
The growth of swine bone marrow cells in the presence of heterologous colony stimulating factor:...
17
Comp. lmmun. Microbiol. infect. Dis., Vol. 6, No. 2, pp. 171-187, 1983. 0147 9571/83/020171-17103.00/0 Printed in Great Britain. All rights reserved Copyright © 1983 Pergamon Press Ltd THE GROWTH OF SWINE BONE MARROW CELLS IN THE PRESENCE OF HETEROLOGOUS COLONY STIMULATING FACTOR: CHARACTERIZATION OF THE DEVELOPING CELL POPULATION PETER MAYER Sandoz Forschungsinstitut Ges.M.B.H., A-1235 Wien, Brunnerstrasse 59, Austria (Received 1 October 1982) Abstract--Conditioned medium prepared from the mouse fibroblast cell line L929 which is known to produce colony-stimulating factor active on mouse bone marrow cells was also able to stimulate the growth of swine bone marrow ceils in a liquid culture system. During the first 4 days of culture mononuclear phagocyte and granulocyte colonies developed. Prolonged cultured cells were classified belonging to the macrophage lineage by morphological and cytochemical criteria. These cells fulfill also functional characteristics for macrophages, like expression of Fc receptors, immune phagocytosis and production of prostaglandins. These bone marrow-derived macrophages could also be activated with LPS and lymphocyte-derived mediators. Key words: Swine bone marrow cell cultures, 'colony stimulating factor', macrophages CROISSANCE DES CELLULES DE MOELLE OSSEUSE AU PORC EN PRESENCE DE 'COLONY STIMULATING FACTOR' HETEROLOGUE: CARACTERISATION DE LA POPULATION R6sum6---La cellule fibroblastique L929 (d'origine murine) est connue comme produisant le 'colony stimulating factor' (CSF) actif sur les cellules de moelle osseuse de souris. Un milieu pr6par6 h partir de cette cellule L929 est aussi capable de stimuler la croissance des cellules de moelle osseuse de pore en syst6me de culture liquide. Pendant les 4 premiers jours de culture, des colonies de phagocytes mononucl6aires et de granulocytes se sont d6velopp6es. Apr6s des p6riodes de culture prolong6es, des cellules ont 6t6 identifi6es comme appartenant h la classe des macrophages sur des crit6res morphologiques et cytochimiques. De plus, ces cellules ont pr6sent6 des caract6ristiques fonctionnelles de macrophage, telles que l'expression des r6cepteurs Fc, la phagocytose immune et la production de prostaglandines. Ces macrophages issus de la moelle osseuse peuvent ~tre aussi stimul6s par du LPS et des m6diateurs d6riv6s des lymphocytes. Mots-clefs: Cellules de moelle osseuse de pore, 'colony stimulating factor', macrophages INTRODUCTION The concept that the continuous formation of blood cells might be under the control of specific humoral regulatory factors received early support from the demonstration that a humoral factor termed erythropoietin, was involved in the regulation of erythropoiesis [1]. However, it was not until the introduction in 1966 of methods for growing colonies of normal hemopoietic cells in semisolid culture that progress was made in identifying specific regulators for other hemopoietic populations. The first hemopoietic colonies to 171 C.LM.t.D. 6/2--E
-
Upload
peter-mayer -
Category
Documents
-
view
213 -
download
0
Transcript of The growth of swine bone marrow cells in the presence of heterologous colony stimulating factor:...

Comp. lmmun. Microbiol. infect. Dis., Vol. 6, No. 2, pp. 171-187, 1983. 0147 9571/83/020171-17103.00/0 Printed in Great Britain. All rights reserved Copyright © 1983 Pergamon Press Ltd
T H E G R O W T H O F S W I N E B O N E M A R R O W C E L L S I N
T H E P R E S E N C E O F H E T E R O L O G O U S C O L O N Y
S T I M U L A T I N G F A C T O R : C H A R A C T E R I Z A T I O N
O F T H E D E V E L O P I N G C E L L P O P U L A T I O N
PETER MAYER
Sandoz Forschungsinstitut Ges.M.B.H., A-1235 Wien, Brunnerstrasse 59, Austria
(Received 1 October 1982)
Abstract--Conditioned medium prepared from the mouse fibroblast cell line L929 which is known to produce colony-stimulating factor active on mouse bone marrow cells was also able to stimulate the growth of swine bone marrow ceils in a liquid culture system. During the first 4 days of culture mononuclear phagocyte and granulocyte colonies developed. Prolonged cultured cells were classified belonging to the macrophage lineage by morphological and cytochemical criteria. These cells fulfill also functional characteristics for macrophages, like expression of Fc receptors, immune phagocytosis and production of prostaglandins. These bone marrow-derived macrophages could also be activated with LPS and lymphocyte-derived mediators.
Key words: Swine bone marrow cell cultures, 'colony stimulating factor', macrophages
CROISSANCE DES CELLULES DE MOELLE OSSEUSE AU PORC EN PRESENCE DE 'COLONY STIMULATING
FACTOR' HETEROLOGUE: CARACTERISATION DE LA POPULATION
R6sum6---La cellule fibroblastique L929 (d'origine murine) est connue comme produisant le 'colony stimulating factor' (CSF) actif sur les cellules de moelle osseuse de souris. Un milieu pr6par6 h partir de cette cellule L929 est aussi capable de stimuler la croissance des cellules de moelle osseuse de pore en syst6me de culture liquide. Pendant les 4 premiers jours de culture, des colonies de phagocytes mononucl6aires et de granulocytes se sont d6velopp6es. Apr6s des p6riodes de culture prolong6es, des cellules ont 6t6 identifi6es comme appartenant h la classe des macrophages sur des crit6res morphologiques et cytochimiques. De plus, ces cellules ont pr6sent6 des caract6ristiques fonctionnelles de macrophage, telles que l'expression des r6cepteurs Fc, la phagocytose immune et la production de prostaglandines. Ces macrophages issus de la moelle osseuse peuvent ~tre aussi stimul6s par du LPS et des m6diateurs d6riv6s des lymphocytes.
Mots-clefs: Cellules de moelle osseuse de pore, 'colony stimulating factor', macrophages
INTRODUCTION
The concept that the continuous formation of blood cells might be under the control of specific humoral regulatory factors received early support from the demonstration that a humoral factor termed erythropoietin, was involved in the regulation of erythropoiesis [1]. However, it was not until the introduction in 1966 of methods for growing colonies of normal hemopoietic cells in semisolid culture that progress was made in identifying specific regulators for other hemopoietic populations. The first hemopoietic colonies to
171 C.LM.t.D. 6/2--E

