
The Fate of Inflorescence Meristems Is Controlled by Developing Fruits in Arabidopsis
14
Plant Physiol. (1994) 106: 863-876 The Fate of Inflorescence Meristems Is Controlled by Developing Fruits in Arabidopsis' linda 1. Hensel, Michelle A. Nelson, Todd A. Richmond, and Anthony B. Bleecker* Botany Department, Birge Hall, University of Wisconsin, Madison, Wisconsin 53706 ~~~ The relationshipbetween fruit development and the proliferative capacities of inflorescence meristems has been examined in Ara- bidopsis fhaliana. In the wild-type Landsberg erecta (Ler) line, flower production ceases coordinately on all inflorescence branches by a process we have designated global proliferative arrest (CPA). Morphological studies indicate that CPA involves a cessation of proliferative activity at the meristems, but a retention of the structural characteristicsof the proliferating meristems. CPA does not occur in the male-sterile (msl-7) line, nor in wild-type Ler when fruits are surgically removed. In these cases, inflorescence meristems continue to proliferate, ultimately terminating by a different process, designated terminal differentiation, in which disruptions in patterning at the apex are followed by the loss of the inflorescence meristem. We present an argument that CPA is mediated by a specific communication system between infloresc- ence meristems and developing fruits. Analysis of reduced-fertility mutants provided evidence that CPA is dependent on seed devel- opment specifically. Mutations conferring hormone deficiency or insensitivity did not disrupt the correlative interactions leading to CPA. The term 'monocarpy" refers to the life history pattem, common to many annual and some perennial plant species, in which the plant senesces and dies after a single reproduc- tive effort. The monocarpic habit is of particular interest agriculturally in that the majority of crop varieties exhibit this trait (reviewed by Noodén, 1988b).Early observations of monocarpic behavior led Hildebrand (1881) to postulate that the whole plant senescence associated with reproduction occurs as a consequence of the diversion of nutrients from vegetative to reproductive structures. Although this premise is still generally accepted, the underlying regulatory mecha- nisms that determine the fate of the parent plant's tissues during the reproductive phase in monocarpic plants are not well understood. Two important processes are generally associated with monocarpy: (a) the suppression of vegetative growth, and (b) the senescence of existing somatic tissues during the repro- ' The work presented here was funded by-grantsfrom the National Science Foundation (No. DMB-9005164), the National Institute on Aging (No. 5F32AG05542-02), and the Department of Energy/ National Science Foundation/U.S. Department of Agriculture Col- laborative Research in Plant Biology Program (No. BIR92-20331). M.A.N. is supported by the Genetics Department Training grant (National Institutes of Health No. GM07133-181). T.A.R. is sup- ported by a National Science Foundation Graduate Fellowship. Corresponding author; fax 1-608-262-7509. ductive phase. The suppression of vegetative development in some monocarpic species results from the conversion of all indeterminate vegetative meristems into determinate flower meristems (Woolhouse, 1983).In other cases, vegetative mer- istems are retained but their proliferative activity is sup- pressed at the time of flowering (Lockhart and Gottschall, 1961; Proebsting et al., 1976). On the other hand, the senes- cence of existing somatic tissues in monocarpic plants, partic- ularly of photosynthetic tissues, occurs through the action of a programmed cell-death process referred to as the senes- cence syndrome (see Noodén, 1988a). The senescence syn- drome is associated with the induction of salvage pathways that convert macromolecules and lipids into transportable metabolites prior to the death of the senescing tissues (Wool- house, 1984; Gut and Matile, 1988; Noodén, 1988a). Both the suppression of vegetative growth and the timely senes- cence of existing somatic tissues may have adaptive value in terms of providing resources for developing seeds. However, in the case of monocarpy these processes also lead to the death of the whole organism. Observations that surgical removal of developing fruits delayed the onset of monocarpic senescence in a number of plants provided the first evidence for the existence of some form of communication system between developing fruits and the tissues of the parent plant (Murneek, 1926; Molisch, 1928). These interactions were referred to as correlative con- trols, a term that continues to describe accurately the extent of our understanding of the relationship between cause and effect for these interactions. Correlative control models in- voking existing plant hormones (Woolhouse, 1983; reviewed by Noodén, 1988b),unidentified 'death hormones" (Noodén, 1988b; Engvild, 1989), and source-sink relationships (Mol- isch, 1928; Sinclair and de Wit, 1975; Kelly and Davies, 1988a) have been suggested-but never satisfactorily verified or refuted. Woolhouse (1983) has pointed out that, given the polyphyletic origin of monocarpic senescence, it is likely that a variety of different control mechanisms may have evolved independently in different plant groups. An excellent example of a monocarpic plant is the crucifer Arabidopsis thaliana, which may produce upward of 20,000 offspring within 10 weeks of germination. This high level of fecundity is achieved at the expense of the soma; the parent plant exhibits symptoms of advanced senescence as the seeds reach maturity (Hensel et al., 1993). The specific develop- mental processes that contribute to monocarpic senescence Abbreviations: GPA, global proliferative arrest; Ler, Landsberg erecta; SEM, scanning electron microscopy; WS, Wassilewskija. 863 Downloaded from https://academic.oup.com/plphys/article/106/3/863/6068706 by guest on 09 December 2021
Transcript of The Fate of Inflorescence Meristems Is Controlled by Developing Fruits in Arabidopsis

Plant Physiol. (1994) 106: 863-876
The Fate of Inflorescence Meristems I s Controlled by Developing Fruits in Arabidopsis'
linda 1. Hensel, Michelle A. Nelson, Todd A. Richmond, and Anthony B. Bleecker*
Botany Department, Birge Hall, University of Wisconsin, Madison, Wisconsin 53706
~~~
The relationship between fruit development and the proliferative capacities of inflorescence meristems has been examined in Ara- bidopsis fhaliana. In the wild-type Landsberg erecta (Ler) line, flower production ceases coordinately on all inflorescence branches by a process we have designated global proliferative arrest (CPA). Morphological studies indicate that CPA involves a cessation of proliferative activity at the meristems, but a retention of the structural characteristics of the proliferating meristems. CPA does not occur in the male-sterile (msl-7) line, nor in wild-type Ler when fruits are surgically removed. In these cases, inflorescence meristems continue to proliferate, ultimately terminating by a different process, designated terminal differentiation, in which disruptions in patterning at the apex are followed by the loss of the inflorescence meristem. We present an argument that CPA is mediated by a specific communication system between infloresc- ence meristems and developing fruits. Analysis of reduced-fertility mutants provided evidence that CPA is dependent on seed devel- opment specifically. Mutations conferring hormone deficiency or insensitivity did not disrupt the correlative interactions leading to CPA.
The term 'monocarpy" refers to the life history pattem, common to many annual and some perennial plant species, in which the plant senesces and dies after a single reproduc- tive effort. The monocarpic habit is of particular interest agriculturally in that the majority of crop varieties exhibit this trait (reviewed by Noodén, 1988b). Early observations of monocarpic behavior led Hildebrand (1881) to postulate that the whole plant senescence associated with reproduction occurs as a consequence of the diversion of nutrients from vegetative to reproductive structures. Although this premise is still generally accepted, the underlying regulatory mecha- nisms that determine the fate of the parent plant's tissues during the reproductive phase in monocarpic plants are not well understood.
Two important processes are generally associated with monocarpy: (a) the suppression of vegetative growth, and (b) the senescence of existing somatic tissues during the repro-
' The work presented here was funded by-grants from the National Science Foundation (No. DMB-9005164), the National Institute on Aging (No. 5F32AG05542-02), and the Department of Energy/ National Science Foundation/U.S. Department of Agriculture Col- laborative Research in Plant Biology Program (No. BIR92-20331). M.A.N. is supported by the Genetics Department Training grant (National Institutes of Health No. GM07133-181). T.A.R. is sup- ported by a National Science Foundation Graduate Fellowship.
Corresponding author; fax 1-608-262-7509.
ductive phase. The suppression of vegetative development in some monocarpic species results from the conversion of all indeterminate vegetative meristems into determinate flower meristems (Woolhouse, 1983). In other cases, vegetative mer- istems are retained but their proliferative activity is sup- pressed at the time of flowering (Lockhart and Gottschall, 1961; Proebsting et al., 1976). On the other hand, the senes- cence of existing somatic tissues in monocarpic plants, partic- ularly of photosynthetic tissues, occurs through the action of a programmed cell-death process referred to as the senes- cence syndrome (see Noodén, 1988a). The senescence syn- drome is associated with the induction of salvage pathways that convert macromolecules and lipids into transportable metabolites prior to the death of the senescing tissues (Wool- house, 1984; Gut and Matile, 1988; Noodén, 1988a). Both the suppression of vegetative growth and the timely senes- cence of existing somatic tissues may have adaptive value in terms of providing resources for developing seeds. However, in the case of monocarpy these processes also lead to the death of the whole organism.
