
The evolution of drought escape and avoidance in natural...
8
Plant Science 234 (2015) 155–162 Contents lists available at ScienceDirect Plant Science j ourna l ho me pa ge: www.elsevier.com/locate/plantsci Review The evolution of drought escape and avoidance in natural herbaceous populations Nicholas J. Kooyers ∗ Department of Biology, University of Virginia, Charlottesville, VA 22904, USA a r t i c l e i n f o Article history: Received 31 October 2014 Received in revised form 4 February 2015 Accepted 19 February 2015 Available online 25 February 2015 Keywords: Adaptation Plasticity Drought resistance Water-use efficiency Gene by environment interactions Flowering time a b s t r a c t While the functional genetics and physiological mechanisms controlling drought resistance in crop plants have been intensely studied, less research has examined the genetic basis of adaptation to drought stress in natural populations. Drought resistance adaptations in nature reflect natural rather than human- mediated selection and may identify novel mechanisms for stress tolerance. Adaptations conferring drought resistance have historically been divided into alternative strategies including drought escape (rapid development to complete a life cycle before drought) and drought avoidance (reducing water loss to prevent dehydration). Recent studies in genetic model systems such as Arabidopsis, Mimulus, and Pan- icum have begun to elucidate the genes, expression profiles, and physiological changes responsible for ecologically important variation in drought resistance. Similar to most crop plants, variation in drought escape and avoidance is complex, underlain by many QTL of small effect, and pervasive gene by environ- ment interactions. Recently identified major-effect alleles point to a significant role for genetic constraints in limiting the concurrent evolution of both drought escape and avoidance strategies, although these con- straints are not universally found. This progress suggests that understanding the mechanistic basic and fitness consequences of gene by environment interactions will be critical for crop improvement and forecasting population persistence in unpredictable environments. © 2015 Published by Elsevier Ireland Ltd. Contents 1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 156 2. The evolution of drought escape in natural populations . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 157 2.1. Prevalence . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 157 2.2. Fitness consequences . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 157 2.3. Genetic basis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 157 3. The evolution of drought avoidance in natural populations . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 158 3.1. Prevalence . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 158 3.2. Fitness consequences . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 159 3.3. Genetic basis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 159 3.4. Resource reserves as drought avoidance adaptations . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 159 3.5. Genetic tradeoffs in the evolution of drought resistance . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 160 4. A path toward understanding drought resistance adaptations in natural populations . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 160 Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 161 References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 161 ∗ Tel.: +1 434 924 0162. E-mail address: [email protected] http://dx.doi.org/10.1016/j.plantsci.2015.02.012 0168-9452/© 2015 Published by Elsevier Ireland Ltd.
Transcript of The evolution of drought escape and avoidance in natural...

R
Tp
ND
a
ARRAA
KAPDWGF
C
h0
Plant Science 234 (2015) 155–162
Contents lists available at ScienceDirect
Plant Science
j ourna l ho me pa ge: www.elsev ier .com/ locate /p lantsc i
eview
he evolution of drought escape and avoidance in natural herbaceousopulations
icholas J. Kooyers ∗
epartment of Biology, University of Virginia, Charlottesville, VA 22904, USA
r t i c l e i n f o
rticle history:eceived 31 October 2014eceived in revised form 4 February 2015ccepted 19 February 2015vailable online 25 February 2015
eywords:daptationlasticityrought resistanceater-use efficiency
a b s t r a c t
While the functional genetics and physiological mechanisms controlling drought resistance in crop plantshave been intensely studied, less research has examined the genetic basis of adaptation to drought stressin natural populations. Drought resistance adaptations in nature reflect natural rather than human-mediated selection and may identify novel mechanisms for stress tolerance. Adaptations conferringdrought resistance have historically been divided into alternative strategies including drought escape(rapid development to complete a life cycle before drought) and drought avoidance (reducing water lossto prevent dehydration). Recent studies in genetic model systems such as Arabidopsis, Mimulus, and Pan-icum have begun to elucidate the genes, expression profiles, and physiological changes responsible forecologically important variation in drought resistance. Similar to most crop plants, variation in droughtescape and avoidance is complex, underlain by many QTL of small effect, and pervasive gene by environ-
ene by environment interactionslowering time
ment interactions. Recently identified major-effect alleles point to a significant role for genetic constraintsin limiting the concurrent evolution of both drought escape and avoidance strategies, although these con-straints are not universally found. This progress suggests that understanding the mechanistic basic andfitness consequences of gene by environment interactions will be critical for crop improvement andforecasting population persistence in unpredictable environments.
© 2015 Published by Elsevier Ireland Ltd.
ontents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1562. The evolution of drought escape in natural populations . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 157
2.1. Prevalence . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1572.2. Fitness consequences . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1572.3. Genetic basis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 157
3. The evolution of drought avoidance in natural populations . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1583.1. Prevalence . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1583.2. Fitness consequences . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1593.3. Genetic basis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1593.4. Resource reserves as drought avoidance adaptations . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1593.5. Genetic tradeoffs in the evolution of drought resistance . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 160
4. A path toward understanding drought resistance adaptations in natural
Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
∗ Tel.: +1 434 924 0162.E-mail address: [email protected]
ttp://dx.doi.org/10.1016/j.plantsci.2015.02.012168-9452/© 2015 Published by Elsevier Ireland Ltd.
populations . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 160. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 161
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 161

1 ience
1
femsdatuesauiDub
btpiHpafgsdteppgt
retetieodsawsaDatdsmamoiaaao
56 N.J. Kooyers / Plant Sc
. Introduction
Climate models forecast an increase in both the severity andrequency of drought in the coming 50 years [1]. This change isspecially difficult for sessile organisms such as plants, whichust be able to respond to wide fluctuations in growing sea-
on conditions while still maintaining the ability to correctly timeevelopmental processes in response to environmental cues. At
population level, increasing aridity and drought should leado strong directional selection for plants with higher fitnessnder drought conditions (i.e., drought-resistant plants); how-ver, a more nuanced understanding of genes and traits underelection is limited by an incomplete knowledge of the mech-nisms that plants use to resist drought stress [2]. Withoutnderstanding the innate resistance mechanisms plants possess,
t is difficult to accurately assess future population persistence.etermining the prevalence and variation in the mechanismsnderlying stress resistance and adaptation is a key goal for plantiologists.
Unlike in natural populations, responses to drought stress haveeen widely studied in a few major crop plants [3–7]. This litera-ure has resulted in an improved understanding of the physiologicalathways involved in drought perception and response as well as
dentified major-effect genes controlling drought resistance [3].owever, wild populations often harbor large pools of genetic andhenotypic diversity that can provide insights into novel mech-nisms of acquiring drought resistance. These insights can rangerom characterizing new phenotypes to identifying new roles forenes involved in abiotic stress-response pathways. While under-tanding the diversity and prevalence of mechanisms underlyingrought resistance in natural populations clearly benefits evolu-ionary biologists, these results can also help agronomists moreffectively improve or develop crops. Here I synthesize recentrogress describing how drought resistance has evolved in naturalopulations of herbaceous plants. I focus on studies that identify theenetic basis of drought strategies as well as describe the evidencehat these strategies are advantageous in natural populations.
