The Influence of Stress Hormones on Fear Circuitry
27
The Influence of Stress Hormones on Fear Circuitry Sarina M. Rodrigues, 1 Joseph E. LeDoux, 2, ∗ and Robert M. Sapolsky 3, ∗ 1 Institute of Personality and Social Research, University of California, Berkeley, California 94720; Address correspondence to Department of Psychology, Oregon State University, Corvallis, Oregon 97331; email: [email protected] 2 Center for Neural Science and Department of Psychology, New York University, New York, New York 10003; Emotional Brain Institute Labs of the Nathan Kline Institute, Orangeburg, New York 10962; email: [email protected] 3 Departments of Biological Sciences and Neurology and Neurological Sciences, Stanford Medical Center, Stanford, California 94305-5020; email: [email protected] Annu. Rev. Neurosci. 2009. 32:289–313 First published online as a Review in Advance on March 24, 2009 The Annual Review of Neuroscience is online at neuro.annualreviews.org This article’s doi: 10.1146/annurev.neuro.051508.135620 Copyright c 2009 by Annual Reviews. All rights reserved 0147-006X/09/0721-0289$20.00 ∗ These authors contributed equally to this work. Key Words fear conditioning, glucocorticoids, norepinephrine Abstract Fear arousal, initiated by an environmental threat, leads to activation of the stress response, a state of alarm that promotes an array of autonomic and endocrine changes designed to aid self-preservation. The stress re- sponse includes the release of glucocorticoids from the adrenal cortex and catecholamines from the adrenal medulla and sympathetic nerves. These stress hormones, in turn, provide feedback to the brain and influ- ence neural structures that control emotion and cognition. To illustrate this influence, we focus on how it impacts fear conditioning, a behavioral paradigm widely used to study the neural mechanisms underlying the acquisition, expression, consolidation, reconsolidation, and extinction of emotional memories. We also discuss how stress and the endocrine mediators of the stress response influence the morphological and elec- trophysiological properties of neurons in brain areas that are crucial for fear-conditioning processes, including the amygdala, hippocampus, and prefrontal cortex. The information in this review illuminates the behavioral and cellular events that underlie the feedforward and feed- back networks that mediate states of fear and stress and their interaction in the brain. 289 Annu. Rev. Neurosci. 2009.32:289-313. Downloaded from arjournals.annualreviews.org by Universiteit van Amsterdam on 03/25/10. For personal use only.
-
Upload
leandervoets -
Category
Documents
-
view
26 -
download
1
description
The Influence of Stress Hormones on Fear Circuitry
Transcript of The Influence of Stress Hormones on Fear Circuitry

ANRV379-NE32-13 ARI 13 May 2009 8:31
The Influence of StressHormones on Fear CircuitrySarina M. Rodrigues,1 Joseph E. LeDoux,2,∗
and Robert M. Sapolsky3,∗
1Institute of Personality and Social Research, University of California, Berkeley,California 94720; Address correspondence to Department of Psychology, Oregon StateUniversity, Corvallis, Oregon 97331; email: [email protected] for Neural Science and Department of Psychology, New York University,New York, New York 10003; Emotional Brain Institute Labs of the Nathan Kline Institute,Orangeburg, New York 10962; email: [email protected] of Biological Sciences and Neurology and Neurological Sciences,Stanford Medical Center, Stanford, California 94305-5020; email: [email protected]
Annu. Rev. Neurosci. 2009. 32:289–313
First published online as a Review in Advance onMarch 24, 2009
The Annual Review of Neuroscience is online atneuro.annualreviews.org
This article’s doi:10.1146/annurev.neuro.051508.135620
Copyright c© 2009 by Annual Reviews.All rights reserved
0147-006X/09/0721-0289$20.00
∗These authors contributed equally to this work.
Key Words
fear conditioning, glucocorticoids, norepinephrine
AbstractFear arousal, initiated by an environmental threat, leads to activation ofthe stress response, a state of alarm that promotes an array of autonomicand endocrine changes designed to aid self-preservation. The stress re-sponse includes the release of glucocorticoids from the adrenal cortexand catecholamines from the adrenal medulla and sympathetic nerves.These stress hormones, in turn, provide feedback to the brain and influ-ence neural structures that control emotion and cognition. To illustratethis influence, we focus on how it impacts fear conditioning, a behavioralparadigm widely used to study the neural mechanisms underlying theacquisition, expression, consolidation, reconsolidation, and extinctionof emotional memories. We also discuss how stress and the endocrinemediators of the stress response influence the morphological and elec-trophysiological properties of neurons in brain areas that are crucialfor fear-conditioning processes, including the amygdala, hippocampus,and prefrontal cortex. The information in this review illuminates thebehavioral and cellular events that underlie the feedforward and feed-back networks that mediate states of fear and stress and their interactionin the brain.
289
Ann
u. R
ev. N
euro
sci.
2009
.32:
289-
313.
Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
Uni
vers
iteit
van
Am
ster
dam
on
03/2
5/10
. For
per
sona
l use
onl
y.

ANRV379-NE32-13 ARI 13 May 2009 8:31
Contents
INTRODUCTION . . . . . . . . . . . . . . . . . . 291WHAT IS FEAR, AND HOW IS IT
ORGANIZED IN THE BRAIN? . . 291Studying Fear in the Laboratory
with Fear Conditioning . . . . . . . . . 291Phases of Fear Conditioning . . . . . . . . 292Neural Circuitry Underlying
Fear Conditioning . . . . . . . . . . . . . . 292FEAR AROUSAL AND THE
STRESS RESPONSE:OVERVIEW OF THISFEEDFORWARD ANDFEEDBACK NETWORK . . . . . . . . . 294
HOW DO STRESS,NOREPINEPHRINE, ANDGLUCOCORTICOIDSINFLUENCE DIFFERENTPHASES OF FEARCONDITIONING?. . . . . . . . . . . . . . . 295
ACQUISITION OF FEARCONDITIONING. . . . . . . . . . . . . . . . 297The Effects of Stress on the
Acquisition of FearConditioning . . . . . . . . . . . . . . . . . . . 297
The Effects of NE on theAcquisition of FearConditioning . . . . . . . . . . . . . . . . . . . 297
The Effects of GCs on theAcquisition of FearConditioning . . . . . . . . . . . . . . . . . . . 297
CONSOLIDATION OFLONG-TERM MEMORY . . . . . . . . 297The Effects of Stress on the
Consolidation of LTM ofFear Conditioning . . . . . . . . . . . . . . 297
The Effects of NE on theConsolidation of LTMof Fear Conditioning . . . . . . . . . . . . 298
The Effects of GCs on the
Consolidation of LTMof Fear Conditioning . . . . . . . . . . . . 298
RECONSOLIDATION . . . . . . . . . . . . . . 299The Effects of Stress on the
Reconsolidation of FearConditioning . . . . . . . . . . . . . . . . . . . 299
The Effects of NE Manipulationon the Reconsolidation of FearConditioning . . . . . . . . . . . . . . . . . . . 299
The Effects of GC Manipulationon the Reconsolidationof Fear Conditioning . . . . . . . . . . . . 299
EXTINCTION . . . . . . . . . . . . . . . . . . . . . . 299The Effects of Stress on the
Extinction of FearConditioning . . . . . . . . . . . . . . . . . . . 299
Involvement of NE in the Extinctionof Fear Conditioning . . . . . . . . . . . . 299
Involvement of GCs in theExtinction of FearConditioning . . . . . . . . . . . . . . . . . . . 300
SUMMARY OF THE EFFECTSON FEAR CONDITIONING . . . . 300
HOW STRESS AFFECTSFEAR CIRCUITRY ONA CELLULAR LEVEL. . . . . . . . . . . . 300The Influence of Stress on
the Morphology of Neuronsin Fear Circuitry . . . . . . . . . . . . . . . . 300
The Influence of Stress on theElectrophysiological Propertiesof Neurons in Fear Circuitry . . . . 302
The Influence of NE on theElectrophysiological Propertiesof Neurons in Fear Circuitry . . . . 304
The Influence of GCs on theElectrophysiological Propertiesof Neurons in Fear Circuitry . . . . 304
CONCLUSIONS . . . . . . . . . . . . . . . . . . . . 304
“The oldest and strongest emotion of mankind isfear.”
—H.P. Lovecraft
“A crust eaten in peace is better than a banqueteaten in fear.”
—Aesop
290 Rodrigues · LeDoux · Sapolsky
Ann
u. R
ev. N
euro
sci.
2009
.32:
289-
313.
Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
Uni
vers
iteit
van
Am
ster
dam
on
03/2
5/10
. For
per
sona
l use
onl
y.

ANRV379-NE32-13 ARI 13 May 2009 8:31
INTRODUCTION
Evolution has endowed each species with an ar-ray of instinctual defense mechanisms to helporganisms cope with environmental threatsand other challenges to safety and well-being.Skunks spray offensive odors, blowfish inflate tolook bigger than they actually are, and roachesscurry away into crevices for safety. Althoughhumans in modern societies usually have theluxury of not having to worry about escapingfrom predators (except for the odd shark, bear,or axe murderer), the emotion of fear and thedefensive behaviors connected with it help tosave us from peril both in everyday situationsas well as under rare but hazardous conditions:We duck for cover, slam on the brakes, run forthe hills, or scream for help.
Fear arousal is one of the most reliableroutes for activating the stress response (LaBar& LeDoux 2001), an array of peripheral au-tonomic and neuroendocrine changes that aidsurvival (McEwen 2003, Sapolsky et al. 2000).At the same time, the peripheral changes in-duced by fear impact the brain, altering theway emotional and cognitive systems processinformation. Although such responses enhancesurvival in the short run, chronic activationof stress responses contributes to the devel-opment of pathological states such as anxiety,phobias, depression, and post-traumatic stressdisorder (PTSD), as well as a host of physi-cal ailments, including compromised immunefunction, hypertension, and insulin resistance(McEwen 2003, Sapolsky et al. 2000).
The aim of this article is to explore howfear arousal, manifested on the neurobiologicallevel, leads to activation of peripheral stress re-sponses and how stress responses, in turn, alterbrain systems that control emotion and cogni-tion. In the interest of space, we cannot coversome important topics, including how early-lifeexperiences, genetics, or gender might inter-act with stress in altering fear (DeRijk & deKloet 2005, Kalin & Shelton 2003, Luine 2002,Lyons & Parker 2007, Meaney 2001, Weinstock2007).
CS: neutralconditioned stimulus
US: aversiveunconditionedstimulus
NE: norepinephrine
GC: glucocorticoid
CR: conditionedresponse
WHAT IS FEAR, AND HOW IS ITORGANIZED IN THE BRAIN?
The term fear refers to both a psychologicalstate and a set of bodily responses that occurin response to threat (LeDoux 1996). Muchprogress has been made in understanding howfear is organized in the brain through studiesof Pavlovian fear conditioning, which we fo-cus on here (Fanselow & Poulos 2005; Lang &Davis 2006; LeDoux 1995, 1996, 2007; Maren2001, 2005; Pare et al. 2004; Rodrigues et al.2004). There are a variety of ways to assesslearned fear besides measuring Pavlovian con-ditioned response effects. For example, a num-ber of studies have examined the role of brain ar-eas in passive and active avoidance conditioning(Gabriel et al. 2003, McGaugh 2003, Sarter &Markowitsch 1985). In such studies, fear ismeasured indirectly in terms of instrumentalbehaviors that avoid danger. Because studies ofavoidance conditioning have not led to a co-herent view of the neural system that mediatesavoidance (Cain & LeDoux 2008), we empha-size the effects of stress on fear conditioning inthis review. However, we mention some find-ings from other procedures where relevant.
Studying Fear in the Laboratorywith Fear Conditioning
Pavlovian fear conditioning is a behavioral pro-cedure in which an emotionally neutral condi-tioned stimulus (CS), such as an auditory tone,is paired with an aversive conditioned stimu-lus (US), typically a foot shock. After one orseveral pairings, the CS comes to elicit de-fensive behaviors, including freezing behav-ior, as well as increased arousal in the brainand secretion of norepinephrine (NE) andglucocorticoids (GCs) peripherally. Theseconditioned responses (CRs) are hard-wiredand occur in response to both innate andlearned threats. Thus, fear learning does notcreate conditioned fear responses but allowsenvironmental stimuli to come to elicit suchresponses.
www.annualreviews.org • Stress Hormones and Fear Circuitry 291
Ann
u. R
ev. N
euro
sci.
2009
.32:
289-
313.
Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
Uni
vers
iteit
van
Am
ster
dam
on
03/2
5/10
. For
per
sona
l use
onl
y.

