
The Ecology of Rubble Structure of the South Atlantic ... · Cover: Fish (top leflj, anemnes (top...
79
Transcript of The Ecology of Rubble Structure of the South Atlantic ... · Cover: Fish (top leflj, anemnes (top...


Cover: Fish (top leflj, anemnes (top center), and seaweeds (top right) are common organisms associated with rubble siklctures such as the jetties at Ani~rrells inlet, South Carolina (bottom photograph).

Bio log ica l Report 85 (7 .20) September 1988
THE ECOLOGY OF RUBBLE STRUCTURES OF THE SOUTH ATLANTIC BIGHT: A COMMUNITY PROFILE
Mark E . Hay Univers i ty o f North Carol ina a t Chapel H i l l
I n s t i t u t e o f Marine Sciences Morehead City, NC 28557
and
John P. Sutherland Duke Univers i ty
Marine Laboratory Beaufort, NC 28516
Pro jec t Of f i ce r
Mary C. Watzin National Wet1 ands Research Center
U.S. Fish and Wi ld l i f e Serv ice 1010 Gause Boulevard Slidell, LA 70458
Performed f o r
U.S. Department of the I n t e r i o r Fish and Wildlife Service
National Wet1 ands Research Cent e r Washington, DC 20245

DISCLA! MER
The op in ions , f i n d i n g s , conc lus ions , o r recommendations expressed i n t h i s r e p o r t o r p roduc t a re those o f t h e au thors and do n o t n e c e s s a r i l y r e f l e c t t h e t h e views o f Research and Development, F i s h and W i l d l i f e Serv ice, o r t he U.S. Department o f t h e I n t e r i o r . The ment ion o f t r a d e names o r commercial p roduc ts does n o t c o n s t i t u t e endorsement o r a recommendation f o r use by t h e Federal Government.
Library of Congress Cataloging-in-Publication Data
Hay, Mark f. The ecology of rubble structures of the South Atlantic
bight.
(Biological report ; 85-7.20) "Performed for U.S. Department of the Interior, Fish
and Wildlife Service, National Wetlands Research Center." Bibliography: p. Supt. of Docs. no.: !49.89/2:85(7.20)
1. Marine ecology--South Atlantic Bight. 2. Biotic wmmraities--South Atlantic Bight. I. Sutherland, John P. I t . Watzin, Mary C. ill. National Wetlands Research Center (U.S.) 1V. Series: Biological report (Washinglon, i3.G.) ; 85-7.20. V. Title.
Suggested c i t a t i o n :
Hay, M.E,, and J.P. Suther land. 1988. The ecology o f r u b b l e s t r u c t u r e s o f t h e South At lan t i c B i g h t : a community profi le . U S , F i s h Wildl. Serv. B i o l . Rep. 85(7.20). 67 PC.

PREFACE
This community profi 1 e provides an introduction t o the ecology of communities on rubble structures in the South Atlantic Bight (Cape Hatteras, North Carolina, t o Cape Canaveral , F l o r i d a ) . The most prominent rubble s t ruc tures i n t h i s area are j e t t i e s b u i l t a t the entrances t o major harbors. We concentrate much of o u r discussion on these types of s t ruc tu res s ince most of the avai lable l i t e r a t u r e concerns j e t t i e s or biological communities s i m i l a r in species composi t ion t o those tha t occur on j e t t i e s . However, we a lso discuss the ecology of natural hard- subs t ra t e hab i t a t s in general and how these compare with the communities t h a t develop on rubble s t ruc tures . I t i s our hope t h a t t h i s t e x t w i l l s e r v e as a general, yet thorough, review of why such s t r u c t u r e s a r e b u i l t , t h e i r g e n e r a l e f f e c t s on near shore sediment dynamics, and what forces affect the organisms that l i v e in c lose associat ion with these structures.
After an i n i t i a l discussion of the d i f f e r e n t t y p e s of rubble s t ruc tu res (Chapter 1) and the physical factors that affect the organi sms associated with them (Chapter 2), we devote a major portion of our t e x t t o t h e ecology of r u b b l e - s t ruc tu re habi ta ts . In Chapter 3 , we
d e s c r i b e t h e community compos i t ion , distribution, seasonal i ty, and recruitment pa t terns of the major types of organisms found on rubble s t ruc tu res (p1 ankton, seaweeds, i n v e r t e b r a t e s , f i shes , and b i r d s ) . We a1 so d e s c r i b e t h e major species within most of these groups a n d review some aspects of their basic natural history. In Chapter 4 , we discuss the major physical and biological f ac to r s a f fec t ing the organization of intertidal communities, sun1 i t subtidal communities, and shaded subtidal communities. We also evaluate the potential effects of complex, and o f t e n i n d i r e c t , i n t e r a c t i o n s in s t r u c t u r i n g t h e s e communit ies . The effects of rubble structures on shoreline evol ution and engineering are considered i n t h e f i n a l c h a p t e r (Chapter 5 ) on management considerat ions.
Questions or comments concerning th i s publicat ion o r others in the p r o f i l e series should be directed to:
Information Transfer Specialist National Wet1 ands Research Center U.S. Fish and Wildlife Service lOlO Gause Boulevard Stidel l , LA 70458

CONVERSION FACTORS
Multiply millimeters (mm) centimeters (cm) meters (m) meters (m) kilometers (km) kilometers (km)
square meters (m2f square kilometers (km2) hedares (ha)
liters (I) cubic meters (m3) cubic meters (m3)
milligrams (mg) grams (g) kilograms (kg) metric tons (!) metric tons (1)
k~localories (kcal) Colsiiis degrees (OC)
tnches ~nches feet (ft) fathoms statute miles ( m ~ ) nautical miles (nmi)
square feet (!t") square m~les (mi'.) acres
gallons (gal) cubic ieet (ft3) acre-feet
ounces (02) ounces (07) pounds jib) pounds (ib) short tons iton)
Rl~t!sh thermal units (6f i f j Fahrenheit degrees ("F)
Metric to U.S. Customary
U.S. Customary to Metric
To Obtain inches inches feet fathom:; statiito miles nautical miles
sqiiare feet square miles acres
gallons c i~b ic feet acre-feet
ounces ounces pourids pounds short tons
British thermal units Fahrenheit degrees
millimeters centimeters meters meters kilometers kliometers
square meters square kilometers hectares
liters cubic rnaters cubic meters
mi!!!grams grams kilograms metric 'tons metric tons
kiiocalories i;eis;iis deg:ees

CONTENTS
PREFACE ..........................s........................... iii CONVERSION FACTORS ................................................ i v FIGURES ........................................................... v i
............................................................ TABLES v i i i ................................................... ACKNOWLEDGMENTS i x
CHAPTER 1 . INTRODUCTION ........................................... 1
1.1 D e f i n i t i o n o f Rubb le S t r u c t u r e s ......................... 1 1 .2 Rubb le S t r u c t u r e s o f t h e South A t l a n t i c B i g h t ........... 2
CHAPTER 2 . PHYSICAL ENVIRONMENT .................................. 5
2 . 1 Temperature and S a l i n i t y ................................ 5 2.3 l i g h t and T u r b i d i t y ..................................... 5 2 .3 N u t r i e n t s .............................................a. 5 2.4 C u r r e n t s ................................................ 6 2. 5 T i d e s and Waves ....................................... 7 2.6 Sediments ............................................... 7
CHAPTER 3 . COMMUNITY DESCRIPTION ................................. 8
3 . 1 P l a n k t o n ................................................ 8 3.2 Seaweeds ................................................ 8 ........................................... 3.3 I n v e r t e b r a t e 5 15 3.4 F i s h e s .................................................. 2 1 3 .5 B i r d s .................................................. 32
CHAPTER 4 . ECOLOGICAL PATTERNS AND PROCESSES ..................... 36
.................................... 4 . 1 Conceptua l Framework 36 4.2 O r g a n i z a t i o n o f I n t e r t i d a l C o m u n i t i e s .................. 36 4 .3 O r g a n i z a t i o n o f S u n l i t . S u b t i d a l Communit ies ............ 38
............ 4 .4 O r g a n i z d t i o n o f Shaded. S1~bt.j da l Coniinuni t i e s 44 ................................ 4.5 Complex I n t e r a c t i o n s .... 45
..............,...*.. .. CHAPTER 5 . MANAGEMENT CONSJ DERATIONS .. .... 47
.................... ............... 5 . 1 Shore1 i ne Evo l u t i on .. 47 ................................... 5.2 S h o r e l i n e E n g i n e e r i n g 49
5.3 E f f e c t s o f J e t t i e s on Nearby Ben th i c C o ~ i ~ ~ i r i i t i e s ........ 51 5 . 4 J e t t i e s as F i s h i n g S i t e s ................................ 51
................................. 5.5 J e t t i e s as D i v i n g S i t e s 52
REFERENCES ...............e...................a...b.es..........*. 53 ..

FIGURES
Number Paqe
1 J e t t i e s a t Murre l 1 s I n l e t . SC ............................... 2
2 Location of major j e t t i e s along t h e South Atlant ic Bight .... 3
3 How irradiance changes with depth a t a turbid inshore v e r s u s a l e s s t u r b i d o f f s h o r e l o c a t i o n ...................... 6
4 Mean t i de level and mean t ida l range a t various l o c a t i o n s throughout t h e South A t l a n t i c Bight ............... 7
5 Vertical d i s t r ibu t ion of s e s s i l e organisms common on j e t t i e s .................................................. '3
6 U1 va and E n t e r o m o ~ @ ~ p l a n t s ................................ 10
7 Hypnea musciformi s a lone and a t t a c h e d t o Sagas sum ........... 10
..................................... 8 Common s u b t i d a l s eaweeds 12
............................. 9 Common i n t e r t i d a l i n v e r t e b r a t e s 15
10 Common m o b i l e i n v e r t e b r a t e s ................................. 16
11 Common s p o n g e s .............................................. 17
12 Common c o e l e n t e r a t e s ........................................ 18
13 Common b r y o z o a n s ............................................ 19
14 Common t u n i c a t e s ........................................... 19
....... 15 Temporal p a t t e r n s of ben th i c i n v e r t e b r a t e recruitment 21
.......... 16 G i l l ne t de t e rmina t i ons of s e a s o n a l f i s h abundance 26
17 Visual de t e rmina t i ons of s e a s o n a l f i s h abundance ............ 26
18 Unmodified crab t r ap determinat ions of seasonal f i sh a b u n d a n c e ................................................... 26
19 Hrid-if ied crab t r ap determinations o f seasonal f i sh a b u n d a n c e ................................................... 26
.......... 20 Rotenone de t e rmina t i ons of s e a s o n a l f i sh abundance 26

Number Pase
21 S e a s o n a l i t y o f common f i s h e s caught in g i f 1 n e t s ............. 2 7
Seasonal i t y of common f i s h e s observed by d i v e r s ..............
Seasonal i ty of common f j shes caught in unmodified c r a b t r a p s ................................... .... ........... Seasonality of common f ishes caught !n modified crab t raps ... Seasonal i t y of common f i shes taken in rotenone co l lec t ions ...
A group of s p o t t a i l p inf i s h feed ing on j e t t y seaweeds ........ Feeding preferences of two f i shes and two small inver tebrates tha t consume seaweeds on j e t t i e s in t h e Sou th At1 a n t i c B i g h t ..................................... Sea - l eve l r i s e dur ing t h e p a s t 17. 000 yea r s ..................
Beach f l a t t e n i n g i n response t o a s torm ...................... The 1 i g h t h o u s e a t Morr i s I s 1 a n d . SC ..........................
J e t t y a t M u r r e l l s I n l e t . SC .................................... Number of boats and bank-anglers using the Murrells Inle t . SC. j e t t i e s d u r i n g d i f f e r e n t s e a s o n s ......................... The types of f i shes wanted by anglers using j e t t i e s versus t h e types of f i s h e s they caught .......................

Number
1 Number of f i sh species collected o r counted during different seasons on the j e t t i e s a t
........................................... M u r r e l 1 s In1 e t , S C 26
2 Foods of f ishes using the j e t t i e s a t Murrells I n l e t , SC, a s determined by g u t con ten t ana lys i s ............. 30
3 Types of birds common on rubble s t ruc tu re s in the S o u t h A t l a n t i c B i g h t ......................................... 35
4 G u t contents of spot tai l pinfish captured on the j e t t y a t Radio I s l a n d , N C .................................... 43

ACKNOWLEDGMENTS
Order of authorship was decided by the flip of a coin.
Sections of this profi 1 e were reviewed by J.E. Duffy, P.F. Hay, S. Ortega, P.E. Renaud, and 5 anonymous reviewers. Illustrations were produced with the help o f S. Ortega, H. Page, V . Page, P . E . Renaud, and S . Taylor. R. Forward provided the data for Figure 3. 0. Pilkey, T. Clayton, and L. Taylor were especially
helpful in providing data for Chapters 1 and 5. Information on invertebrates was provided by W . Nelson, W . Kirby-Smith, and R. Van Dolah. Information on birds was provided by C. Marsh, J . Parnell , and W . Hon. To a1 1 we are grateful.
The funding for this project was provided by the U.S. Army Corps of Engineers, Waterways Experiment Station.

CHAPTER 1. lMYRQDUCTlON
The South A t l a n t i c Bight borders t h e Un i t ed S t a t e s c o a s t 1 i n e from Cape Ha t t e r a s , NC, t o Cape Canaveral , F L . The s h o r e l i n e along t h e Bight i s sandy and cha rac t e r i zed by numerous b a r r i e r i s l ands separa ted by t i d a l in1 e t s . I n t e r spe r sed a l o n g t h i s c o a s t l i n e , e s p e c i a l l y a t i n l e t s , a r e var ious a r t i f i c i a l s t r u c t u r e s cornposed of hard rubble materi a1 s . These rubble s t r u c t u r e s a r e t h e focus of t h i s p r o f i l e .
1.1 DEFINITION OF RUBBLE STRUCTURES
Rubble s t r u c t u r e s a r e mounds o f random-shaped and r andom-p laced s t o n e s pro tec ted with a cover l a y e r of s e l e c t e d s tones o r s p e c i a l l y shaped conc re t e armor u n i t s (Whal in e t a l . 1984). Most commonly they a r e cons t ruc ted of 1 a rge boulders , but they can be b u i l t from a wide v a r i e t y of m a t e r i a l s , inc lud ing s t e e l , conc re t e , p i l i n g s , wood t imbers , and p l a s t i c bags f i l l e d with sand (Whalin e t a l . 1984). Rubble s t r u c t u r e s can be d iv ided i n t o two g e n e r a l c a t e g o r i e s based upon t h e i r pos i t i on r e l a t i v e t o t h e s h o r e l i n e and t h e i r p u r p o s e . The f i r s t c a t e g o r y i n c l udes t h o s e s t r u c t u r e s bui 1 t p e r p e n d i c u l a r t o t h e s h o r e l i n e and d e s i g n e d t o i n t e r r u p t t h e l i t t o r a l t r a n s p o r t o f s e d i m e n t . T h i s c a t e g o r y i n c l u d e s j e t t i e s , w e i r j e t t i e s , and g r o i n s . The second ca tegory of rubble s t r u c t u r e s incl udes t hose b u i l t para? l e l t o t h e s h o r e l i n e and designed t o prevent waves from reaching t h e h igher e l e v a t i o n s of t h e beach. This category inc ludes b r e a k w a t e r s , s e a w a l l s , b u l k h e a d s , and revetments .
S t trlictyres Perpendi ctll ar t o t h e S h o r o l i 12fi
J e t t i e s a r e s t r u c t u r e s used a t i n l e t s t o s t a b i l i z e t h e p o s i t i o n o f t h e
naviga t ion channel , t o s h i e l d ve s se l s from wave f o r c e s , and t o con t ro l t h e movement of sand along t h e ad j acen t beaches so a s t o minimize t h e movement of sand i n t o t h e channel .
Weir j e t t i e s a r e u p d r i f t j e t t i e s wi th -. a low s e c t i o n o r wei r . L i t t o r a l d r i f t moves o v e r t h e w e i r s e c t i o n i n t o a p r e d r e d g e d d e p o s i t i o n b a s i n which i s dredged pe r iod i ca l 1 y .
G r o i n s a r e s h o r e p r o t e c t i o n s t r u c t u r e s b u i l t t o t r a p l i t t o r a l d r i f t o r r e t a r d e ro s ion of t h e sho re . They a r e u sua l l y s h o r t e r than j e t t i e s and a r e used along t h e beach away from i n l e t s .
S t r u c t u r e s P a r a l l e l t o t h e Shore l ine
Breakwaters a r e wave energy b a r r i e r s designed t o p r o t e c t any landform o r water a r ea behind them from t h e d i r e c t a s s a u l t of waves.
S e a w a l l s a r e s t r u c t u r e s s e p a r a t i n g land and water a r ea s , p r imar i l y designed t o prevent e ro s ion and o t h e r damage due t o wave a c t i o n . Seawalls a r e desdgrred t o r ece ive t h e impact of t h e sea a t l e a s t once dur ing each t i d a l c y c l e .
Bul kheads a r e s t r u c t u r e s bu i 1 t h igher on t h e shore than a seawall o r a revetment t o r e t a i n o r prevent s l i d i n g of t he l and . A secondary purpose i s t o p r o t e c t t h e upland a g a i n s t damage from wave a c t i o n dur ing s torms.
Revetments a r e f a c i n g s o f s t o n e , conc re t e o r wood b u i l t t o p r o t e c t a s ca rp , embankment, o r s h o r e s t r u c t u r e a g a i n s t e r o s i o n by wave a c t i o n Car c u r r e v t s . Revetments & r e a p ro i ec t I ve armor, ra t$er tban a r e t a a n ~ n g s t r u c t u r e .

1.2 RUBBLE STRUCTURES OF THE SOUTH ATLANTIC BIGHT
A1 though many small rubble s t r u c t u r e s e x i s t throughout t h e South At1 a n t i c b i g h t , t he most prominent rubble s t r u c t u r e s i n t h i s a rea a r e t h e j e t t i e s cons t ruc t ed t o p r o t e c t t h e e n t r a n c e s t o t h e r e g i o n ' s major harbors (F igures 1 and 2 ) . Some of t h e l a r g e s t j e t t i e s a r e descr ibed below.
Beaufort , North Carol i na
There a r e two s m a l l j e t t i e s n e a r Beaufort I n l e t . Radio I s land j e t t y was b u i l t p r i o r t o 1939 (C.G. Bookhout, Duke Univers i ty Marine Laboratory; pe r s . comm. ) t o prevent t h e shoal i ng o f Bul khead Channel l ead ing t o Beaufort Harbor. Early surveys (U.S. Coast Survey Chart No. 874, 1874; U.S. Coast and Geodet ic Survey No. 3387,
1913) suggest t h a t Shackleford j e t t y was cons t ruc ted near t h e turn o f t h e cen tury i n an e a r l y at tempt t o s t a b i l i z e Beaufort I n l e t . Nei ther of t h e s e j e t t i e s i s c u r r e n t l y more than 300 m in l eng th . Although sma l l , t h e i r l o c a t i o n near t h e Duke Univers i ty Marine Laboratory and t h e I n s t i t u t e of Marine Sciences of t h e Univers i ty of North Caro l ina a t Chapel Hi l l has made them among t h e be s t s tud ied j e t t i e s i n t he South A t l a n t i c Bight . For t h i s reason they a r e included here .
Masonboro I n l e t , North Carol i na
This i n l e t i s between Wr igh t sv i l l e Beach t o t h e nor th and Masonboro I s l and t o t h e s o u t h . The n o r t h j e t t y o f f Wr igh t sv i l l e Beach i s 1,140 rn long and was cons t ruc t ed in 1965- 1966 (Kies l i ch 1981). The continued t r a n s p o r t of sand i n t o t h e
Figure 1. The jetties at Murrells Inlet, SC, in various stages of construction (A and B) and completed (C). (D) shows a close-up of the large boulders used to form oceanic jetties (photos courtesy of U.S. Army Corps of Engineers)-

--%
1 Winyah Bay. A photograph in t he o f f i c e of ,A Senator b i l l Doar, Georgetown, SC, shows
them under cons t ruc t ion in 1898. The north j e t t y o f f North Is land i s some 1,938 m in l eng th , while t he south j e t t y o f f Sand
1 - ~ ~ ~ ~ ~ ~ ~ ~ ~ I w E T Is1 and i s 4,060 m long. LITTLE R I V E Q I N L E T
M URRELLS 'NLE T
c H ~ R L E S T O N
SA\/fiWN4H R l V E F i
- S T JOHNS RIVER
P O N C E DE L E O N INLET
--- Cope Conovarol
Figure 2. Major jetties in t h e S o u t h Atlantic Bight.
channel from t h e south neces s i t a t ed t h e c o n s t r u c t i o n of a s o u t h e r 0 j e t t y o f f Masonboro Beach in 1979-1980. The southern J p t t y i r 560 m 1 0 ~ g (Whal i n e t a l . 1984).
L i t t l e R i v e r Inlet. , between Nor th and South Carol i na
Two j e t t i e s , both approximately 3,090 m in length , were cons t ruc ted between 1981 and 1983 (Hansen and Ward 1985). They a r e designed t o p ro t ec t t h e channel between Bird Is land t o t h e north ar~d Waites Is land t o t h e south .
Murrell s In1 e t , South Carol ina
The north j e t t y extends some 1,040 m o f f Garden City Beach and was conr t ruc ted between 1977 and 1979 (Van Dolah e t a ? . 1984). Ttie south j e t t y , o f f Huntington Beach, i s approximately t h e save length and was cons t ruc ted between 1979 and 1990.
pinyah Bay, Soutfi Caroiji
Two j e t t i e s were cons t ruc ted around the turn of t he century a t t h e en t rance t o
Char les ton , South Carol ina
Construct ion on two j e t t i e s , 4,060 t o 4,689 R in l enq th , was completed in 1895 (Neal e t a l . 1984). Since t h a t t ime S u l l ' i v a n s I s l a n d Lo t h e n o r t h h a s experienced a ne t accumulation of sand , w h i l e Mor r i s i s l a n d t o t h e s o u t h has suf fered severe e ros ion .
Savannah River , GeorqB --
There a r e two j e t t i e s c o n s t r u c t e d between 1890 and 1898 t h a t p ro t ec t t h e Savannah River Harbor ( G r i f f i n and Henry 1 9 8 2 ) . These j e t t i e s ex t end seaward approximately 3,658 m and have in t e r rup ted l i t t o r a l t r a n s p o r t of sand t o t h e sou th . This and continued dredging of t h e channel have r e s u l t e d i n cons iderable e ros ion on Tybee I s l a n d , l o c a t e d s o u t h o f t h e Savannah River .
S t . Marys Entrancet-between Georsi a and Flor ida --
Thp e n t r a ~ c e i s bordered on t h e nor th by Cumberland Is land and on t h e south by Amelia I s land . Work began on t h e two j e t t i e s i n 1881 and continued u n t i l they r eached t h e i r p r e s e n t form i n 1927 (Parchure 1982). The north j e t t y i s 5,980 m long and t h e south j e t t y i s 3 ,500 m i n l ength . Since t h e i r cons t ruc t ion , sana has accumulated on both t h e north and south s i d e s of t h e en t r ance .
S t . ,lohns--Rjver, F lo r ida
The i n i t i a l j e t t i e s were cons t ruc ted between 1880 and 1895. The northern j e t t y extended 3,500 m seaward froni Fort George I s l and . The southern j e t t y extended from Guano I s l and and was '2,659 m lor:g, Rork continued pe r iod i ca l ly u n t i l 1951 when t h e j e t t i e s a t t a i n e d l e n g t h s o f 4 , 4 3 0 and 3 , 4 9 0 rn, r e s p e c t i v e l y ( P j ? keji f t t a a ? , I 'jgGj. TI;? j s t t j s s have i n t e y f c r e d ~ " t ! : t h e southward t r a n s p o r t o f sand ar~d h a v e caused severe e ros ion t o t h e south o f t he j n l e t .

Ponce de Leon I n l e t , F7!or-ida
The i n l e t l i e s between b a r r i e r i s l a n d s on vihich a r e loca ted Daytona Beach t o t h e nor th and New Smyrna Beach t o t h e south . Two j e t t i e s , approximately 1 , 2 5 0 m long, were b u i l t between 1968 and 1972 ( J o n e s and Mehta 1 9 7 8 ) . T h e i r cons t ruc t i on has s t a b i l i z e d t h e i n l e t , but has apparen t ly i n t e r r u p t e d t h e northward movement of sediment. Beaches t o t h e nor th have eroded while those t o t h e south have experienced considerabl e a c c r e t i on.
Natural System C o u n t e r ~ a r t s
There a r e few na tu ra l coun te rpa r t s t o t h e hard s u b s t r a t e provided by open ocean j e t t i e s i n t h e South A t l a n t i c B i g h t . However, low re1 i e f rocky outcrops a r e found in t h e nearshore zone a t a small a r ea nor th of Cape Fear in North Caro l ina and along t h e northerr , ha l f of t h e South Caro l ina c o a s t . Rock outcrops a l s o occur n e a r Mar ine l and i n n o r t h e r n F l o r i d a
(S tephenson and Stephenson 1972; Sua r l e s 1984) . In s h e l t e r e d wa te r s , f l o r a and f a u n a s i m i l a r t o t h o s e o f a r t i f i c i a l s t r u c t i j r e s a r e found on submerged v e g e t a t i o n (Thayer e t a l . 1984; Keoayh and Chernof f 1987), o y s t e r r e e f s (We1 1 s 1961 ; Dame 1979) and shelf r ubb l e .
O t h e r Ar t i f i c i a1 S t r u c t u r e s
S imi l a r organisms a r e a1 so found on a l m o s t any hard s u b s t r a t e placed in t h e w a t e r , inc lud ing pi1 ings , docks, boa t s , a n d r e f u s e such a s c a n s and b o t t l e s . I ndeed , t h e ep i fauna l foul ing community i s renowned f o r t h e t r o u b l e i t causes when g r o w i n g on human made s t r u c t u r e s , p a r t i c u l a r l y boa t s . Much of what we know a b o u t t h e i n v e r t e b r a t e community comes f r o m s t u d i e s conduc t ed on a r t i f i c i a l s e t t l i n g p l a t e s ( e . g . , S u t h e r l a n d and Kar l son 1977). There i s no doubt t h a t t h e a c t i v i t i e s of people i n t h e South A t l a n t i c B i g h t have increased t h e h a b i t a t space f o r t h e s e ep i ben th ic organisms.