172 PETER MAYER
be grown in such cultures were composed of neutrophilic granulocytes and/or macro- phages [-2]. Cells were induced to form colonies by using a variety of cell feeder layers or substrates conditioned by various cells [2, 3]. With the demonstration that colony forma- tion is dependent on an active factor present in such materials the operational term 'colony stimulating factor' 'CSF' was coined [4]. Parallel studies showed that certain murine and human cell lines of normal or neoplastic origin could also produce CSF, the most frequently used system being the L929 transformed mouse fibroblast cell line [5, 6]. The serum-free media conditioned by some of these cell lines are the most desirable starting material for the purification of CSF. CSF from L-cell cultures has now been purified to homogeneity and antibodies against it have been raised [7].
In this study we are able to show that cells in the swine bone marrow respond to L cell conditioned medium with proliferation and differentiation. The cell population develop- ing in these cultures after 10-14 days fulfill the morphological, cytochemical and func- tional criteria of macrophages.
MATERIALS AND METHODS
Animals
Female piglets of the large white breed, weighing 4-5 kg at an age of 2 3 weeks, were separated from their dam and fed with an artificial sow milk preparation (Piglac).
L-cell conditioned medium
L929 cells were maintained in minimal essential medium (MEM) supplemented with 10~o heat-inactivated FCS and passaged by trypsinization (0.25~ trypsin in MEM). Supernatants from confluent monolayers were removed after 10 days, centrifuged at 1000rpm and added to a final concentration of 30~o to Dulbecco's H-21 medium con- taining 15~o heat-inactivated horse serum. This L-cell conditioned medium was used as the source of colony stimulating factor (CSF) [8]. The L-cell medium produced 4600 granulocyte-macrophage colonies/ml when assayed in soft agar with 105 mouse bone marrow cells [9].
Culturing of swine bone marrow cells (SBMC)
Swine bone marrow was obtained by breaking femora from piglets with bone forceps into pieces: the pieces were then transferred into a glass beaker containing Dulbecco's H21 medium and stirred for 30min at room temperature. The medium was poured through a stainless-steel mesh (12 x 0.3 mm wires/cm), the resultant cell suspension sucked into a syringe and then pressed through a 26 gauge needle to get a single cell suspension. 100ml of L-cell conditioned medium (30~o L cell supernatant and 70~o Dulbecco's H-21 with 15~o horse serum) containing 1.5 x 105 SBM cells/ml were sealed in bags of Teflon FEP fluorocarbon film (type 100 A Du Pont de Nemours and Co., Geneva, Switzerland) and cultured at 37°C in an incubator gassed with 95~o air and 5~o COz [10]. The medium was not changed throughout the culture period. The cells were recovered from the Teflon bags by gentle shaking and aspirating the culture fluid. They were centrifuged, suspended in Dulbecco's H21 medium containing 15~o horse serum, the number of cells counted and the viability tested by the trypan blue exclusion test. Using the same conditioned medium as described above, SBMC cultures were also established in Leighton tubes (Bellco Glass, Inc., Vineland, NY) containing a 'flying cover slip'

Swine bone marrow cell cultures, macrophages 173
(10 x 35 mm) and held at 37°C in a water-saturated atmosphere of 5 ~ CO2 in air. 5 x 104 SBMC were seeded in 2 ml medium per tube. For light microscopy the cover slip was rapidly air-dried, fixed in absolute methanol for 10 min and stained with Giemsa's stain for 15 min. Photographs were taken with a Reichert Univar photomicroscope.
Separation of SBMC on a Ficoll 9radient The gradients were prepared by mixing 30 ml Ficoll 1.077 g/ml (Seromed, Munich,
FRG) and different amounts (250-1000 /A) of Metrizoate solution 75~o w/v (Nyegaard A/S, Oslo, Norway). Twenty millilitres of 2 x 10V/ml SRBC suspension in Dulbecco's H21 medium were layered over 20ml Ficoll gradients of different densities in 50 ml plastic centrifuge tubes (Nunc, Denmark) and centrifuged at 420 g for 40 min. The separ- ated cell populations were washed twice in Dulbecco's H21 medium and counted prior to use .
[3H]-Thymidine incorporation into ceils of SBM cultures
[3H]-Thymidine incorporation into DNA was determined as described in detail else- where [11]. Briefly, SBMC seeded at a concentration of 2 x l05 cells/well in microtiter plates (Nunc, Denmark) were labelled by incubation at 37°C for 24hr with 0.3 pCi [3H]-methyl thymidine ([3H]-TdR; 25 Ci/nmol; Amersham, England) per well diluting the concentrated [3H]-TdR solution in Dulbecco's H21. At the end of the labelling period the medium was sucked off and 100 pl 0.1~o sodium docecyl sulfate (SDS) added. The lysate was put on filter paper disks (Whatman Ltd England, grade 3 mm). After drying, the filter disks were washed with 10~o trichloroacetic acid solution and then in methanol. The dried filter disks were put in scintillation vials and after adding scintilla- tion fluid, counted in a Packard scintillation counter.
Cytochemistry Cytocentrifuge preparations were made as described eslewhere [12]. For non-specific
esterase staining, cells were fixed in acetone for 10 min. Esterase activity was determined as reported by others [13] using ct-naphthyl acetate (Sigma Chemical Co. Ltd, London) as the substrate at pH 7.0; the incubation time was 10 min. For quantitation of esterase- positive cells 500 cells were examined.
Quantitation of immune phagocytosis
The phagocytic capacity of SBM-derived cells was determined by using IgG coated sheep red blood cells (SRBC). Opsonization of SRBC was performed as previously reported by Scheetz et al. [14]. As antibody, antisheep 'hemolysin ambozeptor 6000' (Behringwerke AG., Marburg, FRG) at a final dilution of 1 : 80 was used. Immune phago- cytosis was determined by measuring the hemoglobin of internalized SRBC after lysis of SBM-derived cells. Briefly, 2 x 105 SBM-derived cells suspended in 200 #1 Dulbecco's medium with 15~o inactivated horse serum were seeded per well in flat-bottomed micro- titer plates (Nunc, Denmark) and incubated for 2 hr at 37°C in an atmosphere of 5~o CO2. Then the cells were rinsed 3 times with medium to remove non-adherent cells. Using this washing procedures, it could be shown that 91 + 3~o (N = 20) adhered. To each well 2 x 107 opsonized SRBC in 100 #1 medium were added. The microtiter plates were incubated for 90 min. After that the plates were rinsed 3 times with saline to remove non-phagocytosed SRBC. SRBC non-phagocytosed by cultured SBM-derived cells but

