
The feedforward control of posture and movementdigitool.library.mcgill.ca/thesisfile114142.pdf ·...
219
The feedforward control of posture and movement Julia Anne Schaefer Leonard Department of Kinesiology and Physical Education Faculty of Education McGill University, Montreal August 2012 A thesis submitted to the faculty of Graduate Studies and Research In partial fulfilment of the degree of Doctor of Philosophy © Julia Leonard 2012. All rights reserved
Transcript of The feedforward control of posture and movementdigitool.library.mcgill.ca/thesisfile114142.pdf ·...

The feedforward control
of
posture and movement
Julia Anne Schaefer Leonard
Department of Kinesiology and Physical Education
Faculty of Education
McGill University, Montreal
August 2012
A thesis submitted to the faculty of Graduate Studies and Research
In partial fulfilment of the degree of
Doctor of Philosophy
© Julia Leonard 2012. All rights reserved


i
Table of Contents
LIST OF FIGURES ....................................................................................... v
LIST OF TABLES ........................................................................................ ix
Abstract ......................................................................................................... xi
Résumé ........................................................................................................ xiii
Statement of originality ............................................................................... xv
Acknowledgements .................................................................................... xvii
Contributions of authors ............................................................................ xix
List of symbols and abbreviations .............................................................. xxi
Chapter 1 ........................................................................................................ 1 1.1 Scientific Background ..................................................................................2 1.2 Rational ........................................................................................................5 1.3 General Aim .................................................................................................5 1.4 Scientific Objectives and Hypotheses ..........................................................6
Chapter 2 ........................................................................................................ 9 2.1 How are voluntary movements and posture controlled? ......................... 10
2.1.1 The neuroanatomical basis of movement execution .................................. 10 2.1.2 Circuitry of the spinal cord provides a basis for coordinating movement ... 12 2.1.3 Somatotopic organization of spinal cord ................................................... 13 2.1.4 Anatomical organization of the descending pathways for the control of movement ......................................................................................................... 15 2.1.5 Integration of central commands for the global planning of movement and
posture .............................................................................................................. 19 2.2 Postural Control ........................................................................................ 25
2.2.1 Biomechanical requirements for equilibrium control................................. 25 2.2.2 Behavioural goals of the postural system .................................................. 26 2.2.3 Sensorimotor control of posture ................................................................ 27 2.2.4 The problem of motor redundancy ............................................................ 30
2.3 Mechanisms of postural control ................................................................ 32 2.3.1 Overview ................................................................................................. 32 2.3.2 Intrinsic mechanical properties for stability .............................................. 33 2.3.3 Feedback postural responses ..................................................................... 34 2.3.4 Feedforward postural adjustments ............................................................ 37
2.4 Models for movement control ................................................................... 41 2.4.1 Goal-directed movements require both feedback and feedforward control
mechanisms ...................................................................................................... 42 2.4.2 Internal models ........................................................................................ 43
2.5 The control of voluntary arm movements ................................................ 45 2.5.1 Online control of visually-guided reaching movements ............................. 46 2.5.2 Standing imposes equilibrium constraints during perturbed reaching ........ 47
2.6 Summary and direction for future investigation ...................................... 48

ii
Chapter 3 ...................................................................................................... 51 3.1 Rational for experimental protocol ........................................................... 51 3.2 Overview of experimental protocol ........................................................... 53
3.2.1 Experimental apparatus ............................................................................ 53 3.2.2 Behavioural task....................................................................................... 55 3.2.3 Protocol specific to SA1 and SA2 ............................................................. 55 3.2.4 Protocol specific to SA3 ........................................................................... 56 3.2.5 Data collection and analysis ..................................................................... 57
3.3 Significance of the experimental paradigm provides basis for further
exploration .......................................................................................................... 57
Chapter 4 ...................................................................................................... 59 4.1 PREFACE ..................................................................................................... 59 4.2 ABSTRACT................................................................................................... 60 4.3 INTRODUCTION ......................................................................................... 60 4.4 MATERIALS AND METHODS .................................................................. 63
4.4.1 Subjects ................................................................................................... 63 4.4.2 Experimental apparatus and set-up ........................................................... 63 4.4.3 Experimental Procedures .......................................................................... 65 4.4.4 Data analysis ............................................................................................ 68
4.4.5 Statistical analysis .................................................................................... 69
4.5 RESULTS ...................................................................................................... 70 4.5.1 Kinematics of reaching movements during standing ................................. 70 4.5.2 EMG activity in relation to the forces produced: pPA period .................... 74 4.5.3 EMG activity in relation to the forces produced: aPA period .................... 76 4.5.4 Feedforward postural adjustments show directional tuning and are synergic
......................................................................................................................... 76 4.5.5 Spatial patterns of force differ between preparatory and associated postural
adjustments ....................................................................................................... 82 4.6 DISCUSSION ................................................................................................ 88
4.6.1 The roles of preparatory and associated postural adjustments for reaching
during stance .................................................................................................... 88 4.6.2 Tuned, synergic muscle activity characterizes feedforward postural
adjustments ....................................................................................................... 89 4.6.3 Clearly constrained force patterns are seen during preparatory but not
during associated feed- forward postural adjustments ........................................ 90 4.6.4 Implications for the neural control of balance: shared control of feedforward
and feedback postural adjustments .................................................................... 93 4.7 ACKNOWLEDGEMENTS .......................................................................... 94
Chapter 5 ...................................................................................................... 95 5.1 PREFACE ................................................................................................. 95 5.2 ABSTRACT ............................................................................................... 95 5.3 INTRODUCTION ..................................................................................... 96 5.4 METHODS ................................................................................................ 99
5.4.1 Subjects ................................................................................................... 99 5.4.2 Experimental apparatus and set up ............................................................ 99 5.4.3 Data processing and analysis .................................................................. 100
5.5 RESULTS ................................................................................................ 105 5.5.1 Feedforward postural muscle activity is directionally tuned, but shows
variability between trials ................................................................................. 105 5.5.2 Composition and tuning of muscle synergies .......................................... 108

iii
5.5.3 Muscle synergies accurately predict muscle activity patterns .................. 113 5.5.4 Comparison of muscle synergy structure between subjects ..................... 116
5.6 DISCUSSION .......................................................................................... 118 5.6.1 Modular organization of feedforward postural adjustments ..................... 118 5.6.2 Similar organization for feedforward and feedback postural control ........ 120 5.6.3 Conclusions ........................................................................................... 122
5.7 ACKNOWLEDGEMENTS .................................................................... 123
Chapter 6 .................................................................................................... 125 6.1 PREFACE ............................................................................................... 125 6.2 ABSTRACT ............................................................................................. 126 6.3 INTRODUCTION ................................................................................... 127 6.4 METHODS .............................................................................................. 129
6.4.1 Subjects ................................................................................................. 129 6.4.2 Experimental apparatus and set up .......................................................... 129 6.4.3 Experimental procedures ........................................................................ 131 6.4.4 Data analysis .......................................................................................... 135 6.4.5 Statistical analysis .................................................................................. 138
6.5 RESULTS ................................................................................................ 139 6.5.1 Unperturbed reaching and characteristics of online corrections ............... 139 6.5.2 Corrective forces and electromyographic activity accompanying online
corrections of arm movements ........................................................................ 144 6.5.3 Arm-muscle activity responsible for corrections of finger trajectory ....... 147 6.5.4 Corrective postural adjustments in leg muscles lead arm muscle corrections
during online corrections of arm trajectory to unexpected shifts of target position
....................................................................................................................... 147 6.6 DISCUSSION .......................................................................................... 153
6.6.1 Methodological considerations ............................................................... 154 6.6.2 Postural adjustments contribute to the execution of voluntary movement 155 6.6.3 Effects of standing on the characteristics of online corrections of the arm
....................................................................................................................... 156 6.6.4 Implications for the control of posture and movement ............................ 158 6.6.5 Conclusions ........................................................................................... 162
6.7 ACKNOWLEDGEMENTS .................................................................... 162
Chapter 7 .................................................................................................... 163 7.1 Characterization of feedforward postural adjustments during multi-
directional reaching movements ....................................................................... 164 7.1.1 Role feedforward postural activity .......................................................... 164 7.1.2 Independent or parallel commands for global planning of posture and movement ....................................................................................................... 166 7.1.3 Strategies for simplifying the control of posture and movement .............. 167 7.1.4 Force constraint strategy: neural strategy or geometry? ........................... 168 7.1.5 The importance of muscle tuning and synergic organization for feedforward
postural control ............................................................................................... 170 7.2 Central control of posture and movement: integration of feedback and
feedforward postural commands ...................................................................... 173 7.3 Predictive motor control: internal model of posture............................... 174 7.4 Justification for understanding disorders of posture and balance ......... 176 7.5 Conclusions and future directions ........................................................... 177
7.5.1 Do the elderly differ in the spatial and temporal organization of feedforward
postural control? ............................................................................................. 177

iv
7.5.2 Are feedforward muscle synergies robust and how does their recruitment
relate to task-level goals? ................................................................................ 179 7.5.3 Online control of posture: effects of direction and time of visual
perturbation .................................................................................................... 180
Chapter 8: References ............................................................................... 183

v
LIST OF FIGURES
Figure 3. 1: General experimental set-up. Plan view of target light array illustrating subject
orientation on the force plate relative to the array. Targets are arranged from right (0°) to
left (180°). ................................................................................................................... 54
Figure 4. 1: Plan view of the target array and temporal sequence of data collection. A. Subjects
stood on 2 force plates, 1 under each foot and were centered in a 180° light target array,
adjustable for each subject in height and distance (see Methods). Targets (light emitting
diodes, LEDs) were placed at 15o intervals from right to left sides with the position of
each LED set to exactly 130% of their outstretched arm length at shoulder height. Fy =
anterioposterior force, Fx = mediolateral force and Fz = vertical force. B. Temporal
sequence of the data collection period. An auditory tone 500 ms in length sounded to
inform subjects of an impending target illumination. A period of 1000 ms preceded the
onset of the target light upon which subjects were required to reach and point to the
target. The total acquisition period was 3000 ms. A representation of an approximate
movement length (movement time, MT) is shown. ....................................................... 67
Figure 4. 2: 3D kinematic representations of reach to point movements to 3 principal target
directions. (A. 0°, B. 90° and C. 180°) for 1 subject (S5). Stick figures are shown as if
being viewed from the front and slightly to the subject’s left (A) and right (B and C)
sides. Body movements are shown at 2 different times: At the onset of finger movement
(grey sticks) and at the end of the movement (black sticks) when the finger touched the
target. Shown are the following segments bilaterally: foot, shank, thigh, shoulder, upper
arm and lower arm. Markers at the level of the 7th cervical and 10th thoracic vertebrae, the clavicle and sternum form a single segment that represents the torso in the sagittal plane.
The head is represented by markers placed at 4 locations on the left and right temples and
at the same level at the back of the head. Finger trajectory is shown in red. The body
centre of mass (CoM) is shown as a grey and black circles (onset and end of the
movements, respectively). The ground reaction force vector is represented at the onset of
movement in red, and at target attainment in blue. D. Trajectories of the CoM from the
onset of the focal movement (open squares) to when the finger touched the target (open
circles). Trajectories are represented for 5 trials in each direction and are colour-coded as
per the legend across the bottom of the figure. .............................................................. 73
Figure 4. 3 Electromyographic activity, changes in force and vertical torque (Tz) under each
foot for reaching movements to 3 principal directions (A. 0°, B. 90° and C. 180°). Traces
are shown for a period of 500 ms preceding movement onset until the end of each
movement for one typical trial in subject 5. On each plot, the full grey vertical line
indicates the onset of the light target (Light on). The dashed grey line to the left of
movement onset (Movt on) indicates the onset of force and Tz changes during the pPA
period. The dashed grey line to the right of movement onset (between 500 and 750 ms)
indicates the end of the arm movement (Movt end). Forces are shown as forces exerted
against the ground. TFLr and TFLl = tensor facia latae muscles (right and left, respectively), RFr and RFl = rectus femoris, BFr and BFl = biceps femoris, GasLr and
GasLl = gastrocnemius lateralis, , Perr and Perl = peroneus longus, TAr and TAl = tibialis
anterior, Solr and Soll = soleus. Fx = mediolateral force, Fy = anterioposterior force, Fz =
vertical force and Tz = torque exerted around the vertical axis. Left =leftwards, back =
backwards, load = loading and CCW = counterclockwise. Left and right foot forces are
represented by solid and dashed traces, respectively (see legend). Shaded regions

vi
represent the 2 periods of 250 ms under study (pPA and aPA). Successive squares
underneath the time axes pictorially represent changes in Tz at each foot during the two
periods. ....................................................................................................................... 75
Figure 4. 4 Representative EMG traces for 14 selected muscles for S5 across the 13 directions
of pointing. Muscle activity is shown for a total duration of 500 ms, 250 ms before and
after the onset of the pointing movement. Muscle name conventions are as described in
Figure 4.3. The shaded area to the left of time zero on each muscle plot represents the 250
ms preparatory period. Unless shown, muscles have the same scaling for the left leg (top
row) as they do for the right leg (bottom row). ............................................................. 78
Figure 4. 5 Muscle tuning curves for the EMG activity of all 14 postural muscles during the
final 3 bins of the preparatory and associated periods for the representative subject S5.
Differences in tuning and recruitment of the muscle studied can be observed by
comparing the activity of the muscles over the 3 5 equivalent bins (left to right columns).
Dots indicate amplitudes from each trial measured and the solid lines the mean responses.
Muscle name conventions are as described in Figure 4.3............................................... 80
Figure 4. 6 Muscle tuning curves in the final bin of the pPA (A) and aPA (B) periods for each
of the 8 subjects studied. Muscles have been grouped into the 3 major groups that
activated for similar directions of reach. Tuning curves and individual trials are
represented as in Figure 4.5. Schema (C) summarizing the approximate range of
directions of reach to which each identified group contributed. ..................................... 81 Figure 4. 7 Individual resultant horizontal ground reaction force vectors and average values of
Fz produced during the pPA period (A, B, respectively) and the aPA period (C, D) for
subject 5. Forces are shown for each consecutive bin during each period in successive
rows from top to bottom. Black and grey arrows represent the approximate direction of
exerted force under the loaded and unloaded feet, respectively. In B and D, bars above the
top of the plots marked ‘L’ indicate directions of reach for which Fz under each
respective foot was loaded. For reference, the directions of reach used are indicated on the first plot (left foot) for the pPA1 period in Fig. 4.6A. .................................................... 84
Figure 4. 8 Average direction and magnitude of horizontal ground reaction force change during
each bin of the pPA and aPA periods under both feet. .................................................. 86
Figure 5. 1 Representative traces of muscle activity in seven muscles recorded bilaterally
during a 500 ms period preceding movement onset (Movt On) to movement end. The
pPA period is indicated by the shaded vertical grey area to the left of movement onset.106
Figure 5. 2 Muscle tuning curves for all 14 muscles and all 5 time bins during the 250 ms
period preceding the onset of reaching movements in a representative subject (S011). .107
Figure 5. 3 Variability accounted for (VAF) for different number of muscle synergies for the
entire data set for a representative subject (S010). To choose an appropriate number of
muscle synergies (Nsyn), the following criteria were met: (A) Overall VAF attained a
threshold of > 90%; (B) VAF by muscle as a function of muscle synergy number. (C)
VAF by synergy number as a function of muscle was plotted to confirm that chosen Nsyn
met the criteria of 75%. (D) VAF by direction and bin shows that directions were well
characterized. .............................................................................................................109
Figure 5. 4 A: Muscle synergy vectors (W) and B: recruitment coefficients (C) for a
representative subject (S010) across all 5 time bins of the pPA. Synergy activation
coefficients for individual trials are shown by a dot, and average muscle synergy
recruitment is shown by a solid line that illustrates its directional tuning. .....................112

vii
Figure 5. 5 Reconstruction of mean muscle tuning curves using the muscle synergies shown in
Fig. 5.4 for a representative subject (S010). Dashed lines represents observed data and
solid lines represent reconstructed data. Each muscle synergy’s contribution is shown by
the corresponding colored line. Combined, these result in the total reconstruction.
Goodness of fit (VAF, r2) of the reconstruction to the observed EMG is indicated. ......114
Figure 5. 6: Postural muscle activity in the REM trials for reaching to the 75° target is
reconstructed using the extracted muscle synergies for a representative subject (S010).
Variations in the activation levels of the muscles between trials is well accounted for by
the modulation of the muscle synergies. Muscle activation amplitudes for all recorded
muscle are grouped along the x-axis within a trial. Star (✴) represents the observed EMG
and open circle ( ○) is the reconstructed activation amplitude. The relative contributions
of the muscle synergies to the overall muscle activity is shown by the colours in the
vertical histogram for each muscle. .............................................................................115
Figure 5. 7 Muscle synergy structure compared between subjects. Muscle synergies that are
shared between subject are indicated by a significant VAF and r2. Muscle synergies
whose backgrounds are shaded gray are specific to that subject. ..................................117
Figure 6. 1: Experimental set-up and data collection schema. A. Subjects stood on 2 force
plates reached to a central target, aligned with their xiphoid process. Unperturbed ‘reach’
trials were interspersed with online correction (‘corr’) trials involving unexpected illuminations of 1 of 3 other targets placed successively at 15° increments to the right of
centre. B. Explanation of the changes in voltage related to the sequence of light changes.
When the signal rose to 5V each light was illuminated. L1 = light one, L2 =light 2, chest
= chest switch attached around the subjects sternum that acted as a signal from which L2
illumination could be triggered. C. A histogram showing the distribution of L2 onset as a
percentage of mean ‘reach’ peak velocity. Trials from all ‘corr’ conditions have been
pooled (n=652). rFin = right finger. .............................................................................133
Figure 6. 2: Determination of the online correction of finger trajectory (fcorrect). A. Plan view
(x,y) of rFin average ‘reach’ trajectories +/- 1SD (dashed line with shaded grey area) in
relation to one ‘corr45’ trial (full black trajectory). Filled black circle is the onset of light
2 (L2 onset) and the open circle is the time at which the corr45 x,y trajectory exceeded
the average ‘reach’ trajectory plus 1SD for subject S6. B. Average (dashed line) plus 1SD
of curvilinear rFin velocity for a reach movement and one ‘corr45’ trial(full black line).
Black vertical line is light 2 (L2) onset, grey vertical line is the time of online correction
(fcorrect). Each corr condition has been displaced rightwards and downwards for clarity,
but the starting position was the same for each. C, D: Explanation of how the correction
of the EMG activity and GRFcorrect associated with online corrections were determined.
C. Calculation of EMGcorrect. The muscle shown is the left soleus muscle, but the same
procedure was used with all other muscles studied (see Methods). The dashed trace and grey traces represent respectively, the mean ‘reach’ soleus muscle activity ± 2SDs above
and below the mean. The dark full trace represents the soleus muscle activity produced
during an online correction movement, in this example a corr45 movement. The open
circle indicates the time at which the corr45 soleus muscle activity exceeded the
mean+2SD ‘reach’ activity level. This time was taken as the EMGcorrect time (for more
detailed explanation, see Methods and Results). Abbreviations as previous figures, except
Movt end=movement time. D. Calculation of GRFcorrect. Method for determining
GRFcorrect is shown for the left shear force (Fx). The dashed trace and grey traces
represent respectively the mean ‘reach’ Fx and ± 2SDs above and below the mean. The
dark full trace represents the Fx exerted during an online correction movement (in this
example corr45). The open circle indicates the time at which the exerted force was significantly different from the mean forces exerted in a ‘reach’ trial. ..........................136

viii
Figure 6. 3: Reaching movement kinematic characteristics and profiles of curvilinear velocity.
Shown are averages plus 1SD for all trials for subject 2 in each of the 4 conditions
studied. A. Plan view (x,y) kinematics of rFin trajectory for ‘reach’ trials and each of the
correction conditions. B. rFin curvlinear velocity also for all 4 conditions.
fcorrect=kinematic correction of finger trajectory, mvt end=end of the focal movement
(reach and corrected movements). ...............................................................................141
Figure 6. 4 Typical arm and leg muscle activity in relation to the 3D ground reaction forces
produced for a ‘reach’ movement (A) and an online correction movement to the target
placed 45° to the right of midline, ‘corr45’ (B). In each, the muscles plotted in grey are
those recorded in the right leg. The vertical dashed line indicates initial movement onset
(‘Mvt Onset’) and the full vertical black line, movement end (‘Mvt End’). In B., the
shaded grey area indicates the area in which arm and postural adjustments occurred. For
muscle abbreviations, see Methods. L2 onset=light 2 onset, fcorrect=time of kinematic correction of the rFin marker. Fx=mediolateral force, Fy=anteriorposterior force and
Fz=vertical force. ........................................................................................................146
Figure 6. 5: Linear regressions calculated between the four arm muscles recorded in the right
am and the fcorrect latencies calculated using the curvilinear kinematics of the rFin
marker. A. right posteior deltoid, B. right triceps, C. right anterior deltoid and D. right
biceps. Yi=the value of the Y intercept when X is zero. ...............................................149
Figure 6. 6: Multiple comparisons of differences between EMGcorrect values of arm and leg
muscles. A. Average differences EMGcorrect differences per ‘corr’ target. Values for all
3 arm and leg muscles have been pooled (averages for each ‘corr’ target +/- 95%
confidence interval, CI). Positive differences indicate postural muscle corrections before
arm corrections (see direction of arrow, top right of figure). B. Average differences (+/-
810 .................... 95% CI) EMGcorrect (all arm muscles pooled) per leg muscle. Values for
EMGcorrect measures were pooled for all arm muscles and expressed as differences with
each leg muscle in turn (positive differences also indicate postural muscle corrections before arm muscles). C. EMGcorrect differences (+/- 95% CI) per phase of reach, i.e.
before peak velocity (Acceleration) or after peak velocity (Deceleration). Filled circles
show mean EMGcorrect differences for all targets, while open circles show only data for
corr45; shaded area represents 95% confidence interval. D. Average differences (+/- 95%
CI) EMGcorrect (all arm muscles pooled and all leg muscles pooled) per subject.........150

ix
LIST OF TABLES
Table 4. 1 Mean (±1SD) of movement times for pointing movements in all 13 directions ...... 72
Table 4. 2 Results of the broken-axis approach (Holmquist & Sandberg, 1991) ..................... 87
Table 6. 1 Breakdown of total number of trials collected and retained after trial selection
procedure. ..................................................................................................................143
Table 6. 2 Slope (m), Y intercept (Yi), r2 values, p-value for the strength of the regression fit
(p) and p-value for the intercept (p-int) for linear regressions conducted between the leg
and arm muscles selected to characterize the online corrections to all targets for each subject ........................................................................................................................152

x

xi
Abstract
Goal-directed arm movements performed in the standing position
potentially disturb the body's equilibrium as a result of the multi-linked
structure of the musculoskeletal system. To compensate for these disturbances
and ensure that stability is maintained, the central nervous system (CNS)
organizes postural adjustments preceding and accompanying the voluntary
movement in a feedforward manner (Massion 1992) using knowledge of the
dynamics of the body (Bouisset and Zattara 1981). To date, most studies
investigating the control of posture during voluntary movements in humans
have focused on either the role of the postural activity preceding the movement
or on the temporal structure of these anticipatory postural adjustments (APAs)
with respect to the focal movement. As such, detailed knowledge about the
spatial organization of postural activity is lacking. Furthermore, it is not clear
how posture is coordinated when the goal of a voluntary movement changes
online. Therefore, the studies in this thesis were aimed at addressing these
questions to develop a greater understanding of the organization of feedforward
postural control during voluntary movements.
Muscle activity, kinetics and kinematics were recorded as subjects
performed unperturbed and perturbed reaching movements to targets located in
multiple directions while standing. Feedforward postural control strategies
preceding and accompanying the reaching movements were quantified.
Characterization of the spatial and temporal patterns of muscle activity and
ground reaction forces of postural adjustments preceding reach movements
revealed that muscle activity was directionally-tuned to reach direction and
forces that were constrained to two principal directions. Also, muscle synergies
were able to explain the spatial and temporal variability in postural muscle
activity in the period preceding the reaching movements, suggesting that a
modular organization of muscle recruitment is adopted for this task. Overall,
these strategies are similar to those observed for feedback postural responses,

xii
suggesting that the CNS relies on shared neural structures for controlling
posture in both modes of control. Lastly, the nature of postural control was
examined when reaching movements were perturbed with a shift of the visual
target after the reaching movement was initiated. Here, muscle activity in the
legs was consistently modulated prior to changes in the muscle activity related
to the online correction of the arm trajectory.
Taken together, the findings of this thesis provide important insights
into how the brain coordinates the control of posture and movement. This work
provides a measure of feedforward postural control strategies in healthy, young
adults as a first step to understanding how and why deficits in balance control
may occur during the execution of voluntary movements in fall-prone
individuals.

xiii
Résumé
Les mouvements volontaires effectués dans la position debout peuvent
engendrer des perturbations de l’équilibre en raison de la structure complexe du
système musculo-squelettique. Pour amorcer ces perturbations et s’assurer que
l’équilibre est maintenu, le système nerveux central (SNC) amorce le
déplacement du centre de masse (CM) par la mise en jeu d’ajustements
posturaux avant et accompagnant les mouvements programmés en mode
proactif (Massion 1992) en utilisant des représentations internes du corps et de
l’environnement. À ce jour, la majorité des études portant sur le contrôle de la
posture lors des mouvements volontaires chez l’homme ont comme but soit
l’identification du rôle ou la caractérisation de la structure temporelle de ces
ajustements posturaux anticipateurs. Cependant, une connaissance approfondie
concernant l’organisation spatiale de l’activité posturale est manquante. De
plus, ce n’est pas évident comment la posture est coordonnée lorsque le but du
mouvement change après le commencement du mouvement. Ainsi, les études
présentées ici ont comme but de répondre à ces questions pour développer une
meilleure compréhension de l’organisation centrale de la posture et le
mouvement.
Les signaux électromyographiques, les forces de réaction au sol et la
cinématique tridimensionnelle ont été enregistrés pendant que les sujets
effectuaient des mouvements de pointage vers des cibles distinctes dans la
position debout. Les stratégies posturales organisées en mode proactif ont été
quantifiées sans pertubations et avect des pertubations visuomotrices des
movements d’atteinte. La caractérisation de l’organisation spatiale et temporelle
de l’éléctromyographie et des forces appliquées au sol ont révélé que l’activité
des muscles était biaisée vers la direction de pointage (‘directionally-tuned’)
mais que les forces au sol étaient appliquées dans un nombre de directions
limitées (‘force constraint strategy’). De plus, la variabilité spatiale et
temporelle de l’activité des muscles posturaux était expliquée par les synergies

xiv
musculaires. Ceci suggère qu’une organisation modulaire est utilisée par le
SNC pour faciliter la tâche de contrôle de la posture. Ces stratégies sont
similaires à celles observées pour les ajustements posturaux compensatoires (à
base de ‘feedback’ ou rétroaction), ce qui suggère que le SNC dépend des
mêmes structures neuronales pour contrôler la posture dans la mode proactif et
rétroactif. Par la suite, la nature du signal pour le contrôle de la posture a été
examinée lors des mouvements de pointage qui ont été perturbés avec un
déplacement de la cible visuelle après que le mouvement ait été commencé. Ici,
l’activité musculaire dans les jambes était modulée avant la modulation de
l’activité musculaire liée à la correction de la trajectoire du bras.
Ensemble, les conclusions de cette thèse fournissent un aperçu
important sur la façon dont le cerveau coordonne le contrôle de la posture et du
mouvement. Les résultats présentés supportent la conclusion que les
commandes centrales pour la posture et le mouvement interagissent dans le
SNC, et que les structures neuronales sont partagées pour la posture organisée
de façon anticipatoire, ou proactif, et compensatoire. Les stratégies posturales
typiques dans les jeunes adultes en santé sont quantifiées et forment une base de
données pour la comparaison avec des gens sujets au déséquilibre lors de la
performance des mouvements volontaires.

xv
Statement of originality
This thesis incorporates the outcome of research undertaken under the
supervision of Dr. Stapley in the department of Kinesiology and Physical
Education, at McGill University for the requirements of Doctor of Philosophy. I
certify that this thesis, and the research to which it refers are the product of my
own work and has not been published elsewhere except where specific
references are indicated. The manuscripts presented in chapters 4, 5 and 6
represent original material and contribute to the advancement of knowledge in
the fields of posture and movement control. To my knowledge, the studies
presented within this thesis are the first to investigate the organization of feed-
forward postural adjustments during multidirectional reaching performed in
standing in human subjects.
All data presented in this thesis were collected in the BVML (Balance
and Voluntary Movement Laboratory), located in the department of
Kinesiology and Physical Education at McGill University. The protocols used
in the studies herein were approved by the McGill University Research Ethics
Board.

xvi

xvii
Acknowledgements
Over the course of my studies at McGill, I have had the opportunity to
interact with and learn from many talented and inspiring individuals. I am
deeply thankful for all of these experiences, as they have been influential in
shaping my ideas about science, my ability to ask questions and test reasonable
hypotheses.
In particular, I would like to thank my thesis supervisor, Dr. Paul
Stapley. It goes without saying that the work presented in this thesis would not
have been possible without his unrelenting support and patience. His guidance
has been instrumental in my development as a scientist from a naive first-year
Master’s student. In particular, he encouraged independence while maintaining
a level of support I could always count on. I will be forever grateful for the
skills I developed while working in his laboratory. He helped me create a
foundation that I hope to build upon as I continue to mature as a scientist.
I would like to thank my committee members: Drs. Trevor Drew, David
Ostry and Ted Milner for their insight and feedback. In particular, I wish to
thank Dr. Trevor Drew, who provided important feedback on the manuscripts
published in this thesis. I am also very thankful for the extensive support Dr.
Drew has provided as I transition beyond my doctoral studies. I admire his
passion for scientific inquiry and I look forward to his guidance and
mentorship. I wish to also thank Dr. Milner for maintaining a space where I
could work in the final stages of the preparation of this thesis.
I am deeply grateful for the assistance of many people in the
Department of Kinesiology and Physical Education. In particular, Ryan
Ouckama, provided invaluable technical knowledge and accessibility for
trouble-shooting for the experimental set-up; J.J. Loh, shared his love of Matlab
and taught me foundations in computer programming; past and present
members of the BVML lab, including Ryan Brown, Alicia Hilderley, Silvia

xviii
Hua and Will Lee-Shanok, thank you for sharing such an important part of my
intellectual and personal development.
And to the many others, friends and collaborators beyond BVML, thank
you. Dr. Lena Ting, for graciously welcoming me to her lab to learn the
techniques for the synergy analysis; ‘Team Synergy’ - in particular, Drs. Stacie
Chvatal and Seyed Safavynia, thank you for your prompt and thorough
responses to my many questions. I would like to thank Dr. Rob Kearney for his
insight on data analysis and Dr. Jane Macpherson for her comments on the
studies presented in this thesis. And on a personal level, thank you Dr. Karen
Lomond, Dr. Catherine Sabiston and Marilee Nugent for all your support along
this tremendous journey. Also, this work would not have been possible without
the financial support from Fonds de la Recherche en Santé du Québec (FRSQ),
National Science and Engineering Research Council (NSERC) and Canada
Foundation for Innovation (CFI; infrastructure).
Finally, this thesis is the outcome of hard work and dedication that
would not have been possible without the unrelenting support and
unconditional love from my family and close friends. In so many ways, this
thesis reflects the efforts and contributions from so many. Firstly, thank you
Eric. You have been steadfast in your words of affirmation and belief in me.
Thank you for all the sacrifices you have made so that I may pursue this goal.
You are an amazing father and I am truly blessed to share this life with you.
Mom and Dad, you modeled perseverance and commitment to a task. I would
not be where I am today without the generous and unwavering support you
provided. This thesis, and so much more, could not have been completed
without you. To my siblings, Erica, Andrew and David, thank you for your
constant encouragement. Sue and Dave, thank you for the countless hours of
babysitting and belief in me. And lastly, thank you to my two beautiful girls,
Theresa and Madeline. You inspire me to work hard, to focus and be a good
example for you. I have already learnt so much from you both; I love you
dearly.
Julia

xix
Contributions of authors
The work presented in this thesis is primarily the work of the author,
Julia Leonard, including the conception of ideas, development of the
experimental protocol, data collection, data analysis, presentation of results and
preliminary drafts of manuscripts for publication. The outcome of this research
has resulted in two published manuscripts (Chapters 4 and 6) and provides the
basis for a third one in preparation for publication (Chapter 5).
Chapter 4 contains a published manuscript: Leonard JA, Brown RH,
Stapley PJ. Reaching to multiple targets when standing: the spatial organization
of feedforward postural adjustments. J Neurophysiol 101: 2120–2133, 2009.
For this study, I built the experimental set-up, collected and analyzed the data,
prepared figures for presenting the results and prepared and revised the
manuscript following peer-review. Ryan Brown is listed as second author for
his contributions in building the experimental set-up and initial data collection
and preliminary analyses. Contributions from JJ Loh for technical assistance
and Drs. Jane Macpherson and Trevor Drew for insightful discussions are noted
in the acknowledgments section of the manuscript. As my thesis supervisor, Dr.
Stapley provided guidance and insight in all stages of the study and preparation
of the final manuscript.
Chapter 5 contains a manuscript in preparation for the Journal of
Neurophysiology: Leonard JA, Chvatal S, Ting LH, Stapley PJ. Muscle
synergy characterization of feed-forward postural adjustments during reaching
in standing humans. For this study, I built the experimental set-up, collected
and analyzed the data, prepared figures for presenting the results and prepared
the manuscript for submission to peer-review. Dr. Stacie Chvatal provided the
basic Matlab NNMF algorithms, which I customized to my analysis, as well as
important advice about the data analysis and interpretation of results. Dr. Lena
Ting provided advice about interpretation of results and collaborated in
preparing the final version of the manuscript. Dr. Safavynia provided additional

xx
advice about the data analysis and is mentioned in the acknowledgements
section of the manuscript. As my supervisor, Dr. Stapley provided guidance and
insight in all stages of the study and preparation of the final manuscript.
Chapter 6 contains a published manuscript: Leonard JA, Gritsenko V,
Ouckama R, Stapley PJ. Postural adjustments for online corrections of arm
movements in standing humans. J Neurophysiol 105: 2375–2388, 2011. For this
study, I built the experimental set-up, collected and analyzed the data, prepared
figures for presenting the results and prepared and revised the manuscript
following peer-review. Dr. Gritsenko assisted with portions of the data analysis
and collaborated in preparing the final version of the manuscript. Ryan
Ouckama provided technical expertise in programming the experimental set-up.
Dr. Rob Kearney provided advice about the data analysis and is mentioned in
the acknowledgements section of the manuscript. As my thesis supervisor, Dr.
Stapley provided guidance and insight in all stages of the study and preparation
of the final manuscript.

xxi
List of symbols and abbreviations
APA Anticipatory postural adjustment
aPA Associated postural adjustments
APR Automatic postural response
BoS Base of support
CM Centre de masse
CNS Central nervous system
CoM Center of mass
CoP Center of pressure
CPG Central pattern generator
CST Corticospinal tract
DoF Degree of freedom
Fx Mediolateral force
Fy Anteroposterior force
Fz Vertical force
EMG Electromyography
GRF Ground reaction force
ICA Independent component analysis
LED Light emitting diode
NNMF Non-negative matrix factorization
PCA Principal component analysis
pPA Preparatory postural adjustment
PMRF Pontomedullary reticular formation
SNC Système nerveux central
STA Spike-triggered averaging
Tz Moment of force about the vertical (z) axis

xxii

1
Chapter 1
General Introduction
The excitement felt by a parent watching their infant stand for the first
time, and the sense of accomplishment radiating from the infant’s face, suggest
that there is an inherent understanding that the seemingly simple act of
standing, or balancing on two feet, is in fact, a major accomplishment. Armed
with the skill of upright stance, the infant enters an entirely new world of
discovery and exploration, using their hands to grasp, manipulate and move
objects while standing. Stance control, as developed from infancy, is a
fundamental skill that affords the ability to interact with our environment in
very intimate, creative and useful ways. Moreover, while most people take the
skill of reaching while standing for granted, an aging person afflicted with
balance deficits will tell you that performing such a dynamic task is challenging
and poses significant threats to their stability. This is particularly apparent in
situations where the task may change once a movement has already been
initiated, such as reaching for a support that may change positions after the
reach began. In fact, falls, which have significant consequences on health and
lifestyle, are typically experienced in these dynamic situations (Cavanagh et al.
1992; Horak et al. 1989b). To better understand why balance deficits occur in
these dynamic situations, knowledge about the fundamental strategies used by
the central nervous system (CNS) for coordinating the control of balance and
voluntary movements is needed. This information will provide a framework for
understanding balance deficits and the creation of novel rehabilitation
programs.

2
1.1 Scientific Background
Many of our daily activities involve reaching towards objects in our
extrapersonal space while maintaining upright stance. This apparently simple
task actually involves two divergent goals that must be coordinated by the
nervous system (Hess 1943; Massion et al. 2004). Specifically, the CNS must,
on the one hand, plan the trajectories of the limb segments to the goal
(movement), and on the other, maintain the stability of the limbs and balance of
the whole body (posture) (Horak and Macpherson 1996; Massion 1992;
Massion 1998). This duality between posture and movement, and their
underlying coordination, is non-trivial, and has been stressed since the early
studies of Babinski (1899), Hess (1943), Bernstein (1967), and several others
(Cordo and Nashner 1982; Massion 1992; Nardone and Schieppati 1988).
Moreover, the challenge of controlling these two divergent tasks is further
complicated by the need to control the many limbs, joints, and muscles
involved in a specific motor behaviour in the context of a given set of task
constraints. This problem is commonly referred to as the degree of freedom
(DoF) problem (Bernstein 1967). Consequently, it is thought that the CNS must
somehow simplify the complex task of controlling balance and movement with
suitable strategies so that skillful motor performance is achieved.
Due to the mechanical properties of the musculoskeletal system,
movement of any limb segment causes a shift in the position of the centre of
mass (CoM) that is disturbing to equilibrium (Hollerbach and Flash 1982;
Winter and Eng 1995). Furthermore, goal-directed, or focal, movements
involve complex spatial and temporal patterns of muscle activity in multiple
muscles throughout the body that result in net reaction forces that may also
have a perturbing effect on the body (Bouisset and Zattara 1981; 1987; Brown
and Frank 1987; Crenna et al. 1987; Friedli et al. 1988). To compensate for
these disturbances to equilibrium arising from the voluntary component of
actions, the CNS relies on postural muscle activity to stabilize the body (Horak
and Macpherson 1996; Massion 1992).

3
In the case of a voluntary movement, the CNS is able to predict the
mechanical consequences of the goal-directed movement and organizes the
appropriate muscle activity in the supporting limbs to compensate for the
disturbance (Bouisset and Zattara 1987; Cordo and Nashner 1982; Massion
1992). These postural adjustments are planned in advance of the movement and
are programmed using a feedforward mode of control (Gahery and Massion
1981; Massion 1992). In contrast, when balance is unexpectedly perturbed, the
CNS uses afferent information from the visual, vestibular and somatosensory
systems to shape feedback-based automatic postural responses (APRs) to
restore equilibrium (Nashner 1977).
Studies investigating postural responses to unexpected perturbations of
quiet stance have revealed much about the underlying control of posture. In
particular, specific strategies for controlling balance have been identified and
are thought to serve as a general mechanism by which the CNS simplifies the
coordination of motor behaviour (Ting 2007). For example, in response to
unexpected translations of the support surface in multiple directions, both cats
and humans restore equilibrium by using a ‘force-constraint’ strategy
(Macpherson 1988a), which involves exerting forces at the ground by each
supporting limb in only one of two directions irrespective of the direction of
translation (Henry et al. 1998a; Macpherson 1988a). In contrast, the
corresponding postural muscle activity is characterized by broad tuning across
perturbation direction, with maximal activity typically occurring for a single
characteristic direction (Carpenter et al. 1999; Henry et al. 1998b; Macpherson
1988b). More recent investigations of the coordination of muscle activity
underlying the feedback postural responses have revealed that the CNS appears
to simplify the task of balance control by constraining muscles to be activated
in fixed groups, or synergies (Ting and Macpherson 2005; Torres-Oviedo et al.
2006; Torres-Oviedo and Ting 2007). According to this model, the CNS sends
a command to activate a group of muscles, rather than controlling each muscle
individually. These muscle synergies correlate to the endpoint force vector

4
exerted under each supporting limb (Chvatal et al. 2011; McKay and Ting
2008; Ting and Macpherson 2005).
Muscle synergies are proposed to be a mechanism by which task-level
commands, such as CoM stabilization, are translated into execution-level
commands, which are the spatial and temporal patterns of muscle activity
(Chvatal et al. 2011; Ting 2007; Ting and Mckay 2007). Furthermore, muscle
synergies provide an attractive solution to the problem of controlling the
overabundant DoF associated with the multi-segmental arrangement of the
human body (Chiel et al. 2009; Ting 2007). It is not known, however, whether
similar strategies exist for feedforward postural control, in which postural
commands are organized in a predictive manner using knowledge of the body’s
dynamics and the external world (Bouisset and Zattara 1981; 1987).
During the execution of voluntary movements in the standing position,
preparatory anticipatory postural adjustments (pPA) occur prior to the onset of
the movement and serve to either stabilize the body or initiate movement,
whereas associated anticipatory postural adjustments (aPA) occurring during
the movement ensure a stable transition from one body configuration to
another. This postural activity is programmed in feedforward (Massion 1992)
and relies on prior knowledge of the arm and body dynamics (Bouisset and
Zattara 1981; 1987). Furthermore, anticipatory muscle activity in the supporting
limbs preceding a voluntary movement suggests that the CNS relies on a
predictive estimate of the future state of the body (Davidson and Wolpert
2005). This feedforward postural muscle activity is typically studied in
paradigms where the final goal of the movement is planned and therefore the
disturbance due to the movement can be predicted. For example, during a
planned voluntary movement, such as an arm raise, it is possible to predict the
dynamic consequences of the focal movement on equilibrium and therefore
plan the commands for posture accordingly (Massion 1992). However, if the
goal of the movement changes after the movement has been initiated, it is not
known how the CNS modifies postural control in relation to the online focal
correction in order to ensure that the target goal is met and equilibrium is

5
maintained. Investigating this question will provide fundamental insights about
how posture and movement commands interact, which has important
implications for understanding the deficits of balance control observed in
clinical settings.
1.2 Rational
To date, most studies investigating the control of posture during the
execution of voluntary movement have focused on either the role of the
anticipatory postural adjustments (APAs) preceding the movement or on the
temporal structure of these APAs with respect to the focal movement.
Consequently, there is still a lack of detailed information regarding the
organization of these postural adjustments when voluntary movements are
executed in multiple directions. Knowledge about how postural muscle activity
is coordinated with respect to movement direction is necessary to understand
how the nervous system may simplify the task of coordinating posture and
movement. Furthermore, no study to date has specifically addressed the online
control of posture in situations where the task goal changes after the movement
is initiated. These issues need to be clarified, both for a fundamental
understanding of the brain and motor system, as well as for a better
understanding of the postural deficits observed in several pathologies and
disease.
1.3 General Aim
As highlighted above, several specific questions remain about how
posture is controlled in relation to voluntary movement. Thus, the general aim
of the present thesis was to develop a greater understanding of the feedforward
adjustments of posture during voluntary movement. This was achieved with a
series of studies that (1) examined how postural adjustments are organized for
reach movements executed in multiple directions, and (2) investigated how
muscle activity for postural and movement related components of reaching

6
tasks are organized in relation to one another. This thesis was meant to go
beyond merely describing the role of feedforward postural control, but to
examine the spatial and temporal patterns of postural activity in order to make
supported inferences about how these postural adjustments may be organized
by the CNS.
1.4 Scientific Objectives and Hypotheses
Studies of postural control during voluntary movements in humans and
cats have provided evidence for the hypothesis that central commands for
maintaining balance interact with those for controlling movement (Massion
1992). Within this general framework, the main goal of this thesis is to explore
the organization of feedforward postural mechanisms in relation to voluntary
reaching movements performed in humans while standing. Specifically, this
thesis presents a series of experiments to (1) characterize the strategies adopted
for postural adjustments preceding and accompanying reaching movements in
multiple directions in standing (Specific Aim 1, SA1); (2) determine whether
muscle synergies explain the coordination of feedforward postural muscle
activity (SA2); and (3) explore the nature of postural control associated with
online corrections of reaching movements executed while standing (SA3). This
was achieved in Chapters 4, 5, and 6, respectively. Together, these studies
provide an important contribution to our knowledge about how the CNS
coordinates the control of posture and movement. To perform these
experiments, an experimental setup was designed to measure muscle activity,
kinetics and kinematics during whole-body reaching movements in multiple
directions, as detailed in Chapter 3. A description of the general objectives of
each study are presented below.
In Chapter 4, I quantify the strategies adopted by humans for
controlling posture during multidirectional reaching movements performed in
standing. I investigate whether predictive feedforward postural adjustments for
reaching are characterized by similar spatial organization of horizontal forces
exerted at the ground and muscle activity to those seen in reactive postural

7
control, despite their different modes of neural control. In Chapter 5, I extend
the findings of the previous chapter by identifying muscle synergies to explain
the spatial and temporal coordination of feedforward postural muscle activity.
Together, Chapters 4 and 5 provide the first characterization of feedforward
postural adjustments with the aim of drawing parallels with postural strategies
adopted for feedback postural control. Finally, in Chapter 6, I provide the first
examination of online postural control in relation to corrections in arm
movements resulting from visual perturbations of reaching while standing.
Specifically, I determine the mode of control for posture in relation to the signal
for the arm movement. Overall, the studies presented in this thesis further our
knowledge about feedforward postural control and contribute novel insight
about the coordination of posture and movement.

