
THE EMBRYOLOGY AND NUTRITIONAL REQUIREMENTS OF ...
90
THE EMBRYOLOGY AND NUTRITIONAL REQUIREMENTS OF REPRODUCTIVE STRUCTURES IN VITRO OF SOME SOLANACEAE DISSERTATION SUBMITTED IN PARTIAL FULFILMENT OF THE REQUIREMENTS FOR THE DEGREE OF MASTER OF PHILOSOPHY IN BOTANY FAIQ AHMAD KHAN DEPARTMENT OF BOTANY ALIGARH MUSLIM UNIVERSITY ALIGARH 1980
Transcript of THE EMBRYOLOGY AND NUTRITIONAL REQUIREMENTS OF ...

THE EMBRYOLOGY AND NUTRITIONAL REQUIREMENTS OF REPRODUCTIVE STRUCTURES
IN VITRO OF SOME SOLANACEAE
DISSERTATION SUBMITTED IN PARTIAL FULFILMENT OF THE REQUIREMENTS
FOR THE DEGREE OF MASTER OF PHILOSOPHY
IN BOTANY
FAIQ AHMAD KHAN
DEPARTMENT OF BOTANY
ALIGARH MUSLIM UNIVERSITY
ALIGARH
1980

C E R T I F I C A T E
This Is to c e r t i f y t ha t the d i s s e r t a t i o n e n t i t l e d
"The embryology and n u t r i t i o n a l requi reaents of reproductive
s t ruc tu re s i a v i t r o of some Solanaceae" i s a bonafide work
ca r r i ed out under my supervision by Hr. Faiq Ahmad Khan and
can be submitted in p a r t i a l fulf i lment of the requiremoits
for the award of degree of Master of Philosophy in Botany*
( SABED A. SIDDIi UI ) READ£a
Department of Botany,

ACKNOWLEDGEMENT
I deem pleasure in expressing my debt of g ra t i tude
to Dr. Saeed A, Siddiqui , Reader, Departmait of Botany,
Aligarh Kuslim Univers i ty , Aligarh, for suggesting the
problem, help and guidance during the preparat ion of t h i s
d i s s e r t a t i o n .
My s incere thanks are also due to Prof. M,M.R.K.
Afr id i , Head, Department of Botany, Aligarh Muslim Universi ty,
Aligarh, for providing research f a c i l i t i e s ,
X remain indebted to my laboratory colleagiws
Halsuddin Ahmad, Shama Parveen Siddiqui , Saeed Ahmad and
K.Y.K, Ansari for the i r help and encouragement in var ious
ways.
( F A I Q AHMAD KHAN )

PREFACE
The research problem dea ls with two aspects - the
embryology and the n u t r i t i o n a l requlreiaents of reproductive
s t r uc tu r e s 4a v i t r o of some Solanaceae, The present
d i s s e r t a t i o n dea ls with the embryology of 3planum khaslanum
Clarke as Par t A and a review on. the nul3>ltlonal requirements
and factors affecting the growth of embryos In a r t i f i c i a l
culture as Part B.

C O N T E N T S
Mi.=_A fags
In t roduct iah Material and methods Previous l i t e r a t u r e cm Soianaceae External morphology-Mi cr osp or oi gi um Microsporogenesi s I'iale gametophyte Megaspor angium Megasporogffiiesls Female gametophyte Po l l ina t ion and course of pollen tube F e r t i l i z a t i o n Development of endosperm Embryogeny Seed Discussion Suamary L i t e r a tu re c i t ed
1 3 4 8 9
11 13 14 IS 16 17 18 19 20 21 22 27 30
PART . B
Introducticm Laboratory technique of embryo cu l tu re Nu t r i t i ona l requirements Factors af fec t ing the growth of embryos in v i t r o . Importance of embryo cu l tu re Limitat ions of embryo cu l tu re Conclusicm L i t e r a t u r e c i t ed Plan of work
36 38 41 50
54 58 61 63 71

PJUECT - A

INTRODUCTION
The family bolanaceae belongs to the 77th order
Solanales of the phylum angiospermae according to Hutchinscm
(1959) and the 6th order of Dicotyledonae, Synqpetalae, the
Tubiflorae according to iiigler and Pran t l (1899-1935) and
the 8th order of Gamopetalae, h i ca rpe l l a t a e , the Polemcmlales
according to Bentham and Hooker (1862-1383),
The family Solanaceae comprises 90 goiera and about
2000 spec ies , whose d i s t r i b u t i o n i s ubiquotous. The genus
Solanum comprises 1700 species (Wi l l i s , 1966), A number of
p l an t s of t h i s family have great economic Importance and are
known for medicinal, food and ornamental va lues . The genus
Solan um has a t t r a c t e d the a t t en t ion of bo tan i s t s for a long
time due to i t s ornamental, food, medicinal values and
morphological p e c u l i a r i t i e s .
In sp i t e of morphological and embryological p e c u l i a r i t i e s
of the inves t iga ted Solanums about a score of i t s species
have he&a. worked out embryologically by Nanett i (1912),
Soueges (1922, 1936), Young (1922, 1923), Kostoff (1926),
Bhaduri (1932,1935, 1936), Kruger (1932), Beamish (1956),
Dnyansagar and Cooper (1960), Miller (1969), Saxoia and
Singh (1969), Karuna (1970), Singh and Mahana (1977) and
Ahmad and Siddiqui (1980). However, the account about the

2
embryology of iaolaamn is rather scanty. Considering the
embryological peculiarities and the number of species
worked out Dr, Saeed A. Siddiqui suggested me to investi
gate some more species of the genus Splanum and see if
embryological features could be helpful in tracing the
evolutionary trends within the genus itself and the
relationships with the allied families.

3
KATERIAL MD METHODS
Hower buds, open flowers and fruits of different
developnental stages were collected and fixed in P.A*A.
The material was dehydrated in alcohol xylol series and
embedded in paraffin wax. The sections were cut at
10-12/U, mounted on the slides, stained with safranin
and fast green combination and mounted in canada balsam.

4
PREVIOUS LITSIATIBE ON SOLAN AC EAE
The e a r l i e s t cwi t r ibut ion oa the embryology of
Solanaceae Is the work of Hofmelster (1858), who observed
a mature embryo sac In Hyoscyamas grifflWiffi ^gOB9i4Aft
a t ropoldes and Sa lp l e lo s s l s p l c t a . Jonsson (1881) studied
the development of embryo sac in Saraeha Jaltomato and
reported tha t the megaspore mother c e l l undergoes meiosis
and produces l i nea r megaspore t e t r a d , the inner-most of
which develops in to eight*nucleate polygonum type of embryo
sac . Similar type of exBljryo sac development has been
recorded by Guignard (1882), Soueges (1907), Palm (1922),
Svensscm (1926), Cooper (1931), Bhaduri (1932), Sanerji
and Jtoaduri (1933), Gtoodspeed (1947), Bernard (1949), Walker
(1955) and Ahmad and Siddiqui (1930) in solanaceous p l a n t s .
On the contrary Nanet t i (1912) in Splanum aairicatum and
Young (1923) in S., tuberosum described Lilium type of
embryo sac development. Bhaduri (1932) c r i t i c i z e d the
observat ions of Nanet t i (1912) and Young (1923) and recorded
polygonum type of femede gametophyte development in Solanum
B|g3.0PKfflft and S. gJ fflTWfi. I'ater Shaduri (1935) emphasized
t h a t the polygcaium type of female gametophyte development
i s a common feature in the genus Solanum. Bhaduri (1932),
Rees-leonard (l935) and Ahmad and Siddiqui (1980) reported
more than one archespor ia l c e l l s in the same nucellus in
S o l i u m mg^oy^sfflft, S. tuberosum and S. triauetrum respect ively

l3
In B r m f e l s l a (Bhaduri, 1935) the megaspore mother
c e l l undergoes meiosis but without wall formation and leads
to the formation of 4-nucleate eaibryo sac . This embryo sac
does not show any further growth, Aruger (1932) recorded
t h a t in S_. nigrum and S. turbigense the embryo sac develops
from the micropylar megaspore ins tead of chalazal one.
However, the findings of Kruger (1932) have been refuted
by Bhaduri (1935).
Modilewski (1935) observed b l spor i c , 8-nucleate s c i l l a
type of embryo sac development in N^gotl^afift SiAn£&* In
3Planum demlssum Beamish (1955) recorded c e l l u l a r type of
endosperm developmait and solanad type of embryogeny.
Dny-ansagar and Cooper (1960) have given de ta i l ed account of
the development of male and female gametophytes, endosperm
and embryo in Solanum phureja. They a lso reported tha t
the polar nuc le i fuse before f e r t i l i z a t i o n and the endosperm
i s a b - i n i t i o c e l l u l a r . The embryogeny conforms to the
Solanad type, Crete (1961) recorded anagrad type of embryo
geny in Capsicum aBQM&» Mohan Ram and Kamini (1964)
recorded tha t the embryo sac in Withania somnifera i s usually
monosporic, one ins tance of a tendency towards bisporic
development has a l so been not iced and the embryogeny follows
Solanad type.

a
Saxoia and Singh (1969) inves t iga ted the embryology
of ?^olMum iy,S£M« tL* aPtyl<?m^ffi» ^* JaaifiM* ^* nodiflorum^
'±» sarachoidea and ^ , villosum and recorded the development
of anther wall l ayers and mierosporogenesis. Ihe pollen
gra ins are t r i - c o l p o r a t e with a smooth axine. They are
shed a t 2-cel led s t age . The ovules are anacaapylotropous,
unitegmic and tenulnuce l la te with one archespor ia l c e l l
but occasional ly two, The development of embryo sac i s of
polygcmum type, twin embryo sac have a l so been recorded,
iiccumulatian of s ta rch grains in the embryo sac has been
observed in ^ , aaericanum only. Polar nuclei fuse be ore
f e r t i l i z a t i o n , endosperiB i s a b - i n i t i o c e l l u l a r and the
embryogeny conforms to the iiolanad type, / d l l e r (1969) in
^olanuia mammosum studied the development oi seed from anatro-
pous ovule, where the coi led embryo i s enclosed by fleshy
endosperm and the cotyledons and hypotyl r a d i c l e axis sure of
equal loagth . The unique maixuilate appendages develop in
the f r u i t of S. a«ffla2Saffi« iiaruna (1970) studied the develop
ment of male and female gametophytes, endosperm and embryo
in offXWW Mffym^^W* 3:n PhYgalika IXgcaraa ^Ingh and i.ahana
(1977) studied the development of anther in normal and induee<i
with gaim&a r ays , dimethyle sulphate and d ie thy le sulphate .
The ea r ly development was s imilar in both but during l a t e r
s tages t ape ta l alanormalities l i k e hypertrophy of t ape ta l c e l l s ,

7
i t s ear ly degenerat ion, formation of periplasmodium and
delayed degeneration were recorded in the anthers of induced
p l a n t s .
Ahmad and Siddiqui (1980) studied the embryology of
^Qlanvm triouetrum and recorded tha t the development of
anther wall l ayers conforms to the Dicotyledonous type of
Davis (1966). The endothecium i s devoid of fibrous thicken
ings and the anther dehisces by ap ica l pore, i he microspore
t e t r a d s are t e t r a h e d r a l , i s o b i l a t e r a l , decussate and
rhomboidal. The pol len grains are shed a t 3-nucleate s tage .
Usually c»ie and sometimes more than cxie archespor ia l c e l l s
are present in the same nuce l lus . Development of female
gametophyte i s of monosporic, 8-nucleate polygonum type
and the mature emoryo sac i s enclosed by integumentary
tapetum. Development of endosperm i s free nuclear and the
embryogeny i s of iolanad type.

EXTHINAL MORPHOLOGY
SPlanum khaslanum Clarke i s an erect shrub,
75-90 cm t a l l . Stem woody, greenish-brown in colour
with woody roo t s tock. Leaves simple, spiny, a l t e r n a t e ,
p e t i o l a t e , p e t i o l e spiny, 5-9 cm long. Lamina with
much variatictti in shape and s i z e , margin spiny with
prominent mid-r ib , l l , 5 - 1 3 . 6 cm long and 8-10 cm broad.
Inf lorescence cymose, flowers with spiny ped ice l ,
eb rac tea te , r egu la r , bisexual , white in colour. Calyx
with 5 sepa ls , gamosepalous, 2-2,5 mm Icaag and 1-1,5 mm
broad, ha i ry and green in colour. Corolla with 5 -pe ta l s ,
1-2 era long 0 ,2-0 ,3 cm broad, gamopetalous and white in
colour . Stamen 5 , epipeta lous , f i lamai t shor t , 2 mm long
and 1 mm th ick . Anther 1-1.2 nm long and 1 mm broad,
bas i f ixed , l i g h t yellow with porous dehiscence. Ovary
b i ca rpe l l a ry , syncarpous, bilobed, ax i le p lacen ta t ion .
S ty le 1,1-1.3 cm Icaig and 1 mm broad, stigma s l i g h t l y
c a p i t a t e , f r u i t a berry, globose, 2 ,5-3,0 cm in diameter,
changing green to yellow during the r ipen ing , seeds many,
I lowering and f ru i t i ng - November-Dec ember
lebruary-Apr i l .