174 PETER MAYER
adherent to the cell surface were lysed by the addition of distilled water for 5 sec. After adding 300 #1 1N NaOH to each well for 2 hr to lyse the SBM-derived cells, the concen- tration of the released hemoglobin was determined photometrically at a wavelength of 405 nm using a Titertek Multiscan photometer (Flow Laboratories, England). For stimu- lation of phagocytosis a stock solution of Lipopolysaccharide B (LPS) of S. typhosa 0901 (Difco Lab., Detroit, U.S.A.) in saline containing 80 #g/ml was prepared. Further dilu- tions were performed with Dulbecco's H21. The indicated amounts of LPS were added to the cells in a volume of 100/A.
Fc receptors
To test for Fc receptors the rosette method was used [15]. The cells were harvested from the Teflon bags and suspended at 4 x 106/ml in PBS-BSA. 100/A of the cell suspension was mixed with 100 #1 1 x l0 T opsonized SRBC in a plastic tube. This tube was placed on a slow rotator with horizontal axis at 37°C for 15 min. A wet-mount slide was prepared by sealing a drop of the rosette mixture under a coverslip with nail polish. 500 cells were looked at and rosettes were counted using phase contrast optics. A 'rosette' is defined as having three or more attached erythrocytes.
Nitroblue-tetrazolium (NBT) test
The assay for NBT was performed as described elsewhere [16]. Briefly, 1 x 106 SBM- derived cells were incubated in 24 well plastic trays (Nunc, Denmark, 15 mm dia) in 250 #1 Dulbecco's H21 medium without phenol red and 200/ll of a solution of p-nitro- tetrazolium (Sigma Chemical Co. Ltd, London) at a concentration of 1 mg/ml Dulbecco's H21 was added. In addition, 50 pl of a zymosan suspension (Sigma Chemical Co. Ltd, London) containing 1 x 109 particles/ml was added to stimulate the cells to perform phagocytosis and the mixture was incubated for 60 min. Then the cells were washed with saline and lysed with 500 #1 1N NaOH. The OD of which reduction of the yellow dye produced a deep blue color was measured at 550nm using a Titertek Multiscan photometer.
Prostaglandin assay
Prostaglandin levels in the culture supernatant (1 x 106 cells/ml) were determined by a coated charcoal radioimmunoassay [-17] utilizing anti-prostaglandin E2 serum (Miles Laboratories, Inc., Elkhart, Ind.). According to the manufacturer the immunoglobulins also reacted strongly with PGE1, but only weakly with PG F2, (100 and 5% cross- reactivity, respectively). PGE2 (Sigma Chemical Co. Ltd, London) was used as the standard. The results were reported as PGE equivalents.
Preparation of lymphocyte mediators
ConA- and PMA-induced lymphokines were prepared as described [18]. 1 x 107/ml swine spleen cells were incubated with different amounts of Concanavalin A (Pharmacia AB, Uppsala, Sweden; ConA, 2.5-20/~g/ml) or phorbol 12-myristate 13-acetate (Sigma Chemical Co. Ltd, London; PMA, 2-20 ng/ml) in Dulbecco's H21 containing 15% horse serum. After 24 hr the supernatants were collected, filtered and frozen at - 70°C. Control supernatants were prepared in the absence of ConA and PMA in a similar way.

Swine bone marrow cell cultures, macrophages 175
Activation of SBM-derived cells
Cultured SBM cells were collected from the Teflon bags and adjusted to 2 x 106
cells/ml in Dulbecco's H21 medium containing 15~o horse serum. 100/~1 of this suspen- sion was pipetted into flat-bottomed microtiter plates. For activation ConA or PMA induced lymphocyte supernatants (100 pl) were added to each well. SBM-derived cells serving as control received 100 #1 of supernatant from swine spleen cells cultured without ConA or PMA for 24hr. After 24 hr of incubation of the SBM-derived cells with the supernatants, they were washed once and immune phagocytosis performed as described above.
RESULTS
Kinetics of the proliferative response of SBMC L-cell conditioned medium
To examine the conditioned medium dependence of SBMC proliferation, SBMC were incubated in the presence and absence of L-cell supernatant and thymidine incorporation was monitored as a function of time. When SBMC were cultured in the absence of L-cell conditioned medium a steady decline of [3H]-TdR incorporation from day 1 to day 4 of the culture period could be observed. From day 4 on, only background values of [3H]-TdR could be detected. In the presence of L-cell supernatant a slight decrease in thymidine incorporation could be observed during the first 3 days of culture. On day 4, however, thymidine incorporation was significantly higher than in control cultures (P < 0,01). The highest level of thymidine incorporation was found on days 6 and 7 of the culture period. On days 8 and 9 only half of the amount of the labelled material was incorporated into the DNA of the cells compared to days 6 and 7 (Fig. 1).
This pattern of [3H]-TdR incorporation was also reflected in the kinetics of cell proliferation when SBMC were seeded into Teflon bags. The number of cells obtained on day 6 of the culture period in the L-cell conditioned medium was significantly higher than on day 5 (P < 0.05). The highest cell numbers were recovered on days 9 and l0 where the cell number was 10-fold higher than the original number of SBMC, seeded into
20-
15-
lO-
1 2 3 4 5 days of culture
8 9
Fig. 1. [3H]-Thymidine incorporation into SBM cultures in the presence and absence of L-cell conditioned medium. 2 x 105 SBM cells were cultured in microtiter plates. Cell cul- tures were labelled with [3H]-thymidine for 24 hr at days indicated. Shaded blocks show cpm of quadruplet cultures in the presence of L-cell supernatant (30~ L-cell supernatant + 70~o Dulbecco's H21 with 15~o horse serum). Unshaded blocks show cpm of cultures without
L-cell supernatant + SD.

176 PETER MAYER
10-
9-
8-
~ 7 -
x 5 -
,~4-
.~3
"~2-
1! ] 1
2 4 6 8 10 12 14 16 days of culture
Fig. 2. Growth kinetics of SBM in the presence (A A) or absence of L-cell culture super- natants (~, 0). 100 ml of conditioned medium (30% L-cell supernatant + 70% Dulbecco's H21 with 15% horse serum) containing 1.5 x 105 SBMC/ml was sealed in Teflon bags. At times indicated cells were collected and counted in a Thomae hemocytometer. Average cell
number of 4 experiments _+ SD.
the bags. After that time cell numbers declined and on day 16 only half of the day 10 cell population could be harvested. When SBMC were cultured in the absence of the L-cell supernatant a steady decline of the cell numbers could be found and on day 9 no cells could be obtained from the culture bags at all (Fig. 2).
16'
14.
1 2 . ? c~
x 10" E
-~8. OE
.=
Z
U,
K 1:20000 1:2000 1:200 1:20 1:2 Dilution of L-cell supernatant
Fig. 3. [3H]-Thymidine incorporation into SBMC separated on a Ficoll-Metrizoate gradient in response to L-cell conditioned medium. SBMC cells were fractionated on Ficoll-Metri- zoate (density 1.084 g/ml) and 5 x l0 s cells/well were cultivated in microtiter plates for 6 days in the presence of L-cell-conditioned medium (30~o L-cell supernatant + 70~o Dulbecco's H21 with 15% horse serum). L-cell supernatant was diluted 1:2-1:20,000 with Dulbecco's H21 containing 15% horse serum. On day 6 the cells were pulsed with [3H]-thymidine for 24 hr. Averages of quadruplets + SD. Shaded blocks indicate cultivation of cells forming a band;
unshaded blocks indicate cultivation of cells forming a pellet.