ANRV379-NE32-13 ARI 13 May 2009 8:31
STM: short-termmemory
LTM: long-termmemory
LA: lateral nucleus ofthe amygdala
CE: central nucleus ofthe amygdala
Phases of Fear Conditioning
In studies of fear conditioning, responseselicited by the CS are often measured duringacquisition and retrieval tests. Acquisition is ini-tial learning of the association between the CSand US during the training portion of fear con-ditioning, when the animal first learns to pairthe two. Retrieval involves a test of the CS-USassociation in which the CS is presented alone.Retrieval tests that occur within a few hoursafter acquisition measure short-term memory(STM), whereas those that occur later (typ-ically 24 hours after learning) measure long-term memory (LTM). STM and LTM tests areused to study memory consolidation, the pro-cess through which an unstable STM is con-verted into a more stable and enduring LTMtrace (Dudai 2004, McGaugh 2000, Rodrigueset al. 2004).
Retrieval tests are also used to study recon-solidation. Reconsolidation occurs when fearmemories are retrieved and enter a new la-bile state that requires stabilization via proteinsynthesis to persist as an enduring LTM trace(Dudai 2006, Nader 2003, Nader et al. 2000). Inaddition, retrieval tests are utilized to measureextinction, which is the gradual reduction in theability of the CS to elicit fear CRs that occurwhen the CS is presented repeatedly in the ab-sence of the US (Quirk & Mueller 2008, Sotres-Bayon et al. 2004). Unlike the disruption of re-consolidation, which is robust and long lasting,extinction involves a new learning process. Asa result, after extinction training, the CS canspontaneously produce CRs again after the an-imal is reexposed to the US or is placed in anovel context (Bouton et al. 2006, Ji & Maren2007).
Neural Circuitry UnderlyingFear Conditioning
Fear conditioning is especially useful for study-ing the neural basis of fear because the cir-cuit can be described in terms of pathwaysprocessing the CS and controlling the CRs. In-deed, the pathways have been mapped from the
CS sensory processing to the CR motor con-trol systems. A key link in this circuitry is theamygdala, which sits between the sensory in-put systems on one hand and the motor outputsystems on the other (Fanselow & Poulos 2005;Lang & Davis 2006; LeDoux 1995, 1996, 2007;Maren 2001, 2005; Pare et al. 2004; Rodrigueset al. 2004). However, other brain structuresalso contribute to fear conditioning.
Amygdala contributions to fear condition-ing. The amygdala contains a heterogeneity ofdistinct nuclei, differing by cell type, density,neurochemical composition, and connectivity(LeDoux 2007, Pitkanen et al. 2000, Swanson2003).
The lateral nucleus (LA) is typically viewedas the sensory gateway to the amygdala be-cause it receives auditory, visual, gustatory,olfactory, and somatosensory (including pain)information from the thalamus and the cor-tex (olfactory and taste information is transmit-ted to other nuclei, as well). The LA receivessensory information about the CS from tha-lamic and cortical projections. The thalamo-amygdala pathway is a shortcut of sorts, trans-mitting rapid and crude information about thefear-eliciting stimulus without the opportu-nity for filtering by conscious control (LeDoux1995, 1996). The cortico-amygdala pathway, incontrast, provides slower, but more detailed andsophisticated sensory information. Neurons inthe LA respond to both the CS and the US,and damage to, or a disruption of, the LA pre-vents fear conditioning. It appears that the con-vergence takes place in the dorsal half of theLA (Romanski et al. 1993), and indeed single-unit recordings show that cellular plasticity oc-curs in the dorsal LA during fear conditioning(LeDoux 2007, Maren & Quirk 2004). More-over, molecular changes in the LA consolidatethe fear memory, converting STM into LTMtraces (Dudai 2004, Rodrigues et al. 2004).
The central nucleus (CE), in contrast, isviewed as the major output region of theamygdala. The CE controls the expressionof the fear reaction, including behavioral,
292 Rodrigues · LeDoux · Sapolsky
Ann
u. R
ev. N
euro
sci.
2009
.32:
289-
313.
Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
Uni
vers
iteit
van
Am
ster
dam
on
03/2
5/10
. For
per
sona
l use
onl
y.

ANRV379-NE32-13 ARI 13 May 2009 8:31
autonomic, and endocrine responses via projec-tions to downstream areas, including hypotha-lamus, central gray, and the dorsal motor nu-cleus of the vagus (Fanselow & Poulos 2005;Lang & Davis 2006; LeDoux 1995, 1996, 2007;Maren 2001, 2005; Pare et al. 2004; Rodrigueset al. 2004). Plasticity also occurs in the CE(Pare et al. 2004, Wilensky et al. 2006), but thisis likely based on LA plasticity.
The LA communicates with the CE directly,but the connections between these two nucleiare somewhat modest and other amygdaloidnuclei also help mediate between these two.For instance, the LA projects to the basal nu-cleus (B), which in turn projects to CE (LeDoux2000, Pitkanen et al. 1997). The LA and B alsoproject to the intercalated region (ITC), an in-hibitory network that connects with the CE(Likhtik et al. 2008, Pare et al. 2004). Projec-tion neurons in the CE tend to be inhibitory,thus LA and B projections to the ITC maystimulate the ITC’s inhibition of CE neuronsto allow the expression fear responses (LeDoux2007). In addition, the B projects to the striatum(McDonald 1991) to orchestrate instrumentalbehaviors, such as escape to safety (LeDoux2007). Furthermore, CE neurons project to themedial peri-locus coeruleus dendritic region,resulting in increased NE release (Aston-Jones2004), as well as to other monoamine systems inthe brainstem and forebrain (Fudge & Emiliano2003, Gray 1993, Pare 2003).
Information about the context of a fear-ful situation is conveyed to the LA andB by the hippocampus ( Ji & Maren 2007,Kim & Fanselow 1992, Phillips & LeDoux1992, Pitkanen et al. 2000). Thus, althoughfear conditioning to discrete sensory cues,such as a tone or a light, are hippocam-pal independent, conditioning to contex-tual information, including details about theenvironment, are hippocampal dependent. TheLA and B are also interconnected to a variety ofcortical areas, such as the prefrontal and poly-modal association cortices (Amaral & Insausti1992, McDonald 1998, McDonald et al. 1996,
B: basal nucleus of theamygdala
mPFC: medialprefrontal cortex
IL: infralimicsubregion of themPFC
Pitkanen et al 2000, Price 2003, Stefanacci &Amaral 2002). These connections allow higher-order cognitive processes, such as emotion reg-ulation, complex associations, imagination, andreactions to the news of an abstract state, toinfluence amygdala functions.
The LA, B, and accessory basal nuclei aresometimes grouped together as the BLA (baso-lateral amygdala). It is important to keep thesenuclei separate when possible.
Although the LA is required for fear con-ditioning under standard training conditions,it appears that, with overtraining, fear condi-tioning can be mediated by circuits that do notdepend on the LA (Lee et al. 2005, Maren1999). With overtraining, weak connectionsto the CE appear to induce fear conditioning(Zimmerman et al. 2007), but such pathwaysare not normally used. Moreover, although theB is not required for conditioning, it neverthe-less appears to contribute to the appearance ofthe fear response (Anglada-Figueroa & Quirk2005). Some authors suggest that under someconditions, especially those involving the useof instrumental responses to assess Pavlovianconditioning, the CE can mediate condition-ing independent of LA (Balleine & Killcross2006, Cardinal et al. 2002). However, this con-clusion is extrapolated largely on the basisof appetitive conditioning findings and needsto be studied more systematically in aversiveconditioning.
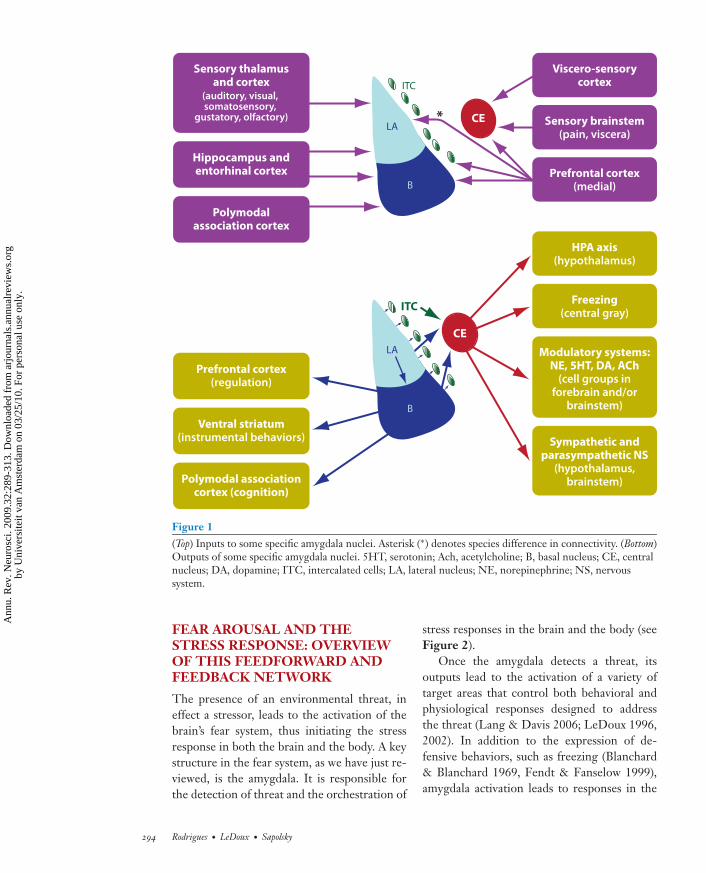
The expression of fear responses can be reg-ulated by the medial prefrontal cortex (mPFC)via projections to the LA, B, and ITC. The in-fralimbic subregion (IL) of the mPFC is es-pecially important for fear extinction (Quirket al. 2006, Rosenkranz et al. 2003, Sotres-Bayon et al. 2004) because it inhibits amygdalaoutput (Vidal-Gonzalez et al. 2006). However,investigators have debated the nature of therole of prefrontal amygdala interactions in ex-tinction (Quirk et al. 2006, Rosenkranz et al.2003, Sotres-Bayon et al. 2004). The varioussensory and higher-order influences on theamygdala are shown in Figure 1.
www.annualreviews.org • Stress Hormones and Fear Circuitry 293
Ann
u. R
ev. N
euro
sci.
2009
.32:
289-
313.
Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
Uni
vers
iteit
van
Am
ster
dam
on
03/2
5/10
. For
per
sona
l use
onl
y.
ANRV379-NE32-13 ARI 13 May 2009 8:31
ITC
LA
B
CE
Sensory thalamusand cortex
(auditory, visual, somatosensory,
gustatory, olfactory)
Viscero-sensorycortex
Sensory brainstem(pain, viscera)
Prefrontal cortex(medial)
Hippocampus andentorhinal cortex
Polymodalassociation cortex
ITC
LA
B
CE
HPA axis(hypothalamus)
Freezing(central gray)
Modulatory systems:NE, 5HT, DA, ACh
(cell groups inforebrain and/or
brainstem)
Sympathetic andparasympathetic NS
(hypothalamus,brainstem)
Ventral striatum(instrumental behaviors)
Polymodal associationcortex (cognition)
Prefrontal cortex(regulation)
Figure 1(Top) Inputs to some specific amygdala nuclei. Asterisk (∗) denotes species difference in connectivity. (Bottom)Outputs of some specific amygdala nuclei. 5HT, serotonin; Ach, acetylcholine; B, basal nucleus; CE, centralnucleus; DA, dopamine; ITC, intercalated cells; LA, lateral nucleus; NE, norepinephrine; NS, nervoussystem.
FEAR AROUSAL AND THESTRESS RESPONSE: OVERVIEWOF THIS FEEDFORWARD ANDFEEDBACK NETWORK
The presence of an environmental threat, ineffect a stressor, leads to the activation of thebrain’s fear system, thus initiating the stressresponse in both the brain and the body. A keystructure in the fear system, as we have just re-viewed, is the amygdala. It is responsible forthe detection of threat and the orchestration of
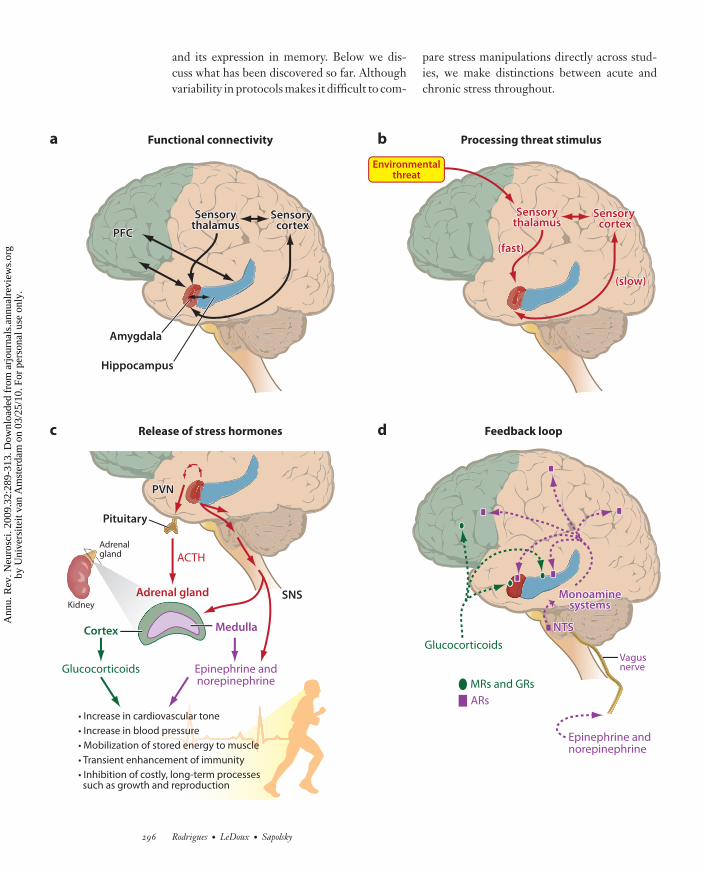
stress responses in the brain and the body (seeFigure 2).
Once the amygdala detects a threat, itsoutputs lead to the activation of a variety oftarget areas that control both behavioral andphysiological responses designed to addressthe threat (Lang & Davis 2006; LeDoux 1996,2002). In addition to the expression of de-fensive behaviors, such as freezing (Blanchard& Blanchard 1969, Fendt & Fanselow 1999),amygdala activation leads to responses in the
294 Rodrigues · LeDoux · Sapolsky
Ann
u. R
ev. N
euro
sci.
2009
.32:
289-
313.
Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
Uni
vers
iteit
van
Am
ster
dam
on
03/2
5/10
. For
per
sona
l use
onl
y.

