,
f u n c t i o n a l AChE f o r m s a r e r e l a t i v e l y s t a b l e , w h i l e p r e c u r s o r f o rm s
e x h i b i t more r a p i d t u r n o v e r r a t e . In v i v o s t u d i e s on t h e t u r n o v e r o f
t h e s e v a r i o u s f o r m s s h o u l d p r o v i d e k n o w l e d g e on t h e s i z e o f t h e p o o l
d e s t i n e d f o r t h e s y n t h e s i s o f t h e e n d p l a t e f o r m s .
- 177 -

Heavy form o f AChE a r e f o u n d e x t r a j u n c t i o n a l l y i n t y p e I m u s c l e s
and t h e y a r e t h e r e f o r e n o t e n d p l a t e s p e c i f i c . The p h y s i o l o g i c a l
s i g n i f i c a n c e o f t h e s e a s y m m e t r i c fo rm s i s n o t c l e a r . H ow e v e r , b e c a u s e
t h e y a r e f o u n d g e n e r a l l y i n m u s c l e s w h ic h e x h i b i t a c t i v e c o n t r a c t i l e
a c t i v i t y , i t may be s p e c u l a t e d t h a t a r e l a t i o n s h i p e x i s t s b e t w e e n t h i s
e x t r a j u n c t i o n a l p o p u l a t i o n and t h e p h y s i o l o g i c a l s t a t e o f t h e m u s c l e . In
a c c o r d a n c e w i t h t h i s e v i d e n c e , an e x t r a j u n c t i o n a l p o p u l a t i o n o f AChE
r e c e p t o r ca n a l s o be f o u n d i n t h e t y p e I m u s c l e f i b r e s . T h e s e
e x t r a j u n c t i o n a l ACh r e c e p t o r s and e s t e r a s e enzyme c o u l d p o s s i b l y b e : ( a )
a s u r p l u s p o o l o f p r o t e i n s i n d u c e d by m u s c l e a c t i v i t y f o r r e p l e n i s h i n g
t h e j u n c t i o n a l co m p o n e n t when r e q u i r e d ; o r ( b ) a d i s c r e t e p o p u l a t i o n
w i t h a s p e c i f i c f u n c t i o n a l l o c a l i s a t i o n . The i n t e r e s t i n g q u e s t i o n a s t o
w h ic h o f t h e s e p o s s i b i l i t i e s a r e c o r r e c t r e m a i n s t o be s e t t l e d .
U 1 t r a s t r u c t u r a l l o c a l i s a t i o n u s i n g m o n o c l o n a l a n t i b o d i e s
s p e c i f i c f o r a s y m m e t r i c AChE i s p r o b a b l y t h e m o s t s e l e c t i v e p r o b e f o r
t h i s t a s k . I m m u n o c y t o c h e m i s t r y u s i n g t h e s e m o n o c l o n a l a n t i b o d i e s a l s o
o f f e r s a p o w e r f u l t e c h n i q u e f o r t h e d i r e c t d e m o n s t r a t i o n o f t h e
a s s o c i a t i o n o f h e a v y AChE w i t h t h e b a s e m e n t membrane c o m p o n e n t s o f t h e
s y n a p s e . M o n o c lo n a l a n t i b o d i e s r a i s e d a g a i n s t t h e b r a i n form o f c h i c k
AChE i n t h i s s t u d y i s b e i n g c u r r e n t l y a p p l i e d f o r t h e e n r i c h m e n t o f t h e
a s y m m e t r i c form i n m u s c l e w h ic h i n t u r n ca n be u s e d f o r t h e p r o d u c t i o n
o f m o n o c lo n a l a n t i b o d i e s s e l e c t i v e f o r t h i s fo rm .
The e n h a n c e m e n t o f t h e amount o f e x t r a j u n c t i o n a l H f o r m s i n
normal PLD and t h e r e d u c t i o n o f e x c e s s L f o r m s i n d y s t r o p h i c PLD by
c h r o n i c s t i m u l a t i o n s u g g e s t s t h a t m u s c l e a c t i v i t y i s i m p o r t a n t i n t h e
r e g u l a t i o n o f e x t r a j u n c t i o n a l AChE. The e l u c i d a t i o n o f t h e a s s o c i a t i o n
o f t h i s c o m p o n e n t w i t h t h e e x t r a j u n c t i o n a l ACh r e c e p t o r , and i t s
m e t a b o l i c t u r n o v e r c o m p a r i n g w i t h t h e j u n c t i o n a l AChE p o o l s h o u l d s h e d
f u r t h e r l i g h t on t h e f u n c t i o n o f t h i s e x t r a j u n c t i o n a l H c o m p o n e n t and
- 178 -

i t s r e l a t i o n s h i p w i t h t h e j u n c t i o n a l AChE. How m u s c l e a c t i v i t y r e g u l a t e s
t h e m e t a b o l i s m o f AChE r e m a i n s a p r im e i n t e r e s t f o r f u r t h e r r e s e a r c h .
#
- 179 -

APPENDICES
- 180 -

A p p e n d ix 1 . R a p id p r o c e s s i n g o f s k e l e t a l M u s c le c r u d e e x t r a c t s .
To show t h a t AChE c a n b e e x t r a c t e d e f f i c i e n t l y from m u s c l e
t i s s u e s by r a p i d p r o c e s s i n g , t h e s o l e u s and EDL m u s c l e s w e r e
d i s s e c t e d from t h e h i n d l i m b s o f t h e r a t , w e i g h e d and h o m o g e n i s e d
i n 10 v o l o f t h e u s u a l h i g h s a l t / T r i t o n b u f f e r ( s e e s e c t i o n
2 . 2 . 4 ) . A s m a l l a l i q u o t from e a c h h o m o g e n a te was k e p t f o r
s u b s e q u e n t a s s a y . The r e s t was c e n t r i f u g e d a t 4 3 , 5 0 0 x g f o r 15
min and t h e s u p e r n a t a n t s w e r e k e p t . The p e l l e t s w e r e
r e h o m o g e n i z e d i n t h e same b u f f e r t o g i v e t h e same f i n a l d i l u t i o n
a s t h e f i r s t h o m o g e n i s a t i o n , and t h e s u s p e n s i o n s r e c e n t r i f u g e d .
The p e l l e t s o b t a i n e d from t h i s s e c o n d e x t r a c t i o n w e r e a g a i n
r e s u s p e n d e d i n t h e same b u f f e r a s f o r t h e s e c o n d h o m o g e n i s a t i o n
s t e p . The s u p e r n a t a n t s and h o m o g e n a t e s o b t a i n e d w e r e t h e n
a s s a y e d f o r AChE a c t i v i t y by t h e r a d i o m e t r i c a s s a y m ethod
( s e c t i o n 2 . 2 7 ) . The s u p e r n a t a n t s o b t a i n e d from t h e f i r s t and
s e c o n d e x t r a c t i o n w e r e s u b j e c t e d t o d e n s i t y g r a d i e n t a n a l y s i s
( s e c t i o n 2 . 2 . 6 ) .
I t was f o u n d t h a t t h e f i r s t e x t r a c t i o n y i e l d e d 90-100% o f
t h e enzyme a c t i v i t y p r e s e n t i n t h e m u s c l e s ( s e e T a b l e ) . The
d i s t r i b u t i o n o f t h e m o l e c u l a r fo rm s o f AChE o b t a i n e d from t h e
f i r s t e x t r a c t i o n was t y p i c a l o f t h a t o b s e r v e d f o r t h e two
m u s c l e s , w h e r e a s t h e s e c o n d e x t r a c t s c o n t a i n e d r e s i d u a l AChE,
m a i n l y L a nd H f o rm s and c o n s t i t u t e d f o r l e s s t h a n 8% o f t o t a l
AChE a c t i v i t y i n t h e m u s c l e s ( s e e F i g u r e ) . I t i s c l e a r ,
t h e r e f o r e , t h a t a l l t h e m o l e c u l a r f o rm s a r e e f f i c i e n t l y e x t r a c t e d
i n t h e h i g h s a l t / T r i t o n b u f f e r d u r i n g r a p i d p r o c e s s i n g o f t i s s u e
c r u d e e x t r a c t s .
- 1 8 1 -

FRACTIONS FRACTIONS
S u c r o s e g r a d i e n t p r o f i l e s o f AChE i n c r u d e e x t r a c t s o f r a t EDL
and s o l e u s
S o l i d l i n e r e p r e s e n t s s u c r o s e g r a d i e n t p r o f i l e o f AChE from t h e
f i r s t e x t r a c t i o n ; a c t i v i t y i n d i c a t e d by t h e y - a x i s on t h e l e f t
hand s i d e . D o t t e d l i n e r e p r e s e n t s s u c r o s e g r a d i e n t p r o f i l e o f
AChE from t h e s e c o n d e x t r a c t i o n ; a c t i v i t y i n d i c a t e d by t h e y - a x i s
on t h e r i g h t hand s i d e . A l l e x t r a c t s w e r e p r e p a r e d i n t h e
p r e s e n c e o f p r o t e a s e i n h i b i t o r s .
AChE a c t i v i t y i n v a r i o u s e x t r a c t s o f r a t EDL and s o l e u s
[AChE] ( U n i t s / g )
M u s c l e Hom ogenate 1 s t e x t r a c t 2nd e x t r a c t p e l l e t
EDL 1 . 1 2 1 . 2 1 0 . 1 0 0 . 0 6
s o l e u s 0 . 8 9 0 . 9 0 0 . 0 5 0 . 0 5
- 1 8 2 -

A p p e n d ix 2 . M o le c u la r s i z e d e t e r m in a t io n o f ACHE b y r a d i a t i o n
i n a c t i v a t i o n
T a r g e t s i z e a n a l y s i s by h i g h e n e r g y r a d i a t i o n i n a c t i v a t i o n
h a s b e e n u s e d f o r d e t e r m i n i n g t h e minimum m o l e c u l a r w e i g h t
a s s o c i a t e d w i t h a b i o l o g i c a l a c t i v i t y i n a p r o t e i n ( f o r r e v i e w ,
s e e Kempner a n d S c h l e g e l , 1 9 7 9 ) . T h i s t e c h n i q u e h a s b e e n u s e d t o
d e t e r m i n e t h e m o l e c u l a r s i z e o f H2 c AChE i n c h i c k e n m u s c l e
METHOD
The s o u r c e o f H2 c AChE was e i t h e r p r e p a r e d a s a membrane
s u s p e n s i o n o r i n s o l u b l e fo rm . F o r membrane p r e p a r a t i o n s , o n e
day o l d c h i c k p e c t o r a l m u s c l e was h o m o g e n i s e d i n 10 v o l o f 10 mM
^ 4^ 3/ ^ EDTA> PH 7 . 0 c o n t a i n i n g , a s p r o t e a s e i n h i b i t o r s ,
b a c i t r a c i n ( l m g / m l ) , b e n z e m i d i n e ( 2 n t l ) , NEM ( 5 nM) a n d S o y a b e a n
t r y p s i n i n h i b i t o r ( 0 . 1 m g / m l ) , and c e n t r i f u g e d a t 4 3 , 5 0 0 x g f o r
30 m in . The same p r o c e d u r e was r e p e a t e d o n c e and t h e f i n a l
p e l l e t o b t a i n e d w a s r e s u s p e n d e d i n 10 v o l o f 10 mM NH^COg/O.l mM
ntl EDTA, pH 7 . 0 , and f i l t e r e d t h r o u g h g l a s s w o o l . For s o l u b l e
H2 c AChE p r e p a r a t i o n s , p e c t o r a l m u s c l e s w e r e h o m o g e n i s e d w i t h
10 v o l o f 10 nM p h o s p h a t e / l n M EGTA, pH 7 . 0 c o n t a i n i n g p r o t e a s e
i n h i b i t o r s a s a b o v e and c e n t r i f u g e d i n a 45 Ti r o t o r (Beckman) a t
28 K rpm f o r 3 0 m in . T h i s p r o c e d u r e was r e p e a t e d a nd t h e f i n a l
p e l l e t o b t a i n e d w a s e x t r a c t e d w i t h 3 v o l o f 1M N a C l / 2 0 mM sodium
p y r o p h o s p h a t e / 1 mM MgCl2 / l mM EGTA/10 nM p h o s p h a t e , pH 8 . 0 ,
c o n t a i n i n g p r o t e a s e i n h i b i t o r s a s a b o v e . The h o m o g e n a te was
c e n t r i f u g e d a t 26 K rpm i n a Beckman SW 6 0 Ti r o t o r f o r 3 0 min
- 183 -