8

9
Chapter 2
Review of Literature
Imagine reaching forward to grasp an object located just beyond arm’s
length while standing. Most healthy people perform this skill with ease and
efficiency, many times a day, without any conscious thought to the underlying
control mechanisms. However, this seemingly simple behavior is complex, and
requires highly coordinated contributions between many structures distributed
throughout the CNS and musculoskeletal system that ensure the maintenance of
balance while performing the task. The underlying complexity in controlling
posture and movement arises from the multi-segmental structure of the body,
which affords the CNS significant redundancy and flexibility on the one hand,
but is also considered to be a computational challenge for the central controller.
Furthermore, the CNS must also consider how to accommodate the stability
requirements in the face of the disturbance of balance associated with the goal-
directed component of a movement. To simplify control, it is thought that the
CNS relies on neural strategies to transform task-level goals into appropriate
motor commands that specify appropriate muscle activation patterns (Ting and
Mckay 2007).
To establish a framework for understanding how the CNS coordinates
the control of posture and movement, the following review of literature will
first provide an overview of the underlying neurophysiology for the control of
movement and posture, with an emphasis on those circuits involved in the
coordination of movement and posture. Then, the biomechanical principles and
sensorimotor contributions governing the control of postural equilibrium and
orientation will be presented. Next, a discussion of the different modes of
postural control (feedback and feedforward) will follow. Finally, current
models of predictive motor control will be discussed.

10
2.1 How are voluntary movements and posture controlled?
The ability to move our body, limbs, head and eyes in order to
communicate, gesture and navigate our environment, while maintaining posture
and balance, is dependent upon the organization and complex interactions of
the motor and sensory systems (Ghez and Krakauer 2000). Successful planning
and execution of purposeful movement is largely facilitated by the hierarchical
organization of the sensorimotor systems distributed throughout the brain and
spinal cord. For example, the spinal cord contains the local circuitry that form
the building blocks used for both reflexive and goal-directed motor behaviours
(Rossignol et al. 2006). Higher levels of the CNS, such as areas of the
brainstem and motor cortex, provide descending modulation, which then
specify how the lower circuits are activated according to the task goals and
environmental context (Drew et al. 2004; McVea and Pearson 2009).
2.1.1 The neuroanatomical basis of movement execution
The production of movement involves four distinct, yet highly
interrelated neural subsystems. Each provides unique contributions to the
overall performance and control of movements. The four systems include the (i)
the motor neurons located in the spinal cord and brainstem, (ii) the motor
neurons of the brainstem and motor cortex, (iii) the cerebellum and (iv) the
basal ganglia (Purves et al. 2012).
The lowest level of the motor hierarchy involves the motor neurons
located in the spinal cord and brainstem. Their cell bodies are located in the
ventral horns of the grey matter of the spinal cord and the tegmentum of the
brainstem, and they synapse terminally on muscle fibers. These motor neurons
can influence the behaviour of their effector muscles directly via a pathway
involving its direct innervation of an alpha-motor neuron. Alternatively, these
motor neurons can serve as a relay for transmitting neural commands from
other motor neurons. As a result, the motor neurons of the spinal cord are
commonly referred to as the ‘final common pathway’ (Sherrington 1961). The
classification of motor neurons depends on the type of muscle fiber it

11
innervates. Alpha-motor neurons innervate extrafusal muscle fibers, which are
the skeletal fibers recruited for generating motion of the skeleton; and gamma-
motor neurons innervate intrafusal fibers, which are the contractile elements of
the muscle spindles (Pearson and Gordon 2000).
The second subsystem encompasses the motor neurons originating in
the motor areas of the frontal cortex and brainstem that subsequently synapse
on motor neurons and interneurons of the brainstem and spinal cord. These
motor neurons do not synapse directly with effector muscles, but rather shape
motor behaviour by modulating other motor neurons and the interneurons of the
spinal cord (Alstermark and Isa 2012; Ghez and Krakauer). These pathways are
important for the control of movement and posture. Notably, the corticospinal
tract is the primary descending pathway for planning and executing purposeful
movements (Ghez and Krakauer 2000). This tract may also influence posture
indirectly via collaterals that branch at different levels of the CNS (Kably and
Drew 1998; Massion 1992). Additionally, the reticulospinal tract has been
shown to have important contributions for the maintenance of balance (Drew et
al. 2002; Luccarini et al. 1990; Mori).
The cerebellum and the basal ganglia form the third and fourth
subsystems, respectively. Although these structures do not innervate muscle
fibers directly, they contribute to motor control via their modulation on the
activity of motor neurons. In particular, the cerebellum is thought to detect and
signal motor error (Desmurget and Grafton 2000), which is the difference
between the intended and actual movements. As such, the cerebellum is critical
for processes related to motor adaptation and long term learning (Bastian 2006).
The basal ganglia, which forms the fourth subsystem, is a collection of
structures located in the deep forebrain. The structures of the basal ganglia are
responsible for inhibiting unwanted movements and preparing the upper motor
neurons for the initiation of motor action (Mink 1996).
Together, these subsystems, along with the associated sensory systems,
form a complex hierarchical system that enables the performance of a vast
range of movements. For the purpose of the present discussion, only the

12
circuitry of the spinal cord, brainstem and motor cortex will be reviewed in
more detail.
2.1.2 Circuitry of the spinal cord provides a basis for coordinating
movement
Reductionist approaches to the study of motor behaviour provided
considerable insight into the circuitry of the spinal cord and how it contributes
to motor behaviour (Eccles and Sherrington 1930; Sherrington 1910). Using
reduced animal preparations involving the transection of the spinal cord or
brain at different levels (Sherrington 1909), or by deafferentation of a muscle of
interest, these studies identified many of the important components of the spinal
cord. Moreover, these studies have explained how the underlying circuitry of
the spinal cord forms the basis of coordinated motor behaviours, including
reflexes (Eccles and Sherrington 1930) and rhythmic behaviour such as
swimming (Grillner 2003) and locomotion (Rossignol et al. 2006).
A classically cited example to explain the spinal circuitry is the stretch
reflex (Pearson and Gordon 2000). The stretch (myotatic) reflex involves the
most basic elements of the spinal cord for producing a mechanical behaviour
without input from higher centers (Liddell and Sherrington 1924). For example,
when a muscle is passively stretched, this deformation is sensed by a muscle
spindle, which is a sensory receptor located within the muscle belly. Increases
in stretch in the muscle result in increased firing of the Ia-afferent sensory fiber,
which in turn excites the alpha-motor neuron of the muscle being stretched via
a direct synapse as well as the alpha-motor neurons of the synergistic muscles.
The Ia-afferent also inhibits the activity of the alpha-motor neuron innervating
the antagonist muscle. The net result of this activity is to resist the stretch on
the muscle by contracting the stretched muscle, thus generating force in that
muscle to cause flexion of the joint crossed by the involved musculature
(Pearson and Gordon 2000). This mechanism illustrates how complex motor
behaviour involving more than one muscle can be achieved by a simple
negative feedback loop mechanism.

13
The neural networks housed in the spinal cord are known as central
pattern generators (CPGs). They are capable of controlling the timing and
coordination of muscle activation patterns appropriate for the environment,
even if those environmental conditions change (Rossignol et al. 2006). CPGs
form the building blocks responsible for generating locomotor or rhythmic
behaviour in many species, including the lamprey (Grillner 2006), cats
(Rossignol et al. 2006), and frogs (Tresch et al. 1999). For example, in the
lamprey, swimming behaviour is achieved by coordinated activation and
inhibition of muscle segments along the length of the animal’s body to produce
sinusoidal motion (Grillner 2006). The neural mechanisms responsible for this
coordinated activity has been studied extensively (Grillner 2006).
The modular organization of the spinal cord is not limited to lower
vertebrates, but rather appears to be fundamental for controlling motion of the
skeleton in a variety of behaviours in several species (Tresch et al. 1999). For
example, several different types of behaviour can be elicited in a spinalized frog
by recruiting the same neural circuits (Cheung et al. 2005; Tresch et al. 1999).
Also, in a spinalized cat, locomotor behaviour can be elicited with the
appropriate sensory input and sufficient postural support to compensate for the
lack of postural tone (Rossignol et al. 2002). This suggests that higher neural
centres provide important modulatory control on the activity of the lower spinal
circuits. However, spinalized cats lack the capacity to maintain balance in
response to unexpected perturbations (Macpherson and Fung 1999). Whether
this is due to the trauma endured by the spinal cord or due to the lack of input
from higher centers remains to be determined (Honeycutt and Nichols 2010). In
support of this view, Schepens and Drew (2006) have proposed that the same
networks identified for the control of locomotion are also recruited in the
control posture.
2.1.3 Somatotopic organization of spinal cord
The spatial distribution of the motor neuron pools in the spinal cord has
been mapped by injecting different muscles with retrograde tracers that label

14
the cell bodies of the motor neurons that innervate that muscle (Levine et al.
2012). Subsequently, histological analyses were used to determine the specific
spatial mappings that exist along both the superior-inferior and medial-lateral
aspects of the spinal cord and brain stem.
Motor neuron pools innervating the upper limbs are found in the
cervical enlargement of the spinal cord, whereas the lower limbs are innervated
by motor neurons in the lumbar regions of the spinal cord (Ghez and Krakauer
2000). Furthermore, along the medial-lateral aspect, motor neuron pools are
organized such that the axial muscles, which are important for postural control,
are located in the medial and anterior regions of the ventral horn of a spinal
cord segment. Accordingly, motor neurons located more laterally within the
ventral horn innervate muscles placed more laterally in the body. Finally, the
most distal muscles of the body extremities, such as the digits, are innervated
by motor neurons that are the most laterally placed from the midline of the
spinal cord. Thus, an apparent functional grouping of musculature from axial,
proximal and distal can be observed as a result of the somatotopic map present
even at the lowest level of organization within the CNS (Levine et al. 2012).
The somatotopic organization of the local circuitry of the spinal cord
along the longitudinal axis also reflects functional differences of the networks
involved in controlling posture and those for skilled movement. In particular,
the local circuits in the medial regions of the intermediate zone synapse on the
motor neurons in the medial ventral horn. The axons involved in these circuits
have some projections that span multiple spinal segments enabling the
coordination of upper and lower limbs, while others synapse the length of the
spinal cord to assist in controlling posture. Some still cross the midline in the
commissure of the spinal cord providing a means for coordinating left and right
axial muscles (Ghez and Krakauer 2000). The local circuitry for the lateral
motor neurons, however, is increasingly differentiated with projections
contained mainly to the ipsilateral side of the spinal cord and spanning no more
than five spinal segments (Ghez and Krakauer 2000). This connectivity is
important for the highly differentiated control of distal muscles recruited for

15
skilled motor behaviour, such as independent control of the digits during
manipulation tasks (Alstermark and Isa 2012). The differences in the local
circuitry of the medial and lateral networks, as well as in the somatotopic
organization of the motor neurons in the ventral horn results in functional
differences in the control of muscles for either postural or goal-directed tasks.
The anatomical organization of the spinal circuits and their effector muscles
provide an anatomical substrate for understanding how the CNS might achieve
the complex coordination of many muscle groups across various parts of the
body.
2.1.4 Anatomical organization of the descending pathways for the control
of movement
The second subsystem consists of the upper motor neurons located in
the brainstem and cerebral cortex. These supraspinal networks provide
important modulation of the lower circuits in the brainstem and spinal cord
upon which they synapse via direct and indirect connections (McVea and
Pearson 2009). These descending pathways are divided into the lateral and
medial systems, according to differences in their spatial organization in the
spinal cord and functional connectivity (Drew et al. 2002). The lateral pathway,
which encompasses the lateral corticospinal tract and the rubrospinal tract, is
located laterally within the spinal cord and is primarily involved in movement
initiation and specifying the complex temporal coordination of muscle activity
for skilled voluntary movements (Ghez and Krakauer 2000). In contrast, the
medial pathways, which include the reticulospinal and vestibulospinal tracts,
have fairly vast innervation patterns of the axial and proximal musculature and
are critical for regulating posture (Drew et al. 2002) and orienting the head,
trunk with respect to vestibular, somatic, auditory and sensory information
(Ghez and Krakauer 2000).

16
2.1.4.1 Lateral descending pathway
The primary tract of the descending pathway is the corticospinal tract.
It, along with the corticobulbar tract, originate from the pyramidal cells of layer
5 of the motor cortex (Areas 4 and 6 of the frontal lobe) and terminate in the
spinal cord and brainstem nuclei, respectively. The corticospinal tract synapses
with motor neurons and interneurons of the brainstem and spinal cord, whereas
the corticobulbar tract projects to brainstem nuclei responsible for the control of
the cranial muscles.
As the corticospinal and corticobulbar tract descend the nervous system,
both have collaterals that branch at various levels to innervate the cranial
nuclei, the reticular formation, the red nucleus, and the pons. At the caudal level
of the medulla, only corticospinal tract (CST) axons remain. At this point,
approximately 90% of the CST axons cross the pyramidal decussation to form
the lateral CST on the contralateral side of the spinal cord and modulate the
lateral spinal motor neurons. The remaining 10% of CST axons continue to run
ipsilaterally or bilaterally, and form the ventral (anterior) CST, which is a part
of the ventral descending system. This portion of the CST originates in the
dorsal and medial regions of the motor cortex and terminates mainly on the
circuits serving axial and proximal musculature. Interestingly, this group of
CST neurons also give rise to the projections terminating in the reticular
formation, providing them with a privileged role in the control of posture.
Overall, the terminal distribution of the CST axons suggests functionally
different roles of the lateral and ventral divisions of the CST, whereby the
lateral CST provides important contributions to the fine control of the upper
extremities and the ventral CST for the control of posture (Ghez and Krakauer
2000; Purves et al. 2012).
The rubrospinal tract arises from the red nucleus in the midbrain and
immediately crosses to the contralateral side of the brain to descend via the
brainstem and innervate spinal motor neurons at several levels of the spinal
cord (Ghez and Krakauer 2000). Similar in function to the CST, the rubrospinal
tract is an indirect pathway for the control of voluntary movements. In

17
primates, however, much of the function of the rubrospinal tract has been
assumed by the CST (Ghez and Krakauer 2000).
2.1.4.2 Medial descending pathway
The diffuse action of the medial descending system is achieved by the
action of four descending pathways: the vestibulospinal tract, the tectospinal
tract and pontine (medial) reticulospinal tract and medullary (lateral)
reticulospinal tract. These pathways arise from various nuclei distributed
throughout the brainstem, including the vestibular nuclei, the tectum, the pons
and medulla, respectively, and project to the ventromedial regions of the spinal
cord gray matter. Together, they are critical for maintaining balance and
orienting the body and gaze (Drew et al. 2002).
For example, the vestibular nuclei give rise to the vestibulospinal
pathway, which has direct connections to the spinal cord, and is therefore able
to rapidly recruit the appropriate postural networks in response to a disturbance
of balance detected by the vestibular apparatus (McVea and Pearson 2009). The
tectospinal tract originates from the superior colliculus, which integrates visual
input received directly from the retina with somatosensory and auditory
information, and terminates in the upper segments of the spinal cord (Purves et
al. 2012). Consequently, it has been proposed that the tectospinal pathway may
have a role in the orientation of gaze. Finally, the reticular formation of the
brainstem forms a complex network of circuits originating in either the pons or
the medulla to form the pontine (medial) reticulospinal tract and the medullary
(lateral) reticulospinal tract. These two pathways have been shown to be
involved in a vast number of functions, including breathing, sleeping, posture
and locomotion (McVea and Pearson 2009).

18
2.1.4.3 Anatomical organization of reticular formation of the brainstem
relevant for postural control
Amidst the many functions of the reticular formation of the brainstem, it
has notable contributions in the control of posture (Drew et al. 2002), likely as a
result of its diffuse projections throughout the CNS. Although both the medial
and lateral reticulospinal tracts have been shown to be involved in organizing
the spatial and temporal coordination of the activation patterns of the trunk and
limb muscles (Drew et al. 2002), the medial pathway appears to have a more
significant role (Brustein and Rossignol 1998). In part, this can be explained by
differences in the connectivity of the two pathways. The medial brainstem
pathways terminate in the ventromedial areas of the spinal cord, providing
important modulatory effects on the axial and proximal musculature. These
pathways are vast in their projections, enabling the modulation of several
functionally related groups of motor nuclei (Drew et al. 2002). In contrast, the
lateral brainstem pathways project to the lateral areas of the spinal cord and are
consequently involved in the control of the distal muscles of the extremities
required for skills such as reaching and grasping (Ghez and Krakauer 2000).
In summary, knowledge of the basic neuroanatomical pathways related
to the initiation and execution of movement and the control of posture is crucial
for understanding how the CNS coordinates the postural and focal demands of a
task. Notably, the primary descending pathway, the corticospinal tract, has
collaterals that branch to the vestibular and reticular nuclei of the brainstem
prior to the decussation (Kably and Drew 1998). These areas are known to have
privileged involvement in controlling the body’s posture and equilibrium (Drew
et al. 2004). Therefore, it is plausible that these circuits provide the basis for the
neural computing required for integrating these two dichotomous aspects of
motor behaviour.

19
2.1.5 Integration of central commands for the global planning of movement
and posture
Cortically-initiated voluntary movements are preceded and accompanied
by postural adjustments that ensure balance is maintained (Massion 1992).
Questions related to the mechanisms through which these two aspects of motor
behaviour are integrated, and the localization of the networks responsible for
integrating posture and movement have received considerable attention in the
literature (Massion 1992). In the following paragraphs, the modes of control for
coordinating posture and movement will be explored, followed by a discussion
of the localization of the neural substrates involved in this coordination.
2.1.5.1 Modes of control for the integration of posture and movement
In consideration of several studies in humans and animals, Massion
(1992) proposed a classical theoretical model to explain how posture and
movement commands may be organized in the CNS. In this framework, two
possible modes of control have been identified (see Fig 16, (Schepens and
Drew 2004).
In the first, posture and movement are controlled via parallel pathways.
Here, it is postulated that posture and movement are controlled independently;
with separate descending neural commands that target either the postural
network or the movement related circuits. The feedforward postural
adjustments preceding movements are typically attributed to this mode of
control, such as during arm (Bouisset and Zattara 1981; 1987) or leg
movements (Mouchnino et al. 1992) in humans and reaching movements in cats
(Schepens and Drew 2003).
In general, temporal decoupling of the signal for the postural adjustment
and the movement is cited as evidence that these two commands are controlled
independently (Massion 1992). For example, the latency of the postural
adjustment preceding an arm movement is modulated as a function of the time
constraints imposed by the task. When the movement is self-paced, postural
activity occurs prior to the movement whereas in a reaction time paradigm,

20
postural and focal activities occur simultaneously (Lee et al. 1987).
Furthermore, the latency of the feedforward postural adjustment prior to the
movement onset increases as the load to be lifted increases (Bouisset and
Zattara 1981). It is hypothesized that the movement command is inhibited until
the postural system reaches a desired reference point optimal for the movement
execution (Cordo and Nashner 1982; Massion 1992). However, the mechanisms
of this inhibition and the nature of the command signals remain largely
unknown.
Alternatively, posture and movement commands may be controlled
hierarchically by a shared descending command that modulates the postural
networks via collateral branches (Gahery and Massion 1981; Gahery and
Nieoullon 1978). This mode of control appears to be used for organizing the
feedforward postural adjustments accompanying the movement (Massion
1992), such as during a bimanual load-lifting task (Paulignan et al. 1989). For
example, when the onset of the postural adjustment is time-locked to the focal
movement, it suggests that posture and movement commands are controlled
together within a hierarchical organization (Gahery and Nieoullon 1978;
Paulignan et al. 1989).
The theoretical model proposed by Massion (1992) has been
substantiated and extended by a series of behavioural and electrophysiological
studies examining the nature of the control signals for posture and movement
during reaching movements in the cat (Schepens and Drew 2006; 2004; 2003;
Yakovenko and Drew 2009). These findings will be discussed in detail in the
next section, but overall, these studies provide evidence that both hierarchical
and parallel modes of control are used by the CNS. Which mode of control is
adopted, however, may depend mainly on the role of the postural adjustment
and the nature of the voluntary task (Massion 1992).
2.1.5.2 Central organization of feedforward postural adjustments
Beyond the neuroanatomical evidence that points to the involvement of
several cortical and subcortical structures in coordinating posture and

21
movement, evidence from lesion, microstimulation and single-unit recording
studies have elucidated many of the central pathways responsible for the
coordination of posture and movement. There is general agreement that the
supraspinal regions of the CNS are involved in planning and initiating
feedforward postural adjustments (Gahery and Nieoullon 1978; Horak and
Macpherson 1996; Mackinnon et al. 2007; Massion 1992; Viallet et al. 1992;
Yakovenko and Drew 2009), whereas the brainstem is thought to be vital for
ensuring the appropriate scaling of the postural adjustments in relation to the
movement by integrating information from several cortical and subcortical
structures (Drew et al. 2004). Finally, it has been hypothesized that the
descending commands recruit the same CPG networks used in locomotion for
coordinating the muscle activity of the postural response (Honeycutt et al.
2009; Schepens and Drew 2006).
Evidence that these postural adjustments have contributions from
descending pathways arising from the cortex was demonstrated by Gahery and
Nieoullon (1978), who performed microstimulation of the motor cortex in the
cat to induce flexion movements of a limb. They found that the induced flexion
movement, whether it was of the fore- or hindlimb, was associated with a
decrease in the loading force under the limb diagonal to the flexed limb and a
concurrent increase in the loading forces of the other two limbs (termed
‘diagonal strategy’). Analysis of the latencies of the force changes revealed that
the postural adjustments for hindlimb flexion clearly preceded the limb flexion,
whereas those for forelimb flexion occurred concurrently or just after the
loading change in the moving limb. These data were interpreted to indicate that
the postural adjustments accompanying the flexion movements are centrally
driven and not the result of feedback driven EMG from peripheral afferents,
although feedback from the periphery may be used at a later stage of the
postural adjustment (Gahery and Nieoullon 1978). Moreover, it was
hypothesized that the pathways for the movement command send collaterals to
the brain stem or spinal cord to modulate the lower level networks responsible
for postural control (Gahery and Massion 1981; Massion 1992).

22
A cortical contribution to the control of posture during intentional
movement is also supported by studies in humans (Mackinnon et al. 2007;
Viallet et al. 1992). Using a bimanual load-lifting task (Dufosse et al. 1985;
Hugon et al. 1982; Paulignan et al. 1989) in patients with lesions in the medial
frontal areas, Viallet and colleagues (1992) demonstrated that both hemispheres
of the cortex are required for producing appropriate postural adjustments during
voluntary movements to maintain the stability of the limb. In this task, a load is
placed on the subject’s forearm and they are required to remove it by
voluntarily lifting the load with the other arm. In control subjects, the position
of the loaded arm remains fairly stable, even as the load is removed. Analysis
of the muscle activity revealed anticipatory inhibition of the loaded forearm’s
flexors that was time locked to the voluntary contraction of the muscles of the
moving segment (Paulignan et al. 1989; Viallet et al. 1992). However,
hemiparetic patients lacked appropriate anticipatory postural adjustments
(APAs) in the arm contralateral to the lesion (Viallet et al. 1992). The authors
suggested that the supplementary motor areas and associated motor cortical
areas contralateral to the loaded arm are necessary for organizing the phasic
anticipatory postural adjustment, whereas the cortical motor areas contralateral
to the moving limb recruit the networks for the focal muscle activity and, via
collaterals, the networks for the postural adjustments accompanying the
movement.
Also, Mackinnon and colleagues (2007) demonstrated that the
amplitude of both the postural activity preceding step initiation and the focal
muscle activity for stepping were increased in a time-locked manner as a result
of TMS applied to the leg area of the motor cortex in humans. These results
further suggest that the commands for posture and movement may be shared.
Finally, evidence from neural recordings in the motor cortex during
reaching movements in the cat have shown a specific contribution of the
descending tracts from the motor cortex to the postural adjustments preceding
movement onset (Yakovenko and Drew 2009). Here, cats performed reaching
movements in an instructed delay task while standing. The discharge of several

23
pyramidal tract neurons was shown to be time-locked to the cue for the ‘GO’
signal and not to the onset of the goal-directed component of the reaching
movement. Furthermore, the activation latency of these neurons was correlated
to the onset of the preparatory postural activity, indicated by a change in
vertical pressure under the supporting limbs. Finally, the discharge
characteristics of the recorded cortical neurons were identical to those
previously recorded in the pontomedullar reticular formation (PMRF)
(Schepens and Drew 2006; 2004; Schepens et al. 2008), a structure known to
have important functions in postural control (Drew et al. 2004) rather than in
movement planning. These findings are consistent with previous lesion and
stimulation studies (Perfiliev 2005) that have suggested that the motor cortex
contributes to the descending signal for anticipatory postural adjustments, but
suggests that it’s contributions are not involved in the global planning of
posture and movement processes (Yakovenko and Drew 2009).
Beyond the cortex, the brainstem reticular formation has been attributed
a critical role in the integration of neural commands for posture and movement
(Schepens and Drew 2003; Schepens et al. 2008). Lesion studies in cats and
primates have shown that damage to the reticulospinal and vestibulospinal
tracts result in deficits in balance control whereas fine control of the distal arm
is maintained (Lawrence and Kuypers 1968). Also, inactivation of the pontine
reticular formation by a cholinergic agonist during cortical stimulations to elicit
feedforward postural adjustments result in deficits in the postural response
(Luccarini et al. 1990). Finally, selective lesions of the medial reticular tract in
cats results in deficits in posture and weight support during locomotion, where
the magnitude of the postural deficit is related to the severity of the lesion
(Brustein and Rossignol 1998).
The importance of the brainstem to the control of balance is supported
by studies of reactive postural control. For example, decerebrate cats are
capable of restoring balance following unexpected perturbations of balance
with similar strategies to those observed in intact animals (Honeycutt et al.
2009; Honeycutt et al. 2012; Honeycutt and Nichols 2010), suggesting that the

24
primary networks for postural control are located at lower levels of the motor
hierarchy. However, for postural responses to be appropriately scaled in time
and magnitude to the parameters of the perturbation, descending input from the
cortex is required (Drew et al. 2004). For example, cats with lesions to the
spinal cord are able to restore balance following a perturbation, but lack the
precise spatial organization of muscle activity in several muscles that is
typically observed in controls (Macpherson and Fung 1999), lending support to
the view that the brainstem has a privileged role in equilibrium. However, some
authors have argued that the deficits in postural control observed in spinalized
animals may be due to the trauma of the lesion and subsequent reorganization
of signaling pathways rather than due to the loss of descending input from the
brainstem or cortical areas (Honeycutt and Nichols 2010).
Recently, a series of electrophysiological studies recorded neurons in
the brainstem reticular formation during reaching movements and unexpected
perturbations in cats (Schepens and Drew 2006; 2004; 2003; Schepens et al.
2008; Stapley and Drew 2009). These have provided considerable insight about
how and where commands for posture and movement are controlled within the
CNS. Specifically, three populations of cells in the PMRF were identified that
were related to either the postural adjustment preceding the movement,
accompanying the movement, or the movement itself (Schepens and Drew
2004; Schepens et al. 2008). Furthermore, discharge of PMRF neurons occurred
for reaches performed with either limb, although was increased for reaches
made with the limb ipsilateral to the recording site (Schepens and Drew 2006).
Quantitative analyses in the form of regressions and spike triggered averaging
(STA) revealed that the bilateral signal is asymmetric in nature and the final
descending signal is modified, or gated, depending on which limb performs the
reaching movement (Schepens and Drew 2006). Interestingly, changes in the
discharge of the neurons of the PMRF were also observed when the support
surface under one of the limbs was unexpectedly removed (Stapley and Drew
2009). Together, these studies demonstrate that the PMRF contributes
significantly to the coordination of posture during both goal-directed

25
movements and unexpected perturbations of the limb. These findings suggest
that the PMRF gates neural signals for both feedback and feedforward signals
and is the site of integration of postural control organized via these two
mechanisms.
2.2 Postural Control
2.2.1 Biomechanical requirements for equilibrium control
Humans are faced with a unique set of equilibrium constraints as a
result of their upright, bipedal stance. In particular, the CNS must deal with a
relatively high position of the center of mass (CoM) within a narrow base of
support (BoS), which consequently affords a limited region of stability (Hayes
1982). To maintain stability and equilibrium during quiet standing and while
executing complex multi-joint movements, the human nervous system must
balance all external and internal forces acting on the body (Horak and
Macpherson 1996; Massion 1992). The forces acting on the body include
external constraints associated with the environment and task, such as the force
of gravity, the reaction forces from the supporting surfaces, and any imposed
accelerations or obstacles in the environment (Massion 1992). In addition,
internal constraints refer to those constraints that arise within the body, such as
the geometry and inertial characteristics of the body segments, and the internal
forces associated with muscle contractions (Massion 1992). For the skillful
execution of movement, the CNS must choose an optimal neural strategy in
consideration of the task and biomechanical, and musculoskeletal constraints
that optimally balances all these forces in order to meet the task goals (Horak
and Macpherson 1996; Massion 1992).
Equilibrium is achieved by maintaining the position of the body’s
CoM, defined as the weighted sum of the body’s segments, within a relatively
small region of stability afforded by the supporting limbs, known as the BoS
(Winter 2009). The tight regulation of the CoM for balance control is illustrated
by the phenomenon of body sway that occurs naturally in quiet stance. In quiet

26
upright stance, the control of balance can be modeled approximately as an
inverted pendulum (Pai and Patton 1997; Winter et al. 1998). The net
acceleration on the body is determined by the moment of force resulting from
the perpendicular difference between line of gravity through the CoM and the
ground reaction force (GRF). The position of the centre of pressure (CoP;
defined as the point of application of the GRF (Horak and Macpherson 1996;
Winter 2009)) is actively modulated by the ankle musculature (Morasso and
Schieppati 1999), which in turn, results in adjustments of the distance between
the line of gravity through the CoM and the GRF (van Ingen Schenau et al.
1992). The perpendicular difference between these two resultant force vectors
creates a moment of force that causes a net acceleration of the whole body
about the ankle joint (Winter 2009). In summary, the CNS actively modulates
the point of application of the external GRF, which corresponds to the CoP, to
achieve the appropriate resultant moment of force that will cause the desired
whole body acceleration. In addition to sway, this control mechanism has been
demonstrated for a number of dynamic tasks, including cycling (van Ingen
Schenau et al. 1992), whole body lifting (Commissaris et al. 2001; Toussaint et
al. 1995), and locomotion (Lepers and Brenière 1995).
2.2.2 Behavioural goals of the postural system
The act of maintaining balance both during quiet stance, and when
transitioning from one body configuration to another during voluntary
movements, engages several systems distributed throughout the CNS. Together,
these processes serve the goals of the postural system. In the context of goal-
directed motor behaviour, the postural system provides an essential foundation
for motor coordination by controlling the position of the CoM, thus providing
stability for transitioning between body configurations and orienting the body
segments with respect to each other, the environment or both (Horak and
Macpherson 1996).
Broadly, the behavioural goals of the postural system can be categorized
as being involved either in specifying the orientation or in maintaining

27
equilibrium of the body and its segments. Specifically, postural orientation
refers to those processes that maintain the alignment of the body segments with
respect to themselves and to the environment (Horak and Macpherson 1996;
Massion and Woollacott 2004). For any given postural configuration, postural
variables such as the alignment of the head, trunk, and the geometry of the
limbs are stabilized according to the task and context (Horak and Macpherson
1996). In contrast, postural equilibrium ensures that all forces acting on the
body are balanced so that the body remains balanced or moves in a controlled
fashion (Horak and Macpherson 1996). This is achieved with coordinated
postural strategies that respond to disturbances of balance resulting from either
internal or external perturbations.
2.2.3 Sensorimotor control of posture
Feedback from a number of sensory modalities is utilized by the CNS to
create a representation of the body in relation to its external environment
(Horak and Macpherson 1996; Mergner and Rosemeier 1998). In particular, the
visual, vestibular and somatosensory, which includes muscle proprioception,
joint and cutaneous afferents, systems each detects an ‘error’ signal relating the
amount of deviation of the body orientation from a reference point (Peterka
2002). These signals are then thought to be integrated and weighted according
to the task, context and availability of sensory information (Horak et al. 1990).
The integrated sensory input is subsequently used to calculate the orientation of
the head, trunk and limbs in space, providing an internal representation of the
body and its surroundings (Horak and Macpherson 1996; Mergner and
Rosemeier 1998).
The relative contributions of the different sensory modalities to the
overall control of postural orientation and equilibrium have been investigated
experimentally with a number of paradigms. Typically, these paradigms involve
systematically manipulating each of the modalities and measuring the effects on
balance control in quiet stance (Diener et al. 1984b; Nashner et al. 1982), in
response to unexpected perturbations of balance (Horak et al. 1990) or during

28
voluntary movements (Macpherson et al. 2007). Overall, these studies have
shown that each sensory modality is unique and responds optimally for a given
behaviour (Horak, 1996 (Horak and Macpherson 1996; Horak et al. 1990;
Mergner and Rosemeier 1998).
For example, when pressure cuffs are applied to the ankles to limit the
input from somatosensory afferents arising from the feet, little effect on
postural sway is observed when vision is available. However, when subjects are
asked to close their eyes, increased sway at the hips is apparent (Diener et al.
1984b). Similarly, patients with vestibular loss are able to compensate for their
sensory loss when visual input is available. However, when vision and
somatosensory inputs are inaccurate, these patients experience a loss of
equilibrium (Nashner et al. 1982). In general, these studies suggest that
somatosensory and vestibular information have critical roles in controlling
balance during upright stance (Horak et al. 1990).
While the study of sway during quiet stance has provided important
insight into the sensorimotor processes of postural control, this approach is
limited in its ability to specify the mechanisms for dynamic postural control,
such as in response to an unexpected loss of balance or during the execution of
goal-directed movements while standing. Accordingly, investigators have
examined how sensory processes shape postural behaviour by quantifying the
postural responses to dynamic perturbations of balance when one or more
sensory inputs is disrupted in cats (Inglis and Macpherson 1995; Macpherson et
al. 2007; Stapley et al. 2002; Stapley et al. 2006) and humans (Horak et al.
1990). Overall, these studies have shown that not only is the detection of the
postural disturbance affected by sensory loss, but so is the selection and
organization of an appropriate postural strategy (Horak et al. 1990).
In particular, studies in cats have provided important insight about the
relative contributions of vestibular and somatosensory inputs. In one such
study, Inglis and Macpherson (1995) subjected cats to support surface
translations in the horizontal plane before and after bilateral labyrinthectomy.
Following the lesion, the postural strategies were similar in spatial and temporal

29
organization to the control responses even when executed in complete darkness.
However, increases in the amplitude of the responses were observed. These
results suggest that vestibular information is not critical for the organization of
the postural response, but rather influences its scaling. Somatosensory
information, however, appears to have a more important role in shaping the
temporal and spatial characteristics of the postural response (Maurer et al. 2001;
Stapley et al. 2002; Ting and Macpherson 2004).
The role of somatosensory information in shaping the postural responses
to unexpected perturbations of balance was examined specifically in cats given
large doses of pyridoxine to induce selective large-fiber deafferentation
(Stapley et al. 2002). The results of this study provide evidence that the large
afferent fibers carrying somatosensory information from the periphery are
responsible for the correct timing of the coordinated postural strategies that
restore equilibrium following an unexpected perturbation. Furthermore, Ting
and Macpherson (2004) demonstrated that somatosensory information is critical
in shaping the postural response by comparing postural responses to multi-
directional pitch/roll rotations and linear translations in the horizontal plane of
the support surface in freely standing cats. They found that the only reliable
parameter of the perturbation in predicting the organization of the postural
response was the ratio of the vertical loading force and horizontal slip force
components of the GRF. This signal presumably reflects the horizontal motion
of the CoM. Based on their findings, the authors proposed that cutaneous
sensors in the feet detect the change in force angle at the ground and
subsequently provide critical information regarding the nature of the
perturbation necessary for organizing the postural response (Ting and
Macpherson 2004). The importance of somatosensory cues from the feet for
controlling balance has also been demonstrated for humans (Maurer et al.
2001).
In summary, the postural control system encompasses all neural,
muscular and sensory processes related to maintaining the equilibrium and
orientation of the body (Horak and Macpherson 1996). The sensory modalities

30
are critical for providing cues about the orientation of the limbs and body in
relation to a reference point. The inputs are thought to be summed (Peterka
2002) and weighted according to the task and context (Jeka et al. 2006;
Mergner and Rosemeier 1998). In the event of the loss or disruption of a
sensory modality, redundancy across the sensory systems, as well as the ability
to re-weight the incoming sensory input, ensure that at least partial recovery of
a representation of the body schema is possible (Black et al. 1989).
2.2.4 The problem of motor redundancy
2.2.4.1 Degree of Freedom Problem
To control balance and posture while standing, the CNS must coordinate
the activity of many muscles crossing a large number of joints capable of
moving in multiple planes (Bunderson et al. 2010). As a result, there is
significant redundancy in the motor system for defining a unique pattern of
joint motion (Kuo 2005; Todorov 2004; Yang and Pai 2007) and muscle
activation patterns (Gottlieb 1998; Lockhart and Ting 2007) for a given motor
task. This classical problem of motor control was first formalized by Bernstein
(1967) and labeled the degree of freedom (DoF) problem (Bernstein 1967).
Faced with this redundancy, it is thought that the CNS must compute an
appropriate set of motor commands to generate muscle activity that is
appropriate for the task and environmental constraints, all while considering the
complex kinematic chains of the musculoskeletal system and the inertial
properties of the body’s segments (Chiel et al. 2009). How the CNS maps task-
level goals into coordinated patterns of muscle activity remains a topic of
considerable debate amongst researchers (Latash 2012).
A widely accepted hypothesis is that the CNS is organized to control
several functionally related DoF together, as modules (Bernstein 1967;
Krishnamoorthy et al. 2003; Krishnamoorthy et al. 2004; Ting 2007; Ting and
Mckay 2007). Within this framework, higher centers of the CNS serve to
constrain global parameters for movement, such as task-level goals, whereas

31
lower levels of the hierarchy specify the coordinated spatial and temporal
patterns of the muscle activations (Ting 2007; Ting and Mckay 2007). This
hierarchical organization is thought to decrease the complexity of control by
diminishing the dimensionality of the control problem (McKay et al. 2007;
McKay and Ting 2008), while providing an attractive mechanism to explain
how the CNS maps task-level goals into appropriate muscle activation patterns
(Chvatal et al. 2011; Ting 2007).
Support for a hierarchical organization of the motor system is evident
from several psychophysical, biomechanical and neurophysiological studies
that have demonstrated that task-level variables are often constrained with
greater rigidity than lower level goals (Chiel et al. 2009; Ting 2007). For
example, in a reaching task, end-point trajectory of the finger is more tightly
regulated than the angular displacements of the individual joint angles.
Furthermore, task level parameters, such as direction, force and velocity of the
hand during reaching, are encoded in the motor cortex in primates
(Georgopoulos 1986). Similarly, in human locomotion, changes in joint angles
are correlated, suggesting that the CNS constrains several joints together to
control CoM motion (Bernstein 1967; Cavagna et al. 1977). Together, these
findings provide support for the hypothesis that the CNS simplifies the control
of redundant systems by grouping DoFs together as ‘synergies’ or ‘modules’
and serve as a mechanisms to control task level goals (Chvatal et al. 2011;
McKay and Ting 2008; Ting 2007; Ting and Mckay 2007; Yakovenko et al.
2011). However, it remains to be determined at what level of the motor system
this redundancy is resolved.
2.2.4.2 Muscle synergies simplify the coordination of muscle activity
Recently, this model of hierarchical control has been developed further
in the context of muscle coordination. It is proposed that the CNS solves the
problem of redundancy at the level of the muscles through the flexible
recruitment of muscle synergies (Lockhart and Ting 2007; Ting 2007; Ting and
Mckay 2007). A muscle synergy is defined as invariant patterns of muscle

32
activation with fixed spatial scaling (Ting 2007; Ting and Macpherson 2005). It
is postulated that the nervous system specifies a neural command that
determines the temporal recruitment of a muscle synergy.
Within this framework, the overall muscle activity is expected to reflect
the linear sum of each of these synergies multiplied by their respective
activation coefficient, which are the purported neural commands (Ting 2007).
Mathematically, these synergies can be identified with factorization techniques
(Tresch et al. 2006), such as non-negative matrix factorization (NNMF) (Lee
and Seung 1999; Ting and Macpherson 2005; Tresch et al. 2006), principal
component analysis (PCA) (Krishnamoorthy et al. 2003; Krishnamoorthy et al.
2004) or independent component analysis (ICA) (Hart and Giszter 2004).
This modular organization of muscle activity has been demonstrated for
several motor behaviours in many species (Cappellini et al. 2006; d'Avella et al.
2006; Drew et al. 2008; Flash and Hochner 2005; Hart and Giszter 2004a; Hart
and Giszter 2004b; Krishnamoorthy et al. 2004; Krouchev et al. 2006; Latash et
al. 2005; Ting and Macpherson 2005; Torres-Oviedo et al. 2006; Torres-Oviedo
and Ting 2007; Yakovenko et al. 2011). Furthermore, the muscle synergies
extracted for one task frequently generalize and accurately reconstruct the
muscle activity patterns in another task, as has been shown for frog scratching,
jumping and locomotion (Cheung et al. 2005), human locomotion and running
(Cappellini et al. 2006) and forwards and backwards cycling motions (Ting et
al. 1999). Typically, a group of synergies is shared across tasks, although some
task-specific synergies may emerge according to the task (Chvatal et al. 2011;
Torres-Oviedo and Ting 2010). Also, the overall recruitment of the synergies
may change (Safavynia et al. 2011).
2.3 Mechanisms of postural control
2.3.1 Overview
Humans encounter numerous predictable and unpredictable
perturbations of balance during their everyday lives. In order to maintain