Adaptation to soil water availability is common across theanges of plant species and is associated with the formation ofcotypes [8,9]. Adaptations conferring drought resistance have his-orically been divided into three alternative strategies: droughtscape, drought avoidance, and drought tolerance [10]. Each ofhese strategies may evolve as a constitutive response that occursndependently of environmental cues such as water deficit, or canvolve as a heritable plastic response that is dependent on oner more environmental cues. Drought escape occurs when plantsevelop rapidly and reproduce before drought conditions becomeevere. Cession of vegetative growth may or may not accompany
drought escape response. In contrast, drought avoidance occurshen plants increase water-use efficiency (WUE) by reducing tran-
piration, limiting vegetative growth, or increasing root growth,nd avoid dehydration during transient periods of drought stress.rought avoidance has also been referred to as dehydration avoid-nce in recent literature. Finally, drought-tolerant plants are ableo withstand dehydration through osmotic adjustment and pro-uction of molecules that stabilize proteins (Fig. 1; [10]). Thesetrategies are coordinated physiological syndromes that involveany physiological and structural traits [11]. For instance, drought
voidance through increased WUE is mediated by lowering sto-atal conductance, which in turn can be influenced by a number
f different potentially correlated traits such as leaf area, leaf lob-ng, succulence, or stomatal density. Here, I will focus on recent
dvances understanding drought escape and avoidance. Thesedvances are largely limited to studies examining flowering times a measure of drought escape and leaf-level WUE as a measuref drought avoidance as these are the traits that have received234 (2015) 155–162
the most attention. Mechanisms of drought tolerance have beencovered in detail elsewhere [3,12].
Although each of these strategies is predicted to evolve in areasof frequent drought stress, they are often viewed as alternativestrategies or syndromes that can be optimally employed in spe-cific seasonal contexts for plants with specific life history strategies(Fig. 1; [6,13]). For instance, drought escape may be optimal forannual plants in environments with short growing seasons that areended by severe terminal drought; whereas drought avoidance maybe more optimal if the growing season is punctuated by transientdroughts. These strategies are unlikely to evolve together becauseplants devoting all of their resources to rapid reproduction needto have high rate of carbon fixation and thus also high stomatalconductance. However, plants typically avoid drought by loweringstomatal conductance to conserve water and thus reducing the rateof carbon fixation and growth. The literature has largely supportedthis view with the most detailed examples pointing toward theindependent evolution of drought escape and avoidance strategies[14,15]. There is limited evidence in some systems that suggeststhat there are not genetic constraints to the concurrent evolutionof both strategies within individual populations [16]. The environ-mental conditions that favor evolution of specific strategies is stillan open topic and identifying the genetic constraints and fitnessramifications associated with each strategy is an area of stronginterest.
While phenotypes associated with escape or avoidance strate-gies have often been studied (e.g., [14,17,18]), obtaining a detailedunderstanding of the genetic and physiological mechanisms thatplants use to escape or avoid drought in natural populations hasbeen challenging. Recreating realistic drought conditions in anexperimental setting is difficult and may not necessarily reflect fieldconditions [19]. Drought can combine the effects of water deficitand possible heat stress. Manipulating water availability is compli-cated in dry-down experiments because water uptake is greater inbigger plants; a problem that can create heterogeneity in the tim-ing of water deficits [20]. An additional challenge is finding specieswith populations that thrive across a range of aridity conditions andthat also possess a genetic toolbox amenable to exploring the genesand pathways responsible for adaptive divergence in morphologyand physiology. In model genetic species where the genetic basisof drought escape or avoidance has been characterized, there areoften multiple QTL (quantitative trait loci), each of small effect thatunderlie variation in drought resistance. This makes it difficult toidentify the phenotypic effects of a given locus [21,22]. Further,drought escape and avoidance can both be dependent on environ-mental context where a water deficit or other environmental cuemay induce rapid flowering or changes in WUE [23,24]. This inher-ent plasticity can complicate linkage mapping and make it difficultto predict drought resistance and fitness consequences in a seasonalenvironment.
Nevertheless, development of new ecological model systemssuch as Mimulus guttatus, Avena barbata, and Panicum hallii as wellas renewed efforts to study Brassica rapa and Arabidopsis sp. inecological contexts has begun to provide new insights into thegenetics underlying adaptive drought escape and avoidance strate-gies. Specifically, work in these systems has begun to addresslongstanding questions about ecophysiological traits regarding theprevalence, adaptive value, and genetic architecture underlyingvariation in these traits in natural populations [6]. Here I reviewthe advances made in the last decade toward identifying the fit-ness benefits and genetic basis of drought escape and avoidancestrategies as well as the constraints that limit concurrent evolution
of both strategies. This large body of the literature establishes thatvariation in drought escape and avoidance traits is prevalent andadaptive, and highlights promising systems where QTLs and genesresponsible for this variation are known. In addition, this review
N.J. Kooyers / Plant Science 234 (2015) 155–162 157
F ought
b comps
hidt
2
2
cpogtfl(ecgihlaTi
tittte[tpttfuimse[
ig. 1. Schematic diagram describing the three ways herbaceous plants evolve dreen associated. The phenotypes listed here are examples and do not represent a
trategy.
ighlights the importance of gene by environmental interactionsn drought escape and avoidance—a useful insight for engineeringrought resistance in crop plants or predicting population persis-ence in natural populations.
. The evolution of drought escape in natural populations
.1. Prevalence
Plants can escape drought by developing rapidly and reprodu-ing before the onset of drought. Thus, flowering time is often therimary trait associated with drought escape in annual populationsf herbaceous plants. Earlier flowering during a drought-shortenedrowing season presumably results in relatively greater fitnesshrough higher seed set or greater seed mass. Extensive variation inowering time in natural populations has been well documentede.g. [25]), and several studies have demonstrated that earlier flow-ring time evolves due to selection caused by drought [26,27]. Aentral line of evidence for drought escape comes from commonarden experiments where plants from xeric areas flower earliern a common garden than plants from mesic areas. This approachas yielded patterns consistent with drought escape in Arabidopsis
yrata [28], Arabidopsis thaliana [29], Panicum hallii [21], Helianthusnomalus [30], Mimulus guttatus [16], and Boechera holboellii [31].hese results have demonstrated that extensive genetic variationn drought escape exists in natural populations.