and i t s s u p e r n a t a n t a p p l i e d t o d e n s i t y g r a d i e n t c e n t r i f u g a t i o n ,
a s d e s c r i b e d i n S e c t i o n 2 . 2 . 6 (5% a nd 20% s u c r o s e s o l u t i o n s w e r e
made up i n 1M N a C l / 1 0 nW p h o s p h a t e , pH 7 . 0 ) . The g r a d i e n t
f r a c t i o n s w h ic h c o n t a i n e d t h e 20 S peak w e r e p o o l e d .
Membrane s u s p e n s i o n o r p o o l e d 20 S f r a c t i o n s w e r e a l i q u o t e d
i n t o P y r e x s a m p le i r r a d i a t i o n t u b e s ( 1 2 x 1 0 0 mm) ( C o r n i n g G l a s s
L t d . ) , ADH (E C . l 1 1 . 1 ) (1 u n i t / s a m p l e t u b e ) and p y r u v a t e k i n a s e
( E . C . 2 . 7 . 1 . 4 0 ) ( 2 0 u n i t s / s a m p l e t u b e ) w e r e r o u t i n e l y u s e d a s
i n t e r n a l p r o t e i n s t a n d a r d s . The s a m p l e s 7 t r i p l i c a t e d f o r e a c h
r a d i a t i o n d o s a g e and n o n - i r r a d i a t e d c o n t r o l s , w e r e f r o z e n i n dry
i c e . f l u s h e d w i t h g a s and t h e t u b e s s e a l e d w i t h an o x y g e n
f l a m e t o r c h . The s a m p l e s w e r e i r r a d i a t e d w i t h 8 MeV e l e c t r o n s a t
t h e Hammersmith H o s p i t a l l i n e a r e l e c t r o n a c c e l e r a t o r , a t a d o s e
r a t e o f 0 . 4 M r a d /m in , w h ic h was m o n i t o r e d w i t h e l e c t r o n
c o l l e c t o r s a b o v e and b e l o w t h e sa m p le a r e a , t o d e t e r m i n e t h e
t o t a l d o s e and t h e d o s e r a t e r e s p e c t i v e l y . I r r a d i a t e d s a m p le
t u b e s w e r e s t o r e d a t - 7 0 ° C u n t i l o p e n e d f o r a s s a y .
A l i q u o t s o f n o n - i r r a d i a t e d c o n t r o l and i r r a d i a t e d membrane
p r e p a r a t i o n w e r e e x t r a c t e d w i t h a 1% T r i t o n / I M NaCl/lmM EDTA/
p h o s p h a t e b u f f e r , pH 7 . 0 , c e n t r i f u g e d a t 4 3 , 5 0 0 x g f o r 30 min and
t h e AChE a c t i v i t y i n t h e s u p e r n a t a n t s m ea s u r e d by t h e m ethod o f
E l l man e t a l . ( 1 9 6 1 ) a s i n S e c t i o n 2 . 2 . 7 . N o n - i r r a d i a t e d
c o n t r o l and i r r a d i a t e d 20S f r a c t i o n s w e r e th aw ed and a l i q u o t s
w ere a s s a y e d f o r AChE a c t i v i t y by t h e r a d i o m e t r i c m eth od ( S e c t i o n
2 . 2 . 7 ) . T h i s work w as c a r r i e d o u t i n c o l l a b o r a t i o n w i t h A. L a i ,
who p e r f o r m e d t h e i r r a d i a t i o n p r o c e d u r e and t h e s u b s e q u e n t a s s a y s
o f t h e i n t e r n a l p r o t e i n s t a n d a r d s .
- 184 -

R e s u l t s
The d o s e - d e p e n d e n t i n a c t i v a t i o n o f t h e 20S fo rm o f AChE i s
shown i n F i g . 1 . The i n a c t i v a t i o n o f o n e o f t h e i n t e r n a l p r o t e i n
s t a n d a r d s i s a l s o sho wn. The m o l e c u l a r s i z e o f AChE d e t e r m i n e d
fr o m t h e c a l i b r a t i o n c u r v e o f A . 1_ v s . m o l e c u l a r w e i g h t o fAo D o se
p r o t e i n s t a n d a r d s was 1 1 0 , 0 0 0 d , w h ic h a g r e e s w i t h t h e s u b u n i t
m o l e c u l a r mass e s t i m a t i o n o f p u r i f i e d c h i c k AChE ( R o t u n d o , 1 9 8 4 ) .
A n a l y s i s o f t h e i r r a d i a t e d membrane p r e p a r a t i o n g a v e i d e n t i c a l
r e s u l t s . T h a t s u c h o b s e r v a t i o n i s n o t due t o d e g r a d a t i o n o f t h e
20S fo rm i s i l l u s t r a t e d i n F i g . 2 . N o n - i r r a d i a t e d c o n t r o l
s a m p l e s o f a membrane p r e p a r a t i o n c o n t a i n s p r e d o m i n a n t l y
AChE. T h u s , t h e s e a n a l y s e s show t h a t t h e minimum m o l e c u l a r s i z e
a s s o c i a t e d w i t h t h e h y d r o l y s i s o f ACh i s i d e n t i c a l t o t h e
m on om er ic c a t a l y t i c u n i t m o l e c u l a r m a s s , s u g g e s t i n g t h a t t h e
monomers a c t a s i n d e p e n d e n t e n z y m i c u n i t s , and t h e r e i s i n s u f f i c i e n t
e n e r g y t r a n s f e r b e t w e e n t h e c a t a l y t i c s i t e s w i t h i n t h e m u l t i m e r
s u c h a s t h e H f o r m . P r e v i o u s o b s e r v a t i o n s on t h e s t r u c t u r e and
s t a b i l i t y o f t h e H2 c ^ ] 2 ^ ^orm E^e c t r °P*10rus S *10W t h a t
l i t t l e i n t e r a c t i o n e x i s t s b e t w e e n t h e t e t r a m e r i c u n i t s l i n k e d t o
a common c o l l a g e n t a i l ( M a s s o u l i e and Bon, 1 9 8 2 ) . The r e s u l t s o f
t h i s s t u d y s u g g e s t t h a t t h e r e a p p e a r s t o be l i t t l e a l l o s t e r i c
i n t e r a c t i o n w i t h i n t h e t e t r a m e r s . The m o l e c u l a r s i z e o f t h e
AChE fo rm i n c h i c k e n m u s c l e , t h e r e f o r e , c a n n o t be r e s o l v e d by
t a r g e t s i z e a n a l y s i s .
- 185 -

FRACTIONS
F I G . 2 S u c r o s e g r a d i e n t p r o f i l e o f AChE e x t r a c t e d from a n o n -
I r r a d i a t e d c o n t r o l s a m p l e o f m u s c l e membrane p r e p a r a t i o n .
-186-

1-0
4 8 12 16Mrad
F I G . 1 D o s e - d e p e n d e n t I n a c t i v a t i o n o f 20S form o f AChE from
c h i c k m u s c l e . A, enzyme a c t i v i t y a f t e r i r r a d i a t i o n ; AQ, o r i g i n a l ( n o n - i r r a d i a t e d ) enzym e a c t i v i t y . PK, p y r u v a t e k i n a s e .
- 187 -

REFERENCES
- 188 -

A A n g l i s t e r , L . , and S i l m a n , I . ( 1 9 7 8 ) , J . M ol . B i o l ., 12 5 , 2 9 3 - 3 1 1 .
A r i a n o , M . A . , A r m s t r o n g , R .B . a n d E d g e r t o n , V .R . ( 1 9 7 3 ) , J .
H i s t o c h e m . C y t o c h e m . , 2 1 , 5 1 - 5 5 .
A r n d t , I . a n d P e p e , F . ( 1 9 7 5 ) , J . H i s t o c h e m . C y t o c h e m . , 2 3 ,
1 5 9 - 1 6 8 .
A sh m ore , C .R . ( 1 9 8 2 ) , Am. J . P h y s i o l . , 2 4 2 , C 1 7 8 - C 1 8 3 .
A sh m ore , C.R. a nd D o e r r , L . ( 1 9 7 1 ) , Exp. N e u r o l . , 3 0 , 4 3 1 - 4 4 6 .
Asm und son , V .S and J u l i a n , L.M. ( 1 9 5 6 ) , J . H e r e d . , 47 ^ 2 4 8 - 2 5 2 .
A u g u s t i n s s o n , K .B. a n d Nachmansohn, D. ( 1 9 4 9 ) , S c i e n c e , 1 1 0 , 9 8 ;
A u s t i n , L. and B e r g , W.K. ( 1 9 5 3 ) , B io c h e m . J . , 5 4 , 6 9 5 - 7 0 0 .
B B a c o u , F . , V i g n e r o n , P . and M a s s o u l i e , J . ( 1 9 8 2 ) , N a t u r e , 2 9 6 ,
6 6 1 - 6 6 4 .
Bandman, E. ( 1 9 8 4 ) , S c i e n c e , 2 2 7 , 7 8 0 - 7 8 2 .
B a r a n y , M. ( 1 9 6 7 ) , J . Gen. P h y s i o l . , 5 0 , ( s u p p l . 2 ) , 1 9 7 - 2 1 8 .
B a r a n y , M . , B a r a n y , K . , R i c k a r d , T. and V o l p e , A. ( 1 9 6 5 ) , Arch.
B i o c h e m . B i o p h y s . , 1 0 9 , 1 8 5 - 1 9 1 .
B a r a n y , M. and C l o s e , R . I . ( 1 9 7 1 ) , J . P h y s i o l . , 2 1 3 , 4 5 5 - 4 7 4 .
B a r n a r d , E . A . , B a r n a r d , P . J . a nd J a r v i s , J . ( 1 9 8 5 ) , 'Low f r e q u e n c y
c h r o n i c e l e c t r i c a l s t i m u l a t i o n o f normal and d y s t r o p h i c
c h i c k e n m u s c l e ' J . P h y s i o l , ( s u b m i t t e d f o r p u b l i c a t i o n ) .
B a r n a r d , E . A . , B a r n a r d , P . J . , J a r v i s , J . J e d r z e j c z y k , J . , L a i ,
J . , P i z z e y , J . A . , R a n d a l l , W.R. and S i l m a n , I . ( 1 9 8 4 a ) , I n :
C h o l i n e s t e r a s e s ( B r z i n £ t al^. e d s . ) , pp . 4 9 - 7 1 . W a l t e r de
G r u y t e r C o . , B e r l i n , N.Y.
B a r n a r d , E . A . , L a i , J . and P i z z e y , J . A . ( 1 9 8 4 b ) , In: N e u r o
m u s c u l a r d i s e a s e s ( S e r r a t r i c e , G. e t a l , e d s . ) , p p . 4 5 5 - 4 6 3 ,
Raven P r e s s , N .Y .
- 1 8 9 -