ANRV379-NE32-13 ARI 13 May 2009 8:31
brain and the body that support the fear reac-tion. In the brain, monoaminergic systems areactivated, resulting in the release of neurotrans-mitters such as NE, acetylcholine, serotonin,and dopamine throughout the brain. Theseneurotransmitters lead to an increase in arousaland vigilance and, in general, an enhancementin the processing of external cues (Aston-Jones& Cohen 2005, LeDoux 2007, Ramos &Arnsten 2007, Talarovicova et al. 2007).Detection of threat by the amygdala also hasendocrine consequences. Amygdaloid signalingcauses secretion of corticotropin-releasing hor-mone (CRH) from the periventricular nucleusof the hypothalamus. CRH, along with otherhypothalamic secretagogs, causes the release ofadrenocorticotropic hormone from the pitu-itary, which in turn stimulates the secretion ofGCs from the adrenal cortex. Circulating GCsbind to the high-affinity mineralocorticoidreceptor (MR) and the low-affinity glucocor-ticoid receptor (GR) in tissues throughout thebody and the brain (de Kloet 2004, Korte 2001).
Finally, detection of threat by the amyg-dala has autonomic consequences. Connectionsfrom the amygdala to the brainstem lead tothe activation of the sympathetic nervous sys-tem, involving the release of epinephrine andNE from the adrenal medulla and NE fromthe terminals of sympathetic nerves through-out the body. Adrenal medullary hormones andsympathetic nerves produce an array of effects,including increasing blood pressure and heartrate, diverting stored energy to exercising mus-cle, and inhibiting digestion (Goldstein 2003,McCarty & Gold 1996, Sapolsky 2004). Com-ing full circle, fear circuitry is profoundly influ-enced by these endocrine and autonomic stressresponses. Thus, GCs travel in the circulationto the brain and affect the amygdala and a vari-ety of other structures. Although NE does notcross the blood-brain barrier, it may affect thebrain indirectly by binding to visceral afferentnerves, which transmit to the brain (McGaugh2003). Thus, the detection of threat and theconsequent activation of the fear system elicit avariety of effects that feed back onto the system
GR: glucocorticoidreceptor
that initiates and modulates emotional process-ing (McEwen 2003, Pare 2003, Sapolsky 2003).With this overview we now turn to a more de-tailed consideration of how the fear system in-teracts with stress.
HOW DO STRESS,NOREPINEPHRINE,AND GLUCOCORTICOIDSINFLUENCE DIFFERENT PHASESOF FEAR CONDITIONING?
In the remainder of this review we examinehow stress and its biochemical concomitants(Charmandari et al. 2005, Sapolsky 2004) in-fluence the acquisition, consolidation, recon-solidation, and extinction of conditioned fear,as well as the underlying neural mechanismsresponsible for different elements of fear pro-cessing. Throughout, we compare animal andhuman studies and their implications for stress-induced psychopathology. Instead of an exhaus-tive survey of all pertinent studies, we highlightoverarching trends in the literature. To distin-guish effects on acquisition from consolidation,it is necessary to compare pretraining manipu-lations with immediate posttraining manipula-tions (Rodrigues et al. 2004). This comparisonis necessary because the effects of stress or drugstend to last beyond the short training sessionin such studies (sometimes only a single train-ing trial). If pretraining manipulations disruptSTM but posttraining manipulations do not,the effect is said to be on acquisition (thoughan effect on STM itself cannot be ruled out).If posttraining manipulations leave STM intactbut disrupt LTM, then the effect is said to be onLTM consolidation. Another important manip-ulation involves treatments given immediatelybefore testing LTM. This treatment is called anexpression test. However, many studies exam-ine pretraining or postraining effects only onLTM but not on STM or on acquisition. Sim-ilarly, relatively few studies observe effects onexpression. Therefore, more investigations areneeded to fully determine how stress, NE, andGCs influence different phases of fear learning
www.annualreviews.org • Stress Hormones and Fear Circuitry 295
Ann
u. R
ev. N
euro
sci.
2009
.32:
289-
313.
Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
Uni
vers
iteit
van
Am
ster
dam
on
03/2
5/10
. For
per
sona
l use
onl
y.
ANRV379-NE32-13 ARI 13 May 2009 8:31
and its expression in memory. Below we dis-cuss what has been discovered so far. Althoughvariability in protocols makes it difficult to com-
pare stress manipulations directly across stud-ies, we make distinctions between acute andchronic stress throughout.
Amygdala
Hippocampus
PFCPFC
Sensory Sensory thalamus thalamus
Sensory Sensory cortexcortex
PFC
Sensory thalamus
Release of stress hormones
Environmentalthreat
(slow)(slow)(slow)
MonoamineMonoaminesystemssystems
Monoaminesystems
Feedback loop
MedullaCortex
PVN PVN PVN
ACTH
SNS
Epinephrine andnorepinephrine
Epinephrine andnorepinephrine
• Increase in cardiovascular tone
• Increase in blood pressure
• Mobilization of stored energy to muscle
• Transient enhancement of immunity
• Inhibition of costly, long-term processes such as growth and reproduction
Glucocorticoids
Glucocorticoids
Pituitary
Functional connectivity Processing threat stimulus
c d
a b
ARs
(fast)(fast)(fast)
Sensory Sensory thalamus thalamus
Sensory Sensory cortexcortex
Sensory thalamus
Sensory cortex
Sensory cortex
Vagusnerve
NTSNTSNTS
MRs and GRs
Kidney
Adrenalgland
Adrenal glandAddren
296 Rodrigues · LeDoux · Sapolsky
Ann
u. R
ev. N
euro
sci.
2009
.32:
289-
313.
Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
Uni
vers
iteit
van
Am
ster
dam
on
03/2
5/10
. For
per
sona
l use
onl
y.

ANRV379-NE32-13 ARI 13 May 2009 8:31
ACQUISITION OF FEARCONDITIONING
The Effects of Stress on theAcquisition of Fear Conditioning
Although many studies have looked at the ef-fects of pretraining stress on the LTM of fearconditioning (see below), influences of pre-training stress on acquisition, as tested in STM,have not been formally assessed. However,postshock freezing levels during fear condi-tioning are greater in chronically stressed ratsthat are classified as highly reactive, as indexedby their locomotion in a novel environment(Cordero et al. 2003a). This classification sug-gests that at least in some conditions, stress mayinfluence behavior during the learning pro-cess. However, this possibility needs furtherstudy because postshock freezing during acqui-sition reflects contextual conditioning as muchas cued.
The Effects of NE on the Acquisitionof Fear Conditioning
Locus coeruleus lesions or pretraining infu-sion of propranolol, a beta adrenergic receptorantagonist, into the amygdala (LA/B) blocksSTM, and consequently LTM (Bush et al. 2006;Neophytou et al. 2001). This finding is consis-tent with an effect on acquisition and its expres-sion in STM.
The Effects of GCs on the Acquisitionof Fear Conditioning
The contribution of GCs to fear acquisitionhas been studied by either removing their ef-fects via adrenalectomy, blocking GRs in theamygdala with an antagonist, or infusing a
viral vector into the amygdala prior to training.Although more STM evaluations are needed todetermine the precise time course of GCs onfear, adrenalectomized rats show reduced con-textual LTM but no immediate memory im-pairments (Pugh et al. 1997b). Likewise, GRblockade in the amygdala (LA/B) or hippocam-pus disrupts contextual LTM but leaves post-shock levels of freezing intact (Donley et al.2005). A similar pattern emerges from pretrain-ing intra-amygdala (LA/B) administration of aviral vector expressing a gene to blunt GR sig-naling, which effectively disrupts both cued andcontextual LTM without affecting acquisitionor postshock levels of freezing (Rodrigues &Sapolsky 2009). These studies suggest that GCsmay not be involved in acquisition so much asin the consolidation of fear LTM. We discussadditional results that support this claim in thefollowing section.
CONSOLIDATION OFLONG-TERM MEMORY
Studies assessing memory consolidation typ-ically administer manipulations immediatelyafter training (Dudai 2004, McGaugh 2000,Rodrigues et al. 2004). If such manipulationsfail to disrupt STM but do disrupt LTM, con-solidation (conversion of STM to LTM) is saidto be affected.
The Effects of Stress on theConsolidation of LTM ofFear Conditioning
Pretraining exposure to both acute and chronicstressors can enhance LTM of both cuedand contextual fear conditioning for at least
←−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−Figure 2Overview of fear arousal and the stress response. Upper left: Black arrows indicate the functional connectivity of brain structuresinvolved in fear conditioning. Upper right: Red arrows represent the activation of brain areas recruited during the processing ofthreatening stimuli. Bottom Left: The stress response leads to the release of stress hormones. Bottom Right: Dashed lines indicatefeedback of the stress response to neural structures involved in fear conditioning. ACTH, adrenocorticotropic hormone; CRH,corticotropin-releasing factor; GRs, glucocorticoid receptors; MRs, mineralocorticoid receptors; NTS: nucleus of the solitary tract;PVN, periventricular nucleus of the hypothalamus; SNS, sympathetic nervous system.
www.annualreviews.org • Stress Hormones and Fear Circuitry 297
Ann
u. R
ev. N
euro
sci.
2009
.32:
289-
313.
Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
Uni
vers
iteit
van
Am
ster
dam
on
03/2
5/10
. For
per
sona
l use
onl
y.

ANRV379-NE32-13 ARI 13 May 2009 8:31
3 months (Conrad et al. 1999; Cordero et al.2003a,b; Kohda et al. 2007; Rau et al. 2005; Rau& Fanselow 2008). Acute posttraining stressalso enhances LTM for cued fear conditioning(Hui et al. 2006). Furthermore, preexposureto stress sensitizes fear-conditioning responsessuch that less-intense stressors can produce ro-bust fear behaviors, which may help explainwhy individuals with PTSD react strongly tomild stimuli and quickly form new fears (Rauet al. 2005). Because measurements of pretrain-ing stress on acquisition and STM are lackingin the literature, more research is needed to ad-dress the temporal dynamics of stress on fearconditioning.
The Effects of NE on theConsolidation of LTMof Fear Conditioning
Central NE depletion or pretraining infu-sions of propranolol into the LA/B priorto conditioning impair auditory fear LTM(Bush et al. 2006, Selden et al. 1990). Inaddition, systemic injections of epinephrine,which leads to the release of NE in the brain(McGaugh 2003), enhance contextual fearLTM (Frankland et al. 2004, Hu et al. 2007) viaa facilitation of AMPA (α-amino-3-hydroxyl-5-methyl-4-isoxazole-propionate) receptor in-sertion in the hippocampus (Hu et al. 2007).However, posttraining administration of pro-pranolol systemically or into the LA/B does notaffect LTM consolidation of auditory or contex-tual fear conditioning (Bush et al. 2006, Debiec& Ledoux 2004, Lee et al. 2001). Thus, NEin the amygdala does not seem critical for theconsolidation of fear conditioning, but it doesseem to interact with GCs to fortify memories(Roozendaal et al. 2006).
Posttraining intrahippocampal propranololinfusions given immediately, but not six hoursafter training, block LTM, but not STM, ofcontextual fear conditioning ( Ji et al. 2003).Furthermore, systemic or intrahippocampal in-fusions given before testing one day after condi-tioning impair the retrieval of contextual mem-ories (Murchison et al. 2004).
In humans, systemic NE blockade dis-rupts the consolidation of contextual, but notcued, fear conditioning LTM (Grillon et al.2004). Studies involving avoidance condition-ing, an indirect means of measuring fear, showthat posttraining systemic and intra-amygdala(LA/B) pharmacological manipulations showthat NE is crucial to consolidate avoidancelearning (Ferry & McGaugh 1999, Gallagheret al. 1977, Liang et al. 1986, Quirarte et al.1997, Roozendaal & McGaugh 1997). More-over, NE levels following avoidance trainingcorrelate with subsequent retention (McGaughet al. 2002, McIntyre et al. 2002). Thus, directmeasurement of fear responses with fear condi-tioning gives a different pattern of results thando indirect measures with avoidance condition-ing. More work systematically examining theseeffects is needed.
The Effects of GCs on theConsolidation of LTMof Fear Conditioning
Pretraining systemic, intrahippocampal, orintra-amygdala (LA/B) manipulation of GCsinfluences the LTM of fear conditioning; con-textual memories are more vulnerable (Conradet al. 2004; Cordero et al. 2002; Donley et al.2005; Pugh et al. 1997a,b) likely because con-textual fear memory is generally weaker (i.e.,harder to learn and easier to disrupt) than cuedfear memory (Phillips & LeDoux 1992).
Posttraining systemic manipulations of GCsalso impact both auditory and contextualfear memory consolidation (Corodimas et al.1994, Hui et al. 2004, Pugh et al. 1997a,Zorawski & Killcross 2002). Moreover, post-training GR blockade in the LA/B affects au-ditory fear LTM but not STM ( Jin et al.2007), providing more evidence that GCs aremost important for consolidation processes.Furthermore, posttraining GCs administeredimmediately, but not 3 hours, after auditoryfear conditioning facilitates freezing to a tonepreviously paired with a US but does not al-ter responses to unpaired tones or to toneor shock alone, suggesting a selective and
298 Rodrigues · LeDoux · Sapolsky
Ann
u. R
ev. N
euro
sci.
2009
.32:
289-
313.
Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
Uni
vers
iteit
van
Am
ster
dam
on
03/2
5/10
. For
per
sona
l use
onl
y.