Fig. 4. Granulocyte colony on the 4th day of liquid culture. Note the tight structure of the colony (Giemsa stain, x 400).
Fig. 5. Mononuclear phagocyte colony on the 4th day of liquid culture. Note the loose, single-layered colony structure. Most of the cells spread to the cover slip (Giemsa stain,
x 250).
177

Fig. 6. Mononuclear cells on day 12 of liquid culture. Note the presence of round cells with oval nucleus, but also cells spreading to the cover slip can be observed ( x 250).

Swine bone marrow cell cultures, macrophages 179
Separation procedure of SBMC on a density gradient Different densities of the Ficoll-Metrizoate solution for the formation of the discon-
tinuous gradient were compared and a density of 1.084 g/ml chosen. An optimal gradient for separation of SBMC was formed by centrifugation at 420 g for 40min. Two cell fractions could be recovered, one forming a pellet and the other forming a band located on top of the gradient. When cells of the pellet were stained with Wright Giemsa it was found that predominantly granulocytes prevailed.
Effect of L-cell supernatant on proliferation of SBMC separated on a Ficoll gradient
When equal numbers of cells (5 × 105/well) from the pellet and the cell fraction form- ing the band were cultivated in the presence of L-cell supernatant (1:2 final dilution) and [3H]-TdR incorporation determined on day 6 after seeding, it could be shown that the cells forming the band incorporated 5 times more [3H]-TdR than the cells of the pellet. This effect could also be observed with higher dilutions of the L-cell supernatant (1:20 and 1:200). When the L-cell supernatant was diluted to 1 : 2000 and 1 : 20,000 the SBMC cultures did not respond any longer (Fig. 3).
Morphological characterization of the colonies of SBMC cultures
Up to day 14 of culture period two kinds of colonies were observed, one consisting of granulocytic cells and the other of mononuclear phagocytes. The structure of the granulocyte colonies was characterized by the close proximity of the cells (Fig. 4). In contrast, in the mononuclear phagocyte colonies the cells formed a monolayer on the glass surface and showed more separation; the round cells were located more centrally and the elongated cells swarmed towards the periphery (Fig. 5). The ratio of granulocyte to mononuclear phagocyte colonies was of the order of 1:4 when checked on day 4 of culture period. Between days 5 and 8 this ratio changed to 1:8. From day 8 on, only mononuclear phagocyte cells formed a confluent monolayer on the glass surface and were morphologically indistinguishable from macrophages (Fig. 6).
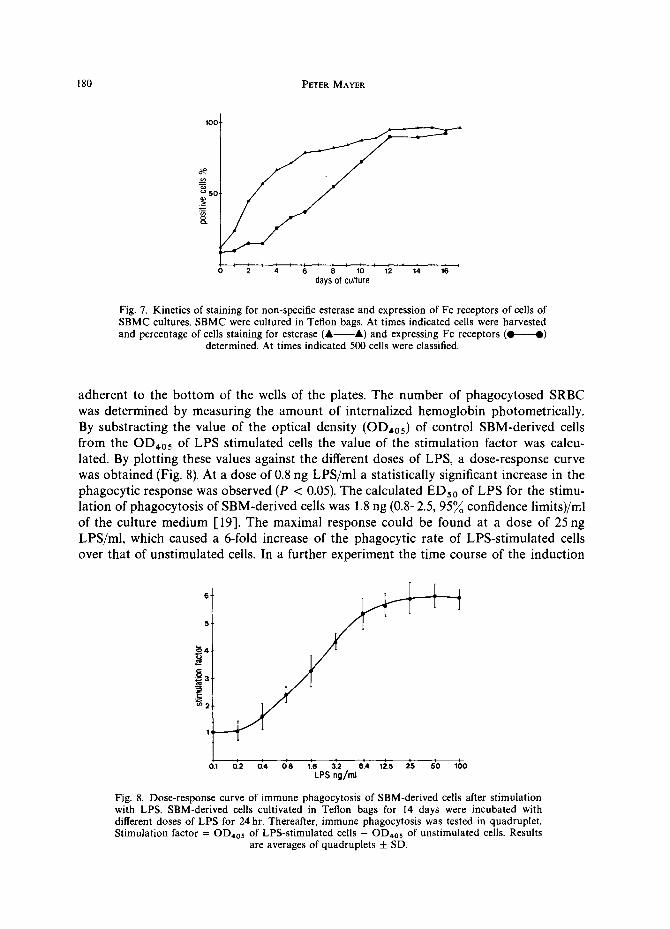
Quantitation of cells staining for esterase and bearing Fc receptors throughout the culture period of SBMC
When SBMC cultures were performed in Teflon bags, the percentage of esterase- positive cells increased from 10~o cells being present in original bone marrow, to 60~o on day 4 and 80~o on day 8 after seeding. From day 12 of the culture period onwards the percentage of esterase-positive staining cells remained in the range of 95~o.
When the percentage of cells expressing membrane receptors for the Fc fragment of the IgG molecule was determined it was found that there was a steady increase in the percentage of Fc receptor-positive cells from 10~o present in the SBM to 90~o on day 12 of culture period (Fig. 7).
Functional characteristics of SBM-derived cells
Quantitation of immune phagocytosis. In a primary experiment phagocytosis was per- formed using SBMC cultivated for 12 days, which thereafter were incubated with differ- ent doses of LPS for 24 hr in microtiter plates. By counting the number of cells in the washing fluid it could be shown that 91 _+ 3~o (N = 20) of the SBM-derived cells were
100'
r.n
~ 5 0 "
0 ~ '~ ' a, ' 6 ' 8 ' 10 ' 12 ' 14 ' 16 ' days of culture
180 PETER MAYER
Fig. 7. Kinetics of staining for non-specific esterase and expression of Fc receptors of cells of SBMC cultures. SBMC were cultured in Teflon bags. At times indicated cells were harvested and percentage of cells staining for esterase (A A) and expressing Fc receptors (.- e )
determined. At times indicated 500 cells were classified.
adherent to the bottom of the wells of the plates. The number of phagocytosed SRBC was determined by measuring the amount of internalized hemoglobin photometrically. By substracting the value of the optical density (OD4o5) of control SBM-derived cells from the OD405 of LPS stimulated cells the value of the stimulation factor was calcu- lated. By plotting these values against the different doses of LPS, a dose-response curve was obtained (Fig. 8). At a dose of 0.8 ng LPS/ml a statistically significant increase in the phagocytic response was observed (P < 0.05). The calculated EDso of LPS for the stimu- lation of phagocytosis of SBM-derived cells was 1.8 ng (0.8-2.5, 959/00 confidence limits)/ml of the culture medium [19]. The maximal response could be found at a dose of 25 ng LPS/ml, which caused a 6-fold increase of the phagocytic rate of LPS-stimulated cells over that of unstimulated cells. In a further experiment the time course of the induction
6"
5 .
~4.
i i o.1 0.2 0.4 o'8 ;.6 ~2 6.4' 1~.s 2'5 5'0 I ;o LPS ng/ml
Fig. 8. Dose-response curve of immune phagocytosis of SBM-derived cells after stimulation with LPS. SBM-derived cells cultivated in Teflon bags for 14 days were incubated with different doses of LPS for 24 hr. Thereafter, immune phagocytosis was tested in quadruplet. Stimulation factor = OD,o5 of LPS-stimulated ceils - OD,os of unstimulated cells. Results
are averages of quadruplets + SD.