B a r n a r d , E . A . , L y l e s , J . M . and P i z z e y , J . A . ( 1 9 8 2 ) , J . P h y s i o l .,
3 3 1 , 3 3 3 - 3 5 4 -
B a r n a r d , R . J . , E d g e r t o n , V . R . , Furukawa, T. and P e t e r , J . B .
( 1 9 7 1 ) , Am. J . P h y s i o l . , 2 2 0 , 4 1 0 - 4 1 4 .
B a r n e t t , P . and R o s e n b e r r y , T . L . ( 1 9 7 9 ) , B . B . A . , 5 6 7 , 1 5 4 - 1 6 0 .
B a r o n e , R . , P a v a u x , C . , B l i n , P .C . and Cuq, P . ( 1 9 7 3 ) , A t l a s
D ' a n a t o m i e du L a p i n , Mason and C o . , P a r i s .
B a r r e t t , A . J . and D i n g l e , J . T . ( 1 9 7 2 ) , B io c h e m . J . 1 2 7 , 4 3 9 - 4 4 1 .
B a r r e t t , A . J . and S t a r k e y , P .M. ( 1 9 7 3 ) , B io c h e m . J . , 1 3 3 ,
7 0 9 - 7 2 4 .
B e n d i t t , E . P . ( 1 9 5 6 ) , Am. S o c . Exp. P a t h . , 15_, 5 0 7 . ( a b s . 1 6 4 6 ) .
Berman, J . D . ( 1 9 7 3 ) , B i o c h e m i s t r y , \ 2 , 1 7 1 0 - 1 7 1 5 .
Berman, J . D . and You ng , M. ( 1 9 7 1 ) , P . N . A . S . , 6 8 , 3 9 5 - 3 9 8 .
B e t z , H . , B o u r g e o i s , J . P . and C h an geux , J . P . ( 1 9 8 0 ) , J . P h y s i o l .
3 0 2 , 1 9 7 - 2 1 8 .
B i r k , Y. ( 1 9 7 6 ) , M e th o d s i n E n z y m o l . , XLV, 7 0 0 - 7 0 7 .
B o i s , P . ( 1 9 6 4 ) , E x p e r i e n t i a , 2 0 , 1 4 0 - 1 4 1 .
Bon, S . ( 1 9 8 2 ) , N eu ro ch em . I n t . , 4 , 5 7 7 - 5 8 5 .
Bon, S . C a r t a u d , J . a nd M a s s o u l i e , J . ( 1 9 7 8 ) , E u r . J . B io c h e m .
8 5 , 1 - 1 4 .
Bon, S . , H u e t , M . , L e m o n n i e r , M. R i e g e r , F . and M a s s o u l i e , J .
( 1 9 7 6 ) , E u r . J . B i o c h e m . , 6 8 , 5 2 3 - 5 3 0 .
Bon, S . and M a s s o u l i e , J . ( 1 9 7 6 ) , FEBS L e t t . , 71 , 2 7 3 - 2 7 8 .
Bon, S . and M a s s o u l i e , J . ( 1 9 7 8 ) , Eur . J . B i o c h e m . , 8 9 , 8 9 - 9 4 .
Bon, S . V i g n y , M. and M a s s o u l i e , J . ( 1 9 7 9 ) , P N . A . S . , 76 ,
2 5 4 6 - 2 5 5 0 .
B r a n d a n , E. and I n e s t r o s a , N .C. ( 1 9 8 4 ) , B i o c h e m . J . , 2 2 1 ,
4 1 5 - 4 2 2 .
- 190 -

B r a u n - F a l c o , 0 . and S a l f e l d , K. ( 1 9 5 9 ) , N a t u r e , 1 8 3 , 5 1 - 5 2 .
B r e n n e r , H . R . , M e i e r , Th. and Widmer, B . ( 1 9 8 3 ) , N a t u r e , 3 0 5 ,
5 3 6 - 5 3 7 .
B r i m i j o i n , S . ( 1 9 8 3 ) , P r o g . N e u r o b i o l . , 2 1 , 2 9 1 - 3 2 2 .
B r o o k e , M.H. and K a i s e r , K.K. ( 1 9 7 0 ) , A r c h s . N e u r o l . , 2 3 ,
3 6 9 - 3 7 9 .
B r z i n , M. and M a j c e n - T k a £ e v , Z . ( 1 9 6 3 ) , J C .B . ,1 9 ^ , 3 4 9 - 3 5 8 .
B u l g e r , J . E . , R a n d a l l , W . R . , N i e b e r g , P . S . , P a t t e r s o n , G . T . ,
McNamee, M.G. and W i l s o n , B W. ( 1 9 8 2 ) , Dev . N e u r o s c i . , 5_,
4 7 4 - 4 8 3 .
B u l l e r , A . J . , E c c l e s , J . C . and E c c l e s , R.M. ( 1 9 6 0 a ) , J .
P h y s i o l . , 1 5 0 , 3 9 9 - 4 1 6 .
B u l l e r , A . J . , E c c l e s , J . C . and E c c l e s , R M. ( 1 9 6 0 b ) , J . P h y s i o l .
1 5 0 , 4 1 7 - 4 3 9 .
B u rd en , S . J . , S a r g e n t , P .B and McMahan, U . J . ( 1 9 7 9 ) , J . C . B . , 8 2 ,
4 1 2 - 4 2 5 .
B u r k e , R.E , L e v i n e , D . N . , Z a j a c , F E . , I l l , T s a i r i s , P . and
E n g e l , W . K . , ( 1 9 7 1 ) , S c i e n c e , 1 7 4 , 7 0 9 - 7 1 2 .
B u t l e r , I . J . , Drachman, D . B . and G o l d b e r g , A M . ( 1 9 7 8 ) , J .
P h y s i o l . , 2 7 4 , 5 9 3 - 6 0 0 .
C a b i b , E. and U l a n e , R. ( 1 9 7 3 ) , B i o c h e m . B i o p h y s . R e s . Comm.
5 0 , 1 8 6 - 1 9 1 .
C a m p b e l l , J . B . ( 1 9 7 9 ) , M . S c . t h e s i s : "A q u a n t i t a t i v e
h i s t o c h e m i c a l s tucty o f t h e c h a n g e i n a c i d p h o s p h a t a s e w i t h
a g e i n normal and d y s t r o p h i c s k e l e t a l m u s c l e . " D e p a r t m e n t o f
Anatomy, U n i v e r s i t y o f D u n d ee , D u n d ee , S c o t l a n d .
C a n g i a n o , A . , L0mo, T . , L u t z e m b e r g e r , L. and S v e e n , 0 . ( 1 9 8 0 ) ,
A c t a P h y s i o l . S c a n d . , 1 0 9 , 2 8 3 - 2 9 6 .
C a r s o n , S . , Bon, S . , V i g n y , M . , M a s s o u l i e , J . and F a r d e a u , M.
( 1 9 7 9 ) , FEBS L e t t . , 9 6 , 3 4 8 - 3 5 2— -191-

C a r t e r , J . L . and B r i m i j o i n , S . ( 1 9 8 1 ) , J . Neurochem. , 3 6 ,
1 0 1 8 - 1 0 2 5 .
Cha n, S . L . , S h i r a c h i , D . Y . , B h a r g a v a , H . N . , G a r d n e r , E . and
T r e v o r , A . J . ( 1 9 7 2 ) , J . N e u r o c h e m . , 1 9 , 2 7 4 7 - 2 7 5 8 .
C h a n g e u x , J . P . ( 1 9 6 6 ) , M o l . P h a r m a c o l . , 2 , 3 6 9 - 3 9 2 .
C h i u , H. a n d L a g u n o f f , D. ( 1 9 7 2 ) , H i s t o c h e m . J . , 4 , 1 3 5 - 1 4 4 .
Chubb, I . W . , Goodman, S . and S m i t h , A .D . ( 1 9 7 6 ) , N e u r o s c i e n c e , 1_,
5 7 - 6 2 .
C i s s o n , C.M. and W i l s o n , B . W . . ( 1 9 7 7 ) , B io c h e m . P h a r m a c . , 2 6 ,
1 9 5 5 - 1 9 6 0 .
C i v i n , C . I . and B a n q u e r i g o , M.L. ( 1 9 8 3 ) , J . Imm. M e t h o d s , j r t ,
1- 8 .C l o s e , R . I . ( 1 9 7 2 ) , P h y s i o l . R e v . , 5 2 , 1 2 9 - 1 9 7 .
C o h en , S . ( 1 9 6 7 ) , In : Nobel S ym po sia n o . 3 . P r o c . 3 r d Nobel
S y m p o s i a . ( K i l l a n d e r , J . e d . ) , p p . 2 1 - 3 2 . A l m q u i s t and
W i k s e l l , S t o c k h o l m ; John W i l e y , Lo n do n.
C o l l i n s , R .L . and Y o u n k i n , S . G . ( 1 9 8 2 ) , J . B . C . , 25 7 , 1 3 6 3 8 - 1 3 6 4 4 .
C o o p e r , G. and S c h i l l e r , A . L . ( 1 9 7 5 ) , Anatorny o f t h e g u i n e a p i g ,
H a rv a r d U n i . P r e s s , C a m b r id g e , M a s s .
C o r n e l i u s , C . E . , Law, R . J . , J u l i a n , L.M. and Asmund son, V . S .
( 1 9 5 9 ) , P r o c . S o c . Exp. B i o l . M e d . , 1 0 1 , 4 1 - 4 4 .
C os m os , E . , B u t l e r , J . , A l l a r d , E . P . and M a z l i a h , J . ( 1 9 7 9 ) ,
Ann. N .Y. A c a d . S c i . , 3 1 7 , 5 7 1 - 5 9 3 .
C o t t e r , M . , H u d l i c k a , 0 . , P e t t e , D . , S t a u d t e , H.W. a n d V r b o v a .
G. ( 1 9 7 3 ) , J . P h y s i o l . , 2 3 0 , 3 4 P .
C o u ra u d , J . Y . and Di G i a m b e r a r d i n o , L . ( 1 9 8 0 ) , J . N e u r o c h e m . ,
3 5 , 1 0 5 3 - 1 0 6 6 .
- 192 -