PLATE - 1
The whole p lan t


!)
MICROSPORANGIUM
The young anther I s composed of homogenous c e l l s
bounded by a well defined epidermis. The hypodermal
archesporium d i f f e r e n t i a t e s a t the foxir corners of the
young anther and thus the anther becoaies four chambered
( H g , 1 ) , The archesporium cons i s t s of a row of c e l l s
having dense cytoplasm and prominent nuc le i (F igs . 1, 2 ) .
The a rchespor ia l c e l l s divide p e r i c l i n a l l y producing a
primary p a r i e t a l l a y ^ towards the epidermis and the
primary sporogenous layer towards the inner side (Fig, 3 ) ,
The c e l l s of priniary sporogenous layer a re polygonal with
dense cytoplasm and prominent n u c l e i . The c e l l s of primary
p a r i e t a l layer d iv ide p e r i c l i n a l l y producing two layers
(P ig . 4 ) , the inner one d i f f e r e n t i a t e s as tapetum and the
outer layer d iv ides again in a s imilar plane forming two
l aye r s ( i i g . 5 ) , The outer one of which d i f f e r e n t i a t e s
i n t o the ^idothecium and the inner one d i f fc i rent ia tes
as middle l aye r , i'he c e l l s of endothecium and middle
l aye r s d iv ide p e r i c l i n a l l y producing 2-3-layered endo-r
thecium and middle l ayers (Pig . 6 ) . The r a d i a l walls of
a l l the wall l ayers a re in a l i n e showing the i r conmon
o r i g i n . Thus the development of anther wall l ayers conforms

to the Dicotyledonous type of Davis (1966),
The epidermal cells of the developing anther also
undergo repeated anticlinal divisions in order to cope up
with the rapidly enlarging internal tissues. The endo-
thecial layers attain maximum development at pollen grain
stage, and the cells are devoid of fibrous thickenings.
The cells of middle layers possess vacuolated cytoplasm
and are in the form of narrow strips. The tapetum is single
layered, composed of glandular cells with dense cytoplasm.
The tapetal cells become binucleate (I'ig, 6), The anther
dehisces by apical pore.

MICROSPORO GENES IS
Prior to the initiatijKi of meiosis a secondary wall
is deposited within the original wall of the microspore
mother cells. The division in microspore mother cells in
an anther are not synchronous, thus different divisicKial
stages of microsporogenesis ranging from prophase 1 to
microspore tetrad may be present in the same anther (Pigs,
7-19), During diakinesis the nuclear membrane disappears
and the chromosomes show tetravalents (Fig, 7), At meta-
phase I, the chromosomes condense and are arranged at the
equator of the cell forming the metaphase plate (lig. 8),
At anaphase I the bivalents begin to move towards the
Opposite poles (Fig, 9), At this stage lagging chromosomes
are also seen (Pig, 10). The chromoso^aes reaoh their
respective poles at telophase I stage (Fig, ii) and loose
their identity and finally dyad is formed ( Pig, 12 ),
The seccHid meiotic division occurs in dyad cells.
At metaphase II, the two metaphase plates of the chromo
somes are formed ( Pig, 13 ), At anaphase II the chromosomes
of metaphase II move towards opposite poles (Pig, 14) and at
telophase II four distinct groups of chromosomes are formed
(Pig. 15), which ultimately lead to the formation of tetrads

1 3
by successive cy tok ines i s . Usually the t e t r ads a r e t e t r a -
hedra l ( f i g , 16 ) , occasional ly rhomboidal, decussate and
i s o b i l a t e r a l ( l i g s , 17-19 ) • Decussate type of te t rad
i s more conunon as compared to the i s o b i l a t e r a l and rhomboidal.
The microspores develop the i r own wai i , although they
continue to l i e for some time within the o r i g i n a l wal l .

i:
MALE GAMfilOPHYTE
Microspore represents the beginning of the male
gametophyte, Ihe wall of the microspore mother c e l l breaks
off and the young microspores are l i be ra t ed in the anther
l o c u l e . The young microspores are somewhat t r iangular in
ou t l i ne with a l a rge nucleus and vacuolated cytoplasm.
Later i t becomes spher ica l and the vacuoles disappear
( F ig , 20 ) , The microspore nucleus d iv ides mi to t i ca l ly
producing a l a rge vegeta t ive and a small g«tierative nucleus
(Pig . 21) , A small generat ive c e l l i s organized a t one side
of the pol len grain which i s del imi ted by a hyaline wall
( J i g , 22) . The pollen grains are shed a t t h i s s tage . The
generat ive nucleus d iv ides ins ide pollen tube forming two
male gametes. The mature pol len grains are spher ica l in
o u t l i n e with a smooth exine. They have three germ pores
i , e . the pollwi grains are three co lpora te . The average
diameter of 20 pol len grains i s 39,2/U.

Explanation of f igures
l i g s . 1-22, Solanum khasianum; raicrosporanglum,
microspor©genesis and male gametophyte,
i^ig. 1, T , s , of young anther showing four archesporia .
Pig, 2 . A pa r t of t . s , of anther showing hypodermal archesporium,
} ig , 3 , A p a r t of t . s , of anther showing sporo-genous and primary p a r i e t a l l a y e r s .
Fig. 4 . T .s , p a r t of antherj the primary p a r i e t a l layer has divided p e r i c l i n a l l y ,
l i g , 5 , T . s . p a r t of anther showing epidermis, endothecium, middle l aye r , t a p e t a l c e l l s and sporogenous t i s s u e .
l i g . 6. T . s . of mature anther showing epidermis 2-layered endothecium, four middle l aye r s , tapeturn and microspore t e t r a d s ,
Pigs, 7-9, Microspore mother c e l l s showing d i a k i n e s i s , metaphse I and anaphase I s tages of micro-sporogenesis r e spec t ive ly .
l i g . 10. Anaphase I showing lagging chromosomes.
l i g . 11 . Microspore mother c e l l a t telophase I s t age .
t i g * 12. Dyad.
t i g s . 13-15. Metaphse I I , anaphase I I and telophase I I s tages of microsporogeaesis.
l i g s . 16-19, Tetrahedral , rhomboidal, decussate and i s o b i l a t e r a l microspore t e t r ads r e spec t ive ly ,
l i g s , 20-22, Uninucleate, b inucleate and 2-cel led pollen grains r e spec t ive ly .


11
MEGASPOHiiNGIUM
The in i t ia t iorx of the ovule takes place when the
anther i s a t the microspore mother c e l l s t age . The ovular
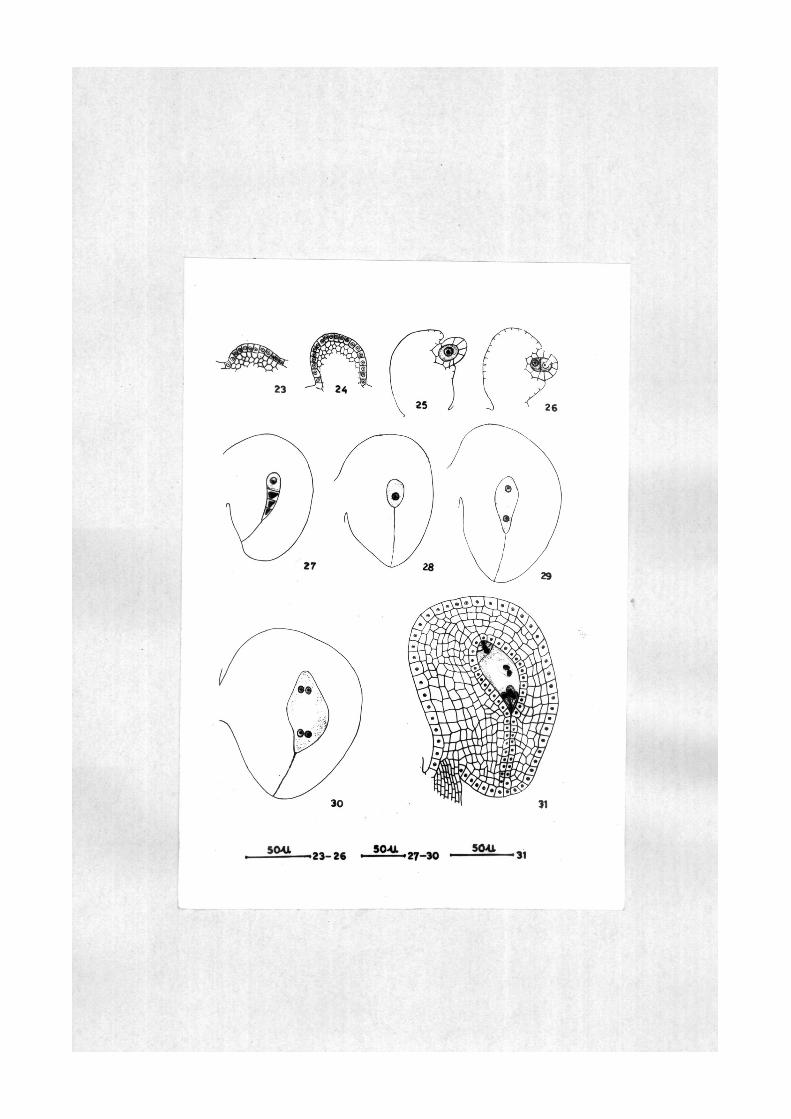
priffiordium i n i t i a t e s as a cy l i nd r i ca l out growth ( l i g s . 23,
24 ) . The young ovule i s composed of homog«mous c e l l s .
The rudiment of integument a r i s e s a t the archespor ia l s tage.
The ovule s t a r t s i t s curvature a t megaspore mother c e l l or »
dyad stage ( Figs , 25, 26 ) . The integument grovs and
reaches the apex of nucel lus forming a narrow niioropyle
a t the megaspore t e t r a d stage ( Fig. 27 ) , The ovule
ccKitinues i t s curvature and a t megaspore t e t rad stage i t
becomes almost hemianatropous ( Figs, 27-30), At mature
embryo sac stage the ovule assumes anatropous configuraticjn
( i i g , 31) , The integumait on the ant i raphe side i s 5-6
layered thick while on the funicular s ide i t i s qui te massive.
The inner most layer of integument d i f f e r e n t i a t e s as
integumentary tapetum and surrounds the embryo sac , Tne
c e l l s of the integumentary tapetum are r a d i a l l y elongated
with glandular contents and prominent n u c l e i . Thus, the
Ovules in Solanum khasianum are anatropous, unitegmic and
t e n u i n u c e l l a t e .

E?^p;^atj^oi^ of flRuy^s
i i g s , 23-31. Solanum khaslanums development of ovule.
i g s . 23, 24. Ovular primordia.
l i g . 26. L , s . ea r ly developmental stage of the ovule with one archespor ia l c e l l and Integumentary prlmordium.
I l g . 26. L . s . of ovule a t dyad s tage .
l i g . 27. L . s , of ovule a t l inea r megaspore t e t r ad stage showing curvature .
l i g s , 28-30, L . s , of anatropous ovules a t functional megaspore, 2-nucleate guid 4-nucleate embryo sac s tages r e spec t ive ly .
i l g . 31 . L . s . of anatropous ovule a t mature embryo sac stage showing integumentary tapetum and micropylar cana l .

MEOASPOROGJiKESIS
The female archesporium is hypodermal in origin.
It possesses dense cytoplasm and prominent nucleus ( !ig.
32 ). The archesporium is generally single celled, occa
sionally two or three celled ( i'lgs, 33, 34 ). In multi
cellular archesporium the cells may be situated side by
side or one above the other ( Figs, 33, 34 ), In one case
in an ovule a dyad is situated towards the chalazal end of
the linear megaspore tetrad ( Fig, 37 ). The archesporial
cell functions directly as megaspore mother cell. It
enlarges considerably and the cytoplasm becomes vacuolated.
The megaspore mother cell undergoes meiosis and produces a
linear megaspore tetrad ( Pigs. 35, 36 ). Usually the
chalazal megaspore remains healthy and the three micropylar
one degenerate (Fig, 38 ), Variations in the number of
healthy megaspores have been observed. Some time the seccmd
megaspore from the chalazal end remains healthy ( lig, 39 ).
In an exceptional case the two chalazal irogaspores remain
functional and rest two degenerate ( Fig. 40 ).