ANRV379-NE32-13 ARI 13 May 2009 8:31
time-dependent role for GC-facilitated mem-ory of the tone-shock association (Hui et al.2004). In addition, elevation of GCs sys-temically or in the LA/B enhances the re-trieval of fear memories in avoidance paradigms(Izquierdo et al. 2002, Roozendaal & McGaugh1997), and this enhancement can be effectivefor memories acquired from one day to manymonths before (Izquierdo et al. 2002). Finally,in humans, high GC levels positively correlatewith fear memory consolidation (Zorawski et al.2006).
RECONSOLIDATION
The Effects of Stress on theReconsolidation of Fear ConditioningPreliminary evidence suggests that acute stressbefore, but not after, memory reactivation dis-rupts auditory fear reconsolidation LTM (Bushet al. 2004). It would be interesting to see howchronic stress or intra-amygdala manipulationsinfluence this phenomenon and the time courseof reconsolidation processes.
The Effects of NE Manipulationon the Reconsolidation of FearConditioningAlthough the effects of prereactivation NE ma-nipulations on reconsolidation are unknown,systemic or intra-amygdala (LA/B) adminis-tration of propranolol after reactivation dis-rupts reconsolidation of auditory and contex-tual fear LTM in rats (Abrari et al. 2008,Debiec & Ledoux 2004). The same trend holdstrue for avoidance conditioning (Przybyslawskiet al. 1999). These findings are consonant withreports that propranolol can effectively treatPTSD when administered after aversive mem-ory activation (Brunet et al. 2008, Orr et al.2006, Pitman & Delahanty 2005, Pitman et al.2002).
The Effects of GC Manipulationon the Reconsolidationof Fear ConditioningHow alterations of GC activity before reactiva-tion might influence reconsolidation has yet to
be investigated. After reactivation, GR antago-nism in the LA/B disrupts LTM but not STMof auditory fear reconsolidation; this effect oc-curs if blockade is immediate, but not 6 hours,after reactivation. Furthermore, this immedi-ate postreactivation blockade is effective both1 day and 10 days after conditioning ( Jin et al.2007). Likewise, GR blockade in the LA/B im-mediately after retrieval blocks reconsolidationin an avoidance paradigm (Tronel & Alberini2007).
EXTINCTION
The Effects of Stress on the Extinctionof Fear Conditioning
Chronic stress prior to conditioning impairsthe LTM of auditory and contextual extinctionLTM (Garcia et al. 2008, Miracle et al. 2006);similarly, patients with PTSD show impairedcued fear extinction (Blechert et al. 2007, Wessa& Flor 2007). However, reports conflict regard-ing the influence of stress on the learning ofextinction (Izquierdo et al. 2006, Miracle et al.2006), perhaps because of differences in stresstypes or protocols or species (mice versus rats).Stress induction after fear conditioning but be-fore extinction does not influence extinctionLTM (G. Quirk, unpublished results). If a stres-sor is controllable, the mPFC inhibits activationof brainstem nuclei (Amat et al. 2005). More-over, stress and environmental enrichment pro-duce opposite effects on fear renewal in a newcontext; the influence of enrichment outweighsthat of stress (Mitra & Sapolsky 2009).
Involvement of NE in the Extinctionof Fear Conditioning
NE in the LA/B after extinction train-ing enhances the consolidation of extinctionmemories of contextual fear LTM (Berlau &McGaugh 2006). Furthermore, NE efflux in-creases in the PFC during presentation of aCS previously paired with a US (Feenstra et al.2001). Similarly, administration of propranololin the IL subregion of the mPFC before, but
www.annualreviews.org • Stress Hormones and Fear Circuitry 299
Ann
u. R
ev. N
euro
sci.
2009
.32:
289-
313.
Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
Uni
vers
iteit
van
Am
ster
dam
on
03/2
5/10
. For
per
sona
l use
onl
y.

ANRV379-NE32-13 ARI 13 May 2009 8:31
not after, extinction training impairs retrievalof extinction the following day while leavingwithin-session extinction intact. Furthermore,NE signaling in the IL strengthens extinctionmemory (Mueller et al. 2008).
Involvement of GCs in the Extinctionof Fear Conditioning
Chronic GC exposure, which decreasesendogenous GC secretion, prior to fear condi-tioning impairs contextual extinction LTM butnot STM and results in a decrease of NMDA(N-methyl-d-aspartate) and AMPA (α-amino-3-hydroxyl-5-methyl-4-isoxazole-propionate)excitatory receptor subunits in the mPFC(Gourley et al. 2008). In addition, systemicand intra-amygdala (LA/B) GC manipulationsprior to extinction training suggest that GRactivation is key for extinction learning in afear-potentiated startle paradigm (Yang et al.2006). Although systemic GC administrationbefore extinction training does not produceextinction learning or LTM impairments (G.Quirk, unpublished results), metyrapone,a GC synthesis inhibitor, affects extinctionLTM while leaving learning intact (Barrett &Gonzalez-Lima 2004). Moreover, adrenergicreceptor (AR) blockade or GR activation afterretrieval disrupts recall of contextual fear con-ditioning, but only the effects of GR activationare reversed by a reminder shock (Abrari et al.2008). Along these same lines, GR-mediateddisruptions of fear memories show sponta-neous recovery and renewal of CRs (Barrett& Gonzalez-Lima 2004, Cai et al. 2006).However, GR inactivation impairs CRs in anavoidance task, and these do not reemergeafter strong reminders (Tronel & Alberini2007), possibly because of the involvement ofother processes in this paradigm. Nonetheless,the results from animal studies that show GCactivity may promote extinction (Barrett &Gonzalez-Lima 2004, Cai et al. 2006) parallelfindings that PTSD and phobia symptoms,but not general anxiety, can improve with GCtreatment (Aerni et al. 2004, de Quervain 2008,Schelling et al. 2004, Soravia et al. 2006).
SUMMARY OF THE EFFECTSON FEAR CONDITIONING
Table 1 summarizes results from animal stud-ies presented in this review. This informationcomes with a number of caveats: First, data wereoften gathered from only a very small number ofpapers for each topic; second, few studies exam-ined both contextual and cued fear condition-ing; third, it was rarely the case that both STMand LTM were examined; fourth, little workhas examined effects of stress, NE, or GCs onthe expression of fear; and fifth, the importantmodulatory effects of age, genetics, and genderhave not been discussed throughout this reviewin the interest of space.
HOW STRESS AFFECTSFEAR CIRCUITRY ONA CELLULAR LEVEL
The Influence of Stress onthe Morphology of Neuronsin Fear Circuitry
Stress can alter the morphology of pyramidalneurons in structures involved in fear circuitry(see Figure 3). To begin, chronic stress causesan increase in the dendritic arborization, spineformation, and synaptic connectivity in princi-pal neurons in the LA/B (Vyas et al. 2002, 2006)but not in the CE (Vyas et al. 2003). Moreover,chronic immobilization stress, but not chronicunpredictable stress, induces this dendritic re-modeling in the LA/B (Vyas & Chattarji 2004).In addition, the duration of stress directly im-pacts the spatiotemporal patterns of spine for-mation such that both acute and chronic stres-sors increase spines in LA/B neurons, but onlythe latter has an effect on dendritic arbors(Mitra et al. 2005). Furthermore, a single, acutedose of GCs is sufficient to produce anxietyand hypertrophy of LA/B neurons (Mitra &Sapolsky 2008). Contrasting with these ef-fects, stress decreases dendritic branching andspine count in the hippocampus (for review, seeMcEwen & Magarinos 2001).
Neurons in the mPFC endure morpholog-ical changes due to stress and GCs, including
300 Rodrigues · LeDoux · Sapolsky
Ann
u. R
ev. N
euro
sci.
2009
.32:
289-
313.
Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
Uni
vers
iteit
van
Am
ster
dam
on
03/2
5/10
. For
per
sona
l use
onl
y.

ANRV379-NE32-13 ARI 13 May 2009 8:31
Tab
le1
Loc
aliz
atio
nof
man
ipul
atio
ns:s
yste
mic
/bra
in-w
ide
(bla
ck),
LA
/B(r
ed),
hipp
ocam
pus
(blu
e),a
ndm
PFC
(gre
en)a,
b.P
osts
hock
free
zing
findi
ngs
incl
uded
inco
ntex
tual
STM
resu
lts
Con
diti
onin
gR
econ
solid
atio
nE
xtin
ctio
nP
retr
ain-
ing
Pos
ttra
in-
ing
Pre
test
-in
gP
rere
acti
va-
tion
Pos
trea
ctiv
a-ti
onP
reco
ndit
ion-
ing
Pre
exti
ncti
ontr
aini
ngP
oste
xtin
ctio
ntr
aini
ngST
RE
SSC
ued
STM
??
??
?×∗
×?
LTM
↑↑
?↑
×↓
×?
Con
text
ual
STM
+?
??
??
??
LTM
↑?
??
?↓
??
NE
Cue
dST
M↑
??
??
?×
×LT
M↑↑
קק
??
↑↑?
↑×
Con
text
ual
STM
↑×
??
??
??
LTM
↑×↑
↑↑?
↑?
?↑
GC
sC
ued
STM
××
??
×?
×?
LTM
×∗↑
↑↑?
?↑
?×∗
?C
onte
xtua
lST
M×
××
??
??
×?
?LT
M↑↑
↑?
??
↓?
↑↑
a Abb
revi
atio
ns:L
A/B
,lat
eral
/bas
alnu
cleu
sof
the
amyg
dala
;GC
,glu
coco
rtic
oids
;LT
M,l
ong-
term
mem
ory;
mP
FC,m
edia
lpre
fron
talc
orte
x;N
E,n
orep
inep
hrin
e;ST
M,s
hort
-ter
mm
emor
y.bK
ey:
↑,St
ress
,NE
,or
GC
sfa
cilit
ate
fear
mem
ory
here
.↓,
Stre
ssor
GC
sim
pair
fear
mem
ory
here
.×
,Str
ess,
NE
,or
GC
sm
anip
ulat
ions
prod
uce
noin
fluen
ces
atth
istim
e.?,
Eff
ects
unkn
own.
∗ ,con
flict
ing
repo
rts.
§,e
ffec
tivel
ydi
srup
tsco
nsol
idat
ion
ofav
oida
nce
lear
ning
,but
notf
ear
cond
ition
ing.
www.annualreviews.org • Stress Hormones and Fear Circuitry 301
Ann
u. R
ev. N
euro
sci.
2009
.32:
289-
313.
Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
Uni
vers
iteit
van
Am
ster
dam
on
03/2
5/10
. For
per
sona
l use
onl
y.