D o a v e y , B . , Y o u n k i n , L .H . and Y o u n k in , S G. ( 1 9 7 9 ) , J . P h y s i o l . ,
2 8 9 , 5 0 1 - 5 1 5 .
Di G i a m b e r a r d i n o , L . f c C o u r a u d J . Y . ( 1 9 7 8 ) , N a t u r e 27 1 1 7 0 - 1 7 2 .
Drachman, D . B . ( 1 9 7 2 ) , J . P h y s i o l . , 2 2 6 , 6 1 9 - 6 2 7 .
Drachman, D .B . a n d W i t z k e , F . ( 1 9 7 2 ) , S c i e n c e , 17 6 , 5 1 4 - 5 1 6 .
D u b o w i t z , Y. and B r o o k e , M.H. ( 1 9 7 3 ) , M u s c l e B i o p s y : A modern
a p p r o a c h . London: W.B. S a u n d e r s Co. L t d .
D u d a i , Y . , H e r z b e r g , M. a nd S i l m a n , I . ( 1 9 7 3 ) , P . N . A . S . ( U S A ) , 70
2 4 7 3 - 2 4 7 6 .
D u d a i , Y. and S i l m a n , I . ( 1 9 7 4 ) , B io c h e m . B i o p h y s . R e s . Comm.
5 9 , 1 1 7 - 1 2 4 .
D u t t a - C h o u d h u r y , T .A . and R o s e n b e r r y , T .L ( 1 9 8 4 ) , J . B . C . , 2 5 9 ,
5 6 5 3 - 5 6 6 0 .
F■— E c c l e s , J . C . , E c c l e s , R.M. and Lu ndberg , A. ( 1 9 5 8 ) , J . P h y s i o l .,|
1 4 2 , 2 7 5 - 2 9 1 .
E e r b e e k , 0 . , K e r n e l l , D. and V e r h e y , B .A . ( 1 9 8 4 ) , J . P h y s i o l . , 3 5 2
7 3 - 9 0 .
E i s e n b e r g , B .R . and S a l m o n s , S . ( 1 9 8 1 ) , C e l l T i s s u e R e s . , 2 2 0 ,
4 4 9 - 4 7 1 .
E l l i s m a n , M . H . , R a s h , J . E . , S t a e h e l i n , L . A . and P o r t e r , K.R.
( 1 9 7 6 ) , J . C . B . , 6 8 , 7 5 2 - 7 7 4 .
E l l m a n , G . L . , C o u r t n e y , K . D . , A n d r e s , V . J r . and F e a t h e r s t o n e ,
R.M. ( 1 9 6 1 ) , B io c h e m . P h a r m a c o l 8 8 - 9 5 .
Ey, P . L . , P r o w s e , S . J . and J e n k i n , C. R. ( 1 9 7 8 ) , I m m u n o c h e m is t r y ,
1 5 , 4 2 9 - 4 3 6 .
- 193 -

F F a l l o n , J . R . , N i t k i n , R . M . , R e i s t , N . E . , W a l l a c e , B .C . and
McMahan, U . J . ( 1 9 8 5 ) , N a t u r e , 3 1 5 , 5 7 1 - 5 7 4 .
Fam bor oug h, D . M . , E n g e l , A .G. and R o s e n b e r r y , T . L . ( 1 9 8 2 ) ,
P . N . A . S . , 7 9 , 1 0 7 8 - 1 0 8 2 .
F e r n a n d e z , H .L. and D u e l ! , M .J . ( 1 9 8 0 ) , J . N eu r o c h e m . , 3 5 ,
1 1 6 6 - 1 1 7 1 .
F e r n a n d e z , H . L . , D u e l l , M . J . and F e s t o f f , B.W. ( 1 9 7 9 ) , J .
N e u r o c h e m . , 3 2 , 5 8 1 - 5 8 5 .
F r a n k , E . , J a n s e n , J . K . S . , L0mo, T. and W e s t g a a r d , H. ( 1 9 7 5 ) , J .
P h y s i o l . , 2 4 7 , 7 2 5 - 7 4 3 .
F u l t o n , M .P . and M o g ey , G .A . ( 1 9 5 4 ) , B r i t . J . P h a r m a c o l . , 9^,
1 3 8 - 1 4 4 .
G G a u t h i e r , G . F . ( 1 9 6 9 ) , Z. Z e l I f o r s c h . , 95 , 4 6 2 .
G a u t h i e r , G .F . a n d P a d y k u l a , H. ( 1 9 6 6 ) , J . C . B . , 2 8 , 3 3 3 - 3 5 4 .
G i n s b o r g , B . L . ( 1 9 6 0 ) , J . P h y s i o l . , 1 5 4 , 5 8 1 - 5 9 8 .
G i s i g e r , V. a n d S t e p h e n s , H.R. ( 1 9 8 3 ) , J . N e u r o c h e m . , 4 1 ,
9 1 9 - 9 2 9 .
G i s i g e r , V. and V i g n y , M. ( 1 9 7 7 ) , FEBS l e t t . , 8 4 , 2 5 3 - 2 5 6 .
G i s i g e r , V . , V i g n y , M . , G a u t r o n , J . R i e g e r , F . ( 1 9 7 8 ) , J .
N e u r o c h e m . , 3 0 , 5 0 1 - 5 1 6 .
G l e n n e r , G.G. and C o h e n , L .A . ( 1 9 6 0 ) , N a t u r e , 1 8 5 , 8 4 6 - 8 4 7 .
G o l d s p i n k , G. a n d Ward, P . S . ( 1 9 7 9 ) , J . P h y s i o l . , 2 9 6 , 4 5 3 - 4 6 9 .
G ord on, T. and V r b o v a , G. ( 1 9 7 5 ) , P f l i i g e r s . Arch., , 3 6 0 , 1 9 9 - 2 1 8 .
G r a f i u s , M.A. a nd M i l l a r , D .B . ( 1 9 6 5 ) , B io c h e m . B i o p h y s . A c t a ,
1 1 0 , 5 4 0 - 5 4 7 .
G r a s s i , J . , M a s s o u l i e , J . and T i m p l , R. ( 1 9 8 3 ) , E u r . J . B i o c h e m . ,
1 3 3 , 3 1 - 3 8 .
-194-

G r a s s i , J . , V i g n y , M. and M a s s o u l i e , J . ( 1 9 8 2 ) , J . N eu r o c h e m . ,
3 8 , 4 5 7 - 4 6 9 .
G r e e n , J . H . , K l u g , G . A . , R e ic hm an n, H. , S e e d o r f , U . 3 W i e h r e r , W.
a n d P e t t e , D. ( 1 9 8 4 ) , P f l t i g e r s A r c h . , 4 0 0 , 4 3 2 - 4 3 8 .
G r o s w a l d , D .E . and D e t t b a r n , W.D. ( 1 9 8 3 a ) , N e u r o c h e m . R e s . , 8 ,
9 8 3 - 9 9 5 .
G r o s w a l d , D .E . and D e t t b a r n , W.D. ( 1 9 8 3 b ) , E x p . N e u r o l . 3 79 ,
5 1 9 - 5 3 1 .
G u th , L . ( 1 9 6 8 ) , P h y s i o l . R e v . , 4 8 6 4 5 - 6 8 7 .
Gutmann, E. a nd Zak, R. ( 1 9 6 1 ) , P h y s i o l . B o h e m o s l o v . , 1 0 ,
4 9 3 - 5 0 0 .
U* * H a b e r l a n d , G. and McConn, R. ( 1 9 7 9 ) , F e d . P r o c . , 3 8 , 2 7 6 0 - 2 7 6 7 .
H a l l , Z.W. ( 1 9 7 3 ) , J . N e u r o b i o l . , 4 , 3 4 3 - 3 6 1 .
H a r r i s , A . J . ( 1 9 8 1 ) , P h i l . T r a n s . R. S o c . L o n d . , 2 9 3 , 2 7 9 - 2 8 6 .
H a y n e s , R. and F e e n e y , R . E . ( 1 9 6 7 ) , J . B . C . , 2 4 2 , 5 3 7 8 - 5 3 8 5 .
H e i l i g , A . and P e t t e , D. ( 1 9 8 3 ) , FEBS L e t t . , 1 5 1 , 2 1 1 - 2 1 4 .
H e i lm a n n , C . , M u l l e r , W. a n d P e t t e , D. ( 1 9 8 1 ) , J . Membr. B i o l . ,
5 9 , 1 4 3 - 1 4 9 .
H e i lm a n n , C. and P e t t e , D. ( 1 9 7 9 ) , Eur . J . B i o c h e m . , 9 3 , 4 3 7 - 4 4 6 .
H e s s , A. ( 1 9 6 1 ) , J . P h y s i o l . , 1 5 7 , 2 2 1 - 2 3 1 .
H e s s , A. ( 1 9 6 7 ) , I n v e s t . O p h t h a l m o l . , 6 , 2 1 7 .
H e s s , A. ( 1 9 7 0 ) , P h y s i o l . R e v . , 5 0 , 4 0 - 6 2 .
H n ik , P . , J i r m a n o v a , I . V y k l i c k y , L. and Z e l e n a , J . ( 1 9 6 7 ) , J .
P h y s i o l . , 1 9 3 , 3 0 9 - 3 2 5 .
H o d g e s o n , A . J . and Chubb, I .W. ( 1 9 8 3 ) , J . N e u r o c h e m . , 4 1 ,
6 5 4 - 6 6 2 .
H o l l i d a y , T A . , V a n m e t e r , J . R . , J u l i a n , L.M. and A sm un d son , V . S .
( 1 9 6 5 ) , Am. J . P h y s i o l . , 2 0 9 , 8 7 1 - 8 7 6 .
- 1 9 5 -

H o l l u n g e r , E .G . and N i k l a s s o n , B .H. ( 1 9 7 3 ) , J . N e u r o c h e m . , 2 0 ,
8 2 1 - 8 3 6 .
H o l m s t e d t , B. ( 1 9 5 7 ) , A c t a P h y s i o l . S c a n d . , 4 0 , 3 2 2 - 3 3 0 .
H o p f f , W .H . , R i g g i o , G. and W a s e r , P .G . ( 1 9 7 3 ) , FEBS L e t t . , 3 5 ,
220- 222.
Hummel, B . C . W . , ( 1 9 5 9 ) , Can. J . B i o c h e m . P h y s i o l . , 3 7 , 1 3 9 3 .
H u s t o n , R .B . a n d K r e b s , E . G . ( 1 9 6 8 ) , B i o c h e m i s t r y , 7_, 2 1 1 6 - 2 1 2 2 .
^ I n e s t r o s a , N . C . , M a t t h e w , W . D . , R e i n e s s , C . G . , H a l l , Z.W. and
R e i c h a r d t , L . F . ( 1 9 8 5 ) , J . N e u r o c h e m . , 4 5 , 8 6 - 9 4 .
I n e s t r o s a , N . C . , R a m i r e z , B U. and F e r n a n d e z , H.L. ( 1 9 7 7 ) , J .
N e u r o c h e m . , 2 8 , 9 4 1 - 9 4 5 .
I n e s t r o s a , N . C . , S i l b e r s t e i n , L. and H a l l , Z.W. ( 1 9 8 2 ) , C e l l ,
2 9 , 7 1 - 7 9 .
I s h i k a w a , H. ( 1 9 8 3 ) , In: C e l l and m u s c l e a c t i v i t y , v o l . 4
(Dowben, R.M. a nd S h a y , J .W . e d s . ) p p . 1 - 5 9 . P lenum P r e s s ,
N .Y . and L on do n.
J e d r z e j c z y k , J . S i l m a n , I . , L a i , J . and B a r n a r d , E .A . ( 1 9 8 4 ) ,
N e u r o s c i . L e t t . , 4 6 , 2 8 3 - 2 8 9 .
J e d r z e j c z y k , J . , S i l m a n , I . , L y l e s , J . M . and B a r n a r d , E.A.
( 1 9 8 1 ) , B i o s c i . R e p . , ^ , 4 5 - 5 1 .
J i r m a n o v a , I . , H n ik , P . and Z e l e n a , J . ( 1 9 7 1 ) , P h y s i o l .
B o h e m o s l o v . , 2 0 , 1 9 9 - 2 0 4 .
J o h n s o n , C .D . and R u s s e l l , R .L . ( 1 9 7 5 ) , A n a l y t . B i o c h e m . , 6 4 ,
2 2 9 - 2 3 8 .
J o l e s z , F . and S r e t e r , F . A . ( 1 9 8 1 ) , Ann. R ev . P h y s i o l . , 4 3 ,
5 3 1 - 5 5 2 .
- 1 9 6 -