1 '«
FEMALE GAHETOPHYTE
The developmoit of female gametophyte i s of monosporlc,
8-nucleate and polygonum type. The functional megaspore
enlarges considerably and the cytoplasm becomes vacuolated
( l i g , 41 ) . The nucleus of the megaspore d iv ides mi to t i -
c a l l y fOTming two n u c l e i , which move towards opposite poles
r e s u l t i n g in to 2-nucleate embryos sac ( l i g . 42 ) . The
nuc l e i of the two nuclea te embryo sac d ivide mi to t i ca l ly
r e s u l t i n g i n to 4-nucleat« embryo sac ( H g . 43 ) . Side by
s ide the embryo sac enlarges and two nuc le i of each pole again
d iv ide m i t o t i c a l l y producing 8-nucleate unorganized embryo
sac ( i i g . 44 ) . Out of the micropylar quar te t one nucleus
migrates to the cen t re as micropylar polar nucleus and
remaining three organize i n t o 3-cel led egg apparatus . From
the chalazal quar te t three nuc le i organize i n t o 3-antlpodal
c e l l s , while the fourth nucleus migrates to the centre as
cha laza l polar nucleus ( F igs . 46, 46 ) . Normally the
ant ipodals are arranged in a way tha t one c e l l l i e s close
to the chalazal wall and the two facing the centre of the
embryo sac ( Fig . 45 ) . Some time the ant ipodals are
arranged l i n e a r l y ( F ig . 46 ) .

ligs. 32-46, SPlanum khaslanum^ megasporogenesis and
female gatnetophyte.
lig, 32. roung ovule showing hypodermal megaspore mother cell.
Figs, 33,34, Young ovules with two and three celled archesporium respectively.
Dyad stage.
Linear megaspore t e t r a d .
Ovule showing a dyad and l inear megaspore t e t r ad in the same nuce l lus .
Three fficropylar degenerated aegaspores and the chalazal heal thy megaspore.
Second megaspore from the chalazal end i s heal thy and r e s t tliree degenerated.
Two chalazal megaspores are heal thy and r e s t two degenerated.
Functional megaspore.
Two nucleate embryo sac .
Four nucleate embryo sac .
Eight nucleate unorganized embryo s ac .
Mature embryo sac with integumentary tapetum and micropylar canal .
Fig , 46. Mature embryo sac with l i n e a r l y arranged ant ipodal c e l l s .
^ig.
l i g .
J ig .
Fig.
Fig.
Fig.
Fig.
l i g .
Fig.
J ig .
Fig.
3 5 .
3 6 .
3 7 .
3 8 ,
39 .
40 ,
4 1 ,
4 2 .
4 3 ,
4 4 .
4 6 ,

32 36'38 46

r/
POLLINATION AND COURSE OF POLLEN TUBE
The pollinaticm in Solanum khasianum is anemophilous.
The pollen grains germinate oa. stigma by absorbing stigmatlc
secreaticai and produce pollen tubes ( Fig. 47 ). The pollen
tubes enter the stigmatic tissue, grow down the style through
the Intercellular spaces without causing any damage to its
cells and finally reach the top of the ovarian cavity. The
pollen tubes enter the ovarian cavity, reach the placenta
and caater the ovules through the micropylar end of the
embryo sac, where the egg apparatus is situated.

! ^
FERTILIZATION
lertilizatlon is of porogamous type. During the
course of entry of pollen tube into the sac, one synergid
is generally destroyed ( Fig, 48 ), but sometimes both
the synergid8 remain healthy even after fertilization
( Fig, 49 ), Before the entry into the embryo sac the
pollen tube is a delicate tubular structure but inside it
becomes considerably conspicuous. The tip of the pollen
tube inside the embryo sac degenerates and the male gamete:
are released. One of the male gametes fuses with the
egg nucleus resulting into zygote. The second male gamete
fuses with the sec<»idary nucleus forming primary endosperm
nucleus ( H g , 48 ), Thus double fertilization has been
observed in SPlanum khasianum.

If!
DEVELOPMMT 01 ENDOSPBRM
The developmesnt of endosperm i s of c e l l u l a r type.
The f i r s t d iv i s ion of the primary endosperm nucleus i s
t r ansverse producing a primary micropylar and a primary
chaiaza l endosperm chambers ( Fig , 49 ) . The d iv is ion in
the primary chaiazal endosperm chamber precedes tha t in
micropylar chamber ( f i g , 50 )• The d iv is ion in both the
priniary aadosperm chambers i s longi tud ina l forming 4-cel led
endosperm ( l i g s , 50, 51 ) , Some time the primary chaiazal
chamber d iv ides t ransverse ly and the micropylar longi tud ina l
ly forming 4 -ce l l ed endosperm ( H g , 52 ) , Later , the division
in the endosperm c e l l s are qui te i r r egu l a r and lead to the
formation of mul t i ce l lu la r endosperm ( t i g s , 53-58 ) , The
d i v i s i o n in the per iphera l c e l l s of the endosperm are more
frequent as coa^jared to the cen t ra l por t ion . The c e l l s
of the endosperm in the ear ly s tages of development are
highly vacuolated while a t mul t i ce l lu l a r s tage the vacuoles
disappear and the cytoplasm becomes r i c h in s t a rch grains
( r i g , 59 ) , The mature curved dicotyledonous embryo
remains embedded in the c e l l u l a r endosperm ( Fig. 58 ) .

ligs, 47-59. SPlanum khasianum^ pollination, fertiliza
tion and development of endosperm.
Fig, 47, L.s. of stigma showing pollination; the pollen grains are seen on the stigma and the pollen tubes 14 the stylar tissue.
Fig, 48, Embryo sac showing double fertilization.
ligs.49, 50. Two and three celled endosperm respectively.
Fig. 51, Jour celled endosperm, both the primary endosperm chambers have divided longitudinally.
Fig, 52. lour celled endosperm, primary chalazal endosperm chamber has divided transversely and primary micropylar endosperm chamber longitudinally,
five, six and seven celled endosperm respectively.
Multi-cellular endosperm at four celled proembryo and at early globular stages respectively,
Kulticellular endosperm s orro mdlng the mature curved dicotyledonous embryo,
Sndosperm cells showing starch grains.
Figs ,
Figs,
rig.
rig.
53-56,
53,57,
5 8 .
5 9 ,


m. .1
EMEBYOGMY
The zygote enlarges considerably and rena lns
undivided upto 7-cel led endosperm s t a g e . The zygote
d iv ides t ransverse ly producing a terminal c e l l , Si&. ^^^ a
basal c e l l , sik ( ^ i g s . 60, 61 ) . The c e l l fia d ivides
t ransverse ly producing the t i e r s , i and 1 ' ( l l g . 62 ) .
The c e l l £^ a l so d iv ides in the s imi la r plane producing
the c e l l s m and jgi* " ii s the four ce l led proembryo has
l i n e a r d i spos i t ion of i t s c e l l s ( F ig , 52 ) , The c e l l s
ffl and si_ again d iv ide t ransverse ly forming the c e l l s d, f,
a and a l which c o n s t i t u t e the suspensor of the embryo
( l i g . 63 ) . The c e l l s 1 and 1 ' d iv ide long i tud ina l ly
forming quadrant s t age . The quadrant c e l l s are arranged
in two t i e r s of two c e l l s each ( F igs , 64, 65 ) . The
quadrant c e l l s d iv ide long i tud ina l ly forming the octant
s tage ( Fig, 66 ) . The octant s tage by i t s repeated
d i v i s i o n s forms the globular proembryo ( r i g s . 67, 68 ) .
The globular proembryo gradually d i f f e r e n t i a t e s in to heart-
shaped ( t i g . 69 ) , torpedo-stage and f i n a l l y to mature
curved dicotyledonous embryo ( l i g , 70 ) which remains
completely surrounded by c e l l u l a r endosperm. Thus, the
embryogeny in the taxcai conforms to Solanad type.

') I
SSi D
The f r u i t i s a berry with numerous seeds. Xhe seeds
a re small, often f l a t t ened , discoid and brownish yellow
in colour . During the development of endosperm and embryo
the nucel lus and integumentary tapetum a re t o t a l l y absorbed,
The Integument which i s 6-10 c e l l layered a t mature embryo
sac s t age , becomes 15-20 c e l l layers a t mature seed stage
by repeated d iv i s ion of i t s c e l l s . Anatomically the mature
seed c a i s i s t s of seed coa t , p e r s i s t e n t aidosperm and mature
curved dicotyled(mous embryo, At t h i s stage the endosperm
c e l l s are r i ch in s t a r c h g ra ins . The seed coat cons i s t s
of an epidermis and 1 5 - ^ layers of parenchymatous c e l l s
of integument. The epidermis i s the wily mechanical layer
which forms an inner s c l e r o t i c and outer thin walled zcaae
with rod l i k e thickenings <»i r a d i a l and inner tangent ia l
wal ls ( l i g , 71 ) .

ai;pAa|:^aU9B 9^ UKW^^
Figs, 60-71. SPlanum khaslanmnf embryogeny and seed,
}ig. 60. Zygote.
- I g . 61, Two ce l led proerobryo.
J ig , 62. Linear proembrycmic t e t r a d .
Tig. 63 . Six ce l led l inea r proembryo,
i i g . 64. Tier 1 has divided long i tua ina l ly ,
l i g s . 65,66, Quadr^it and octant s tages r e spec t ive ly ,
I i g . 67. Early globular proembryo,
I i g . 68. Globular proembryo.
Fig. 69. Heart-shaped proembryo,
I i g . 70. Mature curved dicotyledonous embryo,
J i g , 71, L , s , of seed coat showing epidermis and c e l l s of integument.

69 70
5CVU SOU SOU s o u
-•60-68' '69—rC—71

DISCUSSION
The present discussion is mainly confined to the
Interesting embryological features of the investigated
species of Solanum and other genera of Solanaceae.
The development of anther wall layers in Solanum
khasianum Clarke conforms to the dicotyledonous type of
Davis (1966) as described in other investigated species of
Solanum. Multilayered nc»i-flbrous endothecium as described
here has been reported in Hicotiana tabacum' and g., rustica
(Jos and Singh, 1968 ) and . triauetrum ( Ahmiad and
Siddiqui, 1980 ), while in Withania somniiera ( Mohan Kam
and Kamini, 1964) and S., nigrum^ £. amerlcanumT S.. nodi-
florum, S_, sarachoides. S,, uteum and §,, vUbsum ( Saxena
and Singh, 1969 ) the endothecium is fibrous. Prom outside
the family multilayered endothecium has been recorded in
Argostemma sarmentosum ( Rublaceae) by Inamuddin (1970) and
Utricularia dichotoma (Lentibulariaceae) by Siddiqui (1978).
3-4 middle layers as described in S,, khasianum have not been
recorded earlier in the genus. However, 1-2 middle layers
have been described in the investigated species of the genus
Solanum ( Saxena and Singh, 1969 ) and 3 middle layers in
Wlthania somnifera ( Mohan Ram and Kamini, 1964 ), Glandular
tapetum as observed here has been described earlier in