Evolution of drought escape occurs not only through a constitu-ive difference in developmental timing, but also through changesn the mechanisms by which populations respond to environmen-al cues, i.e., a plastic drought escape response. Plant time theransition to flowering through temperature, vernalization, pho-operiod, and water availability cues [32–34]. Adaptive droughtscape responses occur through temperature and photoperiod cues35,36]. The role of water availability as an environmental cue forhe transition to flowering is typically considered secondary tohotoperiod and vernalization cues; but may be the most impor-ant for breeding a drought escape response because it provideshe most accurate environmental assessment of drought. Evidenceor an adaptive drought escape response to water deficit in nat-ral populations is limited. For example, the degree of plasticity
n flowering time between wet and dry experimental environ-ents was constant for several Arabidopsis lyrata populations that
panned a latitudinal gradient indicating that enhanced droughtscape response has not evolved within any of these populations28].
resistance. Each strategy has specific traits and phenotypes that have historicallylete list. Arrows designate the environmental context that would most favor each
2.2. Fitness consequences
Evidence for drought escape as an adaptive strategy comes fromtemporally replicated series of observations examining changesin populations following drought events (longitudinal studies;e.g.[26]), phenotypic selection analyses under water-limiting con-ditions (e.g., [18]), and reciprocal transplant experiments (e.g. [37]).Adaptive drought responses have been well-characterized in mul-tiple species. For instance, there was strong selection for earlierflowering in two Brassica rapa populations following drought thatresulted in flowering an average of 1.9 days and 8.6 days earlierin each population [26]. However, the general importance of aplastic drought escape response in natural populations is debat-able given the limited number of species with a documenteddrought escape response. The drought escape response in the sameBrassica rapa populations described above was also examined,and there was no evidence for increased plasticity for floweringtime after the aforementioned drought event [38]. While theseexperiments do not identify plastic drought escape responses asadaptive, few experiments have explicitly tested the adaptive valueof plastic responses to drought. Further empirical and theoreticalstudies are necessary to identify conditions when such acclima-tion may be advantageous. For instance, evolving a highly plasticdrought escape response may be more likely where severe droughtshave occurred infrequently over the evolutionary history of a lin-eage, whereas evolving a constitutive drought escape responsemay occur if droughts have occurred frequently over the sametimespan.
2.3. Genetic basis
The genetic architecture underlying variation in drought escapehas been elucidated in a number of species, although the specificgenes and their physiological effects remain relatively uncharacter-ized. One promising system is Panicum hallii, a switchgrass relativefound along an aridity gradient in the U.S. desert southwest. Botha xeric ecotype (P. hallii var. hallii) and a mesic ecotype (P. halliivar. filipes) exist. There is extreme divergence in flowering timebetween these ecotypes where var. hallii flowers in approximatelyhalf the time (∼44 days) it takes var. filipes (∼85 days) in a commonmesic environment. QTL mapping experiments have identified two
regions that account for small but significant portions of this dif-ference [21]. This extensive variation in flowering time betweengeographically close populations provides a promising system tostudy the ecological genetics of drought resistance.
1 ience
tlbaetitnsttaectitptMfsCrggtltt
delwtWicobedpArSoagobnsaa
ttnicdn
58 N.J. Kooyers / Plant Sc
Studies in more established systems have progressed to identifyhe molecular genetic basis and physiological mechanisms under-ying drought escape. For instance, Mimulus guttatus is quicklyecoming a model plant for studying drought escape and avoid-nce in natural populations [39]. The growing season of annualcotypes of M. guttatus is ended by terminal drought; however,here is extensive variation in the timing and duration of the grow-ng season. A survey of flowering time in a common environmenthat utilized 51 populations collected along 10 separate altitudi-al transects demonstrated that populations with shorter growingeasons flower earlier under inductive conditions [16]. Reciprocalransplant experiments between annual and perennial M. gutta-us ecotypes reveal a clear connection between earlier floweringnd fitness in a population with a short growing season typicallynded by drought [40]. QTL mapping following up on these recipro-al transplant experiments identified several regions contributingo fitness through earlier flowering and higher survival, includ-ng the candidate gene DIV2 [41]. Subsequent work demonstratedhat a single inversion that includes DIV2 is responsible for a largeortion of divergence in flowering time and fitness between eco-ypes and that this inversion is widespread throughout the range of. guttatus [42]. Interestingly, this QTL represented only 1 of 6 QTL
ound for flowering time, indicating that many additional loci ofmaller effect also contribute to drought escape in this system [41].ollectively, this work demonstrates a clear fitness advantage forapidly developing plants in populations with drought-shortenedrowing seasons, as well as identifies an inversion as the molecularenetic basis for variation in drought escape. QTL studies such ashe ones described above provide information about which genes orocations in the flowering regulatory network can be successfullyweaked to induce earlier flowering without producing negativeradeoffs in growth or reproduction.
The above examples target the genes involved in constitutiveifferences in drought escape, but less is known about droughtscape as a plastic response to water deficit. Two experiments uti-izing genetic knockouts in Arabidopsis thaliana have connected
ater availability cues to abscisic acid (ABA) signaling as well aso several genes in the flowering gene regulatory network [33,43].
ater deficit is largely perceived by the roots and promotes anncrease in ABA levels throughout the plant. ABA promotes trans-riptional upregulation of FT, TSF, and SOC1 leading to floweringnly under long days [33]. The expression of GIGANTEA (GI), a mem-er of the photoperiod pathway, is key in ensuring that droughtscape response only occurs under long day conditions. Under longay conditions, GI is expressed and interacts independently of otherhotoperiod pathway members such as FKF1, ZTL, or CO to enableBA-mediated expression of FT and SOC1. It is hypothesized that GIegulates chromatin accessibility and blocks repressors of FT andOC1 (such as SVP and FLC) to allow drought escape responses toccur [33]. However, GI does have other roles in starch metabolismnd carbon signaling in Arabidopsis as well as interactions with theibberellic acid signaling pathway that could promote expressionf FT and lead to a drought escape response [44,45]. These linksetween the ABA signaling network, the flowering gene regulatoryetwork, and water availability signals have not been fully synthe-ized into a coherent framework. Development of this connection is
major priority for understanding how drought escape responsesre elicited and able to vary in natural populations.
While the role for heritable genetic variation in the evolu-ion of constitutive and plastic drought escape is well established,he role of epigenetic variation is more controversial. There areumerous examples detailing how DNA methylation, histone mod-
fication, and chromatin remodeling regulate gene expressiononstitutively and in response to environmental cues such asrought [46]. However, researchers have yet to identify an epige-etic variant that underlies phenotypic differences in phenology
234 (2015) 155–162
and fitness differences within natural populations. In a set ofArabidopsis epiRILs (lines with nearly identical genomes but con-trasting DNA methylation profiles), variation in DNA methylationhas been associated with heritable variation in flowering time aswell as several other important ecological traits [47,48]. This exper-iment demonstrates the potential of epigenetic variation as anunderlying molecular basis for adaptive evolution. Additionally,drought may act to environmentally induce epigenetic changesand lead to a drought escape response. Evidence that epigeneticvariants create adaptive variation in drought escape in naturalpopulations is currently nonexistent, but this does not mean itdoes not exist. Identifying the role of this variation is an openand important question in the drought literature given the sig-nificant role that epigenetic modification plays in eliciting stressresponses [46].