K a p la n , N.O. and Cahn, R .D . ( 1 9 6 2 ) , P . N . A . S . , 4 8 , 2 1 2 3 - 2 1 3 0 .
K a r n o v s k y , M .J . and R o o t s , L . ( 1 9 6 4 ) , J . H i s t o c h e m . C y t o c h e m . ,
12^, 2 1 9 - 2 2 1 .
Katunuma, N. and Kominami, E. ( 1 9 7 7 ) , I n : P r o t e i n a s e s i n
mammalian c e l l s and t i s s u e s . ( B a r r e t t , A . J . e d . ) p p . 1 5 1 - 2 0 8 .
N o r t h - H o l i and p u b l i s h i n g Co.
Kempner, E . S . a nd S c h l e g e l , W. ( 1 9 7 9 ) , A n a l y t . B i o c h e m . , 9 2 ,
2- 10 .
K e n n e t t , R . H . , D e n i s , K . A . , Tung, A . S . and K l i n m a n , N.R. ( 1 9 7 8 ) ,
C u rr . Top . M i c r o b i o l . I m m u n o l . , 81^, 7 7 - 9 1 .
Khan, M.A. ( 1 9 8 0 ) , C y t o b i o s . , 2 8 , 8 5 .
K i r s c h k e , H . , L a n g n e r , J . , W i e d e r a n d e r s , B . , A n s o r g e , S . and
B o h l e y , P . ( 1 9 7 7 ) , E u r . J . B i o c h e m . , 7 4 , 2 9 3 - 3 0 1 .
K i t z , R . J . , B r a s w e l l , L.M. a nd G i n s b u r g , S . ( 1 9 7 0 ) , M o l .
P h a r m a c o l . , 6 t 1 0 8 .
K l u g , G . , W i e h r e r , W. , R e ic hm an n, H . , L e b e r e r , E. and P e t t e , D.
( 1 9 8 3 ) , P f l u g e r s . A r c h . , 3 9 9 , 2 8 0 - 2 8 4 .
K n i g h t , C.G. ( 1 9 8 1 ) , B i o c h e m . J . , 1 8 9 , 4 4 7 - 4 5 3 .
K o e n i g , J . , O ren , M. and M e lo n e , M .A .B . ( 1 9 8 2 ) , D e v . N e u r o s c i . ,
5 , 3 1 4 - 3 2 5 .
K o e n i g , J . and V i g n y , M. ( 1 9 7 8 ) , N a t u r e , 2 7 1 , 7 5 - 7 7 .
K o e l l e , G .B. and Ruch, G.A . ( 1 9 8 3 ) , P . N . A . S . , 8 0 , 3 1 0 6 - 3 1 1 0 .
K o h l e r , G . , and Mil s t e i n , C. ( 1 9 7 5 ) , N a t u r e . 2 5 6 , 4 9 5 - 4 9 7 .
K r e u t z b e r g , G.W., S c h u b e r t , P . , T o t h , L . and R i e s k e , E. ( 1 9 7 3 ) ,
B r a i n R e s . , 6 2 , 3 9 9 - 4 0 4 .
K u n i t z , M. and N o r t h r o p , J . H . ( 1 9 3 6 ) , J . Gen. P h y s i o l . 19^, 9 9 1 .
L a em m li , U.K . ( 1 9 7 0 ) , N a t u r e , 2 2 7 , 6 8 0 - 6 8 5 .
L a g u n o f f , D. and B e n d i t t , E . P . ( 1 9 6 3 ) , Ann. N.Y. A c a d . S c i . , 1 0 3 ,
1 8 5 - 1 9 7 .
-197-

L a g u n o f f , D . , B e n d i t t , E P . and W a t t s , R. ( 1 9 6 2 ) , J . H i s t o c h e m .
C y t o c h e m . , 10 , 6 7 2 - 6 7 3 .
Lai J . , J e d r z e j c z y k J . , P i z z e y J . A . , Green D. and B a r n a r d E .A .
( 1 9 8 5 ) N e u r o n a l c o n t r o l o f t h e fo rm s o f AChE i n s l o w mammalian
m u s c l e s . N a t u r e , s u b m i t t e d f o r p u b l i c a t i o n .
L a z a r , M . , S a l m e v o n , E . , V i g n y , M. and M a s s o u l i e , J . ( 1 9 8 4 ) ,
J . B . C . , 2 5 9 , 3 7 0 3 - 3 7 1 3 .
L a z a r , M. and V i g n y , M. ( 1 9 8 0 ) , J . N e u r o c h e m . , 3 5 , 1 0 6 7 - 1 0 7 9 .
L e e , S . L . , H ein em an n, S . a n d T a y l o r , P . ( 1 9 8 2 a ) , J . B . C . , 2 5 7 ,
1 2 2 8 3 - 1 2 2 9 1 .
L e e , S . L . , Camp, S . J . and T a y l o r , P . ( 1 9 8 2 b ) , J . B . C . , 2 5 7 ,
1 2 3 0 2 - 1 2 3 0 9 .
L e e , S . L . and T a y l o r , P . ( 1 9 8 2 ) , J . B . C . , 2 5 7 , 1 2 2 9 2 - 1 2 3 0 1 .
L e n t z , T . L . ( 1 9 7 4 ) , Ann. N.Y. A cad. S c i . , 2 2 8 , 3 2 3 - 3 3 7 .
L e n t z , T . L . , A d d i s , J . S . and C h e s t e r , J . ( 1 9 8 1 ) , Exp. N e u r o l . ,
7 3 , 1 5 4 2 - 1 5 5 7 .
L e w i s , D . M . , K ea n , C .J C. and M c G a r r i c k , J D. ( 1 9 7 4 ) , Ann. N.Y.
A ca d . S c i . , 2 2 8 , 1 0 5 - 1 2 0 .
L e w i s , P . R . and S h u t e , C .C .D . ( 1 9 6 6 ) , J . C e l l . S c i . , 1 9 3 8 1 - 3 9 0 .
L i , Z . Y . a nd Bon , C. ( 1 9 8 3 ) , J . N eu r o c h e m . , 4 0 , 3 3 8 - 3 4 9 .
L i n k h a r t , T . A . and W i l s o n , B.W. ( 1 9 7 5 ) , Exp. N e u r o l . , 4 8 ,
5 5 7 - 5 6 8 .
L o c k r i d g e , 0 . ( 1 9 8 4 ) , I n : C h o l i n e s t e r a s e s ( B r z i n , M. e t a l . e d s )
p p . 5 - 1 2 . W a l t e r d e G r u y t e r , B e r l i n , N.Y.
L o c k r i d g e , 0 . and La Du, B.N . ( 1 9 8 2 ) , J . B . C . , 2 5 7 , 1 2 0 1 2 - 1 2 0 1 8 .
L0mo, T. and S l a t e r , C .R . ( 1 9 8 0 ) , J . P h y s i o l . , 3 0 3 , 1 9 1 - 2 0 2 .
- 1 9 8-

L o n g , J . P . ( 1 9 6 3 ) , In : C h o l i n e s t e r a s e s and A n t i c h o l i n e s t e r a s e
a g e n t s ( K o e l l e , G .B . e d . ) , p p . 3 7 4 - 4 2 7 , S p r i n g e r , B e r l i n .
L o r a n d , L. ( 1 9 7 6 ) , M e th o d s i n Enzymol. , 80^, S e c t i o n I —I V -
L o r a n d , L. ( 1 9 8 2 ) , M e th o d s i n Enzymol. , 8 0 , 6 6 4 - 6 8 0 .
Low ry , H . O . , R o s e n b r o u g h , N . J . , F a r r , A. and R a n d a l l , R J .
( 1 9 5 1 ) , J . B . C . , 1 9 3 , 2 6 5 - 2 7 5 .
L u t h e r t , P . , V r b o v a , G. and W ar d, K. ( 1 9 8 0 ) , J . N e u r o l .
N e u r o s u r . P s y . , 4 3 , 8 0 3 - 8 0 9 .
L w ebu ga-M uk asa , J . S . , L a p p i , S . and T a y l o r , P . ( 1 9 7 6 ) ,
B i o c h e m i s t r y , 15 , 1 4 2 5 - 1 4 3 4 .
L y l e s , J . M . ( 1 9 8 0 ) , P h .D . t h e s i s , " C h o l i n e s t e r a s e s i n normal and
d y s t r o p h i c c h i c k e n s " , I m p e r i a l C o l l e g e o f S c i e n c e and
T e c h n o l o g y , U n i v e r s i t y o f London.
L y l e s , J .M . and B a r n a r d , E .A . ( 1 9 8 0 ) , FEBS L e t t . , 9 - 1 1 .
L y l e s , J . M . , B a r n a r d , E .A . and S i l m a n , I . ( 1 9 8 0 ) , J . N e u r o c h e m . ,
3 4 , 9 7 8 - 9 8 7 .
L y l e s , J . M . , S i l m a n , I . and B a r n a r d , E .A . ( 1 9 7 9 ) , J .
N e u r o c h e m . , 3 3 , 7 2 7 - 7 3 8 .
L y l e s , J . M . , S i l m a n , I . , Di G i a m b e r a r d i n o , L . , C o u ra u d , J . Y .
and B a r n a r d , E.A. ( 1 9 8 2 ) , J . N e u r o c h e m . , _38_, 1 0 0 7 - 1 0 2 1 .
M c A r d l e , J . J . and A l b u q e r q u e , E .X . ( 1 9 7 3 ) , J . Gen. P h y s i o l . , 6 1 ,
1 - 2 3 .
McCaman, M.W. ( 1 9 6 6 ) , L i f e S c i . , 5_, 1 4 5 9 - 1 4 6 5 .
McMahan, U . J . , S a n e s , J . R . and M a r s h a l l , L.M. ( 1 9 7 8 ) , N a t u r e ,
2 7 1 , 1 7 2 - 1 7 4 .
M a b u c h i , K . , S z v e t k o , D . , P i n t e r , K. and S r e t e r , F . A . ( 1 9 8 2 ) ,
Am. J . P h y s i o l . , 2 4 2 , C 3 7 3 - C 3 8 1 .
M a c h n e , X. and Unna, K.R.W. ( 1 9 6 3 ) , I n : C h o l i n e s t e r a s e s and
A n t i c h o l i n e s t e r a s e a g e n t s . ( K o e l l e , G .B. e d . ) , p p . 6 7 9 - 7 0 0 ,
S p r i n g e r B e r l i n .
- 1 9 9 -

