
The migration patterns of the European flounder Platichthys flesus (Linnaeus, 1758) (Pleuronectidae,...
12
The migration patterns of the European flounder Platichthys flesus (Linnaeus, 1758) (Pleuronectidae, Pisces) at the southern limit of its distribution range: Ecological implications and fishery management Pedro Morais a,b, ⁎ ,1 , Ester Dias b,1 , John Babaluk c , Carlos Antunes b,d a ICCE — International Center for Coastal Ecohydrology, Palácio do Capitão-Mor, Horta das Figuras, EN 125, 8005-518 Faro, Portugal b CIMAR/CIIMAR — Centro Interdisciplinar de Investigação Marinha e Ambiental, Universidade do Porto, Rua dos Bragas 289, 4050-123 Porto, Portugal c Fisheries and Oceans Canada, 501 University Crescent, Winnipeg, Manitoba R3T 2N6, Canada d Aquamuseu do Rio Minho, Parque do Castelinho s/n, 4920-290 Vila Nova de Cerveira, Portugal abstract article info Article history: Received 4 February 2010 Received in revised form 13 October 2010 Accepted 4 November 2010 Available online 24 November 2010 Keywords: Platichthys flesus Temperate estuary Migration Strontium Otolith microchemistry LA-ICP-MS This study aims to determine the diversity of migration patterns of the European flounder (Platichthys flesus (Linnaeus, 1758)) present in the Minho estuary and in the adjacent coastal area (NW-Iberian Peninsula). Assessing the diversity of flounder migration patterns at the southern limit of its distribution allows the determination of characteristics of the species' ecology and provides useful information for fishery managers, since it is a regionally important estuarine fishery. An unexpected result of our study was that flounder appears to spawn in both estuarine and coastal areas and not just in coastal areas as was previously widely accepted. Our interpretation of otolith strontium distribution patterns from flounder specimens collected in the freshwater tidal area of the Minho estuary and in the lower estuary suggested that the flounders hatched in the estuary, while only 6.7% of those captured in the coastal area hatched in the coastal area. Ultimately, studies aimed at collecting larval stages and adult flounders must be made to confirm that flounders spawn in the estuary and to define new and better scientifically supported fishing policies, or simply to confirm the existing ones regarding temporal and spatial closures for each gear used in the Minho estuary. © 2010 Elsevier B.V. All rights reserved. 1. Introduction The European flounder (Platichthys flesus Linnaeus, 1758) (Pleur- onectidae, Pisces) is a demersal species, with its distribution range along the NE-Atlantic Ocean, from Norway to Portugal, both in coastal and estuarine ecosystems. It is also present in the adjacent seas, such as the Baltic, North and White seas and in the Mediterranean and Black seas (Nielsen, 1986). Between 1950 and 2006, the reported landings of this species increased from 9700 to 22,379 tonnes and varied between 7407 (1970) and 24,461 tonnes (2005) (FAO, 2008). In 2006, Poland (42.1%), Netherlands (18.0%) and Denmark (15.1%) accounted for 75.2% of the total catch (22,739 tonnes) (FAO, 2008). These three countries are located along the northern distribution range of flounder, while in the southern range of the species (Cabral et al., 2001), Portugal and Spain accounted for only 0.06% and 0.18%, respectively (FAO, 2008). However, flounder is relevant for local artisanal fisheries in northern Portuguese estuaries and captures there show a slight increasing trend (Teixeira and Cabral, 2009). Some of these estuaries were already identified as important nursery grounds for flounder, in particular Ria de Aveiro lagoon and Mondego, Douro, Lima and Minho estuaries (Cabral et al., 2007; Vasconcelos et al., 2008; Freitas et al., 2009; Ramos et al., 2010). Among these, the Minho estuary has the highest density of flounder juveniles, 293 ind. 1000 m −2 (Freitas et al., 2009), which is 32.2, 12.6, 9.8 and 6.6 times greater than the maximum densities registered in Douro estuary (Vinagre et al., 2005), Ria de Aveiro lagoon (Pombo et al., 2007), Mondego estuary (Martinho et al., 2007) and Lima estuary (Ramos et al., 2010), respectively. It was established (Summers, 1979), and then widely accepted for other ecosystems (e.g. Borsa et al., 1997; Minier et al., 2000; Martinho et al., 2007; Froese and Pauly, 2009), that flounder spawn in marine waters during winter and early spring, then move to estuarine and other shallow waters (near-shore coastal areas and coastal lagoons) for feeding. Larvae and young-of-the-year also move into estuaries. These juveniles remain in estuaries until the end of autumn before dispersing to the coast. However, this might not be the case for the entire distribution range of this species as differences have been observed in migration patterns in the Minho estuary (NW-Portugal; SW-Galicia, Spain), where juvenile and/or adult specimens were caught 70 km from the river mouth (Antunes, unpublished data). Journal of Sea Research 65 (2011) 235–246 ⁎ Corresponding author. ICCE — International Center for Coastal Ecohydrology, Palácio do Capitão-Mor, Horta das Figuras, EN 125, 8005-518 Faro, Portugal. Tel.: +351 289800900x7263; fax: +351 289800069. E-mail address: [email protected] (P. Morais). 1 The first two authors made equal contributions for this work. 1385-1101/$ – see front matter © 2010 Elsevier B.V. All rights reserved. doi:10.1016/j.seares.2010.11.001 Contents lists available at ScienceDirect Journal of Sea Research journal homepage: www.elsevier.com/locate/seares
-
Upload
pedro-morais -
Category
Documents
-
view
213 -
download
0
Transcript of The migration patterns of the European flounder Platichthys flesus (Linnaeus, 1758) (Pleuronectidae,...

Journal of Sea Research 65 (2011) 235–246
Contents lists available at ScienceDirect
Journal of Sea Research
j ourna l homepage: www.e lsev ie r.com/ locate /seares
The migration patterns of the European flounder Platichthys flesus (Linnaeus, 1758)(Pleuronectidae, Pisces) at the southern limit of its distribution range: Ecologicalimplications and fishery management
Pedro Morais a,b,⁎,1, Ester Dias b,1, John Babaluk c, Carlos Antunes b,d
a ICCE — International Center for Coastal Ecohydrology, Palácio do Capitão-Mor, Horta das Figuras, EN 125, 8005-518 Faro, Portugalb CIMAR/CIIMAR — Centro Interdisciplinar de Investigação Marinha e Ambiental, Universidade do Porto, Rua dos Bragas 289, 4050-123 Porto, Portugalc Fisheries and Oceans Canada, 501 University Crescent, Winnipeg, Manitoba R3T 2N6, Canadad Aquamuseu do Rio Minho, Parque do Castelinho s/n, 4920-290 Vila Nova de Cerveira, Portugal
⁎ Corresponding author. ICCE — International CenPalácio do Capitão-Mor, Horta das Figuras, EN 125, 8005289800900x7263; fax: +351 289800069.
E-mail address: [email protected] (P. Morais).1 The first two authors made equal contributions for
1385-1101/$ – see front matter © 2010 Elsevier B.V. Aldoi:10.1016/j.seares.2010.11.001
a b s t r a c t
a r t i c l e i n f oArticle history:Received 4 February 2010Received in revised form 13 October 2010Accepted 4 November 2010Available online 24 November 2010
Keywords:Platichthys flesusTemperate estuaryMigrationStrontiumOtolith microchemistryLA-ICP-MS
This study aims to determine the diversity of migration patterns of the European flounder (Platichthys flesus(Linnaeus, 1758)) present in the Minho estuary and in the adjacent coastal area (NW-Iberian Peninsula).Assessing the diversity of flounder migration patterns at the southern limit of its distribution allows thedetermination of characteristics of the species' ecology and provides useful information for fishery managers,since it is a regionally important estuarine fishery. An unexpected result of our study was that flounderappears to spawn in both estuarine and coastal areas and not just in coastal areas as was previously widelyaccepted. Our interpretation of otolith strontium distribution patterns from flounder specimens collected inthe freshwater tidal area of the Minho estuary and in the lower estuary suggested that the flounders hatchedin the estuary, while only 6.7% of those captured in the coastal area hatched in the coastal area. Ultimately,studies aimed at collecting larval stages and adult flounders must be made to confirm that flounders spawn inthe estuary and to define new and better scientifically supported fishing policies, or simply to confirm theexisting ones regarding temporal and spatial closures for each gear used in the Minho estuary.
ter for Coastal Ecohydrology,-518 Faro, Portugal. Tel.: +351
this work.
l rights reserved.
© 2010 Elsevier B.V. All rights reserved.