3. The evolution of drought avoidance in naturalpopulations
3.1. Prevalence
Drought avoidance adaptations can occur through a myriad ofmorphological and physiological traits. Accessions adapted to xericconditions may have lower specific leaf area [49], greater succu-lence [50], increased leaf reflectance [51], accentuated leaf lobing[52], and altered stomatal size and density [53], although thesetraits do not necessarily coordinately evolve in a drought avoid-ance syndrome. Many of these traits are expected to contribute tovariation in transpiration efficiency, with drought-avoiding plantsfrom xeric populations having greater WUE through higher photo-synthetic capacity and lower transpiration. Transpiration efficiencyis a measure of performance, and is approximated at leaf-levelthrough both instantaneous WUE and an integrated measure of life-long transpirational efficiency (�13C; [54] for review). In addition toreducing water loss, plants may slow or cease growth in response toincipient drought as a method of avoiding dehydration [55]. Adap-tations conferring drought avoidance may also involve higher ratiosof root to shoot growth prior to drought conditions [31]; this adap-tation may be more advantageous for longer-lived perennial plantsas a larger root system may provide a cost-effective advantage inmultiple growing seasons [56,57]. Unfortunately, root to shoot ratiohas proven difficult to measure in natural populations and bettermethodology is a key need for deciphering the importance of thisphenotype. Here I focus on a drought avoidance strategy achievedby regulating WUE as this phenotype has constituted the majorityof studies.
Similar to drought escape strategies, drought avoidance canevolve as a constitutive change in WUE or as a plastic response towater deficit or other environmental cues. Despite strong evidencethat traits conferring drought avoidance are prevalent in popula-tions located in xeric areas, there are few cases where xeric-adaptedecotypes have consistently higher WUE than mesic-adapted eco-types across several environments [17,31,37,39]. InstantaneousWUE appears to be a more plastic phenotype than flowering timewith a generally low broad sense heritability (h < 0.1) [6]. Someplants with high instantaneous WUE in adequately watered con-ditions have low instantaneous WUE in water-limited conditions[23,58]. Indeed, accessions from xeric areas are frequently reportedto have higher WUE than accessions from mesic areas only underdrought conditions indicating that improved WUE is the productof a gene by environment interaction contingent on the presenceof a water deficit [17,37,59]. Thus, although many studies have
measured water-use efficiencies in lines from natural populations,few conclusions can be drawn about the prevalence of droughtavoidance adaptations in nature. Future experiments need to exam-ine the degree of plasticity in natural populations to draw more
ience
na
3
iRhtmfrdscidhmssptfatidocptrediaPefgu
3
twAugfiTwpadc[
es[ta
N.J. Kooyers / Plant Sc
uanced conclusions about the frequency of drought avoidancedaptations.
.2. Fitness consequences
The innate plasticity in WUE has made it difficult to character-ze relationships between WUE and fitness in natural populations.esults from those that have measured fitness are mixed. Boecheraolboellii from xeric environments had higher instantaneous WUEhrough lower stomatal conductance as well as increased invest-
ent in root biomass and lower leaf mass per unit area [31]. Plantsrom the xeric area were more fit in their native environment in aeciprocal transplant experiment suggesting that these phenotypicifferences led to higher fitness in xeric conditions. Phenotypicelection analyses also suggest that plants with lower stomatalonductance are more fit in xeric environments [17]. However,n the natural populations where these experiments were con-ucted, plants from the xeric environment do not necessarily haveigher WUE or lower stomatal conductance than plants from theesic environments. This suggests that while higher WUE may be
elected in one particular year, directional selection is not con-istently strong enough to cause adaptive differentiation betweenopulations from xeric and mesic areas. The results from other sys-ems are more nuanced and context dependent. There is evidenceor selection for high WUE varieties of Impatiens capensis during
normal relatively dry growing season [60], but low WUE plantshat flower earlier were selected during an early season droughtn a subsequent experiment [61]. This suggests that the type ofrought strategy under selection depends on when drought stressccurs in development and the intensity of the drought. The fitnessonsequences of having higher WUE are often unintuitive in annuallants. A series of papers contrasting xeric- and mesic-adapted eco-ypes of Avena barbata find that plants that are able to acquireesources, develop quickly, and flower early are favored in bothnvironments [62–64]. Notably, plants with lower stomatal con-uctance, such as that found in the xeric ecotype, had no advantage
n any environment, but those with elevated rates of carbon fixationcquire resources and flower more quickly in both environments.erhaps the xeric ecotype would be favored during years with morextreme drought conditions, and reciprocal transplant experimentsailed to catch this variation. Collectively, these experiments sug-est that WUE is not a simple performance phenotype that can besed easily for breeding.
.3. Genetic basis
Given the phenotypic complexity of WUE it is not surprisinghat the physiological mechanisms connecting genetic variationith phenotypic variation are just beginning to be elucidated [53].
limited number of genes underlying variation in WUE in nat-ral populations have been identified. Two notable examples ofenes underlying variation in WUE have recently been identi-ed in divergent Arabidopsis thaliana accessions, Tus-1 and Kas-1.hese accessions have distinctly different physiological strategieshere Tus-1 plants flower earlier and have lower WUE than Kas-1lants. This physiological difference is caused by loss of functionllele at FRI (FRIDIGA) in Tus-1 plants conferring an enhancedrought escape strategy. The loss of a function FRI allele alsoauses decreased WUE through increased stomatal conductance15,65,66].
A separate cross between divergent Arabidopsis lines Landsbergrecta (Ler) and Cape Verde Islands (CVI) have uncovered several
mall-effect QTL that underlie variation in stomatal conductance22]. One of these QTL has a single non-synonymous substitu-ion at a highly conserved glycine residue in the gene MPK12,nd is responsible for a sizable portion of variance in stomatal234 (2015) 155–162 159
conductance [59]. MPK12 encodes a protein that regulates stoma-tal guard cell physiology. Plants with this substitution have largerguard cells and stomatal openings. Interestingly, this substitutionalso has environment-dependent effects; when subjected to ABAtreatment (as occurs following perception of water deficit), plantswith this substitution are not able to retain stomatal closure aslong as plants without this substitution. This suggests a partiallyimpaired ability to respond to a critical environmental cue. All ofthe work conducted on drought avoidance has demonstrated theneed for a greater understanding of gene by environment interac-tions [24]. Future progress in identifying the adaptive significanceand genetic basis of drought avoidance will rely on developing abetter understanding about how water deficit is perceived, as wellas the pathways involved in signal transduction and developmentalresponses. A solid understanding of both the stomatal developmen-tal pathway [67] and the ABA response pathway [68] already existsand connections are rapidly being synthesized. This framework isnecessary for candidate gene approaches, such as those utilized inthe above Arabidopsis research.
3.4. Resource reserves as drought avoidance adaptations
Much of the drought avoidance research has focused on transpi-ration efficiency despite the existence of other drought avoidancemechanisms. The production of compounds that act as stress pro-tectants or stabilize critical proteins during water deficits arenormally associated with the evolution of a drought tolerancestrategy. However, plants may rely on either developing largestores of nutrients in prime conditions and/or diverting resourcesmaintaining homeostasis during initial water deficits as a droughtavoidance strategy. For example, Lupinus alba diverts organic nitro-gen and carbohydrate resources from stems to seed pods at thefirst sign of water deficit. This reallocation results in similar seedset as plants that have not been subjected to drought stress [69].Enhanced carbohydrate storage may be beneficial to all plants thatfrequently undergo drought stress as closing stomata to reducewater loss also reduces CO2 uptake and growth. Whether theseshort-term reserves are used to maintain homeostasis or supporta reduced level of metabolism over longer stretches may dependon the history of drought intensity and frequency within a givenpopulation. Populations adapted to frequent short-term droughtsmay be able to maintain homeostasis, while those accustom tolonger droughts may reduce growth rates and adjust metabolismaccordingly.