CHAPTER 2. PHYSICAL EMVIROMIMENT
T h i s c h a p t e r c h a r a c t e r i z e s t h e physical environment of t h e South A t l a n t i c B i g h t . The p h y s i c a l v a r i a b l e s most important t o rubble communities i nc lude t h e temperature and s a l i n i t y o f t h e water , and t h e amount of a v a i l a b l e l i g h t and n u t r i e n t s . Water movement i n t h e form of c u r r e n t s , t i d e s , and waves i s a l s o important , p a r t i c u l a r l y because of t h e way i t a f f e c t s sediment t r a n s p o r t a long t h e beach. In t h i s chap t e r we cons ide r each of t h e s e physical v a r i a b l e s in t u rn .
2.1 TEMPERATURE AND SALlNlTY
The American A t l a n t i c Tempera te Region e x t e n d s from Cape Cod, M A , t o s o u t h e r n F l o r i d a (Gosner 1 9 7 9 ) " Cape H a t t e r a s i s a n a t u r a l b i o g e o g r a p h i c boundary along t h e e a s t c o a s t d i v i d i n g t h i s region i n t o t h e Vi rg in ian province i n t h e nor th and t h e Caro l in ian province i n t h e south . The southern boundary of t h e C a r o l i n i a n p r o v i n c e i s Cape C a n a v e r a l . Thus, t h e Carol i n i an province co inc ides with t h e a r e a t r e a t e d i n t h i s p r o f i l e .
Cape Ha t t e r a s and Cape Canaveraf mark s i g n i f i c a n t temperature t r a n s i t i o n zones. In t h e nor thern po r t i on of t h e Ca ro l i n i an p r o v i n c e ( N o r t h Caro l i n a ) w a t e r t empera tures can exceed 30 O C i n summer and drop t o 0 'C i n Winter (Suther land and K a r l s o n 1 9 7 7 ; W . K i rby -Smi th , Duke U n i v e r s i t y Mar ine L a b o r a t o r y ; p e r s . comm. ) . As 1 a t i tude decreases , w in t e r t e m p e r a t u r e s e s p e c i a l l y a r e g r a d u a l l y amel i o r a t e d . In c e n t r a l F lo r ida , water tem e r a t u r e s range from 30 '6 in summer t o g 14 C i n w in t e r (Mook 1980).
Ir! t h i s p r o f i l e we r e s t r i c t o u r a t t e n t i o n t o t h e o u t e r c o a s t and t o t h e sounds and e s t u a r i e s where t h e s a l i n i t y g e n e r a l l y remains above 20 p p t . The f l o r a
and fauna of t h e s e reg ions a r e b a s i c a l l y marine.
2.2 LIGHT AND TURBlDlTY
In t h e South A t l a n t i c Bight , t i d e s , waves, and wind-generated tu rbulence r e - suspend bottom sediments i n t he shal low waters on t h e Continental She l f . This sediment resuspens ion , combined wi th high e s t u a r i n e and n e a r s h o r e p h y t o p l a n k t o n p r o d u c t i v i t y , p r o d u c e s t u r b i d i n s h o r e w a t e r s t h a t d r a s t i c a l l y r e d u c e f i g h t p e n e t r a t i o n . Reduced l i g h t pene t r a t i on appears t o be t h e major f a c t o r r e s t r i c t i n g p l a n t growth t o t he shal low po r t i ons o f most j e t t i e s . F i g u r e 3 shows t h e r e l a t i o n s h i p between i r r a d i a n c e and wave l eng th t a k e n on t h e same d a y f o r d i f f e r e n t dep ths a t an inshore s i t e i n Bogue Sound, NC ( t h e t u rn ing bas in a t Morehead Ci t y j , and a s i t e approximately 6 km o f f s h o r e from Bogue Sound. As can be seen , 7 i g h t decreases dramatical 1 y wi th depth in nearshore waters , where rubble s t r u c t u r e s a r e g e n e r a l l y l oca t ed .
2.3 NUTRIENTS
N u t r i e n t c o n d i t i o n s s u r r o u n d i n g j e t t i e s a r e important because they a f f e c t t h e growth of t h e seaweeds a t t a ched t o the j e t t i e s and t h e growth of phytoplankton in t h e over1 y ing waters . These phytopf ankton a r e consumed by benth ic f i l t e r f e e d e r s on t h e j e t t i e s . On j e t t i e s s u b j e c t t o s t rong wave o r t i d a l a c t i o n , p l a n t s may be m i n i m a l l y a f f e c t e d by 7 ow n u t r i e n t s because new water i s c o n s t a n t l y flowing by and b e c a u s e t u r b u l e n c e i n t e r r u p t s t h e formation of d i f f u s i o n b a r r i e r s around the seaweed t h a l l u s , N i t r o g e n i s most

INSHORE SITE, BOGUE SOUNC, N.C. 1 0 ' ~ . - 1 @ " ' hrry-T
6 krn SEAWARD OF BOGUE SOUND, N.C. r ' ' ' i 2 ' ~ ' ' ~
Wavelength (nm )
Figure 3. lrradiance versus wavelength for an inshore site (left) in Bogue Sound, NC, and for a site 6 krn offshore (right). The numbers on each line indicate depth in meters. (Data provided by R. Forward, Duke University Marine Laboratory.)
commonly t h e n u t r i e n t l i m i t i n g p l a n t growth in c o a s t a l wa t e r s . In summers, t h e a v a i l a b i l i t y o f d i s s o l v e d i n o r g a n i c n i t r ogen may change by more than an o rde r of magnitude on a d ie1 c y c l e , r i s i n g a t rrjght and f a l l i n g d r ama t i ca l l y dur ing t h e day as i t i s consumed d u r i n g p h o t o s y n t h e s i s ( i i t a k e r e t a l . 1 9 8 7 ) . Highly product ive seaweeds l i k e t h e s e a l e t t u c e , w a , can t ake advantage of t h e s e n i t r o g e n s p i k e s by r a p i d l y s t o r i n g n i t r o g e n f o r ' i a t e r u s e . O t h e r more macsive and s lower growing seaweeds 1 i ke Codjum do not have t h i s a b i l i t y (Ramus and Venable 1987), b u t may ga in add! t i o n a l n i t r o g e n by e s t a b l i s h i n g s y m b i o t i c a s s o c i a t i a n s with n i t r ogen f i x i n g blue- green a lgae (Rosenberg and Paerl $980).
2.4 CURRENTS
The most important o f f s h o r e c u r r e n t i n t h e region i s t h e F lo r ida Curren t , which o r i g i n a t e s i n t h e F l o r i d a S t r a i t s between F lo r ida and Cuba. I t i s joined by t h e north flowing A n t i l l e s Cur r en t , which r u n s a l o n g t h e o u t e r edge o f t h e Continental She1 f . The F lo r ida Curren t moves o f f sho re a t Cape Ha t t e r a s t o become t h e Gulf Stream, al though t h i s l a t t e r name i s o f t en appl ied t o t h e F lo r ida Curren t a s we1 I . The p o s i t i o n of t h e F lo r ida Curren t v a r i e s s ea sona l ly . In t h e summer i t moves inshore , b r ing ing warm, c l e a r water t o t h e s h e l f . In t h e w i n t e r i t i s d r i v e n o f f sho re by n o r t h e r l y winds. Whe~ t h i s happens co ld water from t h e nor th may move

Tidal Height (m) 0 A h)
L I
Cope Hotteros +------@A
Caps Lookout I------@-
Cape Feor ------- @ ------.
Charleston p@------'
Savannah I @ +
Jacksonville +-----@-
Cepe Conavorol c------e--------c
3 -4.3 Eu 2 X 0 i. " 3 m - c w o - 4 3cn3S
m u i m 0 v, -37,. r+ $ 3 7,
W m l D 0 rc
3 500 ;;: A . r z 2
O Q W 0 L O W r+ I - J . W " c u Cuz .-J. w-tl-5 072-a " "-3 -5 " E" r+ 0 J. 4. m -5 Z E , ( D c D a , '5 a,
3 -1 ?LC r-z lu < m r + z o w 3 m
-1. ,Q<a e m c n - Imm, r - - 2 2 o (D -0 W U 3 c 3-2 d*
2 (D -,-,+Z.r* y , , " * 3 S
3 zIfe w a z w g ; -43 -10
-5 z.":g' ;,-,. V) <
3 r t g rt m 2. &. A. go , , aJ3 2 A.
W ( D < + ' 3
4 . v - 3-" ,-", 0 w -h
3 * 2 r + = 3 2 -0 IT a m
o w m r + m ;f& -Amg;q
0 a J. f l a f i n S P
s 32 S2-d r + m =r, " > E k S (Do r c m b 4 m -5 I A d d . 3 d. * 3 Cf. m$kf:;;'OI O- 2 - ,,rcE", r c c-t T.$Lw ..a- d. 0 ,p., c-7 -5 g,
0 -4.za 3 KJ 3
.I m a
-5 2 a O m 0- rtA.-hEuQ. _.; ZD3 =lcD
2 Z,1 ,3.2,, ( D O 0 -5w
cu T T - ' . W , 3 V r + O 0 --'m ~ N S I m - ,

CHAPTER 3. COMMUNITY DESCRiPTlON
Rubbl e - s t r u c t u r e communit ies c o n s i s t I n w i n t e r , d i e l changes i n phy top l ank ton o f , and i n t e r a c t w i t h , a w ide v a r i e t y o f abundance a r e s m a l l . I n summer, an d i f f e r e n t f l o r a and f a u n a i n c l u d i n g : ou tgo i ng t i d e i n l a t e a f t e rnoon can have p lank ton , seaweeds, i n v e r t e b r a t e s , f i s h e s , t w i c e t h e abundance o f phy top l ank ton as an and b i r d s . T h i s chap te r desc r i bes t h e ou tgo i ng t i d e i n e a r l y morn ing ( L i t a k e r e t mos t a p p a r e n t g r o u p s o f o r g a n i s m s a l . 1987). a s s o c i a t e d w i t h r u b b l e s t r u c t u r e s and, whe re a p p r o p r i a t e , d i s c u s s e s p r o m i n e n t , Zooplankton communit ies a r e composed spec ies and t h e i r p a t t e r n s o f r e c r i l i t m e n t o f permanent zoop lank te rs ( ho l op l ank ton ) , and d i s t r i b u t i o n . F e e d i n g h a b i t s and such as copepods, and o f t h e l a r v a e o f b a s i c n a t u r a l h i s t o r y a re a1 so t r e a t e d b e n t h i c o r g a n i sms (me rop l a n k t o n ) , here. D e t a i l e d aspects o f t h e eco logy o f i n c l u d i n g t h o s e on r u b b l e s t r u c t u r e s . t h e s e o r g a n i s m s and c o m m u n i t i e s a r e There a r e l a r g e f l u c t u a t i o n s i n d e n s i t y d i s c u s s e d a t l e n g t h i n t h e f o l l o w i n g and spec ies compos i t i on o f h o l o p l a n k t o n i c c h a p t e r o n e c o l o g i c a l p a t t e r n s and organisms. These a re due t o seasonal and processes. d i e 1 changes i n temperature, and p r e d a t i o n
by f i s h and o t h e r zoop lank te r s (Fu l t o n 1983, 1985 ) . F l u c t u a t i o n s i n t h e
3.1 PLANKTON abundance o f l a r v a e f rom t h e benthos c o u l d be a f f e c t e d by t h e same f a c t o r s b u t w i l l
B o t h p h y t o p l a n k t o n and z o o p l a n k t o n a l s o be s i g n i f i c a n t l y a f f e c t e d by t h e s e r v e as i m p o r t a n t f o o d s f o r b e n t h i c t i m i n g o f l a r v a l r e l ease . f i 1 t e r f e e d e r s and f o r some j u v e n i 1 e f i s h e s on j e t t i e s . The a v a i l a b i l i t y of these foods may change on t i m e sca l es o f 3.2 SEAWEEDS h o u r s , days , o r seasons ( H a r r i s 1980; L i t a k e r e t a1. 1987). Th i s i s e s p e c i a l l y Community Com~os i t i o n t r u e on i n l e t j e t t i e s , which a re a f f e c t e d by oceanic wa te rs a t h i g h t i d e and by more Most o f t h e South A t l a n t i c B i g h t i s p roduc t i ve , e s t u a r i n e wate rs a t l o w t i d e . an i n h o s p i t a b 1 e h a b i t a t f o r seaweeds There a re a l s o s i g n i f i c a n t seasonal and because o f t h e l a r g e expanses o f d i e l e f f e c t s caused by i n t e r a c t i o n s among unconso l ida ted sands, s i l t s , atid muds t o r a i n f a l l , evapora t ion , t e r r e s t r i a l r u n o f f , w h i c h mos t seaweeds c a n n o t a t t a c h . and d i e l p a t t e r n s i n phy top l ank ton g rowth N a t u r a l i n t e r t i d a l r o c k s a r e r a r e , ( L i t a k e r e t a1. 1987). As an example, i n o c c u r r i n g a t o n l y a few p l aces near t h e w i n t e r t h e e s t u a r i n e wate rs beh ind t h e bo rde r between No r t h and South Ca ro l i na , O u t e r Banks o f N o r t h C a r o l i n a a r e and a t Mar ine land, FL. On t h e Con t i nen ta l dominated by r i v e r i n e i n p u t s because o f S h e l f , t h e r e a r e ou tc rops o f sedimentary h i g h r a i n f a l l and l o w e v a p o r a t i o n . r o c k s t h a t s t a r t j u s t s o u t h o f Cape Grow th - ' l im i t i ng n i t r o g e n i s s u p p l i e d as H a t t e r a s and r u n a l l t h e way t o F l o r i d a . n i t r a t e and ammonium by r u n o f f f r om t h e However , mos t o f t h e s e o u t c r o p s a r e d ra inage bas i n . I n summer, t h e l owe r c o v e r e d by s e d i m e n t and so a r e n o t r a i n f a l l and h i g h e r e v a p o r a t i o n r a t e s a v a i l a b l e f o r a t t a c h m e n t b y seaweeds. cause t h i s area t o f u n c t i o n more 1 i k e a Hard subs t r a t es t h a t a re a v a i l a b l e f o r l agoon . Most n i t r o g e n i s suppl i e d as at tachment occur most abundant ly i n Onslow ammanium due t o b i o l o g i c a l r egene ra t i on . Bay, NC, and on t h e coas t nea r Palm Beach,
8

F L (Sea r l e s 2984). Because na"tur1 hard s u b s t r a t e s a r e rare i n t h e b i g h t , most seaweeds a r e a t tached t o s h e l l f ragments , o t h e r a lgae , s eag ra s se s , o r t o in t roduced s u b s t r a t e s such a s s eawa l l s , j e t t i e s , and docks.
Between Long Is land Sound and Cape H a t t e r a s , t h e r e a r e a p p r o x i m a t e l y 150 s p e c i e s o f r e d (Rhodophy ta ) , brown ( P h a e o p h y t a ) , and g r e e n ( C h l o r o p h y t a ) seaweeds (Sear1 e s 1984). Between Cape Ha t t e r a s and Cape Canaveral , t h e r e a r e approximately 320 s p e c i e s ; 303 o f t h e s e a r e known from North CaroSina ( S e a r l e s 1984). Nine ty- f ive s p e c i e s occur i n South Caro l i n a (Wiseman and S c h n e i d e r 1976 ; Wiseman 1978; B l a i r and Ha17 1981), 81 spec i e s occur i n Georgia (Chapman 1971, 1973; S e a r l e s 1981, 1984), and only 43 a r e r e p o r t e d i n F l o r i d a n o r t h o f Cape Canave ra l (Humm 1 9 5 2 ) . However, H u m m (1952) probably underest imates t h e number of spec i e s in t h e a r ea s i n c e 234 spec i e s o c c u r between Cape Canave ra l and Palm Beach ( K e r r 1976 ; Eiseman 1976, 1979 ; Eiseman and Moe 1981; Eiseman and Norr i s 1981; Hall and Eisemarl 1981). There a r e severa l reasons why North Caro l ina appears t o have 3 t o 4 t imes t h e number of seaweed s p e c i e s a s Sou th C a r o l i n a o r G e o r g i a . These inc lude (1) t h e l o c a t i o n of North Caro l ina i n a t r a n s i t i o n a l zone between t h e temperate seaweeds of New England and t h e t r o p i c a l seaweeds of t h e Caribbean, ( 2 ) t h e g r e a t e r abundance o f h a r d
s u b s t r a t e s o f f t h e North Caro l ina c o a s t , and (3) t h e g r e a t e r number of seaweed s p e c i a l i s t s t h a t have i n v e s t i g a t e d t h e marine f l o r a of North Caro l ina . Because of t h e ex t ens ive f l o r i s t i c i n v e s t i g a t i o n s conducted in North Caro l ina by S e a r l e s , Schneider , and Kapraun (Schneider 1976; S e a r l e s and S c h n e i d e r 1978 , 1980 ; Schneider and S e a r l e s 1979; Kapraun 1980a, b , 2984; Kapraun and Zechman 1982; S e a r l e s 19841, t h e seaweeds of t h i s a r ea a r e much b e t t e r known than t hose i n any o t h e r p a r t o f t he South A t l a n t i c Bight . The pauc i t y of d a t a from o t h e r r eg ions f o r c e s us t o f o c u s most o f o u r d i s c u s s i o n on t h e seaweeds of t h e Caro l inas . For keys and i 1 l u s t r a t i ons of seaweeds of t h e South A t l a n t i c B i g h t , s e e T a y l o r (1960) and Kapraun (1980a, 1984).
The seaweeds growing h ighes t in t h e i n t e r t i d a l zone a r e u s u a l l y b l u e - g r e e n a l g a e t h a t appear a s a da rk ly colored band on t he rocks (F igure 5 ) . The most common seaweeds immediately below t h e bl ue-green zone a r e u sua l l y t h e green a lgae , W, Ente romoraha , and C ladophora , and , a t t imes , t h e red a lga P o r ~ h v r a . and Enterornor~ha (F igure 6 ) a r e b r i g h t g reen seaweeds t h a t o f t en grow intermixed. They a r e d i s t i ngu i shed p r imar i l y on t h e b a s i s o f f rond morphology. has a f l a t membranous f r o n d composed of two c e l l 1 aye r s ; Enteromoraha f ronds a r e simi 1 a r except t h a t they a r e t u b u l a r , a t l e a s t i n p a r t . S ince some spec i e s of Enteromowha
ALGAE BARNACLES MOLLUSCA HYDROZOA ASClDlACEA BRYOZOA
1 ' Line Transects \ /------7 /-\ /7 A
Figure 5. Vertical distribution (L=level with respect to Mean Tide Level) of the 20 mas% abundant sessile species cbsenred at north jetty stations at Mclrretis Inlet, SC. Band width indicates abundance (Van Dolah ef a!. 1984).

The lower i n t e r t i d a l zone i s u sua l ly occupied by a mixed spec i e s group of red seaweeds ( F i g u r e 5 ) . S e v e r a l s m a l l f i lamentous forms such a s Polvsiuhonia, H e r p o s i ~ h o n i a , Audouinel l a , and E r y t h r o t r i c h i a a r e common, but t h e s e a r e d i f f i c u l t t o i d e n t i f y w i t h o u t magni f ica t ion . Hvunea musciformi s i s a very common l a r g e r seaweed with bushy, cyl i n d r i c a l branches t h a t o f t en end in f i sh-hook l i k e t e n d r i l s t h a t a r e used t o a t t a , ch s e c o n d a r i l y t o o t h e r s eaweeds ( F i g u r e 7 ) . Like many o f t h e r e d (Rhodophyta) seaweeds , i t may be d e e p g r e e n , r e d , s t r a w - c o l o r e d , o r some
Figure 6. The common green seaweeds (A) Ulva, and (B) Enreromorpha.
have broad f l a t b l a d e s w i t h t u b u l a r morphology only along t h e margins ( i . e . s u p e r f i c i a l l y l i k e m), some members of t h i s genus a r e d i f f i c u l t o r impossible t o d i s t i n g u i s h f rom Ulva w i t h o u t a microscope. In c u l t u r e , some spec i e s produce tubul a r progeny, suggest! ng t h a t d i s t i n c t i o n s between t h e two genera a r e ques t ionab le . Cladophora i s one of t h e most common q e n e r a o f sma l l g r e e n f i 1 amentous a l g a e . Xt can e a s i iy be Figure 7. (A) The common red seaweed Hypnea confused with a number of o t h e r a l g a e by mrrsciformis. its hook-shaped branch tips often nonspeciaf i s t s , The red a l g a Porohvra i s facilitate itsattachmentto otherseaweeds. (B)shows m - 1 ! ke i n msrphol ogy but i s usual 1 y a close-up of a Hypnea tip that has attached around brown t o purp le i n c o l o r . the base of a Sargassum f loat.

combination of t h e t h r e e . Lomentaria i s a r o se t o red-co lored p l a n t t h a t looks 1 i k e Hypnea but does not have recurved t i p s . I t forms dense ly branched c reep ing mats, has hollow axes except a t t h e bases , and i t s b r anch l e t s tend t o arch back toward t h e main a x i s . G r a c i l a r i a t i k v a h i a e (formerly G . f o l i i f e r a ) i s 01 ive green t o dark red and has f l a t t e n e d , s t r a p - l i k e b l a d e s w i t h p o i n t e d t i p s ( F i g u r e 8A) . Although t h e e n t i r e p l a n t may a p p e a r i r r e g u l a r l y bushy, t h e branches along each main a x i s a l l branch i n t h e plane of t h e bla,de. Rhod~meni-i_a p ~ e j d o o a l m a t a i s r o s e t o red i n - c o l o r and has s t r a p shaped, d i c h o t o m o u s l y b r a n c h i n g bl a d e s w i t h rounded t i p s . I t u s ~ r a l l y occurs a t , o r below, t h e low t i d e l i n e . Small, wiry t u r f s of reddish- -purp le t o brown seaweeds in t h e genus Gelidium a l s o occur i n t h i s same habi tat . .
On some j e t t i e s , g reen , sponge-1 i ke, dichotomously branching a lgae (F igure 8B) in t h e genus Codium (commonly c a l l e d dead man's f i n g e r s ) a r e common a t t h i s low t i d a l l e v e l . In North Caro l ina , j e t t i e s have t r a d i t i o n a l l y supported popula t ions of Codium d e c o r t i c a t u r n and Codium isthmocladium. Within t h e p a s t decade, t h e introduced spec i e s Codium f r a q i l e has invaded down t h e c o a s t from New England and now makes up a s u b s t a n t i a l por t ion of t h e Codium biomass on l o c a l j e t t i e s (Sea r l e s e t a l . 1984; J . Ramus, Duke Univers i ty Marine Laboratory, Beaufort , NC; pe rs . comm.).
V i r t u a l l y a l l o f t h e g e n e r a and spec i e s mentioned above a l s o occur i n t h e shal low sub t ida l zone dur ing some t imes of t h e y e a r o r i n some l o c a t i o n s . The d i s t i n c t i o n between i n t e r t i d a l and sub t ida l seaweeds i n t h e South A t l a n t i c Bight i s no t always c l e a r and appears t o be useful only dur ing some t imes of t h e year (Kapraun and Zechman 1982). However, s e v e r a l j e t t y seaweeds a r e a l m o s t exc lu s ive ly s u b t i d a l .
The most abundant sub t ida l seaweed on j e t t i e s along much of t h e c o a s t i s t h e brown a lga Sarsassum. I t has a wiry main a x i s , l i n e a r l e a v e s w i th m i d r i b s , and s t a l k e d , sphe r i ca l a i r b ladders f Figure 8C). In summer, t h e brown seaweeds Padina and Dictvota a r e a l s o common. Padina forms a fanshaped, l i g h t l y c a l c i f i e d b lade
( F i g u r e 8D) ; D i c t v o t a h a s membranous, dichotomously branched axes t h a t a r e brown t o golden brown in c o l o r (F igure 8E).
S e v e r a l o t h e r seaweeds a r e occas iona l l y common i n t h e sub t ida l zone inc luding: t h e green a l g a B r v o ~ s i s ; t h e r e d a l g a e Chondr i a ( F i g u r e 8 F ) , C a l l i t h a m n i o n , C h a m ~ i a , Dasva , H v ~ o s l ossum, Cal o n i t o p h v l l urn, and G r i n n e l l i a ; and t h e brown a l g a e E c t o c a r ~ u s , P u n c t a r i a , and P e t a l o n i a . Given t h e d i f f i c u l t i e s a s s o c i a t e d w i t h seaweed i d e n t i f i c a t i o n , r eade r s i n t e r e s t e d i n seaweeds should c o n s u l t Taylor (1960) o r Kapraun (1980a, 1984) before a s s ign ing a name t o any seaweed from t h i s c o a s t .
D i s t r i b u t i o n
When r e s e a r c h e r s s t u d y a1 g a l communities on rubble s t r u c t u r e s i n t h e South A t l a n t i c Bight a t a s i n g l e po in t i n t ime (u sua l l y summer), t hey o f t en d e s c r i b e d i s t i n c t p a t t e r n s o f z o n a t i o n based p r imar i l y on t h e upper l i m i t s o f dominant spec i e s (Hoyt 1920; Wi 11 iams 1949; E a r l e and Humm 1964). When Kapraun and Zechman (1982) i n v e s t i g a t e d seasonal p a t t e r n s of v e r t i c a l zonat ion on j e t t i e s a t Masonboro I n l e t , N C , they noted what appeared t o be r e l a t i v e l y d i s t i n c t i n t e r t i d a l and sub t ida l communities i n t h e summer, bu t dur ing t h e remainder of t h e y e a r t h e r e was no c l e a r s epa ra t i on of communities a t t h e low t i d e l i n e . During w in t e r and e a r l y sp r ing , so c a l l e d i n t e r t i d a l spec i e s 1 i ke P o r ~ h v r a c a r o l i n e n s i s and E n t e r o m o r ~ h a pro1 i f e r a became abundant i n t h e s u b t i d a l z o n e , bu t d u r i n g t h e summers, t h e y r e t r e a t e d back t o t h e i n t e r t i d a l . Kapraun and Zechman (1982) hypothesized t h a t t h e s e changes occurred i n response t o changing compet i t ive i n t e r a c t i o n s among t h e p l a n t s . However, t h e s e changes a r e a1 so c o n s i s t e n t w i t h t h e h y p o t h e s i s t h a t f i s h g r a z i n g dur ing t h e warm p o r t i o n s o f the year, when f i s h e s a r e most numerous, exclude t h e s e p a l a t a b l e seaweeds from t h e sub t ida l zone. The 1 imi t ed a v a i l a b l e d a t a sugges t t h a t f i s h e s d r i v e s eve ra l pal a t a b l e seaweeds t o nea r e x t i n c t i o n on s u b t i d a l po r t i ons of j e t t i e s i n t h e summer (Hay 1986) . A d d i t i o n a l l y , r e c e n t work i n o u t d o o r microcosms h a s shown t h a t E ~ t e r o m s r R h a grows yea r round i n t h e sub t ida l zone i f f i s h e s a r e excluded from t h e system (Hay 1 9 8 6 ) . I f f i s h e s a r e a b u n d a n t ,

Figure 8. Some common seaweeds on jetties in the South Atlantic Bight. (A ) Gracllaria tikwahiae, (B) Codjum SP., Sargassurn filipendula, (DJ Padina gymnaspora, ( E ) Dictyota dichotoma, and ( F ) Chondria sp.