1. Introduction
The European flounder (Platichthys flesus Linnaeus, 1758) (Pleur-onectidae, Pisces) is a demersal species, with its distribution rangealong the NE-Atlantic Ocean, from Norway to Portugal, both in coastaland estuarine ecosystems. It is also present in the adjacent seas, suchas the Baltic, North and White seas and in the Mediterranean andBlack seas (Nielsen, 1986).
Between 1950 and 2006, the reported landings of this speciesincreased from 9700 to 22,379 tonnes and varied between 7407(1970) and 24,461 tonnes (2005) (FAO, 2008). In 2006, Poland(42.1%), Netherlands (18.0%) and Denmark (15.1%) accounted for75.2% of the total catch (22,739 tonnes) (FAO, 2008). These threecountries are located along the northerndistribution range offlounder,while in the southern range of the species (Cabral et al., 2001), Portugaland Spain accounted for only 0.06% and 0.18%, respectively (FAO,2008). However, flounder is relevant for local artisanal fisheries in
northern Portuguese estuaries and captures there show a slightincreasing trend (Teixeira and Cabral, 2009). Some of these estuarieswere already identified as important nursery grounds for flounder, inparticular Ria de Aveiro lagoon and Mondego, Douro, Lima and Minhoestuaries (Cabral et al., 2007; Vasconcelos et al., 2008; Freitas et al.,2009; Ramos et al., 2010). Among these, the Minho estuary has thehighest density of flounder juveniles, 293 ind. 1000 m−2 (Freitas et al.,2009),which is 32.2, 12.6, 9.8 and 6.6 times greater than themaximumdensities registered in Douro estuary (Vinagre et al., 2005), Ria deAveiro lagoon (Pombo et al., 2007), Mondego estuary (Martinho et al.,2007) and Lima estuary (Ramos et al., 2010), respectively.
It was established (Summers, 1979), and then widely accepted forother ecosystems (e.g. Borsa et al., 1997; Minier et al., 2000; Martinhoet al., 2007; Froese and Pauly, 2009), that flounder spawn in marinewaters during winter and early spring, then move to estuarine andother shallow waters (near-shore coastal areas and coastal lagoons)for feeding. Larvae and young-of-the-year also move into estuaries.These juveniles remain in estuaries until the end of autumn beforedispersing to the coast. However, this might not be the case for theentire distribution range of this species as differences have beenobserved in migration patterns in the Minho estuary (NW-Portugal;SW-Galicia, Spain), where juvenile and/or adult specimens werecaught 70 km from the river mouth (Antunes, unpublished data).

236 P. Morais et al. / Journal of Sea Research 65 (2011) 235–246
In the Baltic Sea, the largest brackish ecosystem in the world, theEuropean flounder that colonized it after the last glacial period(10,000 years BP) evolved to cope with the low salinity conditions ofthe area. Surface salinity varies between 3 and 8 in the northern andsouthern Baltic Sea, respectively, and there is a strong halocline in thedeepest basins (south and central Baltic Sea), where salinity variesbetween 10 and 20 (Florin andHöglund, 2008 and references therein). Inthe Baltic Sea there are two sympatric flounder populations using twodifferent spawning habitats (Florin and Höglund, 2008). One populationproduces pelagic eggs mainly in areas of higher salinities and in thedeepestbasins,while theotherproducesdemersal eggs in lowsalinities incoastal areas and on banks (Nissling et al., 2002). There are also migra-tionsbetweensomeof theseareas, sinceflounders share the same feedingcoastal areas during summer–autumn (Florin and Höglund, 2008).
Instead of using traditional tagging and recapturing methods, themigration patterns of fishes can also be assessed by analyzing tracechemical elements incorporated in the sagittal otoliths of fish(Campana, 1999; Campbell et al., 2002; Gillanders, 2005). The traceelements are incorporated in the otolith proportionally to theirconcentration in the ambient water, as a function of fish metabolism,or dependent of their food items (Campana, 1999). The absorptionpathways of inorganic elements are through branchial uptake orintestinal assimilation into the plasma and then to the crystallizingotolith (Campana, 1999). As a result, otoliths are particularly suitableto delineate stocks, to reconstruct environmental history and to infermigration patterns (Campana, 2005). For example, strontium (Sr)concentration distribution in otoliths has been used to trackmigrationpatterns along salinity gradients (Edmonds et al., 1999; Elfman et al.,1999, 2000; Zlokovitz et al., 2003). The general assumption is thatthe concentration of Sr is substantially higher in seawater than inestuaries or freshwater ecosystems (Secor and Rooker, 2000; Krausand Secor, 2004; Elsdon and Gillanders, 2005). In this study, we usedlaser ablation-inductively coupled plasma-mass spectrometry (LA-ICP-MS) analysis to measure and determine the pattern of Srdistribution in otoliths of European flounder from the southern limit
Fig. 1. Sampling areas within the Minho estuary (Portugal/Spa
of its distribution range (Minho estuary and in the adjacent coastalarea, Portugal/Spain) to assess the diversity of the species' migrationpatterns in the area. Moreover, the resultant analysis of floundermigration patterns can reveal more characteristics of flounder ecologyand provide useful information for fishery managers.
2. Material and methods
2.1. Study area
The Minho estuary and its adjacent coastal area, located in theNW-Iberian Peninsula (Europe) (Fig. 1), occupy 23 km2, with only 9%of it composed of intertidal areas. It is a mesotidal estuary, with tidesvarying between 0.7 m and 3.7 m (Alves, 1996), and it tends to form asalt wedge near the mouth of the estuary during periods of high riverflow (Sousa et al., 2005). The limit of tide penetration is about 40 kminland, with the uppermost 30 km a tidal freshwater wetland (Sousaet al., 2008). Themean depth of the estuary is 2.6 m and themaximumdepth is approximately 26 m. Average annual freshwater inflow rateis 300 m3 s−1 (Ferreira et al., 2003).
The coastal area adjacent to the Minho estuary has a narrowcontinental platform (Palenzuela et al., 2004), which varies between30 km and 75 km in width, and is influenced by the NW-Iberianupwelling system, the northernmost limit of the Eastern NorthAtlantic upwelling system (Alvarez et al., 2008, and referencestherein). The coastal upwelling occurs during spring–summer and isaccompanied by favorable southward winds along the coast, althoughit sometimes occurs during winter. The summer upwelling introducesnutrient-rich and cold deep waters into the coastal estuarineecosystems (Alvarez et al., 2008, and references therein).
2.2. Sample collection and otolith preparation
Juvenile flounder were collected from the Minho estuary betweenOctober 2007 and July 2008 using a beam trawl. Two sampling areas
in) (SA1 and SA2) and in the adjacent coastal area (SA3).
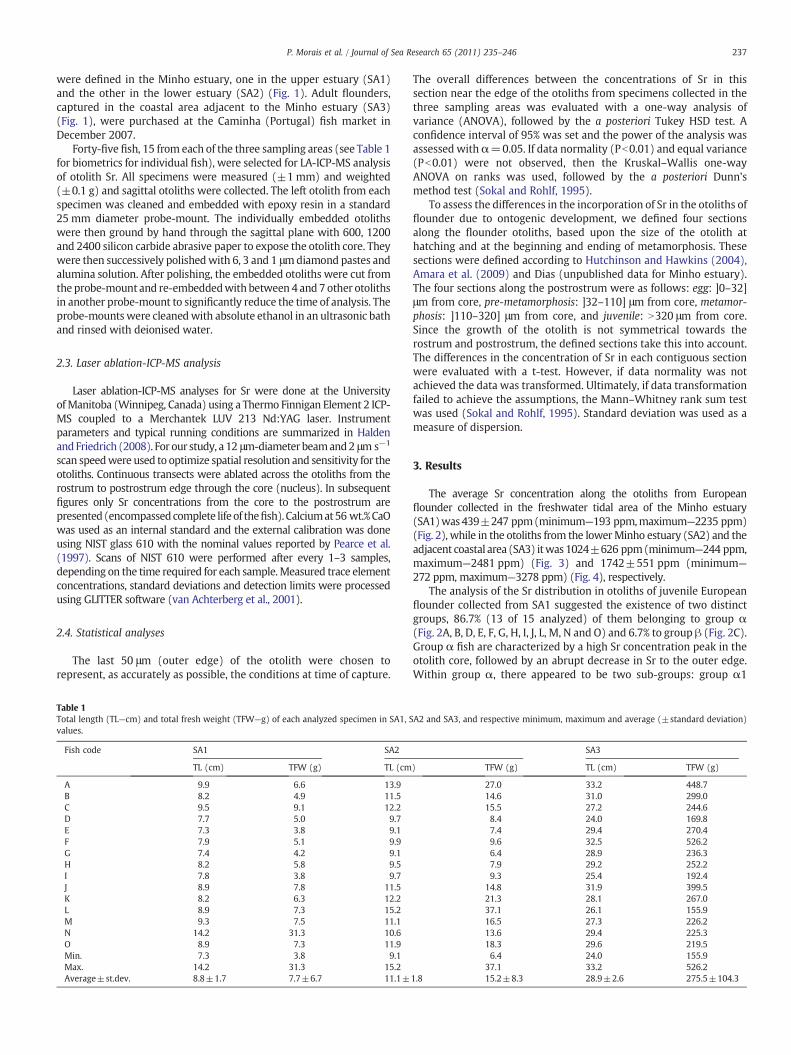
237P. Morais et al. / Journal of Sea Research 65 (2011) 235–246
were defined in the Minho estuary, one in the upper estuary (SA1)and the other in the lower estuary (SA2) (Fig. 1). Adult flounders,captured in the coastal area adjacent to the Minho estuary (SA3)(Fig. 1), were purchased at the Caminha (Portugal) fish market inDecember 2007.