2:^
I»yclum europaeum ( J a in , 1956 ) , Wlthanla soagilfera ( Mohan
Ram and Kamini, 1964 ) , S, nigrum^ §., amerlcanumy S,, nodl»
flormar ^* ^aTftg^oJr^gs, 3 . JjjlfiM and S* yj,ll9?^ffi ( Saxena
and Singh, 1969 ) and a., t r iauetrum ( Ahmad and Siddlqui ,
1980). Presence of 1-2 nuclea te t a p e t a i c e l l s as in S.
khasianum has been recorded in hXSlm gMfPP^^W ( Ja in ,
1956 ) and 3., t r iauet rum ( Ahmad and Siddiqui , 1980 ) .
The microspore t e t r ads a r e general ly t e t r ahedra l in
the family. Occasional occurrence of rhomboidal, decussate
and i s o b i l a t e r a l types of microspore t e t r ads as described
in the present taxon has also been recorded in S., nigruffly
^* americanuffly 3 . nodiflorum^ S. sarachoideSt a.luteum and
§., villosum ( Saxena and Singh, 1969 ) and i . t^rj.q^g^ymn
( Ahmad and Siddiqui , 1980 ) . Pollen morphology i s more or
l e s s same in the described species of the genus.
Anatropous ovules as described here have been recorded
in S.. tuberosum ( Young, 1923 ) . In ^ , nigrum. S.. americanum,
§.* sarachoides . S. nodlflprumf 3 , l^t^uai and S. villosum
( Saxena and Singh, 1969 ) and 3 , t r iauetrum ( Ahmad and
Siddiqui , 1980 ) , the ovules are anacampylotropous and
hemianatropous r e spec t i ve ly . Presence of integumentary
tapetum covering the embryo sac i s the c h a r a c t e r i s t i c feature
of the Solanaceae ( Soueges, 1907 ) . Presence of 1-3 cel led

female archesporium as observed in S,, khaslanum has been
recorded e a r l i e r In Lygqp^ysjrgmn esculentumy fliyyaij^s
peruviana^ Nlcotiana pliai]2&gi&lIiS]J^f ^aiVpi^fi^os^? sj^^^ta
and Qrunfelsla amerlcana ( Bhadurl, 1935 ) , i . aL^Mf
§• amerlcanum. S_. nodlfloruiSf S, sarachoideaT 3., luteum and
S_, vll losum ( Saxena and Singh, 1969 ) , and ^ , t r iauetrum
( Ahmad and Siddiqui , 1980 ) .
The megaspore t e t r ads are general ly l inear and the
embryo sac usual ly develops from the chalazal megaspore in
the family Solanaceae ( Ferguscm, 1927; Bhaduri, 1932,
Rees Lecaiard, 1935; William, 1955; i4ohan Ram and Kamini,
1964; Saxena and Singh, 1969; Karu^a, 1970; Parveen §1 al*
1972 and Ahmad and Siddiqui , 1980 ) . Occasionally in
S,, khasianum the second megaspore from the chalazal end and
sometimes both the chalazal megaspores remain healthy and
r e s t degenerate . Variat ions in the heal thy megaspores as
described here have not been described elsewhere in the
family. Ihe development of female gametophyte in the
inves t iga ted species of Splanum conforms to the polygonum
type (Bhaduri, 1932; Kruger, 1932, Dnyansagar and Cooper,
1960; Saxena and Singh, 1969; Karuna, 1970; Ahmad and
Siddiqui , 1980 ) . Nanet t i (1912) and Young (1923) on the
cont rary found a Liliuiii type of embryo sac development in
SQj^^i^m n yriQat v q and S. tuberosum respec t ive ly . Bhaduri
(1932) has c r i t i c i z e d Nanett i (1912) and Young (1923)

2
for recording the Lilium type of embryo sac development,
for he (1932) f inds i t to be of polygonum type in S,
melongena and S,. nigrum. Later , Bhaduri (1935) emphasized
t h a t the polygonum type of development of female gametophyte
i s a common feature in the genus Solanum.
The development of endosperm in the tazon i s of
c e l l u l a r type as described in the other inves t iga ted species
of the iaolanaceae except in S.. t r iauetrum ( Ahmad and
S iddiqui , 1980 ) where i t i s free nuc lea r .
Soianad type of embryogeny as described in S,, khasianum,
i s c h a r a c t e r i s t i c feature of Solanaceae ( ^oueges, 1922;
Bhaduri, 1936; Beamish, 1955$ Jos and Singh, 1968; Karuna,
1968, 1970; Parveen s i Si* 1972; iihmad and Siddiqui , 1980 ) .
From outside the family Solanad type of embryogoiy has a lso
been reported in Eschscholzi^ gftlXfOTfiJigft (Papaveraceae)
(Sachar and Mohan Ham, 1958 ) and Oldenlandia dichotoma
(Kubiaceae)(Siddiqui and Siddiqui , 1968 ) ,
The mature seed comprises seed coat , p e r s i s t e n t
endosperm and mature curved dicotyledonous embryo in the
described species of Solanuqi. In S . khasianum the seed
coat cons i s t s of an epidermis and 15-20 c e l l l ayers of
integument, while in other inves t iga ted species of Splanum
i t cc«isists of an epidermis and p e r s i s t e n t th ick walled

2 ' '
endothelium ( Saxena and Singh, 1969 ) , In 3 . khasianum
the epidermis of the seed i s the main mechanical layer and
i t s c e l l s form an inner s c l e r o t i c and outer th in walled zone
with rod l i k e thickenings on r a d i a l and inner tangent ia l
w a l l s . However, the thickenings are local ized on the
r a d i a l walls only in S. a i ^ M * ^« awgyigm^?a> §.• GLOdjyisoii,
^» saracholdes . S,, luteum and S_. villosum ( Saxena and Singh,
1969 ) .
Considering the number of the species ( about 1700 )
included in the genus Solanum and the paucity of the
erabryological da ta i t i s ra ther d i f f i c u l t to der ive de f in i t e
conclus ions . However, i t appears tha t the general pa t te rn
of development of embryological fea tures in S.khasianum
resembles g rea t ly with other inves t iga ted species of the
genus Solanum^ but i t has i t s own d i s t i n c t i v e features in
which i t resembles ce r t a in species while showing differences
with the other species of the genus. The comparative embryo-
l o g i c a l observations of Solanum species and other genera of
Solanaceae and a l l i e d families as given above can be of much
value with reference to the taxc«aomy and phylogeny of the
genus because the embryological fea tures of 3, khasianum
not coily resembles with the other genera of Solanaceae but
a l so with the a l l i e d f ami l i e s .

2 "
SUMMARY
The external morphology of Solanum Khasianum has been
descr ibed.
The mlcrosporangium i s four chambered and the develop
ment of anther v a i l l ayers conforms to the Dicotyledcmous
type of Davis (1966), The anther wall cons i s t s of an
epidermis, 2-3-layered endothecium, 3-4-middle layers and a
glandular tapetum, the c e l l s of which are two nuc lea te . The
c e l l s of endothecium are devoid of fibrous thickenings.
Melosis in microspore mother c e l l s has been observed. In
some cases aliHiormal meiosis was observed and laggards were
seen a t anaphase I s tage of microsporogenesls. The micro
spore t e t r ads are general ly t e t r a h e d r a l , occasicaially rhom-
bo ida l , decussate and I s o b i l a t e r a l . Pollen gra ins are
3-furrowed with smooth exine and shed a t 2-nucleate s tage .
The average diameter of pollen grains measure 39.2/U, the
anther dehisces by ap ica l pore .
The ovary i s b i ca rpe l l a ry , syncarpous and bi locular
with ax l l e p l acen ta t ion . Ovules are many, anatropous,
unitegmic and t enu inuce l l a t e . The innermost layer of
integument d i f f e r e n t i a t e s as integumentary tapetum. The
female archesporium i s usual ly s ingle ce l l ed , occasional ly

2-3 ce l l ed . The archespor ia l c e l l d i r e c t l y functions as
megaspore mother c e l l , undergoes meiosis and produces a
l i nea r megaspore t e t r a d . Usually chalazal megaspore i s
funct ional , occasional ly second megaspore from the chalazal
end and some time the two chsdazal megaspores remain healthy
and r e s t degenerate . Some time a dyad and a l i nea r magaspore
t e t r a d may be present in the same nuce l lu s . The development
of female gametophyte i s of monosporlc, 8-nucleate and
polygonum type. Generally the ant ipodals are arranged in
a way tha t two c e l l s are close to the chalazal wall and one
facing the cen t re , but some time these are arranged l i n e a r l y .
Po l l ina t ion i s anemophilous. Double f e r t i l i z a t i o n has
been observed. The development of endosperm i s a b - i n i t i o
c e l l u l a r and the f i r s t divisicwi in the primary endosperm
nucleus i s t r ansve r se . The divislcais in both the prinary
endosperm chambers a re Icmgitudinal , sometimes the primary
chalazal chamber may divide t ransverse ly and micropylar
l ong i tud ina l ly forming 4-ce l led endosperm.
The proemtucyonic t e t r ad i s l i nea r and the embrogeny
conforms to the dolanad type. The mature curved dicotyledCKious
embryo remains surrounded by c e l l u l a r endosperm. The seeds
are small , f l a t and brownish yellow in colour. The nucellus
and integumentary tapetum are absorbed dxiring the seed
developB»nt« The seed coat cons i s t s of epidermis, 15-20

c e l l l ayers of integuments. Epidermis i s only the mechanical
layer having the rod l i k e thickening on r a d i a l and inner
t angen t i a l wal l ,
I'he embryology of Solanum khasianum Clarke has been
con^ared with other described species of the genus Splanum
and other genera of i^olanaceae. I t has been concluded tha t
the embryological fea tures of S., khasianum resemble great ly
with the other inves t iga ted species of the genus Solanum,

0 rS '
LITERATURE CITS)
Ahmad, R. and S.A. Slddiqui , 1980. The embryology of
Solanum trlauetrum oav. Communicated.
Banerj l , I , and Bhaduri, P.N. 1933. Polyembryony In
Solanaceae. Current Sc i . 1 (10) : 310.
Barnard, C, 1949. Microsporogenesis, macrosporogenesis
and development of macrogametophyte and seed of
Dubolsla l l e e h h a r d t i l and D« mvoporoides. Aust.
J . Sc i . Res, B, g : 241-248.
Beamish, K. I . 1955, Seed f a i l u r e following hybridizat ion
between hexaploid Solanum demissum and four diploid
SplaRWI spec ies , Amer. Jour . Bot. 4^: 297-304.
Bentham, G, and Hooker, J .D, 1862-1883. Genera plan tar um
London,
Bhaduri, P,N, 1932, The development of ovule and embryo sac
in Solanum a^^XOBS^^ ^» J . Indian Bot. Soc. 1 1 :
202-224.
1935. Studies on the female gametophyte in
Solanaceae. J . Indian Bot. Soc. H : 133-149.
1936. Studies on the ensbryogaiy of the
Solanaceae I , Bot. Gaz, 2Si j 283-295.
Cooper, D.C. 1931. Macrosporogenesis and development of
macrogametophyte of Lvcopersicum eseulentum. Amer,
J . Bot, 2SLi 739-748,

3 '
Cre te , P« 1961. Smbryogenle des solanacees. Oeveloppemmit
de lembrycMi chez l e Gapslcum annuum. C.R, Acad.
Sc i . P a r i s , S§S « 3104-3106.
Davis, G.L. 1966. Systematic embryology of angiosperms,
John Wiley and S<xis I n c . , New York, London, Sydney.
Dnyansagar, V.R, and D.C. Cooper 1960. Development of the
seed of Solanum phureja. Amer. J . 3ot . £7 : 176-186.
Engler, A. and P r a n t l , K, 1899-1935. Die natur l ichen
Pflanzen famil ien. Leipzig Berl in,
lerguson, M.C. 1927. A cyto logica l and genet ica l study of
Petunia I . Bul l . Torrey bot . Club § i : 657-664.
Goodspeed, T.H. 1947, Maturation of the gametes and f e r t i l i
zat ion in Nicot iana. Madrono. 2 ' 110-120.
Gulgnard, L. 1882. Recherches sur l e sac embryonnaire des
phanerogames Angiospermes. Ann. Sc i . Nat. Bot . (6) .
ia» 136-199.
Hofmeister, W. 1858. Neuere Beobachtungenuber Embryobildung
der Phanerogamen, Jahrb, f. Wiss bot, J.; 82-190.
Hutchinson, J . 1959. The families of flowering p l a n t s .
Vol. 2» Sd. II» Oxford,
Inamuddin, M. 1970, A cont r ibut ion to the embryology of some
Rubiaceae, Doctoral t h e s i s , A.M.D., Aljgarh.
J a i n , I . e . 1956. The gametophytes of Lvcium europaeum Linn,
J . Indian Bot. Soc, 2^ : 181-188,
Jonsson, B. 1881. Y t t e r l i ga re bidrag t i l l kannedomen om
Angiospermernas embryos ackutreckling
Bot, Ho t i se r , 169-187,

d
J o s , J . S . and S.P, Singh, 1968. Gametophyte development
and embryog«ay in the genus Hicot iana. J , Indian
Bot. Soc, i2« 117-128.
Karuna, M. 1968. Korphologlcal s tud ies in Solanaceae I I
morphology, development and s t ruc tu re of seed of
iirowallia demissa Linn, Proc, Nat. I n s t . Sci . India
Par t B. Biol . Sc i . 34(3) : 142-148.
1970. Morphological s tudies in Solanaceae
V. Embryology as well as s t ruc tu re and development
of seed of Solanum macranthum Dun. Agra Univ. J .
Hes, S c i . i a ( 2 ) : 55-56.
Kostoff, D. 1926. Icarmation of pol len by seme v a r i e t i e s of
Capsicum ^nnuum. ioin, Univ. Sofia. Fach. Agric,
4 : 101-124.
Kriager, M. 1932. Vergleich«adeent wicklungs geschicht l iche
untersuchungen (m der ffuchtknoten und fruchten
zwiever Solanum Gt- : pf<|; ij|i|- und ihrer Kl ternar ten,
P lan ta . 22» 372-436.
I4 i l ler , Kobert, H. 1969. A morphological study of Solanum
mamiBOSuin and i t s maiimiform f r u i t . Bot. Gaz. 130
(4)J 230-237.
Modilewski, J . 1935. Cytological investigaticHi of the
genus Nicotianay cytology and embryology of
amphidiploid Nicotiana d i t a g l ^ . Vseukr, Akad.
Nauk, I n s t . Bot. Zhurn. 2 (15) : 7-29.