ANRV379-NE32-13 ARI 13 May 2009 8:31
Control Stress
500 nm
500 nm
500 nm
Amygdala (LA/B)
Hippocampus (CA3)
Prefrontal cortex (IL)
Figure 3Influences of stress on the morphology of neurons in fear circuitry. LA/B,lateral/basal nucleus of the amygdala; CA3, cornu ammonis subfield of thehippocampus; IL, infralimbic subregion of the medial prefrontal cortex.Reconstructed Golgi-impregnated neuron images reproduced with permissionfrom Vyas et al. (2002) and Izquierdo et al. (2006).
atrophy and spine loss (Brown et al. 2005; Cook& Wellman 2004; Czeh et al. 2008; Izquierdoet al. 2006; Radley et al. 2004, 2006, 2008;Wellman 2001). Furthermore, atrophy in ILneurons through brief uncontrollable stress is
accompanied by a resistance to fear extinction(Izquierdo et al. 2006). Stress-induced atrophyof hippocampal (Conrad et al. 1999, McEwen1999, Sousa et al. 2000) and mPFC neurons(Radley et al. 2005) reverses after a stress-free period. However, the same recovery perioddoes not rescue stress-induced hypertrophy ofamygdala neurons (Vyas et al. 2004).
The Influence of Stress on theElectrophysiological Propertiesof Neurons in Fear Circuitry
Stress and its accompanying biological effectsinfluence the electrophysiological activity ofneurons in fear circuitry (see Figure 4). Elec-trophysiological studies often explore neuronalexcitability, as well as long-term potentiation(LTP), an artificial means of inducing synap-tic connectivity between two brain areas, andthus a physiological model of fear learning andmemory (Blair et al. 2001, Rodrigues et al. 2004,Schafe et al. 2001). Stress has differential effectson the amygdala, hippocampus, and prefrontalcortex, which we now review.
LA/B neurons from stressed animals displayincreased firing rates and greater respon-siveness (Garcia et al. 1998, Kavushanskyet al. 2006, Kavushansky & Richter-Levin2006, Rodrıguez Manzanares et al. 2005).Although some stress paradigms, especiallyacute inductions, enhance LTP in the LA/B(Rodrıguez Manzanares et al. 2005, Vouimbaet al. 2004, 2006), other stress protocolssuppress it (Kavushansky & Richter-Levin2006, Kavushansky et al. 2006, Kohda et al.2007). Along these same lines, the type ofstress is also a critical factor in the influenceof stress on hippocampal plasticity such thatchronic, but not acute, stress suppresseshippocampal LTP (Pavlides et al. 2002).Temporal qualities of stressors have oppositeinfluences on the hippocampus, boosting LTPwhen the tetanizing stimulation is in closeproximity to emotional processing and dis-rupting LTP when there is a substantial delaybetween the stimulation and initiation of stress.Furthermore, amygdala (LA/B) stimulation
302 Rodrigues · LeDoux · Sapolsky
Ann
u. R
ev. N
euro
sci.
2009
.32:
289-
313.
Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
Uni
vers
iteit
van
Am
ster
dam
on
03/2
5/10
. For
per
sona
l use
onl
y.

ANRV379-NE32-13 ARI 13 May 2009 8:31
mimics this emotion-induced influence onhippocampal plasticity (LA/B) (Diamond et al.2007, Richter-Levin 2004, Richter-Levin &Akirav 2003). Hippocampal stimulation afterfear extinction training disrupts the recall ofextinction (Garcia et al. 2008). Furthermore,uncontrollable stress drastically impairs hip-pocampal LTP relative to the influence of thesame amount of controllable stress (Shors et al.1989). The mPFC seems to play a role indetermining the controllability of stress (Amatet al. 2005), and neurons here display unique
patterns of firing rates that encode fast andslow responses to stress. Whereas some mPFCneurons respond phasically during the durationof a stressor, others show activity that persistsafter the stress ceases, and yet others respondduring the initiation and termination of stress( Jackson & Moghaddam 2006). Furthermore,stress reduces LTP induction in projectionsfrom both the LA/B (Maroun & Richter-Levin 2003) and hippocampus (Cerqueiraet al. 2007, Rocher et al. 2004) to thePFC.
Dendrite
LA/B neuron
cell body
Decrease of GABA-mediated inhibition
GABA-Areceptor
Axon
GABACl-
Enhancement of neuronalresponses and excitability
Alteration of plasticity thatunderlies fear learning
LTP induction
Time (min)
100
200
Pe
rce
nt
of
con
tro
l
0 50
Figure 4Influence of stress, norepinephrine (NE), and glucocorticoids (GCs) on the electrophysiological propertiesof lateral (LA)/basal nucleus (B) neurons of the amygdala. Representative traces and long-term potentiation(LTP) data reproduced with permission from Tully et al. (2007).
www.annualreviews.org • Stress Hormones and Fear Circuitry 303
Ann
u. R
ev. N
euro
sci.
2009
.32:
289-
313.
Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
Uni
vers
iteit
van
Am
ster
dam
on
03/2
5/10
. For
per
sona
l use
onl
y.

ANRV379-NE32-13 ARI 13 May 2009 8:31
The Influence of NE on theElectrophysiological Propertiesof Neurons in Fear Circuitry
NE increases the spiking of neurons in the LAand facilitates the induction of LTP (Tully et al.2007). What’s more, fear conditioning increasesdepolarization-evoked NE release (Liu et al.2007). Moreover, ARs seem particularly pivotalfor a late phase of LTP in the LA, which persistsfor at least three hours and requires the synthe-sis of new mRNA and protein (Huang et al.2000). The reactivity of the hippocampal neu-rons is enhanced by NE (Harley 2007, Segalet al. 1991), and suppression of GC secretionincreases NE transmission here (Mizoguchiet al. 2008); the relevance of this to fear learningand memory processes is unclear. In the IL ofthe mPFC, fear conditioning decreases, and ex-tinction increases, neuronal excitability (Santiniet al. 2008). Pertinent to this, NE signaling inthis region increases neuron excitability (Barthet al. 2007, Mueller et al. 2008) and strength-ens the memory for extinction (Mueller et al.2008).
The Influence of GCs on theElectrophysiological Propertiesof Neurons in Fear Circuitry
Both high and low GC concentrations en-hance the excitability of LA/B neurons (Du-varci & Pare 2007, Kavushansky & Richter-Levin 2006). In addition, GCs depolarize theresting potential, increase input resistance, andsignificantly decrease spike-frequency adapta-tion of LA/B neurons (Duvarci & Pare 2007).The influences of GCs are different on thehippocampus; there low concentrations (viaMR activation) are excitatory, whereas highconcentrations (via MR plus GR activation)are inhibitory (de Kloet et al. 1999, Joels& de Kloet 1992). A similar biphasic relation-ship of MR and GR activation exists for en-hancing and suppressing effects of GCs onhippocampal LTP, respectively (Pavlides et al.1996, Pavlides & McEwen 1999). We do not
know how GCs directly influence the elec-trophysiological profiles of mPFC neurons.Therefore, more data are needed to illustratehow GCs interact with other factors to influ-ence plasticity in structures that modulate fearconditioning.
CONCLUSIONS
An extensive literature has demonstrated theways in which stress, and the endocrine media-tors of the stress response, can impact explicit,declarative memory processes (McEwen 1999).Though based on a smaller literature, this re-view highlights the ability of stress and arousal-triggered fear to impact emotional learningand memory, as assessed by fear condition-ing. Indeed, stressful experiences, NE, and GCsalter the morphological and electrophysiologi-cal characteristics of neurons in areas that play avital role in fear processing, including the amyg-dala (LA/B), hippocampus, and PFC. As a re-sult, they can have a powerful influence on fearconditioning.
In the laboratory, investigators use a num-ber of models to induce stress, includingexposure to predator odor, restraint, foot shock,forced swimming, saline injections, and coldtemperatures. Furthermore, the duration ofstress manipulations can be acute, moderate, orchronic. Although these differences make directcomparisons between studies difficult, stereo-typical responses to these various stressors dooccur and activate the brain’s fear system,thus initiating the stress response in both thebody and the brain. Nonetheless, because stressmagnitude and duration differentially impactthe morphological and electrophysiologicalproperties of fear circuitry neurons, it wouldbe important for future studies to compareand contrast the influence of different stressprotocols.
Existing studies show that stress before orafter training boosts LTM of fear condition-ing. In addition, stress before, but not after,reactivation enhances reconsolidation. Finally,stress before fear conditioning, but not before
304 Rodrigues · LeDoux · Sapolsky
Ann
u. R
ev. N
euro
sci.
2009
.32:
289-
313.
Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
Uni
vers
iteit
van
Am
ster
dam
on
03/2
5/10
. For
per
sona
l use
onl
y.

ANRV379-NE32-13 ARI 13 May 2009 8:31
extinction training, interferes with extinctionLTM. Altogether, these findings suggest thatduring initial learning, stress activates eventsthat can influence later consolidation and ex-tinction processes. Whether effects exist on theacquisition and expression of fear is less clearowing to the relative paucity of studies.
In regard to NE, cumulative evidence sug-gests that NE is important for fear condition-ing, reconsolidation, and extinction, but not forconsolidation. NE increases neuronal excitabil-ity and LTP in the LA, and its activity there isneeded for both STM and LTM of fear condi-tioning. NE also increases neuronal excitabilityin the IL of the mPFC and is important for theLTM of extinction. Because pretraining but notposttraining intra-amygdala (LA/B) or intra-mPFC infusions of propranolol block LTM offear conditioning and extinction, respectively,it seems that NE must be tonically active dur-ing learning to promote rapid postlearning cas-cades in the LA and IL to facilitate STM re-trieval and to promote the conversion of STMto LTM.
Next, GCs appear to be particularlycrucial for LTM, but not the acquisition,of fear conditioning. Therefore, the overalltrend of systemic or intra-amygdala (LA/B) GCdisrupting LTM is consonant with electrophys-iological findings that GCs have long-term, butnot short-term, enhancing effects on neuronalexcitability in LA/B (Duvarci & Pare 2007).GRs are located both at synaptic and at nuclearsites in the LA. The classic actions of GR, asa steroid receptor, involve binding GCs in thecytoplasm and acting as a transcription factorupon translocating to the nucleus, whereasless traditional GR actions include rapid,nongenomic effects. The fact that GRs seemmore important for fear consolidation than forlearning, and for enduring but not immediateinfluences on the firing of LA/B neurons,
suggests that more traditional genomic effectsunderlie these behavioral and electrophysio-logical findings.
Stress-induced enhancement of LTP in au-ditory inputs to the LA alongside disruption ofLTP in the projections to the PFC from thehippocampus and the LA/B is congruent withthe notion that powerful fear inputs can facil-itate strong emotional pathways at the cost ofactivating more regulatory pathways. Indeed,an abundant literature showing that NE is im-portant particularly for hippocampal and PFCfunction, in addition to LA/B activation, is inline with NE’s role in arousal and cognition.
Given that hippocampal function is dis-rupted by major stressors or exposure to ele-vated levels of GCs, how might it play a pivotalrole in forming fear memories, particularly con-textual ones? This answer might be explained bythe model of the hippocampus shifting from acognitive mode to a flashbulb memory modeunder intense stress, which explains why thecontextual memories of traumatic experiencesare vivid and endure for many years but tend tobe disjointed and fragmented (Diamond et al.2007).
In summary, stress, NE, and GCs appear tobe potent modulators of fear memory forma-tion. Furthermore, the ability of stress, NE,and GCs to decrease GABA (γ -aminobutyricacid)-A receptor-mediated inhibition (Duvarci& Pare 2007, Rodriguez Manzanares et al.2005, Tully et al. 2007), thereby allowing forincreased excitability in LA/B, is harmoniouswith the fact that benzodiazepines act on thisreceptor to decrease anxiety.
During difficult times, we are often advisedto “use our heads” or “follow our hearts” or “gowith our gut.” As this review highlights, ourbodies and our minds really are not separate,but instead mutually inform each other on howto process the emotional events of our lives.
DISCLOSURE STATEMENT
The authors are not aware of any affiliations, memberships, funding, or financial holdings thatmight be perceived as affecting the objectivity of this review.
www.annualreviews.org • Stress Hormones and Fear Circuitry 305
Ann
u. R
ev. N
euro
sci.
2009
.32:
289-
313.
Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
Uni
vers
iteit
van
Am
ster
dam
on
03/2
5/10
. For
per
sona
l use
onl
y.