Forty-five fish, 15 from each of the three sampling areas (see Table 1for biometrics for individual fish), were selected for LA-ICP-MS analysisof otolith Sr. All specimens were measured (±1 mm) and weighted(±0.1 g) and sagittal otoliths were collected. The left otolith from eachspecimen was cleaned and embedded with epoxy resin in a standard25 mm diameter probe-mount. The individually embedded otolithswere then ground by hand through the sagittal plane with 600, 1200and 2400 silicon carbide abrasive paper to expose the otolith core. Theywere then successively polishedwith 6, 3 and 1 μmdiamond pastes andalumina solution. After polishing, the embedded otoliths were cut fromthe probe-mount and re-embeddedwith between 4 and 7 other otolithsin another probe-mount to significantly reduce the time of analysis. Theprobe-mountswere cleanedwith absolute ethanol in an ultrasonic bathand rinsed with deionised water.
2.3. Laser ablation-ICP-MS analysis
Laser ablation-ICP-MS analyses for Sr were done at the UniversityofManitoba (Winnipeg, Canada) using a Thermo Finnigan Element 2 ICP-MS coupled to a Merchantek LUV 213 Nd:YAG laser. Instrumentparameters and typical running conditions are summarized in HaldenandFriedrich (2008). For our study, a 12 μm-diameter beamand2 μm s−1
scan speedwere used to optimize spatial resolution and sensitivity for theotoliths. Continuous transects were ablated across the otoliths from therostrum to postrostrum edge through the core (nucleus). In subsequentfigures only Sr concentrations from the core to the postrostrum arepresented(encompassedcomplete life of thefish). Calciumat56 wt.%CaOwas used as an internal standard and the external calibration was doneusing NIST glass 610 with the nominal values reported by Pearce et al.(1997). Scans of NIST 610 were performed after every 1–3 samples,depending on the time required for each sample.Measured trace elementconcentrations, standard deviations and detection limits were processedusing GLITTER software (van Achterberg et al., 2001).
2.4. Statistical analyses
The last 50 μm (outer edge) of the otolith were chosen torepresent, as accurately as possible, the conditions at time of capture.
Table 1Total length (TL—cm) and total fresh weight (TFW—g) of each analyzed specimen in SA1, Svalues.
Fish code SA1 SA2
TL (cm) TFW (g) TL (cm
A 9.9 6.6 13.9B 8.2 4.9 11.5C 9.5 9.1 12.2D 7.7 5.0 9.7E 7.3 3.8 9.1F 7.9 5.1 9.9G 7.4 4.2 9.1H 8.2 5.8 9.5I 7.8 3.8 9.7J 8.9 7.8 11.5K 8.2 6.3 12.2L 8.9 7.3 15.2M 9.3 7.5 11.1N 14.2 31.3 10.6O 8.9 7.3 11.9Min. 7.3 3.8 9.1Max. 14.2 31.3 15.2Average±st.dev. 8.8±1.7 7.7±6.7 11.1±
The overall differences between the concentrations of Sr in thissection near the edge of the otoliths from specimens collected in thethree sampling areas was evaluated with a one-way analysis ofvariance (ANOVA), followed by the a posteriori Tukey HSD test. Aconfidence interval of 95% was set and the power of the analysis wasassessed withα=0.05. If data normality (Pb0.01) and equal variance(Pb0.01) were not observed, then the Kruskal–Wallis one-wayANOVA on ranks was used, followed by the a posteriori Dunn'smethod test (Sokal and Rohlf, 1995).
To assess the differences in the incorporation of Sr in the otoliths offlounder due to ontogenic development, we defined four sectionsalong the flounder otoliths, based upon the size of the otolith athatching and at the beginning and ending of metamorphosis. Thesesections were defined according to Hutchinson and Hawkins (2004),Amara et al. (2009) and Dias (unpublished data for Minho estuary).The four sections along the postrostrum were as follows: egg: ]0–32]μm from core, pre-metamorphosis: ]32–110] μm from core, metamor-phosis: ]110–320] μm from core, and juvenile: N320 μm from core.Since the growth of the otolith is not symmetrical towards therostrum and postrostrum, the defined sections take this into account.The differences in the concentration of Sr in each contiguous sectionwere evaluated with a t-test. However, if data normality was notachieved the data was transformed. Ultimately, if data transformationfailed to achieve the assumptions, the Mann–Whitney rank sum testwas used (Sokal and Rohlf, 1995). Standard deviation was used as ameasure of dispersion.
3. Results
The average Sr concentration along the otoliths from Europeanflounder collected in the freshwater tidal area of the Minho estuary(SA1)was439±247 ppm(minimum—193 ppm,maximum—2235 ppm)(Fig. 2), while in the otoliths from the lowerMinho estuary (SA2) and theadjacent coastal area (SA3) itwas1024±626 ppm(minimum—244 ppm,maximum—2481 ppm) (Fig. 3) and 1742±551 ppm (minimum—
272 ppm, maximum—3278 ppm) (Fig. 4), respectively.The analysis of the Sr distribution in otoliths of juvenile European
flounder collected from SA1 suggested the existence of two distinctgroups, 86.7% (13 of 15 analyzed) of them belonging to group α(Fig. 2A, B, D, E, F, G, H, I, J, L, M, N and O) and 6.7% to group β (Fig. 2C).Group α fish are characterized by a high Sr concentration peak in theotolith core, followed by an abrupt decrease in Sr to the outer edge.Within group α, there appeared to be two sub-groups: group α1
A2 and SA3, and respective minimum, maximum and average (±standard deviation)
SA3
) TFW (g) TL (cm) TFW (g)
27.0 33.2 448.714.6 31.0 299.015.5 27.2 244.68.4 24.0 169.87.4 29.4 270.49.6 32.5 526.26.4 28.9 236.37.9 29.2 252.29.3 25.4 192.4
14.8 31.9 399.521.3 28.1 267.037.1 26.1 155.916.5 27.3 226.213.6 29.4 225.318.3 29.6 219.56.4 24.0 155.9
37.1 33.2 526.21.8 15.2±8.3 28.9±2.6 275.5±104.3

O
Distance from core (μm)0 200 400 600 800 1000 1200
0
500
1000
1500
2000
2500
3000
L
0 200 400 600 800 1000 12000
500
1000
1500
2000
2500
3000K
0 200 400 600 800 1000 12000
500
1000
1500
2000
2500
3000J
0 200 400 600 800 1000 1200
[Sr]
(pp
m)
0
500
1000
1500
2000
2500
3000
I
0 200 400 600 800 1000 12000
500
1000
1500
2000
2500
3000
H
0 200 400 600 800 1000 12000
500
1000
1500
2000
2500
3000G
0 200 400 600 800 1000 1200
[Sr]
(pp
m)
0
500
1000
1500
2000
2500
3000
F
0 200 400 600 800 1000 12000
500
1000
1500
2000
2500
3000E
0 200 400 600 800 1000 12000
500
1000
1500
2000
2500
3000D
0 200 400 600 800 1000 1200
[Sr]
(pp
m)
0
500
1000
1500
2000
2500
3000
C
0 200 400 600 800 1000 12000
500
1000
1500
2000
2500
3000B
0 200 400 600 800 1000 12000
500
1000
1500
2000
2500
3000A
0 200 400 600 800 1000 1200
[Sr]
(pp
m)
0
500
1000
1500
2000
2500
3000
N
Distance from core (μm)0 200 400 600 800 1000 1200
0
500
1000
1500
2000
2500
3000M
Distance from core (μm)0 200 400 600 800 1000 1200
[Sr]
(pp
m)
0
500
1000
1500
2000
2500
3000
Fig. 2. Strontium profiles from LA-ICP-MS analysis of otoliths from European flounder captured in the freshwater tidal area of the Minho estuary. No data for the first 119 μm of K.