Observations that surgical removal of developing fruits delayed the onset of monocarpic senescence in a number of plants provided the first evidence for the existence of some form of communication system between developing fruits and the tissues of the parent plant (Murneek, 1926; Molisch, 1928). These interactions were referred to as correlative con- trols, a term that continues to describe accurately the extent of our understanding of the relationship between cause and effect for these interactions. Correlative control models in- voking existing plant hormones (Woolhouse, 1983; reviewed by Noodén, 1988b), unidentified 'death hormones" (Noodén, 1988b; Engvild, 1989), and source-sink relationships (Mol- isch, 1928; Sinclair and de Wit, 1975; Kelly and Davies, 1988a) have been suggested-but never satisfactorily verified or refuted. Woolhouse (1983) has pointed out that, given the polyphyletic origin of monocarpic senescence, it is likely that a variety of different control mechanisms may have evolved independently in different plant groups.
An excellent example of a monocarpic plant is the crucifer Arabidopsis thaliana, which may produce upward of 20,000 offspring within 10 weeks of germination. This high level of fecundity is achieved at the expense of the soma; the parent plant exhibits symptoms of advanced senescence as the seeds reach maturity (Hensel et al., 1993). The specific develop- mental processes that contribute to monocarpic senescence
Abbreviations: GPA, global proliferative arrest; Ler, Landsberg erecta; SEM, scanning electron microscopy; WS, Wassilewskija.
863
Dow
nloaded from https://academ
ic.oup.com/plphys/article/106/3/863/6068706 by guest on 09 D
ecember 2021

864 Hensel et al. Plant Physiol. Vol. 106, 1994
in Arabidopsis are as follows: the shift in resource allocation from vegetative to reproductive development begins at the transition to flowering when the shoot apex converts from a leaf-producing meristem to a flower-producing meristem (Vaughan, 1955; Hempel and Feldman, 1994). Axillary mer- istems are subsequently initiated, but they also convert into flower-producing meristems after a brief vegetative phase (Hempel and Feldman, 1994). According to recent reports, the primary inflorescence meristems cease proliferative activ- ity after the production of about 30 flowers (Shannon and Meeks-Wagner, 1991; Alvarez et al., 1992). These stages in the reproductive phase of development are accompanied by the progressive senescence of existing somatic tissues (Hensel et al., 1993). The cessation of shoot meristem activity together with the attrition of existing somatic tissues provide a simple explanation for whole plant senescence in Arabidopsis. The specific regulatory mechanisms that coordinate these proc- esses are the subjects of our research efforts.
This well-defined life cycle of Arubidopsis, coupled with the availability of a large pool of hormonal and develop- mental mutants in isogenic backgrounds (Koomneef et al., 1983; Arubidopsis Biological Resource Center, 1994) and a growing body of research on virtually every aspect of plant development, provide unique opportunities for investigating the genetic mechanisms underlying monocarpic senescence in Arabidopsis. In a previous report we examined the relation- ship between fruit development and somatic tissue senes- cence in Arubidopsis (Hensel et al., 1993). We concluded that the timing of somatic senescence was age related and inde- pendent of the reproductive status of the plant. In this report, we examine the relationship between fruit development and the proliferative activity of inflorescence meristems. Based on initial observations that male-sterile mutants in Arubidopsis show an increase in the number of inflorescence branches and flowers per branch relative to wild type, we postulated that developing fruits normally exhibit correlative control over the capacity of inflorescence meristems to produce ad- ditional flowers. Here we present a comparative analysis of inflorescence development in wild-type Ler and an isogenic male-sterile line (msl-I) . We present morphological and ge- netic evidence that inflorescence meristem activity and thus total fecundity of the plant is regulated by interactions be- tween existing fruits and the inflorescence meristems.
MATERIALS A N D METHODS
Plant Material and Measurements
Seeds for Arabidopsis thaliana Ler and the isogenic mutant lines msl-I, ubi l , abiZ, and abi3 were obtained from A.R. Kranz (Kranz, 1978; Kirchheim and Kranz, 1981). Seeds for the late-flowering lines Id-3, gi, and ld-2 were obtained from R. Amasino (Lee et al., 1994). Columbia seeds were obtained from C. Somerville (Camegie Institute, Stanford, CA). etrl-l was isolated by Bleecker et al. (1988). Seeds for all the lines not mentioned above were obtained from the Arabidopsis Stock Center (Columbus, OH).
We have shown that Arabidopsis plants develop, grow, and senesce uniformly when environmental conditions are main- tained with little fluctuation (Hensel and Bleecker, 1992;
Hensel et al., 1993). Therefore, for the experiments described here, plants were grown in 50 to 75% RH, at 20 to 22OC, under 100 to 150 PIE m-’ s-l continuous light (unless stated otherwise), at a density of one plant per 25 cm’ and with continuous soil moisture as described (Hensel et al., 1993). The light banks were from 2 to 5 feet above the tops of the plants so that the tops and bottoms of the plants received a similar light intensity. Seeds were surface sterilized with 30% bleach/0.5% Triton and stratified for 1 to 2 d at 4OC prior to planting.
To determine plant dry weights, plants grown UI continu- ous light were harvested 50 d after planting. Planí s grown in 8 h light/l6 h dark were harvested 2 to 6 d after the msl-2 primary inflorescences had terminally differentiatc d. The aer- ial portions of plants of a given age were dried at 6OoC for >24 h before weighing. Leaf longevity was measured as described by Hensel et al. (1993).
For plants grown in 16-h and continuous-light cycles, flower production rates were determined by counting the number of siliques or infertile ovaries after petal abscission on the primary inflorescence at 24-h intervals. For plants grown in 8-h light cycles, mature ovaries were counted at 48-h intervals.
For pollination experiments, wild-type flowers were brushed against male-sterile flowers one to two times daily. Greater than 90% of the flowers that were pollinated pro- duced seed. As controls, both wild-type and male-sterile plants were hand pollinated with wild-type or male-sterile flowers, respectively. Floral production in hand-pollinated wild-type and mock-pollinated male-sterile plants was not significantly different from that in the untreated cmontrols.
Branch diagrams were formulated by first counting the number of 2 O , 3O, 4 O , etc. branches per plant and determining an average number of nth order branches per plant. Aerial 2 O
branches are defined as those that emerge from the aerial nodes of the primary inflorescence; basal 2 O branclies emerge from the basal rosette. Second, the average number of mature ovaries per nth order branch was determined. For the surgical experiments depicted in Figure 7, D and E, aborted and excised structures were included in the total number of struc- tures used to determine average branch length. For the sur- gical experiments, structures were removed at their base with surgical scissors, leaving the subtending pedicels or stem bases intact.
Light Microscopy
For general anatomical investigations, samples were fixed in 4% paraformaldehyde, 50 m~ potassium-phosphate, pH 7.0. Samples were dehydrated in a graded ethanol series and embedded in paraffin (Paraplast Plus, Sigma), and 8-PM serial sections were cut on a rotary microtome. The samples were stained with toluidine blue O and photographed using a Zeiss Axioskop.
SEM
For comparative analysis of floral bud development, the inflorescences of both active and arrested Ler were examined by SEM. Primary apical inflorescences were fixed in 2 to 4%
Dow
nloaded from https://academ
ic.oup.com/plphys/article/106/3/863/6068706 by guest on 09 D
ecember 2021

Meristem Fate in Arabidopsis 865
glutaraldehyde in 0.05 M potassium phosphate buffer, pH 7.0, at 4OC ovemight. Samples were rinsed in buffer, dehy- drated in a graded series of ethanol at 4OC, and critical-point dried in liquid carbon dioxide. Samples were partially dis- sected, mounted on stubs, and sputter-coated with gold. The samples were examined in a Hitachi S570 scanning electron microscope at an accelerating voltage of 15 kV. Photographs were taken using Polaroid type 55 film.
RESULTS
A Comparison of the Proliferative Capacity of Inflorescence Meristems from Wild-Type l e r and the Male-Sterile Isogenic l ine msl-1
It has been reported that the primary inflorescence meris- tem of the Arabidopsis plant becomes inactive after the pro- duction of 30 to 50 flowers (Shannon and Meeks-Wagner, 1991; Alvarez et al., 1992). To examine this phenomenon more closely, we monitored the flower production rate of the primary inflorescence meristem over the course of reproduc- tive development. As shown in Figure 1, the rate of flower production by the primary inflorescence meristem of wild- type plants remained constant for about 15 d, with a produc- tion rate of 2.6 flowers per d. At about 15 to 20 d after bolting, the rate of new flower production declined to zero within a few days. This phenomenon was observed in plants grown under constant light, a 16-h photoperiod, and even an 8-h photoperiod.