33
balance in these situations, the CNS relies on feedback and feedforward control
mechanisms to counteract any internal or external forces that may potentially
destabilize the body (Horak and Macpherson 1996; Massion 1992). In the case
of an externally driven perturbation of balance, such as an unexpected slip
when walking on a slippery surface, the CNS relies mainly on feedback
mechanisms to restore balance. In these situations, sensory input from a number
of sensors that signal the direction and amplitude of the perturbation is
integrated by the CNS to shape an appropriate automatic postural response that
effectively accelerates the position of the CoM to a position of stability within
the BoS (Henry et al. 2001; Horak and Macpherson 1996; Macpherson 1988a;
b; Ting and Macpherson 2004). In contrast, voluntary movements are
associated with disturbances to equilibrium due to internal forces related to the
muscle action for limb displacements. Here, the CNS is able to predict the
destabilizing effects of the interaction torques associated with the goal-directed
displacement of limb segments and programs appropriate postural adjustments
in anticipation of the disturbance (Bouisset and Zattara 1981; 1987; Massion
1992). These feedforward postural adjustments are commonly referred to as
'anticipatory postural adjustments' (APAs) and are programmed in feedforward
to compensate for the internally derived disturbance of equilibrium (Bouisset
and Zattara 1981; 1987; Massion 1992).
2.3.2 Intrinsic mechanical properties for stability
Several biomechanical and neural mechanisms are involved in
maintaining a desired postural configuration and equilibrium of the body. The
passive properties of the muscles and joints and the surrounding connective
tissues, as well as tonic muscle activity provide substantial contributions to
postural control (Horak and Macpherson 1996). For example, in upright quiet
stance, the alignment of the limb segments causes the primary muscles for
postural control, such as soleus and illiopsoas, to be stretched beyond their
resting length causing them to be tonically activated (Joseph and Nightingale
1952). When muscles are tonically active, there is an increase in the steepness

34
and decrease in the threshold of their length-tension curve enabling the muscle
to respond almost instantaneously against a disturbing force before any
peripheral or centrally-driven signals are available (Horak and Macpherson
1996). The passive properties of the muscles and their tonic activity provide
important load-compensating mechanisms not only for the maintenance of
posture, but also for providing stability during the execution of movement.
However, for larger, dynamic perturbations of balance, the viscoelastic
properties of muscles, ligaments and tendons, and muscle stretch alone are not
sufficient for restoring balance (Diener et al. 1984a). Thus, additional active
central mechanisms must be recruited for controlling equilibrium and
orientation when faced with unexpected and expected disturbances of balance.
These will be explored next.
2.3.3 Feedback postural responses
While standing on a moving bus, a person may experience a loss of
balance due to an unexpected acceleration or deceleration of the bus. In this
situation, the CNS responds quickly with stereotypical muscle activity in the
supporting limbs to oppose the disturbing force and restore equilibrium.
Postural responses of this nature rely on feedback from a weighted summation
(Mergner and Rosemeier 1998) of the sensory input from the available sensory
modalities, including visual, vestibular, proprioceptive, and somatosensory
systems, to detect the nature of the perturbation (Horak and Macpherson 1996).
These APR have been studied extensively in the laboratory setting and have
provided considerable information about the underlying neural control of
posture (Carpenter et al. 1999; Horak and Nashner 1986; Macpherson 1988a; b;
Nashner 1976; 1977; Torres-Oviedo and Ting 2007; 2010).
A spectrum of postural strategies for restoring balance following a
perturbation have been described in terms of the kinematic, kinetic and
electromyographic (EMG) patterns of the response (Do et al. 1982; Horak and
Macpherson 1996; Horak and Nashner 1986; Maki and McIlroy 1997; McIlroy
and Maki 1993a). For example, an unexpected backward translation of the

35
support surface induces a forward fall of the body. To restore equilibrium, the
CNS sequentially activates the extensor muscles of the body, namely, the
soleus, gastrocnemius, biceps femoris, the paraspinals and muscles of the neck,
to create the necessary GRFs that oppose the forward acceleration of the CoM
and reestablish an upright configuration of the body (Horak and Nashner 1986).
When the restoring torques are mainly focused at the ankle joint, the strategy is
known as an ankle strategy (Horak and Macpherson 1996; Horak and Nashner
1986). Alternatively, a hip strategy may be used to restore balance when the
perturbation is rapid, large in amplitude, or if the support surface is short
relative to the foot length (Horak and Nashner 1986). In this case, rotation at
the hips is accompanied by counter rotation of the ankle and neck to restore a
stable position of the CoM with respect to the BoS. In reality, however, most
foot-in-place postural responses typically involve a mixed strategy with
contributions from both the ankle and hip strategies (Horak and Macpherson
1996; Horak and Nashner 1986). Furthermore, when subjects are not
constrained to maintain their feet in place, an alternative strategy for regaining
balance may be to increase the BoS by taking a step (Do et al. 1982; Do et al.
1999; McIlroy and Maki 1993a; b) or using their hand to hold a support, such
as a handrail or wall (Maki and McIlroy 1997). The observation of these
different strategies led to several questions related to the nature of these
postural strategies: Are these complex movement patterns central programs?
How does the CNS select a particular strategy? Do these strategies reflect a
means by which the CNS simplifies the control of movement?
To gain insight about how the CNS organizes postural responses,
experimental paradigms have been designed where the support surface upon
which a subject is standing is unexpectedly translated or rotated to simulate a
slip. These perturbations elicit stereotypical activity in the muscles of the
supporting limb, typically evoked at latencies of 70-100 ms in humans (Diener
et al. 1988; Horak and Macpherson 1996; Horak and Nashner 1986; Nashner
1977) and 40-60 ms in cats (Horak and Macpherson 1996; Rushmer et al. 1988;
Ting and Macpherson 2004) following the onset of the disturbance. Using

36
perturbations in multiple directions of varying amplitudes and speeds, it has
been shown that the selection of a particular strategy, and the characteristics of
the APR, are shaped by the biomechanics (direction and velocity) of the
disturbance (Diener et al. 1984a; Macpherson 1988b; Ting and Macpherson
2004), as well as the initial body position (Horak and Moore 1993), prior
experience of disturbances (Macpherson 1994) and ‘central set’ (Horak et al.
1989a), which refers to the modulation of a response by descending commands
related to the expectation of what the perturbation and task parameters will be
(Brooks 1984; Evarts 1975; Schmidt 1982).
In response to unexpected perturbations of the support surface, both
humans and cats respond with muscle activity in the supporting limbs that is
broadly tuned to the direction of translation, with maximal amplitudes of
activation occurring for a single direction of translation (Henry et al. 2001;
1998b; Macpherson 1988b; Rushmer et al. 1988) or rotation (Carpenter et al.
1999; 2001; Ting and Macpherson 2004). While most muscles are characterized
by a unique curve, some similarities in the shape and tuning of multiple
muscles can be observed (e.g. (Henry et al. 1998b; Macpherson 1988b; Torres-
Oviedo and Ting 2007), suggesting that these muscles may be controlled as a
unit. Also, muscle activity is broadly tuned and shows some variability within a
reaching direction, indicating there is not a simple mapping between
perturbation direction and muscle activity (Moore et al. 1988). Together, these
findings suggest that the CNS may rely on a synergic organization to simplify
the task of selecting muscle activity patterns appropriate for a perturbation.
Recently, the muscle activation patterns for feedback postural control in
cats and humans have been examined in detail using factorization techniques
(Ting and Macpherson 2005; Torres-Oviedo et al. 2006; Torres-Oviedo and
Ting 2007; 2010). These studies have shown that a small number of spatially-
fixed muscle synergies are sufficient to reconstruct the trial-by-trial variability
of the complex spatial and temporal patterns underlying the postural responses
to perturbations in multiple directions (Ting and Macpherson 2005; Torres-
Oviedo et al. 2006; Torres-Oviedo and Ting 2007). Examination of the

37
relationship between muscle synergy recruitment and GRF patterns has also
shown that muscle synergies produce consistent end-point force patterns
(Krishnamoorthy et al. 2003; Ting and Macpherson 2005; Torres-Oviedo et al.
2006) and are stable in a variety of biomechanical contexts (Torres-Oviedo and
Ting 2010). More recently, shared muscle synergies have been shown for force
control in both non-stepping and stepping reactive postural responses (Chvatal
et al. 2011). Together, these results support the idea that muscle synergies
represent a neural mechanism for translating task-level goal into appropriate
muscle activation patterns for the task (Ting and Mckay 2007).
Characterization of the corresponding GRF for restoring equilibrium has
revealed that both cats and humans only produce force at the ground in one of
two directions aligned with the diagonal irrespective of perturbation direction
(Henry et al. 1998a; 2001; Macpherson 1988a). This strategy has been termed
the force constraint strategy (Macpherson 1988a) and is independent of prior
experience (Macpherson 1994), but is modulated by stance width (Henry et al.
2001; Macpherson 1994a). Specifically, net GRFs appropriate for opposing a
perturbation are achieved by modulating the amplitude, and not the direction, of
the force vector under each limb (Macpherson 1988a). In general, the force
constraint strategy is thought to simplify the control of posture by reducing the
number of parameters that the CNS must control (Macpherson 1988a).
2.3.4 Feedforward postural adjustments
Successful execution of voluntary movement during standing requires
postural activity to stabilize the disturbance of the CoM and compensate for the
disturbing interaction torques associated with the displacement of the focal limb
segments (Massion 1992). These adjustments of posture are termed
‘anticipatory postural adjustments’ (APA) since the postural changes occur
prior to, or at the same time, as the onset of the internal disturbance to posture
due to a voluntary movement and are programmed with a feedforward central
command (Massion 1992). It is traditionally believed that these feedforward
adjustments occurring prior to and during the movement are programmed

38
before feedback from the ongoing movement can influence them (Gahery 1987;
Massion 1992). Preparatory anticipatory postural adjustments (pPAs) occur
before the movement to assist in movement initiation or stabilization, whereas
associated anticipatory postural adjustments (aPAs) stabilize posture against the
internal reactive torques due to the limb movements (Bouisset and Zattara
1981; 1987; Commissaris et al. 2001; Cordo and Nashner 1982; Friedli et al.
1988; Friedli et al. 1984; Lee et al. 1987; Stapley et al. 1998; Stapley et al.
1999). Preparatory and associated postural adjustments will be referred to as
pPA and aPA, respectively, throughout this thesis.
Classically, APAs have been investigated during single arm raises
performed while standing. These studies have shown stereotypical activation
and inhibition of muscles in the supporting limbs, organized in a distal-to-
proximal sequence, occurs some 50-100 ms prior to the activation of the prime
mover (Belenkii et al. 1967; Bouisset and Zattara 1981; 1987; Marsden et al.
1977; Massion 1992). Biomechanical analyses of these movements have
demonstrated that the muscular activity in the legs is correlated to an upward
and forward acceleration of the CoM prior to the onset of the arm acceleration,
suggesting that APAs function to oppose the backward acceleration of the CoM
due to the arm raise itself (Bouisset and Zattara 1981; 1987). These findings led
to the traditional view that APAs occurring before the focal movement serve to
stabilize the position of the CoM in anticipation of the focal arm displacement.
This view is supported by extensive study of arm flexion (Belenkii et al. 1967;
Bouisset and Zattara 1987; Clement et al. 1984; Friedli et al. 1984; Horak et al.
1984; Lee et al. 1987), push or pull movements on a handle (Brown and Frank
1987; Cordo and Nashner 1982), and unilateral leg flexion tasks (Mouchnino et
al. 1992; Rogers and Pai 1990).
Anticipatory postural adjustments have also been attributed a role in
creating the dynamic forces necessary for initiating movement (Stapley et al.
1999). During locomotion, for example, preparatory muscle activity in the legs
serves to accelerate the CoM for gait initiation (Brenière and Do 1987; Brenière
and Do 1986; Lepers and Brenière 1995). Furthermore, the magnitude and

39
duration of the anticipatory EMG is correlated to the velocity of the first step
(Lepers and Brenière 1995), suggesting that the CNS predicts the dynamics of
the focal movement and integrates this information in planning the APA
according to the task constraints. Interestingly, this hypothesis is supported by a
number of studies in humans that have shown that APAs create the dynamic
forces to assist in the focal movement performance even when the BoS remains
fixed (Commissaris et al. 2001; Lee et al. 1987; Lee et al. 1990; Stapley et al.
1998; Stapley et al. 1999), suggesting that the role of APAs is not strictly for
postural stabilization. For example, when subjects are asked to lift an object or
push/pull on a handle during stance, activations across the postural muscles
serve to direct the GRFs to create the angular momentum that facilitates the
task(Commissaris et al. 2001; Lee et al. 1990). Thus, the view that APAs serve
to strictly stabilize the CoM does not necessarily generalize when a broader
range of movements is considered. Rather, it appears that APAs are
programmed as a function of the task requirements, serving to assist the
movement goals, be it stabilization or acceleration of the CoM.
More recently, substantial evidence in humans has accumulated
demonstrating that APAs are shaped by the biomechanics of a focal movement.
For example, in a reaction time paradigm, the temporal organization of the
APAs preceding single leg flexion is tightly linked to the time constraints of a
the focal task (Rogers and Pai 1990). Furthermore, during voluntary reaching or
pointing movements, APAs are programmed as a function of object distance
(Kaminski and Simpkins 2001; Stapley et al. 1998; Stapley et al. 1999), object
size (Bonnetblanc et al. 2004) and movement velocity (Bouisset et al. 2000;
Horak et al. 1984; Lee et al. 1987). Together, these results suggest that the CNS
anticipates and integrates the stability and dynamic requirements of the focal
movement into the postural command (Massion 1992). Thus, using knowledge
of the body and limb dynamics that is gained with experience and learning, the
nervous system is able to predict the dynamic consequences of the intended
movement and shape the postural adjustment appropriately (Bouisset and
Zattara 1981; 1987).

40
While the temporal organization of APAs in relation to the focal
movement has been studied extensively, less is known about the spatial
organization of feedforward postural activity in relation to voluntary
movements executed in multiple directions. Most studies investigating the
spatial organization of APAs have been restricted to focal movements in the
anterior-posterior direction only (Belenkii et al. 1967; Benvenuti et al. 1997;
Bouisset and Zattara 1987; De Wolf et al. 1998; Friedli et al. 1988; Friedli et al.
1984; Gantchev and Dimitrova 1996; Massion et al. 1999; Shiratori and Latash
2001; Slijper et al. 2002). These have shown that postural muscle activity is
sensitive to the direction of the focal movement and tends to be grouped
according to function. For example, groups of either frontal or dorsal muscles
are recruited for posterior and anterior movements, respectively (Bouisset and
Zattara 1987; Cordo and Nashner 1982; Friedli et al. 1984). However, a
detailed characterization of the directional sensitivity of the APAs, and how
muscles may be recruited synergistically is lacking.
To date, only a few studies have characterized the spatial organization
of APAs across a wider range of movement directions (Aruin and Latash 1995;
Santos and Aruin 2008; Vernazza et al. 1996). In particular, Aruin and Latash
(1995a) characterized the APAs preceding bilateral arm raises, performed as
quickly as possible across multiple directions, ranging from shoulder flexion
(forward) to shoulder extension (backward). Notably, they found that postural
muscles were preferentially activated for either forward or backward arm raises.
Differences in the tuning of the proximal and distal muscles were observed,
suggesting the recruitment of groups of functionally-related muscles. However,
in this study, movements were performed bilaterally and disturbances to the
CoM only occurred along the sagittal plane. How the CNS deals with planned
disturbances along the frontal plane as a result of asymmetrical reaching
movements remains to be examined.
The influence of directionality of an expected perturbation on the
organization of APAs was investigated by Santos and Aruin (2008). Here,
subjects were asked to intercept a pendulum with their right or left hand

41
released by the experimenter from a fixed distance and height. In order to test
the effect of directionality, subjects were positioned so as to produce
perturbations in either the frontal, sagittal or oblique planes. The authors
characterized the APAs in muscles distributed across the body, with an
emphasis on the lateral muscles of the body. In general, their findings extend
those of Aruin and Latash (1995a) by demonstrating the directional sensitivity
of APAs holds for the lateral muscles of the body. Furthermore, they show that
complex patterns of muscle coordination are required for adjusting posture in
preparation for intercepting the target, with combinations of muscles acting
together. The authors suggest that the CNS relies on the flexible recruitment of
muscle synergies to produce task appropriate modulation of muscle activity.
Together, the studies of Aruin (1995; 2008) show distinct patterns of
feedforward postural activity that are sensitive to the direction of the impending
movement. Whether these patterns for controlling posture and balance are
similar to those observed for feedback postural control which are triggered by
afferent feedback, is not known.
2.4 Models for movement control
Humans are able to execute complex movement patterns quickly and
with exceptional accuracy, while interacting with objects in their environment
whose dynamic characteristics may or may not be known. In addition, the CNS
must also consider the inherent delays associated with biological feedback and
the nonlinear relationship between muscle and force generation. From a
computational view, a fundamental question is what models are used by the
CNS to compute the appropriate muscle activation patterns that will results in
efficient movements appropriate for the task and environmental context
(Desmurget and Grafton 2000)?

42
2.4.1 Goal-directed movements require both feedback and feedforward
control mechanisms
Given the rapidity and smoothness of human motion, it is widely
accepted that the CNS cannot rely exclusively on feedback mechanisms to
coordinate movements (Kawato 1999). Similarly, a purely feedforward, or
'predictive,' model programmed entirely before the movement onset would also
be limited in it's ability to respond to unexpected perturbations or errors
occurring during the movement. Consequently, it is proposed that the CNS
relies on a 'hybrid' of feedforward and feedback control mechanisms (Gritsenko
et al. 2009), or a dual-mode of control (Milner 1992), for planning and
executing complex goal-directed movements (Desmurget and Grafton 2000).
Early formulations of this model estimated that the CNS programmed the initial
trajectory in a feedforward manner and only accessed sensory feedback in the
late stages of the movement to ensure accuracy to the target goal (Milner 1992).
More recently, these ideas have been developed in terms of internal models, or
representations, that the CNS has of its own dynamics and the external
environment. The CNS organizes an approximate motor plan prior to the
movement, which is then executed under the 'supervision' of internal feedback
loops that may have non-sensory contributions, which can then modulate the
original plan in real time (Desmurget and Grafton 2000).
Historically, a feedforward model for controlling voluntary movements
had received favorable review in the literature on account of the sensorimotor
delays documented for feedback control were too long to effectively control
movement. For example, sensory feedback from the vestibular or visual system
requires a minimum of 80-100 ms but may be as long as 300-700 ms before it
can influence an ongoing movement, such as visually-guided reaching
(Desmurget and Grafton 2000; Jeannerod 1988; Paillard 1996). However, given
that the CNS is able to respond to errors and modify a motor program online, it
is suggested that feedback signals must be accessible and integrated by the
CNS. Also, movements are more accurate when sensory feedback is present,
highlighting the contributions of feedback to the overall control of movements.

43
Current models stress the importance of strong internal feedback loops, which
rely on internal estimates of efferent and afferent signals, as the primary error-
detecting signal (Davidson and Wolpert 2005; Desmurget and Grafton 2000;
Wolpert and Miall 1996).
2.4.2 Internal models
An important insight that has led to current hypotheses about how the
CNS effectively deals with the inherent delays associated with feedback control
and the nature of the feedback signal was postulated by von Holst and
Mittelsteadt (1950). They proposed that the CNS is able to distinguish between
self-induced motion and a passive displacement of the body with respect to the
environment by storing a 'copy' of the motor command for the goal-directed
motion within the brain (Desmurget and Grafton 2000). An extension of this
idea is that the CNS has knowledge of the properties of the body and its
surrounding environment (Davidson and Wolpert 2005; Kawato 1999; Scott
and Norman 2003). Therefore, the CNS can make predictions about the
expected sensory consequences resulting from internally generated movements
and drive corrections when discord between the expected and observed sensory
signals occurs (Davidson and Wolpert 2005; Wolpert and Miall 1996). These
ideas form the basis of the concept of internal models.
Internal models are defined as the neural processes that estimate the
input/output relationships between the motor system and the CNS (Kawato
1999). Two types of internal models have been proposed. A forward model
estimates the sensory consequence of internally driven motion of the body
using an efference copy of the motor command (Wolpert and Miall 1996). In
contrast, an inverse model employs the reverse transformation to generate the
neural commands for movement based on the desired outcomes or
consequences of that movement (Wolpert and Miall 1996). The internal model
hypothesis predicts that the CNS acquires an inverse model of the body's
dynamics through motor learning, enabling the control of movements in a
feedforward manner (Kawato 1999). In turn, a forward model of an efferent-

44
based copy of the future limb state allows for rapid correction of movements
via feedback mechanisms (Davidson and Wolpert 2005; Kawato 1999). The
existence of internal models is not restricted to the motor system and has been
used extensively to explain processes of the sensory system also (Hess and
Angelaki 1999; Merfeld et al. 1999; Roy and Cullen 2004).
2.4.2.1 Inverse internal models
When performing a voluntary movement, such as a reaching movement
to a target, the CNS calculates the motor commands to generate muscle activity
that will compensate for the arm dynamics and result in the desired kinematic
trajectory of the arm (Kawato 1999). However, in situations where the arm is
faced with novel dynamics, the motor program is no longer appropriate and an
erroneous trajectory is produced. To examine this process, subjects have been
studied while performing reaching movements in dynamic force fields that alter
the dynamics of the environment (Lackner and Dizio 1994; Shadmehr and
Mussa-Ivaldi 1994; Shadmehr et al. 1993). In these paradigms, the force fields
generate consistent forces that depend on the state space, such as the position or
velocity of the segment, and cause deviations of the arm trajectory. Over
repeated trials, subjects learn to perform straight trajectories by internalizing the
dynamics of the environment and modifying their muscle activity (Bhushan and
Shadmehr 1999; Flanders 2011; Ioffe et al. 2007). However, when the force
field is unexpectedly removed, a large error is produced by the subject,
providing evidence that subjects have adapted their model of the inverse
dynamics of the arm (Kawato 1999; Shadmehr and Krakauer 2008).
2.4.2.2 Forward internal models
In order to compensate for the delays inherent to sensorimotor
processes, it has been postulated that the execution of skilled voluntary
movements relies on a prediction of the future state of the motor system using
an efferent-based copy of the motor command (Davidson and Wolpert 2005).
This allows for rapid, corrective responses during movement executions

45
(Gritsenko et al. 2009). Evidence from studies examining how grip force on an
object is adjusted in relation to predictable and unpredictable changes in load
force have substantiated the use of forward models by the motor system
(Davidson and Wolpert 2005; Flanagan and Wing 1997; Johansson and Cole
1992; Kawato 1999). For example, when using a precision grip to hold an
object with stable properties between the fingertips and thumb, the CNS
predicts the changes in load force caused by any self-induced changes in load
force due to voluntary movement of the limb and adjusts grip force in parallel
with the change in load force to prevent slippage of the object (Johansson and
Westling 1984). However, if the object's dynamics are unstable or not known,
grip force is adjusted in a reactive manner to changes in load force. These
results suggest that the CNS uses predictive mechanisms, such as a forward
model, to prevent unwanted delays in motor execution and ensure stability of
movements. Similarly, such predictive mechanisms have been documented for
maintaining the equilibrium of the CoM during the execution of goal-directed
arm movements (Bouisset and Zattara 1981; 1987). Recently, such
computational frameworks have predicted that the CNS relies on state
estimation based on an internal model of the body and sensor dynamics to
process afferent information and calculate the orientation of the body (Kuo
2005).
2.5 The control of voluntary arm movements
Goal-directed reaching movements have been studied extensively in
humans and primates to understand how the CNS plans movements and
integrates afferent and efferent signals into the control mechanism (see review,
(Georgopoulos 1986). The kinematic, dynamic and muscle activity for visually-
guided reaching movements are well documented, although the roles of
different sensory inputs to the planning and execution of the reach remain
highly debated (Sarlegna and Sainburg 2009). It appears that reaching
movements to visual targets are planned using information about the initial
position of the hand and the final target position. Likely, initial hand position is

46
encoded by a weighted summation of proprioceptive and visual feedback,
whereas the end of reach position is highly influenced by visual information of
the target (Rossetti et al. 1995). Following illumination of the target, there is a
sequential organization of the eyes, head and arm toward the target (Biguer et
al. 1982). Furthermore, the arm trajectory to the target is characterized by a
bell-shaped velocity curve. Typically, the acceleration and deceleration phases
are of equal duration, however the deceleration phase may be lengthened when
vision of the arm or hand position is restricted (Elliott et al. 1991) or when
increased accuracy is required, such as when reaching to targets reduced in size
(Soechting 1984).
2.5.1 Online control of visually-guided reaching movements
The contributions of the visual and proprioception systems to the
control of aimed reaching movements have been examined more closely in
paradigms where an unexpected shift in target position is introduced. These
double-step paradigms, initially developed to study saccadic eye movements
(Becker and Jürgens 1979; Levy-Schoen and Blanc-Garin 1974; Wheeless et al.
1966), have provided a means to investigate the underlying visuomotor
processes that contribute to the rapid, smooth correction of the arm trajectory
during visual perturbations of reaching movements. When a shift in target
position occurs very early in the reach trajectory, such as immediately before or
at reach onset, very short latency corrections of the reach occur (Day and
Brown 2001; Day and Lyon 2000; Pélisson et al. 1986; Prablanc and Martin
1992). Subjects successfully correct their arm trajectory, as indicated by a lack
of inflection point in the kinematic profile of their reach, at latencies similar to
reaction times for unperturbed reaches without significantly increasing their
movement time. These online corrections are presumed to be automatic in
nature, given that they occur in situations where the subjects are unaware of the
target shift, such as during the saccadic suppression (Bard et al. 1999; Johnson
and Haggard 2005; Pélisson et al. 1986). However, when the target shift occurs
very late in the deceleration phase of the movement, subjects were unable to

47
successfully correct their reaching movement (Komilis et al. 1993). Vision of
the arm is not critical for making correction, however end-point accuracy of the
reaching movement is improved when subjects can visualize their arm and the
target (Komilis et al. 1993; Pélisson et al. 1986; Sarlegna et al. 2003).
2.5.2 Standing imposes equilibrium constraints during perturbed reaching
An interesting question that arises from the literature documenting the
online control of voluntary movement is, how does the CNS resolve
equilibrium constraints when corrective reaches are performed in the standing
position? Answering this question is fundamental to understanding why falls
often occur in dynamic situations where both the movement and postural
systems are challenged. When arm movements are perturbed in the standing
position, the CNS must correct the perturbed arm movement in addition to
dealing with the equilibrium constraints imposed by the upright posture and
decreased BoS.
To date, only two studies have investigated how the CNS corrects goal-
directed movements when standing (Fautrelle et al. 2010; Martin et al. 2000).
Using a modified version of the classic double-step paradigm, Martin and
colleagues (2000) investigated how the likelihood of an impending shift in
target position along the sagittal plane affected the kinematics for reaching
movements executed in the standing position. They found that the kinematics of
the arm movement remained stable across conditions, however trunk flexion
was increased to compensate for the uncertainty of the reaching movement,
suggesting that uncertainty about the movement was integrated in the motor
program for posture and movement in a predictive fashion (Martin et al. 2000).
Similarly, Fautrelle and colleagues (2010) examined the modulation of
postural control during online corrections of reaching movements in standing.
They showed that rapid corrections for arm and leg muscles occurred at
approximately the same latency of 100 ms. At such short latencies, the authors
suggested that the online corrections of posture and movement occur
automatically and are organized at subcortical levels through rapid internal

48
feedback loop mechanisms (Fautrelle et al. 2010). It is not clear from their data,
however, whether the correction in the leg preceded that of the arm. More
detailed information about the temporal organization of the online corrections
in posture and movement will provide additional insight into the central
organization of posture and movement. Therefore, it remains to be seen whether
the CNS updates the commands for posture predicatively of the impending arm
correction, or whether the CNS relies on information from the arm movement
to update posture for the maintenance of balance.
2.6 Summary and direction for future investigation
Several important questions related to the nature and central
organization of postural control during voluntary movements remain to be
addressed. Most studies investigating the central control of these two
behaviours have focused on the temporal coupling between the ‘GO’ signal
triggering a voluntary movement, the onset of the postural changes
accompanying the movement and the onset of the movement related changes
themselves. While the studies detailed in this chapter have provided
fundamental insight into the coordination of feedforward postural adjustments,
a detailed examination of the spatial organization of pPAs and aPAs is lacking.
This information will allow for comparison to other types of postural
behaviour, namely the strategies for maintaining balance organized via a
feedback mode of control. It is expected that this knowledge will contribute to
elucidating the neural structures involved in organizing the general strategies
for posture control.
A second important question is how the CNS resolves the constraints for
equilibrium control when voluntary movements executed in standing are
perturbed and online corrections in the movement are necessary. Specifically,
how are commands for posture and movement controlled in relation to one
another? What models are adopted for ensuring successful execution of the
task? It is commonly accepted that correcting movement errors in the standing
position presents a dynamic and challenging situation for many, as evidenced

49
by the frequency of falls occurring in these types of situations. Therefore, it is
important to investigate the underlying mechanisms for posture and movement
control in order to prevent falls and rehabilitate fall-prone individuals.
The body of work presented here aims to further our understanding the
mechanisms of equilibrium control by examining the strategies for maintaining
balance during goal-directed reaching movements to targets located in multiple
directions by focusing on the above-mentioned topics.

50

51
Chapter 3
General Methodology
The general aim of the thesis was to gain insight into how the CNS
coordinates the control of posture and voluntary movement. To investigate the
questions central to this thesis, an experimental protocol was designed where
subjects were required to reach towards targets in multiple directions while
standing. The rational for adopting this paradigm and the general characteristics
of the experimental paradigm common to each study will be presented here.
The specific experimental details for each are provided in Chapters 4, 5 and 6,
respectively.
3.1 Rational for experimental protocol
Traditionally, questions related to the coordination of posture and
movement have been investigated by characterizing the kinematics, kinetics and
muscle activity generated during arm raising (flexion/abduction) movements
while standing (Bouisset and Zattara 1981; 1987).While this classical approach
allows for a clear distinction between postural and focal components of the
movement, it is limited in it’s ability to fully explore the spatial organization of
APAs, given that the arm motion in these paradigms is generally limited to only
a few directions. Furthermore, it has been previously highlighted that in order
to assess the structure of the data set and determine whether muscle synergies
reflect the organization of the CNS and not the structure of the experimental
design (Macpherson 1991), the experimental paradigm should have more
muscles and experimental conditions than the number of synergies that could be
expected (Ting and Mckay 2007). Thus, in our experiments, to examine the
spatial and synergic organization of feedforward postural adjustments
preceding and accompanying reaching movements, directionality of postural

52
activity was established by requiring subjects to point to multiple targets (a total
of 13), interspersed over a range of 180°.
A primary objective of the thesis is to explore the spatial organization of
feedforward postural activity in the context of what is known about feedback
postural control to generate knowledge about the central control of posture and
movement. This will be addressed in Chapters 4 (SA1) and 5 (SA2). In
response to an unexpected perturbation of balance, APRs accelerate the CoM
and body to a region of stability within a fixed BoS (Horak and Macpherson
1996). These feedback responses depend mainly on afferent signals (Allum et
al. 1994; 1993; Stapley et al. 2002) that shape the timing and amplitude of the
response. In contrast, feedforward postural commands are centrally generated
with descending commands to postural muscles (i.e., rather than being triggered
by an afferent signal) (Massion 1992). In order to make comparisons between
the organization of these modes of control, a paradigm is needed in which the
task goals are similar: to accelerate the CoM and body upon a fixed BoS. In this
regard, a task requiring feedforward postural activity to accelerate the CoM
within a fixed BoS would more closely match the behavioural goals of APRs,
enabling more direct comparisons between feedback and feedforward modes of
postural control. Thus, a reaching task (described below in detail) was designed
where subjects performed reaching movements in standing to targets located
just beyond arm’s length in multiple directions. This paradigm provides an
excellent opportunity therefore, to characterize and quantify feedforward
postural activity in a task where the goal is to accelerate the CoM upon a fixed
BoS.
The third objective (SA3) of the thesis is to examine the nature of the
control signal underlying aPAs accompanying voluntary movements in
situations where the task goal changes after the focal movement has been
initiated. Previous attempts to explore the online control of posture during
voluntary movement have utilized a visual double-step paradigm. However,
these studies have focused mainly on how uncertainty about target location
affects movement performance during standing (Martin et al. 2000) or how

53
quickly the motor system can initiate a correction to the arm movement given
the additional equilibrium constraints of standing (Fautrelle et al. 2010).
Although these studies provide important insight about movement corrections
in standing, neither specifically addressed how posture is updated with respect
to the movement correction. It is important to know how the CNS achieves this
control, and to determine whether the movement correction relies on a postural
correction for successful execution of the movement as people often experience
falls in dynamic situations (Inglis and Macpherson 1988). Thus, borrowing
from the double-step paradigm, we were able to specifically quantify the
postural adjustments occurring just before, and concurrently with the correction
in arm trajectory so as to provide insight regarding the interaction of posture
and movement in these dynamic motor tasks.
3.2 Overview of experimental protocol
3.2.1 Experimental apparatus
The general experimental set-up utilized for SA1 and SA2 is shown in
Figure 3.1. For all experiments, right-handed subjects performed reaching
movements with their dominant arm to targets located in multiple directions
while standing. Subjects stood barefoot on two triaxial force plates (model
FP4060; Bertec, Columbus, OH), which were centred in a custom-built target
light array, fully adjustable in height and target distance. The light array was
suspended from the ceiling and could not provide support to the subject upon
contact with the target. The array contained 13 light-emitting diodes (LED)
targets, interspersed by 15°, with the 0° target located to the subject’s right, 90°
at their midline and 180° to their complete left. Each target consisted of a red
LED encased in a modified gaming switch (model 459512; RP Electronics,
Burnaby, BC, Canada) that produced a 5V pulse upon depression of the switch.
Targets were placed at shoulder height, at a standardized distance of 130% of
the subject’s reach length, as measured to each target. Pilot tests demonstrated
that this distance ensured that significant adjustments in posture were engaged

54
to attain the target, while maintaining a fixed BoS. Reach length was measured
between the subject’s xiphoid process (point from which they initiated their
pointing movements) and the tip of the right index finger when the arm was
extended to each target. Measurements were performed with the subject centred
in the array (xiphoid process aligned with the 90° target) while they stood
straight, and maintained neutral scapular retraction. For leftward targets,
rotation of the trunk was permitted in order to maintain the shoulders square to
the target. This was done to determine the distance to reach the target in
postural configurations that most similarly matched those adopted during the
reaching movements.
Figure 3. 1: General experimental set-up. Plan view of target light array
illustrating subject orientation on the force plate relative to the array. Targets
are arranged from right (0°) to left (180°).

55
3.2.2 Behavioural task
Questions related to the feedforward control of posture were explored
by quantifying the postural adjustments preceding and accompanying a discrete
reaching task performed in standing. For all experiments, subjects adopted their
natural stance width, with one foot on each of the adjacent force plates and their
midline aligned to the 90° target. Subjects began each trial with their left arm
hanging loosely by their side and their right index finger depressing a switch
affixed to their chest, which when released provided a 5V step signal. Upon
random illumination of one of the target LEDs, subjects were instructed to point
and depress the illuminated target at their preferred movement speed.
Immediately upon contact with the target, subjects were asked to release the
target but hold the body configuration with their arm outstretched to the target
for a period of 2 seconds (indicated by extinction of the illuminated LED). By
requiring subjects to stabilize their body at the target, it was possible to
examine whether task goal (acceleration versus stabilization of CoM) affected
the organization of the feedforward postural adjustments. No constraints with
respect to movement speed or finger trajectory were specified.
3.2.3 Protocol specific to SA1 and SA2
In SA1 (Chapter 4) and SA2 (Chapter 5), subjects performed discrete
reach to point movements to one of 13 targets. Testing began with an
acclimatization period where subjects pointed to each target twice, presented in
random order. Following these trials, subjects executed a total of 15 trials to
each target (195 trials) and 15 catch trials where no target illuminated. Catch
trials were presented in order to minimize prediction of the occurrence and
direction of the illuminated target. All target directions were presented in a
pseudorandomized order, in blocks of approximately 50 trials interspaced with
5-minute rest period to reduce the effects of fatigue. Details of the calculated

56
data variables and analysis for SA1 and SA2 are presented in Chapters 4 and 5,
respectively.
3.2.4 Protocol specific to SA3
A modified version of the classic double-step paradigm was developed
for investigating the nature of the control underlying associated postural
adjustments accompanying voluntary reaching movements (SA3; Chapter 6).
Specifically, the basic reaching paradigm of SA1 was modified so that a
voluntary reaching movement could be disrupted by changing the task goal
following onset of the reach. This was achieved by programming a shift in the
illuminated target after the onset of the reaching movement (release of the chest
switch). In these experiments, four targets were used: the central, or 90°, target,
and the three consecutive targets to the right of the central target (45°, 60°and
75°). Two types of trials were presented to subjects: (i) “reach” trials that
consisted of discrete reach movements to the central target (90°) and (ii) “corr”
trials, that required an online correction of the arm trajectory to one of the other
three targets after a reach was initiated to the 90° target. As in SA1, subjects
stood barefoot on two triaxial force plates, with their xiphoid process aligned to
the central target (90°). Each trial began with the subject depressing the chest
switch with their right index finger and their left arm hanging loosely relaxed
by their side. After a variable delay, the central target (90° or L1) would
illuminate. Subjects were instructed to reach and press the target at their natural
speed. Upon contact with the target, subjects were to release the target but
maintain the postural configuration with their arm outstretched to the target
until the target LED extinguished, which corresponded to a period of 2 seconds.
On some trials (corr trials), the target light would shift from L1 to any one of
the other three targets: 75° (corr75), 60° (corr60) or 45° (corr45) after a variable
delay from the reach onset. Subjects were instructed to point to the newly
illuminated target (L2) upon detection of the light shift. The target shift would
occur after a variable delay following online detection of the voltage drop of the
chest switch signal upon release of the chest switch. Subjects performed a block

57
of acclimatization trials consisting of five regular reaches to each target.
Subjects were subsequently presented with trials in random order, including at
least 60 reach trials to the 90° target, 15 trials to each of the corr conditions
(corr75, corr60 and corr45), and 15 catch trials were no target illuminated.
Subjects typically performed blocks of 40 trials, separated by rest periods of 5
minutes.
3.2.5 Data collection and analysis
For all experiments, ground reaction forces (GRF) and moments in the
mediolateral (x), anteroposterior (y) and vertical (z) axes were recorded with
two triaxial force plates (model FP4060; Bertec, Columbus, OH) sampled at
1000-Hz. Three-dimensional body kinematics were collected using a six camera
MX3 motion-capture system (Vicon Peak, Lake Forest, CA) that sampled at
200 Hz. Muscle activity from a total of 16 leg, trunk and arm muscles was
recorded using two DelSys Bagnoli 8-channel systems (Delsys, Boston, MA).
Custom-written programs written in LabVIEW (National Instruments, Austin,
TX) controlled the illumination of the target lights and acquired signals from
the chest and target switch. Synchronization with the analog signals from the
force plates and EMG system was done using the Vicon controller. Custom
programs were written in Matlab (The MathWorks, Natick, MA) for processing
and analyzing the collected data. The details of the data analysis for each
experiment are presented in the respective chapters.
3.3 Significance of the experimental paradigm provides basis for
further exploration
The experimental paradigm developed for this thesis provides a basis
for investigating fundamental questions related to the coordination of posture
and movement. In the first place, reaching tasks in standing provide a motor
task where there is a clear dissociation of postural and focal components of the
movement. Furthermore, by examining postural control strategies adopted

58
when performing reaching movements in multiple directions, we are able to
contribute knowledge about the spatial organization of feedforward postural
adjustments that complement current knowledge about feedback-based postural
responses to rotations and translations of the support surface. Finally, this
paradigm provides a unique opportunity to fully explore the nature of deficits of
postural control observed in populations with pathologies of the motor systems,
such as aging or Parkinson’s Disease. Comparisons of the temporal and spatial
patterns of postural muscle activity to baseline measures determined for healthy
individuals can provide important insight into the mechanisms of the disrupted
motor behaviour.

59
Chapter 4
Reaching to multiple targets when
standing: The spatial organization of
feedforward postural adjustments
4.1 PREFACE
The first study of this thesis was motivated by a perceived gap in
knowledge regarding the strategies for feedforward postural control for
reaching movements in standing. Previous studies of support surface
translations in multiple-directions have demonstrated the existence of a force
constraint strategy (Henry et al. 2001; Macpherson 1988a) and muscle activity
that is directionally-tuned (Henry et al. 2001; Macpherson 1988b) for feedback
postural control. Whether the CNS relies on similar strategies for simplifying
the control of posture organized in feedforward is not known. Therefore, as a
primary aim, this chapter examined the spatial and temporal organization of the
GRF and EMG patterns of feedforward postural adjustments proceeding and
associated with pointing movements in multiple directions executed in
standing. Furthermore, the experimental paradigm developed for the study in
this chapter provides a novel means for investigating the coordination of
posture with respect to voluntary movements.
This chapter was adapted from Leonard JA, Brown RH, and Stapley P.J.
Reaching to multiple targets when standing: The spatial organization of
feedforward postural adjustments. Journal of Neurophysiology 101(4): 2120-
2133, 2009. This manuscript has been reprinted with permission from The
American Physiological Society, publisher of the Journal of Neurophysiology.
The paper is presented in the same format in which it was published with the
exception of formatting to figures and tables to comply with McGill University
thesis formatting guidelines.