3:1
Mohan Ram, H.Y. and I , Kamini 1964, Embryology and f r u i t
development in Wlthania somnifera. Phytomorphology
2^1 574-587.
Nane t t i , A, 1912. Su l l e p r o b l l l cause d e l l a Parteno carpla
c^el Solanum murieatum. Ai t , Kuov, Giorn. 3ot. I t a l ,
N.S,_19: 91 ( Abstract from Schnarf 's vergleichende
Bmbryologie der ^ngiospermen).
Palm, P, 1922, Zaadvormlng en z a a d s t e r i l i t e l t in Deli Tabak,
Bul l . Deli Proefstaion Madan-Sumatra 16:
Parveen, A . , I . Naseem and KJi . Xhaii 1972. The negasporogene-
s i s and development of embryo sac in y</ithania
somnifera. Pak. J , Sc i . And Res. i^ (3) 196-198.
Rees-Leoiara, O.L. 1935. Microsporogenesis and development
of macrogametophyte of iaolanuiP tuberosum. Bot. Qa^z,
2^ s 734-750,
Sachar, R.C, and Mohan Ram, H.Y. 1958. The embryology of
ESGhscholzia ca i^PrAig^ Cham. Phytomorphology ^ :
114-124.
Saxena, I . and Dalbir Singh 1969. Comparative embryology and
seed s t ruc tu r e of oolanum nigrum complex. Seminar
on Morphology, iinatomy and Embryology of land
p l a n t s . Department of Botany, Ohiversity of Delhi
77-78.
S idd iqui , S.xi., and Mrs, Swaleha B. Siddiqui 1968. Studies
in the Rubiaceae I , A contr ibut ion to the embryology
^^ QXdenlandia dichotoma Hook. }, Be i t r . Biol.
Pflanzen, M '• 343-351.

0 i
Sidd iqu i , S,A, 1978, Studies in the Lent ibular iaceae 8.
The developiront of gametophytes in Ut r i cu la r i a
dichotoma L a b i l l . Flora 2^i 111-116.
Singh, D, and S,K. l-!ahana 1977. Development of anther in
normal and induced Phvsal is ixocarpa moT, J .
Indian Bot. Soc. §^: 281-286,
Soueges, H. 1907. Developpeo^nt e t s t ruc tu re du tegument
seminal chez l e s solanacees, Ann, Sc i . Nat. Bot.
Ser (9) ^i 1-124,
1922, Recherches sur lembryogenie des Solanacees.
Bull . Soc. Bot, France ^ : 136-178? 336-341;
352-365; 555-585.
1936, Developpement de L' embryon chez l e
Schizanthus e t Les Petunias , Bull Soc, bot . irance
^ : 570-577.
Svensson, H.G, 1926, Zytologische-embryologische solana-
ceenstudien, I , Uber d i e samenenlvicklung von
Hyoscyamus n ige r . Svensk. bot l i d s k r , 2^: 420-434.
Walker, R . I . 1955, Cytological and embryological s tudies in
SPlanum, Section Juberarium Bull . Torrey bot, CI.
a^i 87-101,
William, E. 1955. Seed f a i l u r e of Chippewa v a r i e t y in
SPlanum tuberosum. Bot. Gaz. HZ* 10 -15 ,

t,* tf
Wil l i s , J .C . 1966. A dictlcaiarjr of the flowering p lants
and f e rns , Camb. Univ. Press Cambridge.
Young, W.J, 1922. Potato ovaries with two embryo sacs .
iimer. J . Bot. ^\ 213-214.
1923. The formation and degeneration of germ
c e l l s in po ta to . Amer.J. Bot. 2Q^t 325-335.

PABT - B

INTRODUCTION
A r t i f i c i a l cu l tu re of excised embryos i s one of the
important aspects of experimental embryology. Embryo cul ture
i s t o t a l l y based on t i s sue cu l tu re technique and i s employed
where the embryo growth i s a r res ted in non-viable seeds.
This i s of considerable i n t e r e s t to the p lant breeders and
h o r t i c u l t u r i s t s for (a) i t enables the rear ing of hybrid
embryos which may otherwise be doomed because of the baneful
inf luence of the maternal t i s sues (b) i t affords a means
of making quick t e s t of the v i a b i l i t y of seeds, and (c) by
t h i s method i t i s poss ib le to byepass the period of dormancy
and reduce the time required for growing a p lan t to maturity,,
To the phys io log is t s i t i s an important tool for understand-
ihg n u t r i t i o n a l requirements of growing embryos.
The older the embryo, the eas ier i t i s to excise and
cu l tu re i t in an a r t i f i c i a l medium. The chief d i f f i c u l t y
l i e s in growing young embryos not only because of the danger
of in jury to the i r t i s sues during d i ssec t ion but because
t h e i r n u t r i t i v e requirements are much more complex than those
of la rger embryos. Young embryos of t«i tend to germinate
precociously without passing through the normal stages of
development and give r i s e to weak seedlings which are of

Q> i
l i t t l e value . However, the foundation for such work has
been l a i d , and i t i s oaly a matter of time tha t even the
youngest embryos w i l l be cul tured successful ly .
In t rac ing the development of t h i s new technique of
embryo cu l tu re , we find tha t Hannig (1904) was the ^ i r s t
to make a successful at tempt. Laibach (1925) showed the
p o s s i b i l i t y of using t h i s method to economic advantage by
cu l tu r ing the hybrid embryos of Linum species . Brink, Cooper
and itusherman (1944) developed a p lant from in tergener ic
cross between Hordeum .lubeturn and Secale cerea le and
Sanders (1948) between the species of Datura. Mukherji
(1951) and Heit (1955) reported tha t embryo cu l tu re t e s t
provides j u s t i f i c a t i o n for the c r i t e r i a of germinabi l i ty .
Therefore, the present inves t iga t ion was undertaken
to find out the n u t r i t i o n a l requirements of embryos ^ v i t r o .

• 3 - '
LABORATORY TBCMIQUE OF adfiKYO CULTURE
tor the cu l tu re of excised embryos i t i s e s sen t i a l to
be acquainted with the laboratory and various techniques
involved, ior the purpose of c u l t u r e , <aie has to prepare a
good number of stock solut ions to cu l tu re the excised embryos.
F i r s t l y the f r u i t s from which the embryos would be excised
are taken to ascept ic chamber which i s espec ia l ly b u i l t to
avoid contamination and equipped with u l t r a v o i l e t lamps
or any other device to make the chamber s t e r i l i z e d .
Although the chamber i s well protected from a l l possible
contaminations, i t i s e s s®i t i a l to s te r i l i sse the walls and
t a b l e s of the chamber by spraying 2 percent mercuric cbloride
so lu t ion . The f r u i t s and seeds are then s t e r i l i z e d by 1,0
percent mercuric chlor ide soluticwi for several minutes
( Lamerts, 1942, Van Overbeek £ i a l* 1942 and Smith, 1944 )
and 70 to 955 ethanol for several seconds, sometime followed
by flaming (Van Overbeek j ^ ^ , , 1942j Haagen-smit jgrt a l .
1945; Mc Lean, 1946). For seeds and embryos, calcium
hypochlori te or sodium hypochlori te solut ion i s often used,
e i the r saturated or at various dilutiCKis for several minutes
depending upon the ma te r i a l s . Their effect iveness may be
increased by addi t ion of a wetting agent as aerosol or
t e r g i t o l .

^ Ci
Instruments to be utilized are also sterilized with
2% mercuric chloride or by flaming, while starting the actual
work, Special attention is paid to avoid contamination and
infection to the material by wearing gown and mask. Silence
is maintained in the asceptic chamber during the whole
operation.
Excision of embryos is made under dissecting microscope
in sterile water, sugar solution, nutrient medium or csi the
moist sterile bits of the blotter to avoid dryness. The
dissected embryos are inoculated in the culture tubes
containing medium. The instruments used once in excisic8i
should be sterilized agedn before using. The mouth of the
culture tubes should be pluged by non-absorbent cotton plugs
to avoid contamination. Finally the cultures are incubated
in culture room where the desired temperature, light and
humidity are maintained. The embryos are usually cultured
at 25'C,
Since the growth of emtaryos and subsequent seedling
formation is usually poorer ia, vitro than under normal
conditions, transfer from aseptic culture to pots of soil
has its difficulties even with healthier plants. Some of
the weaker seedlings are unable to survive transplantation
(Lofland, 1950; Zeibur and Brink, 1951), Seedlings on agar

J, J
media may have few and pcK>r r o o t s , with roo t h a i r s l imited
to areas exposed to a i r (Me Lean, 1946) and probably becon^
es tab l i shed in so i l only af ter new roo t s develop, Honma
(1956) was able to t ransp lan t cul tured Phaseolus seedlings
to s o i l successful ly caily af te r weekly s e r i a l t ransfers to
n u t r i e n t media containing decreasing amount of sucrose.
Seedlings are frequently t ransfer red f i r s t t o s t e r i l i z e d
pots containing sand or other i n e r t porous mater ia l and
watered with a mineral solut ion u n t i l they can be t r ans
planted to s o i l .

4 '
NUmiTIONAL REQUIHEMENIS
The n u t r i t i o n a l requirements of embryos are complex
in a r t i f i c i a l c u l t u r e . The yoiaiger the embryos on excision,
the growth behaviour w i l l be more abnormal. According to
lukey (1934), the s tage of embryo a t the time of excision
i s more important r a the r the d i f f e ren t s a l t solut ions and
same s a l t solut ion of d i f fe ren t concen t ra t ions . Thus, the
n u t r i t i o n a l requirements of excised embryos vary considerably
with the age*
Mineral sa l ts , : Mineral s a l t s were among the f i r s t
substances added to water in the development of cul ture
media for i s o l a t e d embryos. They were f i r s t used according
to formulae derived from experiments CKI n u t r i t i o n of mature
p l a n t s , and include the elements found to be e s s e n t i a l for
p lant growth; potassium, calcium, magnesium, n i t r o g a i ,
phosphorous, sulphur and i ron .
The mineral composition of the most coranonly used
media for growing excised p lant embryos (both pre and post
germinal) i s summarized in Table 1, Some inves t iga to r s
modified s l i gh t l y the composition of the i r media to su i t t he i r
Individual need. The various iron s a l t s used in these media

TABLE 1 - MINERAL SOLUTIONS (Mg/1 of medium)
KNOo Ca(N03)2
KH2PO4 NaHP04H20
Ca3(fo2)2 K2SO4 Na2S04
MgS04 CaSO^
KCl
IeC4H40g
t GwXn
IeP042H20 IeS04H20 162(304)3
H33O3 CUCI2 Kl MnCl2 MnS04 N a2Ho04 ZnCl2 ZnS04
y .-4 1-4 •H
200 800
200 «• • »
-
-
«ft
200 -
mm
-
-
Trace •w
-
.
-
.
--. am
—
n
83 333
83 -•
-
-
-
83 -
42
-
0 . 8 -
-
-
•
-
.
--<••
-
-
5 H
8 1 98
12 • INB
-
-
-
36 «•
65
-
-
-
-
2 .4
.
-
.
-«. mm
-
«•»
0) •p CO
80 200
.
17 -
-
-
200
360 -
65
mm
mm
-
-
2 .5
1.5 -
0 . 8 -
4 . 5 . mm
1.5
Xi
r4 on
•§"3;
85 164
^
•
10 «w
«*
-
18 -
65
-
-
-
1.2 m»
•
-
.
--,m
-
-
^
Mrf sJrt
H
136 -
_
. •
170
-
-
170 170
680
-
-
170 mm
-
>
-
»
-• •
-
«i»
<« 0)
0) r-l
CO "^
202 164
272 138
-
-
349 -
120 -
-
1.8
-
-
-
-
0 .6 0 .2
« B
0 .2 -
0 .2 0 .6
-

have a tendency to p r e c i p i t a t e In prolong cu l tu res and
thereby render the medium iron d e f i c i e n t . Furthermore,
according to S t r e e t , Mc Gonagle and Mc Gregor (1952), t h i s
p rec ip i t a t i cm may also ac t as a ' S t a l l i n g fac to r ' and cause
a d r i f t in the medium's pH towards a l k a l i n i t y , Eappaport
(1954), replaced the r e l a t i v e l y unstable i r a i sulphate
in Kandolph and Cox's cu l tu re medium by a more s tab le iron
complex such as i ron c i t r a t e , ^y the use of such a chelat ing
agent he was able to maintain his embryos in cu l tu re for
longer period without having to renew too frequently the
media.
Very l i t t l e i s known about the ro l e of micro-elements
in the n u t r i t i o n of p lant embryos in cu l tu re media, Nobecourt
(1938) and Gautheret (1939) added berelium, t i tanium, cobal t ,
n i cke l and boron to t he i r cu l tu re media. The necess i ty for
these p a r t i c u l a r elements in p lan t t i s sue cu l tu res seems to
lack experimental b a s i s . Happaport (1954) added Ber the lo t ' s
complete so lu t ion to h i s cu l tu re media and was unable to
de t ec t any influence of the i r presence on the growth of
very young Datura embryos.
The micro-element requirements of embryo cu l tu re media
have not yet beai s tudied thoroughly, and i t i s possible that
fa i lures to grow very young s tages of some embryos success
fu l l y could be a t l e a s t p a r t l y ascribed to the lack of some
group of t r ace elements in the medium.