We investigated the relationship between meristem prolif- erative arrest and fruit development by comparing the Ler wild-type development with that of a male-sterile isogenic line, msl-l (Koomneef et al., 1983). Primary meristems of male-sterile plants initially showed a.flower production rate similar to that of wild-type plants. However, meristems of male-sterile plants continued to proliferate for 1 to 2 weeks longer than those of wild-type plants, producing almost twice as many flowers. Ultimately, the msl-l apical meristem did stop producing new flowers.
The cessation of meristem proliferative activity was not restricted to the primary inflorescence branch. In wild-type plants all lateral meristems became inactive within 2 d of the cessation of primary meristem activity (Fig. 2A). Apical arrest in lateral branches of the male-sterile plants occurred over a much longer time period (Fig. 28). Male-sterile plants also differed from wild-type plants in that they initiated many more higher-order branches. As indicated in Figure 2, fourth- order branches were relatively rare in wild-type plants, whereas fifth-order and higher-order flower-producing branches were common in msl-2.
Visual observations of developing wild-type and male- sterile flowering stalks revealed a number of additional dif- ferences between these lines. Figure 3 shows wild-type Ler plants shortly before (Fig. 3A) and shortly after (Fig. 3C) apical arrest. The lack of flower clusters at the ends of the inflorescence branches in Figure 3C illustrates the global nature of apical arrest in Ler. A closer inspection of the primary apex revealed the presence of floral buds at the arrested apex (Fig. 3D), but no mature flowers. Flowering stalks of male-sterile plants were initially similar in appear-
o male-sterile 0 wild-type
I
C
- 10 20 30
Figure 1. Flower production rates for primary meristems of wild- type (Ler) and male-sterile (msl-1) plants under different photo- periods. A, Plants grown under 8-h-light/l6-h-dark cycles. Between 14 and 28 aborted buds were produced prior to normal-appearing fruits or infertile ovules; these aborted buds are not shown. msl-1, n = 7; wild-type, n = 13. B, Plants grown under 16-h-light/8-h-dark cycles. msl-1, n = 11; wild-type, n = 22. C , Plants grown under continuous light. msl-1, n = 8; wild-type, n = 14. Averages * SE are graphed; SE values <2.0 are not shown.
ance to wild type (compare Fig. 3B with Fig. 3G). However, continued proliferative activity of msl-2 inflorescence mer- istems produced the large, highly branched inflorescence depicted in Figure 3H. The ultimate arrest of msl-l apices was not associated with a terminal cluster of arrested buds, as it was in wild type. Rather, we consistently observed fully developed structures with a variety of morphological abnor- malities at the inflorescence apex (Fig. 3, I and J). The most characteristic abnormalities were single, unfused carpel-like structures at the apex of each branch.
To examine the effects of increased biomass on the prolif- erative capacity of inflorescence meristems, Ler plants were
Dow
nloaded from https://academ
ic.oup.com/plphys/article/106/3/863/6068706 by guest on 09 D
ecember 2021

866 Hensel et al. Plant Physiol. Vol. 106, 1994
A
0.4
B
2
\
6 6
Figure 2. A schematic diagram indicating branching patterns and times of apical arrest for a typical secondary inflorescence branch from wild-type (Ler) and male-sterile (msl-I) plants. A, Ler. B, msl- I . The number above each branch indicates the average number of days between the arrest of the primary inflorescence and the arrest of the designated branch. For clarity only the primary and one secondary branch were included in this analysis. The vertical lines above the last branch point are drawn to scale according to the average number of mature ovaries produced by that order of branch. Branching patterns and times of apical arrest represent averages from 16 plants for Ler and 8 plants for msl-I.
gown under an 8-h photoperiod. As summarized in Table I, SD plants were substantially delayed in flowering time, showed an increase in longevity of somatic tissues, and as a consequence had accumulated much greater vegetative bio- mass at the time of flowering than LD plants (see also Fig. 3K). Despite this much greater total biomass, SD plants underwent apical arrest after the production of about 28 fruits on the primary inflorescence (Fig. 1A).
At about 2 weeks after global arrest of wild-type apices, 10 to 30% of primary inflorescence apices reinitiated prolif- erative activity and began to produce new floral structures (Fig. 3E). In these cases, the arrested buds already present at the apex did not complete development. However, new buds produced by this reinitiated meristem developed into fertile flowers and fruits. The ability to reinitiate inflorescence de- velopment indicates that the arrested meristem in wild-type plants is held in a quiescent rather than a degenerate state. Reinitiation was never observed for the primary apices of male-sterile plants.
Morphological Analysis of Arrested Apices
To gain a better understanding of the processes occurring at the inflorescence meristem associated with apical arrest, we examined the meristems of wild-type and male-sterile plants using SEM. As shown in Figure 4, the arrested Ler meristem retained characteristics of the younger proliferating
meristem. Floral primordia and buds at various stages of development were arranged in the normal spiral p,ittem (Fig. 4B), and the apical dome of the inflorescence meiistem was clearly discernible. Similar observations have been reported previously for the arrested apex of wild-type Arabidopsis plants (Shannon and Meeks-Wagner, 1991). A quantitative analysis of floral developmental stages in active arid arrested wild-type apices is provided in Table 11. All stages of floral development were represented in the arrested aper , although the number of buds at each stage was lower in arrested than in active meristems.
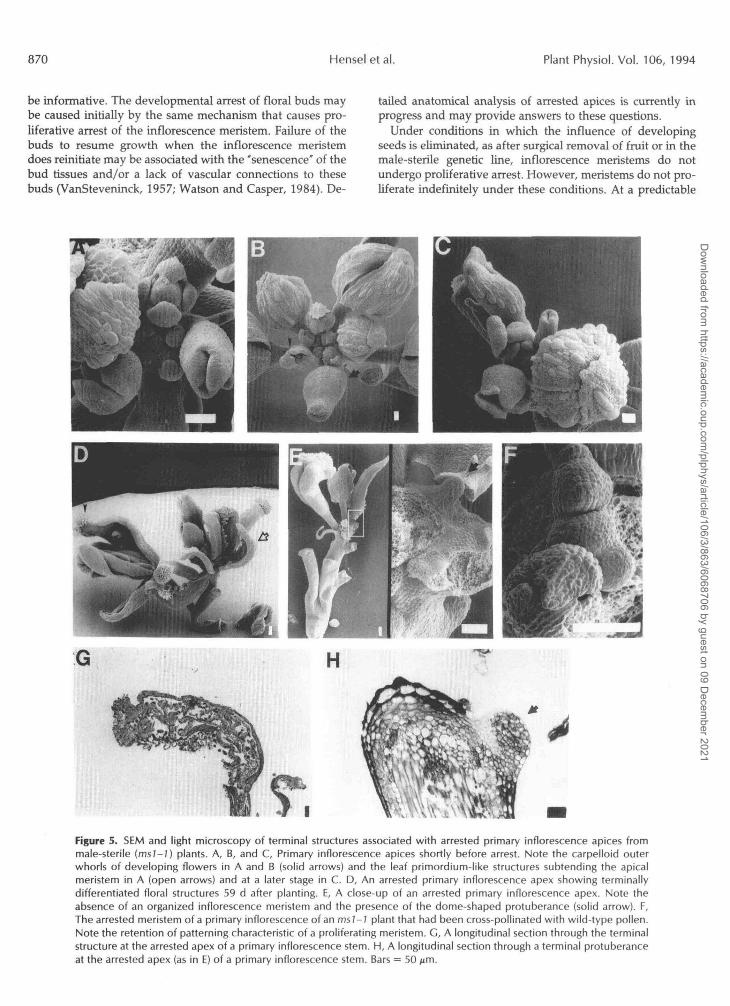
Morphological analysis of the msl-2 mutant line revealed that although proliferating meristems were initially indistin- guishable from wild type, the cessation of apical development was preceded by characteristic aberrations in paí teming at the apex. Altered pattems were manifested first ir^ the floral meristems with homeotic conversion of the oute- whorl in the direction of carpelloidy. In some cases the outer whorl formed a perfectly fused carpel-like structure (Fig. 5, A and B) that often encased well-formed inner-whorl organs (data not shown). Another common feature of older msl-1 meris- tems was the conversion for flower meristems into a pattem characteristic of vegetative meristems, where a single leaf- like primordium arose from the bases of primaq or lateral meristems (Figs. 31, 3J, 5D, and 5E). These primcrdia often developed into feminized leaf-like structures with ovules and stigmatic tissue often observed along the margirls. One to several such structures were usually observed at the termi- nally differentiated apex. The uppermost of these structures appeared to arise laterally from the apical region and had the appearance of unfused carpels (Fig. 5, D and E). Serial sections through nine male-sterile apices failed to reveal the presence of an organized inflorescence meristem (example shown in Fig. 5G). Small protrusions of apparently differen- tiated cells were occasionally observed in the apical region (Fig. 5, E and H). Although these protrusions may represent the residual of the inflorescence meristems, they also shared anatomical characteristics with nectaries, which normally develop at the receptacle of the flower (Davis, 1994).