60
4.2 ABSTRACT
We examined the spatial organization of feedforward postural
adjustments produced prior to and during voluntary arm reaching movements
executed while standing. We wished to investigate if the activity of postural
muscles before and during reaching was directionally tuned and if a strategy of
horizontal force constraint could be observed. To this end, 8 human subjects
executed self-paced reach-to-point movements upon the random illumination of
1 of 13 light targets placed within a 180° array centered along the mid-line of
the body. Analysis was divided into 2 periods: a first corresponding to the 250
ms preceding the onset of the reaching movements (termed pPA period), and a
second 250 ms period immediately preceding target attainment (the aPA
period). For both periods, EMG activity of the lower limb muscles revealed a
clear directional tuning, with groups of muscles being activated for similar
directions of reach. Analysis of horizontal ground reaction forces supported the
existence of a force constraint strategy only for the pPA period, however, with
those in the aPA period being more widely dispersed. We suggest that the
strategy adopted for feedforward pPAs is one where the tuned muscle synergies
constrain the forces diagonally away from the centre of mass (CoM) to move it
within the support base. However, the need to control for final finger and body
position for each target during the aPA phase resulted in a distribution of
vectors across reaching directions. Overall, our results would support the idea
that end-point limb force during postural tasks depends upon the use of
functional muscle synergies, which are used to displace the CoM or decelerate
the body at the end of the reach.
4.3 INTRODUCTION
For multi-joint movements executed during standing such as reaching
forwards, postural adjustments occurring prior to movement onset shift the
CoM within the base of support in order to initiate the movement, and

61
associated postural adjustments overcome the postural disturbances related to
movements of the limbs (Bouisset and Zattara 1981; 1987). To ensure a
controlled transition from one postural configuration to another, these
adjustments of posture must be planned by the central nervous system (CNS) in
advance, and a feedforward mode of neural control sends commands to both
focal and postural muscles to initiate and stabilize posture. Both the preparatory
and associated postural adjustments (pPAs and aPAs, respectively) are
considered to be feedforward in nature as they are produced before feedback
from the ongoing movement can influence them (Gahery 1987; Massion 1992).
The objective of the present study was to examine the spatial organization of
postural muscles and forces produced before and during voluntary reaching
movements in multiple directions to gain insights into the nature of their
underlying control.
Feedback mediated postural responses to unexpected disturbances of
balance have been well characterized. When the surface upon which humans
are standing unexpectedly moves, the body is destabilized in the direction
opposite to that of the surface displacement. In order to regain balance, humans
produce short latency automatic postural responses (APRs) in the supporting
limbs that oppose the perturbation and drive the CoM back towards its initial
position relative to the support surface (Horak and Nashner 1986). The latency
from the initiation of the support surface movement to the onset of the evoked
EMG response is in the order of 80-120 ms in humans (Horak and Macpherson
1996; Nashner 1977; Ting and Macpherson 2004). These compensatory APRs
are triggered by somatosensory feedback from the feet and legs (Bloem et al.
2000; Bloem et al. 2002; Horak and Macpherson 1996; Stapley et al. 2002) and
unless prior warning of the upcoming perturbation is given (Jacobs and Horak
2007; McChesney et al. 1996), they are produced entirely using a feedback
mode of neural control.
Studies in animals and humans have examined feedback-based APRs to
unexpected translations of the support surface in multiple directions with the
aim of identifying strategies that the CNS may adopt to simplify the control of

62
perturbed stance (Fung et al. 1995; Henry et al. 2001; 1998b; Macpherson
1988a; b; Ting and Macpherson 2004). In the cat, regardless of the direction of
horizontal platform translation, force vectors were exerted in one of two main
directions at each limb (termed the ‘force constraint strategy’, (Macpherson
1988a). Muscle activity displayed a tuning across directions with maximal
amplitudes for each muscle arising for one specific direction (Chanaud and
Macpherson 1991; Macpherson 1988b). Similar results have also been obtained
in standing humans subjected to postural perturbations (Henry et al. 2001;
1998b). Overall, results have supported the hypothesis that force vector
production is a high level parameter adopted to reduce the multiple of degrees
of freedom associated with complex postural tasks, and that the production of
the desired vector is solved at a lower level by the synergic recruitment of
muscle groups. More recent studies in the standing cat have elaborated this
hypothesis, and have suggested that an internal model of postural force
generation coordinates functional muscle synergies rather than biomechanical
limb constraints alone (McKay et al. 2007; McKay and Ting 2008). However,
the production of a force constraint upon the relationship between the muscle
synergies produced and the current limb geometry (Torres-Oviedo et al. 2006).
Recent studies have shown that neural commands for feedforward
postural adjustments can be identified in the pontomedullary reticular formation
(PMRF) of the brainstem (Schepens and Drew 2006; 2004; Schepens et al.
2008). Neurons in this area discharged either during the pPA, the aPA or both
during reaching movements in the standing cat (Schepens and Drew 2006;
2004). The activity of this area has also, however, been shown to be implicated
in compensatory, or feedback-based, postural control. Microstimulation of
peripheral afferents, known to be essential for triggering short latency APRs
(Stapley et al. 2002), activate particular subsets of reticulospinal neurons (Drew
et al. 1996) and PMRF neurons are activated strongly during postural
perturbations in the standing cat (Stapley and Drew 2009). If specific neural
sites can modulate postural responses with different underlying modes of
control (i.e., feedforward or feedback), it is plausible to predict that the outward

63
expression of those postural adjustments may show similarities in their
organization. The present study aimed, therefore, to investigate if a similar
spatial organization of horizontal force and muscle activity to that seen with
feedback-based postural responses could be observed for predictive
feedforward postural adjustments in humans, despite their different modes of
neural control. To generate directionality, we asked human subjects to reach
and point to targets in multiple directions throughout 180° at shoulder height
whilst standing. Postural adjustments immediately preceding the onset of the
focal limb movement (pPAs) and those produced before target attainment
(aPAs) were recorded and analyzed. We hypothesized that: 1) feedforward
postural adjustments are directionally-tuned to the current goal of the task
(initiation or termination of the reaching movements), and 2) ground reaction
forces show directional force constraint, supporting the idea that such a strategy
is a high level parameter adopted by the CNS, regardless of the mode
(feedforward or feedback) of neural control.
4.4 MATERIALS AND METHODS
4.4.1 Subjects
Eight (1 female and 7 male) healthy subjects, without any known
neurological, visual or orthopedic disorders, were recruited from the McGill
University student population to participate in the present study. Subjects had a
mean age of 22.5 ± 4.3 (SD) years, a mean height of 1.74 ± 0.04 (SD) m, and a
mean weight of 66.6 ± 5.9 (SD) kg. They were all right-hand dominant. They
gave their informed consent to participate and experiments were conducted with
the approval of the McGill University research ethics board
4.4.2 Experimental apparatus and set-up
Subjects stood barefoot on 2 tri-axial force plates (model FP4060,
Bertec Corp., Columbus, OH, USA) that measured ground reaction force (GRF)
and moments in mediolateral (X), anteroposterior (Y) and vertical (Z) axes at

64
1000 Hz. Stance width (the mediolateral distance between the feet) was taken
as the average of each subject’s natural stance distance as measured after 3
trials of walking the length of the laboratory. Foot position was marked for each
subject and care was taken that subjects kept their feet in the same position for
the duration of the experiments. They were centered in a custom-built 180°
light target array, fully adjustable in height and target distance (see Fig. 1A).
The array contained a total of 13 light emitting diodes (LEDs) each spaced at
15° intervals. Light targets were 2.5 cm in diameter and consisted of 5V red
LEDs encased in modified gaming switches (model 459512, RP Electronics,
Burnaby, BC) that produced a 5V pulse upon contact. The gaming switches
were mounted at the ends of lightweight aluminum dowels adjustable in length,
affixed to a semi-circular aluminium bar suspended from the ceiling. A chest
band worn by all subjects was also equipped with the same switch in order to
detect movement onset (see below).
The activity of 16 muscles was recorded using 2 DelSys Bagnoli 8-
channel systems (Boston, MA, USA) at 1000 Hz. The following muscles were
recorded bilaterally (left and right legs): tibialis anterior (TAl and TAr,
respectively), soleus (Soll, Solr), lateral gastrocnemius (GasLl, GasLr),
peroneus longus (Perl, Perr), biceps femoris (BFl, BFr), rectus femoris (RFl,
RFr) and tensor fascia latae (TFLl, TFLr). In addition, anterior and posterior
deltoid was recorded at the right shoulder. Bilateral kinematic data were
collected using a 6 camera MX3 motion capture system (ViconPeak Inc., Lake
Forest, CA, USA) sampling at 200 Hz. A total of 36 markers were placed at
different locations on the subjects’ whole body as listed in the Plug-in-Gait
model (Vicon Peak ®). This model provides an accurate estimate of CoM
position when compared to the method of ground reaction force integration
(Gutierrez-Farewik et al. 2006). Analog signals from the force plates and EMG
system were captured through the Vicon MX3 controller. A customized
program written in LabView (National Instruments, Austin, TX, USA) was
used to control the illumination of target lights, acquire and synchronize digital

65
signals from the switches (target and chest), and initiate and synchronize data
collection with the Vicon system.
4.4.3 Experimental Procedures
Subjects were centered in the array with their mid-line (xiphoid process)
aligned with the 90° target direction and the 0° and 180° target directions
representing, respectively, each subject’s far right and left side targets (see Fig.
4.1A). Upon the random illumination of 1 of the 13 target lights, subjects were
asked to point and press the lighted switch. Target lights were placed at a
standardized distance of 130% of the outstretched right arm when holding it in
line with each target. The distance was measured between the subject’s xiphoid
process (from where they initiated pointing movements) and the tip of the right
index finger when the arm was extended towards each target. Subjects were
asked to stand straight, maintain neutral scapular retraction during the
measurement, but were permitted to rotate the trunk such that the shoulders
were facing square to the target for leftward targets. This was done in order to
measure the distance to the targets in a postural configuration similar to that
adopted during the reaching movements. During pilot tests, it was found that
the target distance of 130% could be attained comfortably by the subjects using
a combined arm and trunk movement, but did not place them at their limits of
stability at target attainment (determined by measuring centre of pressure
displacement within the base of support). Targets were all at right acromion
height as measured during quiet stance. No constraints of accuracy were given
to subjects, only that they had to point and press the light switch at their natural
speed, immediately following target illumination.
Subjects began their reaching movements with the index finger of their
right hand pressing the chest switch centered at the xiphoid process of the
sternum and their left hand hanging vertically at the side of the body. They
were asked to reach to, press and release the target switch with their right index
finger and hold this body position for 2 s, then return slowly and assume their
initial position. The total acquisition period consisted of a 3 s window. The data

66
collection time line is illustrated in Fig. 4.1B. Once the experimenter was
satisfied that the subject was standing quietly data acquisition was begun. After
a random period of between .5 s and 1 s, a target illuminated and subjects
initiated their movements. The total acquisition period of 3000 ms was
sufficient to record the postural activity preceding and accompanying the
movements. Other than movement speed (natural speed), no other instructions
regarding the strategies to be adopted were given. Subjects were not encouraged
to move as fast as possible (a reaction-time paradigm). Testing began with an
acclimatization period, during which subjects executed 26 trials in random
order, twice to each of the targets. Following the acclimatization period,
subjects were required to execute 15 trials towards each pointing direction, in a
pseudo-randomized order, which included 15 catch trials (no target light
illuminated) to reduce the possible prediction of the upcoming target light.
Thus, a total of 210 trials per subject was recorded and used in the subsequent
analysis. Generally, subjects performed blocks of approximately 50 trials inter-
spaced with 5 min rest periods to reduce fatigue until the required number of
trials at each direction was attained.
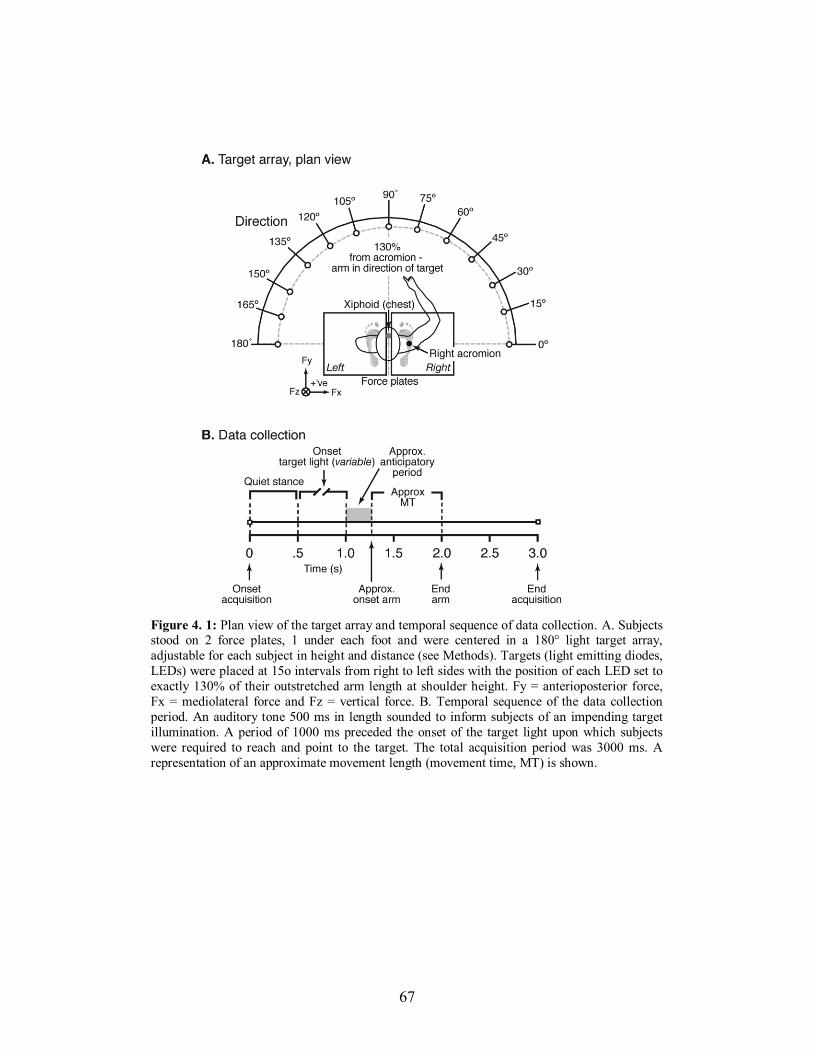
67
Figure 4. 1: Plan view of the target array and temporal sequence of data collection. A. Subjects stood on 2 force plates, 1 under each foot and were centered in a 180° light target array,
adjustable for each subject in height and distance (see Methods). Targets (light emitting diodes,
LEDs) were placed at 15o intervals from right to left sides with the position of each LED set to
exactly 130% of their outstretched arm length at shoulder height. Fy = anterioposterior force,
Fx = mediolateral force and Fz = vertical force. B. Temporal sequence of the data collection
period. An auditory tone 500 ms in length sounded to inform subjects of an impending target
illumination. A period of 1000 ms preceded the onset of the target light upon which subjects
were required to reach and point to the target. The total acquisition period was 3000 ms. A
representation of an approximate movement length (movement time, MT) is shown.

68
4.4.4 Data analysis
Kinetic, kinematic and EMG data were recorded and stored on a PC
computer for further analysis using a series of customized programs written in
MatLab (The Mathworks, Natick, MA, USA). Ground reaction forces and
moments were low-pass filtered using a digital second-order Butterworth filter,
with a 10Hz cut-off frequency. Raw EMG signals were high-pass filtered at
35Hz, de-meaned, rectified, and low-pass filtered at 100Hz (second-order
Butterworth filter). Individual trials were visually inspected for stability during
the quiet stance period (1000 ms prior to target illumination). Any trials that
showed significant variation in Fz and/or CoP during the quiet stance phase (0-
500 ms following data acquisition onset) were eliminated from further analysis,
as were trials in which subjects did not make contact with the target, or used the
target as a support. This was evaluated by checking that the centre of pressure
(CoP) did not leave the limits of the BoS determined using the kinematic
markers on each foot in relation to foot width (included in the Vicon model)
and stance distance and whether switch depression was maintained. Following
the trial selection criteria, a total of 1437 trials from the 8 subjects were retained
for further analysis from a total of 1560 trials.
Focal movement onset and termination were established by chest switch
release and target light depression, respectively. The onset of movement using
the switch was compared to the first deflection above zero of the tangential
velocity of the marker placed on the hand in the Y-axis and movement
termination was also taken as the moment when hand velocity returned to zero.
As no difference greater than 5 ms was found between the onset of motion as
determined by the chest switch and the motion capture data (sampling 1 image
approximately every 5 ms), movement time was taken as the duration between
chest switch release and target depression. Onset of focal movement acted as
time zero upon which all preparatory events were based.
To investigate the temporal and spatial organization of feedforward
preparatory postural adjustments prior to movement onset (hereon called the

69
pPA period) and associated postural adjustments produced during the end phase
of the reaching movements (the aPA period), the analysis was restricted to 250
ms before chest switch release (pPA) and 250 ms before the end of the
movement (aPA). The choice of the period lengths was based upon well
documented changes in anticipatory postural adjustments preceding voluntary
movement, or the pPA period (Belenkii et al. 1967; Bouisset and Zattara 1981;
Crenna and Frigo 1991) and visual inspection of the onset of braking forces and
moments exerted under each foot before the end of the movements for a
number of trials in each direction (aPA period). Both of these periods were
divided into five 50 ms long ‘bins’ chosen to characterize the evolution of the
preparation and the termination of reaching movements (e.g., pPA1, pPA2,
etc.). EMG amplitudes were calculated as the mean of each 50 ms bin for each
period and for each muscle. For each period (pPA and aPA) and muscle, the
highest mean response across the 13 reach directions was used to normalize
muscle activations, so that values ranged between 0 and 1. Normalized
amplitudes were then plotted as muscle tuning curves. Amplitude and direction
of the resultant horizontal force were calculated by summing changes in Fx and
Fy, according to trigonometric vector addition (Zar 1999). Resultant horizontal
plane vectors were plotted in polar coordinates. The torque about the vertical
axis at each foot (Tz) was calculated using the following equation:
Tz = Mz − Xp • Fy + Yp • Fx
where Xp and Yp are the coordinates of the centre of pressure of each
foot, Mz is the moment of the force plate around the vertical (z) axis, and Fy
and Fx are anterior-posterior and mediolateral forces, respectively.
4.4.5 Statistical analysis
The effect of pointing direction upon movement time was examined
using a repeated measures one-way analysis of variance (ANOVA). To
determine if a force constraint strategy characterized the horizontal ground
reaction forces in each of the periods under study, the angle of the average
vector produced in successive 50 ms bins was pooled from all subjects and

70
subjected to circular statistical analysis. Circular statistics refers to a class of
techniques developed for the analysis of directional or cyclic data, assuming an
arbitrary zero and where 0° and 360° represent the same direction. To
determine if pooled vector directions were uniformly distributed or constrained
(defined as a significant clustering of the active horizontal force vector,
Macpherson 1988a), data were examined for a bimodal distribution using the
‘broken- axis approach’ (Holmquist and Sandberg 1991). This approach tests
the null hypothesis that populations of vector direction are uniformly
distributed around a circle against the working hypothesis that they are not so.
Specifically, it evaluates bi-modality, but does not assume that means of
clusters of vectors are separated by 180°. If a bimodal distribution is found, it
returns the mean angles of the two modes. Typically, when analyzing circular
data sets, to evaluate the degree of dispersion mean vector length (r) is
computed. Values may vary between 0 (high dispersion) and 1 (all data
concentrated along a single preferred direction). In the broken axis approach,
rmax represents the mean vector length (r) where the modality of the data set
(k) best fits the data. The value of k specifies the type of modality and no
assumption is made that the data is uni- or bimodal. For example, if k=2, the
data would be symmetrically bimodal (i.e., 2 clusters separated by 180°).
However, when k lies between 1 and 2 the data set would be characterized by
an intermediate clustering of a bimodal distribution where clusters are separated
by 360°/k (e.g., when k=1.6, modes are separated by angle of 225°). This
approach also gives the mean direction (alpha) of any number of clusters that
can be identified. All statistical analyses were custom written and performed in
Matlab.
4.5 RESULTS
4.5.1 Kinematics of reaching movements during standing
Examples of the general kinematic strategies adopted by subjects when
pointing to targets in the array are illustrated in 3D in Figs. 4.2A-C. Three

71
principle directions of reach are shown for 1 representative subject. For all
directions, subjects began from a similar initial quiet stance position with the
right finger pressing the chest switch and the left hand by the side of the body
regardless of reaching direction. The similarities of the initial CoM positions at
movement onset can be seen in Fig. 4.2D (open colored squares). For
movements to 0°, subjects executed a rightward extension of the arm, a
clockwise rotation of the torso and a slight rightward displacement of the pelvis
(Fig. 4.2A). For movements to 90° (Fig. 4.2B) the reaching arm outstretched
approximately along the body mid-line, while the torso rotated forwards and
slightly downwards. Finally, for far leftward movements (Fig. 4.2C), the right
arm rotated leftwards, crossing the mid-line, with the torso and pelvis also
rotating towards the left. For each of the 3 movements shown, the CoM was
displaced from its initial position in the approximate direction of the target
within the base of support. Ground reaction force vectors during the pPA period
(red vectors) were oriented so that the CoM moved in the direction of each
target. At the end of the aPA period the GRF vector was oriented back towards
the CoM (blue vectors) to brake the movement of the body. Although there was
some variability (especially between 90° and 120°), trajectories and end CoM
positions differed for each reaching direction. This is represented for 5 trials for
each direction in Fig 4.2D. Despite the greater amount of body rotation required
by rightward movements, movement times did not significantly increase across
the 13 directions. Average movement times for all subjects are shown in Table
4.I.
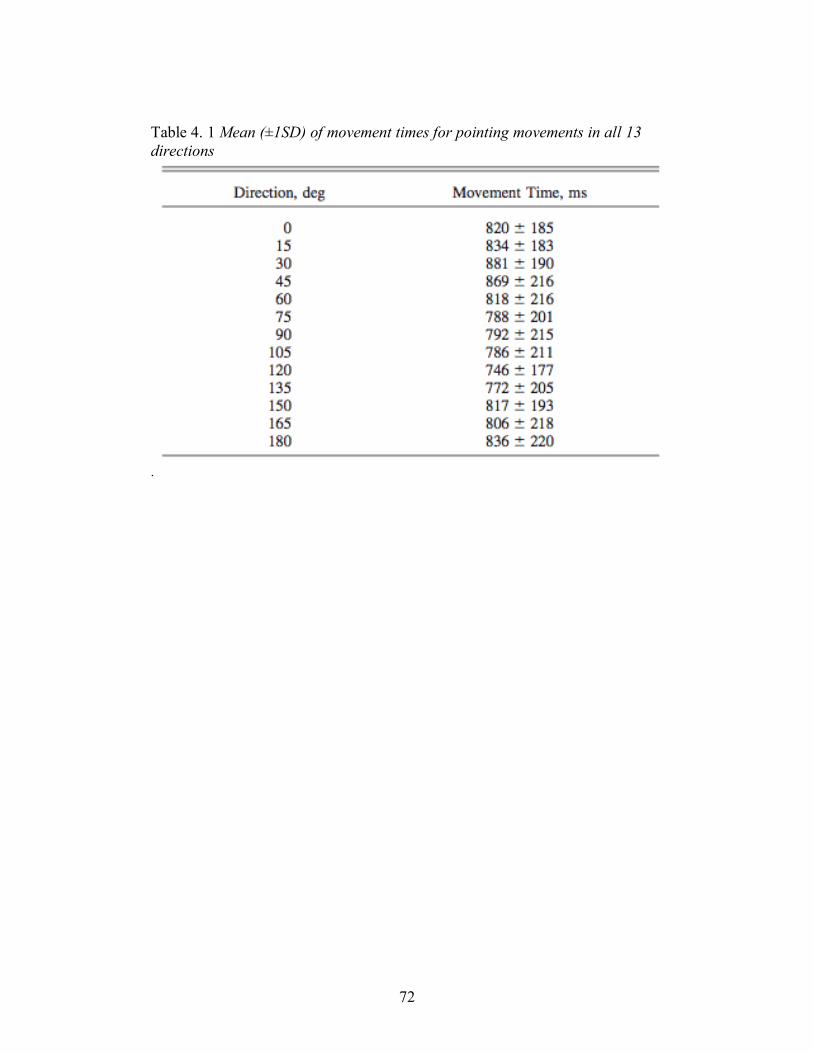
72
Table 4. 1 Mean (±1SD) of movement times for pointing movements in all 13
directions
.

73
Figure 4. 2: 3D kinematic representations of reach to point movements to 3 principal target
directions. (A. 0°, B. 90° and C. 180°) for 1 subject (S5). Stick figures are shown as if being
viewed from the front and slightly to the subject’s left (A) and right (B and C) sides. Body
movements are shown at 2 different times: At the onset of finger movement (grey sticks) and at
the end of the movement (black sticks) when the finger touched the target. Shown are the
following segments bilaterally: foot, shank, thigh, shoulder, upper arm and lower arm. Markers at the level of the 7th cervical and 10th
thoracic vertebrae, the clavicle and sternum form a single
segment that represents the torso in the sagittal plane. The head is represented by markers
placed at 4 locations on the left and right temples and at the same level at the back of the head.
Finger trajectory is shown in red. The body centre of mass (CoM) is shown as a grey and black
circles (onset and end of the movements, respectively). The ground reaction force vector is
represented at the onset of movement in red, and at target attainment in blue. D. Trajectories of
the CoM from the onset of the focal movement (open squares) to when the finger touched the
target (open circles). Trajectories are represented for 5 trials in each direction and are colour-
coded as per the legend across the bottom of the figure.

74
4.5.2 EMG activity in relation to the forces produced: pPA period
There was a clear modulation of EMG activity and force between 0 and
180° target directions during the pPA and aPA periods. Changes in muscle
activity and force for 3 main directions of reaching are shown in Figs. 4.3A-C,
for 1 representative subject. For clarity, the two 250 ms time periods under
study have been shaded. During the pPA period, regardless of pointing
direction, the earliest change in EMG activity was typically an inhibition of
either the right or left soleus muscles. Postural adjustments for far rightward
movements (Fig. 4.3A) began with an inhibition of Solr followed by an
activation of TFLl. This was followed by a leftward and backward push at the
left leg and an unloading of Fz under the right foot. The torque about the
vertical axis (Tz) showed that the left foot exerted a counter-clockwise (CCW)
moment, while the right foot exerted a clockwise (CW) one. Centrally-oriented
movements (Fig. 4.3B) showed a clear bilateral soleus inhibition/tibialis
anterior activation and both peroneus muscles activated, which likely assisted
the initiation of the forward displacement of the body. The TFLl and RFl also
showed anticipatory bursts, as did the BFr. This may have reflected the slightly
asymmetric nature of the movement using the right arm and hand reaching to
the 90° target. There was very little or no change in Fx under either foot but
instead an increase in Fy (backwards push) and Fz (loading) under the right
foot, coupled with a slight forwards push at the left foot. As these movements
involved essentially a forward displacement, there was only a slight CW Tz at
the left foot. For movements to the far leftward 180° target (Fig. 4.3C), which
required the greatest amount of body rotation, there was significant anticipatory
activity in the BFr. Additionally, there was a burst in the Solr. The forces
showed that the right (loaded) foot Fx pushed rightwards exhibiting a CW Tz.
Interestingly, although unloaded, the left foot also showed a CW Tz.

75
Figure 4. 3 Electromyographic activity, changes in force and vertical torque (Tz) under each
foot for reaching movements to 3 principal directions (A. 0°, B. 90° and C. 180°). Traces are
shown for a period of 500 ms preceding movement onset until the end of each movement for
one typical trial in subject 5. On each plot, the full grey vertical line indicates the onset of the
light target (Light on). The dashed grey line to the left of movement onset (Movt on) indicates the onset of force and Tz changes during the pPA period. The dashed grey line to the right of
movement onset (between 500 and 750 ms) indicates the end of the arm movement (Movt end).
Forces are shown as forces exerted against the ground. TFLr and TFLl = tensor facia latae
muscles (right and left, respectively), RFr and RFl = rectus femoris, BFr and BFl = biceps
femoris, GasLr and GasLl = gastrocnemius lateralis, , Perr and Perl = peroneus longus, TAr and
TAl = tibialis anterior, Solr and Soll = soleus. Fx = mediolateral force, Fy = anterioposterior
force, Fz = vertical force and Tz = torque exerted around the vertical axis. Left =leftwards, back
= backwards, load = loading and CCW = counterclockwise. Left and right foot forces are
represented by solid and dashed traces, respectively (see legend). Shaded regions represent the
2 periods of 250 ms under study (pPA and aPA). Successive squares underneath the time axes
pictorially represent changes in Tz at each foot during the two periods.

76
4.5.3 EMG activity in relation to the forces produced: aPA period
During the aPA period, from 250 ms before to the end of the
movements, there were also distinct patterns of muscle activity across the
reaching directions, mostly in the extensors. Rightward (0°) movements (Fig.
4.3A) were characterized by activation of the GasLr, Solr, TAr and Perr. Apart
from the slight activation of TFLr early in this period, there was little activity in
the other muscles. Forces showed that the right foot was loaded, while the
horizontal forces stabilized around their original values. The loaded (right) foot
exerted a CCW Tz. Reaches to 90° (Fig. 4.3B) showed bilateral activity in a
number of extensor muscles (e.g., gastrocnemius, soleus, biceps femoris and, to
a lesser extent, peroneus). The right foot was loaded and pushed forwards (Fy),
while Fx forces steadied. The loaded (right) foot exerted a CCW Tz during this
period, but which reversed shortly after the hand reached the target (see arrow
and asterisk to the right of Movt End). Finally, reaches to 180° (far leftward,
Fig. 4.3C) showed activity in left-side extensors (GasLl and Soll) as well as
right side flexors (BFr and TAr). The loaded left foot pushed in a slightly
rightward and backward direction exerting a CCW Tz.
4.5.4 Feedforward postural adjustments show directional tuning and
are synergic
There was a clear directional tuning of many of the postural muscles
recorded during the 2 periods of study (see Fig. 4.4). In the pPA period,
patterns of EMG activity showed that the tibialis anterior and peroneus, as well
as the rectus femoris (all bilaterally) were activated principally for pointing to
mid-range targets, between approximately 45° and 120°. Other muscles, such as
the tensor facia latae, and soleus showed reciprocal patterns of activity between
reaches to the left and right. The TFLl activated between 0° and 75°, the TFLr
between 105° and 180°, while the Soll displayed inhibition during the pPA
period between 45° and 180° and sustained activity for 0° to 30° movements, as

77
did the Solr between 150° and 180°. The gastrocnemius muscles showed no
activity in the preparatory period, except for GasLl at the extreme rightward
pointing direction (0°). Other muscles, such as the BFr displayed asymmetric
patterns of activity, activating between 60° and 180° on the right side, but not
on the left.
In the aPA period reciprocal patterns of activity could also be noted for
the extensors soleus and gastrocnemius, left side muscles activating for targets
75-180° and those on the right side from 0- 90°. The BFl also activated for 75-
180° targets, while other left side muscles remained largely inactive. On the
right side, the flexor Perr contracted along with Solr as did the TFLr to a
certain extent. Interestingly, BFr showed activity across virtually all reaching
directions during this period

78
Figure 4. 4 Representative EMG traces for 14 selected muscles for S5 across the 13 directions
of pointing. Muscle activity is shown for a total duration of 500 ms, 250 ms before and after the
onset of the pointing movement. Muscle name conventions are as described in Figure 4.3. The
shaded area to the left of time zero on each muscle plot represents the 250 ms preparatory
period. Unless shown, muscles have the same scaling for the left leg (top row) as they do for the
right leg (bottom row).

79
To evaluate the spatial and inter-trial variability characteristics of the
muscular patterns described above, tuning curves were generated for each of the
14 postural muscles recorded during 5 successive 50 ms bins. For both pPA and
aPA periods, each muscle was activated for a range of pointing directions with
many showing directions of maximal recruitment. Typically, during the pPA
period (black lines and open circles) in bins 1 and 2 (pPA1 and pPA2,
respectively) all muscles were generally inactive or showed a baseline level of
activity across directions, hence, for clarity, they are not depicted in Fig. 4.5.
However, in pPA3 (-150 to -100 ms) a directional tuning began to emerge,
despite the evident inter-trial variation that existed. During pPA4 and pPA5, the
tuning curves show that the muscles worked in groups, with TAr/TAl, Perr/Perl
and RFr/RFl activating maximally for targets either side of the central one,
TFLr, BFr and Solr for leftward targets and TFLl and GasLl for rightward
movements. Clear patterns of tuning were also evident for all bins of the aPA
period (grey lines and filled circles) with the patterns emerging from bin 1
onwards (250 to 200 ms before target attainment). Again, for clarity and due to
the constancy of tuning across the 5 bins in this period, only bins 3-5 are
depicted in Fig. 4.5. The muscles Solr, GasLr, Perr and TFLr activated for
similar directions, as did GasLl, Soll and BFl. The BFr showed increasing
activity from 0-180°.
This pattern of muscle tuning was very consistent across the 8 subjects
tested. This can be seen in Fig. 4.6. Here, muscles are depicted for the final bin
of the pPA period (Fig. 4.6A) and aPA period (Fig. 4.6B) and are grouped
according to the directions for which they were recruited to a similar extent.
Figure 4.6C summarizes the recruitment of each of the muscles in the groups
identified in the 2 periods in polar coordinates. The spatial pattern of each
group represents approximately the outermost limits of the EMG tuning.

80
Figure 4. 5 Muscle tuning curves for the EMG activity of all 14 postural muscles during the
final 3 bins of the preparatory and associated periods for the representative subject S5.
Differences in tuning and recruitment of the muscle studied can be observed by comparing the
activity of the muscles over the 3 5 equivalent bins (left to right columns). Dots indicate
amplitudes from each trial measured and the solid lines the mean responses. Muscle name
conventions are as described in Figure 4.3.

81
Figure 4. 6 Muscle tuning curves in the final bin of the pPA (A) and aPA (B) periods for each
of the 8 subjects studied. Muscles have been grouped into the 3 major groups that activated for similar directions of reach. Tuning curves and individual trials are represented as in Figure 4.5.
Schema (C) summarizing the approximate range of directions of reach to which each identified
group contributed.

82
4.5.5 Spatial patterns of force differ between preparatory and
associated postural adjustments
Horizontal GRFs displayed distinct and opposing patterns for the 2
periods studied. The forces produced during the pPA period approached more
of a bi-modal distribution with low dispersion of the vector directions, whereas
those produced in the aPA period, although not unimodal, were more highly
dispersed, suggesting that to control final body position for each direction of
reaching, a unique vectors of force were produced under each foot.
These trends are illustrated in Fig. 4.7 for a representative subject. All
trials for this subject are plotted as horizontal vectors for each successive bin of
50 ms for each period. In the early phases of the pPA period (Fig. 4.7A, pPA1
and pPA2) horizontal forces were of very small amplitude and showed a high
degree of dispersion in terms of their direction. However, from the bin pPA3
onwards, which corresponded approximately to the emergence of EMG tuning
described above and shown in Fig. 4.5, forces, in particular under the loaded
foot shared similar directions (e.g., 0° to 75° for the left foot and 105° to 180°
for the right foot). Clearly constrained directions of force can be seen under the
left foot in pPA4 and pPA5, which was loaded for targets 0°-90° pushing
outwards and leftwards (see different black-blue colored vectors) and under the
right foot, loaded for 105°-180° targets, pushing outwards and rightwards (see
yellow-orange vectors), although right foot vectors were always slightly more
distributed. For clarity in Fig. 4.7A, the black arrows convey the general
direction of the exerted force in the constrained vectors when the limb was
loaded limb and grey arrows when it was unloaded. The constraint observed in
the horizontal vectors during pPA4 and pPA5 is supported by the circular
statistical analysis (see below). Interestingly, the forces produced under the foot
that was unloaded during these bins often also showed a constraint of horizontal
force under the left foot, but was slightly more dispersed under the right.

83
During the aPA period, the feet that were loaded for their respective
directions of reach during the pPA period now became unloaded and vice versa.
In bins pPA4 and pPA5 loaded vectors exerted force backwards and outwards
at the foot contra-lateral to the direction of reach (e.g., left foot, pPA5, blue
vectors). In comparison, during the aPA period, loaded vectors now pushed
outwards and forwards at the limb ipsilateral to the direction of reach (e.g.,
aPA5, orange-red vectors). Such a low dispersion of vectors seen during the
latter stages of the pPA period (especially under the left foot) was not seen
during all bins of the aPA period. Horizontal force vectors produced under the
loaded or unloaded feet in the aPA period were highly distributed, often
throughout a 90° range or greater (see Fig. 4.7D).
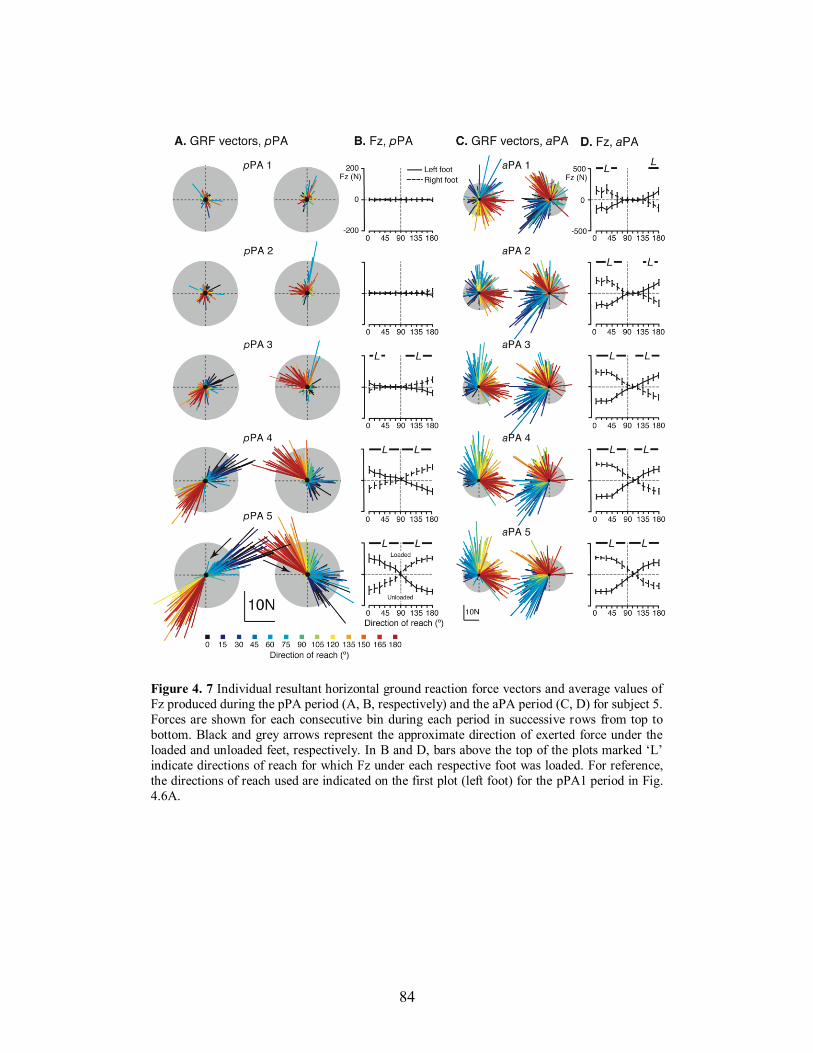
84
Figure 4. 7 Individual resultant horizontal ground reaction force vectors and average values of
Fz produced during the pPA period (A, B, respectively) and the aPA period (C, D) for subject 5.
Forces are shown for each consecutive bin during each period in successive rows from top to
bottom. Black and grey arrows represent the approximate direction of exerted force under the
loaded and unloaded feet, respectively. In B and D, bars above the top of the plots marked ‘L’
indicate directions of reach for which Fz under each respective foot was loaded. For reference,
the directions of reach used are indicated on the first plot (left foot) for the pPA1 period in Fig.
4.6A.

85
Circular statistical analysis (for an explanation, see Methods) supported
this trend of a force constraint strategy during the pPA but less so during the
aPA. The statistical analysis resulting from the broken axis approach is given in
Table II for both feet. During the pPA period, the analysis identified an angular
distribution that was significantly different from uniform for progressive bins.
The distribution of horizontal force was characterized by a clustering of vectors
with k values that increased during pPA4 and pPA5 to >1.5 for the left foot and
1.35 and 1.42, respectively, at the right foot. This indicated that an asymmetric
bimodal distribution existed during those periods. Additionally, rmax values
were > 0.5 (e.g. 0.66 and 0.77 for the left foot and 0.55 ad 0.6 for the right foot
during bins pPA4 and pPA5 respectively) indicating a low degree of dispersion.
Such an evolution of significant bimodal clustering was not seen during the
aPA period. Rather, statistical analysis, revealed that the distribution of force
vectors was significantly different from uniform with rmax values ≤0.5,
indicating higher dispersion than in the pPA period around a single mode.

86
Figure 4. 8 Average direction and magnitude of horizontal ground reaction force change during
each bin of the pPA and aPA periods under both feet.
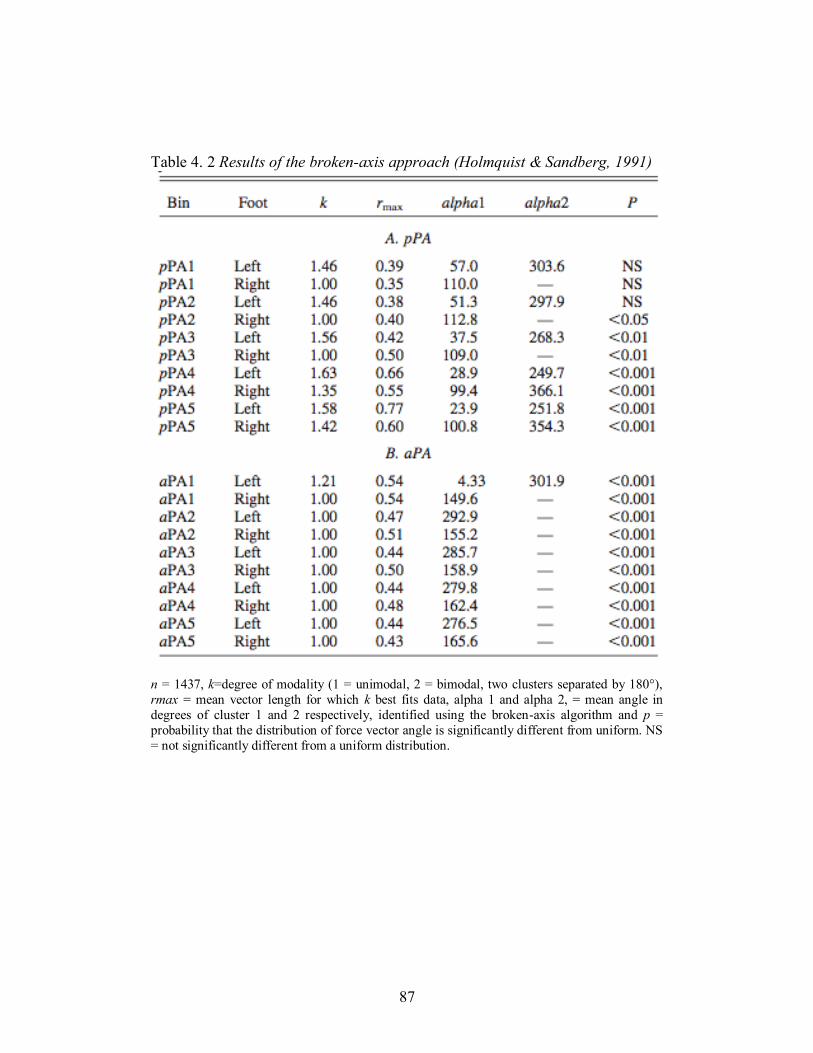
87
Table 4. 2 Results of the broken-axis approach (Holmquist & Sandberg, 1991)
n = 1437, k=degree of modality (1 = unimodal, 2 = bimodal, two clusters separated by 180°),
rmax = mean vector length for which k best fits data, alpha 1 and alpha 2, = mean angle in
degrees of cluster 1 and 2 respectively, identified using the broken-axis algorithm and p =
probability that the distribution of force vector angle is significantly different from uniform. NS
= not significantly different from a uniform distribution.