238 P. Morais et al. / Journal of Sea Research 65 (2011) 235–246

J
0 500 1000 1500
[Sr]
(pp
m)
0
500
1000
1500
2000
2500
3000
G
0 500 1000 1500
[Sr]
(pp
m)
0
500
1000
1500
2000
2500
3000
D
0 500 1000 1500
[Sr]
(pp
m)
0
500
1000
1500
2000
2500
3000
B
0 500 1000 15000
500
1000
1500
2000
2500
3000C
0 500 1000 15000
500
1000
1500
2000
2500
3000
E
0 500 1000 15000
500
1000
1500
2000
2500
3000F
0 500 1000 15000
500
1000
1500
2000
2500
3000
H
0 500 1000 15000
500
1000
1500
2000
2500
3000I
0 500 1000 15000
500
1000
1500
2000
2500
3000
K
0 500 1000 15000
500
1000
1500
2000
2500
3000L
0 500 1000 15000
500
1000
1500
2000
2500
3000
A
0 500 1000 1500
[Sr]
(pp
m)
0
500
1000
1500
2000
2500
3000
O
Distance from core (μm)
0 500 1000 15000
500
1000
1500
2000
2500
3000N
Distance from core (μm)
0 500 1000 15000
500
1000
1500
2000
2500
3000M
Distance from core (μm)
0 500 1000 1500
[Sr]
(pp
m)
0
500
1000
1500
2000
2500
3000
Fig. 3. Strontium profiles from LA-ICP-MS analysis of otoliths from European flounder captured in the lower Minho estuary. No data available for the first 77 μm of A.
239P. Morais et al. / Journal of Sea Research 65 (2011) 235–246

B
0 500 1000 1500 2000 2500 3000 3500 4000 45000
500
1000
1500
2000
2500
3000C
0 500 1000 1500 2000 2500 3000 3500 4000 45000
500
1000
1500
2000
2500
3000A
0 500 1000 1500 2000 2500 3000 3500 4000 4500
[Sr]
(pp
m)
0
500
1000
1500
2000
2500
3000
D
0 500 1000 1500 2000 2500 3000 3500 4000 4500
[Sr]
(pp
m)
0
500
1000
1500
2000
2500
3000E
0 500 1000 1500 2000 2500 3000 3500 4000 45000
500
1000
1500
2000
2500
3000F
0 500 1000 1500 2000 2500 3000 3500 4000 45000
500
1000
1500
2000
2500
3000
G
0 500 1000 1500 2000 2500 3000 3500 4000 4500
[Sr]
(pp
m)
0
500
1000
1500
2000
2500
3000
H
0 500 1000 1500 2000 2500 3000 3500 4000 45000
500
1000
1500
2000
2500
3000
I
0 500 1000 1500 2000 2500 3000 3500 4000 45000
500
1000
1500
2000
2500
3000
J
0 500 1000 1500 2000 2500 3000 3500 4000 4500
[Sr]
(pp
m)
0
500
1000
1500
2000
2500
3000K
0 500 1000 1500 2000 2500 3000 3500 4000 45000
500
1000
1500
2000
2500
3000L
0 500 1000 1500 2000 2500 3000 3500 4000 45000
500
1000
1500
2000
2500
3000
O
Distance from core (μm)
0 500 1000 1500 2000 2500 3000 3500 4000 45000
500
1000
1500
2000
2500
3000
M
Distance from core (μm)
0 500 1000 1500 2000 2500 3000 3500 4000 4500
[Sr]
(pp
m)
0
500
1000
1500
2000
2500
3000N
Distance from core (μm)
0 500 1000 1500 2000 2500 3000 3500 4000 45000
500
1000
1500
2000
2500
3000
Fig. 4. Strontium profiles from LA-ICP-MS analysis of otoliths from European flounder captured in the coastal area adjacent to the Minho estuary.
240 P. Morais et al. / Journal of Sea Research 65 (2011) 235–246

SA3
Ontogenic Stage
Egg Larvae Larvae E. Met. L. Met. Juv.
[Sr]
(pp
m)
0
500
1000
1500
2000
2500P= 0.012 n.s. n.s.
SA1
Egg Larvae Larvae E. Met. L. Met. Juv.
[Sr]
(pp
m)
0
500
1000
1500
2000
2500P ≤ 0.001 P ≤ 0.001
P ≤ 0.001 P ≤ 0.001
n.s.
SA2
[Sr]
(pp
m)
0
500
1000
1500
2000
2500n.s.
241P. Morais et al. / Journal of Sea Research 65 (2011) 235–246
(Fig. 2B, D, G, I, J, L, M and N) which had concentrations of Sr higherthan 1500 ppm in the core and groupα2 (Fig. 2A, E, F, H and O) whichhad slightly lower Sr peaks in the core, varying between 745 and1195 ppm. In group α1, fish 2B shows an intermediate increase of Srconcentration of 1300 ppm around 380 μm from core (Fig. 2B). Thegroup β fish are characterized by low, relatively “flat” Sr distributionprofile (between 300 ppm and 500 ppm) throughout the otolith(Fig. 2C).
Regarding the Sr distribution in otoliths of European flounderjuveniles collected from the lower Minho estuary (SA2), datasuggested that 80.0% (12 of 15 analyzed) had similar Sr profiles tothe groupα fish described for SA1 (Fig. 3B, C, D, E, F, G, H, I, L, M, N andO). The other 20.0% of the analyzed specimens (3 of 15 analyzed) hadvery distinct patters. Thus, one specimen showed relatively high andconstant levels of Sr, between 1260 ppm and 2210 ppm (Fig. 3K).Another specimen had low levels of Sr in the otolith core (540–730 ppm) and then an abrupt increase of Sr concentration up to levelsabove 2000 ppm, after 350 μm from core (Fig. 3J). Finally, a morecomplex pattern was displayed by another fish, which had two peaksof Sr concentration along the otolith, both above 2000 ppm (Fig. 3A).
Regarding fishes collected in the coastal area (SA3), it is possible toidentify three groups according to the concentration of Sr in the core.Thus, one group is formed by 73.3% of the analyzed specimens (11 of15 analyzed), that have Sr core concentrations varying between500 ppm and 2000 ppm (Fig. 4A, B, C, D, G, H, J, L, M, N and O),similarly to the α group identified previously in the Minho estuary.Another group is formed by 20.0% of the analyzed specimens (3 of 15analyzed), whose the Sr concentration in the core is lower than500 ppm (Fig. 4E, F and K) and thus similar to group β fish of SA1 andSA2. A third group, group γ, accounts for 6.7% of the analyzedspecimens (1 of 15 analyzed), which has Sr core concentrations above2000 ppm (Fig. 4I).
The average concentration of Sr along the last 50 μm of theotoliths from SA1, SA2 and SA3 were 363±77 ppm, 1353±603 ppm,and 2080±186 ppm, respectively (Fig. 5). These concentrationswere significantly different among all the sampling sites (H=687.2,d.f.=2, P≤0.001).
The average concentration of Sr in the otoliths from fishescollected in SA1, SA2 and SA3 decreased significantly from the eggto the larvae period by 45% (from 1050±520 ppm to 575±285 ppm)(p≤0.001), 43% (from 1260±405 ppm to 715±385 ppm) (p≤0.001)and 9% (from 1370±520 ppm to 1250±585 ppm) (p=0.012),respectively (Fig. 6). Although there is a significant decrease of theaverage concentration of Sr between the larvae and early metamor-phosis ontogenic stages of fish collected in SA1 and SA2 (p≤0.001), thedecrease is lower than during the previous ontogenic transition, 25% in
Sampling Area
SA1 SA2 SA3
[Sr]
(pp
m)
0
500
1000
1500
2000
2500
Fig. 5. Overall average (dotted line) and median concentration (solid line) of Sr in thelast 50 μm (outer edge) of the otoliths from European flounder captured in each of thethree sampling areas. The 25th and 75th percentiles are shown, as well as the standarddeviation and outliers.
Fig. 6. Average (dotted line) and median (solid line) concentrations of Sr in fourontogenic stages (egg, larvae, early metamorphosis—E. Met., late metamorphosis—L. Met., and early juvenile—Juv.) of European flounder collected in the sampling areas(SA1, SA2 and SA3). The significance level between each consecutive ontogenic stage,determined with Mann–Whitney rank sum test, are shown (n.s.—non-significant). The25th and 75th percentiles are shown, as well as the standard deviation and outliers.