Enteromorph3 i s driven local ly ex t inc t .in the subtidal zone or occurs on ly in small refuge areas.
The most extensive s tudies of the spat ia l d i s t r ibu t ions of seaweeds in the bight focus on the d i s t inc t ions between nearshore and offshore species and on how they are dis t r ibuted north-to-south along the U. S . coast l ine (Schneider 1975, 1976; Searles and Schneider 1980; Searles 1984). Of t h e 303 seaweeds known from North Carol ina, approximately two-thi rds occur in shal low coas t a l h a b i t a t s and a r e potential res idents of rubble s t ructures . Approximately one-ha1 f (109 species) of the 204 species found in shallow water are n o t k n o w n t o occur i n deep o f f s h o r e waters. Of the 194 spec.ies tha t do occur in deep o f f s h o r e wate rs , approximately one-half (108) are known only from those depths. About one-third (96 species) of the to ta l f l o r a have been collected in both shallow and deep hab i ta t s .
For the shallow water species, 21 reach t h e i r northern 1 imit and 27 reach t h e i r southern l imi t of d i s t r ibu t ion in North Carol ina. Within North Carol ina, the Cape Lookout j e t t y appears t o be the northernmost 1 imi t in the i n t e r t i da l zone f o r t r o p i c a l and sub t rop i ca l s e a w e d s (Wi 11 i ams 1948; H u m m 1969; Schneider 1976). The Continental Shelf off Cape Lookout p lays a s i m i l a r r o l e f o r t h e offshore seaweeds (Schneider 1976). A few reach t h e southern l i m i t of t h e i r d i s t r ibu t ion off Cape Lookout (only I%), but many (37%) reach t h e i r northern-most l imi t there. This indicates the more tropical a f f i n i t y of the offshore f l o r a . Of t h e 303 seaweeds known from North Car07 ina, 44% reach t h e i r northern 1 imi t s and 10% reach t h e i r southern 1 imits within the S ta te .
In the Carolinas, seasonal changes in t he occur rence , abundance, and reproduct ion of a l ga l spec i e s can be dramatic (Schneider 1975, 1976; Richardson 1979, 1981, 1982; Kapraun and Zechman 1982; Peck01 1982; Peck01 and S e a r l e s 1983; Van Dolah e t a1. 1984). In a study of seaweeds on an i n t e r t i d a l j e t t y , Kapraun and Zechman (1982) noted t ha t the red algae were most diverse in summer, brown algae were most diverse in winter, and green algae were re1 a t ively aseasonal .
They also suggested t h a t t he re were th ree components of the North Carolina algal community they studied: 1 ) a eurythermal cool - temperate element in w in t e r (comprising 37% of the species a t t h e i r s i t e ) ; 2) a eurythermal tropical element in summer (comprising 18% of the species a t t he i r s i t e ) ; and 3) a l a rger , warm- temperate element tha t occurred year round (comprising 45% of the species a t t h e i r s i t e ) . Schneider (1975, 1976) studied the f l o r a of the Continental Shelf and noted maximal development in mid-summer. The number of spec i e s and t o t a l biomass decreased in f a l l , reached a yearly low in winter, and then increased spring through summer. Ninety-nine percen t of a l 1 species were present in mid-summer, 59% were present in the f a l l , and only 33% were present in the winter.
Successional patterns have not been adequately documented fo r shall ow water seaweeds in the bight. Van Dolah e t a l . (1984) documented the summertime abundance of seaweeds on j e t t i e s a t Murrell s I n l e t , SC, every yea r f o r 4 yea r s a f t e r c o n s t r u c t i o n . Between t h e f i r s t and second year there was an increase in the number of a l g a l s p e c i e s and in a l g a l abundance. Changes between the second and fourth years were nondirectional.
Patterns of Recruitment
Seaweeds may r e c r u i t v ia va r i ous types of spores or , in some species, by fragmentation and reattachment of adul t po r t i ons ( D i x o n 1965) . Given t h e seasonally changing nature of the f lo ra of t h e South A t l a n t i c Bigh t , r e i nvas ion fo l lowing seasons of i n h o s p i t a b l e conditions could be a major problem fo r benthic seaweeds. Many seaweeds appear t o have adapted t o t h e s e cond i t i ons by pers is t ing throughout unfavorable periods as stunted forms or pers is tent holdfas ts ; Sarqassum f i l ioendula , Botrvocladia occidental is , and Graci lar ia mammillaris do t h i s i n Cont inental She l f h a b i t a t s (Schneider 1976) . Several spec i e s on shallow j e t t i e s i n the bight produce ea r ly developmental stages tha t are capable of withstanding Song periods of unfavorabf e cond i t i ons . This phenomenon has been studied in greates t de ta i l by Richardson (1978, 1979, 1981, 1982).

On t h e j e t t y a t Radio I s l and , NC, t h e brown seaweeds Dictvota dichotoma, Padina gvmnosuora (formerly P, vi cke r s i a e ) , and D i c t v o u t e r i s membranacea o v e r w i n t e r a s e a r l y deve lopmen ta l s t a g e s ( R i c h a r d s o n 1978). Of t h e s e t h r e e spec i e s , Dic tvota has been s t u d i e d i n g r e a t e s t d e t a i l (Richardson 1979). Dic tvota i s v i s i b l y p re sen t from mid-April t o December and r e l e a s e s propagules continuously dur ing t h i s growing season; i n most i n s t ances , t h e s e spores r ap id ly germinate and grow i n t o mature p l a n t s . Neither spore re1 ea se nor attachment a r e a f f ec t ed by temperature o r photoperi od. However, germinat ion and es tab l i shment a r e temperature dependent. S p o r e s c a n n o t germi n a t e i n w i n t e r . However, i f t h e r e i s an i n i t i a l warm per iod of 5-6 days, t h e spores germinate and t h e r e s u l t i n g spo r l i ngs can su rv ive s e v e r a l months o f w i n t e r c o n d i t i o n s . Thus, microscopic spo r l i ngs produced a t t h e end of t h e growing season, overwinter and a s s u r e t h e c o n t i n u a t i o n o f t h e populat ion when warmer cond i t i ons r e t u r n .
The red a l g a Dasva ba i l l ouv iana i s apparent on t h e j e t t y between February and May ( i . e . , t h e oppos i t e of t h e p a t t e r n shown by Dic tvo ta ) . In Apri l o r May i t reproduces and d i sappea r s . Like Di c t v o t a , i t p e r s i s t s a s a young developmental s t a g e dur ing those t imes of y e a r when i t i s no t ev iden t (Richardson 1981). Each win t e r , a s i n g l e g e n e r a t i o n o f p l a n t s g rows , r e p r o d u c e s , and r e l e a s e s s p o r e s t h a t s e t t l e but do no t develop i n t o v i s i b l e p l a n t s u n t i l t h e fo l lowing growing season. In Cape Cod, D. ba i l l ouv iana p e r s i s t s through winter a s a spore1 ing but produces mu1 t i p 1 e gene ra t ions dur ing i t s growing season (Sears 1971). In t h e t r o p i c s , D. ba i l l ouv iana grows yea r round a s a - v i s i b l e p l a n t (Mathieson and Dawes 1975).
The g r e e n a l g a B r v o u s i s ulumosa e x h i b i t s a p a t t e r n somewhat s imi l a r t o Dasva and Dic tvota . I t is v i s i b l y p re sen t from J a n u a r y u n t i l May, and p e r s i s t s through the summer and f a l l a s a p r o s t r a t e microthal l u s s t a g e (Richardson 1982).
Amsler and S e a r l e s (1980) i n v e s t i g a t e d t h e d i s t r i b u t i o n o f a l g a l spo re s i n a 20 m water column 30 km o f f t h e c o a s t o f North Caro l ina . Spores of green a lgae were d i s t r i b u t e d throughout t h e column and spores of bangiophycean red
a l g a e ( t h e s i m p l e r r e d a l g a e l i k e P o r ~ h v r a ) were present a t a l l depths but concentrated in g r e a t e s t abundance near t h e bot tom. S p o r e s o f brown and f l o r i d e o p h y c e a n r e d a l g a e ( t h e more complex r e d a l g a e 1 i ke Hmnea and Chondri a) occurred almost exc lus ive ly near t h e bottom. Green and bangiophycean red a lgae tended t o be more o p p o r t u n i s t i c than brown and f l o r i deophycean r e d a1 g a e , s u g g e s t i n g t h a t t h i s d i s t r i b u t i o n o f spores i s adapt ive i n t h a t i t a l lows f o r wide d i s p e r s a l o f t h e o p p o r t u n i s t i c spec i e s ( c a r r i e d with t h e su r f ace c u r e n t s ) and keeps t h e l e s s o p p o r t u n i s t i c spec i e s n e a r h a b i t a t s where t h e p a r e n t s were succes s fu l .
E ~ i p h v t i c Alaae
S e v e r a l s p e c i e s o f a l g a e can u s e o t h e r seaweeds a s s u b s t r a t e s f o r attachment (F igure 7 ) . A hos t of small a lgae (diatoms, f i lamentous bl ue-green, red , brown, and green a lgae) and severa l l a r g e r mac rophy te s ( such a s Hvpnea, Soyridi a , Enteromoroha, Chaetomor~ha, and D i c t v o t a ) commonly o c c u r a s e p i p h y t e s . Growing e p i p h y t i c a l l y can p r o v i d e a mechanism f o r circumventing compet i t ive exc lus ion when a l l primary s u b s t r a t e i s occupied (Hay 1981a; Hawkins and Harkin 1 9 8 5 ) . E p i p h y t e s may a l s o a v o i d h e r b i v o r o u s f i s h e s , which v i s i t 1 a r g e unpala tab le p l a n t s l e s s o f t en than they do sma l l e r , more p a l a t a b l e ones (Hay 1986). Some s p e c i a1 i z ed e p i p h y t e s may o b t a i n n u t r i e n t s from t h e hos t (Harl i n 1973; Goff 1976).
In s i t u a t i o n s where consumption by f i s h e s d o e s n o t s e v e r e l y r e d u c e t h e i r numbers, sma l l c r u s t a c e a n s t h a t g r a z e e p i p h y t e s can o c c u r a t d e n s i t i e s o f s e v e r a l thousand/m2 i n s t a n d s o f macroalgae. In some cases , g raz ing by t h e s e small c rus taceans can keep l a r g e r seaweeds r e l a t i v e l y f r e e o f f o u l i n g ep iphytes (Brawley and Adey 1981a, b ) . These h ighly product ive ep iphytes a r e very important i n maintaining t h e high dens i ty and turnover r a t e o f small c rus taceans t h a t a r e such an important component of t h e d i e t of f i s h e s on rubble s t r u c t u r e s ( s e e l a t e r s e c t i o n s ) .
3.3 INVERTEBRATES
Community Composition
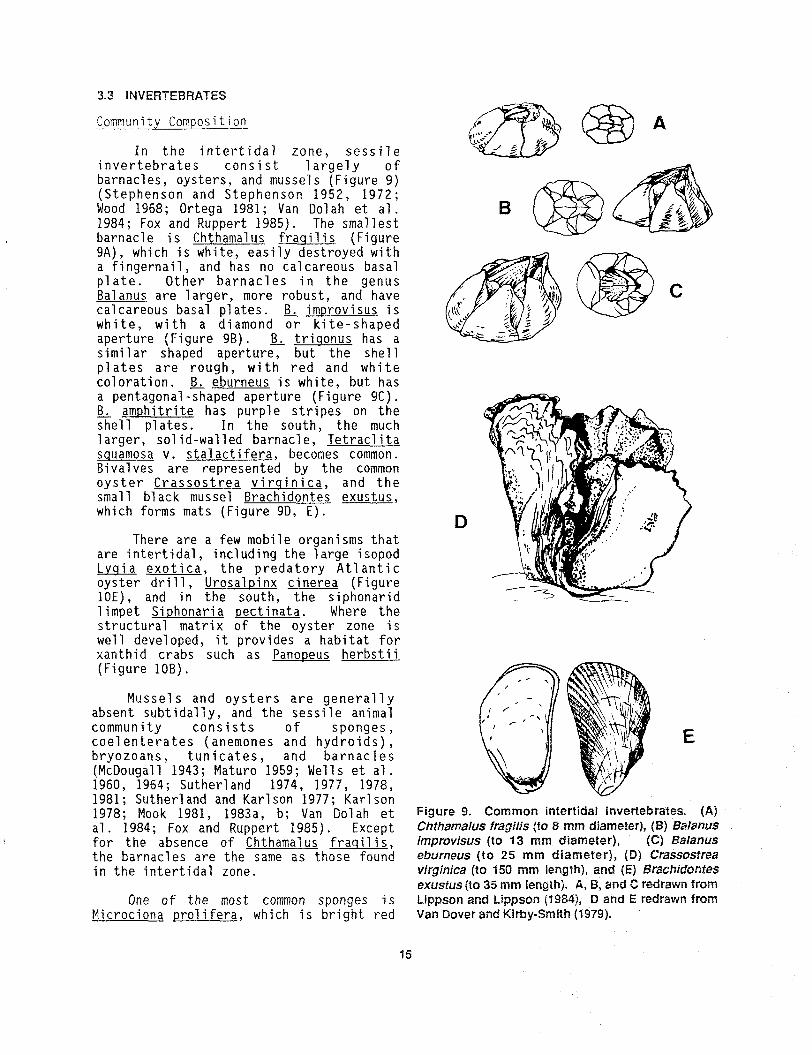
In t h e i n t e r t i d a l z o n e , s e s s i l e i n v e r t e b r a t e s c o n s i s t l a r g e l y o f barnac les , o y s t e r s , and mussels (F igure 9 ) ( S t e p h e n s o n and S t ephenson 1952, 1972; Wood 1968; Ortega 1981; Van Dolah e t a l . 1984; Fox and Ruppert 1985). The sma l l e s t barnac le i s Chthamalus f r a q i l i s (F igure 9A), which i s whi te , e a s i l y des t royed wi th a f i n g e r n a i l , and has no ca l ca r eous basal p l a t e . O t h e r b a r n a c l e s i n t h e genus Balanus a r e l a r g e r , more robus t , and have ca lcareous basal p l a t e s . B, i m ~ r o v i s u s i s w h i t e , w i t h a diamond o r k i t e - s h a p e d ape r tu r e (F igure 9B). B, t r i s o n u s has a s i m i l a r shaped a p e r t u r e , but t h e s h e l l p l a t e s a r e rough , w i t h r e d and w h i t e c o l o r a t i o n . B . eburneus i s whi te , but has a pentagonal -shaped a p e r t u r e (F igu re 9C). B. a m p h i t r i t e has purp le s t r i p e s on t h e - s h e l l p l a t e s . In t h e south , t h e much 1 a r g e r , so l id-wal led ba rnac l e , Te t r ac l i t a sauamosa v. s t a l a c t i f e r a , becones common. Bivalves a r e represen ted by t h e common o y s t e r C r a s s o s t r e a v i r q i n i c a , and t h e small black mussel Brachidontes exus tu s , which forms mats (F igure 9D, E ) .
D There a r e a few mobile organisms t h a t
a r e i n t e r t i d a l , inc lud ing t h e 1 a rge i sopod Lvqia e x o t i c a , t h e p r e d a t o r y A t l a n t i c o y s t e r d r i l l , U r o s a l ~ i n x c i n e r e a (F igure 10E), and in t h e sou th , t h e s iphonar id 1 impet S i ~ h o n a r i a o e c t i n a t a . Where t h e s t r u c t u r a l mat r ix of t h e o y s t e r zone i s well develooed. i t o rovides a h a b i t a t f o r xanthid c r i b s 'such ' a s Panopeus h e r b s t i i (F igure 10B).
Musse l s and o y s t e r s a r e g e n e r a l l y absent s u b t i d a l l y , and t h e s e s s i l e animal community c o n s i s t s o f s p o n g e s , c o e l e n t e r a t e s (anemones and h y d r o i d s ) , E b r y o z o a n s , t u n i c a t e s , and b a r n a c l e s (McDougall 1943; Maturo 1959; Wells e t a1 . 1960, 1964; Sutherland 1974, 1977, 1978, 1981 ; Suther l and and Karl son 1977; Karl son 1978; Nook 1981, 1983a, b; Van Do1 ah e t Figure 9. Common intertidal invertebrates. (A) a1 . 1984 ; Fox and Ruppert 1985) . Except Chthamaius fragilis (to 8 mm diameter), (8) BglarrUS
f o r t h e absence of Chthamalus f r a q i l i s , improwisus (to 13 mm diameter), (C) Balanus t h e barnac les a r e t h e same a s t hose found eburneus ( t o 2 5 rnm d iameter ) , (D) Crassosdrea i n t h e i n t e r t i d a l zone. virginica ( to 150 n m length), and ( E ) Brachidontes
exusfus (to 35 mm length). A, B, and C redrawn from One o f t h e most common sponges i s Lippson and Cippson (t984), D and E redrawn from
M i crociona pro1 i f e r a , which i s b r i g h t red Van Dover and Kirby-Smith (5979).

Figure 10. Common mobile invertebrates. (A) Ampithoe longimana (to 10 mm length), (B) Panopeus herbstii (to 25 mm length), (C) Asterias forbesii(t0 150 mm diameter), (D) Arbaciapunctulata (to 50 mm diameter), and (E) Urosalpinx cinerea (to 25 mm height). All redrawn from Van Dover and Kirby-Smith (1979).
and can be encrusting or erect with finger-like projections (Figure 1 1 C ) . Next in intensity of color is the bright orange, massively encrusting Xestos~onsia halichondrioides. Other sponges are rather drab. Mycale cecila is thinly encrusting, slimy, and a pale yellowish green or ye1 1 owi sh tan. Hal ichondri a bowerbanki assumes a variety of shapes, - starting as 1 ow encrustations, but ofteri developing a mass of ridges or branches (Figure 1 1 B ) . It is straw yellow, beige, or pale orange in color. A number of species of Haliclona (Figure 11A) may be found which are difficult to tell apart without reference to spicules. They are general 7y encrusting, softly spongy, and gray, tan, or pinkish brown in color. Finally, Lissodendorvx isodictyal is is a thickly encrusting sponge with a crisp
consistency and is blue-green or yellow- green in color. When broken open it is said to smell of garlic. Because of the variation in color and shape, spicules should be examined for positive identification (Wells e t al. 1960).
Tubularia crocea is one of the most conspicuous hydroids, forming 1 arge tufts of long, unbranched stalks topped with pink zooids (Figure 1 2 B ) . Other common hydroids are the white, delicate Obelia dichotoma, Eudendrium carneum with its intensely orange colonies, and Halocordvl e disticha (=Pennaria tiarella), which has a long black central stalk with two pinnate rows of side branches bearing polyps (Figure 1 2 A ) . In protected waters Hvdractinia echinata forms a white, fuzzy encrustation over the substrate. Other

Figure 11. Common sponges (A) ffaliclona s p . (colony to 8 cm wide), (B) ffaljchondria bowerbanki (colony to 8 cm high, 30 cm wide), and (C) Microciona prolifera (colony to 20 cm high, 30 cm wide). All redrawn from Lippson and Lippson (1984).
common c o e l e n t e r a t e s a r e t h e l a r g e s ea whip Leptoqorsia v i rqu l a t a (F igu re 12C), t h e small s tony co ra l As t ranqia danae, and t h e 1 a r g e r , b r a n c h i n g c o r a l Ocul i na arbuscul a . Anemones inc lude Bunodosoma c a v e r n a t a which i s l a r g e and w a r t y , A io t a s i a oa l ' l ida (F igure 12D), which i s sma l l and p a l e brown, and Diadumene leucolena , which i s pa l e and t r ans1 ucent .
The most abundant f o l i o s e bryozoan i s Busul a n e r i t i n a , with i t s r ed -pu rp l e bushy co lon i e s (F igure 13B) . B . s t o l o n i f e r a i s s i m i l a r i n morphology to-L n e r i t i n a , bu t whi te i n c o l o r . Ansuine l la palmata i s another f o l i o s e bryozoan but i t has g r ay , nondescr ip t co lon i e s . Alcvonidium hau f f i i s a gray-brown, rubbery bryozoan o f t e n found e n c r u s t i n a t h e s t a l k s o f o t h e r bryozoans and h d r o i d s . There a r e two common enc rus t i ng bryozoans t h a t can be d i s t i n g u i s h e d l a r g e l y on t h e b a s i s o f c o l o r ; w h i t e c o l o n i e s a r e p r o b a b l y Membrani~ora t enu i s and orange c o l o n i e s a r e probably S c h i z o ~ o r e l l a e r r a t a (F igure 13A) .
C o l o n i a l t u n i c a t e s are a1 s o conspicuous f e a t u r e s o f t h e s e s s i l e fauna. Eudis toma c a r o l i n e n s e forms t h i c k , i r r e g u l a r sandy e n c r u s t a t i o n s , whi le i t s congener Eudistoma hepaticum i s purp le and l i v e r - l i k e . Didemnum candidum forms t h i n , p u r e w h i t e e n c r u s t a t i o n s . C l a v e l i n a oblonsa i s a semicolonial t u n i c a t e wi th e l onga te , c l e a r , c o l o r l e s s zooids t h a t a r e j o i n e d t o g e t h e r o n l y a t t h e b a s e . Di s t a ~ l i a bermudens i s comes i n many c o l o r s - - r e d , orange, and purple--and has r e l a t i v e l y l a r g e zooids embedded i n t h e common t u n i c . The i n d i v i d u a l i n t a k e a p e r t u r e s a r e arranged i n c i r c l e s around a common e x h a l e n t o p e n i n g . Aul id ium c o n s t e l 1 atum i s r e d - o r a n g e t o w h i t e , hemispherical and looks 1 i ke a small b r a in a t t a ched t o t h e rocks. I t s congener .\, s t e l l atum forms wh i t i sh , tough, p l a t e -1 i ke co lon i e s with orange-red zooids arranged in a s t e l l a t e p a t t e r n (F igure 14C). Both a r e c a l l e d " sea pork" by l o c a l f ishermen. Peroohora vi r i d i s 1 ooks 1 i ke small , green grapes connected with green s t o l o n s and i s o f t e n found i n t h e canopy o f o t h e r h y d r o i d s and b r y o z o a n s . Di p l osoma macdonaldi forms t h i n , t r a n s p a r e n t s h e e t s which conta in t h e black zooids . Also seen a r e t h e gold-purple-brown, l oose blob-1 i ke r o l l s and lobes of Bot ry l l us sch l o s s e r i , e s p e c i a l l y in North Caro l ina . In South Ca ro l i n a and n o r t h e r n F l o r i d a more t r o p i c a l forms a r e found, such a s t h e p ink i sh , g e l a t i n o u s , enc rus t i ng Svrn~teama vi r i d e , wi th i t s b r i g h t l y cof ored zooids. Sol i t a r y t u n i c a t e s a r e represen ted by t h e g r a y g l o b e s o f Mol su l a m a n h a t t e n s i s (F igure 1481, t h e tough, wrinkled, t a n i n d i v i d u a l s o f S t v e l a pl i c a t a ( F i g u r e

Figure 12. Common coelenterates. (A) the hydroid, Haiocordyle disticha (to 15 cm high), (B) the hydroid, Tubuiaria crocea (to 15 cm high), (C) the sea whip, Leptogorgia virguiata (to 60 cm high), and (Dl the anemone, Aiptasia paflida (to 3 cm high). A, B, and C redrawn from Lippson and Lippson (1984), D redrawn from Spitsbergen (1980).

Figure 13. Common bryozoans. (A) Schizopo~ella errata (colony to 30 cm wide), and (B) Bugula nerifina (to 12 cm high). Redrawn from Spitsbergen (1980).
14A1, and t h e p a l e g r e e n A s c i d i a
Large mo t i l e sub t ida l i n v e r t e b r a t e s i n c l u d e t h e s t a r f i s h A s t e r i a s f o r b e s i i ( F i g u r e 10C) , t h e p u r p l e s e a u r c h i n , Arbacia uunctul a t a (F igure 10D) , t h e s t one c rab , Menippe mercenaria , and t h e blue c r a b , C a l l i n e c t e s s a p i d u s . The most common c r a b i s t h e somewhat s m a l l e r , o rangish mud c r ab N e o ~ a n o ~ e sa-yi-. Hermit c r abs a r e represen ted by t h e f l a t - c l awed P a a u r u s ~ o l l i c a r i s , t h e l o n g - c l a w e d Pasurus l o n s i c a r ~ u s , and t h e much l a r g e r C l i b a n a r i u s v i t t a t u s , which h a s y e l l o w s t r i p e s on t h e walking l e g s . The smal l , tube-bui ld ing gammarid amphipods A m ~ i t h o e (Figure 10A) and Corouhium spp. a r e common i n summer. In w in t e r , c a p r e l l i d amphi pods (Caure l la spp . ) a r e abundant. Many kinds
Figure 14. Common tunicates. (A) Sfyela plicafa (to 13 cm high), (Bf Molgula manhattensis (to 4 cm high), and (C) Aplidlum sfellaturn (colony to >30 cm wide). B redrawn from Lippson and Lippson (1984), @ redrawn from Gosner (1938).
o f s n a i l s a r e p r e s e n t , i n c l u d i n g t h e s m a l l , s e e d - s h a p e d , brown l u n a r dove s n a i l , A s t v r i s l u n a t a , t h e l a r g e r greedy dove s n a i l , Anachis avara , t h e s cu lp tu red t o p s n a i 1 , C a l l i o s toma e u s l rp tum, t h e At1 a n t i c o y s t e r d r i l l , Urosal ~ i n x c i n e r e a and t h e F l o r i d a r o c k s h e l l , T h a i s hemastoma f l o r i d a n a .

D i s t r i b u t i o n
On exposed j e t t i e s t h e supra1 i t t o r a l f r i n g e i s b lackened by b lue-g reen algae, over which wanders t h e occas iona l i sopod L v s i a e x o t i c a (F i gu re 5; Van Dolah e t a l . 1984; Fox and Ruppert 1985). Below t h i s i s a b a r n a c l e zone, u s u a l l y w i t h Chthamalus f r a q i l i s occupying t h e h i g h e s t l e v e l s and v a r i o u s spec ies o f Balanus a t 1 ower 1 eve1 s (F i gu re 5 ) . Barnac les can be abundant, approaching 100% cover (Or tega 1981). Below t h e barnac le zone a t mid- i n t e r t i d a l l e v e l s , i s a zone where t h e o y s t e r Crassos t rea v i r a i n i c a reaches i t s h i g h e s t abundance, a1 though t h i s r a r e l y exceeds 20% c o v e r i n exposed h a b i t a t s ( F i g u r e 5; O r t e g a 1981 ) . Be low t h e ba rnac l e and o y s t e r zones i s a zone w i t h h i g h d e n s i t i e s o f t h e mussel B rach idon tes exustus, which reaches t o mean low wate r ( F i g u r e 5). Th i s p a t t e r n o f zona t i on i s found w i t h m inor m o d i f i c a t i o n s th roughou t t h e S o u t h A t l a n t i c B i g h t , a l t h o u g h i n n o r t h e r n F l o r i d a we beg in t o see a more t r o p i c a l fauna. Balanus spp. g i v e way t o t h e 1 a rger ba rnac l e T e t r a c l i t a squamosa and t h e s i p h o n a r i d l i m p e t S i w h o n a r i a p e c t i n a t a becomes abundant i n t h e mid- i n t e r t i d a l zone (Stephenson and Stephenson 1972).
A s i m i l a r p a t t e r n o f zona t i on i s seen on more p r o t e c t e d r u b b l e s t r u c t u r e s excep t t h a t t h e l o w e r i n t e r t i d a l zone i s d o m i n a t e d b y t h e o y s t e r C r a s s o s t r e a v i r a i n i c a i n s t e a d o f t h e musse l B rach idon tes exus tus (Or tega 1981).
Sub t i da l l y , o u r i n f o r m a t i o n i s b i ased t o w a r d s shaded h a b i t a t s i n p r o t e c t e d wate rs o f N o r t h Ca ro l i na , where most work has been conducted. The ha l lma rk o f t h e sha l l ow s u b t i d a l community i s change. I n t h e i r s t u d i e s a t Beau fo r t , NC, S u t h e r l and and K a r l son (Su the r l and and K a r l son 1977; Ka r l son 1978; Su ther land 1981) found t h a t t h e 1 ongevi t y o f most communi t y members was l e s s t h a n a y e a r . Few s p e c i e s appeared a b l e t o t o l e r a t e t h e e n t i r e 25 OC a n n u a l t e m p e r a t u r e r a n g e a t B e a u f o r t . H y d r o i d s and t u n i c a t e s were e s p e c i a1 l y s e a s o n a l . S h o r t l i f e spans a l s o c o n t r i b u t e d g r e a t l y t o seasonal changes i n spec ies composi t ion.
C h a r a c t e r i s t i c a l l y , mature b e n t h i c assemblages were invaded by t h e t u n i c a t e
S t v e l a p l i c a t a each s p r i n g . Smal l i n d i v i d u a l s o f t e n grew ep i z o o t i c a l l y (on t o p o f o t h e r i n v e r t e b r a t e s ) and a f t e r a summer o f r a p i d growth became t o o heavy f o r t h e i r at tachment s i t e s . S. p l i c a t a commonly sloughed o f f i n t h e f a y , t a k i n g many o t h e r adher ing organisms w i t h i t . T h i s process produced bare spaces on t h e subs t r a t e , which were most o f t e n f i l l e d w i t h newly r e c r u i t i n g 1 arvae. V a r i a t i o n s i n l a r v a l r e c r u i t m e n t p roduced w i n t e r assemblages d o m i n a t e d b y a v a r i e t y o f sponges, hyd ro i ds, and bryozoans. Summer assembl ages were dominated by t h e s o l i t a r y t u n i c a t e S. p l i c a t a and t h e f o l i o s e b r y o z o a n Buqu l a n e r i t i n a ( S u t h e r l and 1981).
Wi th i n c r e a s i n g depth, annual changes i n spec ies abundance a re fewer (Ka r l son 1 9 7 8 ) . Much space i s o c c u p i e d b y r e 1 a t i v e l y 1 ong-1 i v e d o rgan i sms such as t h e h y d r o i d H v d r a c t i n i a e c h i n a t a , t h e sponge Xestoswonsia ha1 i chondro ides , t h e anemone D i adumene 1 e u c o l ena ( K a r l son 1978), and t h e c o r a l Ocul i n a arbusculum (McCl oskey 1970),
I n t h e South A t l a n t i c B i g h t near Cape Canaveral , FL, annual changes i n spec ies abundance a re fewer even i n sha l l ow wate r assembl ages (Mook 1976, 1980, 1981, 1983b ) . B a l a n u s spp . and t h e t u b e b u i l d i n g amphipod Co roph ium l a c u s t r e dominate these communit ies th roughou t t h e year .
Pa t t e rns o f Recru i tment
When s u b s t r a t e has been e x p e r i m e n t a l l y c l e a r e d (Or tega 1981) o r when new j e t t i e s a r e c o n s t r u c t e d (Van Do1 ah e t a1 . 1984), t h e genera l p a t t e r n o f i n t e r t i d a l zona t i on i s r e s t o r e d o r c rea ted by r e c r u i t m e n t w i t h i n a y e a r . N e z r Beau fo r t , NC, Or tega (1981) r e p o r t s most i n t e r t i d a l r e c r u i t m e n t o f B a l a n u s spp . d u r i n g summer and o f Crassos t rea v i r q i n i c a and Brach idon tes exustus i n f a l l , bu t i t i s unknown whether t h i s p a t t e r n i s t y p i c a l f o r t h e A t l a n t i c B i g h t . Recru i tment o f o y s t e r s i s much h i g h e r i n p r o t e c t e d wate rs t han on j e t t i e s and p i l i n g s on t h e open c o a s t , w h i l e t h e r e v e r s e i s t r u e f o r mussels (Or tega 1981). It i s l i k e l y t h a t r e c r u i t m e n t i s l o w e s t f r o m J a n u a r y t o March when temperatures a re mi n ima l (e. g . S u t h e r l and and K a r l son 1977).