88
The bimodal distribution of force vector direction highlighted by the
circular statistics in the pPA period is also reflected when average vector
direction is plotted against each reach direction (Fig. 4.8A). It can clearly be
seen that there was a constraint of force vector direction during pPA4 and
pPA5, especially under the left foot. This constraint of force vector direction
seen in the pPA period was also accompanied by a modulation of force
magnitude with targets at each extremity having the largest GRF (Fig. 4.8B,
pPA4 and pPA5), whereas the aPA period displayed a linearity of GRF vector
and reaching direction from bin 2 onwards (Fig. 4.8C) and a fairly constant
magnitude of GRF vector across all directions (see Fig. 4.8D, all bins).
4.6 DISCUSSION
We investigated the organization of feedforward postural adjustments
produced when standing humans reached with their preferred arm to multiple
targets placed in a semi-circle throughout 180° with centered respect to their
midline. Our objective was to identify spatial tuning of postural muscle activity
and a constraint of horizontal GRF, similar to that shown for feedback- based
postural responses to unexpected perturbations of balance in humans (Henry et
al. 2001) and animals (Macpherson 1988a). The results supported our first
hypothesis; that bilateral EMG activity recorded was directionally-tuned and
served the current goal of the task (initiating or terminating the movements).
However, our second hypothesis; that a generalized force constraint strategy
existed for feedforward postural adjustments, was not completely supported.
4.6.1 The roles of preparatory and associated postural adjustments
for reaching during stance
The postural adjustments studied in the two periods likely performed
functionally different roles. In the first period (pPA), significant EMG activity
emerged approximately between -150 ms to movement onset. The resultant

89
horizontal forces produced under the loaded foot during this period were
consistently in the opposite direction to the desired movement direction, the
vectors being constrained for a number of reaching directions. Moreover, the
moments (Tz) produced indicated that the loaded foot consistently exerted a
torque in the opposite direction to the direction of reach (Fig. 4.3), thus creating
a reaction torque that drove the body towards the target. These results would
support the notion that the pPA created the necessary conditions for CoM
displacement in the direction of the reach, within the base of support (Stapley et
al. 1998; Stapley et al. 1999). This is also supported by the CoM trajectories in
Fig. 4.2D. The pattern of force and torques exerted at the feet reversed during
the aPA period, such that the reaction forces opposed the body displacement as
the hand reached the target. Perhaps the clearest example of this could be seen
under both feet for 180° reaches (Fig. 4.3C), which required the greatest
amount of body rotation. The CCW direction of Tz would have had the effect
of slowing and stabilizing CoM position within the support base as the hand
neared the target. Such a role of associated postural adjustments has been
shown during various arm reaching or lifting tasks (Commissaris and Toussaint
1997; Commissaris et al. 2001; Cordo and Nashner 1982; Krishnamoorthy and
Latash 2005; Schepens and Drew 2003).(Macpherson 1991)
4.6.2 Tuned, synergic muscle activity characterizes feedforward
postural adjustments
The activity of the 14 muscles recorded in the pPA and aPA periods
showed that each muscle was recruited for a range of directions often with a
direction of maximal activation. Muscles were also activated in groups. This
grouping across the two periods would support a synergic organization of
feedforward muscular activity. A synergy has been defined as a group of
muscles constrained to act in a concerted manner (Macpherson 1991;
Sherrington 1961) or ‘activated in synchrony with fixed relative gains and
muscle activation patterns with consistent spatial characteristics’ (Torres-

90
Oviedo and Ting 2007). Directionally tuned feedforward muscle activity has
previously been identified during bilateral arm movements performed when
standing (Aruin and Latash 1995a). When the two arms were moved either
forwards or backwards from a central position, groups of muscles were
activated in functional groups on the dorsal or ventral side of the body to
maintain equilibrium. Direction-specific patterns of feedforward muscle activity
have also been identified in dorsal muscles in anticipation of perturbations to
equilibrium (Latash et al. 1995; Santos and Aruin 2008). The muscle activity
identified in the present study also served a range of directions and was
maximally tuned to a specific reaching direction in both periods studied. We
suggest therefore that the patterns of muscular activation seen in both periods
may belong to the same limited number of robust functional muscle synergies,
despite the inter-trial variations seen in each subject reflecting different levels
of synergy activation. However, APRs are primarily triggered by afferent
feedback from the moving surface, whereas in the present study the postural
adjustments were produced in anticipation of the upcoming movements.
Therefore, similarities in the synergic organization of feedback and feedforward
postural adjustments may support the idea that the CNS adopts functional
synergies for both modes of control to produce different motor behaviors (Ting
2007).
4.6.3 Clearly constrained force patterns are seen during preparatory
but not during associated feed- forward postural adjustments
A force constraint strategy has been defined as a bimodal clustering of
active force vectors where the forces are constrained to act along an
approximately diagonal axis directed roughly toward or away from the CoM at
a single limb (Macpherson 1988a; 1994). Until now, this strategy has been
identified in the horizontal forces comprising the feedback-based postural
adjustments produced when the surface upon which animals or humans were
standing was unexpectedly displaced (Henry et al. 2001; Macpherson 1988a;

91
1994). Our results have shown statistically, that a force constraint strategy
existed for feedforward postural adjustments accompanying a pointing task,
specifically during the latter stages of the pPA period (pPA4 and pPA5) during
which the objective was to displace the CoM in the direction of the target.
During these bins the angular distribution was significantly different from
uniform and rmax values (quantifying the degree of dispersion, see Methods)
supported a low dispersion of vector direction. Our rmax values were
comparable to, if not greater than, those produced by cats perturbed in the
horizontal plane standing at natural stance width (Macpherson 1994). Average
values of k during these same periods (pPA4 and pPA5) were between >1 and
<2, indicating that an asymmetric bimodal distribution existed (less than 180°
separation existed between clusters). We conclude, therefore, that a force
constraint strategy existed for pPA’s. Such an observed pPA period force
constraint strategy may represent a high-level task variable whereby the CNS
groups together the dynamic forces required to execute a number of directions
of reaching in order to simplify the complex control of multijoint movement.
It remains, however, that such a low dispersion of force vector was not
observed in our study for the forces exerted during the aPA period. Values of
rmax were <0.5 (more highly dispersed) and only one single alpha value was
identified. This more dispersed force pattern observed during the aPA period
when compared to the pPA period may be explained by the current nature of
the task. Task instructions were to maintain final finger position at target
position and not to return to the initial position. Thus, the constraints of
displacing the CoM from one position to another, as in the pPA period, did not
apply. Rather, subjects had to maintain their body position and stability at target
attainment. Force constraint strategies have been documented to exist mostly
when humans or animals are required to actively displace the CoM back
towards its initial position following a perturbation (Henry et al. 1998a; 2001;
Macpherson 1988a; Ting and Macpherson 2004). In this instance, the force
constraint strategy would simplify neural control mechanisms to coordinate
force direction and amplitude during these ‘active’ responses (McKay et al.

92
2007). During the aPA period of this study, the task was simply to decelerate
the body to target position and remain in that position. It did not require a
similar ‘active strategy’ of CoM (and body) displacement, but rather a
maintenance of posture. Thus, we suggest that the force constraint strategy
characterizes feedforward postural adjustments when the goal is to displace the
CoM within the base of support (likened to the active adjustment following a
perturbation) but not when task requirements are to precisely control an end
posture and finger position during voluntary reach to point movements.
Finally, it may be asked why the force constraint strategy observed
during the pPA period was weaker at the right foot than at the left? The right
foot, ipsilateral to the reaching arm, showed a less constrained pattern of
horizontal force during pPA4 and pPA5. It is possible that the left foot
(contralateral to the reaching arm) was primarily responsible for producing the
turning moment to targets 0-90°. Indeed, Fig. 4.3A shows that when loaded, the
left foot produced a CCW Tz, while the right foot opposed that torque. The
reverse was not true, however for movements to 180°. Here, the left foot
(although unloaded) exerted a Tz that was in the same direction as the right to
drive the body around to the left. Moreover, at this direction during the aPA
period, the right foot consistently opposed the direction of reach, likely to assist
the braking of the movement. Thus, we tentatively suggest that the leg
contralateral to the arm assists in turning to its contralateral side, whereas the
leg ipsilateral to the reaching arm is perhaps more coordinated with the upper
limb and establishes a base for the limb’s trajectory during the reaching task.
Another possibility is that as all subjects were right-handed, their right leg (and
foot) was also their dominant one. If so, it may be that the dominant limb is
better adapted to controlling the limb’s trajectory by producing more dispersed
vectors, while the left leg ensures a postural transition at the onset of reaching
directions for which it is loaded. A differentiation of trajectory control versus
steady-state limb posture has been identified for dominant and non-dominant
arms (Duff and Sainburg 2007). Our further experiments will vary the reaching
arm used in an attempt to confirm these hypotheses.

93
4.6.4 Implications for the neural control of balance: shared control of
feedforward and feedback postural adjustments
This study has shown that EMG tuning characteristics in the pPA period
are organized in a similar manner to compensatory postural responses produced
via feedback. This raises an important question: To what extent do similar
neural pathways control posture during voluntary movements and following
unexpected perturbations? It is well known that central commands for
feedforward and feedback postural adjustments originate at supraspinal levels.
Commands for APAs arise in different regions of the cortex (Gurfinkel and
Elner 1988; Massion et al. 1989; Wiesendanger et al. 1987). In addition, spinal
cats have difficulty maintaining equilibrium when the support surface is
unexpectedly displaced horizontally (Macpherson and Fung 1999) and do not
exhibit the complex patterns of evoked EMG characteristic of APRs in the
intact animal (Macpherson et al. 1997). Lesion studies have shown that damage
to the brainstem reticulospinal system impairs balance (Gorska et al. 1990;
Gorska et al. 1995; Lawrence and Kuypers 1968). In addition, the injection of
cholinergic agonists, which excite reticular neurons or noradrenergic agents that
block inhibitory neurons to the reticular formation affect feedforward
programming of postural responses accompanying reaching (Luccarini et al.
1990). More recently, it has been shown that signals in the brainstem
pontomedullary reticular formation (PMRF) contribute to feedforward postural
adjustments during reaching with neurons in the PMRF encoding either pPAs
or aPAs, or both (Schepens and Drew 2006; 2004). It has also been shown that
microstimulation of peripheral afferents, known to be essential for the early
triggering of APRs (Stapley et al. 2002), also activate particular subsets of
reticulospinal neurons (Drew et al. 1996). Moreover, the activity of RSNs in the
same anatomical area of the PMRF as those related to feedforward postural
adjustments (Schepens and Drew 2006; 2004), also contribute to the
compensatory postural responses recorded in the limbs following unexpected

94
perturbations (Stapley and Drew 2009). Thus, evidence would suggest that an
integration of feedforward and feedback modes of control may occur in the
brainstem.
What would be the end result of commands sent from the brainstem to
the muscles producing both types of postural adjustments? Recent evidence has
shown that, rather than a unique muscle synergy existing for a particular
direction of postural perturbation, APRs are organized into a few muscle
synergies which represent the general neural strategy that accounts for the
spatio-temporal components of the response (Torres-Oviedo and Ting 2007).
These muscles synergies, or primitives, may be used to control task level
variables such as CoM motion or displacement (Ting and Macpherson 2004) or
CoP displacement (Krishnamoorthy et al. 2003). We propose that the similarity
in organization of preparatory (feedforward) postural adjustments and
compensatory (feedback) postural adjustments may take place in the PMRF.
This structure would then organize the functional muscle synergies required in
either mode of control and the outward expression of those synergies, (the
active force vectors) required to produce whole body movements or active
corrections to balance. The results of this paper do not, however, enable us to
conclude that feedforward and feedback based postural adjustments share
common ‘motor primitives’. Further study is needed therefore, to characterize
the muscle synergy organization during feedforward and feedback postural
tasks in the same muscles of the same subject.
4.7 ACKNOWLEDGEMENTS
This study was supported by a Canadian Foundation for Innovation
(CFI), New opportunities fund grant (#10613) and a Natural Sciences and
Engineering Research Council (NSERC), Individual discovery grant to Paul J.
Stapley. The authors acknowledge the expert technical assistance of JJ Loh. We
also thank Drs. Jane Macpherson and Trevor Drew for helpful discussions
about earlier versions of this work.

95
Chapter 5
Muscle synergy characterisation of
feedforward postural adjustments during
reaching in standing humans
5.1 PREFACE
Chapter 4 investigated the spatial organization of feedforward postural
adjustments during reaching to targets in multiple directions while standing. It
was found that while muscle activity was directionally-tuned to the direction of
reach, forces exerted at the ground were constrained in direction, with an
anisotropic relationship observed between GRF direction and reach direction.
These characteristics bear striking resemblance to the spatial organization of
feedback-based postural responses to unexpected support surface translations in
cats (Macpherson 1988a; b) and humans (Henry et al. 2001; 1998b). In
feedback postural control, the variability in muscle activity has been explained
with muscle synergies. Therefore, we postulated that a limited number of
muscle synergies may underlie the patterns of muscle activity during
feedforward postural adjustments. Thus, to examine this question further,
Chapter 5 investigates whether muscle synergies underlie the feedforward
preparatory postural adjustments (pPA) occurring prior to the onset of multi-
directional reaching movements performed in the standing position.
This manuscript has been prepared for submission to the Journal of
Neurophysiology.
5.2 ABSTRACT
This study investigated whether a limited number of robust muscle
synergies underlie the observed patterns of feedforward postural muscle activity
for reaching movements during stance. We hypothesized that the recruitment of

96
a few muscle synergies could account for the spatiotemporal and inter-trial
variability of preparatory postural adjustments for multidirectional reaching
movements in the standing position. Nine subjects were asked to point to 1 of
13 light targets spaced at 15° intervals from their right to left sides (centered at
midline) while standing on two triaxial force plates. The EMG activity of 14
postural muscles were recorded and quantified in 5 time bins of 50 ms
preceding the onset of the focal movement (pPA period, presumed to be
entirely feedforward in nature) and subjected to a non-negative matrix
factorization (NNMF) analysis (Lee and Seung 1999; Torres-Oviedo and Ting
2007). For the six subjects analyzed, the temporal and spatial patterns of pPA
were well represented by the recruitment of 8 or less muscle synergies, as well
as inter-trial variability of muscle activity. Muscle synergy recruitment was
directionally tuned with respect to reach direction. While these results show
similar features in number of synergies and spatial organization to those
observed for automatic postural responses, robustness of muscle synergies for
feedforward posture control remains to be determined by evaluating whether
the same synergies are preserved for the feedforward postural adjustments
accompanying the reaching movement. Overall, these findings provide a first
step in examining whether a set of robust muscle synergies can be identified
across both feedforward and feedback modes of postural control.
5.3 INTRODUCTION
Voluntary movements executed in the standing position, such as raising
an arm or reaching out to an object, are accompanied by postural adjustments
that occur before and during the execution of the action to stabilize the body
against the perturbing effect of the goal-directed movement (Horak and
Macpherson 1996; Massion 1992). Much of what is known about the
organization of these feedforward or predictive postural adjustments, which
occur in advance or concurrent with voluntary movements, comes from the
study of single arm or leg movements (Massion 1992; Mouchnino et al. 1992).
This previous work has focused largely on the temporal aspects of postural

97
muscle activity in relation to the onset of muscle activity initiating the
voluntary component of the tasks, which were executed primarily along a single
plane. However, everyday movements such as reaching to objects, frequently
involve lateral movements of the arms to the side or across the body midline.
The postural adjustments required to initiate and stabilize posture during the
execution of reaching movements in multiple directions have been studied by a
number of authors. Results have shown that complex patterns of muscle activity
in the legs and trunk that make up functional groups which are spatially ‘tuned’
to the direction of the reaching movement (Aruin and Latash 1995a; Leonard et
al. 2009; Santos and Aruin 2008).
Spatial tuning of postural responses has been demonstrated for
feedback-based postural responses following support surface perturbations
(Carpenter et al. 1999; Henry et al. 2001; 1998b; Macpherson 1988b; Ting and
Macpherson 2005; Torres-Oviedo and Ting 2007; 2010). These automatic
postural responses (APR) represent coordinated patterns of muscle activity
organized to accelerate the center of mass (CoM) to a position of stability with
respect to the base of support (BoS) (Chiel et al. 2009; Lockhart and Ting 2007)
and are shaped by afferent feedback that signal the characteristics of the
postural disturbance (Stapley et al. 2002; Ting and Macpherson 2004). Detailed
analyses of APR have revealed that only a small number of muscle synergies
are necessary to explain the temporal and spatial variability of the complex
patterns of observed postural muscle activity (Ting and Macpherson 2005;
Torres-Oviedo et al. 2006; Torres-Oviedo and Ting 2007). It has been
postulated that the recruitment of muscle synergies simplifies the task of
controlling balance by enabling the CNS to send a single command to recruit a
group of muscles, rather than controlling each muscle individually (Cheung et
al. 2005; Ting 2007; Tresch et al. 1999).
A modular organization of the musculoskeletal system has been
proposed as a mechanism by which the CNS simplifies the control of
movement in both postural and goal-directed motor behavior (Ting 2007; Ting
and Mckay 2007). In this framework, muscle synergies, or M-modes

98
(Krishnamoorthy et al. 2003; Krishnamoorthy et al. 2004), represent groups of
muscles that are constrained to act together (Macpherson 1991; Ting and
Macpherson 2005; Torres-Oviedo et al. 2006; Torres-Oviedo and Ting 2007)
and recruited by the CNS to produce predictable biomechanical functions
(Chiel et al. 2009; Chvatal et al. 2011; Ting and Mckay 2007). The concept of
muscle synergies has been applied in several species to explain the muscle
coordination patterns in a variety of motor behaviors, including balance control
(Chvatal et al. 2011; Krishnamoorthy et al. 2003; Ting and Macpherson 2005;
Torres-Oviedo and Ting 2010), locomotion (Drew et al. 2008), and seated
reaching (d'Avella et al. 2011; Muceli et al. 2010). Although these tasks are, to
some extent, characterized by feedforward muscle activity, an identification of
the synergies produced in the postural muscles during reaching in the standing
position is lacking. Such a characterization of the existence and nature of the
muscle synergies in a postural behaviour that is assumed to be entirely
feedforward in nature would provide us with a means of deducing whether
feedforward and feedback postural behaviours share similar organizational
principles despite their different modes of control.
Therefore, the aim of the present study was to record the postural
adjustments preceding reaching movements of the preferred arm in multiple
directions, so that their spatial organization can be examined, and to
characterize the postural muscle activity using a technique that has been used to
derive synergic relationships between muscles. This experimental set up,
described in detail in Leonard et al. (2009) was chosen as it is known to evoke
spatially-tuned postural muscle activity in the lower limbs. The paradigm was
used therefore to explore whether a muscle synergy control structure can
explain the variability of postural muscle activity when subjects performed
reach movements during standing in multiple directions. Based on our previous
study characterizing feedforward postural adjustments during multidirectional
reaching during stance (Leonard et al. 2009), we hypothesized that postural
muscle activity preceding the onset of goal-directed reaching movements could
be explained by a few muscle synergies.

99
5.4 METHODS
5.4.1 Subjects
Nine subjects (8 male, 1 female), free from any known neurological,
visual or orthopedic disorders were recruited from the McGill University
student population to participate in the present study. All subjects (23.2 ± 5.2
(SD) years, a mean height of 1.69 ± 0.05 m, and a mean weight of 69.2 ± 4.3
kg) were right-hand dominant. They gave their informed consent to participate
and experiments were conducted with the approval of the McGill University
research ethics board. Data from 6 of the 9 subjects is presented here.
5.4.2 Experimental apparatus and set up
The experimental set-up used for this experiment was used previously
and is explained in detail in Chapter 4 (Leonard et al. 2009). In brief, subjects
were centered in a semi-circular target light array containing 13 targets, and
performed multidirectional reaching movements with their right (preferred) arm
while standing. Following a variable delay, one of the 13 target lights,
presented in random order, would illuminate. Subjects were required to point
and depress a target switch at their natural movement speed. Upon contact with
the target, subjects released the switch immediately and stood with their arm
outstretched at the target for a duration of 2 seconds (see Fig. 4.1B). Subjects
were presented with 15 trials to each target and 15 trials where no target
illuminated (‘catch’ trials), presented in a pseudorandom order. Trials where no
target illuminated or ‘catch’ trials were included during data collection to
minimize the anticipation of the illumination of a target light, but were
excluded from further analysis. In total, 195 trials (15 trials x 13 directions) for
each subject were retained for processing and analysis.
Postural adjustments preceding the onset of the reaching movements
were quantified in terms of kinematic, kinetic (force) and EMG activity. The
data analysis presented in this study was restricted to the activity of the
following 14 postural muscles, recorded bilaterally: tibialis anterior (TAl and

100
TAr, respectively), soleus (Soll, Solr), lateral gastrocnemius (GasLl, GasLr),
peroneus longus (Perl, Perr), biceps femoris (BFl, BFr), rectus femoris (RFl,
RFr), and tensor fascia latae (TFLl, TFLr). In addition, anterior and posterior
deltoid muscle activity was recorded at the right shoulder. However, this EMG
activity was excluded from the non-negative matrix factorization (NNMF)
analysis (described in detail below). Raw EMG data were high-pass filtered at
35 Hz, de-meaned, rectified, and low-pass filtered at 40 Hz using custom-
written programs in Matlab (Mathworks, Natick, MA).
5.4.3 Data processing and analysis
The mean activity of each postural muscle was calculated during five 50
ms time bins preceding the onset of the reaching movement, as determined
from the release of a chest switch attached to the subject’s xiphoid process (for
details of chest switch, see Chapter 4). The EMG amplitude values from all
muscles were then assembled into a single, large data matrix, where each row
represented a vector of data from a single muscle for all conditions (time bin,
reach direction and trial). Each vector was organized as follows: 5 time bins x
13 directions x 15 trials for a total of 975 data points, or conditions, for each
muscle. For display purposes, each muscle was then normalized to its maximal
activity across all time bins, pointing directions, and trials so that values fell
between 0 and 1. Normalized muscle activity as a function of reaching direction
was plotted to create ‘EMG tuning curves’ (Fig. 5.2). To ensure that each
muscle was uniformly represented across all conditions and that variations in
muscle activity were weighted equally by the NNMF algorithm, each muscle
vector was normalized to have unit variance. This means that the sum of the
squared values for each row, or muscle, was equal to zero (Ting and Chvatal
2010).
5.4.3.1 The non-negative matrix factorization technique
Non-negative matrix factorization (NNMF) has been identified as a
useful computational tool for examining the coordination between a large

101
number of muscles and biomechanical variables to test hypotheses about how
the CNS might reduce the dimensionality of control (Ting and Chvatal 2010;
Tresch et al. 2006; Tresch and Jarc 2009). It is a linear decomposition technique
that assumes that a data set constitutes a linear combination of a smaller number
of elements (Lee and Seung 1999). While this principle is not new to
computational models of sensorimotor control, NNMF differs from other linear
decomposition techniques, such as PCA or ICA, in that it constrains the
identified elements to fall within a non-negative space (Lee and Seung 1999;
Ting and Chvatal 2010).
When analyzing muscle activation patterns with NNMF, it is assumed
that the observed pattern of muscle activity is the linear sum of a f(Cheung et
al. 2005)ew (Nsyn) muscle synergies (W), each activated by a synergy
recruitment coefficient (c), (see Fig 1A, Ting and Macpherson 2005; (Torres-
Oviedo et al. 2006; Torres-Oviedo and Ting 2007; 2010; Tresch et al. 2006;
Tresch et al. 1999). Using this formulation, the predicted muscle activity (M*)
can be expressed mathematically as:
M*(t) = ∑ci(t)Wi + error
ci ≥ 0 Wi ≥ 0
error =∑∑(Eij - E*
ij)2
where Wi represents the spatial pattern of muscle activity defined by
synergy i, and ci defines the recruitment of synergy i.
A spatially-fixed muscle synergy (Wi) defines a group of muscles that
are co-activated with fixed spatial scaling. Each element, or muscle, of the
vector Wi, has a fixed value between 0 and 1 for all trials, where 1 represents
the maximal level of activity. The synergy, Wi, is recruited across conditions by
a scalar, non-negative value, ci. This recruitment coefficient, ci, is hypothesized
to reflect the neural signal that activates a group of muscles with fixed relative
amplitude ratios defined by Wi. The tuning of Ci defines how the activation of
synergy Wi is modulated as a function of reaching directions and time (Chvatal
et al. 2011; Safavynia and Ting 2011; Torres-Oviedo and Ting 2007). In
summary, the spatial organization, or composition of each synergy, Wi, remains

102
fixed while the recruitment coefficient, ci, may vary in time (Safavynia and
Ting 2011). It is hypothesized that ci represents the neural command that
defines how the muscle synergy is modulated with respect to time and reaching
direction (Ting 2007; Ting and Mckay 2007).
5.4.3.2 Assumptions of non-negative matrix factorization
The assumptions of NNMF are reviewed in detail elsewhere (Ting and
Chvatal 2010). Briefly, a fundamental assumption of NNMF is one of non-
negativity; that is, the weights ci and the components Wi are constrained to be
non-negative (Lee and Seung 1999; Ting and Chvatal 2010). NNMF is a parts-
based decomposition technique; consequently only additive features of the data
set are represented. In the context of the analysis of muscle activity, each part
identified must resemble features of the overall observed muscle activity (Lee
and Seung 1999; Ting and Chvatal 2010; Tresch et al. 2006). As a result of the
non-negative assumption, it is not possible to identify correlated inhibition
patterns between individual muscles (Tresch et al. 2006). However, decreases
in the level of activation of a synergy are interpreted as the inhibition of that
synergy in relation to the other synergies (Torres-Oviedo and Ting 2007). Thus,
relative inhibitory patterns can be examined. When applying NNMF to analysis
of muscle activity it is assumed that (1) any given muscle may be recruited by
more than one synergy (Torres-Oviedo and Ting 2007; 2010) and (2) relative
levels of muscle activation are fixed within a synergy (i.e. spatially-fixed
synergies, (Safavynia and Ting 2011; Ting and Macpherson 2005).
5.4.3.3 Extraction of muscle synergies for feedforward postural adjustments
Using NNMF, spatially fixed muscle synergies (W) and their activation
coefficients (c) were extracted from a data matrix containing the muscle activity
recordings of 14 postural muscles for the pPA period only. The data matrix was
constructed with dimensions m x s, where m is the number of muscles and s the
number of samples (number of time bin x reaching condition x number of

103
trials). For each subject, the components of the data set, W and c, were found
by iterating a number of random elements via a search algorithm (nnmf
function in MATLAB) until a sufficient proportion of the variability in the data
set (at least 75%) was accounted for (Chvatal et al. 2011; Ting and Macpherson
2005). Specifically, the NNMF algorithm randomly selects non-negative
matrices W and c and modifies their composition to minimize the sum of
squared errors between the actual data (M) and reconstructed data (M* = W x
c). This technique has been used successfully to extract muscle synergies for
feedback postural adjustments in cats (Ting and Macpherson 2005; Torres-
Oviedo et al. 2006) and humans (Chvatal et al. 2011; Safavynia and Ting 2011;
Torres-Oviedo and Ting 2007; 2010), as well as in frog scratching, swimming,
and jumping (Cheung et al. 2005; Tresch et al. 1999).
5.4.3.4 Determining the appropriate number of muscle synergies (Nsyn)
The appropriate number of synergies (Nsyn) was established for each
subject by iterating the value of Nsyn from 1 to 14 and determining the least
number of synergies that could adequately reconstruct the muscle activity
observed in each time period and each trial. Goodness of fit between the actual
EMG and reconstructed EMG was quantified using a measure of the variance
accounted for (VAF) in each muscle vector. VAF was defined as 100 x
uncentered Pearson’s correlation coefficient (Torres-Oviedo et al. 2006; Zar
1999).
Several criteria for VAF were used for determining the correct number
of synergies necessary to explain the trial-to-trial and inter-trial variability in
postural muscle activity observed across all time bins and reach directions for
feedforward postural responses. First, a minimum Nsyn was selected when a
global criteria of >90% VAF for the entire data set (overall VAF) was met.
Second, additional local criteria were imposed to ensure that all features of the
data set were reproduced by the extracted synergies. These local criteria were
quantified when only a portion of the entire data set (60% of all pPA trials)
were considered. Specifically, muscle VAF and condition VAF were

104
quantified. Muscle VAF is a measure of how well the extracted synergies
explained the activity of an individual muscles across all time bins, reach
directions and trials, whereas condition VAF considered the activity of the
entire set of recorded EMG for all trials in a single reach direction and time bin
(Chvatal et al. 2011). These local criteria were met when the Nsyn was able to
account for at least 75% of the muscle VAF and condition VAF. Thus, Nsyn
was increased until all criteria were met as a minimum, and further increased if
additional muscle synergies drastically improved local fit criteria. However, if
an additional muscle synergy contributed evenly to the VAF across muscles and
reaching directions, it was assumed to represent noise in the data set and was
therefore not included (Chvatal et al. 2011; Ting and Chvatal 2010; Torres-
Oviedo and Ting 2007).
5.4.3.5 Muscle synergy analysis
The robustness of the extracted muscle synergies was evaluated by
examining how well the muscle synergies extracted from a portion of the data
set (control data, CTRL trials; 60% of all pPA trials) was able to predict the
measured EMG in the remaining trials (remaining 40% of all pPA trials).
Specifically, goodness of fit between predicted and observed EMG was
quantified with centered (r2) and uncentered (VAF) Pearson correlation
coefficients (Chvatal et al. 2011; Safavynia and Ting 2011; Torres-Oviedo and
Ting 2010). These similarity metrics are both defined as the (sum of squared
errors)/(total sum of squares) (Torres-Oviedo et al. 2006; Zar 1999). However,
r2 is calculated with respect to the mean whereas VAF is determined with
respect to zero. As such, r2 compares the shape of two curves without
considering the amplitudes of the curves, whereas VAF considers both shape
and intercept between two data sets. VAF is therefore considered more
stringent, as both shape and intercept must be very similar for high VAF to be
achieved (Safavynia and Ting 2011; Torres-Oviedo et al. 2006).

105
5.5 RESULTS
The results showed that directionally-tuned pPAs that preceded reaching
movements in multiple directions executed during stance could be explained by
a modular organization of muscle activity. Specifically, across all subjects, the
spatial, temporal and inter-trial variability of the postural muscle activity could
be reproduced by between 5 to 8 muscle synergies. Moreover, consistent
muscle synergies extracted from CTRL trials were able to accurately reproduce
the muscle tuning curves as a result of modulation of the synergy recruitment
coefficient (C).
5.5.1 Feedforward postural muscle activity is directionally tuned, but
shows variability between trials
Reaching movements performed in the standing position to targets
located beyond reach and in multiple directions in the horizontal plane were
preceded by postural muscle activity in the supporting limbs. This is shown for
three principal directions of reach (0º, 90º and 180º) in Figure 5.1. Reaching
movements were preceded by direction-specific modulation of EMG in the
supporting limbs occurring as early as 150 ms prior to the onset of the
movement. For rightward targets (e.g., 0º, see Fig. 5.1A), pPAs were
characterized by inhibitory activity in the right limb (Solr and Perr) and
activation of the extensors in the left limb (TFLl and GasLl). Centrally oriented
reaching movements (90º, see Fig 5.1B) showed pPAs to have initial inhibition
of the ankle extensors (principally the soleus muscles) followed by activation of
the flexors (bilateral TA and TFL). Finally, targets located left of the midline
(180º, see Fig 5.1C) involved activation of extensor activity under the right foot
(Solr, BFr and TFLr) during the pPA period.

106
Figure 5. 1 Representative traces of muscle activity in seven muscles recorded bilaterally
during a 500 ms period preceding movement onset (Movt On) to movement end. The pPA
period is indicated by the shaded vertical grey area to the left of movement onset.
The sensitivity of muscle activity with respect to reaching direction and
period of the pPA was quantified by examining the mean activity over a 50 ms
period for five time bins of 50ms each. Figure 5.2 shows tuning curves (mean
trace of 15 trials across directions and the variability across each of the trials)
for one subject (S011). Clear directional tuning of muscle activity occurred in a
number of muscles and became evident 150-100ms before movement onset. For
active muscles, maximal activity occurred for a small range of reaching
directions. While similarities in the shape of the tuning curves were observed
between some muscles, such as TAl and Perl, significant variability in the
timing and amplitude of muscle activity was observed across trials and reaching
directions as shown by the large standard deviations observed within a reaching
direction.

107
Figure 5. 2 Muscle tuning curves for all 14 muscles and all 5 time bins during the 250 ms period preceding the onset of reaching movements in a representative subject (S011).

108
5.5.2 Composition and tuning of muscle synergies
For all subjects, a small number of muscle synergies were sufficient to
reproduce the spatial and temporal patterns of preparatory postural muscle
activity required for initiating reaching movements in multiple directions. The
minimum number of muscle synergies able to account for at least 90% of the
variability of the entire data set (Fig. 5.3A) and at least 75% of the variability in
each muscle (Fig. 5.3B, C) across reach conditions (Fig. 5.3D) and period of
the pPA was chosen (Fig. 5.3E). A range of 5 to 8 muscle synergies/subject was
found to sufficiently reconstruct the EMG patterns across all trials and reaching
conditions, and accounted for 93.33 ± 1.4 of the total VAF.

109
Figure 5. 3 Variability accounted for (VAF) for different number of muscle synergies for the
entire data set for a representative subject (S010). To choose an appropriate number of muscle
synergies (Nsyn), the following criteria were met: (A) Overall VAF attained a threshold of >
90%; (B) VAF by muscle as a function of muscle synergy number. (C) VAF by synergy
number as a function of muscle was plotted to confirm that chosen Nsyn met the criteria of
75%. (D) VAF by direction and bin shows that directions were well characterized.

110
Each muscle synergy (Wi) specified the relative activation levels for the
muscles hypothesized to be activated together by a neural command (Fig. 5.4).
Figure 5.4 shows the composition of the muscle synergies and the temporal
modulation of the synergy activation coefficients, which were directionally-
tuned to the reaching direction. Several muscles were activated by more than
one synergy, although their relative level of activation differed. For example,
Perll was strongly activated by W4 for rightward targets (Fig. 5.4, yellow
synergy) where it likely serve to produce eversion of the left foot and assist in
pushing the body towards the right. However, Perll was also activated for W2
(Fig. 5.4, red synergy) for forward directed targets. Generally, muscle synergies
were composed of muscles from both the left and right limb (W1, Fig. 5.4).
However, groupings were typically more heavily weighted to a single limb
according to muscle function, rather than consisting of an equal distribution of
muscles of the left and right limbs (ie, equal contributions from TAr and TAl
are not observed). For example, for subject S010 represented in Figure 5.4, 6
muscle synergies were selected. During pPA1 and pPA2, the W1 (Fig 5.4, blue
synergy) consisting of extensors Solr, Soll and GasLr (see left side histograms)
were activated, likely providing antigravity function for quiet stance.
Interestingly, the contribution of this W1 (blue) reduced as the pPA period
progressed and quiet stance changed to a preparation for movement onset.
At the bin pPA4, the most obvious changes in recruitment (tuning
curves) illustrate how the synergies were modulated according to reach
direction. The W2 synergy (red histograms, left side Fig. 5.4) subserved
forward reaching targets mainly, being composed of bilateral TA, Per and some
RF. Leftward targets (105º - 180º) involved activation of W3 mainly (pink
synergy; Fig. 5.4) which consisted of RFr, TFLr and Soll mainly. However,
additional contribution from the extensor synergy (W1) was also seen for these
targets. In contrast, targets located to the right of the midline (0º - 75º) recruited
Perl, TFLl, Gasl and Soll (yellow synergy, Fig. 5.4). Initiation of reaches
towards a smaller range of rightward targets (45º - 75º) also recruited some

111
extensor activity in the left (RFl, Soll) and right (BFr and Gasr) legs (magenta
synergy; Fig. 5.4). Finally, a broadly tuned synergy, W6 (green, Fig. 4),
dominated by BFl showed no clear tuning across direction for subject S010 but
may have played a role in the control of body position during the pPA period,
as the other muscles were activated for movement preparation.
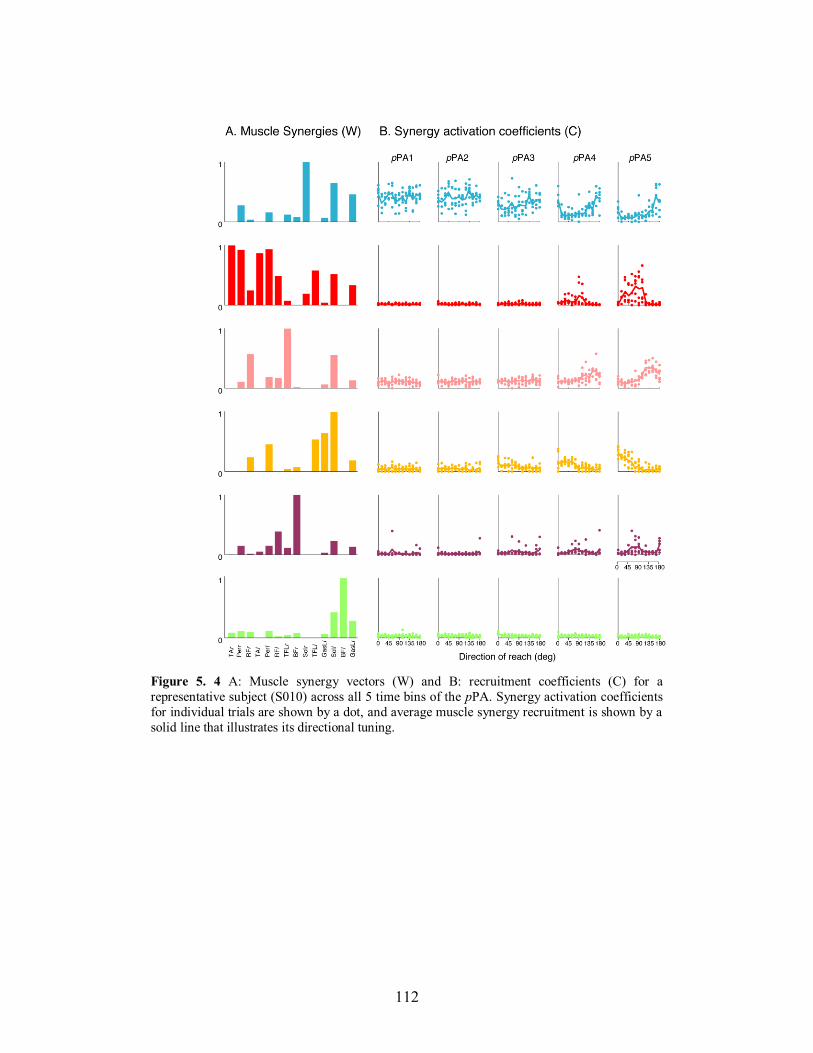
112
Figure 5. 4 A: Muscle synergy vectors (W) and B: recruitment coefficients (C) for a
representative subject (S010) across all 5 time bins of the pPA. Synergy activation coefficients
for individual trials are shown by a dot, and average muscle synergy recruitment is shown by a
solid line that illustrates its directional tuning.

113
5.5.3 Muscle synergies accurately predict muscle activity patterns
To validate the extracted synergies, EMG patterns from REM trials
were reconstructed from the synergies extracted from the CTRL trials. The
reconstruction of EMGs for a representative subject is shown in Figure 5.5. For
this particular subject, the linear summation of 6 muscle synergies could
account for the variation in muscle activity across temporal bins of the pPA and
reach direction. Mean muscle tuning curves demonstrate that the predicted
muscle activity (solid black line) and the recorded EMG (dashed line) for each
postural muscle was well represented, as indicated by the significant goodness
of fit values (Fig 5.5). The mean VAF across all muscles in the pPA5 period for
S010 was 88.84 ± 8.18 (SD). The contribution from each muscle synergy is
depicted with different colored lines. For most muscles, overall activity was
achieved with contributions from mainly from one or two synergies, which is
particularly evident in later bins pPA4 and pPA5.
Muscle activation patterns varied from trial to trial, even within a single
reaching direction, as reflected by the variability in the muscle tuning curves.
The differences in activation could be explained by changes in the activation
coefficients of the muscle synergies, rather than modifications in the muscle
synergy structures (Fig. 5.6). Figure 5.6 shows the reconstruction of pPA4 and
pPA5 of the 6 REM trials for 75° reaching movements for S010 using the
muscle synergies extracted from the CTRL trials (muscle synergies shown in
Fig 5.4). Histograms depict the overall predicted muscle activity of each muscle
as well as showing the relative contribution of each synergy to the muscle’s
activity. Comparison with the actual muscle activation amplitude (indicated by
the stars) shows good predictions by the muscle synergies. Mean VAF across
all trials for pPA4 and pPA5 was 92.19 (±0.78 SD) and 92.66 (±0.77 SD),
respectively.

114
Figure 5. 5 Reconstruction of mean muscle tuning curves using the muscle synergies shown in
Fig. 5.4 for a representative subject (S010). Dashed lines represents observed data and solid
lines represent reconstructed data. Each muscle synergy’s contribution is shown by the
corresponding colored line. Combined, these result in the total reconstruction. Goodness of fit
(VAF, r2) of the reconstruction to the observed EMG is indicated.
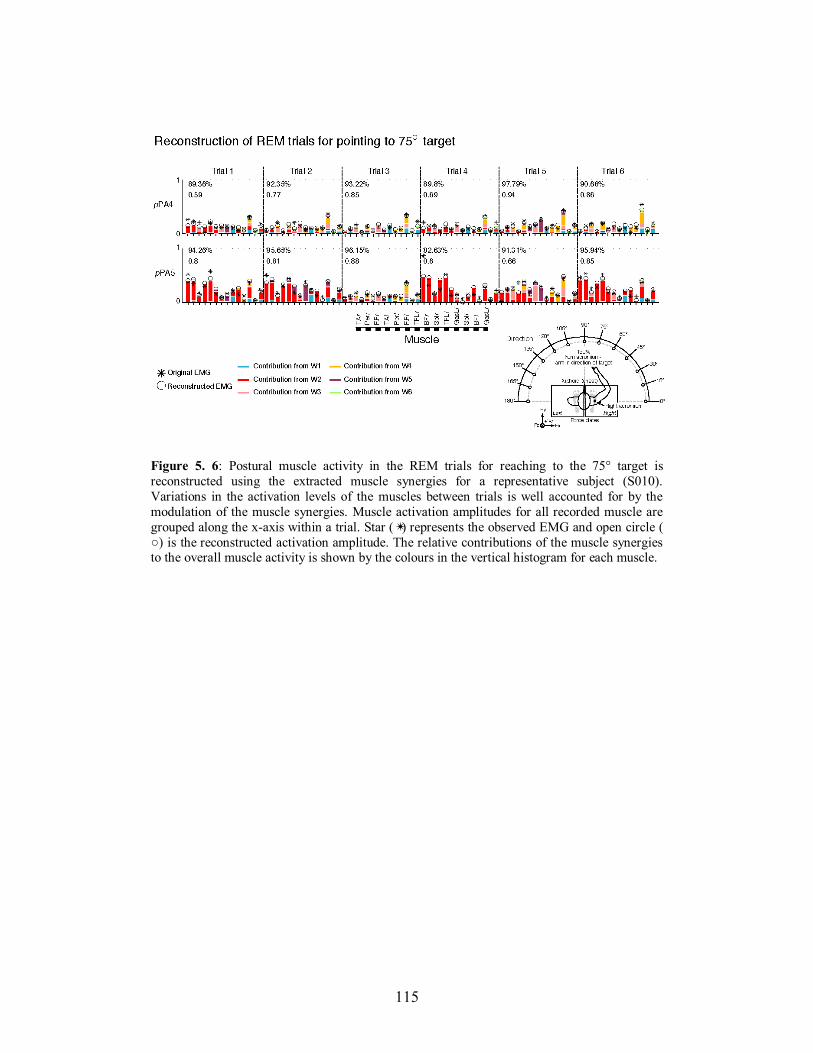
115
Figure 5. 6: Postural muscle activity in the REM trials for reaching to the 75° target is
reconstructed using the extracted muscle synergies for a representative subject (S010).
Variations in the activation levels of the muscles between trials is well accounted for by the
modulation of the muscle synergies. Muscle activation amplitudes for all recorded muscle are
grouped along the x-axis within a trial. Star (✴) represents the observed EMG and open circle (
○) is the reconstructed activation amplitude. The relative contributions of the muscle synergies to the overall muscle activity is shown by the colours in the vertical histogram for each muscle.

116
5.5.4 Comparison of muscle synergy structure between subjects
Across the subjects analyzed, the number of muscle synergies extracted
ranged from a minimum of 5 to a maximum of 8 muscle synergies across all 14
recorded muscles in the pPA period. A comparison of these subjects is shown
in Figure 5.7 and reveals significant variability in the composition of the muscle
synergies between subjects. Across the subjects presented here, only 2-3 muscle
synergies were shared between subjects (Fig. 5.7). Furthermore, 2 subjects
demonstrated completely unique patterns of muscle synergy organization.

117
Figure 5. 7 Muscle synergy structure compared between subjects. Muscle synergies that are
shared between subject are indicated by a significant VAF and r2. Muscle synergies whose
backgrounds are shaded gray are specific to that subject.