During the beginning stages of drought, nitrogen also becomeslimited due to diminished nitrogen reductase activity. Thus, devel-oping basic reserves of usable plant nitrogen would be useful duringshort-term droughts. Cyanogenic glucosides may play this nitro-gen reserve role in white clover (Trifolium repens). The presenceof cyanogenic glucosides is polymorphic in white clover, and thepresence of cyanogenic glucosides has long been thought to actas an herbivore deterrent. New evidence suggests that the abilityto produce cyanogenic glucosides also confers a fitness advan-tage under drought conditions [70]. Cyanogenic glucosides havebeen identified as a nitrogen transport or storage molecules in sev-eral other species [71], and the regulatory network detailing thebreakdown of cyanogenic glucosides in useable plant nitrogen hasbeen characterized in sorghum [72]. Further work must demon-strate that concentrations of cyanogenic glucosides decrease duringdrought, and levels of NO3
− are maintained throughout short-termdroughts. Other common molecules may serve a similar reservoir
or buffering role. For instance, trehalose, a previously identifiedosmoprotectant, may serve as a reserve carbohydrate [73]. Iden-tification of other such short-term reserves may serve as usefuluntapped variation for introducing into crop-breeding programs.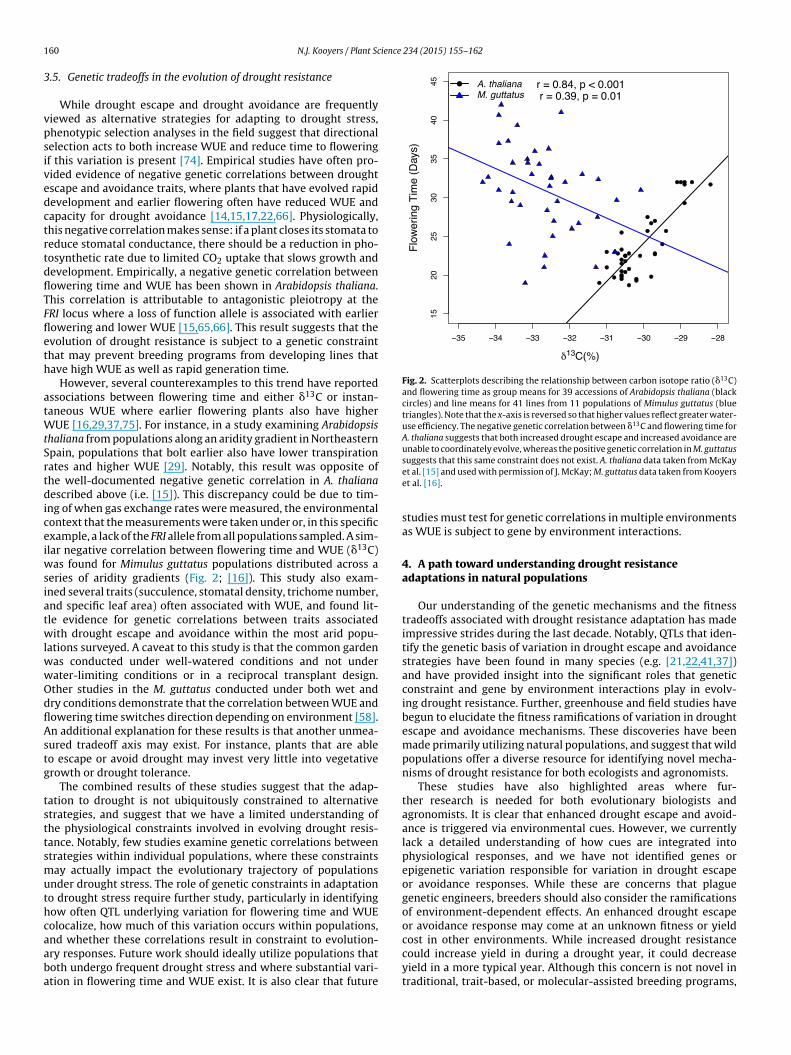
1 ience 234 (2015) 155–162
3
vpsivedctrtdflTFfleth
atWtSrtdiceiwsiatwlwwOdflAstg
tsttsmuthcaaba
δ13 C(%)
Fig. 2. Scatterplots describing the relationship between carbon isotope ratio (�13C)and flowering time as group means for 39 accessions of Arabidopsis thaliana (blackcircles) and line means for 41 lines from 11 populations of Mimulus guttatus (bluetriangles). Note that the x-axis is reversed so that higher values reflect greater water-use efficiency. The negative genetic correlation between �13C and flowering time forA. thaliana suggests that both increased drought escape and increased avoidance areunable to coordinately evolve, whereas the positive genetic correlation in M. guttatus
60 N.J. Kooyers / Plant Sc
.5. Genetic tradeoffs in the evolution of drought resistance
While drought escape and drought avoidance are frequentlyiewed as alternative strategies for adapting to drought stress,henotypic selection analyses in the field suggest that directionalelection acts to both increase WUE and reduce time to floweringf this variation is present [74]. Empirical studies have often pro-ided evidence of negative genetic correlations between droughtscape and avoidance traits, where plants that have evolved rapidevelopment and earlier flowering often have reduced WUE andapacity for drought avoidance [14,15,17,22,66]. Physiologically,his negative correlation makes sense: if a plant closes its stomata toeduce stomatal conductance, there should be a reduction in pho-osynthetic rate due to limited CO2 uptake that slows growth andevelopment. Empirically, a negative genetic correlation betweenowering time and WUE has been shown in Arabidopsis thaliana.his correlation is attributable to antagonistic pleiotropy at theRI locus where a loss of function allele is associated with earlierowering and lower WUE [15,65,66]. This result suggests that thevolution of drought resistance is subject to a genetic constrainthat may prevent breeding programs from developing lines thatave high WUE as well as rapid generation time.
However, several counterexamples to this trend have reportedssociations between flowering time and either �13C or instan-aneous WUE where earlier flowering plants also have higher
UE [16,29,37,75]. For instance, in a study examining Arabidopsishaliana from populations along an aridity gradient in Northeasternpain, populations that bolt earlier also have lower transpirationates and higher WUE [29]. Notably, this result was opposite ofhe well-documented negative genetic correlation in A. thalianaescribed above (i.e. [15]). This discrepancy could be due to tim-
ng of when gas exchange rates were measured, the environmentalontext that the measurements were taken under or, in this specificxample, a lack of the FRI allele from all populations sampled. A sim-lar negative correlation between flowering time and WUE (�13C)
as found for Mimulus guttatus populations distributed across aeries of aridity gradients (Fig. 2; [16]). This study also exam-ned several traits (succulence, stomatal density, trichome number,nd specific leaf area) often associated with WUE, and found lit-le evidence for genetic correlations between traits associatedith drought escape and avoidance within the most arid popu-
ations surveyed. A caveat to this study is that the common gardenas conducted under well-watered conditions and not underater-limiting conditions or in a reciprocal transplant design.ther studies in the M. guttatus conducted under both wet andry conditions demonstrate that the correlation between WUE andowering time switches direction depending on environment [58].n additional explanation for these results is that another unmea-ured tradeoff axis may exist. For instance, plants that are ableo escape or avoid drought may invest very little into vegetativerowth or drought tolerance.