M a g a z a n i k , L . G . , F e d o r o v , V . V . , G i n i a t u l l i n , B . , N i k o l s k y , E E.
a n d S n e t k o v , V .A . ( 1 9 8 4 ) , In : C h o l i n e s t e r a s e s , ( B r z i n , M. e t
a l . e d s . ) p p . 2 2 9 - 2 4 4 . W a l t e r de G r u y t e r , B e r l i n , N .Y .
M a g a z a n i k , L . G . , F e d o r o v , V .V. a nd S n e t k o v , Y .A . ( 1 9 7 9 ) , P r o g .
B r a i n R e s . , 4 9 , 2 2 5 - 2 4 0 .
M a ld o n a d o , M . , R a m i r e z , B . U . , R u i z , G . , Yamuy, J . a nd I n e s t r o s a ,
N .C . ( 1 9 8 4 ) , J . N e u r o c h e m . , 4 3 , 3 7 5 - 3 8 1 .
M a r s h , D . , G r a s s i , J . , V i g n y , M. and M a s s o u l i e , J . ( 1 9 8 4 ) , J .
N e u r o c h e m . , 4 3 , 2 0 4 - 2 1 3 .
M a s k r e y , P . , P l u s k a l , M . G . , H a r r i s , J . B . and P e n n i n g t o n , R . J . T . ,
( 1 9 7 7 ) , J . N e u r o c h e m . , 2 8 , 4 0 3 - 4 0 9 .
M a s s o u l i e ' , J . ( 1 9 8 0 ) , TIBS, 5 , 1 6 0 - 1 6 4 .
M a s s o u l i e , J . and Bon, S . ( 1 9 7 6 ) , E u r . J . B i o c h e m . , 6 8 , 5 3 1 - 5 3 9 .
M a s s o u l i e , J . and B o n , S . ( 1 9 8 2 ) , Ann. R e v . N e u r o s c i . , 5^, 5 7 - 1 0 6 .
M a s s o u l i e , J . a nd R i e g e r , F . ( 1 9 6 9 ) , E u r . J . B i o c h e m . , 1 1 ,
4 4 1 - 4 5 5 .
M a s t r i , C . , S a l m o n s , S . and Thom as , G.H. ( 1 9 8 2 ) , B i o c h e m . J . ,
2 0 6 , 2 1 1 - 2 1 9 .
M ays , C. and R o s e n b e r r y , T .L . ( 1 9 8 1 ) , B i o c h e m i s t r y , 2 0 ,
2 8 1 0 - 2 8 1 7 .
M e t z g e r , J . M . , S c h e i d t , K .B . and F i t t s , R.H. ( 1 9 8 5 ) , J . A p p l .
P h y s i o l . , 5 8 , 1 0 8 5 - 1 0 9 1 .
M e y e r , W . L . , F i s h e r , E .H. and K r e b s , E .G . ( 1 9 6 6 ) , B i o c h e m i s t r y ,
3 , 1 0 3 3 .
M i l e d i , R. and S t e f a n i , E. ( 1 9 6 9 ) , N a t u r e , 2 2 2 , 5 6 9 - 5 7 1 .
M i l e d i , R. and Z e l e n a , J . ( 1 9 6 6 ) , N a t u r e , 2 1 0 , 8 5 5 - 8 5 6 .
M i n t z , K . P . and B r i m i j o i n , S . ( 1 9 8 5 a ) , J . N e u r o c h e m - , 4 4 ,
2 2 5 - 2 3 2 -
M i n t z , K . P . and B r i m i j o i n , S . ( 1 9 8 5 b ) , J . N e u r o c h e m . , 4 5 ,
2 8 4 - 2 9 2 .
-200-

N Nachm ans ohn , D. ( 1 9 5 9 ) , Chemical and M o l e c u l a r b a s i s o f n e r v e
a c t i v i t y , A c a d e m ic P r e s s , N.Y.
N a c h m a n s o h n , D. and R o t h e n b e r g , M .A. ( 1 9 4 4 ) , S c i e n c e , 1 0 0 ,
4 5 4 - 4 5 5 .
Nachm ansohn , D. and R o t h e n b e r g , M.A. ( 1 9 4 5 ) , J . B . C . , 1 5 8 ,
6 5 3 - 6 6 6 .
Nachm ans ohn , D. and W i l s o n , I . B . ( 1 9 5 1 ) , Adv. i n E n z y m o l 1 2 ,
2 5 9 - 3 3 9 .
Newman, J . R . , V i r g i n , J . B . , Y o u n k i n , L .H. and Y o u n k i n , S . G .
( 1 9 8 4 ) , J . P h y s i o l . , 3 5 2 , 3 0 5 - 3 1 8 .
N i e d e r l e , B. and M a y r , R. ( 1 9 7 8 ) , A n a t . E m b r y o l 1 5 3 , 9 - 2 1 .
U o t t , P . and B r o d b e c k , U. ( 1 9 8 4 ) , B . B . A . , 7 7 5 , 7 1 - 7 6 .
O t t , P . , J e n n y , B. and B r o d b e c k , U. ( 1 9 7 5 ) , E u r . J . B i o c h e m . ,
5 7 , 4 6 9 - 4 8 0 .
O t t , P . , L u s t i g , A . , B r o d b e c k , U. and R o s e n b u s c h , J . P . ( 1 9 8 2 ) ,
FEBS L e t t . , 1 3 8 , 1 8 7 - 1 8 9 .
Owens, K . , ( 1 9 7 9 ) , Ann. N Y. A c a d . S c i . , 3 1 7 , 2 4 7 - 2 6 2 .
p1 P a c ty ku la , H.A. a n d G a u t h i e r , G .F . ( 1 9 7 0 ) , J . C . B . , 4 6 , 2 7 .
P a r k , D . C . , P a r s o n s , M.E. and P e n n i n g t o n , R . J . T . , ( 1 9 7 3 ) ,
B i o c h e m . S o c . T r a n s . , U 7 3 0 - 7 3 3 .
P e a c h e y , L . D . and H u x l e y , A.F. ( 1 9 6 2 ) , J . C . B . , V3 , 1 7 7 - 1 8 0 .
P e n n i n g t o n , R . J . T . , ( 1 9 7 7 ) , In: P r o t e i n a s e s i n mammalian c e l l s
and t i s s u e s , ( B a r r e t t , A . J . e d . ) p p . 5 1 5 - 5 4 4 , N o r t h - H o l l a n d
p u b l i s h i n g Co.
P e t e r , J . B . , B a r n a r d , R . J . , E d g e r t o n , V . R . , G i l l e s p i e , C .A . and
S t e m p e l , K .E . ( 1 9 7 2 ) , B i o c h e m i s t r y , 1 1 , 2 6 2 7 - 2 6 3 3 .
-201-

P e t t e , D. ( 1 9 8 0 ) , P l a s t i c i t y o f m u s c l e . W a l t e r de G r u y t e r ,
B e r l i n .
P e t t e , D. ( 1 9 8 4 ) , M e d i c i n e and S c i e n c e i n S p o r t s and E x e r c i s e ,
1 6 , 5 1 7 - 5 2 8 .
P e t t e , D . , M u l l e r , W . , L e i s n e r , E. and V r b o v a , G. ( 1 9 7 6 ) ,
P f l i i g e r s A r c h . , 3 6 4 , 1 0 3 - 1 1 2 .
P e t t e , D . , S m i t h , M.E , S t a u d t e , H.W. and V r b o v a , G. ( 1 9 7 3 ) ,
P f l i i g e r s A r c h . , 3 3 8 , 2 5 7 - 2 7 2 .
P i z z e y , J . A . and B a r n a r d , E .A . ( 1 9 8 3 a ) , N e u r o p a t h .
A p p l . N e u r o b i o l . , j ) , 2 1 - 3 8 .
P i z z e y , J . A . and B a r n a r d , E .A . ( 1 9 8 3 b ) , N e u r o p a t h . A p p l .
N e u r o b i o l 1 4 9 - 1 6 4 .
P o d l e s k i , T . R . , A x e l r o d , D . , R a v d i n , P . , G r e e n b e r g , I . , J o h n s o n
M.M. and S a l p e t e r , M.M. ( 1 9 7 8 ) , P . N . A . S . , 7 5 , 2 0 3 5 - 2 0 3 9 .
Rab a, R . , A a v i k s a a r , A . , Raba, M. and S i i g u r , J . ( 1 9 7 9 ) , E u r . J .
B i o c h e m . , 9 6 , 1 5 1 - 1 5 8 .
R a k o n c z a y , Z . , M a l l o l , J . , S c h e n k , H . , V i n c e n d o n , G. and Z a n e t t a ,
J . P . ( 1 9 8 1 ) , B . B . A . , 6 5 7 , 2 4 3 - 2 5 6 .
R a l s t o n , J . S . , R u sh , R . S . , D o c t o r , B . P . and W o l f e , A .D . ( 1 9 8 5 ) ,
J . B . C . , 2 6 0 , ( 7 ) , 4 3 1 2 - 4 3 1 8 .
R a n d a l l , W . R . , L a i , J . and B a r n a r d , E .A . ( 1 9 8 5 ) , rM o n o c lo n a l
a n t i b o d i e s t o p u r i f i e d a c e t y l c h o l i n e s t e r a s e from a v i a n b r a i n
and t h e d i s c l o s u r e o f two a l l e l i c fo rm s o f t h e c a t a l y t i c
s u b u n i t ' Eu r . J . B i o c h e m . ( s u b m i t t e d f o r p u b l i c a t i o n ) .
R e a v i l l , C . A . , W o o s t e r , M .S . and Plummer, D .T . ( 1 9 7 8 ) , B io c h e m .
J . , 1 7 3 , 8 5 1 - 8 5 6 .
R e i m e r d e s , E .H. and K l o s t e r m e y e r , H. ( 1 9 7 6 ) , M eth od s i n E n z y m o l . ,
XLV, 2 6 - 2 8 .
-202-

Renau d, D . , l e D o u a r i n , G.H. and K h a s k i y e , A. ( 1 9 7 8 ) , Exp.
N e u r o l . , 6 0 , 1 8 0 - 2 0 0 .
R i e g e r , F . , F a i v r e - B a u m a n , A . , Bend a, P . and V i g n y , M. ( 1 9 7 6 ) ,
J . N e u r o c h e m . , 2 7 , 1 0 5 9 - 1 0 6 3 .
R i e g e r , F . , K o e n i g , J . and V i g n y , M. ( 1 9 8 0 ) , D e v . B i o l . , 7 6 ,
3 5 8 - 3 6 5 .
R i e g e r , F . and V i g n y , M. ( 1 9 7 6 ) , J . N e u r o c h e m . , 7J_t 1 2 1 - 1 2 9 .
R i t o n j a , A . , T u rk , V. and G u b e n se k , F . , ( 1 9 8 3 ) , Eu r . J .
B i o c h e m . , 1 3 3 , 4 2 7 - 4 3 2 .
Rod an, S . B . , H i n t z , R . L . , S h a a f i , R . I . and Rod an, G .A . ( 1 9 7 4 ) ,
N a t u r e , 2 5 2 , 5 8 9 - 5 9 1 .
Rom an ul , F . C . A . , S r e t e r , F . A . , S a l m o n s , S . and G e r g e l y , J .
( 1 9 7 4 ) , I n : E x p l o r a t o r y c o n c e p t s i n m u s c u l a r d y s t r o p t \ y , I I
( M i l h o r a t , A .T . e d . ) p p . 3 4 4 - 3 4 8 , E x p e r t p a M e d i c a , Amsterdam.
R o m e r - L u t h i , C . R . , O t t , P . and B r o d b e c k , U. ( 1 9 8 0 ) , B . B . A . , 6 0 1 ,
1 2 3 - 1 3 3 .
R o s e n b e r r y , T . L . ( 1 9 8 2 ) , I n : Membranes & T r a n s p o r t V o l . 2
( A . N . M a r t o n o s i e d . ) n o . 1 3 5 , pp. 3 3 9 - 3 4 8 , P lenum P r e s s ,
N . Y . , London.
R o s e n b e r r y , T . L . , B a r n e t t , P . and M a y s , C. ( 1 9 8 0 ) , N eu ro ch em .
I n t . , 2 , 1 3 5 - 1 4 8 .
R o s e n b e r r y , T . L . , C h a n g , H.W. and C h en , Y .T . ( 1 9 7 2 ) , J . B . C . , 2 4 7 ,
1 5 5 5 - 1 5 6 5 .
R o s e n b e r r y , T . L . and R i c h a r d s o n , J . M . ( 1 9 7 7 ) , B i o c h e m i s t r y , 1 6 ,
3 5 5 0 - 3 5 5 8 .
R o s e n b e r r y , T . L . and S c o g g i n , D.M. ( 1 9 8 4 ) , J . B . C . , 2 5 9 ,
5 6 4 3 - 5 6 5 2 .
R o t h , J . , B e n d a y a n , M. and O r c i , L. ( 1 9 7 8 ) , J . H i s t o c h e m .
C y t o c h e m . , 2 6 , 1 0 7 4 - 1 0 8 1 .
-203-