The results of morphological analysis indicated that the mechanisms of apical arrest. in wild-type and male-sterile plants were different. Arrest of wild-type apices appeared to involve a cessation of proliferative activity but a rc tention of the basic structural pattem of the younger, active1 y prolifer- ating meristem. On the other hand, termination of infloresc- ence development in male-sterile plants was characterized by disruptions in pattem formation resulting in abeirations in floral development and in the apparent terminal differentia- tion of all tissues at the apex.
To distinguish the two types of apical arrest observed, we have designated the term “proliferative arrest” to describe the process characteristic of wild-type plants, and “terminal dif- ferentiation” to describe the process characteristic of male- sterile plants. The term GPA is used to indicate the coordinate proliferative arrest of all inflorescence meristems that occurs in wild-type Ler. GPA is not restricted to the Ler strain of Arabidopsis; we also observed this phenomena in the Colum- bia and WS strains (data not shown).
Dow
nloaded from https://academ
ic.oup.com/plphys/article/106/3/863/6068706 by guest on 09 D
ecember 2021

Meristem Fate in Arabidopsis 867
Figure 3. A comparative analysis of inflorescence developmental patterns in wild-type (Ler) and male-sterile (msJ-))plants. A, A wild-type plant 21 d after planting. B, A wild-type primary inflorescence branch with proliferating meristem.C, A wild-type plant 35 d after planting. All inflorescence branches have arrested. D, A wild-type primary inflorescencein the state of arrested development. E, A wild-type primary inflorescence that has reinitiated proliferative activity afterarrest. F, A male-sterile plant 21 d after planting. G, A male-sterile primary inflorescence branch with proliferatingmeristem. H, A male-sterile plant 49 d after planting. The primary inflorescence is no longer producing flowers. I and J,A male-sterile primary inflorescence that has terminally differentiated. K, Wild-type (left) and male-sterile (right) plantsgrown in an 8-h photoperiod. The photograph was taken at the time of msl-1 primary inflorescence meristem arrest.
Dow
nloaded from https://academ
ic.oup.com/plphys/article/106/3/863/6068706 by guest on 09 D
ecember 2021

868 Hensel et al. Plant Physiol. Vol. 106, 1994
~ ~ ~~
Table 1. Effects of photoperiod on selected developmental processes Leaf longevity was measured as t h e time of visual emergence (>1 mm) to the time of >50% degreening. Averages f SE (n) are shown. ND,
Not determined. Hours of Light in 24-h Cvcle Days to Bolt Leaf 5 Total Mature Ovaries on Plant
Longevitv 1 Inflorescence Drv Weieht (el
d g Wild-type Ler
Continuous 18.1 f 0.4 (44) 19.3 f 0.2 (161) 38.3 f 1.0 (44) 0.7 f 0.1 15) 16h 19.9 f 0.7 (47) 19.9 2 0.6 (1 37) 34.3 f 1.0 (19) ND 8 h 55.0 f 0.8 (20) 30.7 f 1.2 (20) 28.5 f 1.3 (13) 1.55 * 0.2 ( 1 1 )
Male-sterile Continuous 16.8 f 0.3 (25) 20.2 -C 0.5 (37) 63.2 f 2.9 (12) 1.2 f 0.2 :4) 16 h 18.9 f 0.8 (20) 21.8 f 0.9 (145) 68.6 k 2.4 (1 1 ) ND 8 h 54.0 f 0.9 (8) 32.4 f 2.7 (7) 71.8 f 2.8 (7) 1.51 f 0.1 :8)
The Role of Fruit Development in the GPA of Inflorescence Meristems
The fact that proliferative arrest did not occur in male- sterile plants led us to the hypothesis that f e w a t i o n and subsequent fruit development were responsible in some way for the GPA observed in wild-type plants. To test this hy- pothesis, flowers of the msl-1 line were hand pollinated with pollen from wild-type plants. Pollination of msl-1 plants led to a reduction in the total number of flowers produced (Fig. 6) and also resulted in the GPA of inflorescence meristems in a manner similar to that in wild-type plants (Fig. 5F). The cross-pollination of all flowers did not completely reverse the effects of the msl - l mutation on proliferative capacity. This result could be explained by the fact that the fruits on these plants showed a lower average seed set than wild-type fruits. Table I11 shows that when only flowers on the primary inflorescence stem were fertilized, the number of flowers produced by both primary and axillary meristems was re- duced, indicating that the influence of fertilization and fruit development on meristem proliferative capacity was global rather than locally restricted.
To further determine if fruit development plays a role in the timing of proliferative arrest, parts of wild-type plants were surgically removed from the developing inflorescence as shown in Figure 7. When inflorescence branches arising from the rosette were excised shortly after emergence, the proliferative capacity of the primary and aerial secondary inflorescence meristems was increased, but proliferative ar- rest still occurred (Fig. 7B). When all secondary branches were removed, the primary meristem produced over 60 flow- ers and terminated in structures similar to those observed in the male-sterile plants (Fig. 7C). To differentiate between the effects of flower fertilization and fruit development, an ex- periment was performed in which siliques were continuously removed from Ler plants 2 to 3 d after fertilization. In this case, both primary and secondary meristems produced more flowers, all of which terminated in male-sterile-like structures (Fig. 7D). This result indicates that fruit development rather than fertilization per se is responsible for proliferative arrest.
Under the standard growth conditions used in these -ex- periments, arrested meristems of Ler occasionally became reactivated 2 to 3 weeks after apical arrest had occurred (Fig. 3E). Thus, proliferative arrest appears to be a reversible state.
To explore this possibility further, surgical experiments were performed on Ler plants that had undergone GPA. When all fruits were removed from Ler plants 2 d after (PA, inflo- rescence meristems were reactivated and eventually produced new viable fruit (Fig. 7E).
Fruit Production in Hormone and Developmenta I Mutants
To examine the possible involvement of plant hormones in the interaction between inflorescence meristem and de- veloping fruit, the proliferative capacity of the primary inflo- rescence meristem was measured in mutant lines that are hormone deficient or insensitive (Table IV). In a1 cases, the primary inflorescence branch terminated in the bud clusters characteristic of wild-type proliferative arrest, indicating that none of the mutations disrupted the process of arrest. With the exception of the auxin-resistant axr2 and the GA-deficient gul-2, little or nó effect of the hormone mutants on the timing of proliferative arrest was observed. Although both axr2 and gal -2 lines showed a 30% reduction irk meristem productivity, these plants also displayed a number of mor- phological aberrations such as reduced size and increased branching that may have indirectly influenced thc prolifera- tive capacity of the inflorescence. Thus, the analysis of hor- mone mutants provided no compelling evidence ibr a direct role for auxins, GAS, ABA, or ethylene in the interactions of fruits and inflorescence meristems leading to GPA .
We also examined the effects of single-gene mutations that delay flowering time in the Ler and WS backgrounds. Late- flowering mutants in the Ler background showed slight to moderate increases in meristem proliferative capacity com- pared to the early-flowering wild type. On the other hand, late-flowering mutants in the WS background showed a doubling of proliferative capacity over the WS wild-type. Thus, the genetic background appears to influence the effects of delayed flowering on proliferative capacity.