SA1 and 15% in SA2, while in SA3 there was an average non-significantincrease of 2% (Fig. 6). There are no significant differences between latemetamorphosis and juvenile ontogenic stages for fishes collected in allsampling areas (Fig. 6).
4. Discussion
The trend of Sr content along the otoliths of European floundercollected in the Minho estuary and adjacent coastal area seemed to begenerally governed by salinity, as observed for the stone flounder(Platichthys bicoloratus) (Yamashita et al., 2000). According to theaverage concentration of Sr along the last 50 μm of the otoliths(Fig. 5), we interpreted European flounder otolith Sr concentrationslower than 500 ppm as indicative of residence in a freshwater

242 P. Morais et al. / Journal of Sea Research 65 (2011) 235–246
environment (i.e., freshwater tidal area of the Minho estuary),between 1300 and 1700 ppm as estuarine residency (i.e., lowerMinho estuary), and above 2000 ppm as indicative of fish entering anoceanic habitat (i.e., adjacent coastal area). Therefore, the followingdiscussion is based on these assumptions.
4.2. Migration patterns of European flounder collected in the freshwatertidal area of Minho estuary based on otolith Sr distribution
For specimens collected in the freshwater tidal area of the Minhoestuary, one possible interpretation of Sr patterns for our group α fishis that they hatched in more saline waters than where they werecollected. Fish belonging to group α1 would have hatched in the lowestuary/adjacent coastal area, while those of group α2 would havehatched in the brackish estuarine area. Nevertheless, these recentlyhatched, pre-metamorphic larvae would have had to use selectivetidal stream transport, as a mechanism to enter in the estuary andprogressively migrate upstream until reaching the freshwater regionof the Minho estuary, similar to that observed for juvenile flounder inthe Elbe River (Germany) (Bos, 1999), i.e. Minho larvae would haveactively controlled their position in the water column in order to takeadvantage of the flood. However, the anatomical structures that allowflounders to actively control their position in the water column arenot developed at these early development stages (Hutchinson andHawkins, 2004). Therefore, we hypothesize that the strong MinhoRiver inflow that exists during spawning and recruitment periods,usually from February toMay, which varied during thesemonths from95 m3 s−1 to 2070 m3 s−1 between 1991 and 2008 (INAG, 2008), doesnot support the occurrence of passive migration from the coast/lowestuary to the freshwater tidal area.
Thus, we could attributed the abrupt decrease in the concentrationof Sr to a shift in the metabolic regulation of free Sr2+ (Kalish, 1989),as observed in many species that undergo profound metamorphicprocesses during their ontogenic development, such as European eel(Anguilla Anguilla) (Wang and Tzeng, 2000), whitespotted conger(Conger myriaster) (Arai et al., 2002a), daggertooth pike conger(Muraenesox cinereus) (Ling et al., 2005) and reticulated moray(Gymnothorax reticularis) (Ling et al., 2005). Thus, the pattern of Srdecrease would not be interpreted as an indication (proxy) of migra-tion to a less saline environment but might be related to metamor-phosis as was shown for the related species common sole (Solea solea)(de Pontual et al., 2003). Nevertheless, the abrupt decrease of otolithSr concentration for most fish collected in SA1, and also in SA2, wasnot concomitant with the onset of metamorphosis but with hatching(Figs. 2, 3 and 6).
The high concentration of Sr in the core of groupα flounders couldthen reflect the location where the female progenitor hydrated theeggs and not the location where they hatched, as inferred from theotoliths of brown trout (Salmo trutta) (Elfman et al., 1999; Arai et al.,2002b) and Dolly Varden (Salvelinus malma) (Campbell et al., 2002). Ifwe consider that the female progenitor migrated from a highersalinity area to a lower salinity area to spawn, then these flounderscould have hatched in the freshwater tidal area of the Minho estuary.The highest concentration of Sr in the otolith core suggested that eggswere hydrated in the low estuary/adjacent coastal area (group α1)and intermediate Sr concentrations suggested that the eggs werehydrated in a brackish environment (group α2). Specimen B(group α1) shows an intermediate increase of Sr concentration,from 465 ppm up to 1300 ppm and then another decrease to520 ppm, suggesting that this individual migrated from a freshwaterenvironment to a brackish estuarine area, and then back to thefreshwater area of the estuary (Fig. 2B).
From the otolith Sr distribution pattern for group β fish, ourinterpretation was that these flounders hatched in a freshwaterenvironment and remained there until collected. Our interpretationcontradicts the general assumption that European flounders repro-
duce exclusively in marine waters (Borsa et al., 1997; Minier et al.,2000; Martinho et al., 2007; Froese and Pauly, 2009).
Our interpretation of the otolith Sr distribution results is that all thefish collected in SA1 hatched in the freshwater tidal area of the Minhoestuary. Those with patterns of high concentration of Sr in the core ofthe otolith indicated the recent migration of the female progenitorfrom the adjacent coastal area/low estuary to the estuarine freshwatertidal area to spawn, hereafter called “sea-run mother”. One fish(Fig. 2B) appeared to have migrated from the freshwater tidal area ofthe estuary to the low estuary (intermediate maximum Sr concentra-tion ~1300 ppm) and then returned to freshwater. The mechanismsused during this migration are unknown, but a combination of activemigration strategies and selective tidal stream transport are those thatwe considered the most likely to have occurred (Bos, 1999). This typeof migration pattern might explain the presence of freshwater ecto-parasites in flounders collected in the low estuary (Dias, unpublisheddata). The putative advantage of migrations along salinity gradients isthe possibility that it might change the host–parasite relationship,since it might force ectoparasites to detach due to the osmotic stress.Hence, this might allow floundermeaningful short-term physiologicalbenefits across a range of osmoregulatory, metabolic and stressmarkers (Wells et al., 2007). However, it was also reported for anotherspecies that the physiological cost associated with parasitized speci-mens did not constitute a serious threat to individuals (Marks et al.,1996) and thus the population structure and dynamics. Similarly, itseems that the low incidence of parasite-infected flounder from theMinho estuary and the reduced number and diversity of the ecto-parasites in each fish (Dias, unpublished data) might also suggest thattheir growth and mortality are not conditioned by ectoparasites.
4.3. Migration patterns of European flounder collected in the lowerMinho estuary based on otolith Sr distribution
The group α fish collected in the lower Minho estuary (SA2) wereprobably spawned by “sea-run mothers” and hatched in thefreshwater tidal area and then migrated to the low estuary duringthe juvenile phase (Fig. 3B, C, D, E, F, G, H, I, L, M, N and O), with theexception of one fish, which may have moved or been flushed to thelower estuary during itsmetamorphosis (Fig. 3H). Two fishes probablystarted their migration to the lower estuary several days beforecapture (Fig. 3B and C) and another had not begun its migration(Fig. 3G).
One fish hatched in the low estuary and spent its entire lifemigrating between the coast just off the estuary and the lower estuary(Fig. 3K). Another, appeared to have migrated from the low estuary tothe coast as a juvenile (700 μm to 900 μm from the core), then backinto the estuary without reaching the freshwater tidal area (minimumSr concentration—715 ppm), and finally to the transitional area be-tween the estuary and coast before being captured (Fig. 3A). A thirdindividual, hatched in the transitional area between the freshwatertidal area and the low estuary and then made a fast migration to thecoast (350 μm to 450 μm from the core), where it remained beforereturning back to the low estuary prior to capture (Fig. 3J).
These fishes, whichmigrated along broad salinity gradients,must bephysiologically adapted to cope with the osmoregulatory stress(Varsamos et al., 2005). Thus, the migration of group α fishes, fromfreshwater to brackish areas, only after metamorphosis, is possibly astrategy that allows the formation of chloride cells in gill lamellae, fromthe undifferentiated cells in gill filaments. The formation of chloridecells also in gill lamellae, and not only in gill filaments, usually needsa previous acclimation to freshwater; which will probably conferflounder with a better physiological adaptation, namely osmoregula-tory, to freshwater environments after metamorphosis turning themmore adapted to migrate along salinity gradients (seeWada et al., 2007for a review).

243P. Morais et al. / Journal of Sea Research 65 (2011) 235–246
The maximum abundance of flounder was observed in thefreshwater tidal area (Freitas et al., 2009), however the migration ofjuvenile flounders to the low estuary suggests that this area is apreferred nursery area. The low estuary is utilised by older juvenilesmore than the freshwater tidal area (Dias, unpublished data), whichwas also observed in estuarine ecosystems around the GermanWadden Sea (Kerstan, 1991). In the low estuary, juvenile floundersfind the appropriate environmental requirements for optimal growthrates (Gutt, 1985), a high abundance of prey (Wouters and Cabral,2009), and appropriate space in saltmarshes and shallow areas(Peterson, 2003), where they are no longer vulnerable to predation bycommon shrimp (Crangon crangon) (van der Veer et al., 1991; Sousaet al., 2008).