R o t u n d o , R .L . ( 1 9 8 4 a ) , P . N . A . S . 8 ^ , 4 7 9 - 4 8 3 .
R o tu n d o , R .L . ( 1 9 8 4 b ) , J . B . C . , 2 5 9 , 1 3 1 8 6 - 1 3 1 9 4 .
R o t u n d o , R .L . and Fambrough, D.M. ( 1 9 7 9 ) , J . B. C . , 2 5 4 ,
4 7 9 0 - 4 7 9 9 .
R o t u n d o , R .L . and Fambro ugh , D.M. ( 1 9 8 0 a ) , C e l l , 22^ 5 8 3 - 5 9 4 .
R o tu n d o , R .L . and Fambrough, D.M. ( 1 9 8 0 b ) , C e l l , 2 £ , 5 9 5 - 6 0 2 .
R u b in , L . L . , S c h u e t z e , S . M . , W e i l l , C .L . and F i s c h b a c h , 6 . D .
( 1 9 8 0 ) , N a t u r e , 2 8 3 , 2 6 4 - 2 6 7 .
R u b i n s t e i n , N . , M ab u ch i , K . , P e p e , F . , S a l m o n s , S . , G e r g e l y , J .
and S r e t e r , F . ( 1 9 7 8 ) , J . C . B . , 7 9 , 2 5 2 - 2 6 1 .
S a h e k i , T. and H o l z e r , H. ( 1 9 7 4 ) , E u r . J . B i o c h e m . , 42^, 6 2 1 - 6 2 6 .
S a l m o n s , S . ( 1 9 8 0 ) , TIN S, 3 , 1 3 4 - 1 3 7 .
S a l m o n s , S . , G a l e , D .R. and S r e t e r , F . ( 1 9 7 8 ) , J . A n a t . , 1 2 7 ,
1 7 - 3 1 .
S a l m o n s , S . , and H e n r i k s s o n , J . ( 1 9 8 1 ) , M u s c le and N e r v e , 4_,
9 4 - 1 0 5 .
S a l m o n s , S . and S r e t e r , F . A . ( 1 9 7 6 ) , N a t u r e , 2 6 3 , 3 0 - 3 4 .
S a l m o n s , S . and V rb o v a , G. ( 1 9 6 9 ) , J . P h y s i o l . , 2 1 0 , 5 3 5 - 5 4 9 .
S a n e s , J . R . ( 1 9 8 3 ) , Ann. R e v . P h y s i o l . , 4 5 , 5 8 1 - 6 0 0 .
S a n e s , J . R . and C h en ey , J . M . ( 1 9 8 2 ) , N a t u r e , 3 0 0 , 6 4 6 - 6 4 7 .
S a n e s , J . R . , F e ld m an , D . H . , C h e n e y , J . M . , and L a w r e n c e ,
J . C . J r . , ( 1 9 8 4 ) , J . N e u r o s c i . , 4_, 4 6 4 - 4 7 3 .
S a n e s , J . R . and H a l l , Z.W. ( 1 9 7 9 ) , J . C . B . , 8 3 , 3 5 7 - 3 7 0 .
S a n e s , J . R . , M a r s h a l l , L.M. and McMahan, U . J . ( 1 9 7 8 ) , J . C . B . , 7 8 ,
1 7 6 - 1 9 8 .
S a r t o r e , S . , P i e r o b o n B o r m i o l i , S . and S c h i a f f i n o , S . ( 1 9 7 8 ) ,
N a t u r e 2 7 4 , 8 2 - 8 3 .
S a u n d e r s , R . , D y c e , B . J . , V a n n i e r , W.E. and H a v e r b a c k , B . J .
( 1 9 7 1 ) , J . C l i n . I n v e s t . , 5 0 , 2 3 7 6 - 2 3 8 3 .
-204-

S c h a f f e r , N . K . , M i c h e l , H.O. and B r i d g e s , A . F . ( 1 9 7 3 ) ,
B i o c h e m i s t r y , 1 2 , 2 9 4 6 - 2 9 5 0 .
S c h e c h t e r , N .M . , F r a k i , J . E . , G e e s i n , J . C . and L a z a r u s , G . S .
( 1 9 8 3 ) , J . B . C . , 2 5 8 , 2 9 7 3 - 2 9 7 8 .
S e a l o c k , R . , Wray, B . E . and F r o e h n e r , S . C . ( 1 9 8 4 ) , J . C . B . , 9 8 ,
2 2 3 9 - 2 2 4 4 .
S e n n i , M . , B i a g i o n i , S . , M a r i n i , P . t S c a r s e l l a , G. ( 1 9 8 1 ) ,
N eu ro ch em . R e s . , 6 , 1 2 9 1 - 1 2 9 8 .
S i l m a n , I . , L y l e s , J . M . and B a r n a r d , E .A . ( 1 9 7 8 ) , FEBS L e t t . , 9 4 ,
1 6 6 - 1 7 0 .
S i l v e r , A. ( 1 9 7 4 ) , The b i o l o g y o f c h o l i n e s t e r a s e s ( N e u b e r g e r , A.
a n d Tatum, E .L . e d s . ) , N o r t h - H o l l a n d publishing Co.
S k a u , K.A . and B r i m i j o i n , S. ( 1 9 8 0 ) , J . Neurochem., 3 5 ,
1 1 5 1 - 1 1 5 4 .
S k e t e l j , J . and B r z i n , M. ( 1 9 7 9 ) , H i s t o c h e m i s t r y , 6 ^ , 2 3 9 - 2 4 8 .
S k e t e l j , J . and B r z i n , M. ( 1 9 8 0 ) , Neuroche m . R e s . , 5_, 6 5 5 - 6 6 0 .
S k e t e l j , J . , G r u b i c , Z . , K l i n a r , B. and B r z i n , M. ( 1 9 8 0 ) ,
I n : S y n a p t i c c o n s t i t u e n t s i n h e a l t h and d i s e a s e ( B r z i n , M. e t a l
e d s ) p p . 586 Perga m on , O x f o r d .
S k e t e l j , J . , McNamee, M.G. and W i l s o n , B.W. ( 1 9 7 8 ) , Exp.
N e u r o l . , 6 0 , 6 2 4 - 6 2 9 .
S o r e n s e n , K . , G e n t i n e t t a , R. and B r o d b e c k , U. ( 1 9 8 2 ) , J .
N e u r o c h e m . , 3 9 , 1 0 5 0 - 1 0 6 0 .
S t a r k e y , P .M. and B a r r e t t , A . J . ( 1 9 7 7 ) , I n : P r o t e i n a s e s i n
mammalian c e l l s and t i s s u e s . ( B a r r e t t , A . J . e d . ) p p . 6 6 3 - 6 9 6 .
N o r t h - H o l l a n d P u b l i s h i n g Co .
S te d m a n , Edgar a nd St ed m a n , E l l e n , ( 1 9 3 5 ) , B io c h e m . J . , 2 9 ,
2 1 0 7 - 2 1 1 1 .
-205-

S te d m a n , E d g a r , St ed m a n , E l l e n , a nd E a s s o n , L.H. ( 1 9 3 2 ) ,
B i o c h e m . J . , 2 6 , 2 0 5 6 - 2 0 6 6 .
S t e r n b e r g e r , L . A . ( 1 9 7 9 ) , I m m u n o c y t o c h e m i s t r y 2 n d . e d . Jo h n
W i l e y , N.Y.
S t e r z , R . , P a g a l e , M. and P e p e r , K. ( 1 9 8 3 ) , P f l t i g e r s A r c h . , 3 9 8 ,
4 8 - 5 4 .
S t r a c h e r , A . , McGowan, E . B . and S h a f i g , S . A . ( 1 9 7 7 ) , S c i e n c e ,
2 0 0 , 5 0 - 5 1 .
S u g i y a m a , H. ( 1 9 7 7 ) , FEBS L e t t . , 8 4 , 2 5 7 - 2 6 0 .
S u n g , S . C . a nd R u f f , B . A . ( 1 9 8 3 ) , Neurochem . R e s . , 8 , 3 0 3 - 3 1 1 .
S y r o v y , I . , Gutmann, E. and M e l i c h n a , J . ( 1 9 7 1 ) , E x p e r i e n t i a , 2 7 ,
1 4 2 6 - 1 4 2 7 .
T1 T am m el in , L . E . ( 1 9 5 7 ) , A c ta Chem. S c a n d . , 11_, 1 3 4 0 - 1 3 4 9 .
T a y l o r , P . B . , R i e g e r , F . , S h e l a n s k i , M.L. a n d G r e e n e , L .A.
( 1 9 8 1 ) , J . B . C . , 2 5 6 , 3 8 2 7 - 3 8 3 0 .
T e n n y s o n , V . M . , K re m zn er , L .T . and B r z i n , M. ( 1 9 7 7 ) , J .
N e u r o p a t h . E xp . N e u r o l . , 3 6 , 2 4 5 - 2 7 5 .
T h e s l e f f , S . ( 1 9 7 4 ) , Ann. N.Y. A ca d . S c i . , 2 2 8 , 8 9 - 1 0 4 .
T o m im a tsu , Y . , C l a r y , J . J . and B a r t u l o v i c h , J . J . ( 1 9 6 6 ) , A rch.
B io c h e m . B i o p h y s . , U 5 , 5 3 6 - 5 4 4 .
T o n g e , D.A. ( 1 9 7 4 ) , J . P h y s i o l . , 2 4 1 , 1 2 7 - 1 3 9 .
T o r r e s , J . C . a n d I n e s t r o s a , N .C . ( 1 9 8 3 ) , FEBS L e t t . , 1 5 4 ,
2 6 5 - 2 6 8 .
T o u t a n t , J . P . , T o u t a n t , M . , F i s z m a n , M.Y. and M a s s o u l i e , J .
( 1 9 8 3 ) , Neur oche m . I n t . , 5 , 7 5 1 - 7 6 2 .
T o w e r s , S . S . ( 1 9 3 5 ) , Am. J . A n a t . , 5 6 , 1 - 4 3 .
T w i n i n g , S . S . ( 1 9 8 3 ) , F ed . P r o c . , 4 2 , 1 9 5 1 ( a b s . 1 1 3 2 ) .
-206-