Most s t u d i e s o f recru i tment p a t t e r n s o f s u b t i d a l , s e s s i l e a n i m a l s ( f o u l i n g organisms) have been conducted in North C a r o l i n a (McDougall 1943 ; Maturo 1959; We1 1 s e t a1 . 1964; Suther l and and Karl son 1977; S u t h e r l a n d 1 9 8 1 ) . The most e x t e n s i v e s t u d y i s t h a t o f S u t h e r l and (Suther l and and Karl son 1977, Suther l and 1981) who c o l l e c t e d da t a a t Beaufort , N C , f o r 6 1/2 yea r s (Figure 15 ) . Periods of r ec ru i tment f o r 11 common spec i e s show cons iderable seasonal i t y because o f t h e wide annual temperature ranqe a t Beaufort. - Some s p e c i e s , such a s H a l i c l o n a , Hal i c h o n d r i a , T u b u l a r i a , H a l o c o r d v l e (=Pennari a ) , and Bot rv l l us , r e c r u i t e d only p e r i o d i c a l l y . O t h e r s r e c r u i t e d p r e d i c t a b l y e a c h y e a r ( e . g . , S c h i z o o o r e l l a , S t v e l a , A s c i d i a , and C r a s s o s t r e a ) w h i l e s t i l l o t h e r s ( e . g . , Buqul a and Bal anus spp. ) r e c r u i t e d almost continuously (F igure 15 ) . In any given y e a r r e c r u i tmen t c o u l d be ex t reme1 y v a r i a b l e from month t o month, r e s u l t i n g in d i f f e r e n t p a t t e r n s o f community development on newly submerged s u b s t r a t e (Sutherl and and Karl son 1977).
J u s t south o f Cape Canaveral, F L , s i m i l a r v a r i a t i o n i n p a t t e r n s o f r e c r u i t m e n t has been o b s e r v e d by Mook (1976, 1980, 1983b) even though t h e annual f l u c t u a t i o n in temperature t h e r e i s 1 e s s . Some spec i e s r e c r u i t e d only p e r i o d i c a l l y , some p r e d i c t a b l y each y e a r , and some almost continuously. Organisms t h a t were common throughout t h e South A t l a n t i c Bight t e n d e d t o r e c r u i t more c o n t i n u o u s l y throughout t h e yea r (Mook 1976, 1980). Organisms with more t r o p i c a l a f f i n i t i e s
r e c r u i t e d p r i m a r i l y d u r i n g t h e warmer months.
3.4 FISHES
Community Com~os i t i on
The coas t a l warm-temperate f i s h e s o f t h e Sou th A t l a n t i c B i g h t f l u c t u a t e s e a s o n a l l y i n s p e c i e s c o m p o s i t i o n and abundance (Huntsman and Manooch 1978; Mi l l e r and Richards 1979; Lindquist e t a l . 1985; Van Dolah e t a l . 1986) . Most coas t a l f i s h e s a r e absent i n win ter and gradual ly r e t u r n t o inshore h a b i t a t s a s waters warm in sp r ing . Fishes found on coas t a l j e t t i e s tend t o be a subse t o f those found on inshore o y s t e r r e e f s and o f f sho re hard bottoms. These f i s h e s c a h be grouped i n t o f i v e general c a t e g o r i e s based on t h e i r mob i l i t y , need f o r ben th i c h a b i t a t c o m p l e x i t y , and s e a s o n a l i t y o f j e t t y occupancy. The f i r s t group c o n s i s t s of a l i m i t e d number o f sma l l c r y p t i c s p e c i e s , such a s t h o s e i n t h e b l e n n y (Blenni i dae ) and goby (Gobi idae) famil i es t h a t gene ra l ly do not move over l a r g e d i s t a n c e s and a r e dependen t upon t h e s t r u c t u r a l c o m p l e x i t y o f t h e j e t t i e s . These f i s h e s a r e o f t e n r e s i d e n t y e a r - round. The second group c o n s i s t s o f a l a r g e number o f n u m e r i c a l l y dominant s p e c i e s such a s p i n f i s h , Laqodon rhomboides , s p o t t a i l p i n f i s h , D i ~ l o d u s hol brooki , black sea bass , C e n t r o ~ r i s t i s s t r i a t a , and p i g f i s h , O r t h o o r i s t i s c h r v s o ~ t e r a , t h a t a r e abundan t d u r i n g warmer months but move o f f sho re i n c o l d win ter months. The t h i r d group i s made up
Haliclona I Hal lchondria
Tubularla Pennarla
Buyula
Schi zoporell a
S t y e l a Ascidia
Botrql11us
Figure 15. Recruitment periodicities for 11 common invertebrate species. Bars indicate when recruitment was observed on affificial plates exposed for 1-4 weeks (Sutherland 7981).

of several large predatory species such as bluefish, Pomatomus sal t a t r i x , and Spanish mackerel, Scomberomorus macul a tus , t h a t move over l a r g e d i s t a n c e s but a r e a t t r a c t e d t o j e t t i e s because of t h e inc reased d e n s i t y o f prey t h a t occurs there . The fourth group contains species, l i k e the smooth dogfish, Mustelus canis , t ha t are a t t rac ted t o j e t t i e s during t h e i r northerly migration in spring o r t h e i r southerly migration in f a l l (Van Dolah e t a l . 1984, 1986). The f i f t h and l e a s t important group contains various t ropical f i s h e s ( e - g . , b u t t e r f l y f i s h e s of t h e family Chaetodontidae and surgeonfishes of the family Acanthuridae) tha t occur as s t rays during the warmest months of the year. Only f i shes in the f i r s t and second groups a re t r u l y res idents of rubble s t ructures .
Although t h e f i s h e s of t h e South Atlantic Bight are well known (Bohlke and Chap1 in 1968; Dahl berg 1975; Manooch 1984; Robins e t a l . 1986), very few s tudies have focused spec i f i ca l ly on the f i shes using nearshore rubble s t ructures . Van Do1 ah e t a l . (1984, 1986) conducted e x t e n s i v e investigations of the f i shes associated with the large j e t t i e s a t Murrells I n l e t , SC, and Lindquist e t a l . (1985) studied t h o s e on sma l l e r j e t t i e s a t Masonboro I n l e t , NC. Data collected by Van Dolah e t a l . (1984, 1986) are the most extensive presently available and the pat terns they document agree well with those seen on offshore a r t i f i c i a l reefs (Parker e t a l . 1979; M. Hay, pers. obser . ) . We consider t h e i r f i n d i n g s t o be broadly representative of the patterns t h a t occur on most j e t t i e s in the South Atlant ic Bight. During t he i r investigations a t the Murrells In le t s i t e , Van Dolah e t a1 . (1984, 1986) c o l l e c t e d 93 s p e c i e s of f i shes representing 43 famil ies . A few of t h e most common spec i e s a r e d i s cus sed below. For i l l u s t r a t i o n s and s p e c i e s descriptions of a11 the f i shes discussed in t h i s t e x t , see Robins e t a l . (1986).
The smooth dogfish, Mustel us canis , and the clearnose skate , Raja e s l an t e r i a , are often abundant near j e t t i e s during the spring when they are migrating from deeper t o shallower waters o r from southern t o more n o r t h e r l y wate rs . The c l ea rnose skate i s the most common species of skate in inshore waters between Long Island and
North Carolina and grows t o have a d i sc width of s l i gh t l y l e s s than 1 m (Robins e t a1. 1986). I t feeds primarily on f i shes and l a r g e r c ru s t aceans (Hildebrand and Schroeder 1928; Van Dolah e t a l . 1986) and, as i t s name suggests, has broad c l ea r areas on each s ide of the snout. The smooth dogfish i s a small (up t o 1.5 m in length) shark tha t occurs between the Bay of Fundy and Uruguay. I t has several rows of small, pavement-like teeth and, 1 ike t h e c l e a rnose s k a t e , f e eds on l a r g e r crustaceans and small f i shes (Hildebrand and Schroeder 1928; Van Dolah e t a1 . 1986) .
Blue f i sh , Pomatomus s a l t a t r i x , a r e found between Nova Scotia and Argentina, a1 though they are ra re o r absent in the Caribbean. During the spring and summer, they are common in coastal areas along the South Atlant ic Bight. These large ( u p t o 1 . 1 m i n l eng th and 12 kg in mass) predators form aggregations when act ively feeding tha t often drive schools of prey f i shes in to shallow waters near swimming beaches. On such occasions, swimmers and s u r f e r s have been b i t t e n . Although bluefish are not resident on j e t t i e s , they often feed on the f i shes tha t do res ide there and are often caught by anglers c a s t i n g from j e t t i e s . B luef i sh a r e primarily piscivorous but may a l so consume nereid worms, crustaceans, and cephal opods ( p r i m a r i l y squ id s ) (Hildebrand and Schroeder 1928; Grant 1962; Richards 1976; Gallaway e t a1 . 1981; Van Dolah e t a1 . 1986).
Other common p i s c i v o r e s a r e t h e Spanish mackerel, Scomberomorus macul a tus , and t h e conger e e l , Conqer ocean icus . Li ke b1 uef i s h , Spanish mackerel move widely between many habi ta ts but frequent j e t t i e s t o feed on resident j e t t y f i shes . They a t t a i n lengths greater than 80 cm, weigh u p t o 5 kg, and can be distinguished from most o t h e r mackerel by t h e many la rge , dark brown and brassy spots on t h e i r s ides . The conger eel i s dark brown t o bluish gray, grows t o a s ize of 2.3 m and 40 kg. I t i s often caught by anglers f ishing on j e t t i e s , docks, o r p iers in the mid-At1 ant ic Sta tes (Robins e t a1 . 1986). Spanish mackerel and conger e e l s occur between Cape Cod and the Gulf of Mexico.

Three s p a r i d f i s h e s a re common on r u b b l e s t r u c t u r e s i n t h e South A t l a n t i c B i g h t : t h e p i n f i s h , Lasodon rhomboides, t h e s p o t t a i l p i n f i s h , D io lodus h o l b r o o k i , and t h e sheepshead, A r c h o s a r q u s p r o b a t o c e o h a l u s . D a r c y (1985a , b ) rev iewed a v a i l a b l e i n f o r m a t i o n on p i n f i sh and s p o t t a i l p i n f i s h , and Ogburn (1984) i n v e s t i g a t e d f eed ing by sheepshead on a No r t h C a r o l i n a j e t t y . Desp i t e t h e common p e r c e p t i o n t h a t h e r b i v o r o u s f i s h e s a r e absent f r om temperate communit ies (Bakus 1964, 1968; Montgomery 1977, 1980; Ogden and Lobe1 1978; Montgomery and Gerk ing 1980; Hay 1981b; Ga ines and Lubchenco 1982), a l l t h r e e o f these spec ies can consume l a r g e q u a n t i t i e s o f seaweeds (Ca r r and Adams 1973; Adams 1976a; S toner 1980; Ogburn 1984; S toner and L i v i n g s t o n 1984; Hay 1986; Hay e t a1 . 1987, 1988) and may s i g n i t i c a n t l y a f f e c t t h e s t r u c t u r e o f b e n t h i c seaweed c o m m u n i t i e s ( s e e l a t e r s e c t i o n s ) . I n a d d i t i o n t o seaweeds, sheepshead may consume s i g n i f i c a n t q u a n t i t i e s o f b i v a l v e s and b a r n a c l e s (Parker e t a1 . 1979; Van Dolah e t a l . 1984). I n t h e Caro l inas , s p o t t a i l p i n f i s h and p i n f i s h a r e among t h e most abundant spec ies on j e t t i e s . Sheepshead a re common b u t much l e s s abundant (L indqu i s t e t a1 . 1985; Hay 1986; Van Dolah e t a l . 1986).
I n d i v i d u a l s o f a l l o f these spec ies t end t o be r e l a t i v e l y smal l on coas ta l r u b b l e s t r u c t u r e s compared t o t h e l a r g e r i n d i v i d u a l s t h a t occur on o f f s h o r e r e e f s . Young i n d i v i d u a l s appea r t o c o l o n i z e j e t t i e s i n e a r l y s p r i n g , g row r a p i d l y th roughou t t h e summer, and move t o deeper o f f s h o r e areas as nearshore wa te rs coo l i n t h e l a t e f a l l (Darcy 1985a, b; L i n d q u i s t e t a1 . 1985; Van Dolah e t a1 . 1986) . I n t h e Caro l inas , these f i s h e s a re among t h e m a j o r p r e y s p e c i e s on o f f s h o r e r e e f s . T h e i r i n s h o r e - t o - o f f s h o r e m i g r a t i o n s may be i m p o r t a n t i n t r a n s f e r r i n g e n e r g y be tween p r o d u c t i v e i n s h o r e and d e e p e r o f f s h o r e h a b i t a t s ( D a r c y 1985a, b ) . P i n f i s h occur between Cape Cod and t h e Yuca tan . S p o t t a i l p i n f i s h a r e f o u n d be tween t h e Chesapeake Bay and t h e n o r t h e r n G u l f o f Mexico. Sheepshead a re more w i d e l y d i s t r i b u t e d and occu r f rom Nova S c o t i a t o B r a z i l ( R o b i n s e t a l . 1986).
P i g f i s h , O r t h o o r i s t i s ch rvsoo te ra , show seasonal i t y and m i g r a t o r y p a t t e r n s
t h a t r e s e m b l e t h o s e o f t h e s p a r i d s d i s c u s s e d above . Nea r B e a u f o r t , NC, o f f s h o r e m i g r a t i o n occurs i n l a t e f a l l o r w i n t e r . The l a r g e s t i n d i v i d u a l s a re t h e f i r s t t o l e a v e and t h e f i r s t t o r e t u r n i n t h e s p r i n g (H i ldebrand and Cable 1930). S ince i n d i v i d u a l s r e t u r n i n g i n t h e s p r i n g appear t o be i n poor c o n d i t i o n due t o t h e l e s s t h a n o p t i m a l f e e d i n g c o n d i t i o n s o f f s h o r e , p i g f i s h p r o b a b l y m i g r a t e t o avo i d l o w temperatures r a t h e r t han t o seek b e t t e r f e e d i n g g r o u n d s ( D a r c y 1 9 8 3 ) . P i g f i s h are a t t r a c t e d t o hard s u b s t r a t e and o f t e n school near r e e f s o r j e t t i e s ( H a s t i n g s 1 9 7 2 ) . They a r e g e n e r a l i s t c a r n i v o r e s , w i t h p r e y s i z e and t y p e changing as a f u n c t i o n o f age (H i ldebrand and Cable 1930; Ca r r and Adams 1973). Small f i s h f e e d on p l a n k t o n i c crustaceans. Larger f i s h feed on smal l f i s h e s , b e n t h i c crustaceans, m o l l usks, po lychae tes , and a v a r i e t y o f o t h e r i n v e r t e b r a t e s . P i g f i sh occur f rom Massachusetts t o t h e Yucatan.
Spot, Leiostomus xanthurus, a re popu la r p a n f i s h common on j e t t i e s d u r i n g t h e s p r i n g and f a l l . They a re found be tween M a s s a c h u s e t t s and t h e n o r t h e r n G u l f o f Mex i co , and e a t p r i m a r i l y b i v a l ves , decapods , and s m a l l e r crustaceans (Adams 1976a; Van Dolah e t a l . 1986) .
Both b l a c k sea bass, Cen t rop r i s t i s s t r i a t a , and tautog, Tautoqa o n i t i s , a re abundant p reda to r s t h a t r e s i d e on coas ta l j e t t i e s . B l ack sea bass range f rom Maine t o t h e n o r t h e r n G u l f o f Mexico. Tautog occur as f a r n o r t h as Nova S c o t i a b u t extend southward o n l y t o Georg ia (Robins e t a l . 1986; G i l l i g a n 1987). Both o f t h e s e s p e c i e s a r e commonly t a k e n b y hook and l i n e and by spear. L i n d q u i s t e t a1 . (1985) and Van Dolah e t a l . (1986) found t h a t t a u t o g consumed p r i m a r i l y j e t t y - a s s o c i a t ed b i v a l v e s and crustaceans. B l ack sea bass a l s o consume cons i de rab le q u a n t i t i e s o f crustaceans, b u t t h e ma jo r p o r t i o n o f t h e i r d i e t i s f i s h and t hey r a r e l y consume b i va l ves . J u v e n i l e b l a c k sea bass e a t a m i xed d i e t o f f i s h , decapods , amphipods, and o t h e r b e n t h i c i n v e r t e b r a t e s . As b l a c k sea bass i nc rease i n s i ze , t h e p r o p o r t i o n o f f i s h i n t h e d i e t i nc reases c o n s i s t e n t l y (Van Do1 ah e t a1. 1986).

Oyster t o a d f i s h , O ~ s a n u s tau, s k i l l e t f i sh, Gobiesox strumosus, seaboard gobies , Gobiosoma g i n s b u r s i , c r e s t e d bl ennies, H Y D ~ eurochi 1 us geminatus, and feather blennies, Hv~soblennius hentzi , were a l l common on the South Carolina j e t t i e s s t ud i ed by Van Dolah e t a l . (1986). These f i shes are c ryp t ic and tend t o be 1 ess mobile, and thus l e s s seasonal, than the other f i shes common on rubble s t ruc tures in the South Atlant ic Bight. They appear t o be among the only f i shes t h a t overwinter on the j e t t i e s and t ha t do not migrate seasonally t o warmer waters. Winter dens i t i es of these f i shes appear t o be lower than dens i t i es in warmer months. However, t h i s could be a sampl ing a r t i f a c t s i n c e most b l enn i e s and gobies become inact ive and she l t e r in crevices a t low water temperatures. 'Thi s behavior would i n c r e a s e t h e i r p r o b a b i l i t y of being undersampl ed during the winter.
On the j e t t i e s studied by Van Dolah e t a l . (1986) , o y s t e r t o a d f i s h a t e p r ima r i l y f i s h e s ( A t l a n t i c s i l v e r s i d e , Menidia menidia, menhaden, Brevoor t i a tv rannus , and black s ea ba s s ) and decapods. S k i l l e t f i s h , b l enn i e s , and seaboard gobies consumed a d i e t of mixed invertebrates.
The s k i l l e t f i s h i s the only c l ingf i sh on the U.S. coast tha t occurs north of F l o r i d a , i t s range extending from New Jersey t o Brazil . Oyster toadfish occur between Cape Cod and Florida. Because of t h e i r s i z e and ha rd ine s s ( a b i l i t y t o withstand pollution and other s t r e s s e s ) , they have become important experimental and bioassay organisms. Seaboard gobies occur from Massachuset ts t o Georgia , feather blennies from New Jersey t o Texas, and crested blennies from North Carolina t o Texas (Robins e t a l . 1986).
Large school s of At1 a n t i c s i lve rs ides , Menidia menidia, sometimes aggregate over shallow portions of j e t t i e s and consume the epifaunal amphipods t ha t occur t h e r e (Van Dolah e t a l . 1986) . Since Atlant ic s i lve rs ides are typ ica l ly found along sandy shore l i n e s and a t the mouths of i n l e t s , t h e i r occas iona l association with j e t t i e s i s more l i ke ly explained by the location of j e t t i e s a t i n l e t mouths than by t h e i r a t t r a c t i on t o t h e j e t t i e s themselves . At1 a n t i c
s i lve rs ides occur from the G u l f of S t . Lawrence t o t h e no r thea s t e rn c o a s t of Florida (Robins e t a l . 1986). In addition t o epifaunal amphipods, they consume other small benthic and planktonic crustaceans, polychaetes, bivalves, and juvenile f i shes (Hildebrand and Schroeder 1928; Adams 1976a; Bengston 1984; Van Dolah e t a l . 1986) .
Distribution
The spec i e s composit ion of l a r g e , noncryp t ic f i s h e s on shal low rubble s t ructures i s similar t o the community composition seen on natural and a r t i f i c i a1 reefs tha t occur offshore in the South Atlant ic Bight (Parker e t a1 . 1979; Van Dolah e t a1 . 1984; Sedberry and Van Do1 ah 1984; Lindquist e t a l . 1985; Van Dolah e t a l . 1986). However, inshore j e t t i e s tend t o have a lower d ivers i ty of species than na tura l , offshore reefs . Also the s i z e of individual f ishes tends t o be smaller on j e t t i e s (Buchanan 1973; Van Dolah e t a l . 1986; Wenner e t a1 1986), suggesting t h e i r ro le as nurseries.
When rubble s t r u c t u r e s such a s j e t t i e s are constructed, they are very rapidly colonized by f i shes . The seasonal nature of the inshore f i s h fauna obscures successional patterns in f i s h community s t ructure on new j e t t i e s , i f such pat terns occur (Hastings 1979; Van Dolah e t a l . 1984; Lindquist e t a l . 1985). The rapid movement of f i shes onto new1 y constructed j e t t i e s suggests tha t they are i n i t i a l l y a t t r a c t e d by t h e inc reased s t r u c t u r a l complexity, which provides she1 t e r from predators. However, q u t content analyses of common j e t t y f ishes such as black sea bass , p i n f i s h , s p o t t a i l p i n f i s h , sheepshead, spadef i sh (Chaetodiwterus faber) , tautog, grunts (Haemul idae) , and flounder (Bothidae) show tha t they soon come t o re ly on je t ty-associa ted fauna as a food source (Ogburn 1984; Van Dolah e t a l . 1984, 1986; Lindquist e t a l . 1985). When Lindquist e t a l . (1985) compared the f i shes associated w i t h a new j e t t y (1 year old) t o an older one (15 years old) a t Masonboro I n l e t , N C , they found few s i g n i f i c a n t d i f f e r e n c e s i n s p e c i e s ' dens i t i es . Pigfi sh and sheepshead were more abundant on the new j e t t y but i t i s possi b1 e t ha t differences other than j e t t y age caused t h e s e p a t t e r n s . On North

Carol i n a j e t t i e s , d i f f e r e n c e s in f i s h populations on old versus new j e t t f e s and on ocean versus i n l e t s ides of j e t t i e s appear t o be small compared t o the major d i f f e r e n c e s i n s p a t i a l use p a t t e r n s observed over depth gradients (Lindqui s t e t a l . 1985) . Of t h e n ine abundant species studied by Lindquist e t a l . , s i x were s i g n i f i c a n t l y more abundant a t a dep th of 2 m than a t 4 m ( p i n f i s h , s p o t t a i l p i n f i s h , sheepshead, p i g f i s h , b l u e f i s h , and round scad , D e c a ~ t e r u s puncta tus) , one was more abundant a t 4 m ( b l a c k sea b a s s ) , and two showed no s ign i f i can t changes in abundance between these depths ( tautog and spot) . Van Dolah e t a1 . (1986) noted s imilar pat terns f o r several of these species on the South C a r o l i n a j e t t i e s they s t u d i e d ; our experience with t h e North Carol ina j e t t i e s a t Cape Lookout, Shackleford Banks, and Radio Island suggests t h a t these pat terns occur on those j e t t i e s as well.
The abundance of s p a r i d f i s h e s ( p i n f i s h , s p o t t a i l p inf i s h , and sheepshead) i s known t o be p o s i t i v e l y corre l ated with vegetation cover. Stoner (1980b) demonstra ted a very high corre la t ion ( r = 0.998, p < 0.01) between pinf ish abundance and macrophyte biomass in seagrass beds. Other authors have made s imilar observations (Caldwell 1957; Kil by 1955; Schwartz 1964). I t appears t h a t the s p a r i d s a r e abundant i n shal low a r e a s because of t h e i n c r e a s e d abundance o f seaweeds. Seaweeds are fed on d i r e c t l y and a l s o support popu la t ions of small c r u s t a c e a n s t h a t a r e ano ther important component of the sparid d i e t (Ogburn 1984; Darcy 1985a, b). Pelagic spor t f i shes , such as bluefish and spani sh mackerel, may be a t t r ac ted t o these shallower depths because of t h e i n c r e a s e d abundance of spar ids and other prey.
As mentioned ear l i e r , 1 arge seasonal changes occur in the species composition and abundance o f f i s h e s on r u b b l e s t ruc tu res in the South At lant ic Bight. Numerous s t u d i e s sugges t t h a t most o f these changes are driven by t h e need of f i shes t o avoid t h e colder inshore water t empera tu res t h a t occur i n w i n t e r (Huntsman and Manooch 1978; Miller and Richards 1979; Parker e t a l . 1979; Lindquist e t a l . 1985; Van Dolah e t a l . 1986) . Even though t h e s e seasona l
temperature changes are l e s s pronounced a t 1 ower 1 a t i tudes, t h e same general pat terns of offshore o r southerly migrations appear t o occur throughout t h e e n t i r e South At lant ic Bight. As an example, p inf ish and s p o t t a i l p i n f i s h undergo s i m i l a r pat terns of seasonal migration i n both North Carolina and Florida (Adams 19766; Stoner and Livingston 1984; Darcy 1985a, b ) .
Seasonal p a t t e r n s of abundance on j e t t i e s in the bight have been studied most e x t e n s i v e l y on t h e j e t t i e s a t Murrell s I n l e t , SC (Van Do1 ah e t a1 . 1984, 1986) . Abundance and community composition of f i s h e s frequenting these j e t t i e s were assessed quar ter ly using g i l l n e t s , v i s u a l o b s e r v a t i o n s , c r a b t r a p s modified t o r e t a i n small f i shes (6.4 mm mesh), unmodified c r a b t r a p s , and qua1 i t a t i v e rotenone col lect ions . Gill nets were run from the j e t t y t o a d is tance of 23 m away from the j e t t y t o sample not only r e s i d e n t j e t t y f i s h e s , b u t a1 s o pelagic predators preying on these f i shes . Crab t raps were deployed on, o r very near, the j e t t i e s and thus assessed j e t t y f i shes t h a t would en te r t r aps . Visual counts by divers provided an addit ional assessment of noncryptic j e t t y f i shes t h a t may not have been adequately sampled by t h e other t e c h n i q u e s . The q u a l i t a t i v e ro tenone col l e c t i ons a1 1 owed a crude assessment of small c ryp t ic species l i k e blennies and gobies. Rotenone i s a toxin t h a t s t r e s s e s small f i s h , causing them t o leave t h e i r c ryp t ic hab i ta t s and swim in to t h e open water where they can be col lected.
Data from these s tudies are shown in Table 1 and in Figures 16-25. Most of these data a re reported in the l i t e r a t u r e , and thus in our t ab les and f igures , without an indication of t h c variance (Van Dolah e t a l . 1934, 1986). We have included measurements of variance where they e x i s t . The only other avai lable s tudies dealing w i t h f i shes on j e t t i e s in the bight (Ogburn 1984; Lindquist e t a1 . 1985; Hay 1986) are l e s s extensive b u t show s imilar patterns.
The t o t a l number of f i sh species seen in visual counts, o r col lected using g i l l nets o r modified crab t raps , was highest during warmer seasons o f the year and decreased markedly i n winter (Table 1 ) .