4.4. Migration patterns of European flounder collected in the coastal areabased on otolith Sr distribution
Our interpretation of the otolith Sr distribution results of 93.3% ofthe specimens collected in the coastal area, 73.3% of group α and20.0% of group β fishes, reinforces the hypothesis that flounder maynot spawn exclusively in the marine environment (Summers, 1979).Therefore, the Minho estuary might also be a spawning area, ratherthan just a nursery area and feeding ground (Freitas et al., 2009).
Moreover, all 15 fish collected from the coastal area entered anestuarine ecosystem, at least once during their life cycle, supportingthe existence of a strong connectivity between estuaries and thecoastal area during the life cycle of flounder. Similar to the Minhoestuary, three other northern Portuguese estuarine ecosystems(Mondego and Douro estuaries and Ria de Aveiro lagoon) havepreviously been identified as important nursery areas for the flounderthat is commercially harvested along the coast (Vasconcelos et al.,2008).The diversity of European flounder migration patterns insouthern areas of its distribution range demonstrates its ability toadapt to various habitats and salinities. Our study also suggests thatEuropean flounder is not exclusively a catadromous species, aspreviously thought (Borsa et al., 1997; Minier et al., 2000; Martinhoet al., 2007; Froese and Pauly, 2009), since in theMinho estuary and inits adjacent coastal area flounder shows predominance for anadro-mous behavior. Hence, we suggest that flounder migration patternsmay be different among estuarine ecosystems and that general-izations about the species should be made cautiously. For example,the migration patterns for flounders from the Ythan estuary (Scot-land) (Summers, 1979) would necessarily be different from thosefrom the Minho estuary (this study), since their physiography andhydrology are different (Balls et al., 1995 and references therein,Ferreira et al., 2003) (Table 2). Estuary length and area, river mouthwidth, mean annual flow and continental shelf width are factors thatlikely define the role of an estuary in the life cycles of fishes (Monacoet al., 1992; Elliott and Dewailly, 1995; Pease, 1999; Lobry et al.,2003). This might explain the differences in migration patterns foundin different estuarine ecosystems.
4.5. Fishery management of European flounder in temperate estuaries
The study of the connectivity between estuarine and coastal fishpopulations is essential to promote effective conservation andmanage-
Table 2Physiography and hydrology of the Ythan estuary, Scotland (from Balls et al., 1995 andreferences therein) and the Minho estuary, Portugal/Spain (from Ferreira et al., 2003).
Estuary Length(km)
Mouthwidth(km)
Intertidalarea(km2)
Catchmentarea(km2)
Meanannual flow(m3 s−1)
Continentalshelf width(km)
Ythan 8 0.18 1.85 680 6 N100Minho 70 1.25 2.07 17,080 300 30–75
ment strategies (Gillanders, 2005). This study redefined the role andsignificance of the Minho estuary and of its adjacent coastal area to thelife cycle of the European flounder, disclosing a strong connectivitybetween these environments, but different from the expected one. Theunexpected outcome of our study was that flounders appear to use theestuary as a spawning ground and not only as a nursery area. Thishypothesis needs to be tested with a comprehensive ichthyoplanktonsurvey along theMinho estuary, by using traditional samplingmethods(e.g. plankton tows, Lagrangian and Eulerian samplings), light traps(Chícharo et al., 2009), boats equipped to sample in the extensiveshallow areas of the estuary (Strydom, 2007) and with the use ofelectronic tags, both for juvenile and adult specimens, to disclose the useof different habitats throughout the fishes life and migration patternsand speed (Courtney et al., 2000; Aarestrup et al., 2009; Abecasis et al.,2009).
The purpose of the current management of flounder fishery is toprotect the stock during reproduction (February–March) (Sobral,2008) and recruitment periods. Thus, estuarine fishery managersannually allowed the use of the artisanal fishing gears “varga de solha”and “solheira” between August 1st and October 31st or December15th, respectively, and set 16 cm as the minimum size for capture(Capitania do Porto de Caminha, 2005).
Besides the temporal closure, a spatial closure is also set along thefirst 27 km of the estuary. However, the identification of a putativespawning area within the wide freshwater tidal area, which startsaround 10 km upstream the river mouth, is essential to properlymanage the estuarine flounder fishery. Thus, if there is an adultspawning population in the Minho estuary, similar to those of someBritish estuaries (Beaumont and Mann, 1984; Weatherley, 1989),then it might be needed to amplify or diminish the current closurearea. This area can be reduced if spawning and the presence of adultsare restricted to the downstream limits of the freshwater tidal area,otherwise the area must be enlarged. Therefore, a survey aimedspecifically to capture adult specimens of flounders and investigatetheir age, growth and reproduction is required.
The variability of the estuarine flounder fishery depends both onfishing pressure and on the natural variability of river inflow, whichaccounted for 74% of the variability of flounder abundance in theMondego estuary (Portugal) (Martinho et al., 2009). In this estuary,higher abundance of flounder was registered in years of higher riverdischarge, probably because of the formation of extended riverplumes and stronger chemical and visual cues that enhance theprobability of coastal populations to enter estuaries (Martinho et al.,2009 and references therein). However, this does not necessarilymean that the higher the river inflow, the higher the estuarinecolonization and recruitment (Chícharo et al., 2001), since higherriver inflow causes the advection of larval stages off estuaries (Moraiset al., 2010), while moderate inflows enhance estuarine productivityand favor the retention of larval stages inside estuaries (Chícharoet al., 2001).
Finally, in order to implement an improved management strategyfor this European flounder stock, a combined effort between scientists,fishery managers and Portuguese and Spanish fishermen is required.
5. Conclusions
The most unexpected outcome of this study is the suggestion thatthe European flounder might spawn both in estuarine and coastalareas and not only in coastal areas as previously widely accepted. Itseems that all flounder captured within the estuary were hatched inthe estuary and there is evidence that the majority of femalesspawning in freshwater and brackish areas have recently migratedfrom the coast (Fig. 7A). Among the flounder collected in the coast,only 6.7% of them were hatched in a marine environment and theconnectivity between estuarine ecosystems and coastal areas wasclear (Fig. 7B). The migration of flounders from the coast to the

Fig. 7. Hypothesized migration patterns of the European flounder during its life cycle, based in the Sr concentration distribution along the otoliths of specimens collected in theMinho estuary (A) and in the adjacent coastal area (B). Dashed arrows represent the location where the oocyte hydration might have occurred.
244 P. Morais et al. / Journal of Sea Research 65 (2011) 235–246
estuary and within the estuary might provide ecological benefits tothe flounder population, particularly in regard with their relationshipwith ectoparasites. In order to improve the management of theregionally important estuarine flounder fishery, studies aiming tocollect larval stages and adult flounder must be made to confirmwhether flounder spawn in the estuary and if they do, where in theestuary. Once this is known, new temporal and/or spatial closures canbe developed and implemented or existing policy measures can becontinued.
Acknowledgments
The authors thank Rick Wastle (Fisheries and Oceans Canada,Winnipeg, Canada) for his assistance with otolith processing, ZheSong (University of Manitoba, Winnipeg, Canada) for operating theLA-ICP-MS instrument, Ana Amorim, Francisca Carvalho and TeresaSantos (Universidade do Porto, Portugal) for their assistance in fish
processing and Rute Pinto for providing the map of the study area.Carlo Heip and two anonymous reviewers provided constructivecomments on earlier versions of the manuscript. This work wasfunded by Universidade do Porto (Portugal), through the projectIPG75 and by the scholarship grant of Pedro Morais (SFRH/BPD/40832/2007, Fundação para a Ciência e a Tecnologia, Portugal).
References
Aarestrup, K., Økland, F., Hansen, M.M., Righton, D., Gargan, P., Castonguay, M.,Bernatchez, L., Howey, P., Sparholt, H., Pedersen, M.I., McKinley, R.S., 2009. Oceanicspawning migration of the European Eel (Anguilla anguilla). Science 325, 1660.
Abecasis, D., Bentes, L., Erzini, K., 2009. Home range, residency and movements ofDiplodus sargus and Diplodus vulgaris in a coastal lagoon: connectivity betweennursery and adult habitats. Estuar. Coast. Shelf Sci. 85, 525–529.