ANRV379-NE32-13 ARI 13 May 2009 8:31
ACKNOWLEDGMENTS
We apologize to all the investigators whose research could not be appropriately cited owingto space limitations. We extend our sincere gratitude to our many wonderful collaborators forthoughtful discussions pertaining to the topic and data of this review. Research by S.M.R. andR.M.S. on this topic supported by NIH Grant 5R01 AG020633. J.E.L. supported by NIH GrantsP50 MH058911, R01 MH046516, R37 MH037884, and K05 MH067048.
LITERATURE CITED
Abrari K, Rashidy-Pour A, Semnanian S, Fathollahi Y. 2008. Administration of corticosterone after memoryreactivation disrupts subsequent retrieval of a contextual conditioned fear memory: dependence upontraining intensity. Neurobiol. Learn. Mem. 89:178–84
Aerni A, Traber R, Hock C, Roozendaal B, Schelling G, et al. 2004. Low-dose cortisol for symptoms ofposttraumatic stress disorder. Am. J. Psychiatry 161:1488–90
Amaral DG, Insausti R. 1992. Retrograde transport of D-[3H]-aspartate injected into the monkey amygdaloidcomplex. Exp. Brain Res. 88:375-88
Amat J, Baratta MV, Paul E, Bland ST, Watkins LR, Maier SF. 2005. Medial prefrontal cortex determineshow stressor controllability affects behavior and dorsal raphe nucleus. Nat. Neurosci. 8:365–71
Anglada-Figueroa D, Quirk GJ. 2005. Lesions of the basal amygdala block expression of conditioned fear butnot extinction. J. Neurosci. 25:9680–85
Aston-Jones G. 2004. Locus coeruleus, A5 and A7 noradrenergic cell groups. In The Rat Nervous System, ed.G Paxinos, pp. 259–335. Amsterdam/Boston: Elsevier Acad.
Aston-Jones G, Cohen JD. 2005. Adaptive gain and the role of the locus coeruleus-norepinephrine system inoptimal performance. J. Comp. Neurol. 493:99–110
Balleine BW, Killcross S. 2006. Parallel incentive processing: an integrated view of amygdala function. TrendsNeurosci. 29:272–79
Barrett D, Gonzalez-Lima F. 2004. Behavioral effects of metyrapone on Pavlovian extinction. Neurosci. Lett.371:91–96
Barth AM, Vizi ES, Lendvai B. 2007. Noradrenergic enhancement of Ca2+ responses of basal dendrites inlayer 5 pyramidal neurons of the prefrontal cortex. Neurochem. Int. 51:323–27
Berlau DJ, McGaugh JL. 2006. Enhancement of extinction memory consolidation: the role of the noradren-ergic and GABAergic systems within the basolateral amygdala. Neurobiol. Learn. Mem. 86:123–32
Blair HT, Schafe GE, Bauer EP, Rodrigues SM, LeDoux JE. 2001. Synaptic plasticity in the lateral amygdala:a cellular hypothesis of fear conditioning. Learn. Mem. 8:229–42
Blanchard RJ, Blanchard DC. 1969. Crouching as an index of fear. J. Comp. Physiol. Psychol. 67:370–75Blechert J, Michael T, Vriends N, Margraf J, Wilhelm FH. 2007. Fear conditioning in posttraumatic stress
disorder: evidence for delayed extinction of autonomic, experiential, and behavioural responses. Behav.Res. Ther. 45:2019–33
Bouton ME, Westbrook RF, Corcoran KA, Maren S. 2006. Contextual and temporal modulation of extinction:behavioral and biological mechanisms. Biol. Psychiatry 60:352–60
Brown SM, Henning S, Wellman CL. 2005. Mild, short-term stress alters dendritic morphology in rat medialprefrontal cortex. Cereb. Cortex 15:1714–22
Brunet A, Orr SP, Tremblay J, Robertson K, Nader K, Pitman RK. 2008. Effect of post-retrieval propranololon psychophysiologic responding during subsequent script-driven traumatic imagery in post-traumaticstress disorder. J. Psychiatr. Res. 42:503–6
Bush DEA, Gekker A, LeDoux JE. 2006. Blockade of amygdala beta-adrenergic receptors disrupts fear con-ditioning. Soc. Neurosci. 370.7
Bush DEA, McEwen BS, LeDoux JE. 2004. Acute restraint stress enhances the reconsolidation of fear memory.Soc. Neurosci. 208.4
Cai WH, Blundell J, Han J, Greene RW, Powell CM. 2006. Postreactivation glucocorticoids impair recall ofestablished fear memory. J. Neurosci. 26:9560–66
306 Rodrigues · LeDoux · Sapolsky
Ann
u. R
ev. N
euro
sci.
2009
.32:
289-
313.
Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
Uni
vers
iteit
van
Am
ster
dam
on
03/2
5/10
. For
per
sona
l use
onl
y.

ANRV379-NE32-13 ARI 13 May 2009 8:31
Cain CK, LeDoux JE. 2008. Emotional processing and motivation: in search of brain mechanisms. In Handbookof Approach and Avoidance Motivation, ed. AJ Elliot, pp. 17–34. New York: Psychology Press
Cardinal RN, Parkinson JA, Hall J, Everitt BJ. 2002. Emotion and motivation: the role of the amygdala, ventralstriatum, and prefrontal cortex. Neurosci. Biobehav. Rev. 26:321–52
Cerqueira JJ, Mailliet F, Almeida OF, Jay TM, Sousa N. 2007. The prefrontal cortex as a key target of themaladaptive response to stress. J. Neurosci. 27:2781–87
Charmandari E, Tsigos C, Chrousos G. 2005. Endocrinology of the stress response. Annu. Rev. Physiol. 67:259–84
Conrad CD, LeDoux JE, Magarinos AM, McEwen BS. 1999. Repeated restraint stress facilitates fear condi-tioning independently of causing hippocampal CA3 dendritic atrophy. Behav. Neurosci. 113:902–13
Conrad CD, MacMillan DD 2nd, Tsekhanov S, Wright RL, Baran SE, Fuchs RA. 2004. Influence of chroniccorticosterone and glucocorticoid receptor antagonism in the amygdala on fear conditioning. Neurobiol.Learn. Mem. 81:185–99
Cook SC, Wellman CL. 2004. Chronic stress alters dendritic morphology in rat medial prefrontal cortex.J. Neurobiol. 60:236–48
Cordero MI, Kruyt ND, Merino JJ, Sandi C. 2002. Glucocorticoid involvement in memory formation in arat model for traumatic memory. Stress 5:73–79
Cordero MI, Kruyt ND, Sandi C. 2003a. Modulation of contextual fear conditioning by chronic stress in ratsis related to individual differences in behavioral reactivity to novelty. Brain Res. 970:242–45
Cordero MI, Venero C, Kruyt ND, Sandi C. 2003b. Prior exposure to a single stress session facilitatessubsequent contextual fear conditioning in rats. Evidence for a role of corticosterone. Horm. Behav.44:338–45
Corodimas KP, LeDoux JE, Gold PW, Schulkin J. 1994. Corticosterone potentiation of conditioned fear inrats. Ann. NY Acad. Sci. 746:392–93
Czeh B, Perez-Cruz C, Fuchs E, Flugge G. 2008. Chronic stress-induced cellular changes in the medialprefrontal cortex and their potential clinical implications: Does hemisphere location matter? Behav. BrainRes. 190:1–13
Debiec J, Ledoux JE. 2004. Disruption of reconsolidation but not consolidation of auditory fear conditioningby noradrenergic blockade in the amygdala. Neuroscience 129:267–72
de Kloet ER, Oitzl MS, Joels M. 1999. Stress and cognition: Are corticosteroids good or bad guys? TrendsNeurosci. 22:422–26
de Kloet ER. 2004. Hormones and the stressed brain. Ann. NY Acad. Sci. 1018:1–15de Quervain DJ. 2008. Glucocorticoid-induced reduction of traumatic memories: implications for the treat-
ment of PTSD. Prog. Brain Res. 167:239–47DeRijk R, de Kloet ER. 2005. Corticosteroid receptor genetic polymorphisms and stress responsivity. Endocrine
28:263–70Diamond DM, Campbell AM, Park CR, Halonen J, Zoladz PR. 2007. The temporal dynamics model of emo-
tional memory processing: a synthesis on the neurobiological basis of stress-induced amnesia, flashbulband traumatic memories, and the Yerkes-Dodson law. Neural Plast. 2007:1–33, Art. 60803
Donley MP, Schulkin J, Rosen JB. 2005. Glucocorticoid receptor antagonism in the basolateral amygdala andventral hippocampus interferes with long-term memory of contextual fear. Behav. Brain Res. 164:197–205
Dudai Y. 2004. The neurobiology of consolidations, or, how stable is the engram? Annu. Rev. Psychol. 55:51–86Dudai Y. 2006. Reconsolidation: the advantage of being refocused. Curr. Opin. Neurobiol. 16:174–78Duvarci S, Pare D. 2007. Glucocorticoids enhance the excitability of principal basolateral amygdala neurons.
J. Neurosci. 27:4482–91Fanselow MS, Poulos AM. 2005. The neuroscience of mammalian associative learning. Annu. Rev. Psychol.
56:207–34Feenstra MG, Vogel M, Botterblom MH, Joosten RN, de Bruin JP. 2001. Dopamine and noradrenaline
efflux in the rat prefrontal cortex after classical aversive conditioning to an auditory cue. Eur. J. Neurosci.13:1051–54
Fendt M, Fanselow MS. 1999. The neuroanatomical and neurochemical basis of conditioned fear.Neurosci. Biobehav. Rev. 23:743–60
www.annualreviews.org • Stress Hormones and Fear Circuitry 307
Ann
u. R
ev. N
euro
sci.
2009
.32:
289-
313.
Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
Uni
vers
iteit
van
Am
ster
dam
on
03/2
5/10
. For
per
sona
l use
onl
y.

ANRV379-NE32-13 ARI 13 May 2009 8:31
Ferry B, McGaugh JL. 1999. Clenbuterol administration into the basolateral amygdala post-training enhancesretention in an inhibitory avoidance task. Neurobiol. Learn. Mem. 72:8–12
Frankland PW, Josselyn SA, Anagnostaras SG, Kogan JH, Takahashi E, Silva AJ. 2004. Consolidation of CSand US representations in associative fear conditioning. Hippocampus 14:557–69
Fudge JL, Emiliano AB. 2003. The extended amygdala and the dopamine system: another piece of thedopamine puzzle. J. Neuropsychiatry Clin. Neurosci. 15:306–16
Gabriel M, Burhans L, Kashef A. 2003. Consideration of a unified model of amygdalar associative functions.Ann. NY Acad. Sci. 985:206–17
Gallagher M, Kapp BS, Musty RE, Driscoll PA. 1977. Memory formation: evidence for a specific neurochem-ical system in the amygdala. Science 198:423–25
Garcia R, Paquereau J, Vouimba RM, Jaffard R. 1998. Footshock stress but not contextual fear conditioninginduces long-term enhancement of auditory-evoked potentials in the basolateral amygdala of the freelybehaving rat. Eur. J. Neurosci. 10:457–63
Garcia R, Spennato G, Nilsson-Todd L, Moreau JL, Deschaux O. 2008. Hippocampal low-frequency stim-ulation and chronic mild stress similarly disrupt fear extinction memory in rats. Neurobiol. Learn. Mem.89:560–66
Goldstein DS. 2003. Catecholamines and stress. Endocr. Regul. 37:69–80Gourley SL, Kedves AT, Olausson P, Taylor JR. 2008. A history of corticosterone exposure regulates fear
extinction and cortical NR2B, GluR2/3, and BDNF. Neuropsychopharmacology 34:707–16Gray TS. 1993. Amygdaloid CRF pathways. Role in autonomic, neuroendocrine, and behavioral responses to
stress. Ann. NY Acad. Sci. 697:53–60Grillon C, Cordova J, Morgan CA, Charney DS, Davis M. 2004. Effects of the beta-blocker propranolol on
cued and contextual fear conditioning in humans. Psychopharmacology 175:342–52Harley CW. 2007. Norepinephrine and the dentate gyrus. Prog. Brain Res. 163:299–318Hu H, Real E, Takamiya K, Kang MG, Ledoux J, et al. 2007. Emotion enhances learning via norepinephrine
regulation of AMPA-receptor trafficking. Cell 131:160–73Huang YY, Martin KC, Kandel ER. 2000. Both protein kinase A and mitogen-activated protein kinase are re-
quired in the amygdala for the macromolecular synthesis-dependent late phase of long-term potentiation.J. Neurosci. 20:6317–25
Hui GK, Figueroa IR, Poytress BS, Roozendaal B, McGaugh JL, Weinberger NM. 2004. Memory enhance-ment of classical fear conditioning by post-training injections of corticosterone in rats. Neurobiol. Learn.Mem. 81:67–74
Hui IR, Hui GK, Roozendaal B, McGaugh JL, Weinberger NM. 2006. Posttraining handling facilitatesmemory for auditory-cue fear conditioning in rats. Neurobiol. Learn. Mem. 86:160–63
Izquierdo A, Wellman CL, Holmes A. 2006. Brief uncontrollable stress causes dendritic retraction in infral-imbic cortex and resistance to fear extinction in mice. J. Neurosci. 26:5733–38
Izquierdo LA, Barros DM, Medina JH, Izquierdo I. 2002. Stress hormones enhance retrieval of fear condi-tioning acquired either one day or many months before. Behav. Pharmacol. 13:203–13
Jackson ME, Moghaddam B. 2006. Distinct patterns of plasticity in prefrontal cortex neurons that encodeslow and fast responses to stress. Eur. J. Neurosci. 24:1702–10
Ji J, Maren S. 2007. Hippocampal involvement in contextual modulation of fear extinction. Hippocampus17:749–58
Ji JZ, Wang XM, Li BM. 2003. Deficit in long-term contextual fear memory induced by blockade of beta-adrenoceptors in hippocampal CA1 region. Eur. J. Neurosci. 17:1947–52
Jin XC, Lu YF, Yang XF, Ma L, Li BM. 2007. Glucocorticoid receptors in the basolateral nucleus of amygdalaare required for postreactivation reconsolidation of auditory fear memory. Eur. J. Neurosci. 25:3702–12
Joels M, de Kloet ER. 1992. Control of neuronal excitability by corticosteroid hormones. Trends Neurosci.15:25–30
Kalin NH, Shelton SE. 2003. Nonhuman primate models to study anxiety, emotion regulation, and psy-chopathology. Ann. NY Acad. Sci. 1008:189–200
Kavushansky A, Richter-Levin G. 2006. Effects of stress and corticosterone on activity and plasticity in theamygdala. J. Neurosci. Res. 84:1580–87
308 Rodrigues · LeDoux · Sapolsky
Ann
u. R
ev. N
euro
sci.
2009
.32:
289-
313.
Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
Uni
vers
iteit
van
Am
ster
dam
on
03/2
5/10
. For
per
sona
l use
onl
y.