Swine bone marrow cell cultures, macrophages 181
5
3 4
days of culture
Fig. 9. Time course of immune phagocytosis of SBM-derived cells stimulated with LPS. SBM were cultivated in Teflon bags in the presence of L-cell-conditioned medium. At times indi- cated cells were harvested and immune phagocytosis assay performed. Stimulation factor = OD4o5 of LPS stimulated cells - OD405 of unstimulated cells. Results are averages
of quadruplets _ S.D.
of phagocytosis of SBM-derived cells was investigated. The culture of SBMC was termin- ated at different times. The cells were incubated with 100 ng LPS/ml for 24 hr and thereafter immune phagocytosis was performed. As indicated in Fig. 9, LPS was not able to stimulate phagocytosis up to day 6 of the culture period. Thereafter, a steady increase in the phagocytic index up to day 14 was found.
Quantitative NBT test with SBM-derived cells upon stimulation with Zymosan and LPS
The effect of different doses of LPS in the presence of Zymosan particles on NBT reduction of SBM-derived cells cultivated for 14 days was determined. The results showed that both cell functions, phagocytosis as well as NBT reduction, could be stimu- lated with LPS in a dose-dependent manner (Fig. 10). With a dose of 60 ng LPS/ml, immune phagocytosis and NBT reduction of SBM-derived cells was increased 7-fold above values of control cells.
Production of prostaglandin of the E-series (PGE)
The culture period of the SBM cells was terminated at different intervals and the cells were then incubated with 100 ng/ml LPS for 24 hr. Up to a culture period of 8 days the cells responded with an increased PGE production upon LPS stimulation (Fig. l 1). When cells of a culture period of 6 days were stimulated with LPS, these cells excreted 20 times more PGE into the supernatant than control cells. When, however, cells were collected at 10 days of culture period, they could no longer be stimulated for an increased PGE production. This indicates that only SBM cells of an early stage of culture period responded with PGE production upon stimulation.
Activation of SBM-derived cells by lymphokines
Table 1 demonstrates that SBM-derived cells could be readily activated to increased phagocytosis by lymphokine-rich supernatants. Phagocytosis was also increased when the supernatant of unstimulated spleen cells was added. This could be observed by the increase of the optical density detected at 405 nm (OD4os) from 0.105 for SBM-derived

182 PETER MAYER
OD
1 .5 ¸
1 . 0 -
0 . 5
Phagocytosis
NBT Reduction
o + 1'0 2'0 ,;o 8'o 8'0 l+o- LPS ng/ml
Fig. 10. Reduction of nitrotetrazolium blue by SBM-derived cells upon stimulation with Zymosan and LPS. SBMC cultivated for 14 days were stimulated for phagocytosis by Zymo- san particles and different doses of LPS. Results are averages of quadruplet tests _+ S.D.
Immune phagocytosis was measured at OD405, NBT reduction was detected at ODsso.
cells not incubated with spleen cell supernatant to 0.214 for cells incubated with spleen cell supernatant. When SBM-derived cells were incubated with a lymphokine-rich spleen cell supernatant generated by ConA (2.5 #g/ml), the OD4o5 was increased from 0.214 to 0.601. Incubation with a lymphokine-rich supernatant generated by PMA (20 ng/ml) led to an increase in the O D 4 o 5 from 0.214 to 0.460.
No increase in immune phagocytosis could be observed when ConA or PMA were added to the cell-free supernatant of unstimulated spleen cells. Therefore, the increased phagocytosis was mediated by a substance produced by the spleen cells upon stimulation with ConA or PMA.
5 ¸
= 4 ¸
~ _ 2
o _
f [ r
0 2 4 6 8 10 12 14 16 days of culture
Fig. 11. Time course of prostaglandin (PGE2 and PGE~) production in SBM-derived cells upon stimulation with LPS. SBMC were cultivated in the presence of L-cell-conditioned medium. At times indicated cells were harvested. 4 × 106 cells were either incubated with 100 ng/ml LPS or left as controls. After incubation of cells for 24 hr PGEs were determined in the supernatants. Averages of quadruplet tests + S.D. Unst imulated cells (& A); LPS-
stimulated cells (Q- - -O) .

Swine bone marrow cell cultures, macrophages 183
Table I. Effect of lymphokine-rich supernatant from ConA- or PMA-stimulated porcine spleen cell cultures on immune phagocytosis determined by the amount of hemoglobin of internalized SRBC of SBM-derived
macrophages
Spleen cell culture supernatant induced with ConA #g/ml*
Immune phagocytosis hemoglobin content (OD405)
of SBM macrophages after incubation with spleen
cell culture supernatant:~ Stimulation factor¶
Immune phagocytosis ConA added directly to spleen cell culture
control supernatant**
Control 1.25 2.5 5.0
10.0
Spleen cell culture supernatant induced
with PMA ng/ml
0.214 ± 0.026 0.348 ± 0.071§ 0.601 ± 0.040§ 0.587 ±0.075§ 0.593 ± 0.063§
1.6 2.8 2.7 2.7
0.214 ± 0.026 0.197 ± 0.047 0.205 ± 0.032 0.185 ± 0.049 0.223 ± 0.027
Immune phagocytosis PMA added directly to spleen cell culture control supernatant
Control 0.214 ± 0.026 0.214 ± 0.026 5 0.342 ±0.048§ 1.6 0.231 ±0.037
10 0.401 ± 0.030§ 1.8 0.193 ± 0.049 20 0.460 ± 0.026§ 2.1 0.201 ± 0.019
* Porcine spleen cells (1 × 10~/ml) were cultured in Dulbecco's H21 + 15~o horse serum for 24 hr. t Aliquots of the spleen cell culture supernatants (100#1) were incubated with 200/~1 of a SBM-derived
macrophage cell suspension (12 day culture) containing 2 × 105 cells for 24 hr. Then immune phagocytosis was performed as described in Materials and Methods by measuring the internalized hemoglobin of SRBC at an OD405. Mean of quadruplets ± S.D.
§ Statistically significant to control supernatant P < 0.01.
OD induced supernatant ¶ Stimulation factor =
OD control supernatant '
** ConA or PMA were added in doses indicated to control spleen cells culture supernatant and SBM-derived macrophages were incubated with these compounds for 24 hr; then immune phagocytosis was performed.
DISCUSSION
The present experiments demonst ra ted that cells in swine hemopoiet ic tissues were capable of responding by proliferation when st imulated by the addi t ion of heterologous CSF. The CSF used originated from a mouse con t inuous fibroblast cell line (L929). Proliferat ion was reflected by increased [3H]- thymidine incorpora t ion into the D N A of
the cells as well as by an increase in the cell numbers in the liquid cultures. Al though it is possible to enumera te the cells responding to CSF by colony format ion
in semisolid med ium by using in.vitro cloning techniques [20, 21], unti l now at tempts to differentiate these cells clearly from other cells of the mouse or h u m a n bone marrow have been unsuccessful because of lack of immunologica l ly specific markers [22, 23]. Also in our case using a Ficoll gradient it was impossible to separate the myeloid progeni tor cells
completely from the other cells of SBM. With the separat ion technique used it seemed that we could achieve an enr ichment of CSF responder cells. However, the reduced thymidine incorpora t ion in response to s t imulat ion with CSF into cells sediment ing into the pellet may not only result from a reduced n u m b e r of responder cells, but can also be due to the suppression of their growth. We found that neutrophi ls accumula ted nearly