The combined results of these studies suggest that the adap-ation to drought is not ubiquitously constrained to alternativetrategies, and suggest that we have a limited understanding ofhe physiological constraints involved in evolving drought resis-ance. Notably, few studies examine genetic correlations betweentrategies within individual populations, where these constraintsay actually impact the evolutionary trajectory of populations
nder drought stress. The role of genetic constraints in adaptationo drought stress require further study, particularly in identifyingow often QTL underlying variation for flowering time and WUEolocalize, how much of this variation occurs within populations,
nd whether these correlations result in constraint to evolution-ry responses. Future work should ideally utilize populations thatoth undergo frequent drought stress and where substantial vari-tion in flowering time and WUE exist. It is also clear that futuresuggests that this same constraint does not exist. A. thaliana data taken from McKayet al. [15] and used with permission of J. McKay; M. guttatus data taken from Kooyerset al. [16].
studies must test for genetic correlations in multiple environmentsas WUE is subject to gene by environment interactions.
4. A path toward understanding drought resistanceadaptations in natural populations
Our understanding of the genetic mechanisms and the fitnesstradeoffs associated with drought resistance adaptation has madeimpressive strides during the last decade. Notably, QTLs that iden-tify the genetic basis of variation in drought escape and avoidancestrategies have been found in many species (e.g. [21,22,41,37])and have provided insight into the significant roles that geneticconstraint and gene by environment interactions play in evolv-ing drought resistance. Further, greenhouse and field studies havebegun to elucidate the fitness ramifications of variation in droughtescape and avoidance mechanisms. These discoveries have beenmade primarily utilizing natural populations, and suggest that wildpopulations offer a diverse resource for identifying novel mecha-nisms of drought resistance for both ecologists and agronomists.
These studies have also highlighted areas where fur-ther research is needed for both evolutionary biologists andagronomists. It is clear that enhanced drought escape and avoid-ance is triggered via environmental cues. However, we currentlylack a detailed understanding of how cues are integrated intophysiological responses, and we have not identified genes orepigenetic variation responsible for variation in drought escapeor avoidance responses. While these are concerns that plaguegenetic engineers, breeders should also consider the ramificationsof environment-dependent effects. An enhanced drought escapeor avoidance response may come at an unknown fitness or yield
cost in other environments. While increased drought resistancecould increase yield in during a drought year, it could decreaseyield in a more typical year. Although this concern is not novel intraditional, trait-based, or molecular-assisted breeding programs,
ience
eia
cspsainoadiitsbgretiw
tpctcaosaciarccdsigtch
A
oaMvm
R
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
N.J. Kooyers / Plant Sc
xplicitly assessing these costs across a range of environmentss clearly necessary given the gene by environment interactionsssociated with these traits.
Future progress is also needed in determining how seasonalontext and intensity of drought influences evolution of droughttrategies and fitness of plants in natural populations. While Fig. 1rovides ecological predictions describing the drought resistancetrategy that should evolve, these predictions are largely untestednd need to be addressed through experimental manipulationn field conditions. Much of this review has focused on termi-al droughts that end the growing season as these droughts areften associated with strong selection for drought escape or avoid-nce. Less research has been devoted to transient or intermittentrought events, and understanding how plants respond and adapt
n response would provide an interesting contrast. Given thatntermittent drought events take place within a single genera-ion, plastic responses limiting plant growth and increasing WUEhould be highly beneficial. As greater fluctuations in precipitationecome normal, understanding which drought resistance strate-ies are most efficient will obtain greater importance. This includeselatively understudied drought avoidance phenotypes such asnhanced root:shoot ratios and buffering molecules. If these pheno-ypes are responsible for large increases in fitness during droughtsn natural populations, perhaps these hard to measure phenotypes
ould be worth pursuing for breeders and engineers.A continuing priority is determining the importance of adap-
ive constraints in the evolution of drought resistance. Naturalopulations represent excellent systems to study whether geneticonstraints between drought resistance syndromes exist and whenhey are adaptive. In natural populations different levels of geneticorrelation exist in different populations, and nature provides anrena to study when these combinations are adaptive. Discoveryf these constraints will provide insights into the indirect costs ofelecting for certain traits such as flowering or heading earliness,nd could identify new directions for breeding drought-resistantrops that also have high yield. It is naïve to suggest that andeal crop plant should employ both drought escape and avoid-nce strategies. An ideal variety allocates the maximum amount ofesources to produce the highest yield in optimal conditions, and isapable of switching to a drought strategy during incipient droughtonditions. The optimal drought strategy depends on the specificrought conditions of a region. Thus, focus should be directed toynthesizing or finding varieties that are able to respond to incip-ent drought conditions without sacrificing yield during optimalrowing conditions. These conclusions indicate that understandinghe benefits, costs, and basis of gene by environment interactions isritical for developing the next generation of crops and determiningow populations will respond to unpredictable environments.
cknowledgements
I thank University of Virginia and B. Blackman for gener-us financial and intellectual support while writing this articles well as the Blackman Lab group, Jack Colicchio, Christopheruir, Jonathan Gressel and seven anonymous reviewers for pro-
iding helpful criticism and feedback on earlier versions of thisanuscript.
eferences
[1] A. Dai, Increasing drought under global warming in observations and models,Nat. Clim. Change 3 (2012) 52–58.
[2] J.T. Anderson, A.M. Panetta, T. Mitchell-Olds, Evolutionary and ecologi-cal responses to anthropogenic climate change, Plant Physiol. 160 (2012)1728–1740.
[3] H. Hu, L. Xiong, Genetic engineering and breeding of drought-resistant crops,Annu. Rev. Plant Biol. 65 (2014) 715–741.
[
234 (2015) 155–162 161
[4] A.G. Condon, R.A. Richards, G.J. Rebetzke, G.D. Farquhar, Breeding for highwater-use efficiency, J. Exp. Bot. 55 (2004) 2447–2460.
[5] T.E. Juenger, Natural variation and genetic constraints on drought tolerance,Curr. Opin. Plant Biol. 16 (2013) 274–281.
[6] D.D. Ackerly, et al., The evolution of plant ecophysiological traits: recentadvances and future directions, Bioscience 50 (2000) 979–995.
[7] M.M. Chaves, How plants cope with water stress in the field? Photosynthesisand growth, Ann. Bot. 89 (2002) 907–916.
[8] J. Clausen, W.M. Heisey, Experimental Studies on the Nature of Species IV.Genetic Structure of Ecological Races, Carnegie Institute of Washington, Wash-ington, DC, USA, 1958.