7 ^
Carbohydrate St The main r o l e of carbohyclrates in the
c u l t u r e medium i s to supply aiergy for the maintenance of
the various l i v ing processes in the heterotroph embryos.
Simultaneously the carbohydrates act a lso as osmotic agent.
Ear l i e r inves t iga t ions on the growth of excised organs in
syn the t i c media assumed tha t monosaccharides, espec ia l ly
glucose, would be a su i t ab le source of carbohydrate (Bobbins,
1922 and Tukey, 1934, 1938), Van Overbeek, Siu and Haagen-
Smit (1944) found tha t in cu l tu re of heart-shaped Datura
embryos sucrose i s a be t te r source than dextrose . Hall (1948)
noted tha t 0 .5^ , 1,0^ and 2,0J^ sucrose was proved be t te r
than d,glucose for the growth of Solanurn nigrum embryos
while 2$ sucrose was used for hybrid tomato embryos (Smith,
1944), Mc Lean (1946) reported tha t 2,0^ sucrose was bet ter
for the growth of hybrid Datura embryos, Doerpinghaus (1948)
cu l tured "heart-shaped" embryos of t a i Datura species in the
media containing sucrose, dex t rose , fructose and mannose.
He (1948) reported tha t sucrose proved to be superior for
a l l t es ted species , lor the other tes ted sugars , a
preference was found depending upcm the spec ies . Thus,
embryos of three Datura species grew well in 4.0J^ fructose
whereas D.. metq^oit^^s grew well on a l l t es ted sugars
except on fructose and D, d iscolor grew only cm sucrose
and dext rose . Sanders (1950) found tha t growth in
D, stramonium with 4,0jl sucrose was 42 times tha t obtained

i' ^ A ft
with 0.6J6, whereas over the same range, growth of comparable
embryos of three other Datura species increased only 1.2 to
2 .7 t imes. Rietsema s i Si* (1963a & b) reported that there
i s no need of sucrose for 4 ,5 mm, jD, stramonium embryos
but the growth was l imi ted , 1,4 to 2.0 mm embryos requi re
1.0^ sucrose, hea r t and preheart s tages requi re 2.0" and
0 , 1 to 0.15 mm requ i re 8-12/2 sucrose. Thus, embryos of
var ious p lan t species r e a c t in d i f f e ren t way to various
sugars not cmly q u a l i t a t i v e l y but a lso quan t i t a t i ve ly .
Nitrogen compounds: In course of many inves t iga t ions with
p lan t embryos, i t was shown tha t they can ass imi la te
inorganic n i t rogen . N i t r a t e s have proved to be most sui table
n i t rogen source, though in some cases anmonium s a l t s have
been found to be good ni t rogen supplJars , Nitrogenous
organic compounds have been assumed to be inadequate nitrogen
source or even some time toxic when used in synthet ic
cu l tu re media ( Burgeff, 1936 ) , Sanders and Burkholder
(1948) t r i ed to analyse the growth promoting act ion of amino
acid complex present in casein hydrolysate; they used the
post germinal growth of young Datura embryos as a bloassay.
I'hey reported tha t addi t ion of casein hydrolysate (100-800
ppm) and equal p a r t of cystein and tryptophane (6,67 mg per

per 100 mg casein hydrolysate) to the basal culture medium
resulted in notable growth of embryos. Similar results
were obtained by substituting for the casein hydrolysate
in the medium, a mixture of ^ amino acid approximating the
composition of casein hydrolysate. Single amino acid or
incomplete mixture of amino acids did not promote embryonic
growth at rates equal to those obtained by the complete
mixture. Thus it is conclided that the growth stimulus of
the combination of twenty amino acids results from physio
logical interaction rather than from the summation of
effects of individual acids. Paris gt. al.. (1963) found
evidence that young Datura embryos supplied with beneficial
sources of organic nitrogen tended to grow less in the
presence of nitrate than in its absence and they also noted
that some amino acids were less effective than their amides.
Rappaport, Satina and Blakeslee (1960) reported that nucleic
acid proved even inhibitory (both ia vitro and 4a vivo ) in
post germinal culture of Datura embryos.
Vitamins: Addition of vitamins to the medium has given
variable results in embryo culture. No response to the
vitamins tested was noted by Smith (1944) for Lvcopersicum
embryos, Steinberg (1947) for Nicotiana seedlings, Sanders
(1960) for Datura embryos. Van Overbeek, Caaklin and

•X :
Blakeslee (1942) reported tha t young Datura embryos in
' t o rpedo ' stage fa i led to develop cotyledons when grown on
a medium devoid of supplementary f a c t o r s . Mditicni of an
a r b i t r a r i l y composed mixture of growth fac tors (ppm: glycine
3 .0 ; thiamine, 0 . 1 5 | ascorbic acid , 20.0} pantothenic acid,
0.5} Nicot in ic ac id , 1.0} vitamin fig, 0,2} adenine, 0,2}
succinic acid, 25.0) proved e f fec t ive in promoting growth
of these embryos i n t o v iable seedl ings . This mixture of
growth f a c t o r s , however, proved ine f fec t ive in the i r attempts
to grow younger Datura embryos ( Less than 0.5 mm ) .
OTHER ORGANIC SUPPLIMiiUTS
Coconut milk (CM) : Certain embryos, espec ia l ly when excised
a t a very young s tage , would not grow, even i f many possible
vitamin mixtures and growth fac tors were added to the medium.
Van overbeek, Conklin and Blakeslee (1941, 1942) used coconut
milk in the cu l tu re media for growth of young Datura embryos.
They a lso noted tha t with autoclaved coconut milk young
embryos grew as an unorganized mass, whereas with imautoclaved
CM normal growth of very young embryos was observed. They
concluded tha t there might be two substances in CM - a heat
s t ab l e one which produces abundant c e l l p r o l i f e r a t i o n , and
another heat l a b i l e , which a f fec t s normal formation of embryo.

4,^
In an experiment they a lso noted tha t in autoclaved CM
there i s hea t - s t ab l e root i n h i b i t i n g substance which brings
about suppression of roo t growth. Van Overbeek, Siu and
Haagen-smit (1944) successfully obtained an ex t rac t from CM
which had an increased a c t i v i t y of 170 times as compared to
the o r i g i n a l sap. Hall (1948) noted pronounced growth of
15 day of S,, nigrum embryos on lukey ' s basic medium with
1,0^ sucrose along with CM,
Other p lant e x t r a c t s ! Van Overbeek si &!• (1944) tes ted
var ious n a t u r a l e x t r a c t s for the i r growth promoting a c t i v i t y .
They found tha t yeast ex t r ac t , wheat germs, almond meal and
e x t r a c t s of Datura ovules showed embryo factor a c t i v i t y but
not equal to tha t of CM in prolonged cu l t u r e s , Rappaport
(unpublished) found similar growth-promoting r e s u l t s in very
young Datura embryos by adding excised semi-soiled Datura
endosperm to the cu l tu re medium. Blakeslee and Satina (1944)
repor ted tha t a powdered malt ex t r ac t solut ion s t e r i l i z e d
by s e i t z f i l t e r a t i o n was an ef fec t ive subs t i t u t e for CM
in the cu l tu re of immature Datura embryos. While s t e r i l i z a
t ion by autoclaving caused inh ib i t i on on embryonic growth.
Solomon (1950) elucidated the nature of these growth i n h i b i t
ing p roper t i e s of autoclaved malt e x t r a c t , found tha t the
growth factor in malt i s not destroyed by autoclaving but
ins tead i s masked by the presence of newly formed growth

4!!
i n h i b i t o r . This i nh ib i to r appears to be p r e f e r e n t i a l l y
soluble in water, non-vo la t i l e and h e a t - s t a b l e .
Auxins I Van Overbeek (1942) noted roo t i nh ib i t i on when
CM was added to the cu l tu re medium and he suspected that an
auxin present in CM may be respons ib le for t h i s i nh io i t i on .
Subsequent s tudies on the in f lu^ ice of auxins on the growth
of immature embryos i a v i t r o yielded various r e s u l t s . Sanders
(1950) was not able to de tec t any influence of lndole -3-
ace t i c acid on Datura embryos, whereas Happaport si ^ * (I960)
repor ted a s l i g h t improvement in the growth of g.. stramonium
embryos caused by addit ion of alpha napthalene ace t i c acid
(0,05 and 0 .1 ppm)j a t higher concentrat ions ( .5-10 ppm )
an i r r e g u l a r twis t ing and bending of embryos was observed.
I t has been suggested tha t embryo growth jji vivo may be
regula ted by the balance between indole ace t i c acid and
counteract ing substances ( Van Overbeek, 1942, Kietsema jgt §X.»
1953b), while indo le -ace t l c acid was iden t i f i ed as inh ib i to r
respons ib le for aborticaa of embryo in hybrid seeds (Kietsema
e t ^ . 1954).

t) <?
FACTORS AFIBCTIMG THE (BUWTH Or EMHRYOS J^ VITRO
Besides n u t r i t i o n a l requiremaits a number of physical
fac to rs a lso inf luence the growth of embryos in a r t i f i c i a l
cu l tu re which are given below:
Effect of temperature;
The optimal temperature for the growth of Datura
embryos ia, v i t r o i s r e l a t i v e l y high. Van Ov^RTbeek, Caiklin
and Blakeslee (1942) cul tured mature Datura embryos a t 25*C,
The embryos develop r o o t s , hypocotyl and small cotyledons
and l a t e r develop in to v iable seedl ings . Van Qverbeek, Siu
and Haagai-smit (1944) reported tha t Datura embryos were f i r s t
maintained a t 25*C and l a t e r 32"»C for maximum growth. Smith
(1944) cul tured hybrid (Lycopersicum esculentum x k* oeruvi-
aaSiffl) embryos f i r s t a t moderately high temperature but l a t e r
a t room temperature .Hall (1948) cul tured mature Solanum iiigrum
embryos with and without endosperm a t 26**C and 32**C, I t was
concluded tha t the growth was s ign i f i can t ly be t t e r a t higher
temperature. Presence of endosperm has no s ign i f i can t effect
ongrowth, oolomon (1950) cul tured torpedo and near ly mature
Datura stramcaiium embryos a t 26*»C. Thus, i t may be ccmcluded
t h a t in general the growth of embryos jjx v i t r o i s be t te r
a t 25-30«>C.