118
5.6 DISCUSSION
The primary objective of the present study was to perform a
factorization analysis upon a large set of postural muscles that were activated
using an entirely feedforward mode of postural control. By doing so, it
attempted to identify whether postural muscles were recruited as synergic
groups, as previously documented for feedback postural behaviours (Chvatal et
al. 2011; Safavynia and Ting 2011; Ting and Macpherson 2005; Torres-Oviedo
et al. 2006; Torres-Oviedo and Ting 2007; 2010). Using a well documented and
proven NNMF method (Chvatal et al. 2011; Lee and Seung 1999; Safavynia
and Ting 2011; Ting and Macpherson 2005; Torres-Oviedo et al. 2006; Torres-
Oviedo and Ting 2007; 2010; Tresch et al. 2006), the present study found that
5-8 muscles synergies could account for the spatial and temporal patterns of
muscle activity in the supporting limbs prior to the onset of reaching
movements in multiple directions in the horizontal plane. This study extends
the results of our previous one documenting directionally-tuned muscle activity
for feedforward postural control (Leonard et al. 2009) and suggests that the
CNS could rely upon shared motor modules for simplifying the complex task of
balance control.
5.6.1 Modular organization of feedforward postural adjustments
This study has quantitatively defined groups of muscles recruited
together to serve the coordination of preparatory postural activity preceding
multidirectional reaching movements performed in standing. The variability in
the EMG across reach directions and across trials within a single reach
direction was explained by changes in the synergy activation coefficients, rather
than changes in the synergy structures. This supports the hypothesis that the
CNS relies on a modular structure for organizing motor behaviour (Cheung et
al. 2005; Ting and Macpherson 2005), whereby a motor command defines the
spatial and temporal recruitment of a synergy (Drew et al. 2008; Ting 2007).

119
These findings are consistent with several studies that have shown evidence for
muscle synergies in a variety of tasks (Cappellini et al. 2006; Cheung et al.
2005; Chvatal et al. 2011; d'Avella and Bizzi 2005; Hart and Giszter 2004;
Ivanenko et al. 2005; Safavynia and Ting 2011; Ting and Macpherson 2005;
Tresch et al. 1999).
Our findings are in agreement with previous studies that have shown a
modular organization of postural muscle activity associated with voluntary
movements (Krishnamoorthy et al. 2003; Krishnamoorthy et al. 2004). In these
studies, 3 - 5 principle components (PC) were identified to reflect muscle
synergies, or M-modes, related to consistent directions of force production
(Krishnamoorthy et al. 2003) and recruited according to stability of the support
surface (Krishnamoorthy et al. 2004). Fewer synergistic groups of muscles
were found in these studies as compared to our results for multidirectional
reaching movements. Likely, the differences in the number of synergies
recruited were due to differences in the list of muscles recorded or the task
constraint. Here, we asked subjects to perform reaching movements in 13
directions, whereas in the studies of Krishnamoorthy and colleagues,
perturbations resulting from the movement were limited to the sagittal plane
(Krishnamoorthy et al. 2003; Krishnamoorthy et al. 2004). The methods used in
these studies do not extract muscle synergies in the same manner as those used
here, and therefore the differences in the number of extracted synergies could
be due to this, although there does not appear to be an advantage of using one
methodology over another (Tresch et al. 2006). We specifically chose NNMF in
order to draw comparisons with the organization of feedback-based postural
responses documented in the literature and examine whether similarities for
feedback and feedforward control, as discussed in the next section.
A question raised by our study is how the muscle synergies relate to the
biomechanical parameters of the feedforward postural adjustment, such as end-
point force or motion of the CoM. In balance control, functional muscle
synergies have been analyzed in relation to the end-point forces exerted at the
ground (Krishnamoorthy et al. 2003; Ting and Macpherson 2005; Torres-

120
Oviedo et al. 2006) as well as motion of the CoM (Chvatal et al. 2011). These
results, along with results from biomechanical modeling studies (Welch and
Ting 2008), suggest that the muscle synergies are modulated by global
variables of balance control related to stabilization of the CoM rather than local
joint changes (Chiel et al. 2009; Lockhart and Ting 2007). Whether this is also
the case for the control of balance during the execution of movements, where
both postural and movement goals must be coordinated, remains to be
determined. To assess the organization of pPAs within the context of a
hierarchical control scheme where CNS relies on muscle synergies to translate
task-level goals into the appropriate muscle activation pattern, an analysis of
functional synergies is required and represents a direction of future research.
5.6.2 Similar organization for feedforward and feedback postural control
The finding that muscle synergies can explain the spatial and temporal
patterns of feedforward postural adjustments extends our previous study that
suggested feedforward and feedback postural control may be recruited via
shared neural pathways in the CNS (Leonard et al. 2009). Here, we
demonstrated that variations in both the temporal and spatial characteristics of
pPAs were accounted for by the modulation of the synergy activation
coefficients, which are thought to reflect the neural command that modulates
the synergy (Ting 2007; Ting and Mckay 2007). This strategy has been
consistently demonstrated for feedback postural control (Ting and Macpherson
2005; Torres-Oviedo et al. 2006; Torres-Oviedo and Ting 2007). Moreover, the
muscle synergies for feedback control are related to endpoint force production
(Ting and Macpherson 2005) and CoM motion (Chvatal et al. 2011), and
remain stable when examined across various biomechanical contexts (Torres-
Oviedo and Ting 2010). Whether this is also the case for feedforward postural
adjustments remains to be investigated.
Our analysis has shown that a similar number of directionally-tuned
muscle synergies to those observed in feedback (Torres-Oviedo and Ting 2007;
2010) can explain the feedforward postural muscle activity for pPAs preceding

121
multidirectional reaching movements performed in standing. Also, feedforward
muscle synergies showed clear directional sensitivity for certain reach
directions. In particular, the synergies were spatially tuned to rightward,
forward or leftward directions. Although direct comparisons to the muscle
synergies extracted for APRs (Torres-Oviedo and Ting 2007; 2010) cannot be
made as different muscle groups were recorded, several characteristics
generalize between the two modes of postural control. In both pPAs and APRs,
the task involves recruiting postural muscles to accelerate the CoM within the
BoS. Given that a similar number of directionally-tuned muscle synergies are
activated for both tasks, it is possible that the CNS recruits the same motor
modules for controlling the CoM regardless of the mode of control. Indeed,
based on signals recorded in the pontomedullary reticular formation (PMRF)
for both feedback and feedforward control have been identified in the cat
(Schepens et al. 2008), it is proposed that this structure may be responsible for
organizing postural control across the two modes. Therefore, we propose that
the CNS might rely on shared structures in both feedback and feedforward
postural control and that this integration occurs in the PMRF of the brainstem
(Leonard et al. 2009; Schepens et al. 2008).
It is interesting to note that the composition of the muscle synergies for
feedforward postural control exhibited greater variability between subjects than
those observed for APRs, which were also extracted using NNMF (see Fig 7 in
Torres-Oviedo and Ting 2007). Generally, for feedback balance control tasks,
such as restoring perturbed balance, several (approximately 4) muscle synergies
are shared between human subjects and exhibit consistent similarities in the
tuning of their activation coefficients (Torres-Oviedo and Ting 2007; 2010),
suggesting that the modular organization reflects the underlying neural control
(Ting 2007). Here, we show instead that it was an exception for muscle synergy
structure to be shared across subjects. It is hypothesized that descending
commands from higher neural centers recruit muscle synergies (Roh et al.
2011), therefore, given the voluntary nature of the task, subjects may have
adopted different strategies to perform the task. In this present experiment, no

122
instruction regarding movement speed or pattern were given, therefore subjects
were free to select their own movement strategies. Therefore, it is possible that
subjects recruited functionally equivalent synergies that differed in their
composition due to redundancy of their respective biomechanics (McKay and
Ting 2008). Further study is needed to address these differences.
5.6.3 Conclusions
We have shown that subject-specific muscle synergies explain the
spatial and temporal coordination of postural muscle activity organized in a
feedforward manner preceding reaching movements in multiple directions. In
general, 5-8 muscle synergies were sufficient to reconstruct the feedforward
postural EMG for each subject. However, only a small number of synergies
were shared across subjects. It is unclear whether this is related to the voluntary
nature of the task or whether subjects recruit distinct synergies that may be
functionally equivalent due to redundancy of the musculoskeletal system
(McKay and Ting 2008). These results are consistent with published
observations that muscle synergies represent a modular organization of motor
control, although they fall short of explaining how the synergy recruitment can
explain the force constraint strategy documented in our previous study
(Leonard et al. 2009). Future studies will examine how the synergic muscle
activity relates to the biomechanics of feedforward postural control. While
these results show similar features in number of synergies and spatial
organization to those observed for APRs, robustness of muscle synergies for
feedforward posture control remains to be determined by evaluating whether
the same synergies are preserved for postural adjustments accompanying
reaching movements. Overall, these findings provide a first step in examining
whether a set of robust muscle synergies exists for feedforward and feedback
postural control.

123
5.7 ACKNOWLEDGEMENTS
We thank Dr. Seyed Safavynia (Wallace H. Coulter Dept. of
Biomechanical Engineering, Georgia Inst. of Technology and Emory Univ.) for
advice about data analysis.

124

125
Chapter 6
Postural adjustments for online corrections
of arm movements in standing humans
6.1 PREFACE
The previous two chapters examined the postural strategies and
coordination of muscle activity for feedforward postural adjustments preceding
and accompanying goal-directed reaching movements. In these studies, the final
goal of the movement was planned and the disturbances associated with the
displacement of the limb and body towards the target could be predicted.
Consequently, pPAs and aPAs could be planned in feedforward accordingly
(Bouisset and Zattara 1981; 1987). However, in situations where the target goal
may change once the movement is initiated, the planned postural activity is no
longer appropriate for the updated focal movement. It is not known whether the
CNS updates posture predictively, or whether it relies on feedback from the
moving limb to make the correction in the posture. Therefore, in Chapter 6, I
address this question by determining the nature of the aPA command in relation
to an online correction of reaching movement executed in standing. This is the
first study to specifically examine the online control of posture using a
paradigm of visual perturbations of reaching movements performed in standing.
This chapter was adapted from Leonard JA, Gritsenko V, Ouckama R
and Stapley P.J. Postural adjustments for online corrections of arm movements
in standing humans. Journal of Neurophysiology, 105(5): 2375-2388, 2011.
This manuscript has been reprinted with permission from The American
Physiological Society, publisher of the Journal of Neurophysiology. The paper
is presented in the same format in which it was published with the exception of

126
formatting to figures and tables to comply with McGill University thesis
formatting guidelines.
6.2 ABSTRACT
The aim of this study was to investigate how humans correct ongoing
arm movements while standing. Specifically, we sought to understand if the
postural adjustments in the legs required for online corrections of arm
movements are predictive or rely upon feedback from the moving limb. To
answer this question we measured online correction in arm and leg muscles
during pointing movements while standing. Nine healthy right-handed subjects
reached with their dominant arm to a visual target in front of them and aligned
with their midline. In some trials the position of the target would switch from
the central target to one of the other target locations 15°, 30° or 45° to the right
of the central (midline) target. For each target correction, we measured the time
at which arm kinematics, ground reaction forces, and arm and leg muscle EMG
significantly changed in response to the target displacement. Results show that
postural adjustments in the left leg preceded kinematic corrections in the limb.
The corrective postural muscle activity in the left leg consistently preceded the
corrective reaching muscle activity in the right arm. Our results demonstrate
that corrections of arm movements in response to target displacement during
stance are preceded by postural adjustments in the leg contralateral to the
direction of target shift. Furthermore, postural adjustments preceded both the
hand trajectory correction and the arm muscle activity responsible for it, which
suggests that the CNS does not depend upon feedback from the moving arm to
modify body posture during voluntary movement. Instead, postural adjustments
lead the online correction in the arm the same way they lead the initiation of
voluntary arm movements. This suggests that forward models for voluntary
movements executed during stance incorporate commands for posture that are
produced based on the required task demands.

127
6.3 INTRODUCTION
When standing humans reach out to point at or grasp an object, the
central nervous system (CNS) must resolve two major task constraints: the
production of the correct hand and arm trajectory towards the target and the
necessary associated postural adjustments (aPAs) in the supporting limbs and
trunk for the maintenance of equilibrium. Using prior knowledge of the
dynamics of the moving arm and the internal disturbances that arm movements
inflict upon the body, the CNS can anticipate the impending disturbance and
correctly program the aPAs accordingly. However, what happens when the
final position of a visual target unexpectedly changes after the onset of a
planned voluntary movement that is executed while standing? In this situation,
the CNS must correct online the arm trajectory toward the new target and
modify the required postural adjustments. The objective of this study is to
investigate how posture is modified with respect to arm movements during
visual perturbations of the reaching goal. The online control mechanisms for
arm movements have been extensively investigated during experiments with
seated subjects. These studies have shown that modifications of hand trajectory
in response to target displacements occur at short latencies of 100 to 150 ms
(Day and Lyon 2000; Goodale et al. 1986; Paulignan et al. 1990; Prablanc and
Martin 1992; Prablanc et al. 1986; Soechting and Lacquaniti 1983; Zelaznik et
al. 1983). Other studies have shown that visuomotor corrections are automatic
and occur without voluntary intervention (Day and Lyon 2000; Gritsenko et al.
2009). Because of the inherent delays associated with the use of sensory
feedback, the short latencies of online correction support the notion that the
CNS adopts predictive mechanisms to execute rapid arm movements to visual
targets. Such a prediction involves the formulation of an initial plan of the
movement using a feedforward mode of control, but also a continuous
estimation of the actual state of the system compared to the desired one, which
is achieved using rapid, internal feedback loops (Bhushan and Shadmehr 1999;
Desmurget and Grafton 2000; Mehta and Schaal 2002; Sabes 2000; Shadmehr
and Krakauer 2008). This efference copy-based state estimation ensures that

128
motor commands can be modulated at short latencies and online, automatic
corrections of movement be made without detriment to the resulting movement
(Nijhof 2003; Saunders and Knill 2003).
Anticipatory modulation of muscular activity in the supporting limbs
that precedes the onset of voluntary movements during stance also suggests that
a degree of prediction of the future state of the body occurs in advance of an
impending disturbance to posture (Davidson and Wolpert 2005). The
characteristics of aPAs occurring before the onset of movement depend on the
prior knowledge of arm and body dynamics (Bouisset and Zattara 1981; 1987).
In these paradigms (e.g., arm raising) however, a postural disturbance can be
predicted in advance, and need not be modified online during the execution of
the movement, as the end goal matches that for which the postural adjustments
were initially programmed. Indeed, during reaching to fixed targets in multiple
directions when standing, feedforward postural adjustments follow a consistent
spatial pattern both before and during the movements (Aruin and Latash
1995a). What happens however, when postural adjustments for reaching,
programmed based on an initial state and an expected outcome, must be
modified because of an unexpected change in the visual position of the final
goal? Does the CNS still adopt a predictive mode of control or does it correct
arm trajectory before posture, which is then updated based upon the feedback
obtained from the arm correction? We attempted to investigate these questions
by inducing unexpected shifts in the visual location of the target after the onset
of reaching movements during stance. All targets were placed at a distance such
that subjects could point to them and retain their center of mass (CoM) within
the support base, without the need for a corrective step. Thus, subjects were
aware that they could complete the corrections and not become unstable to the
extent that they would lose balance. Therefore, two mechanisms of postural
control were possible: 1) Postural adjustments occur after the arm movement
corrections for the visual perturbations, and the feedback from the change in
arm and body configuration is used to update the necessary postural
adjustments, or 2) Postural adjustments are predictive and precede online

129
corrections of arm movement. This may indicate that, rather than acting to
reduce the disturbance induced by the upcoming arm correction, the postural
control system participates in the movement component of the action. We
hypothesized that because a change in the trajectory of the arm in response to
an unexpected shift in target position could potentially destabilize one’s
balance, the CNS predicts the postural adjustments necessary to execute smooth
online corrections of arm movements. In other words, when the target shift
occurs, leg muscle activity is updated and precedes the necessary changes in
ground reaction force in advance of changes in arm muscle activity or in
trajectory toward the new target. We show that this is indeed what occurs for
online corrections of arm movements during stance.
6.4 METHODS
6.4.1 Subjects
Nine right-handed subjects (5 females, 4 males) were recruited from the
McGill University student population to participate in the study. Subjects had a
mean age of 22.9 ± 3.1 (SD) years and measured on average 1.68 ± 0.1 m and
62.4 ± 9.6 kg in height and weight, respectively. All subjects were free of any
known neurological, visual, or orthopedic disorders, and provided their
informed consent to participate in this study. The study had ethical approval
from the McGill University research ethics board.
6.4.2 Experimental apparatus and set up
Subjects stood barefoot on two triaxial force plates (model FP4060,
Bertec, Columbus OH) that recorded ground reaction forces (GRFs) and
moments in the mediolateral (x), anteroposterior (y) and vertical (z) axes at
1000 Hz. Each subject stood with their feet positioned according to their
preferred mediolateral stance width, which was based on the average distance
between the 2 heels calculated immediately after 3 trials of walking 15 m across

130
the laboratory. This preferred stance configuration was recorded and marked on
the force plates and subjects maintained it throughout the experiments.
The experimental apparatus consisted of 4 target lights arranged about a
semi-circular radius separated by 15° (see Fig. 6.1A). The apparatus was fully
adjustable to each subject’s reach distance and height. Light targets were 2.5 cm
in diameter and consisted of a 5-V red light-emitting diode (LED) encased in a
modified gaming switch (model 459512; RP Electronics, Burnaby, BC,
Canada) that produced a 5-V pulse upon contact. Targets were mounted on
lightweight aluminum dowels, adjustable in length, affixed to a semicircular
aluminum bar suspended from the ceiling. Subjects wore a chest band with the
same switch that enabled the detection of movement onset upon its release.
Targets were situated at a distance corresponding to 130% of each subject’s
reach length measured to each respective target. Previous experiments adopted
this distance, which was attained comfortably without the centre of pressure of
either foot leaving foot length or width (Leonard et al. 2009). Any trials where
this occurred were rejected from further analysis. The choice of 130% was
especially important as we sought to evoke postural adjustments for online
corrections readily distinguishable above those produced for ongoing (initial)
reaches to the central target. Reach length (100%) was measured as the distance
between the xiphoid process (where the finger tip was held at the start of each
trial) to the tip of their outstretched finger when the arm was extended in the
direction of each of the targets (neutral scapula retraction).
The muscle activity of 16 leg, trunk and arm muscles was sampled at
1000 Hz using two DelSys Bagnoli 8-channel systems (Delsys, Boston, MA).
For all subjects the activity of the following leg muscles was recorded
bilaterally: tibialis anterior (lTA and lrTA), soleus (lSol, rSol), peroneus longus
(lPerl, rPerl), rectus femoris (lRF, rRF), biceps femoris (lBF, rBF) and tensor
facia latae (lTFL, rTFL). Additionally, recordings of muscle activity of the
reaching arm (right) included anterior and posterior deltoid (rADelt and rPDelt,
respectively), long head of the triceps (rTric) and the long head of the bicep
muscle (rBic). Bilateral kinematics were collected using a 6-camera MX3

131
motion-capture system (Vicon Peak, Lake Forest, CA) sampled at 200 Hz. A
custom written program using LabVIEW (National Instruments, Austin, TX)
controlled the illumination of the target lights, and acquired and synchronized
signals from the chest and target switches. Synchronization with the analog
signals obtained from the force plates and EMG system was done using the
Vicon controller.
6.4.3 Experimental procedures
Subjects began each trial standing with the head forwards in the
direction of the central (90°) target, which was aligned along the midline of the
body (see Fig. 6.1A). Before each trial, they depressed the chest switch attached
at the xiphoid process with their right index finger. The left arm was held in a
relaxed downward pointing position at the side of the body. The study
comprised 2 principal types of trials: regular reaches (‘reach’ trials) to the
central (90°) target and trials that required corrections of arm trajectory towards
3 other targets at different times after a ‘reach’ was initiated (online corrections
or ‘corr’ trials).
Subjects were asked to stand quietly before each trial began. Once the
experimenter was satisfied that the subject was stable, he/she initiated data
collection and after a variable delay, the central target light (90° or ‘L1’) would
illuminate. Subjects were instructed to reach and press the illuminated light at
their natural speed (‘reach’ trials, dashed trajectory, Fig. 6.1A), remaining in
that position until told to return (approx. 2 secs). For some trials, the target light
would shift from L1 to any one of the other 3 targets, 75° (‘corr75’), 60°
(‘corr60’), or 45° (‘corr45’) at different times after the onset of the initial reach
movement. All ‘reach’ and ‘corr’ trials were randomly presented as well as
trials when subjects were prepared, but no light illuminated. For the ‘corr’
trials, subjects were instructed to correct arm trajectory when they detected the
light change and point to the newly illuminated target. The target shift could
occur after a variable delay from the online detection of a voltage drop that
occurred when subjects released the chest switch. Figure 6.1B illustrates a

132
representative timeline for a typical corr45 trial at 442 ms after movement
onset. The first light (L1) illuminated approximately 250 ms after the auditory
tone. The subject reacted to L1 onset by initiating a movement of the finger
(‘movt onset’ in Fig. 6.1B). At this time, chest switch voltage dropped to zero.
After 442 ms, L1 turned off and light 2 (L2) illuminated. In this example, the
corr45 movement resulted in a lengthening of movement time
(Balasubramaniam and Turvey) of 322 ms with respect to the mean ‘reach’
movement.

133
Figure 6. 1: Experimental set-up and data collection schema. A. Subjects stood on 2 force
plates reached to a central target, aligned with their xiphoid process. Unperturbed ‘reach’ trials
were interspersed with online correction (‘corr’) trials involving unexpected illuminations of 1
of 3 other targets placed successively at 15° increments to the right of centre. B. Explanation of
the changes in voltage related to the sequence of light changes. When the signal rose to 5V each
light was illuminated. L1 = light one, L2 =light 2, chest = chest switch attached around the
subjects sternum that acted as a signal from which L2 illumination could be triggered. C. A
histogram showing the distribution of L2 onset as a percentage of mean ‘reach’ peak velocity.
Trials from all ‘corr’ conditions have been pooled (n=652). rFin = right finger.

134
A variable delay existed between when the controlling Labview routine
received the switch signal (shift from 5 to 0V indicating movement onset) and
when it simultaneously extinguished L1 and illuminated L2. This was estimated
to be on average 62.1 (± 35.1) ms. Thus, even though standard delays of 200
and 350 ms between movement onset and light (L2) change were used, actual
delays of the light changes were calculated on a trial by trial basis, using the
rising edge of the L2 5V signal (see centre grey line on Fig. 6.1B). In order to
standardize when L2 changes occurred, delays were represented as a percentage
of mean ‘reach’ trial MTs. The distribution of L2 onsets as a percentage of
‘reach’ MT is shown in Fig. 6.1C. Even though light shifts occurred within the
acceleration phase of ‘reach’ trials (open bars), a higher percentage occurred
during the deceleration phase (grey bars).
Each experimental session began with subjects performing a series of
acclimatization trials, consisting of 5 regular reaches to each target in turn.
Following the acclimatization period, trials were organized in a random order.
We required subjects to execute a total of 60 ‘reach’ trials to the central target,
and at least 15 correction trials to each of the 3 other target positions. Fifteen
trials were given during which no target illuminated (catch trials). In some
subjects, more than the standard number of corr trials was collected to ensure
that a large enough database was established. Catch trials were included in an
attempt to prevent subjects from predicting and begin moving to the target
before L1 illuminated. All catch trials were eliminated from subsequent
analysis. For experimental conditions to have adequate controls, at least three
times the number of control trials compared to experimental trials must be
collected (Zar 1999). For our analysis the ‘reach’ trials were to act as controls
for detecting differences in the ‘corr’ trials. Thus, we aimed to collect at least
120 trials from each subject (1080 for all 9 subjects). A breakdown of the total
number of trials collected and retained after trial selection procedures (see Data
analysis) is given in Table 6.1. Subjects performed blocks of 40 trials between
which they would take 5-minute rest periods to reduce fatigue. Each data
collection period lasted 45-60 minutes.

135
6.4.4 Data analysis
Data analyses were performed offline using custom routines written in
Matlab (The MathWorks, Natick, MA). Kinematic data and GRFs were low-
pass filtered using a digital second-order Butterworth filter, with a 10Hz and
100Hz cutoff frequencies, respectively. Raw EMG signals were high-pass
filtered at 35 Hz, demeaned, rectified, and low-pass filtered at 10 Hz, using a
2nd-order Butterworth filter. All trials were visually inspected for stability of
Fz during the background period (500 ms of quiet stance before the first target
light illuminated). Any trials showing significant variation in Fz were
eliminated from further analysis.

136
Figure 6. 2: Determination of the online correction of finger trajectory (fcorrect). A. Plan view (x,y) of rFin average ‘reach’ trajectories +/- 1SD (dashed line with shaded grey area) in relation
to one ‘corr45’ trial (full black trajectory). Filled black circle is the onset of light 2 (L2 onset)
and the open circle is the time at which the corr45 x,y trajectory exceeded the average ‘reach’
trajectory plus 1SD for subject S6. B. Average (dashed line) plus 1SD of curvilinear rFin
velocity for a reach movement and one ‘corr45’ trial(full black line). Black vertical line is light
2 (L2) onset, grey vertical line is the time of online correction (fcorrect). Each corr condition
has been displaced rightwards and downwards for clarity, but the starting position was the same
for each. C, D: Explanation of how the correction of the EMG activity and GRFcorrect
associated with online corrections were determined. C. Calculation of EMGcorrect. The muscle
shown is the left soleus muscle, but the same procedure was used with all other muscles studied
(see Methods). The dashed trace and grey traces represent respectively, the mean ‘reach’ soleus
muscle activity ± 2SDs above and below the mean. The dark full trace represents the soleus muscle activity produced during an online correction movement, in this example a corr45
movement. The open circle indicates the time at which the corr45 soleus muscle activity
exceeded the mean+2SD ‘reach’ activity level. This time was taken as the EMGcorrect time
(for more detailed explanation, see Methods and Results). Abbreviations as previous figures,
except Movt end=movement time. D. Calculation of GRFcorrect. Method for determining
GRFcorrect is shown for the left shear force (Fx). The dashed trace and grey traces represent
respectively the mean ‘reach’ Fx and ± 2SDs above and below the mean. The dark full trace
represents the Fx exerted during an online correction movement (in this example corr45). The
open circle indicates the time at which the exerted force was significantly different from the
mean forces exerted in a ‘reach’ trial.

137
Trials were aligned to movement onset, which was determined on a
trial-by-trial basis to be the time at which the tangential velocity (derivative of x
and y velocity) of the right finger marker surpassed a value of 3% of the peak
velocity in that trial. Movement end was also established as when velocity
reduced below the 3% threshold in that trial. We, and others, have previously
used this threshold value to successfully determine movement onset and end
(Leonard et al. 2009; Shabbott and Sainburg 2009). Correction (‘corr’) trials
were also eliminated if: 1) they showed tangential velocity profiles that did not
illustrate a pronounced ‘trough’ indicating that a significant reacceleration
occurred. In other words, in these trials the subject may have hesitated
sufficiently to execute a reach directly to a ‘corr’ target, or 2) the trough
between the first and second peaks of tangential velocity (see Fig. 6.3B)
descended below the 3% initial peak velocity threshold, indicating that subjects
moved too fast and reached the central target before correcting to the ‘corr’
target.
Once these trial rejection procedures were implemented, a number of
essential measures were determined based on each subject’s average
trajectories. These included: fcorrect (the deviation of a ‘corr’ trajectory from
the mean of all regular reaches), EMGcorrect (the times at which particular
EMG traces during ‘corr’ trials significantly deviated from an average of
‘reach’ trials) and GRFcorrect (times at which forces in the 3 axes significantly
deviated from the average force exerted for ‘reach’ trials). The online correction
of focal movement (fcorrect) was detected on a trial-by-trial basis for all ‘corr’
trials using the tangential velocity of the marker placed on the right index finger
with respect to the original target. First, the mean ±1SD of all ‘reach’ trial x-y
trajectories was computed for a subject (see dashed trajectory and shaded area,
respectively on Fig. 6.2A). Second, the tangential velocity of each ‘corr’ trial
was compared to the mean tangential velocity of all ‘reach’ trials and an
algorithm calculated the time at which the reacceleration of the finger for the
‘corr’ trials (corresponding to the online correction) exceeded 1SD of the mean
‘reach’ trials. This is shown in Fig. 6.2B (fcorrect), occurring for this particular

138
trial at 460 ms, 169 ms after the change in the target light (L2). Similar methods
were used to calculate EMGcorrect and GRFcorrect, but corrections were
detected using a threshold of mean ±2SDs, rather than 1SD for fcorrect
(kinematics). Mean ± 2SDs was found to be more robust for detecting
EMGcorrect and GRFcorrect than mean ± 1SD. An illustration of the
calculation of these variables is given in the relevant section of the Results (see
below). After a stable initial posture, ability to detect significant EMG
corrections in arm or leg muscles and other ‘corr’ trial rejection procedures
were implemented, a total of 652 trials were retained and used for further
analysis for the 9 subjects after the unstable trials were eliminated as well as
those in which the algorithm for EMG or force corrections could not detect a
significant change with respect to regular reaches. Table 6.1 lists the total
number of trials collected and retained in each reaching condition.
A custom-written algorithm detected the time at which the EMG
activity of the muscles identified as participating in the corrections in ‘corr’
trials exceeded that produced for ‘reach’ trials, and values for each ‘corr’ trial
were verified on a trial-by-trial basis. For example, Fig. 6.2C illustrates soleus
muscle activity for the same ‘corr45’ trial as in Figs 6.2A and B (finger
trajectory), and shows how the EMG activity of this muscle increases well
above the mean+2SD of the reach trials, and well before the correction of the
arm trajectory (fcorrect). EMGcorrect was computed for a total of the 3 leg
muscles in the left leg as well as the 3 identified in the right arm (lSol, lTA,
lPerl, rADel, rPDel and rTric) and additionally rBic. A similar algorithm was
used to detect when shear (Fx) and vertical (Fz) force also increased above the
force produced during a regular reach (Fig 6.2D).
6.4.5 Statistical analysis
To detect significant differences in total movement times and fcorrect
values between ‘reach’ and ‘corr’ conditions a one-way analysis of variance
(ANOVA) was used, with experimental condition as the single factor. The
statistical comparison between EMG correction times of arm and leg muscles

139
was done using ANOVA with 4 factors. First, for each individual trial, the
EMGcorrect values of each postural leg muscle were subtracted from the
EMGcorrect values of each arm muscle. Positive differences indicate that leg
muscles change after the target jump before arm muscles. Values (EMGcorrect
of leg muscles minus arm muscle EMGcorrect) were sorted for trials in which
the target jump occurred during the acceleration or deceleration phases of the
initial reach movement. This was done to investigate if the leg/arm muscle
corrections were influenced by the extra time and feedback that may have been
available when target jumps occurred in the acceleration phase. Then ANOVA
was applied to these differences to determine the main effects of the following
factors: Target (corr75, corr60, and corr45), Postural Muscle (lSol, lPerl, and
lTA), Subject (9 subjects), and Phase (acceleration or deceleration of the arm
during the target jump). Post-hoc comparisons were performed with Sidak-
Bonferroni correction for alpha (Abdi 2007). Further post-hoc comparisons
were done using linear regressions between the EMG correction times for arm
muscles deemed as contributing to the online kinematic corrections and fcorrect
values, as well as between postural and arm muscle EMG correction times. For
each type of linear regression we report slope (m, the amount of increase in Y
that accompanies one unit of increase in X) and the Y intercept (Yi), the point
conventionally chosen to define Y coordinates when X=0. Thus, for the
relationship between leg and arm muscle correction times, we sought to
investigate what latency the arm muscle corrected at when the postural muscle
was 0 (or vice versa).
6.5 RESULTS
6.5.1 Unperturbed reaching and characteristics of online corrections
Trials executed to the central 90° target (‘reach’) typically showed an
early phase, up to peak velocity, during which the trajectory curved slightly
rightwards from the midline. This is illustrated by the average trajectory (± 1SD
of the mean) for one representative subject in Fig. 6.3A. The acceleration phase

140
was followed by a longer deceleration phase during which the trajectory curved
inwards towards the target. Trials that required online corrections of trajectory
showed significant deviations (fcorrect) from the reach trajectory, represented
by filled circles on average trajectories to each of the 3 targets. This deviation
became more accentuated as corrections were required further towards the right
target (45°). However, subjects were able to successfully correct their
trajectory, attain the targets and remain balanced.

141
Figure 6. 3: Reaching movement kinematic characteristics and profiles of curvilinear velocity.
Shown are averages plus 1SD for all trials for subject 2 in each of the 4 conditions studied. A.
Plan view (x,y) kinematics of rFin trajectory for ‘reach’ trials and each of the correction
conditions. B. rFin curvlinear velocity also for all 4 conditions. fcorrect=kinematic correction of
finger trajectory, mvt end=end of the focal movement (reach and corrected movements).

142
The curvilinear velocity of the right finger (rFin) marker enabled a very
clear identification of the online correction of the pointing movements
(‘fcorrect’, filled circles). Average velocity profiles for reach, corr75, corr60
and corr45 are shown in Fig. 6.3B for subject S2 (corresponding to the average
trajectories, Fig. 6.3A). Typically, at fcorrect, the finger reaccelerated and
displayed a second peak in velocity before the arm decelerated to the target
(‘mvt end’). The online correction of trajectory (fcorrect) occurred at, on
average 178.1 ± 58.3 ms (corr75), 187.6 ± 58.4 ms (corr60) and 191 ± 48.1 ms
(corr45) after L2 onset (values shown on Fig 6.2B for subject S2 are absolute
values after the onset of the movement). Movement times and fcorrect values
for all ‘corr’ conditions are given in Table 6.1. Movement times increased
between each of the 4 conditions from 815.5 ms (reach) to 1294.6 ms (corr45)
and there was a significant main effect of condition between reach, corr75,
corr60, corr45 (F(3,772)=492, p=0.00). However, there was no significant
difference between the fcorrect times to any of the 3 conditions that required
online corrections (corr75, corr60 and corr45), indicating that, when the whole
range of perturbations was considered, the online corrections were constant
with respect to the onset of the initial reaching movement and L2 onset.

143
Table 6. 1 Breakdown of total number of trials collected and retained after
trial selection procedure.
Condition n Movement Time (ms) L2 onset (ms) fcorrect (ms)
reach 228 877.6 (176.3) n/a n/a corr75 158 1090.7 (141.4) 360.1 (127.4) 567.7 (103.8)
corr60 148 1237.7 (137.8) 348.5 (76.9) 563.5 (87.6)
corr45 118 1318.8 (269.0) 340.1 (91.4) 566.5 (90.1)
Average movement times, light 2 (L2) onset and time to correction of finger
trajectory (the latter 2 for the applicable conditions) for all experimental
conditions tested. Latencies are calculated relative to the onset of the initial
finger (reach) movement. N = Number of trials.

144
6.5.2 Corrective forces and electromyographic activity accompanying
online corrections of arm movements
Typical EMGs and GRFs produced for both a ‘reach’ and ‘corr45’ are
shown in Fig. 6.4. Four arm muscles are shown on the same time scale as 6
bilateral lower limb muscles and the corresponding Fx, Fy and Fz forces.
During a typical ‘reach’ (Fig. 6.4A), the arm movement was initiated by the
activation of the rADel, and also bilateral anticipatory TA activity and Sol
inhibition in the legs. The effect of this postural muscle activity at the
beginning of the movements was to produce a backward directed Fy and a
loading (Fz) of the right foot to induce a forward sway of the body to the target.
Towards the end of the movement, the posterior deltoid and, to a lesser extent
the triceps and biceps muscles, became active. This activation of arm muscles
to brake the arm movement at the target was accompanied by associated
postural adjustments in the extensor muscles of the legs, represented in Fig.
6.4A by bilateral Sol muscle activity, starting approximately 500 ms after
movement onset. Forces showed that during this phase of the movement both
feet exerted force forwards (Fy) and the right foot Fz was loaded. This pattern
of EMG and force activity preceding and during the reach movements was the
same as that described previously (Leonard et al. 2009).
Movements necessitating online corrections of finger trajectory evoked
specific adjustments of both arm and leg muscle activity preceding fcorrect.
The adjustments in the postural muscles were recorded principally in the
muscles of the left limb during the period between light 2 onset (L2) onset and
fcorrect (shaded areas of EMG traces in greyed bar, Fig. 6.4B) and produced
distinct changes in Fx and Fz components of GRF under the left foot, as
compared to ‘reach’ trials (see vertical arrows). The most significant
adjustments in arm muscle activity were evident in the rADel, rPDel and rTric.
In the postural muscles, significant activations between L2 onset and fcorrect
occurred in the lSol, lTA and lPerl. During approximately the same period of
‘reach’ trials, no such postural or arm muscle activity could be seen (see Fig.

145
6.4A, ‘approx. area of fcorrect’). The postural adjustments from L2 to fcorrect
recorded during online corrections led to an increase in leftward-directed shear
force (Fx), and a reloading of Fz also under the left foot. Analysis of the times
to correction of arm and leg muscles corresponding to the online correction of
movement, and their relationship was therefore principally limited to this subset
of left side postural muscles (lSol, lTA and lPerl) and the 3 right arm muscles
(rADel, rPDel and rTric).

146
Figure 6. 4 Typical arm and leg muscle activity in relation to the 3D ground reaction forces
produced for a ‘reach’ movement (A) and an online correction movement to the target placed
45° to the right of midline, ‘corr45’ (B). In each, the muscles plotted in grey are those recorded
in the right leg. The vertical dashed line indicates initial movement onset (‘Mvt Onset’) and the full vertical black line, movement end (‘Mvt End’). In B., the shaded grey area indicates the
area in which arm and postural adjustments occurred. For muscle abbreviations, see Methods.
L2 onset=light 2 onset, fcorrect=time of kinematic correction of the rFin marker.
Fx=mediolateral force, Fy=anteriorposterior force and Fz=vertical force.

147
6.5.3 Arm-muscle activity responsible for corrections of finger trajectory
Of the right arm muscles recorded, we investigated which were related
to the online correction of finger trajectory. Figure 6.5 shows linear correlations
calculated between the latency of onset of fcorrect and EMGcorrect of each of
the recorded arm muscles (rPDel, rTric, rADel and rBic). Of these 4 muscles, it
can be seen that 3 (rPDel, rTric and rADel) showed highly significant
correlations between the onset of the corrective muscle activity of the arm and
the correction of curvilinear finger trajectory (fcorrect). The corrective muscle
activity in these 3 muscles preceded fcorrect by average values of -104.5 ms
(rPDel), -101.8 ms (rTric) and -30.4 ms (rADel), as determined by the intercept
of the regression lines. Therefore, based upon this, we sought to determine if
the left leg postural activity evoking changes in shear (Fx) and vertical force
(Fz) that reoriented the body towards the new target during online corrections,
preceded or not the corrective arm muscle activity in each of the 3 right arm
muscles that were correlated to fcorrect (rPDel, rTric and rADel).
6.5.4 Corrective postural adjustments in leg muscles lead arm muscle
corrections during online corrections of arm trajectory to
unexpected shifts of target position
Most of the EMGcorrect times in the postural leg muscles identified as
participating in the online corrections during ‘corr’ trials (lSol, lPerl and lTA)
were shorter than those in the reaching arm muscles related to fcorrect (rPDel,
rADel and rTric; Fig. 6.5). The average GRFcorrect values for Fx preceded
fcorrect by approximately 80-85 ms (corr45: 84.8 ms ± 40.6; corr60: 79.7 ms ±
85.5; corr75: -85.9 ms ± 69.6). Thus, it was likely that the postural activity
occurred before the changes in GRF and corrections of arm muscular activity
(see Fig. 6.4) and kinematics (fcorrect). ANOVA found a significant main
effect of the Subject factor (F = 26.23, p < 0.001), but not-significant main
effects of Target (F = 0.49, p = 0.61), Phase (F = 1.61, p = 0.20), and Postural
Muscle (F = 2.34, p = 0.10) factors. Post-hoc multiple comparisons show that

148
the EMGcorrect times in the postural muscles were significantly shorter than
those in the reaching muscles for the corr60 target (Fig. 6.6A). However, this
effect is significant for the lSol and lPerl muscles, but not for the lTA muscle
(Fig. 6.6B). Furthermore, the differences between postural and reaching
EMGcorrect times are even stronger for the corr45 target, in which the visual
perturbation was the largest and the inter-trial variability of the EMGcorrect
times was the lowest (Fig. 6.6B). However, the variability of differences
between the postural and reaching EMGcorrect times was large across subjects,
with 2 of 9 subjects showing shorter EMGcorrect times for the arm muscles
(Fig. 6.6C). Lastly, the differences between postural and reaching EMGcorrect
times did not vary between the acceleration and deceleration phases of the reach
(Fig. 6.6D).
Each of the EMGcorrect times in the postural muscles were linearly
correlated with the EMGcorrect times in the reaching muscles. All EMGcorrect
times were calculated with respect to the onset of the initial reach movement
across the entire range of L2 light onset latencies (Fig. 6.1C). Figure 6.7 shows
an example of 9 linear regressions between postural and reaching muscles (data
for all ‘corr’ trials were pooled) for a representative subject (S9), The Y
intercepts (Yi) of the regression lines across most subjects and most conditions
were largely positive, which indicates that most of the corrective postural
muscle activity started before the onset of the corrective reaching muscle
activity (Table 6.2). The advance activation of the leg muscles with respect to
arm muscles ranged from 16.9 ms (lPerl before rPDel) to 390.8 ms (lPerl before
rTric). The slopes (m) of each of the regression lines ranged from 0.256 (lPerl
vs. rPDel) to 1.71 (lSol vs. rTric).
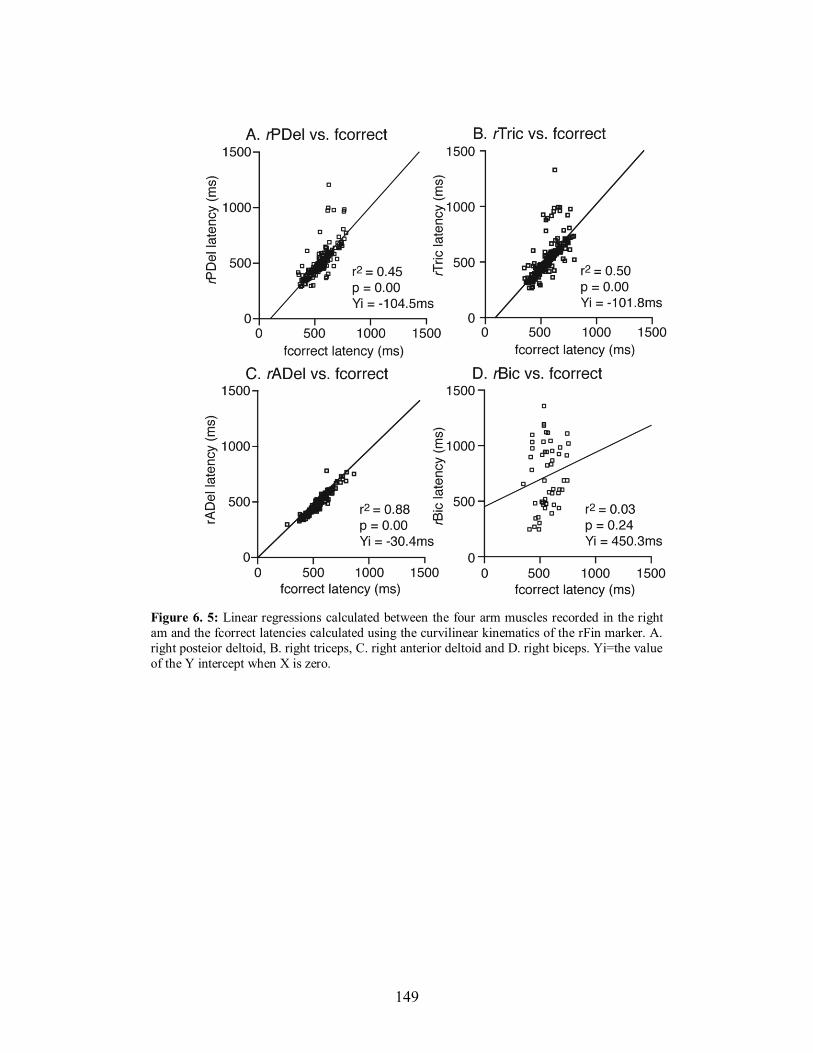
149
Figure 6. 5: Linear regressions calculated between the four arm muscles recorded in the right
am and the fcorrect latencies calculated using the curvilinear kinematics of the rFin marker. A.
right posteior deltoid, B. right triceps, C. right anterior deltoid and D. right biceps. Yi=the value
of the Y intercept when X is zero.