Table 1. Number of fish species collected or counted during different seasonson the jetties at Murrells inlet, SC (data from Van Dolah et a!. 1986).
Met hod Spring Summer Fat 1 Winter
Visual transects 11 2 4 2 2 1
Gill nets 25 3 4 25 6 Modif ied
crab traps 7 5 3 2 Unmodified
crab traps 11 9 8 7 Rotenone 13 11 12 9
Figure 16. Gill net collections from jettles at Murrells Inlet, SC. Plotted are the totals from three nets deployed for 3-hour set periods during each season. One end of the 30.5 m long net was set on the jetty; the other end was about 23 m from the jetty (data from Van Dolah et al. 1986).
04- Spring Summer Fall Wtnter
Flgure 18. Unmodified crab trap collections from the north jetty at Murrells Inlet, SC. Piotted are the total number and mass of fishes collected during each season using 15 traps set for a period of 12 daytime and 12 nighttime hours during each season (data from Van Dolah et al. 1986).
0- -0 indiv~duols 0--0 Moss
r- 0 600 I
> I/,
o do0 (/I
X
0 zoo ?,
-.-- 0 000 S p r q Sumrncr Fall Whntsr
Flgure 19. Modified crab trap (covered with 6.4 mm mesh to retain small fishes) collections from the base of jetties at Murrells Inlet, SC. Plots show means ct 1 standard error for 14 traps that were set for 3 hours during each season (data from Van Dolah et al. 1986).
Figure 57, Diver observations of the ~ e ~ ~ o n a l Figure 20. Rotenone collections from the jetties at abundance of common fishes on the jettiesat Murrells Murrells Inlet, SC. Plotted are the total number of Inlet, SC. Plotted are thetotals from ten 5min Counts fishes In a single qualitative collection made during In each season (data from Van Dolah et al. 1986). each wason (data from Van Dolah et al. 1986).

1001 Raja egianteria (Clearnose Skofe)
cn 500 - Mustelus canis 80 0 3
(Smooth Dogfish) TJ 400 .- > - 60
300 -
40 C
0 200 I a, 13 100
20
E 3 z 0 0
Scomberomorus maculatus Vl 200 - Pornatomus salfatrix 150
0 3 TJ 160 125 .- > . - 100
120 - 'C
75
0 80 L 50 a,
40 E 25 3
0 0
cn 401 Brevoortia fyrannus 350 Leiostomus xanthurus (Atlantic Menhaden) 3oo
250
200
150
100
50
0
Spring Summer Fall Winter Spring Summer Fall Winter
Figure 21. Seasonal abundance of common fishes captured in gill nets set near jetties at Murrells Inlet, SC. Histograms show the totals from 3 nets set for 3 hours during each season (data from Van Dolah et al. 1986).
Lindquist e t a l . (1985) also found a significant correlation between the mean number of species observed each m o n t h (counted in visual transects) and water temperature ( r = 0.82, p < 0.01). Van Dolah e t a l . ' s (1986) collections from unmodified crab t r aps and by rotenone indicated that species number was a t a low in the winter, b u t the relative change was s l i g h t compared to t h a t of the other collection methods (Table 1) . Figures 16- 20 show seasonal change in the t o t a l number, and in some cases total mass, of fishes collected by each method. All of these show large decreases in the winter
even though unmodified crab t r aps and rotenone co l l ec t ions , again, tended t o show smaller re1 ative reductions than did the other collection methods. Rotenone col lec t ions focused primarily on small blennies and gobies that are probably less able than the l a rge r species t o successfully make the long offshore migration to deeper water. Reasons for the reduced seasonal variat ion i n unmodified crab-trap collections are 'less apparent.
Figures 21-25 show seasonal patterns of abundance for the most common species

/ (Spottai l Pinf ish) 500 : Lagodon rhomboides
(Pinfish)
A
o Menidia menidia 0 2 0 ."- ( A t l o n f i c Silverside) X - rn 16 - 0 3 75 .- > 12 .- u c - L 8 0 L
X 4 E 3
0
80 1 Mugil cephalus
! 20 V) - Leiostomus xanthurus 0 j 100 T;! .-> 80 v C -
60 0
40 n E 20 3 Z
0
Spnng Summer Fall Winter Spring Summer Fall Winter
Figure 22. Seasonal abundance of common fishes observed by divers on the jetties at Murrells Inlet, SC. Histograms show the totals from ten 5-min counts during each season (data from Van Dolah et al. 1986).
assessed by each o f t he methods discussed above. All species show peak abundances i n spring, summer, o r f a l l and are absent o r r e l a t i v e l y r a r e i n w in ter .
Feedinq Pat terns o f Fishes On and Near J e t t i e s
Three s t u d i e s have i n v e s t i g a t e d feed ing by f i s h e s on j e t t i e s i n the South A t l a n t i c Bight . Ogburn (1984) q u a n t i f i e d t h e g u t contents o f sheepshead c o l l e c t e d From j e t t i e s a t Masonboro I n l e t , NC, and
L i n d q u i s t e t a l . (1985) i n v e s t i g a t e d feeding by sheepshead, p i n f i sh, s p o t t a i l p i n f i s h , and tautog on t h i s same j e t t y . Van Dolah e t a1 . (1986) have provided t h e broadest data base on t h i s top i c . They q u a n t i f i e d the gut contents o f 55 f i s h species captured near Murrel 1 s I n 1 et , SC. Table 2 shows t h e i r f i nd ings f o r those species and seasons where a t l e a s t t h ree i n d i v i d u a l s t h a t c o n t a i n e d f o o d were examined. About h a l f o f t h e spec ies examined f e d p r i m a r i l y on j e t t y b i o t a du r ing one o r more seasons. Sheepshead,

50, (Oyster Toadfish) 70 -. Centropristis striata
Spring Summer Fall Winter Spring Summer Fall Winter
Figure 23. Seasonal abundance of common fishes captured In unmodified crab traps deployed on the north jetty at Murrells Inlet, SC. Histograms show the totals from 15 traps set for 12 hours during the day and 12 hours during the night for each season (data from Van Dorah et a!. 1986).
bl ack drum (Poaoni as cromi s ) , tautog, and spadefish were among the important recreational f i shes tha t fed almost exclusively on j e t t y biota. Other important recreational f i shes , such as bluefish, black sea bass, spotted seatrout (Cvnoscion neb01 osus) , red drum (Sci aenoas ocell atus), Spanish mackerel, and weakfish (Cvnoscion reqal i s ) , were ind i rec t ly dependent on the jet ty since they fed heavily on smaller fishes that directly consumed jet ty biota (Van Dolah e t a? . 1986) . Patterns of Recruitment
The general pattern of larval recruitment seen for most reef fishes in the South Atlantic Bight i s that large numbers of juveniles invade ree fs , je t t ies , or estuaries starting l a te in the winter and continuing throughout the summer (Hildebrand and Cable 1930; Wang and Raney 1971; Hoss 1974; Thayer e t a l . 1974; Adams 1976b; Parker e t a l . 1979; Boreman and Dean 1980; Van Dolah e t a1 . 1986). In our experience with j e t t i e s in
North Carolina, i t appears that je t t ies on the outer coast a re colonized in the spring by bo th adults and juveniles, while j e t t i e s in the sounds are colonized primarily by juveniles.
Je t t ies as Concentrated Nurseries
Je t t ies often harbor high densities of young fishes that typically l ive on offshore reefs as adul ts . P inf ish , spo t ta i l p inf ish , bl ack sea bass, sheepshead, spadef i sh, and gag (Mvcteroaerca micro1 e ~ i s ] a7 1 provide examples of t h i s . For these f i shes j e t t i e s obviously serve as nurser ies , providing both feeding s i t e s and the s t ruc tu ra l complexity necessary fo r avoiding predators. These fishes can be very dense on j e t t i e s (Figure 26 ) ; spottail pinfish can occur a t 8/m2 (Hay 1986). However, the extreme1 y smaf l area covered by rubble structures compared t o the immense stretches of sandy beaches and estuarine habitats i n the bight, suggests that rubble structures cannot be having a

e a ~ w n j
ea~ep{sXn
epOdejaa
epodo1auoa-j
N - - - N- - - ' emh '4 - U N L D O NID c ; N W m- U V V1 d -(D FlNN V3 N-
w-m w u r n O - N
Gin in 0, - -
V
epodo~lseg
eozoyiuy
~ O Z O J ~ ~ H
eJaj ! J O ~
ep!Aa~ru!ae~oj
~ P ~ L V
u o n a s
u "7 ? --+ Y Y Y N - \D
--,.- N- V V V
--- v Y
OD om,.,
- - v
- -7 Y V Y o m - 7-F
v Y - 6. --N V Q *
n.3 b ASb A A 2 2 %2%2,=2,Zu% % %ZL%ZUSLZ , paukurexa sq~ewoas p?) 41 !n s4JPUOaS
Y ~ P V sip=^ ~ 2 6 u a i 7 ~ o d = i d
4 3 6 ~ ~ ~ t e l o l = i l mua at ~ ~ e p u e a s - i s
:(w) J ~ U P J JZlS
.,, ,,, N on^ n - * CI m z = s ~ 2 z ~ ~ ~ ~ w m m m o L D m u 5 mx>> m~ : 2 n\ w\zxm-..rn\m-..p'N'd~Nx2 $ ~ ~ ~ Y Y . ? ? ~ ? ? T?
N N - N- N - - F w i n h o m r - w m ~ m
NN NU N
k3-.Z-~-Z-~-Z-Z-g-e- ~ - g - ~ - g - z - 3- 2-g-Z-2- ?27'2"?l,Y3,93,7e7&":2";2 y & y d y & y 8 y & y< y 2 y $ T & ; & 2"$-~-k-~-3-&'~-" - z-zuulh--- -- h - ~ - g - ~ - - N - - - - o h m o m - w

e no scion nebulosbs T t T e s e a t r o o t )
Qnoscion r e a l i s (weakfi+-'
Le iosto~nus xanthurus -EiG$---
Ment i c i r rhus arrericanus - 7 G Z K e X kingfish) H e n t i c i r r h u s 1 i t t o r a l i s - 3 u l f g f i s h j
Mic;"pogon~as undulatus A t l a n t i c c r o a k e r )
P o g y i a s cromi s black=
Sciaeno s o c e l l a t u s - & r ~ r - Chaetodi ter us faber +T -- Tab? o n i t i s
t a u t r
Gabiosona i n s b u r i - 7 S s a M ~
Sco~beromorus nacu la tus - (Spanish m a m -
Pe r i l u s a l e i d o t u s *e*
<I 74 1-3.5.15.21-23
100 100 24-26
100 TOO
1 2.4.6.2l.25- 36
I - - a Many o f t he b lack sea bass examined i n f a l l were f rom t r a p s b a i t e d w i t h menhaden. Th is p robab ly i n f l a t e d the p r o p o r t i o n of f i s h i n t h e d i e t .
Code f o r f eed ing references: 1. t i l ldebrand & Schroder 1928 14. 2. Darnell 1958 15. 3. Cdr r & Adam 1973 16. 4. D iene r e t a l . 1974 17. 5. Odurn & Hera ld 1972 18. 6 , Adams 1976a 19. 7. Bengston 1984 20. 8. S te im le h Ogren 1982 21. 9. Grant 1962 22. 10. Gal lawsy e t a l . 1981 23. 11. Richards 1976 24. 12. Darcy 1983 25. 13. Van Ooldh e t a l . 1984 26.
L i n d q u i s t e t a l . 1985 Overs t ree t & Heard 1982 Ogburn 1984 Spr inge r L Woodburn 1960 Hansen 1969 S toner 1 980a Oarcy 1985a Darnel1 1961 f,lercer 1 984a Pearson 1929 M e r r i n e r 1975 Chao & Musick 1977 S t i ckney e t a l . 1975
27. C u r r i n e t al. 1984 28. Govoni et al. 1983 29. Govoni e t a l . 1986 30. Hales 8 Van de A r y l e 1985 31. Hodson e t a1 1981 32. K je l son e t a1 1975 33. Koby l i nsk i & Sheridan 1971 34. Roelofs 1954 35. Sher idan C Tr im 1983 36. Smi th e t a l . 1984 37. Bearden 1963 38. Overs t ree t & Heard 1978b 39. Re id e t a l . 1956
Bass 6 A v a u l t 1975 Mercer 1984b Overs t ree t L Heard 1978a B o o m y & A v a u l t 1971 Randal l 6 Hartman 1968 O l l a e t a l . 1974 L i n d q u i s t 6 Oi l laman 1986 Munroe & Lo tspe ich 1979 Naughton 6 Soloman 1981 B e r r i e n b Finan 1978 Randal l 1967 B i g e l o 6 Schroeder 1953 R a d c l i f f e 1914 Schwartz 6 Outcher 1963

Centroprjstis stria Fa l o 1
(Black Sea Bass) i Conger oceanicus (Conger Eel)
Opsanus tau 300 1 Menidia menidia w - o (Atlantic Silverside) 3 12 -0 . - > .- z 9 - *- 0
6 L 0)
P
E 3 3 Z
0 Spring Summer Fall Winter Spring Summer Fall Winter
Lagodon rhomboides (Pinfish)
Spring Summer Fall Winter
Figure 24. Seasonal abundance of common fishes captured in modified crab traps at Murrells Inlet, SC. Histograms show the total numbers captured in 24 traps set for 3 hours during each season (data from Van Dolah et al. 1986).
s ign i f i can t e f f ec t on the regional abundance of these species.
3.5 BIRDS
Cornunity Comuosition
A great variety of birds in the South Atlantic Bight use rubble structures as loafing cr roasting s i tes . However, birds in a few ecological categories, or "guildsn, feed on or near j e t t i e s and can be considered part of the rubble structure
community. The guilds include: (1) surface-searching shorebirds, ( 2 ) aeri a1 - searching birds, (3) floating and diving water birds, and (4 ) wading birds (Table 3 ) . Surface-searching shorebirds feed primarily on crustaceans, polychaetes, barnacles, moll uscs, and insects. The most common shorebird i s the ruddy turnstone, Arenaria in terures . When feeding on je t t ies i t i s often found in groups of 100 or more ( G . Marsh; pers. comm.) . Purpl e sandpipers, Cal i d r i s mari tima, are a1 so occasionally abundant, in flocks of 40-50. Both the ruddy

Goblosoma ginsburg? 1207 (Seaboard Goby)
100 - Gobiesox strumosus 100
0 3
80 8 0 > - TJ C 60 60
C
O 40 40 L aJ
20 E 2 0
2
0 0
Hypleurochilus geminatus
Spring Summer Foll Winter Spring Summer Fall Winter
Figure 25. Seasonal abundance of common fishes in rotenone collections at Murrells Inlet, SC. A single collection was taken each season (data from Van Dolah et al. 1986).
turnstone and the purple sandpiper use rocks and j e t t i e s as the i r primary feeding habitats. Other shorebirds use them only on occasion, feeding on surrounding mudflats and seagrass beds as well (Peterson and Peterson 1978; Thayer e t a l . 1984) .
Aerial -searching birds include a group of sea gulls (Table 3) which are opportunistic consumers of "anything they can get off the rocks" (C. Marsh; pers. comm. ) . This includes molluscs, crustaceans, f ish, insects, carrion, and refuse. The most common bird in th is group i s the herring g u l l , Larus arsenta tus . Brown pel icans, Pelecanus occidentalis and a variety of terns are also frequently seen roosting on rubble structures, b u t generally do not use them as feeding s i t e s (3. Parnell; pers. comm. ) .
The guild of f loa t ing and diving water birds i s composed largely of a group
of sea ducks (Table 3 ) . Most of these are strong underwater swimmers and feed on echinoids (sand do1 1 a r s ) and bivalves (clams, and s c a l l o p s ) found on sandy bottoms near j e t t i e s . The surf scoter, Melanitta ~ e r s u i c i l l a ta , i s a particularly capable diver, attaining depths of 12 m or more. Fish are also taken by birds in t h i s group, e spec ia l ly by the double- breasted cormorant, Phal acrocorax auri tus. The American wigeon, Anas americana, does not dive; i t i s a surface dabbler, feeding primari 1 y on a1 gae and seagrasses.
Wading birds (Table 3) are commonly found only near rubble structures built in estuaries away from severe wave action, In these quiet, shallow waters they can be seen feeding on small fishes and crabs.
Distribution
Most shorebirds t h a t make use o f je t t ies breed i n the Arctic during June and Ju1 y (Johnsgard 1981 ; Farrand 1983a).

Figure 26. Spottail pinfish, Diplodus holbrooki, on the jetty at Radio Island, NC. The common sea urchin Arbacia punctulatacan be seen in the lower center of the picture.
They can be found along the southeast coast o f the Uni ted States du r ing the r e s t o f t he year. The American oystercatcher and b o a t - t a i l e d g r a c k l e a re y e a r - r o u n d residents. The r i n g - b i l l e d g u l l breeds i n the western Uni ted States and Canada and i s seen i n the South A t l a n t i c Bight only i n w in ter . Other g u l l s are year-round
res idents (Farrand 19836). Except f o r the double-breasted cormorant which i s present year-round, most f l o a t i n g and d i v i n g water b i r d s breed dur ing the summer i n Canada and the A r c t i c (Farrand 1983a). They can be found i n t h e South A t l a n t i c B i g h t dur ing the remainder o f the year. The wading b i r d s are year-round residents.

Table 3. Types of birds common to rubble structures in the South Atlantic ~ ight?
Species Present
Surface-searching shorebirds Black-be1 1 ied plover (Pl uvi a1 i s squatarol a) Semi pal mated plover (Charadri us semi pa1 matus American oystercatcher (Haematopus pal 1 i atus) W i l l e t (Catoptrophorus semi palmatus) Ruddy turnstone (Arenari a i nterpres) Red knot (Cal i d r i s canutus) Sander1 i ng (Cal i d r i s *) Semi pal mated sandpiper (Cal i d r i s pusi 11 a) Western sandpiper (Cal i d r i s mauri) Least sandpiper (Gal i d r i s minuti1 l a ) Purple sandpiper (Cal i d r i s maritima) Dun1 in (Cal i d r i s a lpina) Boat-tailed grackle (Qui sca1 uss ma.ior) Fish crow (Corvus ossi fragus)
Aerial -searching birds Laughing gul l (Larus a t r i c i l l a ) Ring-bi l led gul l (Larus del awarensi s ) Herring gull (Larus argentatus) Great black-backed gul l (Larus marinus)
Floating and diving water birds Common loon (Gavia immer) Horned grebe (Podi ceps aur i tus) Double-crested cormorant (Phal acrocorax auri t u s ) American wigeon (Anas americana) Canvasback (Aythya val i s i ne r i a ) Redhead (Aythya americana) Ring-necked duck (Aythya c o l l a r i s ) Greater scaup (Aythya mari la) Lesser Scaup (Aythya a f f i n i s ) Oldsquaw (Cl angul a hyemal i s ) Black sco te r (Melanitta nigra) Surf sco te r (Me1 ani t t a pe r sp i c i l l a ta ) White-winged sco te r (Melanitta fusca) Common goldeneye (Bucephal a cl angul a) Red-breasted merganser (Mergus s e r r a to r )
Wading birds Great blue heron (Ardea herodias) Great egre t (Casmerodius albus) Snowy egre t (Egret ta thul a) Green-backed heron (Butorides s t r i a tus) Black-crowned night heron (Nycticorax nycticorax) Tricolored heron (Egretta t r i c o l o r )
winter winter res ident res ident winter winter winter spring, fa1 1 winter winter winter winter res ident res ident
res ident winter resident res ident
winter winter res ident winter winter winter winter winter winter winter winter winter winter winter winter
resident res ident res ident res ident res ident resident
a ~ a t a f o r t h i s l i s t were assembled from Pearson e t a1 (1942), Stokes and Shackle ton (1968), Zingmark (1978), Farrand (1983a, b) , S c o t t e t a l . (1983), and o b s e r v a t i o n s of C. Marsh (Un ive r s i t y of South Carol ina, Coastal Carol ina Co1 lege, Conway, SCJ, J. Parnel l (University of North Carolina, Wilmington, Wilmington, N C ) , and W. Hon (University of Georgia, Marine Extension Service, Savannah, G A ) ,

CHAPTER 4. ECOLOGICAL PATTERNS AND PROCESSES
This chap te r d i s cus ses t h e eco log ica l processes t h a t determine t h e d i s t r i b u t i o n and abundance o f f l o r a and f a u n a assoc ia ted with rubble s t r u c t u r e s i n t h e South A t l a n t i c Bight. In gene ra l , we r e s t r i c t our a t t e n t i o n here t o wa te r s o f r e l a t i v e l y high s a l i n i t y , c h a r a c t e r i z e d by p r e d o m i n a t e l y mar ine o r g a n i s m s . I n severa l ca se s , t h e processes organiz ing t h e s e communi t ies have n o t been in t ens ive ly s tud i ed on rubble s t r u c t u r e s , and we mus t , t h e r e f o r e , i n f e r t h e i r importance from s t u d i e s conducted i n o t h e r s i m i l a r h a b i t a t s . In t h i s chap te r , we out1 i n e our general conceptual framework and then concent ra te on t h e o rgan iza t ion o f i n t e r t i d a l communi t i e s , s u n l i t , sub t ida l communities (which tend t o be dominated by s e a w e e d s ) , and s h a d e d , subt ida l communities (which tend t o be dominated by benth ic animals) . We a l s o d i scus s t h e e f f e c t s of i n d i r e c t , complex i n t e r a c t i o n s among organisms s i n c e t h e s e i n t e r a c t i o n s a r e probably more important than has been gene ra l ly apprec ia ted .
4.1 CONCEPTUAL FRAMEWORK
In a d d i t i o n t o the s e a s o n a l temperature e f f e c t s d i scussed previous ly , t h e two major physical g r a d i e n t s a f f e c t i n g t h e p l a n t and animal communities of rubble s t r u c t u r e s a r e t i d a l l e v e l and ava i l a b i l i t y of sunl i gh t .
The animal communities i n p a r t i c u l a r a r e well segregated by t i d a l l eve l f o r reasons d iscussed by Jackson (1977). I n t h e i n t e r t i d a l zone s o l i t a r y s e s s i l e animals dominate. These animals u sua l ly p o s s e s s h a r d e x t e r n a l c o v e r i n g s ( e - g . , s h e l l s ) which confer s u p e r i o r r e s i s t a n c e t o t h e h a r s h e r p h y s i c a l c o n d i t i o n s experienced when exposed dur i ng 1 ow t i d e . Subtidal l y , co1 oni a1 animal s a r e more
abundant because they have inde termina te , v e g e t a t i v e growth and a r e l e s s s u s c e p t i b l e t o ove rg rowth and g r a z i n g . Sol i t a r y a n i m a l s s u r v i v e s u b t i d a l l y t h r o u g h morpho log ica l o r b e h a v i o r a l a t t r i b u t e s (1 a rge s i z e , aggrega t ive behavior) which p r o t e c t them i n competi t ion with co lon ia l animals and from sub t ida l p r eda to r s .
Jackson (1977) has a l s o argued t h a t when l i g h t i s s u f f i c i e n t and a l l e l s e i s e q u a l , c o l o n i a l a n i m a l s l o s e i n competi t ion with p l a n t s . This i s because many p l a n t s , 1 i ke co lonia l animals , have c h a r a c t e r i s t i c s t h a t a r e i m p o r t a n t i n competi t ion ( e .g . , asexual reproduct ion and inde termina te growth). Add i t i ona l ly , wi th adequate l i g h t , p l a n t s have a f u r t h e r a d v a n t a g e i n t h a t t h e y c a n p h o t o s y n t h e t i c a l l y d e r i v e e n e r g y f o r growth and reproduct ion. In c o n t r a s t , co lon ia l animals must depend on ex t e rna l food s o u r c e s . T h e r e f o r e , s eaweeds g e n e r a l l y domina t e s h a l l o w s u n l i t h a b i t a t s , and co l oni a1 animals g e n e r a l l y dominate deeper , da rke r water and shaded h a b i t a t s beneath docks and br idges .
Wave ac t ion appears l e s s important than t i d a l l eve l and s u n l i g h t , but some o r g a n i s m s , f o r example t h e o y s t e r Cras sos t r ea v i r q i n i c a , a r e in to1 e r a n t o f t h e h igher wave ac t ion of t h e open c o a s t (Ortega 1981).
4.2 ORGANIZATION OF INTERTIDAL COMMUNITIES
In s p i t e o f t h e p a u c i t y o f experimental s t u d i e s , i t i s l i k e l y t h a t t h e o r g a n i z a t i o n o f t h e i n v e r t e b r a t e community on r u b b l e s t r u c t u r e s i s by processes s i m i l a r t o those ope ra t ing i n o t h e r r o c k y i n t e r t i d a l communi t i e s (Connell 1972; Menge 1976). On exposed shores , t h e upper l i m i t s of blue-green

algae, barnacles, and mussel s a re probably controlled by physical f ac to rs (e.g. Van Dolah e t a1. 1984). Chthamalus f r a q i l i s i s probably r e s t r i c t e d t o t h e upper l eve l s of the barnacle zone through competition with l a rger , f a s t e r growing Balanus spp. (Wethey 1983, 1984). Simil a r l y , the lower d i s t r i b u t i o n a l l i m i t of b a r n a c l e s i s probably s e t by compet i t ion wi th t h e mussel Brachidontes e x u s t u s s i nce predation on barnacles in the i n t e r t i d a l i s minimal (Ortega 1981; Van Dolah e t a l . 1984). The lower l i m i t of the mussel beds a t mean low water (Figure 5) i s probably a r e s u l t of p r e d a t i o n by t h e s t a r f i s h Asterias f o r b e s i i , t h e sheepshead, (Van Dolah e t a l . 1984) , and t h e A t l a n t i c oyster d r i 11 (Wood 1968). Experimental s tudies have shown t h a t oyster abundance on exposed j e t t i e s i s low because exposure t o heavy wave action r e s t r i c t s growth and s u r v i v a l and because o y s t e r s a r e outcompeted by t h e mussel Brachidontes exustus (Ortega 1981),
In protected waters i t i s a l s o 1 i kely t h a t the upper l i m i t s of each zone a re determined by physical f ac to rs while the lower 1 imi t s a r e determined by biological f ac to rs . Again, Chthamalus f r a q i l i s i s probably excl uded from 1 ower 1 eve1 s through compet i t ion wi th t h e l a r g e r Balanus spp. ( e . g . Wethey 1984) . Brachidontes e x u s t u s i s a b s e n t from protected waters and Ortega (1981) has shown t h a t oysters competitively exclude barnacles from the oyster zone. In waters of r e l a t i v e l y high s a l i n i t y the re i s an abrupt end t o the oyster zone a t mean low water (Wells 1961). This i s probably a r e s u l t of several biological processes, i n c l u d i n g p r e d a t i o n by o y s t e r d r i l l s , U r o s a l ~ i n x c i n e r e a (Chestnut and Fahy 1953) and Thais f lor idana (Wells and Gray 1960), and she l l erosion by t h e boring sponge Cliona c e l a t a (Lunz 1943). Oysters a r e found s u b t i d a l l y on ly where low s a l i ni t y excludes t h e s e o t h e r s p e c i e s (We1 1 s 1961).
Physical and biological disturbances can cause c o n s i d e r a b l e changes in t h e abundance of organisms in each zone. For example, near Beaufort, NC, Brachidontes exustus was absent from p i l ings on the open beach from May through August 1977 (Ortega 1981). Abundance increased t o n e a r l y 100% cover i n September 1977,
remained high unt i l February 1978, and decreased again t o near 0% a f t e r March. Mortality seemed t o be a d i r e c t r e s u l t of wave actiofl on mussels which had increased in s i z e during a winter of growth. I t i s p o s s i b l e t h a t t h e wooden s u b s t r a t e provided a 1 ess secure attachment s i t e than t h e g r a n i t i c rock of which j e t t i e s a r e u s u a l l y made. However, s i m i l a r changes in t h e abundance of mussels were observed on j e t t i e s a t Murrells I n l e t , SC, by Van Dolah e t a l . (1984) , where morta l i ty was a r e s u l t of winter feeding by ruddy turnstones and g u l l s .
The i n t e r t i d a l community of inver tebrates i s r e s i l i e n t (Holling 1973); t h e genera l p a t t e r n o f zona t ion i s res tored by recruitment within a year or two a f t e r experimental c lear ing (Ortega 1981) o r predation by bi rds on mussels (Van Do1 ah e t a1 . 1984).
A1 though exper imental d a t a a r e 1 acking, the most 1 i kely fac to rs af fect ing t h e biomass and species composition of i n t e r t i d a l algal communi t i e s on j e t t i e s in t h e South At1 an t i c Bight a re desiccation during low t i d e s and grazing by f i s h e s during high t ides . Green algae in the genera u, E n t e r o m o r ~ h a , Cl adoahora, Ulothrix, Chaetomoraha, and Brvopsis and r e d a l g a e i n t h e genera Gelidium, E r v t h r o t r i c h i a , and Audouinell a a r e commonly among the more abundant seaweeds i n the i n t e r t i d a l zone (Williams 1949; Kapraun and Zechman 1982) . Feeding preference and gut content s tudies show these algae t o be preferred o r heavily used by omnivorous f i shes common t o the b i g h t . Ogburn (1984) noted t h a t sheepshead tended t o feed in t h e i n t e r t i d a l zone during periods of high t i d e and t h a t more than 70% of the algae and i n v e r t e b r a t e s in t h e i r stomachs occurred primarily in the i n t e r t i d a l zone a t her study s i t e . Hay (1986) documented some of the e f f e c t s of j e t t y f i shes on p a t t e r n s o f seaweed d i s t r i b u t i o n . P a l a t a b l e seaweeds 1 i k e Ulva and Enteromoraha were a lmost complete ly excluded from s u b t i d a l h a b i t a t s d u r i n g warm periods of the year when f i shes were common. During cold seasons when f i shes were r a r e , Ulva and Ente romor~ha were common i n subtidal areas.