[9] G.L. Stebbins, Aridity as a stimulus to plant evolution, Am. Nat. 86 (1952)33–44.
10] M.M. Ludlow, Strategies of response to water stress, in: K.H. Kreeb, H. Richter,T.M. Hinckley (Eds.), Structural and Functional Responses to EnvironmentalStresses, SPB Academic, The Hague, 1989, pp. 269–281.
11] F.S. Chapin, K. Autumn, F. Pugnaire, Evolution of suites of traits in response toenvironmental stress, Am. Nat. 142 (1993) S78–S92.
12] J. Ingram, D. Bartels, The molecular basis of dehydration tolerance in plants,Annu. Rev. Plant Physiol. Plant Mol. Biol. 47 (1996) 377–403.
13] J.P. Grime, Evidence for the existence of three primary strategies in plantsand its relevance to ecological and evolutionary theory, Am. Nat. 111 (1977)1169–1194.
14] M.A. Geber, T.E. Dawson, Genetic variation in stomatal and biochemical limita-tions to photosynthesis in the annual plant, Polygonum arenastrum, Oecologia109 (1997) 535–546.
15] J.K. McKay, J.H. Richards, T. Mitchell-Olds, Genetics of drought adaptation inArabidopsis thaliana: I. Pleiotropy contributes to genetic correlations amongecological traits, Mol. Ecol. 12 (2003) 1137–1151.
16] N.J. Kooyers, A. Greenlee, J.M. Colicchio, M. Oh, B.K. Blackman, Repli-cate altitudinal clines reveal evolutionary flexibility underlies adaptationto drought stress in annual Mimulus guttatus, New Phytol. 206 (2014)152–165.
17] S.A. Dudley, Differing selection on plant physiological traits in response to envi-ronmental water availability: a test of adaptive hypotheses, Evolution 50 (1996)92–102.
18] L.A. Donovan, S.A. Dudley, D.M. Rosenthal, F. Ludwig, Phenotypic selection onleaf water use efficiency and related ecophysiological traits for natural popu-lations of desert sunflowers, Oecologia 152 (2007) 13–25.
19] R.G. Latta, K.M. Gardner, Natural selection on pleiotropic quantitative traitloci affecting a life-history trade-off in Avena barbata, Evolution 63 (2009)2153–2163.
20] H. Poorter, J. Bühler, D. van Dusschoten, J. Climent, J.A. Postma, Pot size matters:a meta-analysis of the effects of rooting volume on plant growth, Funct. PlantBiol. 39 (2012) 839.
21] D.B. Lowry, et al., The genetics of divergence and reproductive isolationbetween ecotypes of Panicum hallii, New Phytol. 205 (2014) 402–414.
22] T.E. Juenger, J.K. Mckay, N. Hausmann, J.J.B. Keurentjes, S. Sen, Identificationand characterization of QTL underlying whole- plant physiology in Arabidopsisthaliana: �13C, stomatal conductance and transpiration efficiency, Plant, CellEnviron. 28 (2005) 697–708.
23] C.E. Edwards, B.E. Ewers, C.R. McClung, P. Lou, C. Weinig, Quantitative vari-ation in water-use efficiency across water regimes and its relationship withcircadian, vegetative, reproductive, and leaf gas-exchange traits, Mol. Plant 5(2002) 653–668.
24] D.L. Des Marais, K.M. Hernandez, T.E. Juenger, Genotype-by-environment inter-action and plasticity: exploring genomic responses of plants to the abioticenvironment, Annu. Rev. Ecol. Evol. Syst. 44 (2013) 5–29.
25] I.M. Ehrenreich, Y. Hanzawa, L. Chou, J.L. Roe, P.X. Kover, M.D. Purugganan,Candidate gene association mapping of Arabidopsis flowering time, Genetics183 (2009) 325–335.
26] S.J. Franks, S. Sim, A.E. Weis, Rapid evolution of flowering time by an annualplant in response to a climate fluctuation, Proc. Natl. Acad. Sci. U. S. A. 104(2007) 1278–1282.
27] J.D. Berger, C. Ludwig, Contrasting adaptive strategies to terminal drought-stress gradients in Mediterranean legumes: phenology, productivity, and waterrelations in wild and domesticated Lupinus luteus L, J. Exp. Bot. 65 (2014)6219–6229.
28] A. Paccard, A. Fruleux, Y. Willi, Latitudinal trait variation and responses todrought in Arabidopsis lyrata, Oecologia 175 (2014) 577–587.
29] M.D. Wolfe, S.J. Tonsor, Adaptation to spring heat and drought in northeasternSpanish Arabidopsis thaliana, New Phytol. 201 (2014) 323–334.
30] L.C. Brouillette, C.M. Mason, R.Y. Shirk, L.A. Donovan, Adaptive differentiationof traits related to resource use in a desert annual along a resource gradient,New Phytol. 201 (2014) 1316–1327.
31] C.A. Knight, H. Vogel, J. Kroymann, A. Shumate, H. Witsenboer, T. Mitchell-Olds,Expression profiling and local adaptation of Boechera holboellii populations forwater use efficiency across a naturally occurring water stress gradient, Mol.Ecol. 15 (2006) 1229–1237.
32] M.J. Kobayashi, et al., Mass flowering of the tropical tree Shorea beccariana waspreceded by expression changes in flowering and drought-responsive genes,
Mol. Ecol. 22 (2013) 4767–4782.33] M. Riboni, M. Galbiati, C. Tonelli, L. Conti, GIGANTEA enables droughtescape response via abscisic acid-dependent activation of the florigens andSUPPRESSOR OF OVEREXPRESSION OF CONSTANS, Plant Physiol. 162 (2013)1706–1719.

1 ience
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
62 N.J. Kooyers / Plant Sc
34] G. Bernier, C. Périlleux, A physiological overview of the genetics of floweringtime control, Plant Biotechnol. J. 3 (2005) 3–16.
35] J.D. Berger, S.P. Milroy, N.C. Turner, K.H.M. Siddique, M. Imtiaz, R. Malho-tra, Chickpea evolution has selected for contrasting phenological mechanismsamong different habitats, Euphytica 180 (2011) 1–15.
36] B.K. Blackman, S.D. Michaels, L.H. Rieseberg, Connecting the sun to floweringin sunflower adaptation, Mol. Ecol. 20 (2011) 3503–3512.
37] M.E. Sherrard, H. Maherali, R.G. Latta, Water stress alters the genetic architec-ture of functional traits associated with drought adaptation in Avena barbata,Evolution 63 (2009) 702–715.
38] S.J. Franks, Plasticity and evolution in drought avoidance and escape in theannual plant Brassica rapa, New Phytol. 190 (2011) 249–257.
39] C.A. Wu, D.B. Lowry, L.I. Nutter, J.H. Willis, Natural variation for drought-response traits in the Mimulus guttatus species complex, Oecologia 162 (2010)23–33.
40] M.C. Hall, J.H. Willis, Divergent selection on flowering time contributes to localadaptation in Mimulus guttatus, Evolution 60 (2006) 2466–2477.