150
Figure 6. 6: Multiple comparisons of differences between EMGcorrect values of arm and leg
muscles. A. Average differences EMGcorrect differences per ‘corr’ target. Values for all 3 arm
and leg muscles have been pooled (averages for each ‘corr’ target +/- 95% confidence interval,
CI). Positive differences indicate postural muscle corrections before arm corrections (see direction of arrow, top right of figure). B. Average differences (+/- 810 95% CI) EMGcorrect
(all arm muscles pooled) per leg muscle. Values for EMGcorrect measures were pooled for all
arm muscles and expressed as differences with each leg muscle in turn (positive differences
also indicate postural muscle corrections before arm muscles). C. EMGcorrect differences (+/-
95% CI) per phase of reach, i.e. before peak velocity (Acceleration) or after peak velocity
(Deceleration). Filled circles show mean EMGcorrect differences for all targets, while open
circles show only data for corr45; shaded area represents 95% confidence interval. D. Average
differences (+/- 95% CI) EMGcorrect (all arm muscles pooled and all leg muscles pooled) per
subject.

151
Figure 6.7 Linear regressions calculated between postural and arm muscles participating in the
online corrections to unexpected visual perturbations of target position. Each graph shows the
postural muscle correction latencies (EMGcorrect) as the dependent variable (A. lSol, B. lPerl
and C. lTA) with the regressions performed between the 3 arm muscle correction latencies
(rPDel, rADel and rTric). Times (in ms) are expressed along each axis from the onset of the
initial reach movement.

152
Table 6. 2 Slope (m), Y intercept (Yi), r2 values, p-value for the strength
of the regression fit (p) and p-value for the intercept (p-int) for linear regressions conducted between the leg and arm muscles selected to characterize the online corrections to all targets for each subject
* and ** = significant at p < 0.05 and < 0.01 respectively. n/a = no significant
modulation of EMG between corr and reach conditions. = Multiple comparisons of intercepts
between subject are significant when p-int is alpha of 0.0057 (Sidak-Bonferroni correction for 9 tests between muscles per subject).

153
When data was pooled for each ‘corr’ condition the activation of the leg
muscles consistently preceded those of the arm (positive Y intercept) and the
leg muscles evolved earlier during the corrections than arm muscle activation
(slope analysis). In order to verify if the predictive activation of leg vs. arm
muscles held across target shifts of different amplitudes (eccentricity), we
performed the same linear regression analysis for the pooled ‘corr’ conditions
for each subject. Table 6.2 provides a complete breakdown of these
relationships. Of the 49 linear regressions performed (3 x 3 muscles for 9
subjects), all but 9 showed significant r2 values. Importantly however, only 5
revealed that arm muscle activation preceded leg muscle activation (Yi values <
0 and m values >1). For most comparisons, values of slope were <1, and as low
as 0.107 (rADel vs. lPerl for S6), confirming, as with the pooled data, that the
activation of postural muscles evolved more rapidly than arm muscles.
Therefore, when all conditions and muscles were considered, our results
supported a predictive control of postural activity in relation to arm muscle
activity across all target directions.
6.6 DISCUSSION
This study investigated how the two components of posture and
voluntary movement interact when changes in ongoing reaching movements are
produced while standing. We proposed two possibilities: 1) when faced with an
unexpected change in visual target position, for which a reaching movement
had been initiated, postural adjustments would occur after the adjustments seen
in the arm muscles correcting the trajectory of the hand towards the target; 2)
postural adjustments would occur before any change in arm muscle correction
and kinematic change of hand position, and posture would therefore be
predictive with respect to the voluntary component of the action. Our results
supported the latter of these two possibilities, i.e. modifications in leg muscle
activity preceded those of arm muscles when target position changed,
regardless of how far a correction was required to be made with respect to the
midline (eccentricity). We will begin by discussing some methodological

154
differences between our paradigm and previous visuomotor corrections
paradigms. Then we will highlight the functional role of the online postural
adjustments, situate our kinematic correction results within existing literature
taken from double-step experiments carried out in seated subjects, and finish by
placing our findings within a theoretical framework of predictive motor control.
6.6.1 Methodological considerations
In this study, we used a digital signal, derived from a switch detecting
the onset of right finger movement, to trigger the change in light target from the
initial central one to one of the other 3 targets. Different delays resulted
between when the finger began to move and when the first and second lights
extinguished and illuminated, respectively. This paradigm was intended to
provide us with a means of investigating how humans adapt posture and
movement when programmed actions must be modified ‘online’ after they have
begun. Our paradigm cannot however, be regarded as a classical ‘double-step’
paradigm, one that involves a change in gaze saccade after an initial one has
been initiated, a paradigm traditionally been used to probe the properties of the
oculomotor system (Becker and Jürgens 1979; Westheimer 1954; Wheeless et
al. 1966). This is principally because our light target changes (from L1 to L2)
were not triggered using the onset of the initial gaze shift, as in other arm
movement studies (Goodale et al. 1986; Gritsenko et al. 2009). Nevertheless,
our paradigm can be likened to a number of arm movement studies that have
used a double step paradigm triggered on the onset of movement, either a 1-step
(amplitude), double-step (Gielen et al. 1984; Megaw 1974), or a 2-step
(amplitude and direction) one (Day and Lyon 2000; Georgopoulos et al. 1981;
Soechting and Lacquaniti 1983). For the purposes of this study however, using
delays from the onset of finger movement provided us with a window of time
sufficiently long in which to investigate changes in arm movements (and the
EMG activity related to them), and their associated postural adjustments. Given
that our L2 latencies averaged between 365-394 ms, and fcorrect latencies
between 556-565 ms, we were sure that we had a window of time (± 200 ms)

155
long enough to perform our analysis of both arm and postural muscles.
Moreover, during this period (and beyond) our focus was on the relationship
between arm and postural muscle activity to produce the corrections.
6.6.2 Postural adjustments contribute to the execution of voluntary
movement
An interesting aspect of our results, with regard to the postural
corrections associated with the change in hand trajectory, was that the postural
muscles showed activity that effectively increased the shear force under the
limb contralateral to the reaching arm prior to fcorrect. The vertical force under
this limb also showed loading during the same period. This would suggest that
the postural corrections acted to push out and down with the left foot, thus
helping to rotate the body rightwards towards the new target (see shaded area,
Fig. 6.4B). With this in mind, the postural corrections can be described as being
a component of the voluntary movement, rather than ensuring only the
maintenance of equilibrium. In other words, they accelerated the body in the
direction of the target and did not stabilize posture or compensate for the
impending arm correction. Had the latter been the primary objective of the
postural adjustments produced between L2 and fcorrect, significant activation
of right limb muscles would have been recorded to counter the impending
rightward rotation of the body when the arm was oriented towards the new
target. In light of this, it would seem difficult to divide so-called ‘posture’ and
‘movement’ components of the motor act, as has often been the case during
voluntary movement studies (Cordo and Nashner 1982; Hess 1954; Saltzman
1979). Rather, our results corroborate earlier work suggesting an integration of
postural and focal commands at higher levels of the CNS (Aruin and Latash
1995b; Commissaris et al. 2001; Stapley et al. 1999). Our results show that
postural adjustments contribute to focal corrections of voluntary movement.

156
6.6.3 Effects of standing on the characteristics of online corrections of the
arm
An interesting question that arises when examining the online correction
of arm movements during stance, is to what extent does upright posture
influence the time taken to initiate a correction once target position changes? It
is out of the scope of this study to make definitive conclusions with regard to
this question from an experimental perspective, as 1) we did not directly
compare online corrections in seated and standing conditions, and 2) in order to
obtain significant postural adjustments, the amplitudes of target shift used by us
far exceeded those adopted in seated studies. Nevertheless, comparisons can be
made between our study and previous ones in terms of the percentage of total
MT taken to initiate a corrective response, even though the mode of target shift
should also be accounted for (see above).
A detailed description of the early kinematic response and its relation to
the amplitude of target displacement has recently been reported by Gritsenko et
al. (2009). They used target shifts of 15 cm amplitudes in 8 possible directions
from a central one. Light changes were triggered during a period of saccadic
suppression of the initial saccade to the first light, and were estimated to occur
on average 50 ms before the onset of initial hand movements, which were
executed at preferred speeds (as in our study). For perturbed movements that
were on average 403 ms in duration, Gritsenko et al. (2009) reported that
corrections occurred at 35% (average 140 ms) of total MT. Longer ranges of
38-61% have however been reported by Prablanc and Martin (1992). Our
correction values occurred at, on average 47% of total MT across the 3 target
positions. Thus, it is likely that the mode of target shift trigger (gaze vs. first
hand movement) plays a role in the onset of the online correction. Despite this,
our results demonstrate that standing does not impede the early onset of the
correction, which is to some extent comparable to online corrections produced
when humans are seated.
Nevertheless, it is evident that both the total MTs and times to online
correction (fcorrect) reported by us were far slower (almost double) than those

157
reported during seated movements (Goodale et al. 1986; Gritsenko et al. 2009;
Komilis et al. 1993; Pélisson et al. 1986; Prablanc and Martin 1992; Sarlegna et
al. 2003). Previous studies using the double-step paradigm to perturb arm
movements during stance have reported correction times as low as 164-168 ms
(Fautrelle et al. 2010). However, these authors used targets changes that
required corrections in the sagittal plane only and triggered target position
change before or just after (50 ms) hand movement onset. To our knowledge,
the only other study of double-step perturbations during stance investigated
differences in reaction time or MT with the likelihood of a double-step
perturbation (Martin et al. 2000). Other recent studies have investigated online
corrections of the foot during walking or a single step (Reynolds and Day
2005a; b). The foot online correction onset has been reported to be between 239
and 402 ms after the foot off the ground for a step (Reynolds and Day 2005b),
while during swing phase of locomotion the foot online correction onset has
been reported to be more similar to that for the arm, 114-151 ms (Reynolds and
Day 2005a). This suggests that be delayed onset of foot online correction
during a step and, possibly, the delayed onsets of online correction observed in
this study are due to the increased information processing of equilibrium
constraints, which may be simplified during a predictive shift of CoM thought
to occur during locomotion (Day et al. 1997). Furthermore, Hollands et al.
(Hollands et al. 2004) showed that during tasks involving body rotation when
standing the onset of saccadic eye movements to targets is also delayed,
possibly by additional information processing of equilibrium constraints.
Interestingly however, taken together with studies of arm correction in seated
subjects and foot correction of stepping subjects, our results provide evidence
for a constancy of times to online corrections of arm movements regardless of
postural configuration. Both Gritsenko et al. (2009) and Prablanc and Martin
(1992) reported no significant differences in the time to correction across
different target amplitudes, and our fcorrect data also showed no significant
differences across the 3 ‘corr’ conditions.

158
It may be debated that the outward (rightward) curvature (rather than a
straight trajectory towards the central target) seen for the ‘reach’ trials was
influenced by the expectation of a change in target position. It is outside the
scope of the present study to definitively rule out if expectation was the cause
or not, of that curvature. However, it is interesting to note that trajectories
displayed for reaches to the same central target during a paradigm of
multidirectional reaching (Leonard et al. 2009) also displayed a similar
curvature (unpublished observations). Moreover, in a seated online correction
task, Day and Lyon (2000) also showed similar curvatures of straight reaches.
However, further study is warranted to investigate the role of target change
expectation in trajectory formation, in both the seated and standing positions.
6.6.4 Implications for the control of posture and movement
Skilled voluntary movement, such as reaching with the arm, relies upon
the prediction of the future state of the motor system because of the inherent
delays associated with information processing. Studies of anticipatory
adjustments of grip force with load during object manipulation (Flanagan and
Wing 1997; Johansson and Cole 1992; Kawato 1999) have shown that the CNS
‘predicts’ the dynamic effects of upcoming movements. Interestingly, such a
grip force/load force predictive relationship is preserved and precedes arm
movement online corrections, which is strong evidence of the use of predictive
motor mechanisms by the brain (Danion and Sarlegna 2007). Other studies of
the initiation of stepping suggest that the CoM is controlled predictively by the
CNS to maximise the efficiency of movement (Day and Lyon 2000; Day et al.
1997). These studies, and others that have examined the execution or learning
of seated arm movements, have proposed that forward, or predictive, internal
models are employed to anticipate the consequences of actions based on
efference copy of outgoing motor commands (Wolpert and Miall 1996).
Efference copy is used to estimate the sensory feedback likely to result from the
motor command, which results in accurate predictions of current motion
(Davidson and Wolpert 2005; Desmurget and Grafton 2000).

159
The use of such predictive models for controlling equilibrium during
arm movements has also been documented. The specific characteristics of
anticipatory postural adjustments occurring before the onset of movement
(APAs) depend on the prior knowledge of arm and body dynamics (Bouisset
and Zattara 1981; 1987). However, APAs are predictive with respect to the
initial onset of movement and are programmed with the CNS anticipating the
consequences of the impending act. Our results have shown that when the
initial goal of a programmed reach movement unexpectedly changes after it has
been initiated, the postural adjustments required to execute the online
corrections consistently lead any changes in arm EMG or kinematics (Figs.
6.6,6.7). Moreover, our results also show that, even for small amplitude
changes in target, requiring smaller corrections of hand trajectory and posture, a
predictive mode of postural control is still largely adopted (leg muscle
corrections led those of the arm consistently across ‘corr’ conditions, Fig. 6.7).
Therefore, we suggest that postural adjustments in this situation are largely
predictive in nature and not based upon feedback from the moving limb. Our
results corroborate those described by Ruget et al (2008) who showed that
human subjects are able to modify components of weight shift online during the
anticipatory period preceding the onset of stepping.
How do our findings fit with what is known about the neural control of
arm movements and posture as well as forward models of reaching movements?
It is known that there is a significant cortical involvement in predictive postural
behaviour. Mackinnon et al. (2007) demonstrated a facilitation of the muscles
involved in APAs whereas anticipatory adjustments of arm muscles were
absent in patients with damage to their motor cortex (Viallet et al. 1992).
Moreover, animal studies have suggested that the cortex contributes to motor
planning of reaching during stance (Martin and Ghez 1985; Perfiliev 2005;
Perfiliev 1998; Vicario et al. 1983) and the feedforward adjustments
accompanying the reach (Yakovenko and Drew 2009). Thus, the CNS may
send a global command that specifies the planning and execution of movement
and posture as one. Even though evidence has been given in support of global,

160
hierarchical signal in which pathways for movement give off collaterals for
posture (Gahery and Nieoullon 1978), the findings of a number of studies
would suggest that parallel, independent commands exist for the postural
adjustments preceding the movement, accompanying the movement, and indeed
the movement itself (Horak et al. 1984; Schepens and Drew 2004; Schepens et
al. 2008).
Although our results do not allow us to elucidate the origin and specific
anatomical structure of the pathways involved in posture and movement, we
can speculate about what happens, in terms of the production of postural
adjustments in relation to arm muscle activity, when the expected outcome of a
reach does not materialize, such as when target location unexpectedly changes
after movement onset. Figure 6.8 proposes a simplified model of how
commands movement and posture would fit with a forward model of arm
movements. The model shows that the motor cortex sends a global planning
command for movement and posture (Gritsenko et al. 2009). Efference copy of
commands for the execution of both the arm movement, APAs for initiating the
reach and aPAs accompanying the movement (right side of figure) ensures that
discrepancies of the expected movement are detected (Desmurget and Grafton
2000). All the time the expected reach movement mirrors that actually being
executed (above the grey horizontal bar), rapid feedback loops adjust and refine
the movement in real-time. When the target shifts, the expected movement no
longer reflects that which must be produced and a delay ensues. It is here that
adjustments of accompanying postural commands (aPA adjustments) must
occur before those of the arm (arm adjustments), in order that the dynamic
constraints of the task to be satisfied. Once posture is updated, predictively of
the arm movement, the online correction can be made and the target attained.
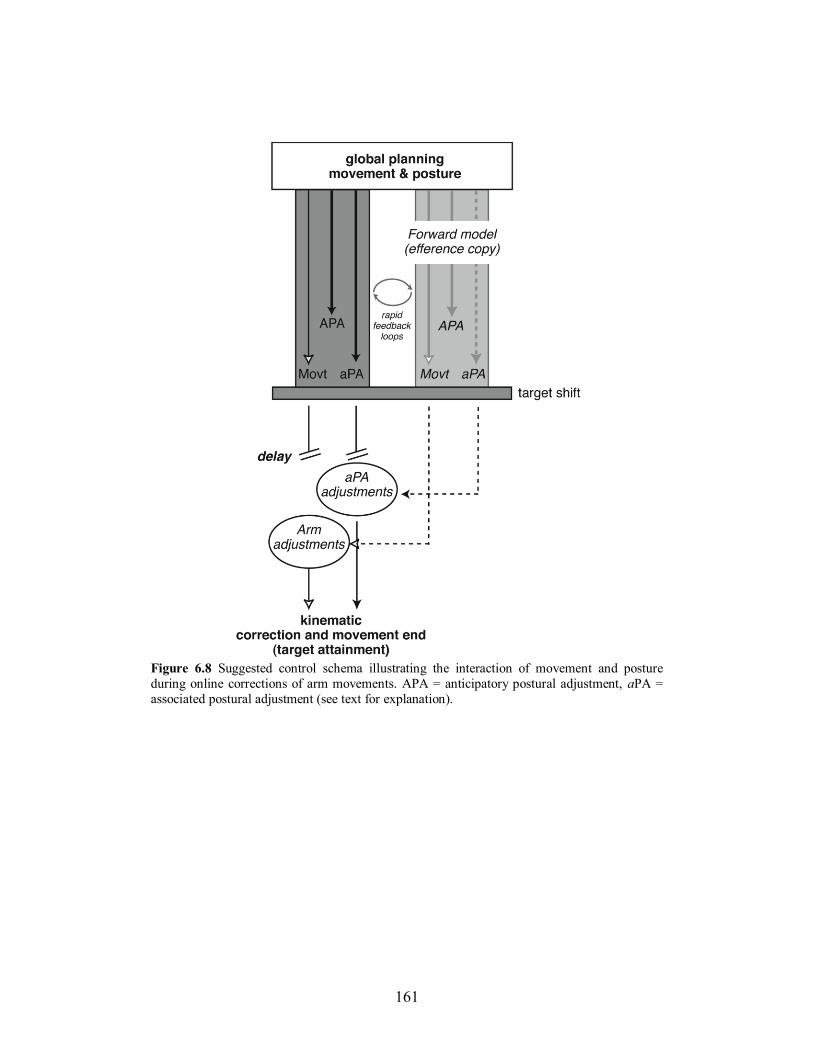
161
Figure 6.8 Suggested control schema illustrating the interaction of movement and posture
during online corrections of arm movements. APA = anticipatory postural adjustment, aPA =
associated postural adjustment (see text for explanation).

162
6.6.5 Conclusions
We have shown that when online corrections of ongoing arm
movements are required, the CNS adopts a predictive mode of postural control,
rather than a purely feedback-based mode. This was shown by the adjustments
of postural muscle activity consistently leading those of the arm muscles
responsible for correcting the trajectory of the arm to the target (Figs. 6.6,6.7).
Had the CNS relied upon information from a change in arm movement to
update posture, adjustments of arm muscle activity would have led those of the
postural muscles. This was not seen even for the smallest amplitude target
corrections (15° to the right), which could have been executed without
significant threats to stability.
6.7 ACKNOWLEDGEMENTS
This study was supported by a grant from the Canadian Foundation for
Innovation and a Natural Sciences and Engineering Research Council (NSERC)
of Canada discovery grant to PJS. The authors thank Prof. Robert Kearney
(Dept. Biomedical Engineering, McGill University) for his advice about data
analysis.

163
Chapter 7
General Discussion
Movements executed while standing incur a disturbance to equilibrium
due to the complex architecture of the skeletal and neuromuscular systems of
land-dwelling animals. For example, reaching to a target located just beyond
reach while standing (Flanders et al. 1999) requires the coordination of arm,
trunk, and supporting limb segment muscles to attain the visual target. It has
been shown that, in this instance the CNS predicts the dynamic consequences of
the motion, and plans postural adjustments preceding and accompanying the
movements to stabilize the body and assist in the performance of the movement
(Massion 1992). Presumably, coordinating these two aspects of a motor
behaviour is complex, and requires coordinated signals for the postural and
movement systems. The central organization of these two components of motor
behaviour remain elusive, therefore, the primary aim of this thesis was to
investigate the patterns of postural activity during goal-directed reaching
movements performed while standing.
To better understand how posture is controlled in relation to goal
directed movements, three studies were performed. The first study (Chapter 4),
examined how feedforward postural adjustments are organized spatially with
respect to movement direction. It demonstrated the existence of a force-
constraint strategy and muscle activity that is directionally tuned for pPAs. In
contrast, aPAs, while tuned in their EMG patterns, were characterized by a
greater dispersion of the direction of the GRF. With the goal of building on the
findings of the first study, the second study (Chapter 5) sought to determine
whether muscle synergies can explain the coordination of muscle activity for
feedforward postural adjustments preceding reaching movements. Results
showed that the coordination of muscle activity in the pPA period can be
explained by a time-varying recruitment of a few synergies. Finally, the third

164
study (Chapter 6) explored the nature of postural control signal associated with
online corrections of reaching movements. It showed that the CNS updates
commands to the postural muscles in advance of corrections to arm muscles
and finger trajectory.
Overall, these findings advance our knowledge of feedforward
(predictive) postural control. In particular, Chapters 4 and 5 showed that certain
characteristics of feedforward postural adjustments are similar to those
observed for feedback postural control. Furthermore, Chapter 6 demonstrated
that posture is prioritized and updated predictively when online corrections to
movement are required. Finally, this body of work provides baseline measures
of feedforward postural control as a first step to understanding why deficits in
balance control during voluntary movement may occur in the elderly, and other
pathological conditions.
In the following discussion, I will first review the significance of the
findings of tuned muscle activity, force constraint and muscle synergies for
feedforward postural control in relation to the strategies observed in feedback
postural control. Then, I will discuss how the findings can be used to interpret
possible modes of central control of posture and movement. Subsequently, I
will address the online control of posture within a broader scope of predictive
motor control. I will also review the relevance of these studies to the study of
balance in clinical populations. Finally, I will conclude by discussing some of
the unanswered questions raised by this thesis and directions of future research.
7.1 Characterization of feedforward postural adjustments during
multi-directional reaching movements
7.1.1 Role feedforward postural activity
Historically, the study of posture control during voluntary movement
has focused on the role of feedforward postural adjustments with respect to the
focal movement to gain insight into how these two components of motor
behaviour may be controlled by the CNS. Early studies of APAs suggested that

165
the postural activity preceding arm movements function to stabilize the body
against the disturbance of the movement (Bouisset and Zattara 1981; 1987;
Crenna et al. 1987; Massion 1992). However, subsequent studies investigating
forward reaching movements involving the trunk (Commissaris et al. 2001;
Stapley et al. 1998; Stapley et al. 1999), or those requiring a change in the
configuration of the BoS (Brenière and Do 1987; Brenière and Do 1986; Lepers
and Brenière 1995), have demonstrated that postural adjustments preceding the
movement onset instead create the dynamic forces required for initiating the
movement. In these situations, it appears that feedforward postural activity
contributes to the execution of the goal-directed component of the movement.
Similarly, pPAs and aPAs during unperturbed and perturbed reaching
movements contributed to the execution of the focal component of the
movement (Leonard et al. 2009; Leonard et al. 2011). For example, during
unperturbed reaching movements, pPAs and aPAs created the forces that
contributed to the initiation and termination of the reaching movement,
respectively (Leonard et al. 2009). In particular, a small forward displacement
of the CoM was observed prior to the onset of the reaching movement, as
defined by the onset of the finger movement. This was associated with
activation of the flexor muscles serving to initiate movement rather than
stabilize the position of the CoM. However, for the aPAs for unperturbed
reaching, strong extensor activity served to decelerate the CoM and stabilize the
position of the body towards the target. In the case of perturbed reaching
movements, the aPAs clearly contributed to overall movement correction by
breaking the forward motion and accelerating the body towards the new target
(Leonard et al. 2011). Together, these data support the view that posture
contributes to a voluntary movement by ensuring that the dynamic conditions
for the movement are met and suggest that task demands are integrated and
used to plan the appropriate postural activity using an internal model of posture.

166
7.1.2 Independent or parallel commands for global planning of posture and
movement
The fact that the postural adjustments preceding and accompanying the
movement contribute to the overall execution of the movement does not
necessarily indicate that these two commands are controlled by a single neural
command. Data showing that pPAs may be time-locked to the onset of a
voluntary movement have suggested that posture and movement are controlled
in a hierarchical fashion, where posture is controlled via collaterals from the
primary descending command for the movement (Massion 1992). However,
situations where the latency of the pPA is variable with respect to the
movement onset have also been observed (Brown and Frank 1987; Cordo and
Nashner 1982; Massion 1992), suggesting that instead posture and movement
are controlled via parallel, but independent neural pathways. There is also
debate concerning the nature of the signals for posture and movement for aPAs.
Until recently, it was generally accepted that postural commands are organized
in a hierarchical fashion in relation to the descending command for movement
via collaterals that recruit postural networks, likely localized to the brainstem
(Gahery and Nieoullon 1978; Massion 1992). This framework was largely
based on knowledge that cortical signals contribute significantly to the aPAs
(Mackinnon et al. 2007; Massion 1992; Viallet et al. 1992).
Recently, a series of studies of reaching movements in the cat have
provided considerable insight into the nature of the command signals for
posture and movement (Schepens and Drew 2006; 2004; 2003; Schepens et al.
2008; Yakovenko and Drew 2009). These studies identified independent signals
in the PMRF related to the pPA, the aPA, the movement, or the aPA and the
movement. What is clear therefore, is that a supraspinal structure, such as the
brainstem, plays a significant role in organizing both the postural adjustments
needed to execute reaching movements in the standing position, as well as the
voluntary movement itself. Interestingly, this same structure has also been
shown to control purely feedback-driven postural adjustments (Stapley and
Drew 2009).

167
The results of Chapter 6 demonstrated that the signal for the corrective
aPA consistently led the modulatory signal of the correction in the arm EMG
(see Fig 6.6 and 6.7). These findings are interpreted in the context of a forward
model of arm and posture (Gritsenko et al. 2009) in which the signals for
movement and posture are controlled via independent, parallel pathways
(Yakovenko and Drew 2009). In this framework, a global signal for posture and
movement is expressed, likely in premotor areas and sent to the motor cortex
(Massion 1992; Yakovenko and Drew 2009). Then, independent signals for the
pPA, aPA and movement are sent to the respective areas of the PMRF that
recruit and scale the activity of the appropriate muscle groups for the different
components of the motor behaviour (Schepens and Drew 2004). Meanwhile,
efference copies of these commands are consistently being evaluated and ensure
any discrepancies of the expected movement are detected (Leonard et al. 2011).
Accordingly, the corrective aPA must occur prior to the focal correction of the
movement and potentially inhibits the movement command until the conditions
for the movement have been met, as has been observed for pPAs and movement
initiation (Cordo and Nashner 1982; Massion 1992).
7.1.3 Strategies for simplifying the control of posture and movement
Several authors have argued that the CNS relies on neuromechanical
strategies to simplify the process of coordinating the many DoF associated with
complex task of controlling balance (Bernstein 1967; Chiel et al. 2009;
Macpherson 1991; Ting 2007; Ting and Mckay 2007). The postural strategies
have been examined in detail for reactive balance control using support surface
perturbations in cats (Macpherson 1988a; b; Ting and Macpherson 2005) and
humans (Carpenter et al. 1999; Henry et al. 2001; Horak and Nashner 1986;
Torres-Oviedo and Ting 2007). These studies have provided detailed analyses
of the spatial and temporal characterization of the muscle activity, force and
kinematic patterns for a range of perturbations spanning many directions, with
varying amplitudes and velocities. The resulting postural responses are
characterized by muscle activity that is directionally-tuned and GRF patterns

168
that are constrained. As well, the coordination of muscle activity across a large
number of muscles is explained by the temporal recruitment of a small number
of muscle synergies (Ting and Macpherson 2005; Torres-Oviedo and Ting
2007) according to task goal (Chvatal et al. 2011). Overall, these data have been
interpreted to support the hypothesis that the CNS relies on a hierarchical
organization to simplify movement control (Lockhart and Ting 2007; McKay
and Ting 2008; Ting 2007; Ting and Mckay 2007).
Prior to the work presented herein, no study had examined feedforward
postural control with the specific goal of drawing parallels to feedback-based
postural strategies. As such, Chapters 4 and 5 characterized the spatial
organization of the GRF and EMG patterns preceding (pPA) and accompanying
(aPA) reaching movements performed in standing across many directions of
reach with the goal of drawing comparisons to those cited in the literature for
APRs. Indeed, the characteristics of the muscle activity preceding reaching
movements (pPAs), i.e. directionally-tuned EMG explained by muscle
synergies and force constraint strategy, bear striking similarities to postural
responses organized in a feedback mode (Henry et al. 2001; 1998b;
Macpherson 1988a; b; Torres-Oviedo and Ting 2007). Interestingly, however,
the patterns of force in the aPA period exhibited greater dispersion yet still
displayed muscle activity that is sensitive to the direction of reach. The
significance of these observations will be discussed next.
7.1.4 Force constraint strategy: neural strategy or geometry?
Whether the force constraint strategy reflects a higher-level neural
strategy, or whether it is simply the outcome of the biomechanics of the limbs
requires some discussion. In studies of APRs, a characterization of the active
restoring force patterns revealed a clear bimodal relationship between direction
of CoM motion consequent to the perturbation and the direction of the GRF
(Henry et al. 1998a; 2001; Macpherson 1988a; Ting and Macpherson 2004).
Based on these findings, it was hypothesized that CNS constrains the direction
of GRF to simplify the control of motion by decreasing the DoF to be

169
controlled (Henry et al. 1998a; 2001; Macpherson 1988a). However, recent
modeling (Bunderson et al. 2010; McKay et al. 2007) and neurophysiological
(Honeycutt and Nichols 2010) studies have provided evidence that suggests the
force constraint may be in part due to the biomechanics and geometry of the
limb. It has also been shown that the CNS modulates the force generating
capabilities of the limb through the flexible recruitment of muscle synergies
(McKay and Ting 2008).
This framework may explain why aPAs were associated with forces that
were distributed relatively uniformly in relation to pointing direction in contrast
to the bimodal distribution observed in the pPA (see Fig 4.7 and Fig 4.8).
During the pPA period, subjects adopted an upright body configuration with
equal weight distributed between the left and right supporting limbs. However,
in the aPA, subjects leaned towards a single target with their right arm
extended. To maintain balance in this position, a greater contribution from a
single limb to the loading force was observed, as the CoM moves towards the
target and towards the limits of the BoS. Thus, it is possible that the orientation
of the limb and its musculature resulted in the distributed force patterns. This
would support the hypothesis that the biomechanics of the limb determine the
set of possible directions of applied force (Bunderson et al. 2010; Honeycutt
and Nichols 2010).
An alternative possibility is that the task goal may influence the patterns
of GRF directions. Modeling studies have shown that the space of possible
force production, which is inherently determined by the limb mechanics
(Bunderson et al. 2010; McKay et al. 2007), is actively constrained by the
recruitment of synergies (McKay and Ting 2008). As discussed in Chapter 4,
the muscle activity and forces generated in the pPA served to accelerate the
body forwards towards the target. In contrast, aPAs functioned to stabilize the
body at each target, which required precise control of the CoM and finger end-
point positions. The biomechanical constraints of this task may require
additional muscle synergies to those recruited for the pPA. Several studies have
shown the existence of ‘shared’ and ‘task-specific’ muscle synergies across

170
tasks (Chvatal et al. 2011; d'Avella and Bizzi 2005; Krishnamoorthy et al.
2004). Therefore, one could argue that when the goal is to accelerate the CoM,
as is the case in pPAs and APRs, the force constraint strategy may provide the
optimal solution. However, in situations where the CoM must be stabilized
uniquely with a different postural configuration, such as when the CoM
excursion approaches the limits of the BoS, additional task-specific muscle
synergies may need to be recruited, which may be related to additional
directions of force production. Further study is required to disambiguate these
relationships.
7.1.5 The importance of muscle tuning and synergic organization for
feedforward postural control
A salient finding of this thesis is that muscle activity was directionally-
tuned during both the pPA and aPA periods and a synergic organization of
muscle activity exists for the pPAs. Most postural muscles were modulated as a
function of reach direction, with maximal activity occurring for a small range of
directions (see Figure 4.5 and Figure 4.6). Within each period, several muscles
shared similar directions of maximal activity, suggesting that they may be
modulated together within a muscle synergy. Analysis of these relationships in
the pPA with NNMF revealed the existence of spatially-fixed muscle synergies
that were modulated as a function of reach direction (see Fig 5.4). These
findings contribute to our understanding of the central problems of motor
control by supporting current hypotheses of dimensional reduction for
simplifying motor control (Cheung et al. 2005; Ting 2007; Tresch et al. 1999).
Furthermore, the apparent similarity to the organization of feedback postural
control suggest that the CNS relies on shared structures to organize the muscle
activity for controlling balance regardless of the control mode.
Muscle tuning has been consistently observed in a variety of motor
tasks, including voluntary isometric contractions of the elbow joint (Buchanan
et al. 1986; Buchanan et al. 1989), feedback postural responses (Carpenter et al.
1999; Henry et al. 2001; 1998b; Honeycutt et al. 2009; Macpherson 1988b;

171
Ting and Macpherson 2004; Torres-Oviedo and Ting 2007), reactive stepping
(Chvatal et al. 2011), postural adjustments during voluntary movements (Aruin
and Latash 1995a), and goal-directed reaching (Flanders and Soechting 1990;
Scott et al. 1997; Sergio and Kalaska 1998; Thoroughman and Shadmehr 1999).
The observation of muscle tuning across such a range of behaviours suggests
that directional tuning is an inherent property of the motor system and largely
reflects the underlying anatomical and mechanical capabilities of a muscle in a
given orientation as a result of its complex architecture (Buchanan et al. 1986;
Herrmann and Flanders 1998). Therefore, the finding of tuned muscle activity
in Chapter 4 is not completely unexpected, and has been demonstrated in other
postural tasks associated with voluntary behaviour (Aruin and Latash 1995a).
However, inspection of the tuning curves in relation to one another can reveal
considerable insight into the contribution of individual muscles to the overall
torque generated and the inter-muscular coordination for different directions of
motor behaviour (Buchanan et al. 1986; Buchanan et al. 1989; Macpherson
1988b). Of interest is the finding that several muscles shared similar shapes of
their tuning curves and were activated together to subserve the postural activity
for a reaching direction (see Figure 4.6). This grouped tuning suggests that a
muscle synergy organization is used by the CNS to simplify the control of
feedforward posture by recruiting groups of muscles together as muscle
synergies. It has been proposed that this synergic organization is a mechanism
to simplify the control of movement by reducing the dimensionality of the
DoFs to control (Ting 2007).
Several studies have suggested that the CNS simplifies the control of the
musculoskeletal system by recruiting a small number of motor modules (Bizzi
et al. 2000; Cheung et al. 2005; Roh et al. 2011; Ting and Macpherson 2005;
Tresch et al. 1999). This has been shown for a number of tasks, including
reactive balance control in cats and humans, locomotor behaviours in the frog
(Cheung et al. 2005; Roh et al. 2011; Tresch et al. 1999), finger movements
(Shim et al. 2005), and balance control during voluntary whole-body sway
(Klous et al. 2012; Krishnamoorthy et al. 2003; Krishnamoorthy et al. 2004)

172
and arm movements (d'Avella et al. 2011; Muceli et al. 2010). In reactive
balance control, muscle synergies have been shown to modulate end-point force
and are thought to be a mechanism by which the CNS controls the CoM. These
relationships have been interpreted to suggest that the CNS recruits muscle
synergies to simplify the task of translating abstract task-level goals, such as
CoM kinematics, into the appropriate muscle activity (Cheung et al. 2005;
Chvatal et al. 2011; Ting and Mckay 2007; Torres-Oviedo and Ting 2010).
The analyses performed in Chapter 5 revealed that a muscle synergy
structure can reconstruct the temporal, spatial and inter-trial variability of pPAs,
which are organized strictly in feedforward (Massion 1992). Indeed, these
findings suggest that a modular organization of muscle activity, as has been
observed in feedback postural control, generalizes to the control of posture in
feedforward. However, in this study, only the muscle activity of the postural
muscles was evaluated without considering the relationship of the muscle
synergies to the forces exerted on the ground or the resulting CoM motion. In
order to make conclusions about whether the extracted muscle synergies are
related to task-level goals and whether a muscle synergy structure supports a
hierarchical organization of posture in feedforward, further study of the
functional muscle synergies is required. Furthermore, the robustness of the
muscle synergies remains to be determined by examining whether the
composition of the muscle synergies remains stable in different biomechanical
contexts, for example by varying stance width or during the aPAs. Nonetheless,
this thesis provides a first step by demonstrating the existence of muscle
synergies for postural adjustments preceding multi-directional reaching
movements. This provides further evidence that similarities exist in the central
control of posture for both feedback and feedforward modes of control,
suggesting that these signals are integrated within the CNS.
In summary, the synergy analysis performed in Chapter 5 shows
modularity as a basis for muscle coordination for pPAs. Together with the
findings of Chapter 4, these results suggest that feedforward postural control is
organized in a similar manner to feedback postural control. Unfortunately,

173
without analyzing the biomechanical variables with respect to the muscle
synergies, these data cannot specifically contribute to the discussion of a
hierarchical control of posture and movement in which task-level goals are
translated into muscle activation patterns. Instead, the similarities observed here
for pPAs and aPAs to reactive postural control strategies suggests that
commands for feedback and feedforward are integrated somewhere in the CNS.
Potential neural substrates where these adjustments may be organized will be
discussed next.
7.2 Central control of posture and movement: integration of
feedback and feedforward postural commands
A primary objective of this thesis was to characterize the organization of
feedforward postural adjustments in order to draw comparisons to the strategies
observed for reactive postural control. The findings of Chapters 4 and 5 present
compelling evidence that the strategies for feedback and feedforward postural
control are similar in their organization. Observation of muscle tuning in pPAs
and aPAs and the existence of muscle synergies for the pPA suggest that an
integration of feedback and feedforward postural commands within the CNS.
Moreover, it is possible that the same neural pathways or motor modules are
recruited for controlling balance in voluntary and reactive postural tasks.
The localization within the CNS of these motor modules, or muscle
synergies, for either feedback or feedforward postural control is not currently
known. Substantial neurophysiological evidence points to the spinal cord (Bizzi
et al. 1991; Hart and Giszter 2004) and brainstem (Deliagina et al. 2008;
Honeycutt et al. 2009; Macpherson et al. 1997) as potential candidates for
encoding motor modules across a wide range of behaviours. For postural
control, however, it appears that the involvement of the brainstem is required.
Supraspinal structures are known to mediate postural responses organized in
both feedback and feedforward (Massion 1992). Furthermore, the lack of
spatial tuning of muscle activity in many postural muscles in spinalized cats

174
subjected to multidirectional support surface perturbations (Macpherson et al.
2007; Macpherson and Fung 1999) suggests that higher levels of the CNS are
required for appropriately mediating the postural muscle activity. Also, lesion
and inactivation studies of the reticulospinal circuitry of the brainstem result in
significant deficits of balance (Gorska et al. 1990; Gorska et al. 1995; Lawrence
and Kuypers 1968; Luccarini et al. 1990).
Recently, neurophysiological studies exploring the nature of the signals
of the brainstem during goal-directed reaching and postural responses to
unexpected disturbances of balance in the cat have shown that signals in the
PMRF contribute to both feedback and feedforward postural control (Schepens
and Drew 2006; 2004; 2003; Schepens et al. 2008; Stapley and Drew 2009).
These findings suggest that the PMRF of the brainstem gates neural signals for
both feedback and feedforward postural control and is the site of integration of
these two signals (Schepens et al. 2008; Stapley and Drew 2009). If so,
similarities in the outward expression of feedback and feedforward postural
behaviour, in terms of patterns of EMG and resulting force, would support this
notion. The brainstem has been proposed as an ideal structure for performing
this integration, and as it receives input from numerous descending commands
and rich afferent feedback from the spinal cord and periphery (Drew et al.
1996). Our results support this hypothesis by providing biomechanical and
electromyographic evidence that posture is organized similarly in both feedback
and feedforward modes of control. We suggest that the muscle synergies for
postural control may be organized at the level of the brainstem.
7.3 Predictive motor control: internal model of posture
A second major aim of the thesis was to examine how posture and
movement components of motor behaviour interact during online corrections of
reaching movements executed in standing. Classic double step paradigms, in
which the light shift is related to a gaze shift, have been used to examine the
visuomotor processes involved in making rapid online corrections to goal-
directed movements (Day and Brown 2001; Day and Lyon 2000; Pélisson et al.

175
1986; Prablanc and Martin 1992). These studies have shown that vision of the
arm improves accuracy by providing a continuous signal of the reach trajectory
in relation to the target goal. Overall, these studies have suggested that the CNS
uses a predictive forward model to rapidly detect deviations from the desired
trajectory and signal a correction (Davidson and Wolpert 2005; Desmurget and
Grafton 2000; Gritsenko et al. 2009). How equilibrium constraints affect the
ability to make such online corrections of reaching movement has received little
attention. Specifically, is it not known if posture is updated in a similar
predictive fashion when online visuomotor perturbations of the reaching
movement are imposed when the subject is standing.
Accordingly, in Chapter 6, a modified double step paradigm was
employed to challenge both the postural and movement systems and examine
the interaction between the two. Although previous studies have shown that
reach uncertainty is integrated in a predictive manner (Martin et al. 2000) and
that corrections to both posture and movement occur automatically (Fautrelle et
al. 2010), neither study systematically addressed the temporal relationships
between the postural and reach behaviours. As such, the analyses performed in
Chapter 6 investigated the temporal relationships between the postural and
focal commands during online corrections of reaching during standing. It was
found that subjects consistently updated their postural commands prior to
correcting their arm movement, even when perturbations were initiated late in
the reaching movement (i.e., deceleration phase of the reach). These results are
interpreted to suggest that the CNS adopts a predictive model of the body and
arm dynamics for controlling posture rather than relying on delayed feedback of
the arm correction. These findings support previous observations that the CNS
is able to modify weight shift during APAs prior to step initiation (Ruget et al.
2008).
The results presented here contribute to a general understanding of the
mechanisms underlying postural control during the execution of voluntary. A
widely accepted view is that the CNS plans goal-directed movements using
knowledge of the dynamics of the musculoskeletal system and the environment

176
(Davidson and Wolpert 2005). Posture is thought to be controlled in a similar
way (Bouisset and Zattara 1981; 1987; Ruget et al. 2008). However, prior to the
study herein, no data could support or refute the hypothesis that the CNS
continued to rely on a predictive internal model during online corrections of the
focal movement. Specifically, the fact that posture was updated prior to the
reach correction would suggest that corrections to posture and movement are
not based upon a purely feedback-driven model of limb movement, as the
postural adjustments occurred in advance of the adjustments to limb trajectory
and muscle activation. Therefore, our results support the notion of a predictive
internal model of voluntary movement, whereby the consequences of the action
are estimated in advance and planned for by the CNS.
7.4 Justification for understanding disorders of posture and
balance
Currently, it is estimated that over one million Canadians experience at
least one fall per year, with associated health care costs estimated at $2.8 billion
annually (Health Canada, 2002). More important, however, are the devastating
consequences of fall-related injuries, which are known to precipitate more
serious, life-threatening co-morbidities. The study of instability and falls has
focused mainly on the mechanisms of reactive postural control in populations
known to exhibit deficits in balance control (Mansfield and Maki 2009; Tokuno
et al. 2010). However, falls commonly occur in situations of dynamic balance,
where both postural and focal components of a movement must be coordinated.
It is not known if individuals fall as a result of deficits in their feedback or
feedforward control mechanisms, or whether they lack the ability to coordinate
posture and movement (Woollacott 2006).
While the emphasis of the presented work has been to understand
feedforward postural control mechanisms associated with reaching movements
in healthy young individuals, the data generated from these studies provide a
basis for characterizing differences in feedforward postural control strategies in

177
people prone to falls. Notably, both the spatial and temporal organization of the
pPAs and aPAs identified in these studies can serve as a benchmark for
comparison with populations afflicted with deficits in balance control.
Furthermore, using the experimental paradigm used here, the online control of
posture during visuomotor perturbations of reaching can be investigated in
different populations. Specifically, the hypothesis that an inability to update
postural commands using a predictive signal when the goal of the focal
movement changes results in a loss of balance could be tested. Thus, both the
experimental paradigm and the results of this thesis have significant
implications for clinical and fundamental applications.
7.5 Conclusions and future directions
Together, the results from this thesis contribute significantly to our
understanding of the central organization of posture and movement.
Furthermore, the experimental paradigm developed for the studies of this thesis
provides a basis for exploring the coordination of posture and movement in
populations suffering from balance deficits. I have shown similarities in the
organization of feedforward postural adjustments to those observed in feedback
postural control, providing biomechanical evidence that the CNS may rely on
similar neural pathways to control posture during predictable and unexpected
disturbances of balance. Subsequently, I demonstrated that the CNS relies on a
predictive control in the aPA when online corrections in movements are
required. While the results presented in this thesis provide important insight
into our understanding of how the CNS coordinates posture and movement, a
number of questions are raised that require further study. Some of these are
reviewed in the following paragraphs.
7.5.1 Do the elderly differ in the spatial and temporal organization of
feedforward postural control?