Umezawa, H. a nd A o y a g i , T. ( 1 9 7 7 ) , I n : P r o t e i n a s e s i n mammalian
c e l l s and t i s s u e s , ( B a r r e t t , A . J . e d . ) , p p . 6 5 3 - 6 5 4 .
N o r t h - H o l l a n d P u b l i s h i n g Co .
U n g a r , G. a n d Damgaard, E. ( 1 9 5 5 ) , J . Exp. M e d . , 1 0 1 , 1 - 1 5 . ,
V a l l e t t e , F . M . , M a r sh , D . J . , M u l l e r , F . and M a s s o u l i e , J . ( 1 9 8 3 ) ,
J . - C h r o m a t o g r a p h y . , 2 5 7 , 2 8 5 - 2 9 6 .
V i g n y , M . , Bon , S . , M a s s o u l i e , J and G i s i g e r , V. ( 1 9 7 9 ) , J .
N e u r o c h e m . , 3 3 , 5 5 9 - 5 6 5 .
V ig n y M . , K o e n i g , J . and R i e g e r , F . ( 1 9 7 6 ) , J . o f N e u r o c h e m . , 2 7 ,
1 3 4 7 - 1 3 5 3 .
V i g n y , M . , M a r t i n , G .R. and G r o t e n d o r s t , G.R. ( 1 9 8 2 ) , J . B . C . ,
2 5 8 , 8 7 9 4 - 8 7 9 8 .
V i r a t e l l e , O.M. and B e r n h a r d , S . A . ( 1 9 8 0 ) , B i o c h e m i s t r y , 1 9 ,
4 9 9 9 - 5 0 0 7 .
V r b o v a , G . , G o rd o n , T. and J o n e s , R. ( 1 9 7 8 ) , N e r v e m u s c l e
i n t e r a c t i o n . Chapman a n d H a l l , London.
V r b o v a , G. and Ward, K. ( 1 9 8 1 ) , J . N e u r o l . N e u r o s u r . P s y . , 4 4 ,
1 0 0 2 - 1 0 0 6 .
W W a l k e r , C R. and S t r o h m a n , R .C . ( 1 9 7 6 ) , J . C . B . , 7 0 , A 1 1 7 .
W a l k e r , C .R . and W i l s o n , B.W. ( 1 9 7 5 ) , N a t u r e , 2 5 6 , 2 1 5 - 2 1 6 .
W a l k e r , C .R . a n d W i l s o n , B.W. ( 1 9 7 6 ) , N e u r o s c i . , 1_, 1 9 1 - 1 9 6 .
W a l l a c e , B . G . , N i t k i n , R . M . , R e i s t , N . E . , F a l l o n , J . R . , M o a y e r i ,
N.N and McMahan, U . J . ( 1 9 8 5 ) , N a t u r e , 3 1 5 , 5 7 4 - 5 7 7 .
W a r n i c k , J . E . , L e b e d a , F . J . a n d A l b u q u e r q u e , E . X . ( 1 9 7 9 ) , Ann.
N .Y . A c a d . S c i . , 3 1 7 , 2 6 3 - 2 8 5 .
W e i n b e r g , C .B . and H a l l , Z.W. ( 1 9 7 9 ) , Dev . B i o l . , 6 8 , 6 3 1 - 6 3 5 .
W ied m er , T . , F r a n c e s c o , C . and B r o d b e c k , U. ( 1 9 7 9 ) , Eur, J .
B i o c h e m . , 1 0 2 , 5 9 - 6 4 .
-207-

W i l s o n , B . W . , M e t t l e r , M.A. and Asmund son, R .V . ( 1 9 6 9 ) , J . Exp.
Z o o l . , 1 7 2 , 4 9 - 5 8 .
W i l s o n , B . W . , N i e b e r g , P . S . , W a l k e r , C . R . , L i n k h a r t , T .A . and
F r y , D.M. ( 1 9 7 3 ) , D ev . B i o l . , 3 3 , 2 8 5 - 2 9 9 .
W i l s o n , B . W . , R a n d a l l , W . R . , P a t t e r s o n , G.T. and E n t r i k i n , R.K.
( 1 9 7 9 ) , Ann. N.Y. A cad. S c i . , 3 1 7 , 2 2 4 - 2 4 6 .
W i l s o n , B.W. and W a l k e r , C .R . ( 1 9 7 4 ) , P . N . A . S . , 7 1 , 3 1 9 4 - 3 1 9 8 .
W it z e m a n n , Y. ( 1 9 8 0 ) , N e u r o s c i . L e t t : , 2 0 , 2 7 7 - 2 8 2 .
W ood bu ry, R . G . , E v e r i t t , M .T. and N e u r a t h , H. ( 1 9 8 1 ) , M e th o d s i n
E n z y m o l . , 8 0 , 5 8 8 - 6 0 9 .
Yamamura, H . L . , R e i c h a r d , D . W . , G a r d n e r , T . L . , M o r r i s e t t , J . D .
and B r o o m f i e l d , C .A . ( 1 9 7 3 ) , B . B . A . , 3 0 2 , 3 0 5 - 3 1 5 .
Y e l l i n , H. ( 1 9 6 9 ) , Exp. N e u r o l . , 2 5 , 1 5 3 - 1 6 3 .
Y o s h i k a w a , A . , Takano-Ohm uro, H. and M a s a k i , T . ( 1 9 8 4 ) , M u s c l e
an d N e r v e , 7_, 7 3 3 - 7 4 0 .
Y o u n k i n , S . G . , B r e t t , R . S . , D a v e y , B. and Y o u n k i n , L .H. ( 1 9 7 8 ) ,
S c i e n c e , 2 0 0 , 1 2 9 2 - 1 2 9 5 . ,
Y o u n k i n , S . G . , R o s e n s t e i n , C . , C o l l i n s , P . L . and R o s e n b e r r y , T . L .
( 1 9 8 2 ) , J . B . C . , 2 5 7 , 1 3 6 3 0 - 1 3 6 3 7 .
Z a n e t t a , J . P * , R a c k o n c z a y , Z . , R e e b e r , A . , K a s a , P . and
V i n c e n d o n , G. ( 1 9 8 1 ) , FEBS L e t t . , 1 2 9 , 2 9 3 - 2 9 6 .
Z e l e n a , J . and J i r m a n o v a , I . ( 1 9 7 3 ) , Exp. N e u r o l . , 3 8 , 2 7 2 - 2 8 5 .
Z i s k i n d - C o n h a i m , L . , I n e s t r o s a , N.C. a n d H a l l , Z.W. ( 1 9 8 4 ) ,
D e v . B i o l . , 1 0 3 , 3 6 9 - 3 7 7 .
-208-

List of publications
1 . E .A . B a r n a r d , P . J . B a r n a r d , J . J a r v i s , J . J e d r z e j c z y k , J . L a i , J . A . P i z z e y , W. R a n d a l l and I . S i lm a n ( 1 9 8 4 ) M u l t i p l e m o l e c u l a r f o r m s o f a c e t y l c h o l i n e s t e r a s e and t h e i r r e l a t i o n s h i p t o m u s c l e f u n c t i o n . In C h o l i n e s t e r a s e s (M. B r z i n e t a l . e d s . ) , W a l t e r de G r u y t e r , B e r l i n - N e w Y o r k , p p . 4 9 - 7 1 .
2 . J . J e d r z e j c z y k , I . S i l m a n , J . L a i , E .A . B arn ard ( 1 9 8 4 ) M o l e c u l a r f o r m s o f a c e t y l c h o l i n e s t e r a s e i n s y n a p t i c and e x t r a s y n a p t i c r e g i o n s o f a v i a n t o n i c m u s c l e . N e u r o s c i . L e t t . , 46_, 2 8 3 - 2 8 9 .
3 . E .A . B a r n a r d , J . Lai and J . P i z z e y ( 1 9 8 4 ) S y n a p t i c and e x t r a s y n a p t i c fo rm s o f a c e t y l c h o l i n e s t e r a s e i n s k e l e t a l m u s c l e s : v a r i a t i o n w i t h f i b r e t y p e and f u n c t i o n a l c o n s i d e r a t i o n s . In N e u r o m u s c u l a r D i s e a s e s (G. S e r r a t r i c e e t a l . e d s ) , Raven P r e s s , New Y o r k , p p . 4 5 5 - 4 6 3 .
4 . W.R. R a n d a l l , J . Lai and E .A . Bar nar d ( 1 9 8 5 ) A c e t y l c h o l i n e s t e r a s e o f m u s c l e and n e r v e . In M o l e c u l a r B a s i s o f N erveA c t i v i t y ( J . P . Changeux e t a l . e d s . ) , W a l t e r de G r u y t e r , B e r l i n - New Y o r k , p p . 5 9 5 - 6 1 7 .
5 . E .A . B a r n a r d , P . J . B a r n a r d , J . C . J a r v i s and J . Lai ( 1 9 8 5 )C ha nge s i n m o l e c u l a r fo r m s o f a c e t y l c h o l i n e s t e r a s e i nc h r o n i c a l l y s t i m u l a t e d normal and d y s t r o p h i c c h i c k e n m u s c l e . J . P h y s i o l . , s u b m i t t e d f o r p u b l i c a t i o n .
6 . J . L a i , J . J e d r z e j c z y k , J . A . P i z z e y , D. G reen and E . A . Ba rna rd( 1 9 8 5 ) N e u r o n a l c o n t r o l o f t h e fo rm s o f AChE i n s l o w mammalian m u s c l e s . N a t u r e , s u b m i t t e d f o r p u b l i c a t i o n .
7 . W.R. R a n d a l l , J . Lai and E .A . B arn ard ( 1 9 8 5 ) M o n o c lo n a l a n t i b o d i e s t o p u r i f i e d a c e t y l c h o l i n e s t e r a s e from a v i a n b r a i n and t h e d i s c l o s u r e o f tw o a l l e l i c fo rm s o f t h e c a t a l y t i c s u b u n i t . E u r . J . B i o c h e m . , s u b m i t t e d f o r p u b l i c a t i o n .
8 . J . L a i , J . P i z z e y and E .A . Barnar d ( 1 9 8 3 ) AChE i n t h e d i f f e r e n t f i b r e t y p e s o f mammalian m u s c l e . I . R e l a t i o n s h i p o f AChE m o l e c u l a r for m s t o t h e f i b r e t y p e s . In: S e c o n d I n t e r n a t i o n a l M e e t i n g on C h o l i n e s t e r a s e s , B l e d , Y u g o s l a v i a .
9 . J . L a i , J . P i z z e y , D. Green and E .A . B a r n a r d ( 1 9 8 3 ) AChE i n t h ed i f f e r e n t f i b r e t y p e s o f mammalian m u s c l e . I I . E f f e c t s o f d e n e r v a t i o n . In: S e c o n d I n t e r n a t i o n a l M e e t i n g onC h o l i n e s t e r a s e s , B l e d , Y u g o s l a v i a .
10 J . C . J a r v i s , J . Lai and P . J . Ba rn ard ( 1 9 8 3 ) AChE i n s t i m u l a t e d f a s t - t w i t c h a v i a n m u s c l e . In: S eco n d I n t e r n a t i o n a l M e e t i n g on C h o l i n e s t e r a s e s , B l e d , Y u g o s l a v i a .
-209-


