184 PETER MAYER
quantitatively in the fraction in which a reduced thymidine incorporation could be detected. It was shown by others that neutrophils exert a negative influence on the development of granulocyte macrophage colonies under semisolid culture conditions 1-24, 25].
In most cases investigating humoral regulation of proliferation and differentiation of hemopoietic cells, semisolid culture systems have been used E26-28]. Despite the proven usefulness of the semisolid culture systems, the precise quantitation of the rates of proliferation, differentiation and functional characterization of the developing cells is unsatisfactory. To overcome these difficulties we used the same liquid culture system for growing SBMC colonies on glass surface as described earlier for the mouse [29, 30]. In this culture system two types of colonies could be identified: mononuclear phagocyte and granulocyte colonies. Most of the cells of the macrophage lineage had Fc receptors on their cell surface and were esterase positive. These features had also been found in both human and mouse macrophages E31-34]. Moreover, the respective granulocyte colonies were formed only during the first days of culture. Using mouse bone marrow as the indicator cell population, the L-cell derived CSF also stimulated preferentially but not exclusively macrophage colony formation [35].
For the functional characterization of these SBM-derived cells, we cultured them in Teflon bags. An advantage of this culture method was that the cells could be easily harvested without damage, cell recovery was almost complete and the cells were functionally unimpaired [36].
These SBM-derived cells performed immune phagocytosis and reduction of Nitro- tetrazolium blue upon stimulation with LPS. Immune phagocytosis is mediated by monocytes, macrophages, polymorphonuclear leukocytes and macrophage-like cell lines [37-39]. The reduction of Nitroblue-tetrazolium is correlated with the stimulation of the activity of the enzyme nicotinamide adenine dinucleotide (NADH) oxidase [40]. The increased oxydase activity indicates a metabolic response of phagocytic cells that is linked to the phagocytic process, yet this response is also performed by both poly- morphonuclear leukocytes and macrophages [41-43].
Macrophages have also been shown to have extensive biosynthetic capabilities result- ing in the secretion of complement components [44], a wide array of enzymes [45, 46], interferon [47] and prostaglandins (PGs) [48]. Available data suggest that among immunocompetent cells, the macrophages are the primary and sole source of PGs pro- duction 1-49]. It has been well accepted that PGs of the E type exert a crucial effect on the induction of the inflammatory response by increasing vascular permeability [50]. PGs and other arachidonic acid metabolites are also generated by macrophages in vitro
[51, 52]. Macrophages release PGE1 and PGE2 in response to various stimuli, e.g. lipo- polysaccharides of Gram-negative bacteria [53]. We also were able to demonstrate that SBM-derived cells secreted PGs in response to stimulation with LPS. It became, how- ever, apparent that only cells of short time cell cultures responded with an increased PGs production upon stimulation with LPS, whereas cells taken from long time cultures did not respond. Yet only these ceils could be stimulated to increased immune phagocytosis by LPS, whereas cells from short time cell cultures showed only a poor response. This divergent effect could possibly be due to different subpopulations of macrophages or to different stages of maturation of these cells in the SBMC culture. Examination of sub- populations of human macrophages isolated by density-gradient fractionation revealed differences among these cells in the amount of PGs synthesized [54].