ANRV379-NE32-13 ARI 13 May 2009 8:31
Kavushansky A, Vouimba RM, Cohen H, Richter-Levin G. 2006. Activity and plasticity in the CA1, the dentategyrus, and the amygdala following controllable vs. uncontrollable water stress. Hippocampus 16:35–42
Kim JJ, Fanselow MS. 1992. Modality-specific retrograde amnesia of fear. Science 256:675–77Kohda K, Harada K, Kato K, Hoshino A, Motohashi J, et al. 2007. Glucocorticoid receptor activation is
involved in producing abnormal phenotypes of single-prolonged stress rats: a putative post-traumaticstress disorder model. Neuroscience 148:22–33
Korte SM. 2001. Corticosteroids in relation to fear, anxiety and psychopathology. Neurosci. Biobehav. Rev.25:117–42
LaBar KS, LeDoux JE. 2001. Coping with danger: the neural basis of defensive behaviors and fearful feelings.In Handbook of Physiology, Section 7: The Endocrine System, Vol. IV: Coping with the Environment: Neural andEndocrine Mechanisms, ed. BS McEwen, pp. 139–54. New York: Oxford Univ. Press
Lang PJ, Davis M. 2006. Emotion, motivation, and the brain: reflex foundations in animal and human research.Prog. Brain Res. 156:3–29
LeDoux J. 2007. The amygdala. Curr. Biol. 17:R868–74LeDoux JE. 1995. Emotion: clues from the brain. Annu. Rev. Psychol. 46:209–35LeDoux JE. 1996. The Emotional Brain: The Mysterious Underpinnings of Emotional Life. New York: Simon &
Schuster. 384 pp.LeDoux JE. 2000. Emotion circuits in the brain. Annu. Rev. Neurosci. 23:155–84LeDoux JE. 2002. Synaptic Self: How Our Brains Become Who We Are. New York: Viking. x. 406 pp.Lee HJ, Berger SY, Stiedl O, Spiess J, Kim JJ. 2001. Post-training injections of catecholaminergic drugs do
not modulate fear conditioning in rats and mice. Neurosci. Lett. 303:123–26Lee JL, Dickinson A, Everitt BJ. 2005. Conditioned suppression and freezing as measures of aversive Pavlovian
conditioning: effects of discrete amygdala lesions and overtraining. Behav. Brain Res. 159:221–33Liang KC, Juler RG, McGaugh JL. 1986. Modulating effects of posttraining epinephrine on memory: in-
volvement of the amygdala noradrenergic system. Brain Res. 368:125–33Likhtik E, Popa D, Apergis-Schoute J, Fidacaro GA, Pare D. 2008. Amygdala intercalated neurons are required
for expression of fear extinction. Nature 454:642–45Liu X, Lonart G, Sanford LD. 2007. Transient fear-induced alterations in evoked release of norepinephrine
and GABA in amygdala slices. Brain Res. 1142:46–53Luine V. 2002. Sex differences in chronic stress effects on memory in rats. Stress 5:205–16Lyons DM, Parker KJ. 2007. Stress inoculation-induced indications of resilience in monkeys. J. Trauma. Stress
20:423–33Maren S. 1999. Neurotoxic basolateral amygdala lesions impair learning and memory but not the performance
of conditional fear in rats. J. Neurosci. 19:8696–703Maren S. 2001. Neurobiology of Pavlovian fear conditioning. Annu. Rev. Neurosci. 24:897–931Maren S. 2005. Synaptic mechanisms of associative memory in the amygdala. Neuron 47:783–86Maren S, Quirk GJ. 2004. Neuronal signalling of fear memory. Nat. Rev. 5:844–52Maroun M, Richter-Levin G. 2003. Exposure to acute stress blocks the induction of long-term potentiation
of the amygdala-prefrontal cortex pathway in vivo. J. Neurosci. 23:4406–9McCarty R, Gold PE. 1996. Catecholamines, stress, and disease: a psychobiological perspective. Psychosom.
Med. 58:590–97McDonald AJ. 1991. Topographical organization of amygdaloid projections to the caudatoputamen, nucleus
accumbens, and related striatal-like areas of the rat brain. Neuroscience 44:15–33McDonald AJ. 1998. Cortical pathways to the mammalian amygdala. Prog. Neurobiol. 55:257–332McDonald AJ, Mascagni F, Guo L. 1996. Projections of the medial and lateral prefrontal cortices to the
amygdala: a Phaseolus vulgaris leucoagglutinin study in the rat. Neuroscience 71:55–75McEwen BS. 1999. Stress and hippocampal plasticity. Annu. Rev. Neurosci. 22:105–22McEwen BS. 2001. Plasticity of the hippocampus: adaptation to chronic stress and allostatic load. Ann. NY
Acad. Sci. 933:265–77McEwen BS. 2003. Mood disorders and allostatic load. Biol. Psychiatry 54:200–7McEwen BS, Magarinos AM. 2001. Stress and hippocampal plasticity: implications for the pathophysiology
of affective disorders. Hum. Psychopharmacol. 16:S7–19
www.annualreviews.org • Stress Hormones and Fear Circuitry 309
Ann
u. R
ev. N
euro
sci.
2009
.32:
289-
313.
Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
Uni
vers
iteit
van
Am
ster
dam
on
03/2
5/10
. For
per
sona
l use
onl
y.

ANRV379-NE32-13 ARI 13 May 2009 8:31
McGaugh JL. 2000. Memory—a century of consolidation. Science 287:248–51McGaugh JL. 2003. Memory and Emotion: The Making of Lasting Memories. London: Weidenfeld & Nicolson.
xi. 162 pp.McGaugh JL, McIntyre CK, Power AE. 2002. Amygdala modulation of memory consolidation: interaction
with other brain systems. Neurobiol. Learn. Mem. 78:539–52McIntyre CK, Hatfield T, McGaugh JL. 2002. Amygdala norepinephrine levels after training predict in-
hibitory avoidance retention performance in rats. Eur. J. Neurosci. 16:1223–26Meaney MJ. 2001. Maternal care, gene expression, and the transmission of individual differences in stress
reactivity across generations. Annu. Rev. Neurosci. 24:1161–92Miracle AD, Brace MF, Huyck KD, Singler SA, Wellman CL. 2006. Chronic stress impairs recall of extinction
of conditioned fear. Neurobiol. Learn. Mem. 85:213–18Mitra R, Jadhav S, McEwen BS, Vyas A, Chattarji S. 2005. Stress duration modulates the spatiotemporal
patterns of spine formation in the basolateral amygdala. Proc. Natl. Acad. Sci. USA 102:9371–76Mitra R, Sapolsky RM. 2008. Acute corticosterone treatment is sufficient to induce anxiety and amygdaloid
dendritic hypertrophy. Proc. Natl. Acad. Sci. USA 105:5573–78Mitra R, Sapolsky RM. 2009. Effects of enrichment predominate over those of chronic stress on fear-related
behavior in male rats. Stress. In pressMizoguchi K, Shoji H, Ikeda R, Tanaka Y, Maruyama W, Tabira T. 2008. Suppression of glucocorticoid
secretion enhances cholinergic transmission in rat hippocampus. Brain Res. Bull. 76:612–15Mueller D, Porter JT, Quirk GJ. 2008. Noradrenergic signaling in infralimbic cortex increases cell excitability
and strengthens memory for fear extinction. J. Neurosci. 28:369–75Murchison CF, Zhang XY, Zhang WP, Ouyang M, Lee A, Thomas SA. 2004. A distinct role for norepinephrine
in memory retrieval. Cell 117:131–43Nader K. 2003. Memory traces unbound. Trends Neurosci. 26;65–72Nader K, Schafe GE, LeDoux JE. 2000. The labile nature of consolidation theory. Nat. Rev. 1:216–19Neophytou SI, Aspley S, Butler S, Beckett S, Marsden CA. 2001. Effects of lesioning noradrenergic
neurones in the locus coeruleus on conditioned and unconditioned aversive behaviour in the rat.Prog. Neuropsychopharmacol. Biol. Psychiatry 25:1307–21
Orr SP, Milad MR, Metzger LJ, Lasko NB, Gilbertson MW, Pitman RK. 2006. Effects of beta blockade, PTSDdiagnosis, and explicit threat on the extinction and retention of an aversively conditioned response. Biol.Psychol. 73:262–71
Pare D. 2003. Role of the basolateral amygdala in memory consolidation. Progress Neurobiol. 70:409–20Pare D, Quirk GJ, Ledoux JE. 2004. New vistas on amygdala networks in conditioned fear. J. Neurophysiol.
92:1–9Pavlides C, McEwen BS. 1999. Effects of mineralocorticoid and glucocorticoid receptors on long-term po-
tentiation in the CA3 hippocampal field. Brain Res. 851:204–14Pavlides C, Nivon LG, McEwen BS. 2002. Effects of chronic stress on hippocampal long-term potentiation.
Hippocampus 12:245–57Pavlides C, Ogawa S, Kimura A, McEwen BS. 1996. Role of adrenal steroid mineralocorticoid and glucocor-
ticoid receptors in long-term potentiation in the CA1 field of hippocampal slices. Brain Res. 738:229–35
Phillips RG, LeDoux JE. 1992. Differential contribution of amygdala and hippocampus to cued and contextualfear conditioning. Behav. Neurosci. 106:274–85
Pitkanen A, Pikkarainen M, Nurminen N, Ylinen A. 2000. Reciprocal connections between the amygdala andthe hippocampal formation, perirhinal cortex, and postrhinal cortex in rat. A review. Ann. NY Acad. Sci.911:369–91
Pitkanen A, Savander V, LeDoux JE. 1997. Organization of intra-amygdaloid circuitries in the rat: an emergingframework for understanding functions of the amygdala. Trends Neurosci. 20:517–23
Pitman RK, Delahanty DL. 2005. Conceptually driven pharmacologic approaches to acute trauma.CNS Spectr. 10:99–106
Pitman RK, Sanders KM, Zusman RM, Healy AR, Cheema F, et al. 2002. Pilot study of secondary preventionof posttraumatic stress disorder with propranolol. Biol. Psychiatry 51:189–92
310 Rodrigues · LeDoux · Sapolsky
Ann
u. R
ev. N
euro
sci.
2009
.32:
289-
313.
Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
Uni
vers
iteit
van
Am
ster
dam
on
03/2
5/10
. For
per
sona
l use
onl
y.

ANRV379-NE32-13 ARI 13 May 2009 8:31
Price JL. 2003. Comparative aspects of amygdala connectivity. Ann. NY Acad. Sci. 985:50–58Przybyslawski J, Roullet P, Sara SJ. 1999. Attenuation of emotional and nonemotional memories after their
reactivation: role of beta adrenergic receptors. J. Neurosci. 19:6623–28Pugh CR, Fleshner M, Rudy JW. 1997a. Type II glucocorticoid receptor antagonists impair contextual but
not auditory-cue fear conditioning in juvenile rats. Neurobiol. Learn. Mem. 67:75–79Pugh CR, Tremblay D, Fleshner M, Rudy JW. 1997b. A selective role for corticosterone in contextual-fear
conditioning. Behav. Neurosci. 111:503–11Quirarte GL, Roozendaal B, McGaugh JL. 1997. Glucocorticoid enhancement of memory storage involves
noradrenergic activation in the basolateral amygdala. Proc. Natl. Acad. Sci. USA 94:14048–53Quirk GJ, Garcia R, Gonzalez-Lima F. 2006. Prefrontal mechanisms in extinction of conditioned fear. Biol.
Psychiatry 60:337–43Quirk GJ, Mueller D. 2008. Neural mechanisms of extinction learning and retrieval. Neuropsychopharmacology
33:56–72Radley JJ, Rocher AB, Janssen WG, Hof PR, McEwen BS, Morrison JH. 2005. Reversibility of apical dendritic
retraction in the rat medial prefrontal cortex following repeated stress. Exp. Neurol. 196:199–203Radley JJ, Rocher AB, Miller M, Janssen WG, Liston C, et al. 2006. Repeated stress induces dendritic spine
loss in the rat medial prefrontal cortex. Cereb. Cortex 16:313–20Radley JJ, Rocher AB, Rodriguez A, Ehlenberger DB, Dammann M, et al. 2008. Repeated stress alters dendritic
spine morphology in the rat medial prefrontal cortex. J. Comp. Neurol. 507:1141–50Radley JJ, Sisti HM, Hao J, Rocher AB, McCall T, et al. 2004. Chronic behavioral stress induces apical
dendritic reorganization in pyramidal neurons of the medial prefrontal cortex. Neuroscience 125:1–6Ramos BP, Arnsten AF. 2007. Adrenergic pharmacology and cognition: focus on the prefrontal cortex. Phar-
macol. Ther. 113:523–36Rau V, DeCola JP, Fanselow MS. 2005. Stress-induced enhancement of fear learning: an animal model of
posttraumatic stress disorder. Neurosci. Biobehav. Rev. 29:1207–23Rau V, Fanselow MS. 2008. Exposure to a stressor produces a long lasting enhancement of fear learning in
rats. Stress 18:1Richter-Levin G. 2004. The amygdala, the hippocampus, and emotional modulation of memory.
Neuroscientist 10:31–39Richter-Levin G, Akirav I. 2003. Emotional tagging of memory formation—in the search for neural mecha-
nisms. Brain Res. Brain Res. Rev. 43:247–56Rocher C, Spedding M, Munoz C, Jay TM. 2004. Acute stress-induced changes in hippocampal/prefrontal
circuits in rats: effects of antidepressants. Cereb. Cortex 14:224–29Rodrigues SM, Sapolsky RM. 2009. Disruption of fear memory through dual-hormone gene therapy. Biol.
Psychiatry 65:441–44Rodrigues SM, Schafe GE, LeDoux JE. 2004. Molecular mechanisms underlying emotional learning and
memory in the lateral amygdala. Neuron 44:75–91Rodrıguez Manzanares PA, Isoardi NA, Carrer HF, Molina VA. 2005. Previous stress facilitates fear mem-
ory, attenuates GABAergic inhibition, and increases synaptic plasticity in the rat basolateral amygdala.J. Neurosci. 25:8725–34
Romanski LM, Clugnet MC, Bordi F, LeDoux JE. 1993. Somatosensory and auditory convergence in thelateral nucleus of the amygdala. Behav. Neurosci. 107:444–50
Roozendaal B, Hui GK, Hui IR, Berlau DJ, McGaugh JL, Weinberger NM. 2006. Basolateral amygdala nora-drenergic activity mediates corticosterone-induced enhancement of auditory fear conditioning. Neurobiol.Learn. Mem. 86:249–55
Roozendaal B, McGaugh JL. 1997. Glucocorticoid receptor agonist and antagonist administration intothe basolateral but not central amygdala modulates memory storage. Neurobiol. Learn. Mem. 67:176–79
Rosenkranz JA, Moore H, Grace AA. 2003. The prefrontal cortex regulates lateral amygdala neuronal plasticityand responses to previously conditioned stimuli. J. Neurosci. 23:11054–64
Santini E, Quirk GJ, Porter JT. 2008. Fear conditioning and extinction differentially modify the intrinsicexcitability of infralimbic neurons. J. Neurosci. 28:4028–36
www.annualreviews.org • Stress Hormones and Fear Circuitry 311
Ann
u. R
ev. N
euro
sci.
2009
.32:
289-
313.
Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
Uni
vers
iteit
van
Am
ster
dam
on
03/2
5/10
. For
per
sona
l use
onl
y.