41] M.C. Hall, D.B. Lowry, J.H. Willis, Is local adaptation in Mimulus guttatus causedby trade-offs at individual loci? Mol. Ecol. 19 (2010) 2739–2753.
42] D.B. Lowry, J.H. Willis, A widespread chromosomal inversion polymorphismcontributes to a major life-history transition, local adaptation, and reproductiveisolation, PLoS Biol. 8 (2010) e1000500.
43] Y. Han, X. Zhang, Y. Wang, F. Ming, The suppression of WRKY44 by GIGANTEA-miR172 pathway is involved in drought response of Arabidopsis thaliana, PLoSOne 8 (2013) e73541.
44] K. Eimert, S.M. Wang, W. Lue, J. Chen, Monogenic recessive mutations causingboth late floral initiation and excess starch accumulation in Arabidopsis, PlantCell 7 (1995) 1703–1712.
45] A. Putarjunan, S. Rodermel, GIGANTEA suppresses immutans variegation byinteractions with cytokinin and gibberellin signaling pathways, Plant Physiol.166 (2014) 2115–2132.
46] P.P. Sahu, G. Pandey, N. Sharma, S. Puranik, M. Muthamilarasan, M. Prasad,Epigenetic mechanisms of plant stress responses and adaptation, Plant CellRep. 32 (2013) 1151–1159.
47] F. Johannes, et al., Assessing the impact of transgenerational epigenetic varia-tion on complex traits, PLoS Genet. 5 (2009) e1000530.
48] Y. Zhang, M. Fischer, V. Colot, O. Bossdorf, Epigenetic variation creates potentialfor evolution of plant phenotypic plasticity, New Phytol. 197 (2013) 314–322.
49] R.E. Quiroga, R.J. Fernández, R.A. Golluscio, L.J. Blanco, Differential water-usestrategies and drought resistance in Trichloris crinita plants from contrastingaridity origins, Plant Ecol. 214 (2013) 1027–1035.
50] U. Eggli, R. Nyffeler, Living under temporarily arid conditions—succulence asan adaptive strategy, Bradleya (2009) 13–36.
51] J. Ehleringer, O. Björkman, H.A. Mooney, Leaf pubescence: effects on absorp-tance and photosynthesis in a desert shrub, Science 192 (1976) 376–377.
52] C.M. Talbert, A.E. Holch, A study of the lobing of sun and shade leaves, Ecology
38 (1957) 655–658.53] J. Masle, S.R. Gilmore, G.D. Farquhar, The ERECTA gene regulates plant transpi-ration efficiency in Arabidopsis, Nature 436 (2005) 866–870.
54] T.E. Dawson, S. Mambelli, A.H. Plamboeck, P.H. Templer, K.P. Tu, Stable isotopesin plant ecology, Annu. Rev. Ecol. Syst. 33 (2002) 507–559.
[
[
The author has requested enhancement of the downloaded file. All in-textThe author has requested enhancement of the downloaded file. All in-text
234 (2015) 155–162
55] K. Baerenfaller, et al., Systems-based analysis of Arabidopsis leaf growth revealsadaptation to water deficit, Mol. Syst. Biol. 8 (2012) 606.
56] K. Mokany, R.J. Raison, A.S. Prokushkin, Critical analysis of root: shoot ratios interrestrial biomes, Glob. Change Biol. 12 (2006) 84–96.
57] A. Fitter, R. Hay, Environmental Physiology of Plants, 3rd ed., Academic Press,San Francisco, California, 2002.
58] C.T. Ivey, D.E. Carr, Tests for the joint evolution of mating system and droughtescape in Mimulus, Ann. Bot. 109 (2012) 583–598.
59] D.L. Des Marais, et al., Variation in MPK12 affects water use efficiency in Ara-bidopsis and reveals a pleiotropic link between guard cell size and ABA response,Proc. Natl. Acad. Sci. U. S. A. (2014) 2–7.
60] M.S. Heschel, K. Donohue, N.J. Hausmann, J. Schmitt, Population differentiationand natural selection for water-use efficiency in Impatiens capensis (Balsami-naceae), Int. J. Plant Sci. 163 (2002) 907–912.
61] M.S. Heschel, C. Riginos, Mechanisms of selection for drought stress toleranceand avoidance in Impatiens capensis (Balsaminaceae), Am. J. Bot. 92 (2005)37–44.
62] R.G. Latta, C.M.C. Cain, Path analysis of natural selection via survival and fecun-dity across contrasting environments in Avena barbata, J. Evol. Biol. 22 (2009)2458–2469.
63] R.G. Latta, K.M. Gardner, A.D. Johansen-Morris, Hybridization, recombination,and the genetic basis of fitness variation across environments in Avena barbata,Genetica 129 (2007) 167–177.
64] M.E. Sherrard, H. Maherali, The adaptive signifigance of drought escape in Avenabarbata, an annual grass, Evolution 60 (2006) 2478–2489.
65] J.K. McKay, et al., Genetics of drought adaptation in Arabidopsis thaliana II. QTLanalysis of a new mapping population, Kas-1 x Tsu-1, Evolution 62 (2008)3014–3026.
66] J.T. Lovell, et al., Pleiotropy of FRIGIDA enhances the potential for multivariateadaptation, Proc. R. Soc. B 280 (2013) 20131043.
67] L.J. Pillitteri, K.U. Torii, Mechanisms of stomatal development, Annu. Rev. PlantBiol. 63 (2012) 591–614.
68] A.S. Raghavendra, V.K. Gonugunta, A. Christmann, E. Grill, ABA perception andsignalling, Trends Plant Sci. 15 (2010) 395–401.
69] M.L. Rodrigues, C.M. Pacheco, M.M. Chaves, Soil-plant water relations, root dis-tribution and biomass partitioning in Lupinus albus L. under drought conditions,J. Exp. Bot. 46 (1995) 947–956.
70] N.J. Kooyers, L.R. Gage, A. Al-Lozi, K.M. Olsen, Aridity shapes cyanogenesis clineevolution in white clover (Trifolium repens L.), Mol. Ecol. 23 (2014) 1053–1070.
71] B.L. Møller, Functional diversifications of cyanogenic glucosides, Curr. Opin.Plant Biol. 13 (2010) 338–347.
72] R. Jenrich, et al., Evolution of heteromeric nitrilase complexes in Poaceae withnew functions in nitrile metabolism, Proc. Natl. Acad. Sci. U. S. A. 104 (2007)18848–18853.
73] A.K. Garg, et al., Trehalose accumulation in rice plants confers high tolerancelevels to different abiotic stresses, Proc. Natl. Acad. Sci. U. S. A. 99 (2002)
15895–15903.74] S. Kimball, et al., Phenotypic selection favors missing trait combinations incoexisting annual plants, Am. Nat. 182 (2013) 191–207.
75] M.A. Farris, M.J. Lechowicz, Functional interactions among traits that determinereproductive success in a native annual plant, Ecology 71 (1990) 548–557.
references underlined in blue are linked to publications on ResearchGate. references underlined in blue are linked to publications on ResearchGate.


