-- i
Kffggt Of URk%i
Generally, the embryos cul tured by various Inves t igators
are kept in dark incubator u n t i l the necess i ty of a l i g h t
source for photosynthet ic a c t i v i t y of the r e su l t i ng young
seedl ings a r i s e s . Younger embryos grow equally v e i l in
dark or l i g h t or grow be t te r in dark (Gorter, 1965) but
s tages approaching germination appear to be stimulated by
or r equ i res l i g h t (Hannig, 1904; La Rue and Avery, 1938;
Randolph, 1946; Mc Lean, 1946 and Konzak a i ^ . 1951). In
dim l i g h t a t 26«»C Datura embryos develop roo t , hypocotyl
and cotyledons (Van Overbeek; Conklin and Blakeslee, 1942).
The hybrid embryos of tomato species were grown at room
temperature under moderate l i g h t (Smith, 1944).
Effect of DH:
The response of Da. ura embryos to pH varies according
to the stage at which they are cultured. During the first
two to four days after inoculation the optimal pH was about
7,0, However, for a greater length of time, the cultures are
maintained at 5,5 pH which appears to be optimum. The
relative growth of Datura embryos increases with the Increase
in pH from 5,0 to 7,0 and above pH 7,0 the growth decreases

f'
and at pH 9,0 the growth is nil. According to Van
Overbeek, Siu and Haagen Smit (1944) the viability of
embryos also increases with the increase in pH upto
about 7,0. They also cultured Datura embryos at pH 5,5 and
6.9 in basic medium with and without coconut milk and
noted that pH 5,6 was better than pH 6.9 for the growth
of older stages. Hall (1948) cultured 16 days old
S. nigrum embryos and recorded that 7.0 pH was optimum
for growth.
Effect of osmotic pressure of the culture medium?
Osmotic values of nutrioit media are variable,
sometime intentionally, but quite often simply as a result
of adding carbohydrates and other substances for better
growth. Hannig (1904) and Dietrich (1924) used 6 to 10
percent sucrose solutions, considering them to be isotonic
to the embryos, Rietsema fit , (1963a) used mannitol to
maintain osmotic value of the medium and confirmed that
sucrose is a carbohydrate source for growing embryos. They
also noted that the osmotic value influences the growth
oi Datura embryos and for younger embryos higher osmotic
values favour the growth. Precocious germination in
Hordeum embryos due to high osmotic value was prevented
by adding casein hydrolysate solution (Ziebur sLii * 1950).

t>
Effect of Agar concentra t ion;
Agar does not play any n u t r i t i o n a l r o l e and i s
genera l ly used to so l i d i fy the iMdiuia. Agar i s generally
used to concentrat ions ranging from 0.6 to 1.6 per cent .
Smith (1944) successful ly cul tured the hybrid tomato
embryos in modified White's medium so l id i f i ed with 0.75
percent agar. Hall (1948) noted the e f fec t of agar con
cent ra t ions ranging from 0.25 to 2,00 percent for the
growth of S . ni£rum embryos. At .25 percent the growth
was best while a t 2.0 p e r c a i t there was no growth. CcHicentra-
ticttis ranging from 0.6 to 1,0 percent gave e r r a t i c r e s u l t s
but were about equal ly good. Datura embryos a t ear ly stages
of development grew be t t e r in a l iquid medium where the
concentrat ion of agar i s low, but with d i f f e r en t i a t i on in to
seed l ing , t rans fe r red to an agar medium was b a i e f i c i a l
(Solomon, 1950),

r
IMPORTANCE or EMBRYO CULTtJRE
The embryo cu l tu re technique for growing mature plants
from otherwise non-viable seeds has made poss ib le the use of
many addi t iona l kinds of p lan t in p r a c t i c a l breeding program
as well as in t h e o r i t i c a l research . Since the time of
Laibach (1925) the technique has been extensively used in
ag r i cu l tu re and h o r t i c u l t u r e where the f a i l u r e to obtain
v iab le seeds i s due to one of the two causes (1) the embryo
of the f i r s t generation abort in the seed, or (2) the i l
hybrid produces seeds which are incapable of supporting the
development of the embryo to the germinable s t age . In
severa l ins tances of in te rgener ic or i n t e r s p e c i f i c matings,
which had been declared ine f fec tua l or discouraging because
of only a low y ie ld of adul t progeny, embryo cu l tu re has
proved a boon. The embryo cu l tu re technique has been used
in ag r i cu l tu re in an i n d i r e c t or d i r e c t ways. In the former
where the embryo abor ts before maturation leaving the seeds
nonviable . Crosses between the p l an t s of d i f f e ren t ploidy
within the same species e . g . , I r i s (Werckmeister, 1934),
2ea (Uttaman, 1949); i n t e r s p e c i f i c hybrids e .g . Galeopsis
(Muntzing, 1930), Prunus and Lilium ( Skirm, 1942; Emsweller
e t a l . , 1962), Datura (Blakesleee and Sat ina , 1944),

.5 J
LycQperslcum ( Smith, 1944), Hordeum ( Konzak e t a l . 1951 ) ,
Cucurblta ( Wall, 1954 ) , Pfiasgol^s ( Haima, 1955 ) , Chrysan
themum ( Kaneko, 1957 ) , Qrvza ( Bouharmwit, 1961 ) e t c . and
in t e rgene r i c hybrids betweaa the species of Hordeum and Secale
(Brink (£t ^ 1 . 1944), between Datura and Bruemansia (Blakeslee
and Sa t ina , 1944), between Triticum and Elvmus (Iwanowskaya,
1946) and between Tripsacum and Zea (1 arquharson, 1957).
Direc t app l ica t ions to ag r i cu l tu re and Hort icul ture
are perhaps most important for measurement of germinabil i ty
and t e s t i ng the v i a b i l i t y of seeds , increase the percentage
germinaticm, breaking dor nancy and induction of adventive
effibryony. Embryo cu l tu re has provided a rapid means of
measuring germinabi l i ty of Prunus seeds since there i s a
good correspaidence between germination of excised embryos
from non-af ter - r ipened seeds and germination of af ter- r ipened
seeds (Tukey, 1944), Similar determinations of germinative
power of 100 kinds of dormant seeds were more r e l i a b l e than
other quick methods (Heit , 1955), and Mukherji (1951) reported
tha t embryo cu l tu re t e s t s provide j u s t i f i c a t i o n for c r i t e r i a
of germinabi l i ty already in use with Zea.
The cu l tu re of embryos from normal seeds may some time
be used to shorten breeding cyc les , p a r t i c u l a r l y where dormant
seeds are concerned. This has been done with rosaceous f ru i t

(,•
t r e e s to obtain in e ight months seedlings which normally
r equ i r e twenty or more months of growth (Von Veh, 1936;
K i c k e l l , 1951), in I r i s to reduce the seed-to-flower in te rva l
from severa l years to l e s s than one year (Randolph and Cox,
1943) and in Rosa to make poss ib le two generations in a year
(Laamierts, 1946) or shortening of the breeding cycle by three
to four months ( Asen, 1948 )•
Application of embryo cu l tu re technique with hetero
logous transplantat icxi i s very in^jortant in ag r i cu l tu re . Hall
(1964, 1956) made a cross between IrlttcWW aestivum and
Secale cereale and f a i l ed . He cul tured the wheat (Triticum)
eiEbryo on rye (Secale) endosperm and the obtained wheat
(Trit icum) plant was crossed with rye (Secale) and produced
the hybrid ' T r i t i c a l e ' .
The response of an embryo to a p a r t i c u l a r n u t r i e n t
medium depends upon the species as well as upc»i the stage
of development (Sanders and Burkholder, 1948; Sanders, 1950*,
Wright and Srb, 1950 and Hijven, 1956 and 1958). Such var ia
t ions make poss ib le the i nves t iga t ions of physiological
d i f fe rences associa ted with genetic d i f fe rences . Most of the
appl ica t ions of embryo cu l tu re technique which i s r e l a t ed
to p l an t physiology are of fundamental na tu re . Plant physio
l o g i s t s study the physiology of adul t p lan t s and when they

want to study the physiology of embryo, they always follow
embryo cu l tu re technique to stuay the n u t r i t i o n a l r equ i re
ments of embryo, fac tors affect ing the growth and d i f fe ren t ia
t i o n , fac tors con t ro l l ing the development of root and shoot
and fac tors respons ib le for dormancy and germination. By
using th i s technique, the embryo can be grown without
endosperm (outs ide the parent p l a n t ) . Tissue d i f f e r en t i a t i on
can a l so be s tud ied .

LIMITATIONS 0} EMBRYO CULTURE
Despite many years of experimentation and i t s applica
t ion in p r a c t i c a l biology and fundamental research , the
problem of the cult"aring of embryos cannot be regarded as
solved. The embryo undergoes a gradual t r a n s i t i o n frcMii a
s tage of dependence in the beginning to tha t of r e l a t i v e
autonomy, Ccmsequehtly, i t s requirements for growth also
change with age. Of i n t e r e s t in th i s conn-ectlon i s
the work of Haghavan and Torrey (1963) on the embryos of
an i n t e r v a r i e t a l hybrid of Ca t t l eva . Ihey observed tha t with
progressive ontogenetic development the emoryo acquired a
capaci ty for ac t iva t ion of ce r t a in enzyme systems, especia l ly
those concerned with the u t i l i z a t i o n of ni t rogen compounds.
The a b i l i t y to ass imi la te n i t r a t e ni t rogen corresponded with
tne appearance of the enzyme n i t r a t e reductase in the embryo.
In the seed the embryo i s nourished by the t i s sue
adjacent to i t , namely the endosperm except in Trapaceae,
Podostemaceae and Orchidaceae, where there i s l i t t l e or no
endosperm formed. I t i s , there fore , log ica l to presume that
as a r u l e the changes in the contents of the endosperm
l a rge ly correspond to the requirements of the embryo. In
t h e i r at tempts to provide the embryos with n a t u r a l n u t r i e n t s ,

f. a
many inves t i ga to r s have added e x t r a c t of endosperms,ovules,
or f r u i t s of the same species to the cu l ture medium. Van
Overbeek g^ j | i , (1944) reported tha t an ex t rac t of the
ovules of Datura favoured the growth of the young embryos.
Likewise, Ziebur and Brink (1951) observed tha t the endosperm
o^ Hordeum vulnare had a high s t imulatory ef fect on the
growth of i t s embryos. Endosperms and embryos of s imilar
as well as of d i f f e r o i t stages were t e s t ed . The endosperms
were excised and placed around the embryos e i t he r d i r e c t l y
or on the surface of the agar medium. I t was found that the
older endosperms were more e f fec t ive in improving the growth
of young embryos. Further , f reshly excised en t i r e l iv ing
endosperms were superior to (a) overnight-frozen endosperm
(b) a crude e x t r a c t of aadospems, and (c) autoclaved
endosperm.
I t i s curious tha t sometimes "foreign" endosperms
support the growth of embryos be t t e r than the endosperms
of the same spec ies . This amply elucidated by the stimula
t i v e effect which the endosperm of coconut has on embryos
of widely separated spec ies . The pict i i re which emerges
from th i s observation and other inves t iga t ions carr ied out
during the l a s t e ight decades i s tha t underlying the develop
ment of zygote in to the embryo there i s an in te rp lay of many
n u t r i e n t s , growth s t imulants and i n h i b i t o r s , whose exact

K M
duplication In artificial culture is by no means easy.
Consequently when freed from the confines of the ovule,
the young embryo often shows an unrestrained behaviour but
also manifests growth responses of a most varied nature.
There are two chief limitations in exploiting the
embryo culture method to fullest advantage. lirst young
embryos, especially of hybrids, often fail to grow in
artificial media. Second, excised embryos tend to germinate
immediately to produce miniature seedlings rather than to
continue their usual embryonal growth and attain full growth
patterns and weaker seedlings. For a solution of these
difficulties we require a better knowledge of the nutritive
requirements of the embryos in terms of known chemical
substances, further, research will no doubt help to clear
some of the present obscurities in this connection and may
well enable us not only to obtain an uninterrupted growth
of embryos in artificial media but also to follow the entire
process of fertilization and embryo^geny in a petri dish.