To explore the quantitative relationship between seed de- velopment and the proliferative arrest of meristems, we meas- ured the proliferative capacity of the primary inllorescence meristem in mutants with reduced fertility. As indicated in Table V, mutations that reduced the number of jeeds pro- duced per fruit by greater than 50% were associated with a substantial increase in proliferative capacity of the primary inflorescence and the absence of proliferative arrest. On the
Dow
nloaded from https://academ
ic.oup.com/plphys/article/106/3/863/6068706 by guest on 09 D
ecember 2021

Meristem Fate in Arabidopsis 869
Figure 4. SEM of proliferating and arrested primary inflorescenceapices from wild-type (Ler) plants. A, A wild-type primary infloresc-ence apex 25 d after planting. The meristem is actively proliferatingand nine floral buds in developmental stages 1 through 5 (Smyth etal., 1990) are present. Bar = 50 urn. B, A wild-type primary inflo-rescence 1 week after proliferative arrest and 48 d after planting.Five buds in developmental stages 1 through 5 are present. Bar =50 jim.
other hand, lines that were heterozygous for embryo lethalmutations and therefore segregated 3:1 for viable seed pro-duced fewer fruits and showed proliferative arrest. We con-clude from this study that it is the number of developingseeds rather than the number of fruits per se that determinesthe occurrence and timing of proliferative arrest. The data
obtained indicate that a threshold of greater than 30% ofwild-type seed per pod is necessary to induce proliferativearrest.
DISCUSSION
Alternative Fates for Inflorescence Meristems
We have presented evidence in this report that infloresc-ence meristems of Arabidopsis are subject to one of twoalternative fates depending on the reproductive status of theplant. In wild-type plants, developing seeds appear to beresponsible for the propagation of some form of systemicsignal that triggers the arrest of all of the inflorescencemeristems on the plant within a 2-d period. The analysis offlower production rate presented in Figure 1 indicates thatcessation of flower production is rather abrupt. This appar-ently sharp transition from proliferating to arrested apexinvolves not only the inflorescence meristem, but also thedeveloping flower buds. The decreased number of budsfound at each developmental stage in the arrested relative tothe proliferating apex (Table II) is consistent with a model inwhich arrest occurs initially at the inflorescence meristemand subsequently affects floral buds only after the develop-ment of some buds has progressed through anthesis. Thearrest of the inflorescence meristem must involve primarily acessation of cell division activity. Given the capacity of thearrested meristem to reinitiate normal inflorescence devel-opment (Fig. 3E), the mechanisms involved in pattern for-mation are either maintained at the arrested apex or can bereinstated. On the other hand, the arrested floral buds thatresult from apical arrest are apparently not capable of resum-ing normal development (as shown in Fig. 3E).
On the basis of these observations, we postulate that atleast two different processes are associated with fruit-inducedarrest: (a) the proliferative arrest of the inflorescence meris-tem involving a reversible cessation of cell division, and (b)the arrested development of floral buds involving an irre-versible cessation of cell expansion and differentiation, aswell as cell division. The mechanisms by which these twoprocesses are controlled are still unknown. For proliferativearrest, it will be important to determine whether cells in themeristem are blocked at a specific stage in the cell cycle.Expression patterns of the cell-cycle-associated genes thathave recently been cloned from Arabidopsis (Ferreira et al.,1991; Hirayama et al., 1991; Hemerly et al., 1992) may also
Table II. Comparative analysis of floral development at active andarrested inflorescence meristems of Ler
Stages of flower development are defined by Smyth et al. (1990).Stage 6 represents the point at which sepals completely enclosethe bud.
Number of BudsMeristem
In stages 1-5 In stages 6-12
Active meristemArrested meristem
Average ± SE (n)
8.9 ±0.1 (16) 19.6 ±0.2 (16)4.0 + 0.3(23) 11.2 ±0.7 (23)
Dow
nloaded from https://academ
ic.oup.com/plphys/article/106/3/863/6068706 by guest on 09 D
ecember 2021
870 Hensel e\. al. Plant Physiol. Vol. 106, 1994
be informative. The developmental arrest of floral buds maybe caused initially by the same mechanism that causes pro-liferative arrest of the inflorescence meristem. Failure of thebuds to resume growth when the inflorescence meristemdoes reinitiate may be associated with the 'senescence" of thebud tissues and/or a lack of vascular connections to thesebuds (VanSteveninck, 1957; Watson and Casper, 1984). De-
tailed anatomical analysis of arrested apices is currently inprogress and may provide answers to these questions.
Under conditions in which the influence of developingseeds is eliminated, as after surgical removal of fruit or in themale-sterile genetic line, inflorescence meristems do notundergo proliferative arrest. However, meristems do not pro-liferate indefinitely under these conditions. At a predictable
H IS*
Figure 5. SEM and light microscopy of terminal structures associated with arrested primary inflorescence apices frommale-sterile (ms/-7) plants. A, B, and C, Primary inflorescence apices shortly before arrest. Note the carpelloid outerwhorls of developing flowers in A and B (solid arrows) and the leaf primordium-like structures subtending the apicalmeristem in A (open arrows) and at a later stage in C. D, An arrested primary inflorescence apex showing terminallydifferentiated floral structures 59 d after planting. E, A close-up of an arrested primary inflorescence apex. Note theabsence of an organized inflorescence meristem and the presence of the dome-shaped protuberance (solid arrow). F,The arrested meristem of a primary inflorescence of an msl-1 plant that had been cross-pollinated with wild-type pollen.Note the retention of patterning characteristic of a proliferating meristem. G, A longitudinal section through the terminalstructure at the arrested apex of a primary inflorescence stem. H, A longitudinal section through a terminal protuberanceat the arrested apex (as in E) of a primary inflorescence stem. Bars = 50 p.m.
Dow
nloaded from https://academ
ic.oup.com/plphys/article/106/3/863/6068706 by guest on 09 D
ecember 2021

Meristem Fate in Arabidopsis 871
u) Q .-
60- B g! a E 40- c
c
L O
al D
z 5 20-
0-
'1 B .- A Figure 6. The affects of pollination o n the proliferative capacity of the primary inflorescence meristem of male-sterile (msl-1) plants. The average numbers f SE of mature ovaries produced by wild- type Ler (n = 84), untreated msl-1 (n = 64), partially pollinated msl-1 (n = 21), and completely pollinated msl-1 (n = 9) are depicted. For partial pollination, only flowers on the primary inflo- rescence were pollinated.
stage of development (i.e. after the production of 60-70 flowers) individual flower meristems will begin to produce aberrant floral structures. These homeotic conversions are often followed by the apparent reversion of the apical mer- istem toward a vegetative mode of patterning. The subse- quent cessation of proliferative activity at the apex is associ- ated with the loss of organization at the apical meristem and the differentiation of cells within the apex. Our examination of a number of reduced-fertility lines (Table V) indicated that apices terminated in structures similar to those observed for msl-1. Similar observations have been reported for male- sterile mutants (Chaudhury, 1993), female-sterile mutants (Modrusan et al., 1994), and flower-development mutants (Schultz and Haughn, 1993). We interpret these observations by considering that, by whatever mechanism, the termination of development characteristic of the male-sterile line is the default fate for the meristems. We consider the proliferative arrest obtained through the influence of developing fruit to be a regulatory process that is superimposed on this default fate for inflorescence meristems.
The mechanism involved in the determinant development of the apex of male-sterile and defruited wild-type plants is not known. The morphology of terminal structures is similar to that of structures caused by mutations in floral organ identity genes. In particular, the observed loss of second- whorl organs and conversion of first-whorl organs to carpe- loid structures are similar to homeotic conversions resulting
from mutations in APZ, AP2, or LFY (Kunst et al., 1989; Irish and Sussex, 1990; Bowman et al., 1991; Schultz and Haughn, 1991). In all of these cases, it may be ectopic expression of the AG gene that drives the feminization of the developing organs. As in terminating msl-1 apices, mutations in LFY also tend to shift the pattem of development toward the vegeta- tive program (Schultz and Haughn, 1991; Weigel et al., 1992). Thus, it may be of particular interest to examine the pattems of expression of LFY and AG in terminating msl-Z infloresc- ence apices. Although the similarities of the phenotypes we have observed to floral mutants are intriguing, it is unlikely that terminal differentiation in male-sterile plants results from genetic lesions in these genes. The predictable timing and phenotypic progression of the terminal differentiation process is indicative of some epigenetic phenomenon that produces consequences similar to those of the homeotic mu- tations. It is certainly possible that the pattem disruptions observed are associated with altered expression levels of floral identity genes.
Comparative Analysis of Apical Arrest in Arabidopsis and Pea
The correlative interactions we have observed in Arabidop- sis between seed development and meristem proliferative capacity share many features in common with correlative interaction in another well-studied monocarpic plant, Pisum sativum. According to the original work of Lockhart and Gottschall (1961), fruit removal delayed the arrest of the vegetative apex in pea. They termed the fruit-induced arrest "senescence," but subsequent work demonstrated that the arrested apex retained a relatively healthy-appearing apical meristem and that the arrested apex could be reactivated by subsequent fruit removal (Wang and Woolhouse, 1982).