178
Clear deficits in postural control in the elderly have been documented
(Woollacott 2006). However, most studies have focused on the differences in
reactive postural strategies compared to healthy controls, with little attention on
feedforward postural control. Therefore, to fully understand why and how
people become destabilized and subsequently fail to recover balance with
increasing age, studies exploring the coordination of posture and movement are
required. Additional studies exploring the organization of predictive postural
control in these populations are required if we are to fully understand why and
how people become destabilized and subsequently fail to recover balance. The
experimental paradigm developed here provides the basis for undertaking such
studies.
The task and methodology developed in this thesis provide a novel
means for examining the coordination of posture and movement in the elderly.
These studies would complement our current knowledge about deficits in
feedback postural control. By it’s very nature, the reaching task executed in
standing allows a clear dissociation between the changes related to either the
postural or focal components of the overall movement strategy. Both the
temporal and spatial organization of muscle activity and GRF patterns can be
quantified in detail, and compared to the baseline measures for healthy young
adults documented in Chapters 4 and 5. This approach will make it possible to
identify differences in control strategies that may contribute to instability and
subsequent falls by making comparisons between healthy and afflicted persons.
Using this knowledge, it is hoped that targeted rehabilitation programs for
improving balance control can be developed and refined.
Moreover, it would be possible to determine whether the elderly are
capable of updating posture in a predictive manner when online changes in the
movement goal are required. It is possible that the elderly rely on a feedback
mode of control in such situations, and may be destabilized by the change in the
focal segment’s trajectory. The experiments performed in Chapter 6 could be
extended to elderly populations to test this hypothesis. This would provide
important insight into the control and coordination of posture in dynamic

179
situations in the elderly, who often become destabilized in these types of
dynamic situations. These methods and findings could then be extended to
other populations suffering from deficits in balance control.
7.5.2 Are feedforward muscle synergies robust and how does their
recruitment relate to task-level goals?
In Chapter 5, it was found that the temporal recruitment of a few muscle
synergies can explain the coordination of muscle activity of the pPAs,
presumed to be controlled strictly in feedforward (Massion 1992). The first
question that arises from this study is related to the robustness of the pPA
muscle synergies: are the same muscle synergies recruited for the pPAs as
aPAs? In feedback postural control, muscle synergy robustness has been
evaluated by examining the composition and activation of muscle in different
postural configurations (Torres-Oviedo et al. 2006; Torres-Oviedo and Ting
2010), sensory contexts (Torres-Oviedo and Ting 2010) and types of postural
reactions (Chvatal et al. 2011). Muscle synergies have also been shown to be
stable across a variety of motor behaviours, although task-specific synergies
may be recruited (Cheung et al. 2005; Chvatal et al. 2011; d'Avella and Bizzi
2005). Therefore, I propose to evaluate muscle synergy robustness by
determining whether the CNS recruits the same muscles synergies in the pPA
and aPA periods. Given that the goal of the postural adjustments differs for the
the pPA and aPA (acceleration and deceleration of the CoM, respectively), I
hypothesize that the CNS will recruit a set of shared synergies in addition to
task-specific muscle synergies in the aPA.
In the event that task-specific muscle synergies are found for the aPAs,
it is possible that these additional synergies explain the additional directions of
GRF produced in the aPA period. To test the hypothesis that stabilization of the
body at the target requires additional muscle synergies related to the GRF, a
functional synergy analysis could be performed. To determine these
relationships, biomechanical parameters, such as GRF direction (Ting and
Macpherson 2005; Torres-Oviedo et al. 2006) and CoM acceleration (Chvatal

180
et al. 2011) would be included in the NNMF algorithm. Also, to increase the
richness and variability of the data set, the reaching experiments could be
performed with a variety of stance configurations, such a wide, narrow, natural,
and single foot. This will permit the examination of the muscle synergies in a
variety of biomechanical contexts.
7.5.3 Online control of posture: effects of direction and time of visual
perturbation
The findings of Chapter 6 demonstrated that the postural adjustments
required for initiating online corrections in arm movements while standing for
target shifts from a central target to the right of the midline are organized
predictively of the arm correction. To make these corrections, postural muscle
activity was programmed to create significant loading under the left foot to
break the counter-clockwise rotation and accelerate the body to the new target
(see Fig 6.4B). In the event of a center-left target shift, however, the body
would already be rotating in the direction of the new target. The breaking and
acceleration of the body observed for center-right corrections would not be
required for successful execution of the task in a center-left perturbation. This
begs the question whether the initial planned postural adjustment to the first
target is robust enough to support the change in focal reach direction?
Specifically, would posture continue to be updated predictively in situations
where the body is already in motion towards the corrected target, or would the
adjustments in posture occur in a reactive mode, relying on afferent feedback
related to the motion of the arm or of the CoM in relation to the BoS? To test
the hypothesis that the CNS continues to rely on a predictive mode of control
irrespective of the direction of the perturbation, the postural activity in relation
to the arm EMG and trajectory for center-left perturbations should be
quantified.
A second issue raised from the study in Chapter 6 is how the latency of
the target shift affects the ability of the CNS to organize the appropriate
predictive postural adjustments. In this study, the majority of the target shifts

181
occurred at or very soon after peak velocity during the deceleration phase of the
reaching movement (see Fig 6.1C). It is possible that target shifts occurring
earlier in the execution of the reaching movement may involve a different mode
of control for organizing the postural adjustment. For example, several studies
of perturbed reaching movements executed in seating have shown that the reach
corrections occur ‘automatically’ when target shifts are presented just before or
immediately after the onset of the first gaze shift (Day and Brown 2001; Day
and Lyon 2000; Pélisson et al. 1986; Prablanc and Martin 1992). It remains to
be seen how posture and movement interact when corrections to the reach are
triggered early in the movement, and in relation to visual information, such as a
gaze shift, rather than the reach onset. Therefore, to confirm that a predictive
mode of postural control is consistently used irrespective of latency of the
target shift, experiments involving target shifts occurring at several latencies
with respect to an initial gaze shift should be performed. The findings of such
studies, when compared to online corrections of reaching in seated, will provide
insight into how equilibrium constraints affect the planning and execution of
online corrections involving posture and movement.

182

183
Chapter 8: References
Abdi H. The Bonferonni and Šidák Corrections for Multiple Comparisons. In: Encyclopedia of
measurement and statistics, edited by Salkind N. Thousand Oaks, CA: Sage, 2007, p. 103-107.
Alstermark B, and Isa T. Circuits for Skilled Reaching and Grasping. Annual review of neuroscience 2012.
Aruin AS, and Latash ML. Directional specificity of postural muscles in feed-forward
postural reactions during fast voluntary arm movements. Experimental brain research 103:
323-332, 1995a.
Aruin AS, and Latash ML. The role of motor action in anticipatory postural adjustments
studied with self-induced and externally triggered perturbations. Experimental brain research
106: 291-300, 1995b.
Babinski J. De l'asynergie cerebelleuse. Rev Neurol 7: 806-816, 1899.
Balasubramaniam R, and Turvey M. The λ-Based Equilibrium Point (EP) Hypothesis 1990-
1999. 1-6, 2005.
Bastian AJ. Learning to predict the future: the cerebellum adapts feedforward movement
control. Current Opinion in Neurobiology 16: 645-649, 2006.
Becker W, and Jürgens R. An analysis of the saccadic system by means of double step
stimuli. VISION RESEARCH 19: 967-983, 1979.
Bernstein N. The Coordination and Regulation of Movements. Oxford: Pergamon Press, 1967,
p. 196.
Bhushan N, and Shadmehr R. Computational nature of human adaptive control during
learning of reaching movements in force fields. Biological Cybernetics 81: 39-60, 1999.
Biguer B, Jeannerod M, and Prablanc C. The coordination of eye, head, and arm movements
during reaching at a single visual target. Experimental brain research 46: 301-304, 1982.
Bizzi E, Mussa-Ivaldi FA, and Giszter S. Computations Underlying the Execution of
Movement - A Biological Perspective. Science 253: 287-291, 1991.
Bizzi E, Tresch MC, Saltiel P, and d'Avella A. New perspectives on spinal motor systems. Nature Reviews Neuroscience 1: 101-108, 2000.
Black FO, Shupert CL, Peterka RJ, and Nashner LM. Effects of unilateral loss of vestibular
function on the vestibulo-ocular reflex and postural control. The Annals of otology, rhinology,
and laryngology 98: 884-889, 1989.
Bonnetblanc F, Martin O, and Teasdale N. Pointing to a target from an upright standing
position: anticipatory postural adjustments are modulated by the size of the target in humans.
Neuroscience letters 358: 181-184, 2004.

184
Bouisset S, and Zattara M. A sequence of postural movements precedes voluntary movement.
Neuroscience letters 22: 263-270, 1981.
Bouisset S, and Zattara M. Biomechanical study of the programming of anticipatory postural
adjustments associated with voluntary movement. Journal of Biomechanics 20: 735-742, 1987.
Brustein E, and Rossignol S. Recovery of locomotion after ventral and ventrolateral spinal
lesions in the cat. I. Deficits and adaptive mechanisms. J Neurophysiol 80: 1245-1267, 1998.
Buchanan TS, Almdale DP, Lewis JL, and Rymer WZ. Characteristics of synergic relations
during isometric contractions of human elbow muscles. Journal of neurophysiology 56: 1225-
1241, 1986.
Buchanan TS, Rovai GP, and Rymer WZ. Strategies for muscle activation during isometric
torque generation at the human elbow. Journal of neurophysiology 62: 1201-1212, 1989.
Bunderson NE, Mckay JL, Ting LH, and Burkholder TJ. Directional constraint of endpoint
force emerges from hindlimb anatomy. The Journal of experimental biology 213: 2131-2141,
2010.
Canada H. Healthy Aging: Prevention of Unintentional Injuries Among Seniors. 2002.
Cappellini G, Ivanenko YP, Poppele RE, and Lacquaniti F. Motor patterns in human
walking and running. Journal of neurophysiology 95: 3426-3437, 2006.
Carpenter MG, Allum JH, and Honegger F. Directional sensitivity of stretch reflexes and balance corrections for normal subjects in the roll and pitch planes. Experimental brain
research 129: 93-113, 1999.
Cavagna GA, Heglund NC, and Taylor CR. Mechanical work in terrestrial locomotion: two
basic mechanisms for minimizing energy expenditure. The American journal of physiology 233:
R243-261, 1977.
Cavanagh P, Derr J, Ulbrecht J, Maser R, and Orchard T. Problems with gait and posture
in neuropathic patients with insulin-dependent diabetes mellitus. Diabet Med 9: 469-474, 1992.
Chanaud CM, and Macpherson JM. Functionally complex muscles of the cat hindlimb. 3.
Differential activation within biceps femoris during postural perturbations. Exp Brain Res 85:
271-280, 1991.
Cheung VCK, d'Avella A, Tresch MC, and Bizzi E. Central and sensory contributions to the activation and organization of muscle synergies during natural motor behaviors. Journal of
Neuroscience 25: 6419-6434, 2005.
Chiel HJ, Ting LH, Ekeberg O, and Hartmann MJZ. The brain in its body: motor control
and sensing in a biomechanical context. Journal of Neuroscience 29: 12807-12814, 2009.
Chvatal SA, Torres-Oviedo G, Safavynia SA, and Ting LH. Common muscle synergies for
control of center of mass and force in nonstepping and stepping postural behaviors. Journal of
neurophysiology 106: 999-1015, 2011.
Commissaris DA, Toussaint HM, and Hirschfeld H. Anticipatory postural adjustments in a
bimanual, whole-body lifting task seem not only aimed at minimising anterior--posterior centre
of mass displacements. Gait & Posture 14: 44-55, 2001.

185
Cordo PJ, and Nashner LM. Properties of postural adjustments associated with rapid arm
movements. Journal of neurophysiology 47: 287-302, 1982.
d'Avella A, and Bizzi E. Shared and specific muscle synergies in natural motor behaviors.
Proceedings of the National Academy of Sciences of the United States of America 102: 3076-
3081, 2005.
d'Avella A, Portone A, and Lacquaniti F. Superposition and modulation of muscle synergies for reaching in response to a change in target location. Journal of neurophysiology 106: 2796-
2812, 2011.
Danion F, and Sarlegna FR. Can the human brain predict the consequences of arm movement
corrections when transporting an object? Hints from grip force adjustments. Journal of
Neuroscience 27: 12839-12843, 2007.
Davidson PR, and Wolpert DM. Widespread access to predictive models in the motor system:
a short review. Journal of neural engineering 2: S313-319, 2005.
Day BI, and Brown P. Evidence for subcortical involvement in the visual control of human
reaching. Brain 124: 1832-1840, 2001.
Day BL, and Lyon IN. Voluntary modification of automatic arm movements evoked by
motion of a visual target. Experimental brain research 130: 159-168, 2000.
Day BL, Severac Cauquil A, Bartolomei L, Pastor MA, and Lyon IN. Human body-segment tilts induced by galvanic stimulation: a vestibularly driven balance protection mechanism. J
Physiol 500 ( Pt 3): 661-672, 1997.
Deliagina TG, Beloozerova IN, Zelenin PV, and Orlovsky GN. Spinal and supraspinal
postural networks. Brain research reviews 57: 212-221, 2008.
Desmurget M, and Grafton S. Forward modeling allows feedback control for fast reaching
movements. Trends in cognitive sciences 4: 423-431, 2000.
Diener HC, Dichgans J, Bootz F, and Bacher M. Early stabilization of human posture after a
sudden disturbance: influence of rate and amplitude of displacement. Experimental brain
research 56: 126-134, 1984a.
Diener HC, Dichgans J, Guschlbauer B, and Mau H. The significance of proprioception on
postural stabilization as assessed by ischemia. Brain research 296: 103-109, 1984b.
Drew T, Cabana T, and Rossignol S. Responses of medullary reticulospinal neurones to
stimulation of cutaneous limb nerves during locomotion in intact cats. Experimental Brain
Research 111: 153-168, 1996.
Drew T, Jiang W, and Widajewicz W. Contributions of the motor cortex to the control of the
hindlimbs during locomotion in the cat. Brain research Brain research reviews 40: 178-191,
2002.
Drew T, Kalaska J, and Krouchev N. Muscle synergies during locomotion in the cat: a model
for motor cortex control. The Journal of Physiology 586: 1239-1245, 2008.
Drew T, Prentice S, and Schepens B. Cortical and brainstem control of locomotion. Progress
in brain research 143: 251-261, 2004.

186
Duff SV, and Sainburg RL. Lateralization of motor adaptation reveals independence in
control of trajectory and steady-state position. Experimental brain research 179: 551-561,
2007.
Eccles JC, and Sherrington CS. Reflex summation in the ipsilateral spinal flexion reflex. The
Journal of Physiology 69: 1-28, 1930.
Elliott D, Garson RG, Goodman D, and Chua R. Discrete vs. continuous visual control of manual aiming. Human Movement Science 10: 393-418, 1991.
Fautrelle L, Prablanc C, Berret B, Ballay Y, and Bonnetblanc F. Pointing to double-step
visual stimuli from a standing position: very short latency (express) corrections are observed in
upper and lower limbs and may not require cortical involvement. Neuroscience 169: 697-705,
2010.
Flanders M, Daghestani L, and Berthoz A. Reaching beyond reach. Experimental brain
research 126: 19-30, 1999.
Flanders M, and Soechting JF. Arm muscle activation for static forces in three-dimensional
space. Journal of neurophysiology 64: 1818-1837, 1990.
Gahery Y, and Nieoullon A. Postural and kinetic coordination following cortical stimuli which
induce flexion movements in the cat's limbs. Brain research 149: 25-37, 1978.
Georgopoulos AP. On reaching. Annual review of neuroscience 9: 147-170, 1986.
Georgopoulos AP, Kalaska JF, and Massey JT. Spatial trajectories and reaction times of
aimed movements: effects of practice, uncertainty, and change in target location. J
Neurophysiol 46: 725-743, 1981.
Ghez C, and Krakauer JW. The organization of movement. In: Principles of neural science,
edited by Kandel ER, Schwartz JH, and Jessell TM. New York: McGraw-Hill, 2000, p. 653-
673.
Gielen CC, van den Heuvel PJ, and van Gisbergen JA. Coordination of fast eye and arm
movements in a tracking task. Experimental brain research 56: 154-161, 1984.
Goodale MA, Pelisson D, and Prablanc C. Large adjustments in visually guided reaching do
not depend on vision of the hand or perception of target displacement. Nature 320: 748-750,
1986.
Gorska T, Bem T, and Majczynski H. Locomotion in Cats with Ventral Spinal Lesions -
Support Patterns and Duration of Support Phases During Unrestrained Walking. Acta Neurobiol
Exp 50: 191-200, 1990.
Gorska T, Ioffe M, Zmyslowski W, Bem T, Majczynski H, and Mats VN. Unrestrained
walking in cats with medial pontine reticular lesions. Brain Research Bulletin 38: 297-304,
1995.
Grillner S. Biological pattern generation: the cellular and computational logic of networks in
motion. Neuron 52: 751-766, 2006.
Grillner S. The motor infrastructure: from ion channels to neuronal networks. Nature Reviews
Neuroscience 4: 573-586, 2003.

187
Gritsenko V, Yakovenko S, and Kalaska JF. Integration of predictive feedforward and
sensory feedback signals for online control of visually guided movement. Journal of
neurophysiology 102: 914-930, 2009.
Gutierrez-Farewik EM, Bartonek A, and Saraste H. Comparison and evaluation of two
common methods to measure center of mass displacement in three dimensions during gait.
Human Movement Science 25: 238-256, 2006.
Hart CB, and Giszter SF. Modular premotor drives and unit bursts as primitives for frog
motor behaviors. Journal of Neuroscience 24: 5269-5282, 2004.
Hayes KC. Biomechanics of postural control. Exercise and sport sciences reviews 10: 363-391,
1982.
Henry SM, Fung J, and Horak FB. Control of stance during lateral and anterior/posterior
surface translations. IEEE transactions on rehabilitation engineering : a publication of the
IEEE Engineering in Medicine and Biology Society 6: 32-42, 1998a.
Henry SM, Fung J, and Horak FB. Effect of stance width on multidirectional postural
responses. Journal of neurophysiology 85: 559-570, 2001.
Henry SM, Fung J, and Horak FB. EMG responses to maintain stance during multidirectional
surface translations. Journal of neurophysiology 80: 1939-1950, 1998b.
Herrmann U, and Flanders M. Directional tuning of single motor units. The Journal of neuroscience : the official journal of the Society for Neuroscience 18: 8402-8416, 1998.
Hess W. Telekinetisches und ereismatisches r ftesystem in der Biomotorik. . Helv Physiol
Pharmacol Acta 1: C62-C63, 1954.
Hess W. Teleokinetisches und ereismatisches Kräftesystem in der Biomotorik? Helv Physiol
Pharmac Acta 1: C62-C63, 1943.
Hollands MA, Ziavra NV, and Bronstein AM. A new paradigm to investigate the roles of
head and eye movements in the coordination of whole-body movements. Experimental brain
research 154: 261-266, 2004.
Hollerbach MJ, and Flash T. Dynamic interactions between limb segments during planar arm
movement. Biological Cybernetics 44: 67-77, 1982.
Holmquist B, and Sandberg R. The broken axis approach - a new way to analyze bi-directional circular data. Experientia 47: 845-851, 1991.
Honeycutt CF, Gottschall JS, and Nichols TR. Electromyographic responses from the
hindlimb muscles of the decerebrate cat to horizontal support surface perturbations. Journal of
neurophysiology 101: 2751-2761, 2009.
Honeycutt CF, and Nichols TR. The decerebrate cat generates the essential features of the
force constraint strategy. Journal of neurophysiology 103: 3266-3273, 2010.
Horak FB, Diener HC, and Nashner LM. Influence of central set on human postural
responses. Journal of neurophysiology 62: 841-853, 1989a.

188
Horak FB, and Macpherson JM. Postural orientation and equilibrium. In: Handbook of
Physiology Section 12: Exercise: Regulation and Integration of Multiple Systems, edited by
Rowell LB, and Shepherd JT. New York, NY: Oxford Univ Press, 1996, p. 255-292.
Horak FB, and Moore SP. The effect of prior leaning on human postural responses. Gait &
Posture 1: 203-210, 1993.
Horak FB, and Nashner LM. Central programming of postural movements: adaptation to altered support-surface configurations. Journal of neurophysiology 55: 1369-1381, 1986.
Horak FB, Nashner LM, and Diener HC. Postural strategies associated with somatosensory
and vestibular loss. Experimental brain research 82: 167-177, 1990.
Horak FB, Shupert CL, and Mirka A. Components of postural dyscontrol in the elderly: a
review. Neurobiology of aging 10: 727-738, 1989b.
Inglis JT, and Macpherson JM. Bilateral labyrinthectomy in the cat: effects on the postural
response to translation. Journal of neurophysiology 73: 1181-1191, 1995.
Inglis JT, and Macpherson JM. The role of the cat vestibular system in the response to linear
acceleration of the support surface. Neurosci Abstr 14: 63, 1988.
Ivanenko YP, Cappellini G, Dominici N, Poppele RE, and Lacquaniti F. Coordination of
locomotion with voluntary movements in humans. Journal of Neuroscience 25: 7238-7253,
2005.
Johansson RS, and Westling G. Roles of glabrous skin receptors and sensorimotor memory in
automatic control of precision grip when lifting rougher or more slippery objects. Exp Brain
Res 56: 550-564, 1984.
Joseph J, and Nightingale A. Electromyography of muscles of posture: Leg muscles in males.
J Physiol 117: 484-491, 1952.
Kably B, and Drew T. Corticoreticular pathways in the cat. I. Projection patterns and
collaterization. Journal of neurophysiology 80: 389-405, 1998.
Kawato M. Internal models for motor control and trajectory planning. Current Opinion in
Neurobiology 9: 718-727, 1999.
Klous M, Mikulic P, and Latash ML. Early postural adjustments in preparation to whole-
body voluntary sway. Journal of electromyography and kinesiology : official journal of the International Society of Electrophysiological Kinesiology 22: 110-116, 2012.
Komilis E, Pélisson D, and Prablanc C. Error processing in pointing at randomly feedback-
induced double-step stimuli. Journal of motor behavior 25: 299-308, 1993.
Krishnamoorthy V, Goodman S, Zatsiorsky V, and Latash ML. Muscle synergies during
shifts of the center of pressure by standing persons: identification of muscle modes. Biological
Cybernetics 89: 152-161, 2003.
Krishnamoorthy V, Latash ML, Scholz JP, and Zatsiorsky VM. Muscle modes during shifts
of the center of pressure by standing persons: effect of instability and additional support.
Experimental brain research Experimentelle Hirnforschung Expérimentation cérébrale 157:
18-31, 2004.

189
Kuo AD. An optimal state estimation model of sensory integration in human postural balance.
Journal of neural engineering 2: S235-249, 2005.
Latash ML. The bliss (not the problem) of motor abundance (not redundancy). Experimental
brain research Experimentelle Hirnforschung Expérimentation cérébrale 2012.
Lawrence DG, and Kuypers HGJM. The functional organization of the motor system in the
monkey. II. The effects of lesions of the descending brain-stem pathways. Brain 91: 15-36, 1968.
Lee DD, and Seung HS. Learning the parts of objects by non-negative matrix factorization.
Nature 401: 788-791, 1999.
Lee WA, Buchanan TS, and Rogers MW. Effects of arm acceleration and behavioral
conditions on the organization of postural adjustments during arm flexion. Experimental brain
research Experimentelle Hirnforschung Expérimentation cérébrale 66: 257-270, 1987.
Leonard JA, Brown RH, and Stapley PJ. Reaching to multiple targets when standing: the
spatial organization of feedforward postural adjustments. Journal of neurophysiology 101:
2120-2133, 2009.
Leonard JA, Gritsenko V, Ouckama R, and Stapley PJ. Postural adjustments for online
corrections of arm movements in standing humans. Journal of neurophysiology 105: 2375-
2388, 2011.
Lepers R, and Brenière Y. The role of anticipatory postural adjustments and gravity in gait
initiation. Experimental brain research Experimentelle Hirnforschung Expérimentation
cérébrale 107: 118-124, 1995.
Levine AJ, Lewallen KA, and Pfaff SL. Spatial organization of cortical and spinal neurons
controlling motor behavior. Current Opinion in Neurobiology 2012.
Liddell EGT, and Sherrington C. Reflexes in Response to Stretch (Myotatic Reflexes).
Proceedings of the Royal Society B: Biological Sciences 96: 212-242, 1924.
Lockhart DB, and Ting LH. Optimal sensorimotor transformations for balance. Nature
Neuroscience 10: 1329-1336, 2007.
Luccarini P, Gahery Y, and Pompeiano O. Cholinoceptive pontine reticular structures
modify the postural adjustments during the limb movement induced by cortical stimulation. Arch Ital Biol 128: 19-45, 1990.
Mackinnon CD, Bissig D, Chiusano J, Miller E, Rudnick L, Jager C, Zhang Y, Mille M-L,
and Rogers MW. Preparation of anticipatory postural adjustments prior to stepping. Journal of
neurophysiology 97: 4368-4379, 2007.
Macpherson J. How flexible are muscle synergies? In: Motor control: Concepts and Issues,
edited by Humphrey DR, and Freund H-J. UK: John Wiley & Sons, 1991, p. 33-47.
Macpherson JM. Strategies that simplify the control of quadrupedal stance. I. Forces at the
ground. Journal of neurophysiology 60: 204-217, 1988a.
Macpherson JM. Strategies that simplify the control of quadrupedal stance. II.
Electromyographic activity. Journal of neurophysiology 60: 218-231, 1988b.

190
Macpherson JM. The force constraint strategy for stance is independent of prior experience.
Experimental brain research Experimentelle Hirnforschung Expérimentation cérébrale 101:
397-405, 1994.
Macpherson JM, Everaert DG, Stapley PJ, and Ting LH. Bilateral Vestibular Loss in Cats
Leads to Active Destabilization of Balance During Pitch and Roll Rotations of the Support
Surface. Journal of neurophysiology 97: 4357-4367, 2007.
Macpherson JM, and Fung J. Weight support and balance during perturbed stance in the
chronic spinal cat. Journal of neurophysiology 82: 3066-3081, 1999.
Macpherson JM, Fung J, and Jacobs R. Postural orientation, equilibrium and the spinal cord.
In: Neuronal Regeneration Reorganization and Repair, edited by Seil F. Philadelphia:
Lippincott-Raven, 1997, p. 227-232.
Maki BE, and McIlroy WE. The role of limb movements in maintaining upright stance: the
"change-in-support" strategy. Physical Therapy 77: 488-507, 1997.
Mansfield A, and Maki BE. Are age-related impairments in change-in-support balance
reactions dependent on the method of balance perturbation? Journal of Biomechanics 42: 1023-
1031, 2009.
Martin O, Teasdale N, Simoneau M, Corbeil P, and Bourdin C. Pointing to a target from an
upright position in human: tuning of postural responses when there is target uncertainty. Neuroscience letters 281: 53-56, 2000.
Massion J. Movement, posture and equilibrium: Interaction and coordination. Progress in
neurobiology 38: 35-56, 1992.
Massion J. Postural control systems in developmental perspective. Neuroscience and
biobehavioral reviews 22: 465-472, 1998.
Massion J, Alexandrov A, and Frolov A. Why and how are posture and movement
coordinated? Progress in brain research 143: 13-27, 2004.
Massion J, and Woollacott MH. Posture and equilibrium. 2004, p. 1-19.
Maurer C, Mergner T, Bolha B, and Hlavacka F. Human balance control during cutaneous
stimulation of the plantar soles. Neuroscience letters 302: 45-48, 2001.
McKay JL, Burkholder TJ, and Ting LH. Biomechanical capabilities influence postural control strategies in the cat hindlimb. Journal of Biomechanics 40: 2254-2260, 2007.
McKay JL, and Ting LH. Functional muscle synergies constrain force production during
postural tasks. Journal of Biomechanics 41: 299-306, 2008.
McVea DA, and Pearson KG. Object avoidance during locomotion. Advances in experimental
medicine and biology 629: 293-315, 2009.
Megaw ED. Possible modification to a rapid on-going programmed manual response. Brain
research 71: 425-441, 1974.
Mehta B, and Schaal S. Forward Models in Visuomotor Control. Journal of neurophysiology
88: 942-953, 2002.

191
Mergner T, and Rosemeier T. Interaction of vestibular, somatosensory and visual signals for
postural control and motion perception under terrestrial and microgravity conditions--a
conceptual model. Brain research Brain research reviews 28: 118-135, 1998.
Milner TE. A model for the generation of movements requiring endpoint precision.
Neuroscience 49: 487-496, 1992.
Mink JW. The basal ganglia: focused selection and inhibition of competing motor programs. Progress in neurobiology 50: 381-425, 1996.
Moore SP, Rushmer DS, Windus SL, and Nashner LM. Human automatic postural
responses: responses to horizontal perturbations of stance in multiple directions. Experimental
brain research Experimentelle Hirnforschung Expérimentation cérébrale 73: 648-658, 1988.
Morasso PG, and Schieppati M. Can muscle stiffness alone stabilize upright standing? J
Neurophysiol 82: 1622-1626, 1999.
Mouchnino L, Aurenty R, Massion J, and Pedotti A. Coordination between equilibrium and
head-trunk orientation during leg movement: a new strategy build up by training. Journal of
neurophysiology 67: 1587-1598, 1992.
Muceli S, Boye AT, d'Avella A, and Farina D. Identifying representative synergy matrixes
for describing muscular activation patterns during multi-directional reaching in the horizontal
plane. Journal of neurophysiology 2010.
Nardone A, and Schieppati M. Postural adjustments associated with voluntary contraction of
leg muscles in standing man. Experimental brain research Experimentelle Hirnforschung
Expérimentation cérébrale 69: 469-480, 1988.
Nashner LM. Fixed patterns of rapid postural responses among leg muscles during stance.
Experimental brain research Experimentelle Hirnforschung Expérimentation cérébrale 30: 13-
24, 1977.
Nashner LM, Black FO, and Wall C. Adaptation to altered support and visual conditions
during stance: patients with vestibular deficits. The Journal of neuroscience : the official
journal of the Society for Neuroscience 2: 536-544, 1982.
Nijhof E. On-line trajectory modifications of planar, goal-directed arm movements. Human
Movement Science 22: 13-36, 2003.
Paulignan Y, Dufossé M, Hugon M, and Massion J. Acquisition of co-ordination between
posture and movement in a bimanual task. Experimental brain research Experimentelle
Hirnforschung Expérimentation cérébrale 77: 337-348, 1989.
Paulignan Y, MacKenzie C, Marteniuk R, and Jeannerod M. The coupling of arm and
finger movements during prehension. Experimental brain research Experimentelle
Hirnforschung Expérimentation cérébrale 79: 431-435, 1990.
Pearson KG, and Gordon J. Spinal reflexes. In: Principles of Neural Science, edited by
Kandel ER, Schwartz JH, and Jessell TM. New York: McGraw Hill, 2000, p. 713-736.
Pélisson D, Prablanc C, Goodale MA, and Jeannerod M. Visual control of reaching
movements without vision of the limb. II. Evidence of fast unconscious processes correcting the
trajectory of the hand to the final position of a double-step stimulus. Experimental brain
research Experimentelle Hirnforschung Expérimentation cérébrale 62: 303-311, 1986.

192
Perfiliev S. Bilateral processing of motor commands in the motor cortex of the cat during
target-reaching. Journal of neurophysiology 93: 2489-2506, 2005.
Peterka RJ. Sensorimotor integration in human postural control. Journal of neurophysiology
88: 1097-1118, 2002.
Prablanc C, and Martin O. Automatic control during hand reaching at undetected two-
dimensional target displacements. Journal of neurophysiology 67: 455-469, 1992.
Prablanc C, Pélisson D, and Goodale MA. Visual control of reaching movements without
vision of the limb. I. Role of retinal feedback of target position in guiding the hand.
Experimental brain research Experimentelle Hirnforschung Expérimentation cérébrale 62:
293-302, 1986.
Purves D, Augustine GJ, Fitzpatrick D, Hall WC, Lamantia A-S, and White LE.
Neuroscience. Sinauer Associates, 2012, p. 858.
Reynolds R, and Day B. Rapid visuo-motor processes drive the leg regardless of balance
constraints. Current biology : CB 15: R48-49, 2005a.
Reynolds R, and Day B. Visual guidance of the human foot during a step. The Journal of
Physiology 569: 677-684, 2005b.
Rogers MW, and Pai YC. Dynamic transitions in stance support accompanying leg flexion
movements in man. Experimental brain research Experimentelle Hirnforschung Expérimentation cérébrale 81: 398-402, 1990.
Roh J, Cheung VCK, and Bizzi E. Modules in the brain stem and spinal cord underlying
motor behaviors. Journal of neurophysiology 106: 1363-1378, 2011.
Rossetti Y, Desmurget M, and Prablanc C. Vectorial coding of movement: vision,
proprioception, or both? Journal of neurophysiology 74: 457-463, 1995.
Rossignol S, Bouyer L, Barthélemy D, Langlet C, and Leblond H. Recovery of locomotion
in the cat following spinal cord lesions. Brain research Brain research reviews 40: 257-266,
2002.
Rossignol S, Dubuc R, and Gossard J-P. Dynamic sensorimotor interactions in locomotion.
Physiological reviews 86: 89-154, 2006.
Ruget H, Blouin J, Teasdale N, and Mouchnino L. Can prepared anticipatory postural adjustments be updated by proprioception? Neuroscience 155: 640-648, 2008.
Sabes PN. The planning and control of reaching movements. Current Opinion in Neurobiology
10: 740-746, 2000.
Safavynia SA, and Ting LH. Task-level feedback can explain temporal recruitment of
spatially-fixed muscle synergies throughout postural perturbations. Journal of neurophysiology
2011.
Safavynia SA, Torres-Oviedo G, and Ting LH. Muscle Synergies: Implications for Clinical
Evaluation and Rehabilitation of Movement. Topics in spinal cord injury rehabilitation 17: 16-
24, 2011.
Saltzman E. Levels of sensorimotor representation. J Math Psychol 20: 91-163, 1979.

193
Santos MJ, and Aruin AS. Role of lateral muscles and body orientation in feedforward
postural control. Experimental brain research Experimentelle Hirnforschung Expérimentation
cérébrale 184: 547-559, 2008.
Sarlegna FR, and Sainburg RL. The roles of vision and proprioception in the planning of
reaching movements. Advances in experimental medicine and biology 629: 317-335, 2009.
Saunders J, and Knill D. Humans use continuous visual feedback from the hand to control fast reaching movements. Experimental brain research Experimentelle Hirnforschung
Expérimentation cérébrale 152: 341-352, 2003.
Schepens B, and Drew T. Descending signals from the pontomedullary reticular formation are
bilateral, asymmetric, and gated during reaching movements in the cat. Journal of
neurophysiology 96: 2229-2252, 2006.
Schepens B, and Drew T. Independent and convergent signals from the pontomedullary
reticular formation contribute to the control of posture and movement during reaching in the
cat. Journal of neurophysiology 92: 2217-2238, 2004.
Schepens B, and Drew T. Strategies for the integration of posture and movement during
reaching in the cat. Journal of neurophysiology 90: 3066-3086, 2003.
Schepens B, Stapley P, and Drew T. Neurons in the pontomedullary reticular formation signal
posture and movement both as an integrated behavior and independently. Journal of neurophysiology 100: 2235-2253, 2008.
Scott SH, Sergio LE, and Kalaska JF. Reaching movements with similar hand paths but
different arm orientations. II. Activity of individual cells in dorsal premotor cortex and parietal
area 5. Journal of neurophysiology 78: 2413-2426, 1997.
Sergio LE, and Kalaska JF. Changes in the temporal pattern of primary motor cortex activity
in a directional isometric force versus limb movement task. Journal of neurophysiology 80:
1577-1583, 1998.
Shabbott BA, and Sainburg RL. On-line corrections for visuomotor errors. Experimental
brain research Experimentelle Hirnforschung Expérimentation cérébrale 195: 59-72, 2009.
Shadmehr R, and Krakauer JW. A computational neuroanatomy for motor control.
Experimental brain research Experimentelle Hirnforschung Expérimentation cérébrale 185: 359-381, 2008.
Sherrington C. The Integrative Action of the Nervous System. New Haven: Yale University
Press, 1961, p. 413.
Sherrington CS. A mammalian spinal preparation. The Journal of Physiology 38: 375-383,
1909.
Shim JK, Olafsdottir H, Zatsiorsky VM, and Latash ML. The emergence and disappearance
of multi-digit synergies during force-production tasks. Experimental brain research 164: 260-
270, 2005.
Soechting JF. Effect of target size on spatial and temporal characteristics of a pointing
movement in man. Experimental brain research Experimentelle Hirnforschung
Expérimentation cérébrale 54: 121-132, 1984.

194
Soechting JF, and Lacquaniti F. Modification of trajectory of a pointing movement in
response to a change in target location. Journal of neurophysiology 49: 548-564, 1983.
Stapley PJ, and Drew T. The pontomedullary reticular formation contributes to the
compensatory postural responses observed following removal of the support surface in the
standing cat. Journal of neurophysiology 101: 1334-1350, 2009.
Stapley PJ, Pozzo T, Cheron G, and Grishin A. Does the coordination between posture and movement during human whole-body reaching ensure center of mass stabilization?
Experimental brain research Experimentelle Hirnforschung Expérimentation cérébrale 129:
134-146, 1999.
Stapley PJ, Ting LH, Hulliger M, and Macpherson JM. Automatic postural responses are
delayed by pyridoxine-induced somatosensory loss. Journal of Neuroscience 22: 5803-5807,
2002.
Thoroughman KA, and Shadmehr R. Electromyographic correlates of learning an internal
model of reaching movements. Journal of Neuroscience 19: 8573-8588, 1999.
Ting LH. Dimensional reduction in sensorimotor systems: a framework for understanding
muscle coordination of posture. Progress in brain research 165: 299-321, 2007.
Ting LH, and Chvatal SA. Decomposing muscle activity in motor tasks. In: Motor Control:
Theories, Experiments, and Applications, edited by Latash ML, and Danion FOxford University Press, 2010, p. 102-140.
Ting LH, Kautz SA, Brown DA, and Zajac FE. Phase reversal of biomechanical functions
and muscle activity in backward pedaling. Journal of neurophysiology 81: 544-551, 1999.
Ting LH, and Macpherson JM. A limited set of muscle synergies for force control during a
postural task. Journal of neurophysiology 93: 609-613, 2005.
Ting LH, and Macpherson JM. Ratio of shear to load ground-reaction force may underlie the
directional tuning of the automatic postural response to rotation and translation. Journal of
neurophysiology 92: 808-823, 2004.
Ting LH, and Mckay JL. Neuromechanics of muscle synergies for posture and movement.
Current Opinion in Neurobiology 17: 622-628, 2007.
Tokuno CD, Cresswell AG, Thorstensson A, and Carpenter MG. Age-related changes in postural responses revealed by support-surface translations with a long acceleration-
deceleration interval. Clinical neurophysiology : official journal of the International Federation
of Clinical Neurophysiology 121: 109-117, 2010.
Torres-Oviedo G, Macpherson JM, and Ting LH. Muscle synergy organization is robust
across a variety of postural perturbations. Journal of neurophysiology 96: 1530-1546, 2006.
Torres-Oviedo G, and Ting LH. Muscle synergies characterizing human postural responses.
Journal of neurophysiology 98: 2144-2156, 2007.
Torres-Oviedo G, and Ting LH. Subject-specific muscle synergies in human balance control
are consistent across different biomechanical contexts. Journal of neurophysiology 103: 3084-
3098, 2010.

195
Tresch MC, Cheung VCK, and d'Avella A. Matrix factorization algorithms for the
identification of muscle synergies: evaluation on simulated and experimental data sets. Journal
of neurophysiology 95: 2199-2212, 2006.
Tresch MC, and Jarc A. The case for and against muscle synergies. Current Opinion in
Neurobiology 19: 601-607, 2009.
Tresch MC, Saltiel P, and Bizzi E. The construction of movement by the spinal cord. Nature Neuroscience 2: 162-167, 1999.
van Ingen Schenau GJ, Boots PJ, de Groot G, Snackers RJ, and van Woensel WW. The
constrained control of force and position in multi-joint movements. Neuroscience 46: 197-207,
1992.
Viallet F, Massion J, Massarino R, and Khalil R. Coordination between posture and
movement in a bimanual load lifting task: putative role of a medial frontal region including the
supplementary motor area. Experimental brain research Experimentelle Hirnforschung
Expérimentation cérébrale 88: 674-684, 1992.
von Holst E, and Mittelstaedt H. Das reafferenzprinzip [Translation of title: The principle of
reafference]. Naturwissenschaften 37: 464-476, 1950.
Welch TDJ, and Ting LH. A feedback model reproduces muscle activity during human
postural responses to support-surface translations. Journal of neurophysiology 99: 1032-1038, 2008.
Westheimer G. Eye movement responses to a horizontally moving visual stimulus. Arch
Ophthalmol 52: 932-941, 1954.
Wheeless LL, Boynton RM, and Cohen GH. Eye-movement responses to step and pulse-step
stimuli. Journal of the Optical Society of America 56: 956-960, 1966.
Winter DA. Biomechanics and Motor Control of Human Movement. John Wiley and Sons,
2009, p. 384.
Winter DA, and Eng P. Kinetics: our window into the goals and strategies of the central
nervous system. Behavioural brain research 67: 111-120, 1995.
Wolpert D, and Miall R. Forward Models for Physiological Motor Control. Neural networks :
the official journal of the International Neural Network Society 9: 1265-1279, 1996.
Woollacott MH. Changes in posture and voluntary control in the elderly: Research findings
and rehabilitation. Topics in Geriatric Rehabilitation 5: 1-11, 2006.
Yakovenko S, and Drew T. A motor cortical contribution to the anticipatory postural
adjustments that precede reaching in the cat. Journal of neurophysiology 102: 853-874, 2009.
Zar JH. Biostatistical Analysis. New Jersey: Prentice Hall, 1999, p. 663.
Zelaznik HZ, Hawkins B, and Kisselburgh L. Rapid visual feedback processing in single-
aiming movements. Journal of motor behavior 15: 217-236, 1983.

