I n t e r t i d a l h a b i t a t s on rubb l e s t ructures may serve as par t i a l refuges f o r p a l a t a b l e a l ga l s p e c i e s t h a t a r e seasonally eaten t o ext inct ion in subtidal habi ta ts by herbivorous f i shes (e .g , , Hay 1981c, 1984a, 1985; Hatcher and Larkum 1983) . S ince seaweed p r o d u c t i v i t y i n subtidal communities i s often much l e s s than in i n t e r t i d a l communities, t h e e f f ec t s of herbivory on seaweeds can be much greater in the subtidal zone (Hay 1981c; Hatcher and Larkum 1983). This occurs because p r o d u c t i v i t y o f t e n dec r ea se s r a p i d l y with depth due t o decreas ing l i g h t and t u rbu l ence , which provides n u t r i e n t s by breaking down d i f f u s i o n g r a d i e n t s around t h e a l g a l tha l lus . Thus, the proportion of net production l o s t t o grazers can be g rea te r in deeper than in shallower waters even i f absolute grazing r a t e s are equal. As an example, Vine (1974) and Hay (1981b) found t h a t seaweed production was 27 t o over 400 times greater a t 2 t o 3 m deep than a t 13 t o 20 m deep on t ropical ree fs . Thus, i f herbivores removed equal amounts of plant material from deep and shallow s i t e s , the e f f ec t s on the deep habi ta t plants would be greater since losses would be a l a rger portion of net growth and take longer t o replace. This pattern i s compounded by the f ac t t ha t seaweeds in deeper areas a re always a v a i l a b l e t o herbivorous f i s h e s whi le t hose in sha l lower a r e a s p e r i o d i c a l l y escape f i s h e s dur ing low t i d e s and per iods o f t u r b u l e n t s e a s . The e f f e c t s of herbivorous f i s h e s a r e d i scussed a t g r e a t e r l eng th i n t h e following section on the organization of s u n l i t , subtidal communities.
The e f f ec t s of other herbivores on i n t e r t i d a l communities have no t been studied. The most common sea urchin, Arbacia ~ u n c t u l a t a , i s very prone t o desiccation and appears t o have l i t t l e impact in in te r t ida l communities. I t i s p o s s i b l e t h a t herbivorous amphipods, i sopods, o r po lychae tes could g r aze in te r t ida l algae and avoid desiccation by shel ter ing in t he bases of algal t u r f s during low t i de . Some of these small mesograzers consume macroalgae (Glynn 1965; Martin 1966; Greze 1968; Nicotri 1977, 1980; Zimmerman e t a l . 1979; Lewis and Kensley 1982; Norton and Benson 1983; D'Antonio 1985; Gunnill 1985; Hay e t a l . 1987, 19881, but t h e i r e f f e c t s i n t h e
i n t e r t i da l zone along this coast have not been studied. Herbivorous gastropods have been shown t o have a substantial impact on i n t e r t i d a l a l g a l communities in New England and elsewhere (see the review by Hawkins and Har tno l l 19831, bu t t h e i r e f f ec t s on i n t e r t i da l hard substra tes in the bight have not been studied.
Competition has been demonstrated t o p lay a s u b s t a n t i a l r o l e i n t h e o rgan i za t i on of i n t e r t i d a l a l g a l communities in New England and on the west coast of the United States (Dayton 1971, 1975; Lubchenco 1978, 1980; and o thers ) . Descriptive s tudies of algal seasonal i t y and zonation on j e t t i e s in the bight have suggested tha t competition among seaweeds, and between seaweeds and invertebrates i s important i n determining community o rgan i za t i on in t h e i n t e r t i d a l zone (Wi 11 i ams 1949; Kapraun and Zechman 1982). However, no experimental evidence i s a v a i l abl e t o e i t h e r suppor t o r r e f u t e these contentions.
4.3 ORGANIZATION OF SUNLIT, SUBTIDAL COMMUNITIES
In t h e shal low s u b t i d a l zone, seaweeds tend t o be the dominant members of the s e s s i l e community. In the one l o c a t i o n where success ion has been studied, the algal community appeared t o be the r e su l t of a 3-year-successional process (Van Dolah e t a1 . 1984). Van Dolah and coworkers found t ha t the mussel Brachidontes exus tus dominated t h e subtidal zone in the f i r s t year a f t e r construction of the j e t t i e s a t Murrells In le t . Predation by the s t a r f i s h Asterias f o r b e s i i , and sheepshead appeared t o r e su l t in the replacement of mussels by hydroids, bryozoans, and tunicates a f t e r the f i r s t year. These groups i n t u r n were replaced by red and green algae by the t h i rd year. This successional process was not en t i r e l y predictable; i t was observed only on the north j e t t y . B . exustus dominated the subtidal zone on-the south j e t t y f o r t h e two yea r s t h a t i t was studied (Van Do1 ah e t a1 . 1984). However, B , exustus i s largely absent from the - subtidal zone of j e t t i e s in North Carolina ( J . Sutherland and M . Hay; pers. obser.) . I t i s l i ke ly t ha t mussels are l imited t o the in te r t ida l zone by predation. The

endpoint of succession appears t o be t h e brown alga , Sargassum f i l i pendul a.
No exper imenta l s t u d i e s have unambiguously demonstrated the importance of t h e v a r i o u s phys ica l f a c t o r s t h a t a f f e c t t h e o r g a n i z a t i o n of sunl i t , subtidal communities in the South At lan t i c Bight. However, the large annual changes in water temperature t h a t occur throughout t h i s region c l e a r l y have major d i r e c t and i n d i r e c t e f f e c t s on b e n t h i c community s t ruc tu re . Changes in water temperature appear t o be d i r e c t l y responsible f o r t h e l a rge - sca le migration of most f i s h e s from inshore waters in the winter and f o r t h e i r re turn i n the spring. These migrations probably have a s u b s t a n t i a l e f f e c t on energy t r a n s f e r from inshore t o offshore hab i t a t s and on inshore and offshore prey p o p u l a t i o n s . As o u t l i n e d i n p r e v i o u s chapters, temperature changes a1 so have major e f f e c t s on seaweed and inver tebra te populations. Some of these organisms must r e i n v a d e r u b b l e s t r u c t u r e s each y e a r , whi 1 e o thers have evolved mechani sms f o r "over-wintering" as r e s t ing s tages .
Wave action a1 so changes seasonal 1 y, and w i n t e r s to rms , o r l a r g e waves generated in o ther seasons, can have a s u b s t a n t i a l impact on s u b t i d a l communities. Evidence of t h i s can be seen in the l a rge mass of subtidal organisms occasionally deposited on beaches in the South At lant ic Bight. Since much of the b i g h t i s devoid of hard s u b s t r a t e i n shallow water, waves may have more impact on rubble s t ruc tu res than on natural hard s u b s t r a t e h a b i t a t s , which a r e u s u a l l y deeper. Waves a lso increase sand scour and t u r b i d i t y . Both of t h e s e f a c t o r s should s i g n i f i c a n t l y a f f e c t b e n t h i c communi t y s t ruc tu re by k i l l ing, slowing growth, o r d e c r e a s i n g r e p r o d u c t i o n o f benthic f l o r a and fauna. Sedimentation and scour might be p a r t i c u l a r l y damaging t o newly s e t t l e d juveniles.
The e f f e c t s of competition on t h e o r g a n i z a t i o n o f sunl i t , s u b t i d a l communities have ra re ly been addressed in t h e South At1 a n t i c Bight . However, experiments have been conducted in hard subs t ra te communities a t a depth of 20 m on t h e Cont inen ta l S h e l f o f f North Carol ina (Peck01 and Sear les 1983). These exper iments i n d i c a t e d t h a t seasona l
p a t t e r n s o f r e c r u i t m e n t and phys ica l disturbance in teracted with competitors and consumers t o a f f e c t the d i s t r i b u t i o n and abundance of both seaweed and benthic inver tebra te populations. When s e t t l i n g p la tes were in cages t h a t excluded large consumers, competition f o r space occurred. However, community development was dependent upon season of submergence and upon t h e seasonal growth and recruitment c h a r a c t e r i s t i c s of the species involved. In t h i s deep and often poorly l i t h a b i t a t , i t appeared t h a t barnacles would have been t h e competitive dominants had they not been s e l e c t i v e l y consumed.
Richardson (1978) conducted a s imi la r study a t a depth of 1.5 m below low t i d e level on the j e t t y a t Radio Island, NC. In h i s cages, mucous/sand-tube building p o l y c h a e t e s dominated, b i v a l v e s and s e r p u l i d po lychae tes i n c r e a s e d , and barnacles and l ea fy algae decreased in abundance compared t o t h e i r abundance on p la tes in open-sided control cages. Some of these changes were in terpre ted as being a consequence of competi t i o n on the caged p l a t e s . However, unoccupied space remained a t about 50% on p la tes i n the c l o s e d cages and i t i s doub t fu l t h a t compet i t ion caused t h e r e d u c t i o n in barnacl e s and 1 eafy a1 gae. Both amphi pods and polychaetes along t h e North Carolina coast have recent ly been demonstrated t o be capab le of consuming s i g n i f i c a n t quan t i t i e s of l a rge r organisms (Hay e t a1 . 1987, 1988). A build-up of these organisms i n t h e cages o f both of t h e above mentioned s tud ies could have s i g n i f i c a n t l y af fected t h e i r r e s u l t s (Brawley and Adey 1981a, b ) . S i n c e amphipods and polychaetes a r e common prey of many f i s h e s (Table 2 ) , t h e i r increased abundance in f i s h exclosures seems 1 i kely.
Both f i e 1 d and microcosm experiments us ing seaweeds from j e t t i e s i n North C a r o l i n a demons t ra te t h a t seaweeds i n c lose associat ion with 1 arger , overstory a1 gae 1 i ke Sarsassum, experience decreased growth r a t e s due t o competition (Hay 1986; P f i s t e r 1987). The consequences o f t h i s f o r community organization have not been adequately evaluated, Given the 1 arge r o l e of competition f o r space, l i g h t , and n u t r i e n t s i n o t h e r seaweed-dominated communities (Pearse and Hines 1979; Dayton 1975; Kastendiek 1982; Reed and Foster

1984; S a n t e l i c e s and O j e d a 1984 ) , t h e impor tance o f c o m p e t i t i o n i n s t r u c t u r i n g sunl it, b e n t h i c communit ies i n t h e South At1 a n t i c B i g h t deserves more a t t e n t i o n .
Compet i t i on among f i s h e s i n t h e South A t l a n t i c B i g h t has n o t been s t ud i ed , b u t g i v e n t h e h i g h degree o f d i e t o v e r l a p among many f i s h e s on j e t t i e s (Tab le 2) and t h e h i g h d e n s i t i e s o f t h e s e f i s h e s , compe t i t i on seems l i k e l y . I f i n t r a - s p e c i f i c c o m p e t i t i o n f o r food occurs, t hen t h e growth o f immature f i s h e s should be d e n s i t y dependent. T h i s p a t t e r n has been documented on severa l occas ions and i s widespread i n mar ine f i s h e s (Anthony 1971; Cushing and Horwood 1977; Legge t t 1977; Rauck and Z i j l s t r a 1978; Jones 1984a, b ) . However, t h e impor tance o f i n t e r s p e c i f i c compe t i t i on i n j u v e n i l e s tages o f temperate r e e f f i s h e s has n o t been i n v e s t i g a t e d .
F o r t h e many j u v e n i l e s t h a t u s e r u b b l e s t r u c t u r e s and e s t u a r i e s as n u r s e r i e s , c o m p e t i t i o n f r o m abundan t omn i vo rous f i s h e s , 1 i k e p i n f i s h and s p o t t a i l p i n f i s h , c o u l d be p a r t i c u l a r l y acute. I n sha l low h a b i t a t s a1 ong p o r t i o n s o f t h e b i g h t , p i n f i s h and s p o t t a i l p i n f i s h may comprise more t han 50% o f t h e t o t a l f i s h s t and ing s t o c k d u r i n g t h e summer and f a l l . T h i s i s a l s o t h e t i m e when f e e d i n g by f i s h e s has reduced t h e abundance o f ep i f auna l p r e y t o y e a r l y lows (Thayer e t a l . 1975; Adams 1976a; Nelson 1979, 1980a, b; Darcy 1985a, b ) . Because t h e d i e t s o f t hese s p a r i d f i s h e s o v e r l a p s u b s t a n t i a l l y w i t h t h e d i e t s o f j u v e n i l e gag, spot , b l a c k sea bass, and o t h e r s (Adams 1976a; L i n k 19801, c o m p e t i t i o n be tween t h e s e spec ies and spa r i ds c o u l d be p a r t i c u l a r i l y impo r t an t .
One advantage t h a t spa r i ds may have i s t h e a b i l i t y t o feed on p l a n t m a t e r i a l when c r u s t a c e a n p o p u l a t i o n s have been dep le ted . T h i s may a l l o w them t o m a i n t a i n h i g h d e n s i t i e s t h a t p reven t t h e r ecove ry o f crustaceans and t h e r e f o r e make t h e a rea o f m a r g i n a l v a l u e f o r o t h e r j u v e n i l e f i s h e s . Jones (1984a, b) p rov i des severa l l i n e s o f ev idence sugges t ing t h a t j u v e n i l e temperate f i s h e s migh t be 1 i m i t e d by t h e abundance o f e p i f a u n a l p rey .
Tempera te i n v e s t i g a t i o n s o f how h e r b i v o r o u s i n v e r t e b r a t e s a f f e c t a l g a l
d i s t r i b u t i o n and t h e o r g a n i z a t i o n o f b e n t h i c communit ies i n genera l , have been remarkably f r u i t f u l . They have p rov i ded b o t h ideas and a da ta base f o r many o f t h e genera l i z a t i o n s i n t h e c u r r e n t e c o l o g i c a l l i t e r a t u r e (Dayton 1971, 1975; Menge and S u t h e r l a n d 1976; Lubchenco 1978; Sousa 1979; Paine 1980; Lubchenco and Gaines 1981; Gaines and Lubchenco 1982). The e f f e c t s o f g r a z i n g i n v e r t e b r a t e s on t h e o r g a n i z a t i o n o f s u n l i t , s u b t i d a l communit ies i n t h e South A t l a n t i c B i g h t , however, a re 1 a r g e l y un i nves t i ga ted . Bo th Richardson (1978) and Peck01 and Sear les ( 1983 ) d e m o n s t r a t e d t h a t e x c l u s i o n o f 1 a rge consumers cou ld s i g n i f i c a n t l y a f f e c t b e n t h i c p r e y popu la t i ons . However, t h e i r exper iments d i d n o t separa te t h e e f f e c t s o f t h e i n v e r t e b r a t e s ( u r c h i n s , l a r g e crabs, e t c . ) f r om those o f t h e f i shes , so t h e e f f e c t s o f i n v e r t e b r a t e s a lone a r e unknown.
The mos t o b v i o u s i n v e r t e b r a t e he rb i vo res on j e t t i e s i n t h e b i g h t a re sea u r c h i n s . No f i e l d exper iments have been c o n d u c t e d t o assess t h e i r e f f e c t s on community s t r u c t u r e . Hay e t a l . (1986) p resen t some d a t a on t h e j e t t y seaweeds t h a t a re p r e f e r r e d and avoided by t h e common sea u r c h i n Arbac ia ounc tu l a ta , and on i t s c h e m o a t t r a c t i o n t o w a r d t h e s e seaweeds. Some l ow p re fe rence seaweeds l i k e t h e brown a l ga D i c t v o t a dichotoma a r e chem ica l l y defended a g a i n s t A rbac i a (Hay e t a1 . 1987). P f i s t e r (1987) has a1 so demonstrated t h a t pa l a t a b l e seaweeds g a i n some p r o t e c t i o n f rom g r a z i n g A rbac i a b y b e i n g c l o s e l y a s s o c i a t e d w i t h unpa la tab l e seaweeds 1 i k e t h e brown a l g a Sarsassum f i 1 i oendul a. The consequences o f t h e s e t y p e s o f i n t e r a c t i o n s a r e d iscussed a t g r e a t e r l e n g t h i n t h e s e c t i o n on complex i n t e r a c t i o n s . G i v e n t h e s i g n i f i c a n t e f f e c t s t h a t sea u r c h i n s have been shown t o have i n o t h e r b e n t h i c mar ine communit ies (Lawrence and Sammarco 1982), i t i s p robab le t h a t sea u r ch i ns , when abundan t , s i g n i f i c a n t l y a f f e c t t h e o r g a n i z a t i o n o f c o m m u n i t i e s on r u b b l e s t r u c t u r e s i n t h e b i g h t .
O t h e r common h e r b i v o r o u s i n v e r t e - b r a t e s i n c l u d e amphi pods, i sopods, c rabs , gast ropods, and po l ychaetes . These can be i m p o r t a n t h e r b i v o r e s i n some h a b i t a t s (Steneck and Wa t l i ng 1982; Hawkins and H a r t n o l l 1983) b u t l i t t l e i s known o f

t h e i r e f f e c t s on s u b t i d a l seaweeds i n t h e South A t l a n t i c B i g h t . S tud ies o f some o f these sma l l e r he rb i vo res show t h a t t h e y a re r a r e l y resource 1 i m i t e d (Zimmerman e t a l . 1979; Stoner 1 9 8 0 ~ ) b u t a r e o f t e n s t r o n g l y a f f e c t e d by t h e i r p r e d a t o r s (Young e t a l . 1976; Young and Young 1978; Nelson 1979, 1980a, b, 1981; Stoner 1980a, b; Brawley and Adey 1981a, b; Edgar 1983). They appear t o have o n l y l i m i t e d impact on mos t seaweeds ( C a r p e n t e r 1986) because t h e i r p reda to r s u s u a l l y keep them w e l l be1 ow c a r r y i n g c a p a c i t y . However, t h e i r p o t e n t i a l impact i s g r e a t . Brawley and Adey (1981a) demonstrated t h a t amphipods c o u l d have a l a r g e e f f e c t on r e c e n t l y e s t a b l i s h e d a l g a l c o m m u n i t i e s , and P. Dayton and M. Tegner (Sc r i pps I n s t i t u t i o n o f Oceanography; p e r s . comm.) h a v e r e c e n t l y o b s e r v e d g i a n t k e l p p l a n t s ( M a c r o c v s t i s ) on t h e wes t c o a s t b e i n g c o m p l e t e l y consumed b y amph ipods when amphi pod-consuming f i s h e s were m i s s i n g f r o m n e a r s h o r e c o m m u n i t i e s because o f even ts r e l a t e d t o t h e E l Nif io phenomenon.
Amphipods and po lychae tes can a l s o s i g n i f i c a n t l y damage seaweeds t h a t a r e ve r y r e s i s t a n t t o f i s h g raz i ng . Hav and coworkers (Hay e t a l . 1987, 1988; Paul e t a l . 1987) have r e c e n t l y shown t h a t seaweeds avoided by omnivorous f i s h e s a r e o f t e n se1 e c t i v e l y consumed by amphi pods and po lychae tes (F i gu re 27) and t h a t t h e seaweed seconda ry m e t a b o l i t e s t h a t e f f e c t i v e l y d e t e r f eed ing by f i s h e s o f t e n do n o t a f f e c t , o r may even s t i m u l a t e , f e e d i n g b y amphipods and p o l y c h a e t e s . They suggest t h e f o l l o w i n g reasons f o r t h e e v o l u t i o n o f t h i s p a t t e r n . Because smal l , r e1 a t i v e l y sedentary he rb i vo res 1 i ke t ube - b u i l d i n g amphipods and po lychae tes l i v e on t h e p l a n t s t hey consume, t h e y shou ld v iew p l a n t s as bo th foods and l i v i n g s i t e s . S ince 1 arge, mob i l e he rb i vo res l i ke f i s h e s commonly move among, and feed on, many p l a n t s , t h e y should v iew p l a n t s p r i m a r i l y as foods and r a r e l y as p o t e n t i a l l i v i n g s i t e s . I n t h e South A t l a n t i c B i gh t , where f i s h e s t h a t consume p l a n t s a r e a l s o i m p o r t a n t p r e d a t o r s on amph ipods and po lychae tes (Tab1 e 21, seaweeds avoided by f i shes shou ld r ep resen t s a f e r l i v i n g s i t e s f o r s m a l l g r a z e r s . Thus, s m a l l , r e l a t i v e l y s e d e n t a r y h e r b i v o r e s s h o u l d evo l ve a p re fe rence f o r seaweeds t h a t a r e w e l l defended a g a i n s t f i s h e s because if they a re l i v i n g on unpa la tab l e seaweeds
- 200.7 LAGODON RHOMBOfDES
160- - 5 120-
3 80- W
40- 3 u 0-
- 700-
i .
50- AMPHIPODS , \ > ' - ,+:H;, ; 2 $&> 1 2.g +KXY
48, 1.; 4% J %<
0
5 560- Z
2 420- u
280- W u (1? 140- u
oa
Figure 27. Feeding preferences of two smnivorous fishes and two invertebrate grazers common in the South Atlantic Bight. Lines through the top of each histogram represent il standard error (data from Hay et al. 1987,1988; M. Hay unpubl.).
DIPLODUS HOL BROOK1
N=8
t h e y should exper ience l e s s p r e d a t i o n t han i f t hey a re l i v i n g on seaweeds p r e f e r r e d b y f i s h e s . T h e r e a r e now s e v e r a l documented cases o f South A t l a n t i c B i g h t o r C a r i b b e a n amphipods and p o l y c h a e t e s

being r e s i s t a n t t o seaweed chemical defenses t h a t de te r co-occurring f i shes . However, t h e r e a r e s t i l l no d a t a t o suggest t ha t these small grazers sequester the algal metabol i t e s and thereby d i r ec t l y reduce t h e i r a ccep t ab i l i t y a s prey t o local f ishes (Hay e t a l . 1987). The e f f e c t s of small g r a z e r s on a l g a l community s t r u c t u r e c l e a r l y wa r r an t s increased a t t en t ion .
Given t h e l a r g e impact t h a t herbivorous invertebrates have been shown t o have on temperate algal communities, i t i s surpr is ing t ha t more a t tent ion has not been focused on the e f f ec t s of temperate herbivorous f i s h e s . These f i s h e s a r e abundant i n t h e South A t l a n t i c Bight (F igu re 2 6 ) , a r e very mobile, sea rch visual ly , and have high metabolic r a t e s r e l a t i v e t o co-occur r ing i n v e r t e b r a t e he rb ivo re s . I t would be s u r p r i s i n g therefore i f they did n o t have a large impact on the organization of subtidal community s t r u c t u r e . Choat (1982) recently reviewed feeding by f i shes in temperate wate rs and i t s e f f e c t s on benthic communi t y s t ructure . He compiled an impressive l i s t of s tudies tha t suggest tha t temperate herbivorous f i shes have the p o t e n t i a l t o a f f e c t ben th i c community s t ructure . However, he concluded t ha t no s tudies had c lea r ly demonstrated extensive modification of temperate, hard-substrate biota by grazing f i shes .
The following famil i e s of herbivorous f i shes occur in the South Atlantic Bight: Sparidae, Bl eni idae, Kyphosidae, Mono- canthidae, Mugilidae, and Pomacentridae. Of these, the Sparidae (pinf ish , spo t ta i l pinfi sh, and sheepshead) probably have the greates t impact on the community organ- ization of rubble s t ructures because they are often the most abundant f ishes in shall ow waters (Adams 1976b; Darcy 1985a, b; Hay 1986). They also consume large quant i t ies of benthic macrophytes (Carr and Adams 1973; Adams 1976a; Ogburn 1984; Darcy li985a, b; Lindquist e t a l . 1985). As an example, Table 4 shows the stomach contents of 21 spo t ta i l pinfish collected from the j e t t y a t Radio Island, NC during l a t e summer. Approximately 98% of the dry mass of stomach contents was plant material . Carr and Adams (1973) reported s imilar data fo r several s i z e c lasses of spo t ta i l pinfish ranging from 26 to 167 mm; 50% t o
90% of the volume of t h e i r stomach contents was algae. When offered a variety of common macrophytes in the lab, both pinfish and spo t ta i l pinfish showed a strong preference fo r some species and consumed very l i t t l e of others (Hay e t a l . 1987, 1988; and Figure 2 7 ) . During mid- t o l a t e - summer, high preference species such as Hyonea and Caloni toohvl lum show dramatic declines in abundance as f i s h numbers and s izes increase. Low preference species such as Sarqassum, Padina, and Dictyota increase or show no change in abundance a t t h i s time ( M . Hay; pers. obser. ) .
To assess the potential e f fec t s of temperate herbivorous f i s h e s on t h e o rgan i za t i on of sub t i da l j e t t y communities, Hay (1986 and work i n p rog re s s ) cons t ruc t ed e i g h t 5,000-L outdoor microcosms t h a t were designed t o mimic the nearby j e t t y a t Radio Island, NC. Each microcosm was divided by 1.5 cm pla3t ic mesh in to two equal sized par ts of 1 m each. The mean wet mass/m2 of a l l common algae from the j e t t y was attached t o the bottom and s ides of each pa r t i t i on , and the mean f i e l d density of each common herbivorous or omnivorous f i sh was placed in one po r t i on of each microcosm. Spot ta ' l pinfish were stocked a t a density
?? of 8/m ; pinfish and planehead f i l e f i s h , Mon~canthus h i s o i d u s , were stocked a t l/mL. Five new cinder blocks were added t o each side of each microcosm and were monitored a t 2- to 4-week intervals f o r presence and percent cover of a l l animal and plant species. The same cinder blocks were monitored without rep1 acement fo r the e n t i r e 4 month study . All microcosms were located immediately adjacent t o Bogue Sound and received continuous inputs of u n f i l t e r e d sound water through an apparatus which also generated waves. The water in the microcosms turned over every 45-90 minutes.
Between t h e i n i t i a t i o n of t h e experiment in ea r ly August and termination in l a t e November, f i s h e s : ( 1 ) s ign i f ican t ly reduced the abundance and r a t e o f es tab l i shment of u, Enteromoroha, small filamentous algae, and a l l of the common red seaweeds ( H ~ ~ n e a , Sovridia, Chondria, Charnoia, Polysiohonia, G r a c i l a r i a , and Neoasardhiel l a ) , ( 2 ) s ign i f ican t ly increased the abundance of