Lockhart and Gottschall also reported that in the absence of fruit, the apical meristem underwent a subsequent arrest that was associated with the development of a terminal flower (Le. terminal differentiation). Grafting experiments led to the conclusion that the second form of apical arrest was a function of the age of the meristem. Since no detailed mor- phological analysis was performed on the terminally differ- entiated apices, the nature of apical arrest is not known. More recently it has been shown that in the determinate det mutant of pea, the inflorescence meristem does not undergo conver- sion to a flower, but rather differentiates into a pubescent rudimentary stub (Singer et al., 1990).
Thus, although the specific mechanism may differ, it would appear that Arabidopsis and some varieties of pea show comparable aspects of correlative interactions between fruits
Table 111. Branch and flower oroduction in male-sterile olants with fruit set
Total Number Dry Weight Number of Branches per 2' Branch
3' 4' 5" 6" 7" Flowers per 2' g
Ler 2.8 f 0.4 2.6 f 0.4 0.6 f 0.4 0.0 f 0.0 0.0f 0.0 68.6 f 6.9 0.7 f 0.1 msl-1 3.8 f 0 . 3 5.8 f 0.5 4.3 f 0.3 1.3 f 0.5 0.5 f 0.3 377.3 f 19.4 1.8 f 0.5 Fertilized msl-la 3.4 f 0.3 5.2 f 0.3 3.2 f 0.7 1.2 f 0.9 0.4 f 0.5 287.8 f 21.6 1.2 f 0.2
~~~ ~~
a All the flowers on the primary inflorescence were hand-pollinated once every 24 to 48 h.
Dow
nloaded from https://academ
ic.oup.com/plphys/article/106/3/863/6068706 by guest on 09 D
ecember 2021

8 72 Hensel et al. Plant Physiol. Vol. 106, 1994
and meristems. In both species, reversible proliferative arrest of apical meristems is controlled by developing fruit. Inter- estingly, the affected apical meristem is vegetative in the case of pea but are inflorescence meristems in the case of Arabi- dopsis. In pea, only a single vegetative meristem is active prior to proliferative arrest. On the other hand, the highly branched inflorescence of Arabidopsis provides the first evi- dence for a global systemic signal that causes the coordinate arrest of all shoot meristems simultaneously. Finally, the terminal development of inflorescence meristems of male- sterile Arabidopsis is similar to the terminal development of the inflorescence meristems in the det mutant in pea (Singer et al., 1990). In both cases the inflorescence meristem appears to degenerate into a relatively differentiated mound of tissue that shows no subsequent signs of proliferative activity.
A number of studies in pea have attempted to address the regulatory mechanisms responsible for correlative interac- tions between fruits and meristems. The most detailed analy- sis comes from work with the G2 variety of peas. Under the influence of the dominant genes Sn and Hr, the G2 variety undergoes apical arrest in long days, but will continue to proliferate almost indefinitely under SD conditions (Reid, 1980). Since the G2 variety initiates flowering in both long and short days at the same time, G2 provides an opportunity to examine a system in which the proliferative activity of the apical meristem is not suppressed by developing fruit. De- tailed developmental studies indicated that the lack of apical arrest in SD-grown G2 is due to an avoidance of resource allocation conflict between fruits and meristem. Under short days, fruits develop more slowly and tend to abort more frequently (Gianfagna and Davies, 1981). Under these con- ditions there appears to be less direct competition for re- sources between fruits and the vegetative meristem, perhaps due to an increase in physical distance between the apex and developing fruit. These developmental effects of the short day can be mimicked in LD plants by GA application (Proebsting et al., 1976; Murfet and Reid, 1987), indicating that GAS play an important role in governing the balance between vegetative and reproductive development in the pea plants.
Correlative Control Models for Fruit/Meristem Interactions
Although the studies of the G2 variety of peas described above provide an example of how heterochronic alterations in vegetative and reproductive development may result in the avoidance of resource allocation conflicts, they do not address the question of what mechanisms operate to resolve these conflicts once they do occur. In both Arabidopsis and LD-grown G2 peas, resolution of these conflicts involves the proliferative arrest of the apical meristem. We consider two different basic models that may account for the mechanisms of apical arrest: (a) the resource limitation model, and (b) the inhibitory factor model. Schematic representations of these two models are provided in Figure 8.
In the resource limitation model, we envision that under conditions of competition between developing seeds and meristems, some resource drops below a threshold level at the meristems, triggering proliferative arrest. The limiting
resource could be a nutrient or a specific growl h factor or hormone. The alternative inhibitory factor modd stipulates that a chemical signal produced by fruits or seeds is respon- sible for suppression of meristem activity when the level of the inhibitor exceeds a specific threshold at the apex. These models are not new, but are rather more specific restatements of Molisch’s original nutrient drain hypothesis (1928) and the senescence factor hypothesis originally suggested and developed by Leopold (1961). The relative merts of these two models continue to be discussed in the literature (Kelly and Davies, 1988a; Noodén, 1988b). However, definitive experiments that would differentiate between these models have not yet been designed.
Using the G2 variety of pea, Kelly and Davies (1988b) showed that plants grown in long days partitioned less assim- ilate to the apical bud than did plants in short days, favoring the resource limitation model. However, these experiments did not prove a causal role for decreased nutrient supply, since it was not determined that the lower level of assimilate supplied to the apex in long days was below the limit nec- essary to sustain apical development. On the other hand, the identification of a chemical senescence factor pioduced by fruits has not been achieved in any plant system. Labeling of G2 pea fruits with 14C02 failed to reveal a specific labeled compound with the appropriate properties of a senescence factor, leading Hamilton and Davies (1988) to conclude that no such factor existed. However, if the presumpti\re chemical factor operated at a concentration relevant to hormones, it is unlikely that the labeling studies used would have identified such a factor.
Although the nature of the signal that triggers proliferative arrest is not known, certain properties may be inferred from our results. The rapid transition from proliferating to arrested meristem that we have observed (Fig. 1) prompts us to suggest that the transition represents a threshold response to the presumptive signal. We also postulate that the signal is a systemic one based on the observation that prolifer ative arrest affects ail inflorescence meristems at about the same time (Fig. 2). In addition, manipulation of individual branches of the plant, such as fruit removal in wild type (Fig. ?) or cross- pollination of male-sterile flowers (Table 111), affects prolif- erative activity of untreated branches. Furthermore, if the interaction between fruits and meristems were localized within individual branches, then surgical removal of second- ary branches should not prevent the arrest of the primary apex. The fact that the primary apex did not arrest under these conditions (Fig. 7C) implies the involvemlznt of the basal stem and/or root in the global interaction. ‘lihe results of this particular surgical experiment could be explained if the basal stem and/or roots acted as a source of the positive growth factor. Removal of secondary branches would elimi- nate a major sink for this pool of positive growth factor, and, as a consequence, the level of the growth factor in the primary stem would never drop below the threshold needec to sustain the proliferative activity at the primary apex. On the other hand, the results obtained could also be interpreted according to the negative regulator model by assuming that the basal stem and/or roots act as a sink for the negative regulator. In this case, the limited production of the negative regulator by the fruits on the primary stem alone would not be sufficient
‘
Dow
nloaded from https://academ
ic.oup.com/plphys/article/106/3/863/6068706 by guest on 09 D
ecember 2021

Meristem Fate in Arabidopsis a73
A
B C
&. D E
Key
\ silique - silique removed 4 infertile ovary o arrested bud 8 arrested bud cluster
branch removed
Figure 7. Schematic diagram depicting the effects of surgical treatments on the proliferative activity of the inflorescence meristems of Ler. A, Untreated Ler plants ( n = 50). B, All basal secondary branches were excised (n = 15). Branches were removed within 48 h of visible emergence. C, All secondary branches were excised ( n = 14). Branches were removed within 48 h of visible emergence. D, Siliques were excised sequentially within 2 to 3 d of fertilization (n = 6). E, All siliques were excised from the plant 3 d after proliferative arrest of the primary inflorescence meristem (n = 4). One-half of the plants ultimately produced terminally differentiated structures at the primary branch. The vertical length of the branches above the last branch point is drawn to scale according to the average number of structures produced. All SE values were ~ 1 0 % of the average number of structures, except the SE for fourth-order wild-type branches, which was 14.3% of the average. Only primary and secondary branches are shown for D and E; higher-order branches behaved in the same manner. Plants were grown in continuous light for A through C , and 16-h light/8-h dark for D and E.
to elevate the level of the regulator above the threshold needed to cause the proliferative arrest of the primary mer- istem. Either mechanism would provide the plant with a means of measuring the extent of fruit production and re- sponding in terms of limiting new seed production before the entire system becomes depleted of general resources. That some such preemptive system does operate in Arabidopsis is evidenced by the fact that the final flowers, which do reach anthesis, invariably go on to produce a full complement of viable seed (Fig. 3D).