I ' *»
CONCLUSION
Culture of plant embryos, although a relatively young
branch of plant physiology, has made valuable contributions
during the last 40 years to our knowledge of certain phases
in the development of such embryos. Outstanding among the
achievements in this field are the atleast two of an applied
nature: (1) Overcoming certain dormancy phenomena in some
seeds, and (2) obtaining viable hybrids which were previously
considered to be unobtainable.
In considering the literature discussed in the review,
one is disappointed in the lack of more specific data on the
physiological functions of various substances and in the
paucity of conclusive explanations of the results derived
from the various treatments of the constituents of the culture
media. No detailed analysis of the function of certain
substances in the developing young embryos has been made.
More than ten years have elapsed since coconut milk was
introduced as a growth-stimulating substance in the culture
of young plant embryos, and even now it is not lonovn what
factor or groups of factors are responsible for this unusual
stimulus. When using this substance, one is still unable to

6
'duplicate nature' in growing very young embryos 'preheart'
and 'proembryos' 4|i vitro. Certainly the foundaticai of such
work has been laid, and it is promising sign that embryo
culture work is being carried out in widely scattered
laboratories. This lack of detailed knowledge is true not
only for the fields of embryo culture but also for a very
closely related and more widely used branch of plant
physiology, tissue culture*

LITERATURE CITED
Asen, 3 , 1948. Embryo cu l tu re of rose seeds. Amer. Rose
Annu. PP. 151-152,
Blakeslee, A.F. and Sat ina , S, 1944. New hybrids from
incoaapatible crosses in Datura through cu l tu re of
excised embryos on malt media. Science 22.* 331-334.
Bouharmcttit, J . 1961. Embryo cu l tu re of r i c e on s t e r i l e
medium. Euphytica lSt» 283-293,
Brink, R.A., Cooper, D.C, and Ausherman, L.E. 1944, A hybrid
between Hordeum JMfeftl? ^ ^ ^ Secale SJS3£MX&» J ^ ^ *
Hered. 2§.» 66-75.
Burgeff, H, 1936. Samenkeimung der Orchideen.
D i e t r i c h , K, 1924, Uber kul tur von KmbrycMiai ausserhalb
des samens. Flora J |^» 379-417,
Doerpinghaus, S.I,, 1948. The e f fec t of various sugars on
the growth of excised embryos in ten species of
Datura. Thesis , Smith College,
Emsweller, S.L, , Asen, S, and Uhring, J . 1962, Tumor
formation in i n t e r s p e c i f i c hybrids of Lilium.
Science iSS* 266,
Farquharson, L . I , 1957, Hybridization of Triosacum and Zea.
Jour , Hered, ^ : 295-299.

i ' f, ! ''• J U I,
Gautheret , R . J . 1939. Sur La p o s s l b l l i t e de r e a l l s e r La
cu l tu re indef in ie des t i s s u s de tubercule de
c a r o t t e . Campt. Rend, Acad. 3 c i . (Par is ) 208: 218.
Gorter, J .C . 1955, J a v i t r o cu l tu re of cyclamen embryos.
Proc. Kon, Ned, Akad. Wet,, Amsterdam 58; 377-385.
Haagen-Smit, A . J . , Siu , R, and Wilson, G, 1945. A method
for cu l tu r ing of excised immature corn embryos
in v i t r o . Science 101: 234,
Ha l l , C.B. 1948, Culture of Splanum nigrum embryos. Proc.
Amer. Soc. ho r t . Sc i . ^ : 343-346.
Hal l , O.L. 1954. Hybridization of wheat and rye af tor
enbryo t r ansp lan ta t ion . Hereditas ^ : 453-458.
1956, Further experiments in embryo t ransplanta
tion* Hereditas 4g: 261-262.
Hannig, E. 1904. Zur physiologic pf lanz l icher Embryonen.
I . Ueber d i e kul tur von cruciferen-Embryonen
ausserhalb des Embryosacks. Zei t schr . 3ot, 62:
45-80.
Hei t , C.S. 1955. The excised embryo method for t e s t ing
germination qua l i ty of dormant seed. Proc, Assoc.
Of f i e . Seed Analysts ^ : 108-117.
Honraa, Shigemi 1955, A technique for a r t i f i c i a l cul tur ing
of bean embryos, Proc, iimer. Soc. ho r t . Sc i , 65;
'K)5-408,

Iwanowskaya, E.V. 1946, Hybrid embryos of cereals grown on
artificial nutrient medium. C.R. (Doklady) Acad,
bci., IBSS §4J 445-448.
Kaneko, K. 1957. Studies on the embryo culture on the inter
specific hybridization of Chrysanthemum. Jap. Jour,
Qenet, gg: 300-305.
Konzak, C.I., Randolph, L.I. and Jensen, K.I. 1951. Emoryo
culture of barley species hybrids. Jour, riered. 42;
125-134.
Laibach, F, 1925, Das Taubwerden von Bastardsam«n und die
kunstliche Aufzucht fruh absterbender Bastardembryo-
nen.. Zeitschr. 3ot, XL* 417-459,
Lanmerts, W.K, 1942, Embryo culture an effective technique
for shortening the breeding cycle of deciduous trees
and increasing germination of hybrid seed. Amer.
Jour. Bot, ^ t 166-171,
1946, Use of embryo culture in rose breeding.
Plants and Gardens 2i 111.
La Kue, C D . and Avery, G.S. Jr. 193B, The development of
the embryo of Zizania aauatica in the seed and in
artificial culture. Bull Xorrey bot. cl. : 11-21.
Lofland, H.d. 1960, la vitrp culture of cotton embryo.
Bot. Gaz, 211* 307-311,
Mc Lean, S.W. 1946, Interspecific crosses involving Datura
ceratocaula obtained by embryo dissection. Amer.
Jour. Sot, aa: 630-638.

0 0
Mukherji, D.K. 1951, Embryo cu l tu re as an aid to seed
t e s t i n g . Proc, n a t , I n s t , dc i , India 3, 17;
253-259.
Muntzing, A, 1930, Outlines to a genetic monograph of the
genus Galeopsis with spec ia l refepotice to the nature
and inher i t ance of p a r t i a l s t e r i l i t y , Hereditas 13;
185-341.
N icke l l , L,G. 1951. Embryo cu l tu re of weeping crabapple,
Proc. Amer. Soc, ho r t . Sc i , §2« 401-405.
Nobecourt, P, 1937, Cultures en se r i e de t i s s u s vegetaux
su^teiilieu a r t i f i c i e l compt. Rend. Acad, (Paris) 205;
521.
P a r i s , D, , Eietsema, J . , Sa t ina , J . and i l akes lee , A,} , 1953,
Effect of amino ac ids , espec ia l ly a spa r t i c and
glutamic acid and the i r amides,on the growth of
Datura stramonium embryos l a v i t r o . Proc, na t ,
Acad. oci,,Wash, SSL^ 1205-1212,
Kaghavan, V. and John, G. Torrey 1963, Grcxrfth and morphogene
s i s of globular and older embryos of Cap s e l l a In
cu l t u r e , Amer, J , itot, §^j 540-551,
Randolph, L.F, 1945. Embryo cu l tu re of I r i s seed. Bull,
.uner. I r i s Soc. 22* 33-45,
and Cox, L.G, 1943, I ac tors influencing the
gerrnination of I r i s seed and the r e l a t i o n of
i n h i b i t i n g substances to embryo dormancy, Proc.
Amer. 3oc, hor t , Sc i . ^ j 248-300.

u /
Rappaport, J , , S, Sat ina and A.F. 31akeslee 1960, Extracts
of ovular tumors and the i r i nh ib i t i on of emrbyo
growth in Datura. Amer, Jour , ,:iot. 22« 686-595,
Rappaport, J , 1964. IQ, v i t r o cu l tu re of p lant embryos and
fac to rs con t ro l l ing the i r growth. Bot.Rev, 20:
201-225.
Rietsema, J . Sa t ina , S. and Blakeslee, A , ! . 1953a, The
effect of sucrose on the growth of Datura stramonium
embryos 4a y i t r o . Amer, Jour , i3ot, ^ : 638-545.
, & 195Sb. The effect
of lndo le -3-ace t ic acid CMI Datura embryos, Proc.
n a t . Acad. S c i . , wash, S^t 924-933,
, & 1954, On the nature
of embryo inh ib i to r in ovular tumors of Datura.
Proc, n a t . Acad. S c i . , Wash, 4^ : 424-431,
Ri;)ven, A.H.G.C. 1956, Glutamine and asparagine as nitrogen
sources for the growth of p lant embryos Xn. v i t r o ;
a comparative study of 12 spec ies . Austra l , Jour.
Biol . Sc i . £x 511-527,
1958, Effects of some inorganic nitrogenous
substances on growth and ass imi la t ion of young plant
embryos i|i, v i t r o . Aus t ra l . Jour . M o l . Sc i , 11 :
142-154,
Robbins, W,J, 1922. CultlvatiDn of excised roo t t i p s and stem
t i p s under s t e r i l e conditicxis, Bot. Ga2. 22.' 376-390,

Sanders, M,E. 1948, Embryo development in four Datura
species following se l f and hybrid p o l l i n a t i o n s ,
Amer. Jour , Sot, 35 t 525-532,
1960. Development of s e l f and hybrid Datura
embryos in a r t i f i c i a l c u l t u r e , Amer, Jour, Bot,
3Zi 6-15,
and Burkholder, P,H. 1948. Influence of amino
acids on growth of Datura embryos in cu l t u r e , Proc,
na t , Acad. S c i , , Wash, 2i« 516-526.
Skirm, G.W, 1942, Embryo cu l tu re as an aid to p lant breeding.
Jour , Hered, 22* 210-215,
Smith, P,G, 1944, Embryo cu l tu re of a tomato species hybrid.
Proc, Amer. Soc. ho r t . S c i . M» 413-416,
Solomon, a, 1960, Inh ib i t ing e f fec t of autoclaved malt
preventing the i a v i t r o growth of Datura embryos.
Amer. Jour . Bot, 22.t 1-6.
S te inberg , R.A. 1947. Growth responses to organic compounds
by tobacco seedl ings in asep t ic c u l t u r e . Jour, Agric.
Bes. 7^: 81-92.
S t r e e t , H.i^., Moira P. Mc Gcaiagle, and Sheila Kc Gregor 1952.
Observat ia is CMI the "S ta l l ing" of White's medium by
excised tomato r o o t s , I I . Iron a v a i l a b i l i t y . Physiol ,
P l an t . §.: 248.

lukey, H.B. 1934. A r t i f i c i a l cu l t u r e methods for Isolated
embryos of deciduous f r u i t s , Proc, Amer. 3oc. hor t ,
Sc i . j2S: 313-322.
1938. Growth pa t t e rn s of p l an t s developed from
inffnature embryos in a r t i f i c i a l c u l t u r e . Sot, Gaz.
§a« 630-665.
1944, The excised embryo method of t e s t i ng the
germinabi l i ty of f r u i t seed with pa r t i cu l a r reference
to peach seed, Proc. Amer, Soc. ho r t , Sc i . 45 :
211-219,
Uttaman, P. 1949, Embryo cu l tu re to obtain Fi p lan ts of
incompatible crosses in corn (maize). Curr. Sc i . 18;
297-299.
Van Overbeek, J . , COaiklin, M.E. and Blakeslee, A.P. 1941.
fac tors in coconut milk e s sen t i a l for growth and
development of very young Datura embryos. Science 94;
350-351.
Van Overbeek, J . 1942, Hormonal cont ro l of embryo and seed
l i n g . Cold Spring Harbor Symp, Quant, Biol , iQt
126-133.
Van Overbeek, J , , Ccmklin, M.E. and 31akeslee, A,?, 1942.
Cul t iva t ion i a v i t r o of small Datura embryos. Amer.
Jour , Bot. S S J 472-477, J . ,
Van Overbeek^^Siu, R, and Haagen-Smlt, A,J, 1944. Factors
a f fec t ing the growth of Datura embryos i a v i t r o .
Amer, Jour , Bot, ^ s 219-224.

7 M
Von Veh, R. 1936. Eine neue methode der Anzucht von
Samllngen, unabhangig von Ruheperioden und
Jah resze i t (bei Apfeln, Blrnen, Qultten, Pflaumen,
Klrschen). Zuchter §,: 145-151.
Wall, J .R. 1954, In t e r spec i f i c hybrids of "Cucurbita"
obtained by embryo c u l t u r e . Proc, Amer, Soc. hor t ,
3 c i . §S« 427-430.
Werckmeister, P. 1934, Uber die kunst l iche aufzucht von
Embryonen aus Iris-.dastard3amen. Gartenbauwiss,
8 : 607-608.
Wright, J .E . and Srb. A.M. 1950. Inh ib i t ion of growth in
maize embryos by canavanine and i t s r e v e r s a l . Bot,
Gaz, 112t 52-57.
Ziebur, N.K,, Brink, R.A., Graf, L.H. and Stahmann, M.A. 1950.
The e f fec t of casein hydrolysate on the growth
in v i t r o of immature Hordeum embryos, Amer, Jour.
Bot. S2» 144-148.
Ziebur, N.K. and Brink, H.A, 1951, The s t imula t ive ef fec t
of Hordeum endosperms on the growth of immature plant
embryos ^ v i t r o . Amer, Jour . Bot. ^ t 253-256.

PLAN OF WORK
The d i f fe ren t developmental stages of flowers and
f r u i t s of SPlanum Integr l fol ium P o i r . , Soj afium a ffrg flt l Hm
Dun, and Solanum hlspldum Pers , have been col lec ted from
the garden of the department of Botany for de t a i l ed
embryological i n v e s t i g a t i o n s . The mater ia ls have been
fixed in F.A.A., dehydrated in a lcohol-xylol s e r i e s ,
embedded in paraff in wax and sect ions have been cut a t
10-12/U, The preparaticais have been stained with the
safranin and fas t green combination.
The ovules and embryos of Solanum khasianum Clarke
and cjPlanum in tegr i fo l lum Poi r , w i l l be cul tured to find
out the i r n u t r i t i o n a l requirements and fac tors affect ing
t he i r growth i a v i t r o .


