ANRV379-NE32-13 ARI 13 May 2009 8:31
Sapolsky RM. 2003. Stress and plasticity in the limbic system. Neurochem. Res. 28:1735–42Sapolsky RM. 2004. Why Zebras Don’t Get Ulcers. New York: Owl Book/Henry Holt. xv. 539 pp.Sapolsky RM, Romero LM, Munck AU. 2000. How do glucocorticoids influence stress responses? Integrating
permissive, suppressive, stimulatory, and preparative actions. Endocr. Rev. 21:55–89Sarter M, Markowitsch HJ. 1985. Involvement of the amygdala in learning and memory: a critical review, with
emphasis on anatomical relations. Behav. Neurosci. 99:342–80Schafe GE, Nader K, Blair HT, LeDoux JE. 2001. Memory consolidation of Pavlovian fear conditioning: a
cellular and molecular perspective. Trends Neurosci. 24:540–46Schelling G, Roozendaal B, De Quervain DJ. 2004. Can posttraumatic stress disorder be prevented with
glucocorticoids? Ann. NY Acad. Sci. 1032:158–66Segal M, Markram H, Richter-Levin G. 1991. Actions of norepinephrine in the rat hippocampus.
Prog. Brain Res. 88:323–30Selden NR, Robbins TW, Everitt BJ. 1990. Enhanced behavioral conditioning to context and impaired be-
havioral and neuroendocrine responses to conditioned stimuli following ceruleocortical noradrenergiclesions: support for an attentional hypothesis of central noradrenergic function. J. Neurosci. 10:531–39
Shors TJ, Seib TB, Levine S, Thompson RF. 1989. Inescapable versus escapable shock modulates long-termpotentiation in the rat hippocampus. Science 244:224–26
Soravia LM, Heinrichs M, Aerni A, Maroni C, Schelling G, et al. 2006. Glucocorticoids reduce phobic fearin humans. Proc. Natl. Acad. Sci. USA 103:5585–90
Sotres-Bayon F, Bush DE, LeDoux JE. 2004. Emotional perseveration: an update on prefrontal-amygdalainteractions in fear extinction. Learn. Mem. 11:525–35
Sousa N, Lukoyanov NV, Madeira MD, Almeida OFX, Paula-Barbosa MM. 2000. Reorganization of themorphology of hippocampal neurites and synapses after stress-induced damage correlates with behavioralimprovement. Neuroscience 97:253–66
Stefanacci L, Amaral DG. 2002. Some observations on cortical inputs to the macaque monkey amygdala: ananterograde tracing study. J. Comp. Neurol. 451:301–23
Swanson LW. 2003. The amygdala and its place in the cerebral hemisphere. Ann. NY Acad. Sci. 985:174–84Talarovicova A, Krskova L, Kiss A. 2007. Some assessments of the amygdala role in suprahypothalamic neu-
roendocrine regulation: a minireview. Endocr. Regul. 41:155–62Tronel S, Alberini CM. 2007. Persistent disruption of a traumatic memory by postretrieval inactivation of
glucocorticoid receptors in the amygdala. Biol. Psychiatry 62:33–39Tully K, Li Y, Tsvetkov E, Bolshakov VY. 2007. Norepinephrine enables the induction of associative long-term
potentiation at thalamo-amygdala synapses. Proc. Natl. Acad. Sci. USA 104:14146–50Vidal-Gonzalez I, Vidal-Gonzalez B, Rauch SL, Quirk GJ. 2006. Microstimulation reveals opposing influ-
ences of prelimbic and infralimbic cortex on the expression of conditioned fear. Learn. Mem. 13:728–33
Vouimba RM, Munoz C, Diamond DM. 2006. Differential effects of predator stress and the antidepres-sant tianeptine on physiological plasticity in the hippocampus and basolateral amygdala. Stress 9:29–40
Vouimba RM, Yaniv D, Diamond D, Richter-Levin G. 2004. Effects of inescapable stress on LTP in theamygdala versus the dentate gyrus of freely behaving rats. Eur. J. Neurosci. 19:1887–94
Vyas A, Bernal S, Chattarji S. 2003. Effects of chronic stress on dendritic arborization in the central andextended amygdala. Brain Res. 965:290–94
Vyas A, Chattarji S. 2004. Modulation of different states of anxiety-like behavior by chronic stress. Behav.Neurosci. 118:1450–54
Vyas A, Jadhav S, Chattarji S. 2006. Prolonged behavioral stress enhances synaptic connectivity in the baso-lateral amygdala. Neuroscience 143:387–93
Vyas A, Mitra R, Shankaranarayana Rao BS, Chattarji S. 2002. Chronic stress induces contrasting patterns ofdendritic remodeling in hippocampal and amygdaloid neurons. J. Neurosci. 22:6810–18
Vyas A, Pillai AG, Chattarji S. 2004. Recovery after chronic stress fails to reverse amygdaloid neuronalhypertrophy and enhanced anxiety-like behavior. Neuroscience 128:667–73
312 Rodrigues · LeDoux · Sapolsky
Ann
u. R
ev. N
euro
sci.
2009
.32:
289-
313.
Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
Uni
vers
iteit
van
Am
ster
dam
on
03/2
5/10
. For
per
sona
l use
onl
y.

ANRV379-NE32-13 ARI 13 May 2009 8:31
Weinstock M. 2007. Gender differences in the effects of prenatal stress on brain development and behaviour.Neurochem. Res. 32:1730–40
Wellman CL. 2001. Dendritic reorganization in pyramidal neurons in medial prefrontal cortex after chroniccorticosterone administration. J. Neurobiol. 49:245–53
Wessa M, Flor H. 2007. Failure of extinction of fear responses in posttraumatic stress disorder: evidence fromsecond-order conditioning. Am. J. Psychiatry 164:1684–92
Wilensky AE, Schafe GE, Kristensen MP, LeDoux JE. 2006. Rethinking the fear circuit: the central nucleus ofthe amygdala is required for the acquisition, consolidation, and expression of Pavlovian fear conditioning.J. Neurosci. 26:12387–96
Yang YL, Chao PK, Lu KT. 2006. Systemic and intra-amygdala administration of glucocorticoid agonist andantagonist modulate extinction of conditioned fear. Neuropsychopharmacology 31:912–24
Zimmerman JM, Rabinak CA, McLachlan IG, Maren S. 2007. The central nucleus of the amygdala is essentialfor acquiring and expressing conditional fear after overtraining. Learn. Mem. 14:634–44
Zorawski M, Blanding NQ, Kuhn CM, LaBar KS. 2006. Effects of stress and sex on acquisition and consoli-dation of human fear conditioning. Learn. Mem. 13:441–50
Zorawski M, Killcross S. 2002. Posttraining glucocorticoid receptor agonist enhances memory in appetitiveand aversive Pavlovian discrete-cue conditioning paradigms. Neurobiol. Learn. Mem. 78:458–64
www.annualreviews.org • Stress Hormones and Fear Circuitry 313
Ann
u. R
ev. N
euro
sci.
2009
.32:
289-
313.
Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
Uni
vers
iteit
van
Am
ster
dam
on
03/2
5/10
. For
per
sona
l use
onl
y.

AR379-FM ARI 20 May 2009 14:8
Annual Review ofNeuroscience
Volume 32, 2009Contents
Neuropathic Pain: A Maladaptive Response of the NervousSystem to DamageMichael Costigan, Joachim Scholz, and Clifford J. Woolf � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 1
Synaptic Mechanisms for Plasticity in NeocortexDaniel E. Feldman � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �33
Neurocognitive Mechanisms in Depression: Implications forTreatmentLuke Clark, Samuel R. Chamberlain, and Barbara J. Sahakian � � � � � � � � � � � � � � � � � � � � � � � � � �57
Using Diffusion Imaging to Study Human Connectional AnatomyHeidi Johansen-Berg and Matthew F.S. Rushworth � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �75
Serotonin in Affective ControlPeter Dayan and Quentin J.M. Huys � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �95
Physiology and Pharmacology of Striatal NeuronsAnatol C. Kreitzer � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 127
The Glial Nature of Embryonic and Adult Neural Stem CellsArnold Kriegstein and Arturo Alvarez-Buylla � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 149
Representation of Number in the BrainAndreas Nieder and Stanislas Dehaene � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 185
Neuronal Gamma-Band Synchronization as a Fundamental Processin Cortical ComputationPascal Fries � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 209
The Neurobiology of Individual Differences in ComplexBehavioral TraitsAhmad R. Hariri � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 225
The Science of Neural Interface SystemsNicholas G. Hatsopoulos and John P. Donoghue � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 249
The Neuropsychopharmacology of Fronto-Executive Function:Monoaminergic ModulationT.W. Robbins and A.F.T. Arnsten � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 267
v
Ann
u. R
ev. N
euro
sci.
2009
.32:
289-
313.
Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
Uni
vers
iteit
van
Am
ster
dam
on
03/2
5/10
. For
per
sona
l use
onl
y.

AR379-FM ARI 20 May 2009 14:8
The Influence of Stress Hormones on Fear CircuitrySarina M. Rodrigues, Joseph E. LeDoux, and Robert M. Sapolsky � � � � � � � � � � � � � � � � � � � � � � � 289
The Primate Cortical Auditory System and Neural Representation ofConspecific VocalizationsLizabeth M. Romanski and Bruno B. Averbeck � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 315
Establishment of Axon-Dendrite Polarity in Developing NeuronsAnthony P. Barnes and Franck Polleux � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 347
Axon Growth and Guidance: Receptor Regulationand Signal TransductionMichael O’Donnell, Rebecca K. Chance, and Greg J. Bashaw � � � � � � � � � � � � � � � � � � � � � � � � � � � � 383
Cerebellum and Nonmotor FunctionPeter L. Strick, Richard P. Dum, and Julie A. Fiez � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 413
Advances in Light Microscopy for NeuroscienceBrian A. Wilt, Laurie D. Burns, Eric Tatt Wei Ho, Kunal K. Ghosh,Eran A. Mukamel, and Mark J. Schnitzer � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 435
Indexes
Cumulative Index of Contributing Authors, Volumes 23–32 � � � � � � � � � � � � � � � � � � � � � � � � � � � 507
Cumulative Index of Chapter Titles, Volumes 23–32 � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 511
Errata
An online log of corrections to Annual Review of Neuroscience articles may be found athttp://neuro.annualreviews.org/
vi Contents
Ann
u. R
ev. N
euro
sci.
2009
.32:
289-
313.
Dow
nloa
ded
from
arj
ourn
als.
annu
alre
view
s.or
gby
Uni
vers
iteit
van
Am
ster
dam
on
03/2
5/10
. For
per
sona
l use
onl
y.