Our examination of meristem fate in lines carrying muta- tions that affect reproductive development leads us to con- clude that proliferative arrest is dependent on the number of seeds produced per fruit and the total number of fruit. The
sink demand produced by developing seed is likely to play a role in determining the timing of proliferative arrest. How- ever, the relationship between existing resources and sink demand by seed is apparently not a simple one. In the Ler background, delayed flowering, whether induced by short days or by mutations, caused a substantial increase in total vegetative biomass for the plant but did not appreciably alter the timing of proliferative arrest. On the other hand, delayed flowering in the WS background resulted in a doubling of proliferative capacity of the primary inflorescence meristem. Genetic analysis of crosses between the Ler and WS lines is currently being performed to determine whether specific modifiers are responsible for the observed differences.
In considering the nature of specific chemical signals that
Dow
nloaded from https://academ
ic.oup.com/plphys/article/106/3/863/6068706 by guest on 09 D
ecember 2021

874 Hensel et al. Plant Physiol. Vol. 106, 1994
Table IV. Fruit production on the primary inflorescence branch in hormone and developmental mutant lines
Line Phenotype Fruits Produced (Average & SE)
Columbia genetic background Col axrl- 12 axr2 ein2- 1 etol-l etrl-1
Ler abil abi2 ga 1-2 ga 1-5 ga4 gai fca-1 ,
fd- 1 fe 1 fha- 1
he- 1
Ler genetic background
ft- 1
fY- 1 Id-3
ws WS genetic background
gi Id-2
wild type 15 Auxin resistant 5 Auxin resistant 7
Ethylene overproducer 7 Ethylene insensitive a
Ethylene insensitive 7
Wild type ABA insensitive ABA insensitive GA deficient GA deficient GA deficient GA insensitive Late flowering Late flowering Late flowering Late flowering Late flowering Late flowering Late flowering Late flowering
58
a
a
a
12 9
7 10
14 9
7 9 9 9 5
Wild type 30 Late flowering 7 Late flowering 7
38 f 2 38 4 25 f 1 37 f 3 37 f 2 3 6 f 1
3 0 f 1 40 f 3 45 f 2 20 f 2 42 f 2 36 f 2 27 f 2 39 f 2 36 f 2 29 f 1 31 f 3 3 5 2 2 33 f 1 3 4 f 1 3 2 f 2
47 f 2 97 f 10 81 f 7
may be responsible for the interaction between developing seeds and inflorescence meristems, the known plant hor- mones are obvious candidates. Our initial studies with hor- mone mutants did not provide compelling evidence for a role for auxin, GAS, ABA, or ethylene. Although these experi- ments do not allow us to rule out a central role for one of these four classes of hormones, our results do indicate that the correlative interaction between developing seeds and proliferating meristems is resilient enough to operate under conditions in which major alterations in hormone levels or sensitivities are occumng. We have initiated additional ex-
-
periments in which cytokinins and GAS have been applied to plants shortly after proliferative arrest. In preliminary experiments, we saw no reinitiation of arrested apices after hormone application (data not shown). Although íhe known hormones have not been ruled out, altemative possibilities should be considered. Recent evidence that peptid1.s may act as systemic signals in plants (Pearce et al., 1991) coupled with the identification in plants of genes that code for growth factor receptor-like proteins (Walker and Zhang, 1990; Stein et al., 1991; Chang et al., 1992) provide at lead a formal possibility that this type of signaling system so prevalent in
Table V. Fruit and seed production on the primary inf/orescence in embryo defective and reduced fertility T-tagged lines - Terminal Structures on the Inflorescen,:e N Seed/Pod’ Fruits Produced
(Average 2 SE) (Average & SE) Line Phenotype
ws Wild type 61 f 2 47 f 2 30 bud cluster CS2330 Reduced fertility 6 f l 69 f 5 4 homeotic-like
Reduced fertility i a f 3 63 f 12 4 homeotic-like CS2333 Reduced fertility 5 + - 1 62 f 6 4 homeotic-li ke CS2334
CS2346 Reduced fertility ND 68 f 7 5 homeotic-like
CS3005 Embryo lethal, globular 2 3 f 2 69 f 2 6 homeotic-like CS3011 Embryo lethal, globular 39 f 2 3 a f i 3 bud cluster CS3039 Embryo lethal, heart 48 f 3 35 f 1 7 bud cluster CS3049 Embryo lethal, heart 48 f 2 3 0 f 1 a bud cluster
csma Reduced fertility 1 2 f l 57 f 4 4 N D ~
a Seeds were counted for siliques from positions 10 through 20 (numbered by order of emergence) o n the primary inflorescence. ND, Not determined.
Dow
nloaded from https://academ
ic.oup.com/plphys/article/106/3/863/6068706 by guest on 09 D
ecember 2021

Meristem Fate in Arabidopsis 875
proliferating inflorescence meristem
I c/ b arrested inflorescence I
time - Model 1
meristem developing fruit
Model 2
Figure 8. Alternative models to explain the influence of developing fruits on the proliferative activity of the inflorescence meristems. The growth-factor diversion model (model 1 ) is based on the assumption that a factor that is needed to sustain proliferative activity at the meristems is sequestered by developing seeds, resulting in arrest. The negative- regulator model (model 2) is based on the idea that a negative regulator produced by developing seeds increases above a threshold level and causes the proliferative arrest of the meristems.
the control of cell-division activities in animal systems is also operating in plants.
CONCLUDING REMARKS
As a consequence of whole plant senescence associated with the monocarpic habit, reproductive development may occur under conditions in which maternal resources become increasingly limited. Maternal investment in reproduction must then represent a trade off between fecundity (the num- ber of offspring produced) and individual investment (the amount of resource apportioned to each offspring). Under conditions of resource limitation, these two requirements will become mutually antagonistic unless mechanisms that limit the number of offspring produced are in place. Evolutionary theorists have developed a number of population genetics models that take these so-called resource allocation conflicts into account (for a review, see Lloyd, 1992). These models predict that alleles of genes that act to resolve resource allocation conflicts in evolutionarily favorable ways will be selected for. The interactions between developing fruits and inflorescence meristems that we have described in this paper may provide a specific test for these theoretical considera- tions. If specific genetic mechanisms are in place in Arabidop- sis that mediate the interactions between developing seeds and inflorescence meristems, then mutational analysis may identify specific genes that mediate these interactions. Fur- thermore, the cloning and characterization of these mutated genes could provide the basis for investigating the biochem- ical mechanisms that mediate these "correlative" interactions.
ACKNOWLEDGMENTS
We thank Claudia Lipke for photographic preparations. We thank Andrea Strzelec for help with plant analyses, and appreaate the assistance with SEM provided by Sara Patterson. We thank Sara Patterson, Rick Amasino, Donna Femandez, and Judy Croxdale for critical review of the manuscript.
Received March 24, 1994; accepted July 8, 1994. Copyright Clearance Center: 0032-0889/94/106/0863/14.