shallow- water ha rd - subs t r a t e communities (Ogburn 1984; Darcy 1985a, b; Hay 1986; Hay e t a l . 1987, 1988).
4.4 ORGANIZATION OF SHADED, SUBTIDAL COMMUNITIES
Organisms growing on shaded h a r d s u b s t r a t e s have long been regarded a s a source of t r o u b l e s i n c e they a1 so grow on t h e bo t tom o f b o a t s and must be p e r i o d i c a l l y scraped o f f . As a r e s u l t t h i s assemblage i s o f t en r e f e r r e d t o a s t h e f o u l i n g communi t y . Low l i g h t avai 1 abi 1 i t y g e n e r a l l y 1 imi t s t h e growth of m a c r o a l g a e on t h e s e s u b s t r a t e s , a l lowing s e s s i l e animals t o dominate.
The s t a r t i n g po in t o f community development on unoccupied s u b s t r a t e i s t h e recru i tment o f l a r v a e t o t h a t s u b s t r a t e . This process i s extremely unpredic tab le ; d i f f e r e n t p a t t e r n s of i n i t i a l development a r e p o s s i b l e from month t o month and from yea r t o yea r (Mook 1976, 1980; Suther land and Karlson 1977). Ins tead o f p repar ing t h e way f o r s u b s e q u e n t a r r i v a l s , most r e s i d e n t a d u l t s s t r o n g l y i n h i b i t t h e recru i tment and growth of o t h e r s p e c i e s ( S u t h e r l and 1974, 1977 , 1978 , 1981; S u t h e r l and and Kar l son 1 9 7 7 ) . T h i s p a t t e r n of development appears t o conform t o what Connell and S l a y t e r (1977) have termed t h e i n h i b i t i o n model of success ion . Spec ies vary in t h e i r a b i l i t y t o r e s i s t subsequent invas ion and l a r v a e vary i n t h e i r a b i l i t y t o invade assemblages of a d u l t o r g a n i s m s . As a r e s u l t , t h e d i r e c t i o n and r a t e o f community development a r e dependent on t h e o rde r of invas ion and a r e d i f f i c u l t t o p r e d i c t .
The endpoint o f community development depends on l o c a t i o n and, a t t imes , on t h e pe r spec t i ve of t h e observer . Su ther l and and Karlson (1977) have argued t h a t near Beaufor t , N C , community composition never s t o p s c h a n g i n g and t h a t no c l i m a x community i s p r e s e n t . As pointed o u t in p r e v i o u s s e c t i o n s , t h e w i n t e r s p e c i e s assembl age is extremely v a r i a b l e from yea r t o y e a r . However, Sutherland (1981) has a l s o a rgued t h a t one e n d p o i n t i s a community domina t ed by t h e s o l i t a r y t u n i c a t e S tye l a p l i c a t a . T h i s s p e c i e s p r ed i c t ab ly dominates summer assemblages,
i n h i b i t s recru i tment by o t h e r spec i e s when p re sen t , and re invades in sp r ing a f t e r sloughing o f f t h e previous f a l l . This i s a n a l o g o u s t o t h e mussel communi t i e s s t ud i ed by Paine (1966, 1974) and Menge ( 1 9 7 6 ) , where p a t c h e s o f m u s s e l s a r e removed by a v a r i e t y of d i s t u rbances , but even tua l l y re invade . Thus, whether o r no t a "climax" i s p r e sen t depends on which period i s chosen as a r e f e r ence po in t and t he l eng th of t h e observa t ion per iod .
O t h e r s h a l low w a t e r f o u l i n g communi t ies n e a r B e a u f o r t i n Nor th C a r o l i n a " t e r m i n a t e " a t d i f f e r e n t endpoin ts . In t h e i r s t u d i e s , Su ther land and Karlson worked p r imar i l y a t t h e dock of t h e Duke Univers i ty Marine Laboratory and t h e nearby p i l i n g s of t h e r a i l r o a d b r i d g e a c r o s s t h e B e a u f o r t c h a n n e l . P i l i n g s near t h e A t l a n t i c Beach br idge i n Bogue Sound a r e dominated by t h e co lon i a l t u n i c a t e b l i d i u m c o n s t e l l a t u m , which apparen t ly can maintain t h i s compet i t ive dominance f o r long per iods o f t ime. Wells e t a1 . (1964) found t h e f o u l i n g community a t Cape Ha t t e r a s t o be dominated by t h e co lon i a l t u n i c a t e Botrvl l u s sch l o s s e r i and va r ious spec i e s of sponges.
Near Cape Canave ra l i n F l o r i d a , community deve l opment i n s h a l l ow w a t e r ended i n a s s e m b l a g e s domina ted by t h e t ub i cu lous amphi pod Coroohium l a c u s t r a and s eve ra l spec i e s of Balanus, i n s p i t e o f d i f f e r e n c e s in i n i t i a l development (Mook 1 9 8 1 ) . The F l o r i d a a s s e m b l a g e was p e r s i s t e n t , showing few annual changes i n spec i e s composition.
On p i1 ings in deeper waters (>2m) near Beaufort , NC, t h e foul ing community i s dominated by long- l ived forms such a s t h e h y d r o i d H v d r a c t i n i a e c h i n a t a , t h e sponge X e s t o s ~ o n s i a ha l ichondro ides and t h e anemone D i adumene 1 eucol ena (Karl son 1978). These spec i e s a r e r e s i s t a n t t o g r a z i n g by t h e s e a u r c h i n A r b a c i a p u n c t u l a t a , which removes o t h e r l e s s r e s i s t a n t forms. Grazer r e s i s t a n t forms tend t o r e c r u i t a t very low i n t e n s i t i e s , but g r adua l ly come t o dominate through v e g e t a t i v e growth ( o r b inary f i s s i o n i n anemones) . I n t h e p r e s e n c e o f A. p u n c t u l a t a , t h e s e g r a z e r r e s i s t a n t e n d p o i n t s would p r e sumab ly be o b s e r v e d r e g a r d l e s s of d i f f e r e n c e s i n i n i t i a l deve lopmen t . I n d e e d , Kar l son (1978 )

documented enhanced r ec ru i tmen t and vegetative growth of H, echinata in the presence of A, punctul a t a .
A nor th -sou th g r a d i e n t i n t h e in tensi ty of f i s h predation on fouling organisms may be present. Near Beaufort, f i s h a r e only occa s iona l l y important predators and have l i t t l e t o do with the eventual endpoint of community deve'l opment (Sutherland 1974). Fish can remove small i n d i v i d u a l s of t he t u n i c a t e Stvel a p l i c a t a , when they s e t t l e on open substra te . However, juveniles of t h i s tunicate commonly find refuges a t the base of e r e c t co lon i e s of hydroids and bryozoans, and adul ts predictably dominate summer assemblages. Near Cape Canaveral , S. p l i c a t a i s predictably removed from - shall ow water assembl ages by sheepshead. This t u n i c a t e dominates only when s u b s t r a t e s a r e experimental l y i so l a t ed from f i s h predators (Mook 1981).
4.5 COMPLEX INTERACTIONS
Experimental marine ecol ogi s t s have been impress ive ly success fu l i n document i ng how competition, predation, and physical disturbances a f fec t community s t ructure (Pai ne 1966; Dayton 1971, 1975; Su ther l and 1974; Connell 1975, 1978; Lubchenco 1978; Sousa 1979; Ayling 1981; Hay 1981b; Hixon and Bros tof f 1983; Dethier 1984). The best of these s tudies have a1 so investigated the in teract ions among the various fac to rs . However, the obvious success achieved by studies focusing p r ima r i l y on compet i t ion , predation, o r physical disturbances may have caused ecologists t o overlook the importance of more complex, and o f t en i n d i r e c t , i n t e r a c t i o n s . These i n t e r a c t i o n s can be counte r i n t u i t i v e ( e . one compet i tor i s dependent o n another), and thus are easy t o overlook. However, in some cases they may have a major impact on how communities function (Dethier and Duggins 1984; Hay 1986). Two examples of complex interact ions t ha t do, o r may, occur on j e t t i e s in the South Atlant ic Bight are described here.
On j e t t i e s in the bight, palatable seaweeds can gain s ign i f ican t protection from herbivorous f i s h e s by a s s o c i a t i n g with abundant competitors t h a t are l e s s
palatable t o these f i shes . In f a c t , when herbivorous f i shes are present, pala table seaweeds a r e completely dependent upon t h e i r unpalatable competi t o r s t o provide mi c r o s i t e s of reduced herb ivory t h a t prevent f i shes from causing t h e i r local e x t i n c t i o n . When f i s h e s a r e excluded, however, t h e growth r a t e of p a l a t a b l e species can be severely decreased (by more than 80%) by t h e i r a s s o c i a t i o n wi th unpalatable ones (Hay 1986). For these palatable seaweeds, the costs of being associated with an unpalatable competitor are much l e s s than the costs of increased consumption i n t h e absence of t h a t competitor. For the North Carolina j e t t y community where t h i s i n t e r a c t i o n was studied, i t appeared t h a t removing the dominant (unpal atabl e ) seaweed competitor from the system would cause a decrease, instead of an increase, in the abundance of co-occurring (pal atabl e ) competitors (Hay 1986) . More recent investigations ( P f i s t e r 1987) have shown t h a t t h e s e unexpected interact ions between compet i ng seaweeds have s i m i l a r e f f e c t s on both foraging f i shes and sea urchins.
Although t h e r e a r e no r i go rous s tudies of the recruitment of juvenile f i shes t o j e t t i e s o r reefs in the South Atlant ic Bight, a New Zealand study (Jones 1984a, b ) may be i n s t r u c t i v e f o r i t s information on the ecology of a temperate reef f i sh and fo r i t s i l l u s t r a t i on of complex ecological in teract ions . Jones' s t udy , a s well a s ex t ens ive work on tropical ree fs , suggests t h a t the spa t ia l and temporal changes in d i s t r ibu t ion and abundance s f many species r e su l t primarily from p a t t e r n s of j u v e n i l e r e c ru i tmen t (Sale 1980; Williams 1980; Williams and Sale 1981; Doherty 1982, 1983a, b). Jones (1984a, b) showed experimental l y t h a t seaweed abundance was c r i t i c a l l y important in t h e rec ru i tment of j u v e n i l e f i s h because i t provided both cover and food in the form of epi fauna? crustaceans. When rec ru i tment was monitored over a wide range o f r e e f h a b i t a t s , j u v e n i l e recruitment a t a s i t e was shown t o be s i g n i f i c a n t l y c o r r e l a t e d with a l g a l abundance. Addi t iona? ly , when seaweeds were removed from some ree f a r e a s , recruitment on those areas decreased by 87% compared with nearby caa t ro l s , When a l g a l abundance was exper imenta l ly inc reased by removing herbivorous s ea

urch ins , rec ru i tmen t o f j u v e n i l e f i s h e s increased approximately s i x f o l d .
The p o t e n t i a l i n t e r a c t i o n s between h e r b i v o r o u s sea u r c h i n s , seaweeds, and j u v e n i l e r e e f f i s h e s may be o f p a r t i c u l a r i n t e r e s t i n h e a v i l y f i s h e d areas o f t he South A t l a n t i c B igh t s ince i t appears t h a t u rch ins occur i n unusua l ly h igh abundance p r i m a r i l y i n areas t h a t have been h e a v i l y f i s h e d by p e o p l e (Es tes and Palmisano 1974; Estes e t a l . 1978; Simenstad e t a1. 1978; Hay 1984b). On some r e e f s i n the b i g h t u rch ins may occur a t d e n s i t i e s o f >30/rn2. On j e t t i e s near Beaufor t , N47 u rch in dens i t y ranges from t l t o 10/m . Predatory f i s h e s on temperate and t r o p i c a l r e e f s have been shown t o a f f e c t sea u r c h i n
d i s t r i b u t i o n , s i z e f requency , and abundance (Tegner and Dayton 1977, 1981; Bernste in e t a l . 1981; Cowen 1983; Hoffman and Robertson 1983), as we l l as behaviora l pa t te rns , f o rag ing range, and d i e t breadth (Nelson and Vance 1979; Vance and Schmit t 1979; Carpenter 1984). Studies from both the east and west coasts o f North America, as w e l l as the Caribbean, have s t r o n g l y suggested t h a t human removal o f u rch in predators i n d i r e c t l y r e s u l t s i n unusual ly h igh u rch in dens i t i es and thus the l o s s o f a l g a l c o v e r upon wh ich many o t h e r organisms may depend (Estes and Palmi sano 1974; Breen and Mann 1976; Estes e t a l . 1978; Simenstad e t a l . 1978; Hay 1984b). This could r e s u l t i n 1 ongterm suppression o f some r e e f f i shes .

CHAPTER 5. MANAGEMENT CONSlDERATlONS
Rubble s t ructures are constructed as part of a management s t ra tegy t o slow coastal erosion and/or in1 e t migration. They a r e designed t o so lve a l oca l problem, but they almost always have broader impacts on t h e c o a s t a l environment. These impacts are the major focus of t h i s chapter. In the At lant ic Bigh t , rubb le s t r u c t u r e s r e p r e s e n t a unique h a b i t a t i n o therwise d i s t i n c t l y d i f fe ren t surroundings. They can a t t r a c t large numbers of f i shes , but on a regional scale , rubble s t ruc tures have very 1 i t t l e impact on f i sh and wi ld l i fe population s i z e s o r d i s t r i b u t i o n s . There a r e , however, some very localized benefits of j e t t i e s t o people who f i sh and t o other r e c r e a t i o n a l e n t h u s i a s t s . These a r e discussed a t the end of the chapter.
5.1 SHORELINE EVOLUTION
In t h e South A t l a n t i c Bight most rubble s t ructures are ins ta l 1 ed on bar r ie r
islands. To understand the e f f ec t s t h a t j e t t i e s and groins have on these islands i t i s necessa ry t o unders tand t h e dynamics of the in teract ion between the land and the sea .
The fac t of overriding importance i s t h a t the level of the sea i s r i s ing . Some 15,000-18,000 years ago ( a t the end of the l a s t ice age) sea level was as much as 100 m lower than a t present because of the amount of water t i ed u p in glacia l icecaps (Figure 28). As the g lac ie r s melted, sea level rose qui te rapidly unt i l about 5,000 years ago. Since then, although the r a t e of r i s e has slowed, i t continues a t about 0.3 m a century (Pilkey e t a l . 1980). Exper ts expect t h i s r a t e of r i s e may a c c e l e r a t e . The National Academy of Science has warned tha t the burning of f o s s i l fue l s and other a c t i v i t i e s have resulted in the presence of extra carbon dioxide and other "greenhouse gases" in the atmosphere. The resul t ing "greenhouse e f fec t " causes the atmosphere t o re ta in
P O 2
5
E ,I wzs
m C1
3 - 17C00 years ago !f
r '? m_
150
Figure 28. The sea-level rise during the past 17,000 years (from Piikey et at. 1984).
47

hea t , which i nc rea se s t h e melt-ing o f t h e po l a r i c e caps and r a i s e s sea 1 eve1 .
As s e a l e v e l r i s e s , t h e b a r r i e r i s l a n d - s o u n d sys t em m i g r a t e s up t h e Continental She l f . This process has been ope ra t i ng f o r t h e l a s t s eve ra l thousand y e a r s and cont inues today. The f r o n t s i d e o f an i s 1 and i s moved backwards by e ro s ion . The back s i d e of t h e i s l a n d grows by two processes (Neal e t a l . 1984). Storm dr iven waves can c a r r y sand from t h e beach ac ros s t h e i s l and t o t h e back s i d e i n a process known a s overwash (Godfrey and Godfrey 1973; Knutson and Finkel s t e i n 1987). In essence , i s l a n d s migra te by r o l l i n g over on themselves l i k e a tank t r e a d . S e c o n d l y , s t o r m s o f t e n form i n l e t s , new passageways between t h e sound and t h e ocean. Tidal c u r r e n t s o f t en c a r r y l a r g e q u a n t i t i e s of sand through t h e s e i n l e t s i n t o t h e sound, forming a t i d a l d e l t a much t h i c k e r t h a n t h e o r i g i n a l i s l a n d . Longshore c u r r e n t s cause t h e s e i n l e t s t o migra te , and a s they do t h e i s l a n d i s th ickened over t h e d i s t a n c e of t h e migra t ion . This process cont inues a s Song a s t h e i n l e t i s open. Overwash a r e a s and t i d a l d e l t a s become s t a b i l i z e d with vege ta t ion and a new sound-side border t o t h e i s l and i s formed.
The d i s t a n c e a b a r r i e r i s l and migra tes wi th a given r i s e i n s ea l eve l i s a func t ion o f the s lope o f the Continental She l f . In much of t h e South A t l a n t i c Bight t h e s l o p e i s so gradual t h a t a 0 . 3 m r i s e i n s e a l eve l produces a landward m i g r a t i o n o f t h e b a r r i e r i s l and-sound system of from 30 t o 300 m (P i lkey e t a l . 1975, 1980). This means t h a t c u r r e n t r a t e s of s e a l eve l r i s e t r a n s l a t e i n t o a 1 andward migra t ion of 0.3 t o 30 m pe r y e a r . T h i s m i g r a t i o n i s what beach eng inee r s c a l l e ro s ion .
The s i z e and shape o f b a r r i e r i s l a n d s depend on t h e ref a t i v e magnitude of t i d a l and wave ene rg i e s (Nummedal e t a l . 1977). The d i f f e r e n c e i n t h e forms of a t i d e - dominated and a wave-dominated c o a s t l i n e r e f l e c t s t h e a b i l i t y of t h e t i d a l c u r r e n t s t o t r a n s p o r t s e d i m e n t s t h r o u g h i n l e t s , v e r s u s t h e a b i 1 i t y o f w a v e - g e n e r a t e d langshore c u r r e n t s t o t r a n s p o r t sediments along t h e c o a s t . Along wave-dominated c o a s t s , t h e l o n g s h o r e c u r r e n t s p roduce l o n g , c o n t i n u o u s b a r r i e r i s l a n d s w i t h
sma l l e b b - t i d a l d e l t a s ( s a n d b o d i e s seaward of i n l e t s ) because waves r a p i d l y d i s p e r s e t h e sediments . Sediments c a r r i e d landward i n t o i n l e t s by t i d a l c u r r e n t s accumulate i n 1 a rge back -ba r r i e r f lood- t i d a l d e l t a s b e c a u s e t h e s e a r e a s a r e she1 t e r e d from wave d i spe r sa l . With an i n c r e a s e i n t i d a l r a n g e a l o n g t i d e - domina ted c o a s t s , t h e t i d a l c u r r e n t s through t h e i n l e t s i n c r e a s e in s t r e n g t h . Consequently, they can suppor t l a r g e r ebb- t i d a l d e l t a s a g a i n s t t h e d e s t r u c t i v e i n f l uence of t h e waves.
In t h e South A t l a n t i c Bight , b a r r i e r i s l a n d s along t h e North Caro l ina c o a s t t y p i f y t h e wave-dominated c o a s t l i ne . The mean t i d e range i s on ly 0 .9-1 .2 m (F igure 4 ) . The i s 1 ands a r e l o n g , g e n e r a l l y narrow, and cu t by widely s epa ra t ed t i d a l i n l e t s w i t h l a r g e f l o o d - t i d a l d e l t a s . They a r e low i n e l eva t i on and f r e q u e n t l y overwashed (Neal e t a1 . 1984). I s l ands a l o n g t h e s o u t h e r n South C a r o l i n a and Georg i a c o a s t s t y p i f y a t i d e - d o m i n a t e d c o a s t l i n e . Here t h e t i d a l range i s 1 .5- 2 . 2 m ( F i g u r e 4 ) . The i s l a n d s a r e r e l a t i v e l y s h o r t and s t u b b y and a r e separa ted by s t a b l e t i d a l i n l e t s . Large ebb - t i da l d e l t a s a r e a s soc i a t ed with a l l i n l e t s . The i s l a n d s u s u a l l y have a we l l - developed row of sand dunes p a r a l l e l t o t h e beach t h a t i s s u f f i c i e n t t o block overwash (Neal e t a l . 1984).
In t h e South A t l a n t i c Bight , ocean waves g e n e r a l l y h i t t h e c o a s t a t an angle which p r o d u c e s l o n g s h o r e c u r r e n t s f rom nor th t o south . This i s t h e d i r e c t i o n i n which sand and i n l e t s migra te , e s p e c i a l l y a1 ong wave-dominated c o a s t l i n e s . However, t h e s e waves can be r e f r a c t e d by l a r g e ebb- t i d a l d e l t a s , producing south t o nor th c u r r e n t s j u s t south o f t h e i n l e t . Ebb- t i d a l d e l t a s may a1 s o produce a wave dead zone j u s t south of t h e i n l e t . I s l ands i n Sou th C a r o l i n a and Georg i a t h a t a r e s u f f i c i e n t l y long have t h e shape of a drumstick a s sand i s l o s t ( t o t h e ebb- t i d a l d e l t a o f t he next i n l e t ) a t t h e south end and sand accumulates a t t h e no r th . In c o n t r a s t , i s l a n d s i n North Carol ina a r e of s i m i l a r width throughout .
For a g e o l o g i s t , beaches extend from t h e base of t h e f i r s t row of dunes t o a d e p t h o f 10 -15 rn o f f s h o r e . What we u s u a l l y walk on i s on ly t h e upper beach.

Beaches are extremely dynamic systems. We have already seen how they respond t o r i s e s in sea level and how t h e i r s i ze and form i s determined by t h e r e l a t i v e importance of wave and t ida l energy. They also respond predictably to the increase in wave energy produced by storms (Figure 29) . During a storm, waves take sand From the upper beach or the f i r s t dune and transport i t t o the lower beach. The beach becomes more f la t tened and storm waves expend t h e i r energy over a broader and more level surface. The upper beach can lose a great deal of sand during a storm. Much of i t i s r ep l en i shed , however, during f a i r weather. Sand i s pushed shoreward by fair-weather waves or carr ied in by long-shore t ransport . The source of sand a f t e r storms i s the same sand tha t was on the upper beach pr ior t o the storm.
First dune
Beach
5.2 SHORELINE ENGINEERING
We have seen t ha t ba r r ie r islands i n the South At1 an t ic Bight are extremely dynamic systems. They migrate landward as sea level r i s e s , are moulded by waves and t i de s , and respond in predictable ways t o storms. All of these responses involve the transport of enormous quant i t ies of sand. The engineer's response t o t h i s movement, labelled erosion, i s t o t r y t o stop i t and " s t ab i l i z e " the shore. The most common method i s with rubb l e s t ructures : j e t t i e s , groins and seawalls.
Both g r o i n s and j e t t i e s are success fu l sand t r a p s . I f longshore t ransport of sand i s s ign i f ican t , sand will p i l e up on the updri f t s ide of the s t ruc ture . However, t h i s accumulation of sand on the updri f t s ide l imi t s the supply
a= 0 w +,
New first dune forming
\ C
* - - \
, , ~allen'house
Gentle slope
0 -t
New profile of adjustment + c
after storm re-storm profile -iV/y/%/lq
Figure 29. Beach flattening in response to a storm. Shaded area In A' is about @quai lo shaded area in A, Hause is not drawn to scale. (From Pilkey et at. 4984).

of sand t o beaches on the downdrift side. The r e s u l t i s t h a t t h e s e s t r u c t u r e s actual ly increase the r a t e of erosion on downdrift beaches. A c l a s s i c example of t h i s i s represented by the j e t t i e s bu i l t i n 1898 t o s t a b i l i z e t h e i n l e t t o Char les ton Harbor (Neal e t a l . 1984) . Since tha t time, sand has accreted a t S u l l i v a n s i s l a n d t o t h e no r th , whi le Morris I s land t o t h e south has been severely eroding. In the mid - 1800rs, Morris Island had dunes 10-12 m high and a well developed f o r e s t of p ines and pa lmet toes . P r e sen t l y i t i s a low, rapidly migrating sand f l a t . The Morris Is1 and Lighthouse, which was approximately 850 m from the shoreline in the l a t e 19th century, now stands 500 m offshore (Figure 30) (Neal e t a1 . 1984).
Recognizing t ha t j e t t i e s often cause e ro s ion of "downstream" beaches, newer j e t t i e s , such as those a t Murrells I n l e t , SC, a re being b u i l t with provisions t o move sand from the updri f t s ide of the
j e t t i e s t o t h e downdrif t s i d e . A t Murrells In le t the inner section of the north j e t t y i s a subtidal weir j e t t y , al lowing sand t o pass over i n t o a deposition basin (Van Dolah e t a1 . 1984). The design was t o allow the basin to be periodically dredged, depositing the sand on the downdrift side of the south j e t t y .
Seawal ls and bulkheads a r e constructions of 1 a s t r esor t (Pi1 key e t a1 , 1980). Seawall s r e f l e c t wave energy a t high t i d e , i nc r ea s ing t h e r a t e o f offshore sand t ranspor t , This steepens the beach p ro f i l e , which in turn increases t h e energy of t h e waves s t r i k i n g t h e s eawa l l . Seawall s a1 so i nc r ea se t h e i n t e n s i t y of longshore c u r r e n t s , which remove even more sand from in f ront of the wall. Ultimately the system i s s e l f - destructing.
Seawall s and bul kheads a1 so prevent the exchange of sand between the beach and the dunes during storms. The beach cannot
Figure 30. The lighthouse at Morris Island, SC (photo taken by 0. Pilkey).
50

.r $ f a t t e n 48 response t o s t o r m generated waves. I n s t ead ; t h e f u l l force o f t h e s e waves i s expended on t he s t r u c t u r e with t h e r e s u l t t h a t -it f r e q u e n t l y f a i l s . U l t i m a t e l y , i f s e a l e v e l c o n t i n u e s t o r i s e , a n y t h i n g t h a t d o e s n ' t m i g r a t e shoreward w i l l be destroyed o r l e f t a t s ea l i k e t h e Morris I s land i i g h t h o u s e (F igu re 30) !
5.3 EFFECTS OF JETTIES Obi NEARBY BENTHlC COMMUNlf IES
Knot e t a l . ( 1984) sampled t h e macrobenthic cammunities of t h e i n t e r t i d a l and n e a r s h o r e s u b t i d a ? e n v i r o n m e n t s a t Murre l l s I n l e t , South Ca ro l i na . This was done dur ing t h e cons t ruc t i on o f t h e j e t t y and once again 5 yea r s l a t e r . They found t h e infaunal community t o be dominated by severa l spec i e s of polychaetes (40% of t h e s p e c i e s and 60% o f t h e i n d i v i d u a l s ) , amphi pods, and pel ecypods. The presence o f t h e j e t t y a p p e a r e d t o a f f e c t t h e
d i s t r i b u t i o n and abundance of on ly one b iva lve and one polychaete . Comparison of spec i e s abundance between y e a r s and among l o c a l i t i e s ( u p d r i f t and downdri f t ) s u g g e s t e d no w i d e s p r e a d i m p a c t s a t t r i b u t a b l e t o j e t t y cons t ruc t i on .
R e c r e a t i o n a l f i s h i n g i s o f t e n c o n c e n t r a t e d a round r u b b l e s t r u c t u r e s ( F i g u r e 31) b e c a u s e o f t h e i n c r e a s e d numbers of f i s h e s t h a t occur t h e r e . Parker e t a l . (1979) es t imated t h a t an a r t i f i c i a1 r e e f c o n s t r u c t e d o f f M u r r e l l s I n l e t , SC, i n c r e a s e d f i s h s tanding s tock i n t h a t immediate l o c a t i o n by a f a c t o r of 1,800. J e t t i e s could have s i m i l a r consequences and o f t en seem t~ i n c r e a s e a n g l e r d e n s i t i e s by s i m i l a r amounts. Figures 32 and 33 show t h e s e a s o n a l p a t t e r n o f a n g l e r s u s i n g t h e j e t t i e s a t Murre l l s I n l e t , SC, and t h e spec i e s of f i s h e s caught r e l a t i v e t o t h e t ypes f o r which they were f i s h i n g .
Figure 31. Large jetties like the one at MurreBIs Inlet, SC, pictured here, provide favored Rearshore fishing sites (photo taken b y Fa. Vain Dolah).
51

- 20 0
5.5 JEU1ES AS DIVING SITES 0
16 - >-
Most of the sport diving industry +- t- 12 along the South Atlantic Bight is centered W 7 around wreck diving on the Continental " 8 5 Shelf. However, training dives for V) 3 beginning SCUBA classes and most r~ 4 recreational shore dives take place at 2 0 CT!
jetties since these provide nearshore - 0 0
- access to deeper water and allow divers to 0 $6 view higher densities of fishes and x - benthic organisms. Given the small * + proportion of the population that dives I- w 7 and the limited number of dives conducted w Z
on jetties, jetties do not represent a R substantial asset for the sport diving
<n ar
industry. Nonetheless, they provide W _ J
inexpensive recreational and educational 9 4 z opportunities to individual s that would 4 x not otherwise be able to view these reef- $ 0 m
like communities.
Figure 32. Estimated total number of boats (A) and bank-anglers (B) using the jetties at Murreils Inlet, SC, during different seasons (data from Van Doiah et al. 1986).
PERCENTAGE OF TOTAL PERCENTAGE OF ANGLERS CATCH BY SPECIES SEEKING THAT SPEC1 ES
20 10 0 10 20 30 40
Red Orurn Flounder Spat Bluefish King Mackerel Sheepshead Spatted Sea Trout Black Drum Croaker Spanish Mackerel Kingfish (Whiting) Black Sea Bass Shark (any species) Florida Pompano Pinfish Skates Tautoq
Figure 33. Types sf fishes caught compared with types desired by anglers using the jetties at Murreils Inlet, SC (data from Van Dolalr et at. 1986).