Swine bone marrow cell cultures, macrophages 185
A further charac ter i s t ic p rope r ty of mac rophages is their abi l i ty to be ' ac t iva ted ' . I t has long been k n o w n tha t mac rophages f rom animals infected with var ious bac te r ia and paras i tes are morpho log ica l ly and funct ional ly different from those of the n o r m a l host. These a l t e ra t ions include increased microb ic ida l and phagocyt ic act ivi ty and increased p roduc t i on of lysosomal enzymes [55-57] . In the course of cel lular immune react ions, mac rophages are though t to be ac t iva ted by med ia to r s der ived from lymphocy tes [58]. Ant igen- and mi togen- s t imula t ed lymphocy tes release soluble med ia to r s cal led lympho- kines which increased the phagocy t i c [59] and microb ic ida l [60] act ivi ty of macro- phages. Biological test ing for ' m a c r o p h a g e a rming factor ' in the mouse system is done using mur ine per i tonea l mac rophages [61].
Recent ly mur ine bone m a r r o w der ived mac rophages have been increasingly used as an a l te rna t ive source of macrophages , especial ly as it cou ld be shown tha t these cells were sensit ive ind ica tor cells for l ymphocy te media to r s [62, 63]. W e too were able to show that SBM-der ived cells r e sponded to h o m o l o g o u s lymphok ine - r i ch superna tan t s of mi togen-s t imula ted lymphocy tes with increased immune phagocytos is .
Us ing the experience of several o ther g roups work ing with o ther an imal species we were able to es tabl ish a re l iable system for the cul ture of S B M C in v i t ro . In long t ime cul tures in the presence of he te ro logous CSF, cells from the SBM prol i fera te and differ- ent ia te into macrophages .
Unt i l now, the only m e t h o d descr ibed for col lect ing swine m a c r o p h a g e s was the lavage of the lung [64]. Less c u m b e r s o m e and more effective, however, could be the use of SBM-der ived m a c r o p h a g e s as an a l te rna t ive source of swine macrophages . Yet this has to be p roven by c o m p a r i n g b iochemica l and funct ional pa rame te r s of porc ine a lveolar and bone m a r r o w der ived mac rophages further.
Acknowledgement--The author is grateful to Dr Fiona Hamilton for reviewing the manuscript.
REFERENCES 1. Carnot, P. and Deflandre, G., Sur l'activit6 hemopoietique du serum au cours de la regeneration du sang.
C.r. Acad. Sci. Paris 143, 384-386 (1966). 2. Bradley, T. R. and Metcalf, D., The growth of mouse bone marrow cells in vitro. Aust. J. exp. Biol. reed. Sci.
44, 287-300 (1966). 3. Ichikawa, Y., Pluznik, D. N. and Sachs, L., In vitro control of the development of macrophage and
granulocyte colonies. Proc. hath. Acad. Sci., U.S.A. 56, 488-495 (1966). 4. Metcalf, B., Studies on colony formation in vitro by mouse bone marrow cells. II. Action of colony
stimulating factor. J. cell. Physiol. 76, 84-100 (1970). 5. Austin, P. E., McCulloch, E. A. and Till, J. E., Characterization of the factor in L-cell conditioned medium
capable of stimulating colony formation by mouse marrow cells in culture. J. cell. Physiol. 77, 121-133 (1971).
6. Yunis, A. A., Arimura, G. K., Wu, M.-C., Gross, M. A. and Purcell, Y., Purification of a colony stimulating factor from culture cell lines propagated from human lung. FEBS Lett. 90, 279 (1978).
7. Stanley, E. R. and Guilbert, L. J., Methods for the purification, assay, characterization and target cell binding of a colony stimulating factor (CSF-1). J. immun. Meth. 42, 253-284 (1981).
8. Austin, P. E,, McCulloch, E. A. and Till, J. E., Characterization of the factor in L-cell conditioned medium capable of stimulating colony formation by mouse marrow cells in culture. J. cell. Physiol. 77, 121 (1971).
9. Bradley, T. R. and Metcalf D., The growth of mouse bone marrow cells in vitro. Aust J. exp. Biol. reed. Sci. 44, 287 (1966).
10. Van der Meer, S. W. M, Bulberman, D., Van Zwet, T. L., Elzenga-Claasen and Van Truth, R., Culture of mononuclear phagocytes on a Teflon surface to prevent adherence, J. exp. Med. 147, 271 (1978).
11. Waithe, W. I. and Hirschhorn, K., Lymphocyte Response to Activators. In Handbook of Experimental Immunolooy, Vol. 2, Cellular Immunology (Ed. by Weir, D. M.), pp. 26.1-26.10. Blackwells, Oxford (1978).
12. Blusse, Van Oud, Alblas A., and Van Furth, R., Origin, kinetics, and characteristics of pulmonary macro- phages in the normal steady state. J. exp. Med. 149, 1504-1518 (1979).

186 PETER MAYER
13. Kaplow, L. S., Cytochemical identification of mononuclear macrophages. In Manual of Macrophage Meth- odology (Ed. by Herscowitz, H. B., Holden, H. T., Bellanti, J. A. and Ghaffar A.), pp. 199-207. Marcel Dekker, New York (1981).
14. Scheetz, M. E., Thomas, L. J., Allemenos, B. K. and Schirnitzky, M. R., A sensitive and reproducible assay for the quantitation of erythrophagocytosis and its correlation with reduction in titrated thymidine uptake in a tumor target cell system modified by immunoenhancing or immunosuppressive agents, lmmunol. Commun. 5, 189 203 (1976).
15. Ross, G. D., Detection of complement receptors and Fc receptors on macrophages. In Manual of Macro- phage Methodology (Ed. by Herscowitz, H. B., Holden, T. H., Bellanti, J. A. and Ghaffor, A.), pp. 209-227. Marcel Dekker, New York (1981).
16. Baehner, R. L. and Nathan, D. G., Quantitative nitroblue tetrazolium test in chronic granulomatous disease. New Engl. J. Med. 278, 971 979 (1968).
17. Jaffe, B. M. and Behrman, H. R., In Methods of Hormone Radioimmunoassay, pp. 19-34. Academic Press, New York (1974).
18. Lohmann-Matthes, M. L., Kolb, B. and Meerpohl, H. G., Susceptibility of malignant and normal target cells to the cytotoxic action of bone marrow macrophages activated in vitro with the macrophage cytotoxic factor (MCF). Cell. lmmunol. 41,231-239 (1978).
19. Spearman-K~irber, H. L., In Statistische Auswertungsmethoden (Ed. Sachs, L.), pp. 357-376. Springer Verlag, Berlin (1969).
20. Iscove, N. N., Sieber, F. and Winterhalter, R. W., Erythroid colony formation in cultures of mouse and human bone marrow: Analysis of the requirement for erythropoietin by gel filtration and affinity chroma- tography on Agarose-Concanavalin A. J. cell. Physiol. 83, 309 320 (1974).
21. Axelrod, A. A., Mc Loed, D. L., Shreeve, H. A. and Heath, D. A., Haemopoesis in Culture (Ed. by Robinson, W.), pp. 226-234. U. S. Government Printing Office, Washington (1974).
22. Cline, M. S. and Billings, R., Antigens expressed by human B lymphocytes and myeloid stem cells. J. exp. Med. 146, 1143-1145 (1977).
23. Janossy, G., Frances, G. E., Capellaro, D., Goldstone, A. H. and Greaves, M. E., Cell sorter analysis of leukemia-associated antigens on human myeloid precursors. Nature 276, 176-178 (1978).
24. Paran, M., Ichikawa Y. and Sachs, L., Feedback inhibition of the development of macrophage and granulo- cyte colonies II., Inhibition by granulocytes. Proc. hath. Acad. Sci., U.S.A. 62, 81-87 (1969).
25. Baker, F. C., Broxmeyer, H. E. and Galbraith, P. R., Control of granulopoesis in man: III Inhibition of colony formation by dense leukocytes. J. cell. Physiol. 86, 337 (1975).
26. Metcalf, D., In Hemopoetic Colonies. In vitro Cloning of Normal and Leukemic Cells, pp. 371-384. Springer Verlag, Berlin (1977).
27. Pike, B. L. and Robinson, W. A., Human bone marrow colony growth in Agar-gel. J. cell. Physiol. 76, 77-84 (1976).
28. Pluznik, D. W. and Sachs, L., The induction of clones of normal mast cells by a substance from conditioned medium. Exp. Cell. Res. 43, 553 563 (1966).
29. Goud, T. J. L. M., Schotte, C. and Van Furth, R., Identification and characterization of the monoblast in mononuclear phagocyte colonies grown in vitro. J. exp. Med. 142, 1180-1190 (1975).
30. Van der Meer, J. W. R., Beelen, W. J., Fluitsma, D. M. and Van Furth, R., Ultrastructure of mononuclear phagocytes developing in liquid bone marrow cultures: A study on peroxydase activity. J. exp. Med. 149, 17 26 (1979).
31. Hay, F. C., Torrigiani, G. and Roitt, L. M., The binding of human Ig subclasses to human monocytes. Eur. J. lmmunol. 2, 257 261 (1977).
32. Heusser, C. H., Anderson, C. C. and Grey, H. M., Receptors for IgG: Subclass specificity of receptors on different mouse cell types and the definition of two distinct receptors on a macrophage cell line. J. exp. Med. 145, 1316 (1977).
33. Kaplow, L. S., Cytochemical heterogeneity of human circulating monocytes. Acta cytologica 19, 358-365 (1979).
34. Berken, A. and Benacerraf, B., Properties of antibodies cytophilic for macrophages. J. exp. Med. 123, 119 144 (1966).
35. Stanley, E. R., Colony stimulating factor (CSF) radioimmunoassay: Detection of a CSF subclass stimulat- ing macrophage production. Proc. natn. Acad. Sci., U.S.A. 76, 2969 2973 (1979).
36. Fleit, H. B. and Rabinovitch, M., Production of Interferon by in vitro derived bone marrow macrophages. Cell. lmmunol. 57, 495-504 (1981).
37. Butz, D. J. and Mills, J., In Basic and Clinical Immunology (Ed. by Fudenberg, H. H., Stites, D. P., Caldwell, J. L. and Wells, J. V.), p. 217. Lange Med. Pub., Los Altos, CA (1978).
38. Walker, W. S. and Demus, A., Antibody dependent cytolysis of chicken erythrocytes by an in vitro- established line of mouse peritoneal macrophages. J. lmmunol. 114, 765 (1975).
39. Snyderman, R., Pike, M. C., Fischer, D. G. and Koren, H. S., Biologic and biochemical activities of continuous macrophage cell lines P 388 D 1 and J 774. J. lmmunol. !19, 2060 (1977).