LITERATURE CITED
Alvarez J, Guli CL, Yu X, Smyth DR (1992) terminal power, a gene affecting inflorescence development in Arabidopsis thuliana. Plant
Arubidopsis Biological Resource Center at Ohio State (1994) Seed and DNA Stock List. Columbus, OH
Bleecker AB, Estelle MA, Somerville C, Kende H (1988) Insensi- tivity to ethylene conferred by a dominant mutation in Arabidopsis thuliana. Science 2 4 1017-1132
Bowman JL, Smyth DR, Meyerowitz EM (1991) Genetic interac- tions among floral homeotic genes of Arabidopsis. Development 112 1-20
Chang C, Schaller GE, Patterson SE, Kwok SF, Meyerowitz EM, Bleecker AB (1992) The TMKl gene from Arubidopsis codes for a protein with structural and biochemical characteristics of a receptor protein kinase. Plant Cell 4 1263-1271
Chaudhury AM (1993) Nuclear genes controlling male fertility. Plant Cell 5: 1277-1283
Davis AR (1994) Wild-type nectary morphology. In J Bowman, ed, Arubidopsis: An Atlas of Morphology and Development. Springer- Verlag, New York, pp 172-173
Engvild KC (1989) The death hormone hypothesis. Physiol Plant
J 2 103-116
77: 282-285
Dow
nloaded from https://academ
ic.oup.com/plphys/article/106/3/863/6068706 by guest on 09 D
ecember 2021

8 76 Hensel et al. Plant Physiol. Vol. 106, 1994
Ferreira P, Hemerly AS, Villarroel R, Van Montagu M, Inzé D (1991) The Arabidopsis functional homolog of the ~ 3 4 ' ~ ' ~ protein kinase. Plant Cell3 531-540
Gianfagna TJ, Davies PJ (1981) The relationship between fruit growth and apical senescence in the G2 line of peas. Planta 152
Gut H, Matile P (1988) Apparent induction of key enzymes of the glyoxylic acid cycle in senescent barley leaves. Planta 176
Hamilton DA, Davies PJ (1988) Sucrose and malic acid as the compounds exported to the apical bud of pea following "CO2 labeling of the fruit. Plant Physiol88: 466-472
Hemerly A, Bergounioux C, Van Montagu M, Inze D, Ferreira P (1992) Genes regulating the plant cell cycle: isolation of a mitotic- like cyclin from Arabidopsis thaliana. Proc Natl Acad Sci USA 8 9
Hempel FD, Feldman LJ (1994) Bi-directional inflorescence devel- opment in Arabidopsis thaliana: acropetal initiation of flowers and basipetal initiation of paraclades. Planta 192 276-286
Hensel LL, Bleecker AB (1992) Arabidopsis as a model system for analysis of leaf senescence and inflorescence-meristem longevity. In RM Amasino, ed, Proceedings of the Twenty-First Steenbock Symposium: Cellular Communication in Plants. Plenum Press, New York, pp.123-130
Hensel LL, Grbic V, Baumgarten DB, Bleecker AB (1993) Devel- opmental and age-related processes that influence the longevity and senescence of photosynthetic tissues in Arabidopsis. Plant Cell
Hildebrand F (1881) Die Lebensdauer und Vegetationsweise der Pflanzen, ihre Ursahe und ihre Entwicklung. Bot Jahrb Syst Pflan- zengesch Pflanzengeogr 2: 51-135
Hirayama T, Imajuku Y, Anai T, Matsui M, Oka A (1991) Identi- fication of two cell-cycle-controlling cdc2 gene homologs in Aru- bidopsis thulinna. Gene 105 159-165
Irish VF, Sussex IM (1990) Function of the apetala-l gene during Arubidopsis floral development. Plant Cell2 741-753
Kelly MO, Davies PJ (1988a) The control of whole plant senescence. CRC Crit Rev Plant S a 7: 139-172
Kelly MO, Davies PJ (1988b) Photoperiodic and genetic control of carbon partitioning in peas and its relationship to apical senes- cence. Plant Physiol86 978-982
Kirchheim B, Kranz AR (1981) New population samples of the AIS- seed bank. Arabidopsis Inf Serv 18: 173-176
Koornneef M, van Eden J, Hanhart CJ, Stam P, Braaksma FK, Feenstra WJ (1983) Linkage map of Arubidopsis thaliana. J Hered
Kranz A R (1978) Demonstration of new and additional populations samples and mutant lines of the AIS-seed bank. Arabidopsis lnf Serv 15: 118-139
Kunst L, Klenz JE, Martinez-Zapater J, Haughn GW (1989) AP2 gene determines the identity of perianth organs in flowers of Arabidopsis thaliana. Plant Cell 1: 1195-1208
Lee I, Aukerman MJ, Gore SL, Lohman KN, Michaels SD, Weaver LM, John MC, Feldmann KA, Amasino RM (1994) Isolation of LUMZNIDEPENDENS a gene involved in the control of flowering time in Arabidopsis. Plant Cell 6 75-83
Leopold AC (1961) Senescence in plant development. Science 134
Lloyd DG (1992). Evolutionarily stable strategies of reproduction in plants: who benefits and how? In R Wyatt, ed, Ecology and Evolution of Plant Reproduction. Chapman and Hall, New York,
Lockhart JA, Gottschall V (1961) Fruit-induced and apical senes-
Modrusan Z, Reiser L, Feldmann KA, Fischer RL, Haughn GW
356-364
548-550
3295-3299
5: 553-564
7 4 265-272
1727-1732
pp 137-168
cence in Pisum sativum L. Plant Physiol36 389-398
(1994) Homeotic transfonnation of ovules into carpd-like shuc- tures in Arabidopsis. Plant Cell6 333-349
Molisch H (1938) The Longevity of Plants. Authonzed English Edition by E Fulling. The Saence P m s Printing Compariy, hcaster, PA
Murfet IC, Reid JB (1987) Flowering in Pisum: gibkjerellins and flowering genes. J Plant Physiol127: 23-29
Murneek. AR (1926) Effects of correlation between vegetative and reproductive functions in the tomato (Lycopersicon, esculentum MiU.). Plant Physiol 1: 3-56
Noodén ILD (1988a) The phenomena of senescence a:id aging. In LD Noodén, AC Leopold, eds, Senescence and Agirig in Plants. Acadenuc Press, San Diego, pp 1-50
Noodén LD (1988b) Whole plant senescence. In LD Noodén, AC Leopold, eds, Senescence and Aging in Plants. Acaliemic Press, San Diego, pp 391-439
Pearce G, Strydom D, Johnson S, Ryan CA (1991) A polypeptide from tomato leaves induces wound-inducible proteinase inhibitor proteins. Saence 253 895-898
Proebsting WM, Davies PJ, Marx GA (1976) PhotopeIiodic control of apical senescence in a genetic line of peas. Planí. Physiol 58:
Reid JB (1980) Apical senescence in Pisum: a direct or indirect role for the flowering genes. Ann Bot 4 5 195-201
Schultz EA, Haughn GW (1991) LEAFY, a homeoec gene that regulates inflorescence development in Arabidopsis. .Plant Cell 3
Schultz EA, Haughn GW (1993) Genetic analysis of the floral initiation process (FLIP) in Arabidopsis. Development 1.19: 745-765
Shannon S, Meeks-Wagner DR (1991) A mutation in the Arabidopsis TFLl gene affects inflorescence meristem developmerit. Plant Cell
Sinclair TR, de Wit CT (1975) Photosynthate and nitrclgen require- ments for seed production by various crops. Sdence 189 565-567
Singer SR, Hsiung LP, Huber SC (1990) Determinate (de t ) mutant of Pisum sativum (Leguminosae: papilionoideae) exhbits an inde- terminate growth pattem. Am J Bot 77: 1330-1335
Smyth DR, Bowman JL, Meyerowitz EM (1990) Earlq flower de- velopment in Arabidopsis. Plant Cell2 755-767
Stein JC, Howlett B, Boyes DC, Nasrallah ME, Nasrallah JB (1991) Molecular cloning of a putative receptor kinase gent' encoded at the self-incompatibility locus of Brassica oleracen. Prcc Natl Acad
VanSteveninck RFM (1957) Factors affecting the abscission of re- productive organs in yellow lupins (Lupinus luteus L.). I. The effect of different pattems of flower removal. J Exp Bot 8: 373-381
Vaughan JG (1955) The morphology and growth of the vegetative and reproductive apices of Arabidopsis thaliana (L.) Heynh., Cap- sella bursn-pastoris (L.) Medic., and Anagallis ntuensis I-. J Linn SOC Lond Bot 5 5 279-301
Walker JC, Zhang R (1990) Relationship of a putative receptor protein kinase from maize to the S-locus glycoproteinii of Brassica. Nature 345 743-746
Wang T, Woolhouse HW (1982) Growth regulators in plant senes- cence. British Plant Growth Regulator Group Monograph 8: 5-25
Watson MA, Casper BB (1984) Morphogenetic constraints on pat- tems of carbon distribution in plants. Annu Rev Eco1 Syst 1 5
Weigel D, Alvarez J, Smyth DR, Yanofsky MF, Meyl?rowitz EM (1992) LEAFY controls floral meristem identity in Aratidopsis. Cell
Woolhouse HW (1983) Hormonal control of senescerice allied to reproduction in plants. Zn Beltsville Symposia in Agricultura1 Re- search-Strategies of Plant Reproduction. Allanheld, Osmun, and Co, Totowa, NJ, pp 201-236
Woolhouse HW (1984) The biochemistry and regulation of senes- cence in chloroplasts. Can J Bot 6 2 2934-2942
800-802
771-781
3 877-892
Sci USA 88: 8816-8820
233-258
6 9 843-859
Dow
nloaded from https://academ
ic.oup.com/plphys/article/106/3/863/6068706 by guest on 09 D
ecember 2021


