Alvarez, I., Gomez-Gesteira,M., deCastro,M., Novoa, E.M., 2008. Ekman transport along theGalician Coast (NWSpain) calculated fromQuikSCATwinds. J. Mar. Syst. 72, 101–115.
Alves, A.M., 1996. Causas e processos da dinâmica sedimentar na evolução actual dolitoral do Alto Minho. PhD Thesis, Universidade do Minho, Portugal, 442 pp.

245P. Morais et al. / Journal of Sea Research 65 (2011) 235–246
Amara, R., Selleslagh, J., Billon, G., Minier, C., 2009. Growth and condition of 0-groupEuropean flounder, Platichthys flesus as indicator of estuarine habitat quality.Hydrobiologia 627, 87–98.
Arai, T., Ikemoto, T., Kunito, T., Tanabe, S., Miyazaki, N., 2002a. Otolithmicrochemistry ofthe conger eel, Conger myriaster. J. Mar. Biol. Assoc. U.K. 82, 303–305.
Arai, T., Kotake, A., Aoyama, T., Hayano, H., Miyazaki, N., 2002b. Identifying sea-runbrown trout, Salmo trutta, using Sr:Ca ratios of otoliths. Ichthyol. Res. 49, 380–383.
Balls, P.W., Macdonald, A., Pugh, K., Edwards, A.C., 1995. Long-term nutrient enrich-ment of an estuarine system: Ythan, Scotland (1958–1993). Environ. Pollut. 90,311–321.
Beaumont, W.R.C., Mann, R.H.K., 1984. The age, growth and diet of a freshwaterpopulation of the flounder, Platichthys flesus (L), in southern England. J. Fish Biol.25, 607–616.
Borsa, P., Blanquer, A., Berrebi, P., 1997. Genetic structure of the flounders Platichthysflesus and P. stellatus at different geographic scales. Mar. Biol. 129, 233–246.
Bos, A.R., 1999. Tidal transport of flounder larvae (Pleuronectes flesus) in the Elbe River,Germany. Arch. Fish. Mar. Res. 47, 47–60.
Cabral, H.N., Costa, M.J., Salgado, J.P., 2001. Does the Tagus estuary fish communityreflect environmental changes? Clim. Res. 18, 119–126.
Cabral, H.N., Vasconcelos, R., Vinagre, C., França, S., Fonseca, V., Maia, A., Reis-Santos, P.,Lopes, M., Ruano, M., Campos, J., Freitas, V., Santos, P.T., Costa, M.J., 2007. Relativeimportance of estuarine flatfish nurseries along the Portuguese coast. J. Sea Res. 57,209–217.
Campana, S.E., 1999. Chemistry and composition of fish otoliths-pathways, mecha-nisms and applications. Mar. Ecol. Prog. Ser. 188, 263–297.
Campana, S.E., 2005. Otolith science entering the 21st century. Mar. Freshwater Res. 56,485–495.
Campbell, J.L., Babaluk, J.A., Cooper, M., Grime, G.W., Halden, N.M., Nejedly, Z., Rajta, I.,Reist, J.D., 2002. Strontium distribution in young-of-the-year Dolly Varden otoliths:potential for stock discrimination. Nucl. Instrum. Meth. Phys. Res. B 150, 260–266.
Capitania do Porto de Caminha, 2005. Edital Nº 139/2005.Chícharo, L., Chícharo, M.A., Esteves, E., Andrade, J.P., Morais, P., 2001. Effect of
alterations in freshwater supply on the abundance and distribution of Engraulisencrasicolus in the Guadiana estuary and adjacent coastal areas of south Portugal.Ecohydrol. Hydrobiol. 1, 341–345.
Chícharo, L., Faria, A., Morais, P., Amaral, A., Mendes, C., Chícharo, M.A., 2009. How tosample larval fish for taxonomical and ecophysiological studies in shallowertemperate coastal ecosystems? Cah. Biol. Mar. 50, 311–318.
Courtney, D.L., Mortensena, D.G., Orsia, J.A., Munk, K.M., 2000. Origin of juvenile Pacificsalmon recovered from coastal southeastern Alaska identified by otolith thermalmarks and coded wire tags. Fish. Res. 46, 267–278.
de Pontual, H., Lagardère, F., Amara, R., Bohn, M., Ogor, A., 2003. Influence ofontogenetic and environmental changes in the otolith microchemistry of juvenilesole (Solea solea). J. Sea Res. 50, 199–210.
Edmonds, J.S., Steckis, R.A., Moran, M.J., Caputi, N., Morita, M., 1999. Stock delineation ofpink snapper and tailor from Western Australia by analysis of stable isotope andstrontium/calcium ratios in otolith carbonate. J. Fish Biol. 55, 243–259.
Elfman, M., Limburg, K.E., Kristiansson, P., Malmqvist, K., Pallon, J., 1999. Application ofmicro-PIXE to fish life history analyses: trace element analysis of otoliths. Nucl.Instrum. Meth. Phys. Res. B 150, 272–276.
Elfman, M., Limburg, K.E., Kristiansson, P., Svedäng, H., Westin, L., Wickström, H.,Malmqvist, K., Pallon, J., 2000. Complex life histories of fishes revealed throughnatural information storage devices: cases studies of diadromous events asrecorded by otoliths. Nucl. Instr. Meth. Phys. Res. B 161–163, 877–881.
Elliott, M., Dewailly, F., 1995. The structure and components of European estuarine fishcommunities. Neth. J. Aquat. Ecol. 29, 397–417.
Elsdon, T.S., Gillanders, B.M., 2005. Temporal variability in strontium, calcium, barium,and manganese in estuaries: implications for reconstructing environmentalhistories of fish from chemicals in calcified structures. Estuar. Coast. Shelf Sci. 66,147–156.
FAO — Fisheries and Aquaculture Information and Statistics Service, 2008. TotalProduction 1950–2006. FISHSTAT Plus — Universal Software for Fishery StatisticalTime Series [online or CD-ROM]. Food and Agriculture Organization of the UnitedNations.
Ferreira, J.G., Simas, T., Nobre, A., Silva, M.C., Schifferegger, K., Lencart-Silva, J., 2003.Identification of sensitive areas and vulnerable zones in transitional and coastalPortuguese systems. Application of the United States National Estuarine Eutrophi-cation Assessment to the Minho, Lima, Douro, Ria de Aveiro, Mondego, Tagus, Sado,Mira, Ria Formosa and Guadiana systems. INAG/IMAR Technical Report.
Florin, A.-B., Höglund, J., 2008. Population structure of flounder (Platichthys flesus) inthe Baltic Sea: differences among demersal and pelagic spawners. Heredity 101,27–38.
Freitas, V., Costa-Dias, S., Campos, J., Bio, A., Santos, P., Antunes, C., 2009. Patterns inabundance and distribution of juvenile flounder, Platichthys flesus, inMinho estuary(NW Iberian Peninsula). Aquat. Ecol. 43, 1143–1153.
Froese, R., Pauly, D., 2009. Fish base. http://www.fishbase.org. Version February 2009.Gillanders, B.M., 2005. Using elemental chemistry of fish otoliths to determine con-
nectivity between estuarine and coastal habitats. Estuar. Coast. Shelf Sci. 64, 47–57.Gutt, J., 1985. The growth of juvenile flounders (Platichthys flesus) at salinities 0, 5, 15
and 35‰. Z. Angew. Ichthyol. 1, 17–26.Halden, N.M., Friedrich, L.A., 2008. Trace-element distribution in fish otoliths: natural
markers of life histories, environmental conditions and exposure to tailingseffluence. Mineral. Mag. 72, 593–605.
Hutchinson, S., Hawkins, L.E., 2004. The relationship between temperature and the sizeand age of larvae and peri-metamorphic stages of Pleuronectes flesus. J. Fish Biol. 65,448–459.
INAG, 2008. Sistema nacional de informação de recursos hídricosAvailable at: http://http://www.inag.pt.
Kalish, J.M., 1989. Otolith microchemistry: validation of the effects of physiology, ageand environment on otolith composition. J. Exp. Mar. Biol. Ecol. 132, 151–178.
Kerstan, M., 1991. The importance of rivers as nursery grounds for 0- and 1-groupflounder (Platichthys flesus L.) in comparison to the Wadden Sea. Neth. J. Sea Res.27, 353–366.
Kraus, R.T., Secor, D.H., 2004. Incorporation of strontium into otoliths of an estuarinefish. J. Exp. Mar. Biol. Ecol. 302, 85–106.
Ling, Y.J., Iizuka, Y., Tzeng, W.N., 2005. Decreased Sr/Ca ratios in the otoliths of twomarine eels, Gymnothorax reticularis and Muraenesox cinereus, during metamor-phosis. Mar. Ecol. Prog. Ser. 304, 201–206.