Swine bone marrow cell cultures, macrophages 187
40. Baehner, R. L. and Nathan, D. G., Leukocyte Oxydase: Defective activity in chronic granulomatous disease. Science 155, 835-836 (1967).
41. Baehner, R. L. and Nathan, D. G., Quantitative nitroblue tetrazolium test in chronic granulomatous disease. New. Engl. J. Med. 278, 971 979 (1968).
42. Rossi, F., Zabucchi, G. and Romeo, D., In Mononuclear Phagocytes in Immunity, Infection and Pathology (Ed. by van Furth, R.), p. 442. Blackwells, London (1975).
43. Yamaguchi, T., Kakinuma, K., Kaneda, M. and Shimada, K., Comparative studies on alveolar macro- phages and polymorphonuclear leukocytes. J. Biochem. 87, 1449-1455 (1980).
44. Littleton, C., Kessler, D. and Burkholder, D. M., Cellular basis for synthesis of the fourth component of guinea-pig complement as determined by a hemolytic plaque technique. Immunology 18, 693-704 (1970).
45. Gordon, S., Regulation of enzyme secretion by mononuclear phagocytes: studies with macrophage plasmin- ogen activator and lysocyme. Fed. Proc. 37, 2754-2758 (1978).
46. Gordon, S., Todd, J. and Cohn, Z. A., In vitro synthesis and secretion of lysozyme by mononuclear phagocytes. J. exp. Med. 139, 1228 1248 (1974).
47. Jullien, P., De Maeyer-Guignard, J. and De Maeyer, E., Interferon synthesis in X-irradiated animals. V. Origin of mouse serum interferon induced by polyinosinic-polycytidilic acid and encephalomyocarditis virus. Infect. Immun. 10, 1023-1028 (1974).
48. Unanue, E. R., Secretory function of mononuclear phagocytes. Am. J. Path. 83, 396 (1976). 49. Goldyn, M. E. and Stobo, J. D., In Critical Reviews in Immunology (Ed. by Atassi, M-Z.), Vol. 2, pp.
189-223 (1981). 50. Williams, T. S. and Morley, J., Prostaglandins as potentators of increased vascular permeability in inflam-
mation. Nature 246, 215-217 (1973). 51. Kurland, J. L. and Bockmann, R., Prostaglandin E production by human monocytes and mouse peritoneal
macrophages. J. exp. Med. 147, 952 (1978). 52. Humes, J. L., Bonney, R. J., Dahlgren, L., Sadowski, M. E., Kuehl, F. A. J. and Davies, P., Macrophages
synthesize and release prostaglandins in response to inflammatory stimuli. Nature 269, 149 (1977). 53. Gemsa, D., B~irlin, E., Leser, H. G., Deimann, W. and Seitz, M., Prostaglandins and leukotrienes: Physio-
logical and Pathophysiological mediators of inflammation. Behring Inst. Mitt. 68, 51-67 (1981). 54. Goldyne, M. E. and Stobo, J. D., Synthesis of prostaglandins by subpopulations of human peripheral blood
monocytes. Prostaglandins 18, 687 (1979). 55, Mackaness, G. B., The immunological basis of acquired cellular resistance. J. exp. Med. 120, 205 (1964). 56, North, R. J., The concept of the activated macrophage. J. Immunol. 121,806 (1978). 57. Cohn, Z. A., The activation of mononuclear phagocytes. J. lmmunol. 121,813 (1978). 58. Rocklin, R. E., In Mediators of Cellular Immunity in Basic and Clinical Immunology (Ed. by Fudenberg,
H. H., Sites, D. P., Caldwell, J. L. and Wells, J. V.). Lange Medical Publ., Los Altos, CA (1978). 59. Nathan, C. F., Karnowsky, M. L. and David, J., Activation of macrophage functions by mediators from
lymphocytes. J. exp. Med. 133, 1356 (1971). 60. Krahenbuhl, J. L. and Remington, J. S., In vitro induction of nonspecific resistance in macrophages by
specifically sensitized lymphocytes, lnfect, lmmun. 4, 337-343 (1971). 61. Patterson, R. J. and Youmans, G. P., Demonstration in tissue culture of lymphocyte-mediated immunity to
tuberculosis. Infect. lmmun. 1,600-603 (1970). 62. Klimitzek, V. and Remold, H. G., The murine bone marrow macrophage, a sensitive indicator cell for
murine migration inhibitory factor and a new method for their harvest. Cell. lmmunol. 53, 257-266 (1980). 63. Meerpohl, H. G., Lobmann-Matthes, M. L. and Fischer, H., Studies on the activation of mouse bone
marrow-derived macrophages by the macrophage cytotoxicity factor (MAF). Eur. J. Immunol. 6, 213-217 (1976).
64. Harmsen, A. G., Birmingham, J. R., Eugen, R. C. and Jeska, E. C., A method for obtaining swine alveolar macrophages by segmental pulmonary lavage. J. immun, Meth. 27, 199-202 (1979).


