Lobry, J., Mourand, L., Rochard, E., Elie, P., 2003. Structure of the Gironde estuarine fishassemblages: a European estuaries comparison perspective. Aquat. Living Resour.16, 47–58.
Marks, R.E., Juanes, F., Hare, J.A., Conover, D.O., 1996. Occurrence and effect of theparasitic isopod, Lironeca ovalis (Isopoda: Cymothoidae), on young-of-the-yearbluefish, Pomatomus saltatrix (Pisces: Pomatomidae). Can. J. Fish. Aquat. Sci. 53,2052–2057.
Martinho, F., Leitão, R., Neto, J.M., Cabral, N.H., Marques, J.C., Pardal, M.A., 2007. The useof nursery areas by juvenile fish in a temperate estuary, Portugal. Hydrobiologia587, 281–290.
Martinho, F., Dolbeth, M., Viegas, I., Teixeira, C.M., Cabral, H.N., Pardal, M.A., 2009.Environmental effects on the recruitment variability of nursery species. Estuar.Coast. Shelf Sci. 83, 460–468.
Minier, C., Levy, F., Rabel, D., Bocquené, G., Godefroy, D., Burgeot, T., Leboulenger, F.,2000. Flounder health status in the Seine Bay. A multibiomarker study. Mar.Environ. Res. 50, 373–377.
Monaco, M.E., Lowery, T.A., Emmett, R.L., 1992. Assemblages of U.S. west coast estuariesbased on the distribution of fishes. J. Biogeogr. 19, 251–267.
Morais, P., Martins, F., Chícharo, M.A., Lopes, J., Chícharo, L., 2010. Merging anchovy eggsabundance into a hydrodynamic model as an assessment tool for estuarineecohydrological management. River Res. Appl. doi:10.1002/rra.1443
Nielsen, J.G., 1986. Pleuronectidae. In: Whitehead, P.J.P., Bauchot, M.-L., Hureau, J.-C.,Nielsen, J., Tortonese, E. (Eds.), Fishes of the North-Eastern Atlantic and theMediterranean, Vol. 3. UNESCO, Paris, pp. 1299–1307.
Nissling, A., Westin, L., Hjerne, O., 2002. Reproductive success in relation to salinity forthree flatfish species, dab (Limanda limanda), plaice (Pleuronectes platessa), andflounder (Pleuronectes flesus), in the brackish water Baltic Sea. ICES J. Mar. Sci. 59,93–108.
Palenzuela, J., Iglesias, G.M., Vilas, L.G., 2004. Pelagic fisheries study using GIS andremote sensing imagery in Galicia (Spain). ICES CM 2004/ P:44.
Pearce, N.J.G., Perkins, W.T., Westgate, J.A., Gorton, M.P., Jackson, S.E., Neal, C.R.,Chenery, S.P., 1997. A compilation of new and published major and trace elementdata for NIST SRM 610 and NIST SRM 612 glass reference materials. Geostand.Newsl. J. Geostand. Geoanal. 21, 115–144.
Pease, B.C., 1999. A spatially oriented analysis of estuaries and their associatedcommercial fisheries in New South Wales, Australia. Fish. Res. 42, 67–86.
Peterson, M., 2003. A conceptual view of environment–habitat–production linkages intidal river estuaries. Rev. Fish. Sci. 11, 291–313.
Pombo, L., Rebelo, J.E., Elliott, M., 2007. The structure, diversity and somatic productionof the fish community in an estuarine coastal lagoon, Ria de Aveiro (Portugal).Hydrobiologia 587, 253–268.
Ramos, S., Ré, P., Bordalo, A.A., 2010. Recruitment of flatfish species to an estuarinenursery habitat (Lima estuary, NW Iberian Peninsula). J. Sea Res. 64, 473–486.
Secor, D.H., Rooker, J.R., 2000. Is otolith strontium a useful scalar of life cycles inestuarine fishes? Fish. Res. 46, 359–371.
Sobral, M.P.C., 2008. Aspectos relativos a' biologia reprodutiva da solha, Platichthysflesus (Linnaeus, 1758), da Ria de Aveiro e litoral adjacente. Relatórios Científicos eTécnicos do IPIMAR, Série Digital, No. 44. 31 pp.
Sokal, R.R., Rohlf, F.J., 1995. Biometry: The Principles and Practice of Statistics inBiological Research, 3rd Edition. W. H. Freeman and Co., New York. 887 pp.
Sousa, R., Guilhermino, L., Antunes, C., 2005. Molluscan fauna in the freshwater tidal areaof the River Minho estuary, NW of Iberian Peninsula. Ann. Limnol. 41, 141–147.
Sousa, R., Dias, S., Freitas, V., Antunes, C., 2008. Subtidal macrozoobenthic assemblagesalong the River Minho estuarine gradient (north-west Iberian Peninsula). Aquat.Conserv. Mar. Freshw. Ecosyst. 18, 1063–1077.
Strydom, N.A., 2007. Jetski-based plankton towing as a new method of sampling larvalfishes in shallow marine habitats. Environ. Biol. Fish. 78, 299–306.
Summers, R.W., 1979. Life cycle and population ecology of the flounder Platichthysflesus (L.) in the Ythan estuary, Scotland. J. Nat. Hist. 13, 703–723.
Teixeira, C.T., Cabral, H.N., 2009. Time series analysis of flatfish landings in thePortuguese coast. Fish. Res. 96, 252–258.
van Achterberg, E., Ryan, C.G., Griffin, W.L., 2001. GLITTER User's Manual: On-lineInteractive Data Reduction for the LA-ICP-MS Microprobe. Version 4. MacquarieResearch Limited, North Ryde.
Van der Veer, H.W., Bergman, M.J.N., Dapper, R., Witte, J.I.J., 1991. Population dynamicsof an intertidal 0-group flounder Platichthys flesus population in the western DutchWadden Sea. Mar. Ecol. Prog. Ser. 73, 141–148.
Varsamos, S., Nebel, C., Charmantier, G., 2005. Ontogeny of osmoregulation in post-embryonic fish: a review. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 141,401–429.
Vasconcelos, R.P., Reis-Santos, P., Tanner, S., Maia, A., Latkoczy, C., Günther, D., Costa, M.J.,Cabral, H., 2008. Evidence of estuarine nursery origin of five coastal fish species alongthe Portuguese coast through otolith elemental fingerprints. Estuar. Coast. Shelf Sci.79, 317–327.

246 P. Morais et al. / Journal of Sea Research 65 (2011) 235–246
Vinagre, C., França, S., Costa, M.J., Cabral, H.N., 2005. Niche overlap between juvenileflatfishes, Platichthys flesus and Solea solea, in a southern European estuary andadjacent coastal waters. J. Appl. Ichthyol. 21, 114–120.
Wada, T., Aritaki, M., Yamashita, Y., Tanaka, M., 2007. Comparison of low-salinityadaptability and morphological development during the early life history of fivepleuronectid flatfishes, and implications for migration and recruitment to theirnurseries. J. Sea Res. 58, 241–254.
Wang, C.H., Tzeng, W.N., 2000. The timing of metamorphosis and growth rates ofAmerican and European eel leptocephali: a mechanism of larval segregativemigration. Fish. Res. 46, 191–205.
Weatherley, N.S., 1989. The diet and growth of 0-group flounder, Platichthys flesus (L),in the river Dee, North Wales. Hydrobiologia 178, 193–198.
Wells, A., Grierson, C.E., Marshall, L., MacKenzie, M., Russon, I.J., Reinardy, H.,Sivertsgård, R., Bjørn, P.A., Finstad, B., Wendelaar Bonga, S.E., Todd, C.D., Hazon,N., 2007. Physiological consequences of ‘premature freshwater return’ for wild sea-run brown trout (Salmo trutta) postsmolts infested with sea lice (Lepeophtheirussalmonis). Can. J. Fish. Aquat. Sci. 64, 1360–1369.
Wouters, N., Cabral, H.N., 2009. Are flatfish nursery grounds richer in benthic prey?Estuar. Coast. Shelf Sci. 83, 613–620.
Yamashita, Y., Otake, T., Yamada, H., 2000. Relative contributions from exposed inshoreand estuarine nursery grounds to the recruitment of stone flounder, Platichthysbicoloratus, estimated using otolith Sr:Ca ratios. Fish. Oceanogr. 9, 316–327.
Zlokovitz, E.R., Secor, D.H., Piccoli, P.M., 2003. Patterns of migration in Hudson Riverstriped bass as determined by otolith microchemistry. Fish. Res. 63, 245–259.