REFERENCES
kdams, S.M. 1976a. Feeding ecology of e e l g r a s s f i s h communities. Trans. Am. F i sh . Soc. 105:514-519.
Adams, S.M. 1976b. The ecology of Zos te ra marina ( L . ) f i s h communities. 11. func t iona l a n a l y s i s . J . Exp. Mar. Biol . E c o ~ . 22:293-311.
Amsler, C . D . , and R . B . S e a r l e s . 1980. V e r t i c a l d i s t r i b u t i o n of seaweed spores i n a water column o f f sho re of North Caro l ina . J . Phycol. 16:617-619.
Anthony, V . C . 1971 . The d e n s i t y dependence of growth i n t h e A t l a n t i c he r r i ng i n Maine. Rapp. P.-V. Reun. Cons. I n t . Explor. Mer. 160: 197-205.
A y l i n g , A . M . 1981. The r o l e o f b io log ica l d i s t u rbance in temperate s u b t i d a l e n c r u s t i n g communi t i e s . E C O ~ ogy 62:830-847.
Bakus, G.J. 1964. The e f f e c t s o f f i s h graz ing on i n v e r t e b r a t e evo lu t i on i n shal low t r o p i c a l waters . A1 1 en Hancock Found. Occas. Pap. 27: l -29.
Bakus, G.J. 1968. Defense mechanisms and ecology of some t r o p i c a l ho lo thu r i ans . Mar. Biol . 2:23-32.
Bakus, G.J. 1969. Energe t ics and feed ing in shal low marine waters . I n t . Rev. Gen. Exp. Zool. 4:275-369.
Bass, R.J., and J.W. Avault , J r . 1975. Food h a b i t s , length-weight r e l a t i o n s h i p , condi t ion f a c t o ~ , and growth of j u v e n i l e r e d drum, S c i a e n o ~ s o c e l l a t u s , i n L o u i s i a n a . T r a n s . A m . F i s h . Soc . 104:35-45.
Bearden, C.M. 1963. A c o n t r i b u t i o n t o t h e biology of t h e king whi t ings genus
Ment i c i r r h u s o f Sou th C a r o l i n a . Cont r ib . Bears Bluff Lab. 38 : l -27 .
Bengs ton , D . A . 1984. Resou rce p a r t i t i o n i n g by Menidia menidia and Menid ia b e r y l 1 i n a ( O s t e i c h t h y e s : A t h e r i n i d a e ) . Mar. Eco l . Prcrg. S e r . 1 8 ~ 2 1 - 3 0 .
Berns te in , B . B . , B . E . Williams, and K . E . Mann. 1981. The r o l e of behavioral r e s p o n s e s t o p r e d a t o r s i n mod i fy ing u r c h i n s ' ( S t r o n s y l o c e n t r o t u s droebachiensi s ) d e s t r u c t i v e graz ing and seasonal forag ing p a t t e r n s . Mar. Biol . 63:39-49.
B e r r i e n , P . , and D . F i n a n . 1978. B i o l o g i c a l and f i s h e r i e s d a t a on S p a n i s h m a c k e r e l , Scomberomorus rnacul a t u s (Mi t c h i 1 1 ) . NMFS Sandy Hook Lab. Tech. Ser . Rep. 9.
Bigelow, H . B . , and W.C. Schroeder. 1953. Fishes of t h e Western North A t l a n t i c . P a r t 2 . Mem. Sears Found. Mar. Res. 15: 1-514.
B l a i r , S.M., and M.O. Ha l l . 1981. Ten new records of deep water marine a lgae from G e o r g i a and Sou th C a r o l i n a . Northeast Gulf Sci . 4: 127-130.
Bohlke, J .E . , and C . C . G . Chaplin. 1958. Fishes of t h e Bahamas and ad j acen t t r o p i c a l wa t e r s . Livingston Pub1 i sh ing . Company, Wynnewood, Pennsylvania. 771 PP-
Bohnsack, J .A . , and D.L. S o u t h e r l a n d . 1985. A r t i f i c i a l r e e f research : a review wi th recommendations f o r f u t u r e p r i o r i t i e s . Sufi, War. S c i . 37~11-39 .
Boothby, R.N., and J.W. Avault, J r . 1971. Food h a b i t s , length-weight r e l a t i o n s h i p ,

a c o n d i t i o n f a c t o r of t h e r e d drum (Sciaenops o c e l l a tus ) i n southeastern Louis iana. Trans. Am. F ish . Soc. 100: 290-295. -
Bozeman, E.L., Jr., and J.M. Dean. 1980. The abundance o f e s t u a r i n e 1 a r v a l and j u v e n i l e f i s h i n a Sou th C a r o l i n a i n t e r t i d a l creek. Es tua r i es 3:89-97.
Brawley, S.H., and W.H. Adey. 1981a. The e f f e c t s o f m i c r o g r a z e r s on a l g a l communi ty s t r u c t u r e i n a c o r a l r e e f microcosm. Mar. B io . 61:167-177.
Brawley, S.H., and W.H. Adey. 1981b. M i c r o g r a z e r s may a f f e c t m a c r o a l g a l d e n s i t y . Nature 292: 177.
Breen , P.A., and K.H. Mann. 1976. Chang ing l o b s t e r abundance and t h e d e s t r u c t i o n o f k e l p beds by sea-u rch ins . Mar. B i o l . 34:137-142.
Buchanan, C.C. 1973. E f f e c t s o f an a r t i f i c i a l h a b i t a t on t h e mar ine s p o r t f i s h e r y and economy o f M u r r e l l s I n l e t , South Ca ro l i na . Mar. F i sh . Rev. 35: 15-22.
Bumpus, D.F. 1973. A d e s c r i p t i o n o f t h e c i r c u l a t i o n on t h e Con t i nen ta l S h e l f of t h e eas t coas t o f t h e U n i t e d S ta tes . Prog . Oceanogr. 6: 11 1-157.
Caldwe l l , D.R. 1957. The b i o l o g y and systemat ics o f t h e p i n f i s h , Laqodon rhomboides (L innaeus) . B u l l . F l a . S t a t e Mus. B i o l . Sc i . 2:77-173.
C a r l i s l e , J.G., Jr., C.H. Turner, and E.E. Eber t , 1964. A r t i f i c i a l h a b i t a t i n t h e mar ine environment. Cal i f . Dep. F ish . Game F i s h B u l l . 124:l-93.
occupying seagrass beds i n t h e e s t u a r i n e zone near C r y s t a l R i ve r , F l o r i d a . Trans. Am. Fish.Soc. 102:511-540.
Chao, L.N., and J.A. Musick. 1977. L i f e h i s t o r y , f e e d i n g h a b i t s , and f u n c t i o n a l mo rpho logy o f j u v e n i l e sc i aen id f i s h e s i n t h e York R i ve r estuary , V i r g i n i a . F i sh . B u l l . 75:657- 702.
Chapman, R.L. 1971. The macroscopic mar ine a lgae o f Sapelo I s 1 and and o t h e r s i t e s on t h e Georgia coas t . B u l l . Ga. Acad. Sc i . 29:77-89.
Chapman, R.L. 1973. An a d d i t i o n t o t h e m a c r o s c o ~ i c mar ine a1 aae o f Georai a. The genus ~l a d o ~ h o r a . B u l l . Ga. gad. Sc i . 31:147-150.
Chestnut, A.F., and W.E. Fahy. 1953. Stud ies on t h e v e r t i c a l d i s t r i b u t i o n of s e t t i n g o f o y s t e r s i n No r t h Ca ro l i na . P roc . G u l f C a r i b b . F i s h . I n s t . 1952:106-112.
Choat, J.H. 1982. F i s h feed ing and t h e s t r u c t u r e o f b e n t h i c communities i n t e m p e r a t e w a t e r s . Annu. Rev. E c o l . Syst . 13:423-449.
C o n n e l l , J .H. 1972. Community i n t e r a c t i o n s on mar ine r ocky i n t e r t i d a l shores. Annu. Rev. Ecol . Sys t . 3:169- 192.
Connel l , J.H. 1975. Some mechani sms producing s t r u c t u r e i n n a t u r a l communi - t i e s . Pages 460-490 i n M.L. Cody and J.M. Diamond, eds. Ecology and evo lu - t i o n o f communities. Belknap Press o f Harvard U n i v e r s i t y Press, Cambridge, Mass.
C a r p e n t e r , R.C. 1984. P r e d a t o r and C o n n e l l , J .H. 1978. D i v e r s i t y i n popu la t i on d e n s i t y c o n t r o l of homing t r o p i c a l r a i n f o r e s t s and c o r a l r e e f s . b e h a v i o r i n t h e C a r i b b e a n e c h i n o i d Science 199:1302-1310. D i aderna a n t i l l a r u m . Mar . B i o l . 82:lOl-108. Connel l . J.H.. and R.O. S l a v t e r . 1977.
~ e c h a n i sms o f success ion - i n n a t u r a l C a r p e n t e r , R. C . 1986. P a r t i t i o n i n g communities and t h e i r r o l e i n community
k e r b i v o r y and i t s e f f ec t s on c o r a l s t a b i l i t y and o rgan i za t i on . Am. Nat. ree f a1 ga l communities . Ecol . Monogr. 111:1119-1144. 56:345-363.
Cowen, R.K. 1983. The e f f e c t s o f Carr , W.E.S., and C.A. Adarns. 1973. Food sheephead ( S e r n i c o s s v ~ h u s p u l c h e r )
' h a b i t s o f j u v e n i l e m a r i n e f i s h e s p r e d a t i o n on r e d sea u r c h i n

(Stronqylocentrotus franciscannus) populations: an experimental analysis. Oecologia (Berl . ) 58:249-255.
Currin, B.M., J.P. Seed, and J.M. Miller. 1984. Growth, production, food consump- tion, and mortality of juvenile spot and croaker: a comparison of tidal and non- tidal nursery areas. Estuaries 7:451- 459.
Cushing, D.H., and J.W. Horwood. 1977. Development of a model of stock and recruitment. Pages 21-35 in Steele, J.H., ed. Fisheries mathematics. Academic Press, London.
Dahlberg, M.D. 1975. Guide to coastal fishes of Georgia and nearby states. University of Georgia Press, Athens. 186 pp.
Dame, R.F. 1979. The abundance, diversity and biomass of macrobenthos on North Inlet, South Carolina, intertidal oyster reefs. Proc. Nat . Shelifish. Assoc. 69:6-10.
D'Antonio, C. 1985. Epiphytes on the rocky intertidal red alga Rhodomel a larix (Turner) C. Agardh: negative effects on the host and food for herbivores? J. Exp. Mar. Biol . Ecol . 86:197- 218.
Darcy, G.H. 1983. Synopsis of biological data on the pigfish, Ortho~ristis chrysoptera (Pisces:Haemulidae). NOAA Tech. Rep., NMFS Circ. 449.
Darcy, G.H. 1985a. Synopsis of biological data on the pinfish, Laqodon rhomboides (Pisces:Sparidae) . NOAA Tech. Rep., NMFS Circ. 23.
Darcy, G.H. 1985b. Synopsis of biological data on the spottail pinfish, D i ~ l odus hol brooki (Pi sces:Sparidae) . NOAA Tech. Rep., NMFS Circ. 13.
Darnell, R.M. 1958. Food habits of fishes and larger invertebrates o f Lake Pontchartrain, Louisiana, an estuarine community. Pub1 . Inst. Mar. Sci. Univ. Tex. 5:353-416.
Darnell, R.M. 1961. Trophic spectrum of an estuarine community, based on studies of Lake Pontchartrain, Louisiana. E C O ~ ogy 42: 553-568.
Davi es, J. L. 1964. A morphogeneti c approach to world shore1 ines. Z. Geomorphol. 8:127-142.
Dayton, P.K. 1971. Competition, disturbance, and commun i ty organization: the provision and subsequent utilization of space in a rocky intertidal community. Ecol. Monogr. 41:351-389.
Dayton, P.K. 1975. Experimental evaluation of ecological dominance in a rocky intertidal algal community. Ecol . Monogr. 45: 137-159.
Dethier, M.N. 1984. Disturbance and recovery in intertidal pool s : maintenance of mosaic patterns. Ecol. Monogr. 54:99-118.
Dethier, M.N., and D.O. Duggins. 1984. An "indirect commensalism" between marine herbivores and the importance of competitive hierarchies. Am. Nat. 124:205-219.
Diener, R . A . , A. Ingl is, and G.B. Adams. 1974. Stomach contents of fishes from Clear Lake and tributary waters, a Texas estuarine area. Contri b. Mar. Sci . 18:7-17.
Dixon, P.S. 1965. Perennation, vegetative propagation and algal f ife histories, with special reference to Asperasopsis and other Rhodophyta. Botanica Gothoburgensia 3:67-74.
Doherty, P.J. 1982. Some effects of density on the juveniles of two species of tropical, territorial damsel f f shes. J. Exp. Mar. Biol. Ecol. 65:249-261.
Doherty, P.J. 1983a, Tropical territorial damsel fishes: is density 1 imi ted by aggression or recruitment? Ecology 643175-190.
Doherty, P.J. 1983b. Coral reef fishes: recrui tment-1 ini ted assembl ages?

Proc . Four th I n t . Cora l Reef Symp., Phil ippines 2:465-470.
Ear le , L . , and H . H u m m . 1964. I n t e r t i d a l zonation of a lgae in Beaufort Harbor. J . El i sha Mitchel l Sc i . Soc. 80:78-82.
Edgar, G.J. 1983. The ecology of south- e a s t Tasmanian p h y t a l animal communi t i e s . IV. Factors a f f e c t i n g t h e d i s t r i b u t i o n of ampi thoid amphi pods among a lgae . J . Exp. Mar. Biol . Ecol . 70:205-225.
Eiseman, N.J. 1976. Benthic p l a n t s of t h e e a s t F lor ida Continental She1 f . Annu. Rep. Harbor Branch Found. 1976:Al-A13.
Fox, R.S. , and E . E . Ruppe r t . 1985. Shal low-water mari ne b e n t h i c macroi nve r t eb ra t e s of South Carol i na. S p e c i e s i d e n t i f i c a t i o n , community composition, and symbiotic a s soc ia t ions . Bel le W . Baruch Library in Marine Science, No. 14. Univers i ty of South Carol i na Press , Col umbi a.
Fulton, R.S., 111. 1983. I n t e r a c t i v e e f f e c t s o f temperature and predat ion on an e s t u a r i n e zooplankton community. J . Exp. Mar. Biol . Ecol. 72:67-81.
Ful t on , R.S., 111. 1985. Predator-prey re1 a t ionsh ips i n an e s t u a r i n e 1 i t t o r a l copepod community. Ecology 66:21-29.
Eiseman, N.J. 1979. Marine a lgae of t h e Gaines, S.D. , and J . Lubchenco. 1982. A e a s t F lor ida Continental Shel f . 1 . un i f i ed approach t o marine p l an t - Some new records of Rhodophyta including h e r b i v o r e i n t e r a c t i o n s . 11. S c i n a i a i n c r e s s a t a s p . nov. Biogeography. Annu. Rev. Ecol. Sys t . (Nemal i a1 e s , C h a a e t a n g i a c e a e ) . 13: l l l -138. Phycologia 18:355-361.
Eiseman, N.J., and R . L . Moe. 1981. M a r i o e l t a a t 1 a n t i c a s p . nov. (Rhodophyta, Rhodomeni a1 e s ) a new deep- water alga from Flor ida . J . Phycol. 17:299-308.
Eiseman, M.J., and J.N. Norris . 1981. Dudresnava pa tu l a sp. nov., an unusual deep water red a lga from F lo r ida . J . Phycol. 17:186-191.
Es tes , J.A., and J.F. Palmisano. 1974. Sea o t t e r s : t h e i r r o l e i n s t r u c t u r i n g n e a r s h o r e communi t ies . S c i e n c e 185: 1058-1060.
E s t e s , J . A . , N.S. Smi th , and J.F. Palmisano. 1978. Sea o t t e r predat ion and community o r g a n i z a t i o n i n t h e w e s t e r n A l e u t i a n I s 1 a n d s , A1 a s k a . E C O ~ ogy 59 : 822-833.
Farrand, J . , J r . , ed. 1983a. The Audubon Socie ty master guide t o b i rd ing . Vol. 1. Loons t o sandpipers . Alfred A . Knopf, New York. 447 pp.
Farrand, ,I.,&., ed. 1983b. The Audubon Socie ty master guide t o b i rd ing . Vol. 2. Gul ls t o d ippers . Alfred A . Knopf, New York. 397 pp.
Gallaway, B. J . , L . R . Martin, R.L . Howard, G.S. Boland, and G.D. Dennis. 1981. E f fec t s on a r t i f i c i a l reef and demersal f i sh and macrocrustacean communities. Pages 237-299 i n B . S . Middlei tch, ed. Environmental e f f e c t s of o f f sho re o i 1 production. Plenum Pub1 i shing Corp., New York, N . Y .
G i l l i gan , M . R . (1987). An i l l u s t r a t e d f i e l d guide t o t h e f i s h e s o f Gray's Reef National Sanctuary. NOAA Tech. Rep. OCRMISPD. In p re s s .
Glynn, P.W. 1965. Community composition, s t r u c t u r e , and i n t e r r e l a t i o n s h i p s i n t h e marine i n t e r t i d a l Endocl adi a muricata- Bal anus a1 andul a a s soc ia t ion in Monterey Bay, Cal i f o r n i a . Beaufort ia 12:l-198.
Godfrey, P .J . , and M . M . Godfrey. 1977. Ba r r i e r i s l and ecosystems o f Cape Lookout National Seashore and v i c i n i t y , North Carol i na. Sci . Monogr. 13, Natl . Park Serv. , Washington, D.C.
Gof f , L . J . 19766. The b i o l o g y o f Harvevel 1 a m i r ab i 1 i s (Cryptonemi a1 es, Rhodophyceae) . V . Host r e s p o n s e t o p a r a s i t e i n f e c t i o n . J . Phycol . 12:313 328.

Gosner, K . L . 1979. A f i e l d gu ide t o t h e At1 a n t i c seashore . Houghton Miff1 i n Co. Boston., 329 pp.
Govoni, J . J . , D . E . Hoss, and A.J. Ches te r . 1983. Comparative feed ing of t h r e e s p e c i e s of l a r v a l f i s h e s in t h e nor thern Gul f of Mexico: Brevoort i a patronus, Leiostomus xanthurus, and Mic ro~oqon ia s undul a tu s . Mar. Ecol . Prog. Ser . 413:189-199.
Govoni, J . J . , P . B . O r tne r , F . Al-Yamant, and L . C . H i l l . 1986. S e l e c t i v e feed ing o f s p o t , Le ios tomus x a n t h u r u s , and A t l a n t i c c rocke r , Mi croooqoni a s undulatus , l a r v a e in t h e nor thern Gulf o f Mexico. Mar. E c o l . Prog . S e r . 28: 175-183.
Grant . G . C . 1962. Preda t ion of b l u e f i s h on young A t l a n t i c menhaden i n Indian River , Delaware. Chesapeake Sci . 3:45- 47.
Grant , J . J . , K .C . Wilson, A . Grover, and H . A . Togstad. 1982. Early development of P e n d l e t o n a r t i f i c i a l r e e f . Mar. F i sh . Rev. 44:53-60.
Gray, I .E . , and M.J. Cerame-Vivas. 1963. The c i r c u l a t i o n of s u r f a c e wa te r s i n Raleigh Bay, North Caro l ina . Limn01 . Oceanogr. 8:330-337.
Greze , I . I . 1986. Feeding h a b i t s and food requirements of some amphipods i n t h e Black Sea. Mar. Biol . 1:316-321.
G r i f f i n , M.M., and V.J. Henry. 1982. H i s t o r i c a l changes i n t h e mean h igh water shore1 i ne of Georgia 1857-1982. Georg i a Dep t . Nat . Res . , Env. P r o t . D i v . , Geo rg i a G e o l o g i c Su rvey B u l l . 98: 1-78.
Gunnil 1 , F . C . 1985. Growth, morphology and mi c roherb i vore faunas of Pel ve t i a f a s t i q i a t a (Phaeophyta, Fucaceae) a t La J o l l a , C a l i f o r n i a , USA. Bot . Mar. 28: 187-199.
Hales, L.S., and M. Van Den Avyle. (1985.) Species p r o f i l e s : 1 i f e h i s t o r i e s and environmental requirements of coas t a l f i s h e s and i n v e r t e b r a t e s . South At1 a n t i c Bight . Spot (Leiostomus xanthurus) . U.S. Fish W i f d l . Serv. Biol .
Rep. 8 2 ( ) U.S. Army Corps of Engineers , TR EL-82-4. In p r e s s .
Ha l l , M.O., and N.J. Eiseman. 1981. The s e a g r a s s e p i p h y t e s o f t h e I n d i a n River , F lo r ida . I . Spec ies l i s t wi th d e s c r i p t i o n s and seasonal r e f e r ences . Bot. Mar. 24:139-146.
Hansen, D . J . 1969. Food, g r o w t h , reproduct ion , and abundance of pi nf i s h , Lasodon rhombo ides , and a t l a n t i c c r o a k e r , Microooqon u n d u l a t u s , n e a r Pensaco l a , F l o r i d a . F i s h . B u l l . 68:135-146.
Hansen, M . E . , and D . L . Ward. 1986. Response of e b b - t i d a l d e l t a s t o j e t t y cons t ruc t i on . Pages 38-39 in Aubrey, D . G , L . Weishar, D. Duane, and L. Bu t l e r , eds . Hydrodynamics and sediment dynamics of t i d a l i n l e t s . Woods Hole Oceano- graphic I n s t i t u t i o n symposium. Woods Hole Oceanographic I n s t i t u t e , U.S. Army Engineer Waterways Experiment S t a t i o n , Coastal Engineering Research Center .
Ha r l i n , M . M . 1973. T r a n s f e r o f p roducts between e p i p h y t i c marine a l g a e and hos t p l a n t s . J. Phycol . 9:243-248.
Har r i s , G.P. 1980. Temporal and s p a t i a l s c a l e s i n phy top l a n k t o n e c o l o g y . Mechanisms, methods , model and management. Can. J . F i sh . Aquat. S c i . 37:877-900.
Hastings, R.W. 1972. The o r i g i n and s e a s o n a l i t y o f f i s h fauna on a new j e t t y i n t h e n o r t h e a s t e r n Gul f o f Mexico. P h . D . D i s s e r t a t i o n . F lo r ida S t a t e Un ive r s i t y , Ta l l ahas see . 124 pp.
Hast ings, R.W. 1978. Rock j e t t y f i s h fauna a s an endangered shore based f i s h e r y . Mar. Recreat ional Fish. 3:29- 36.
Hastings, R.W. 1979. The o r i g i n and seasonal i t y o f t h e f i s h fauna on a new j e t t y i n t h e n o r t h e a s t e r n Gul f o f Mexico. Bull . F7a. S t a t e Mus. B io l . S c i . 24:l-122.
Hatcher, B . G . , and A.W.D, Larkurn. 1983. An e x p e r i m e n t a l a n a l y s i s o f f a c t o r s c o n t r o l l i n g t h e s tanding crop of t h e

e p i l i t h i c a lga l community on a cora l r e e f . J . Exp. Mar. B io l , Ecol. 69:61- 84.
Hawkins, S . J . , and E . H a r k i n . 1985. Prel iminary canopy removal experiments i n a lga l dominated communities low on t h e shore and in t h e shal low sub t ida l on t h e I s l e of Man. Bot. Mar. 28:223- 230.
Hawkins, S . J . , and R . G . Hartnol l . 1983. Grazing of i n t e r t i d a l a lgae by marine i n v e r t e b r a t e s . Oceanogr . Mar. Bi 01 . Annu. Rev. 21:195-282.
Hay, M . E . 1981a . The f u n c t i o n a l morphology of tu r f - forming seaweeds: p e r s i s t e n c e i n s t r e s s f u l m a r i n e h a b i t a t s . Ecology 62:739-750.
Hay, M . E . 1981b. H e r b i v o r y , a l g a l d i s t r i b u t i o n , and t h e maintenance of between-habi t a t d i v e r s i t y on a t r o p i c a l f r i n g i n g r e e f . Am. Nat. 118:520-540.
Hay, M . E . 1981c. Spa t i a l p a t t e r n s of graz ing i n t e n s i t y on a Caribbean b a r r i e r r e e f : herbivory and a lga l d i s t r i b u t i o n . Aquat . Bot . 11 : 97- 109.
Hay, M . E . 1984a. P red i c t ab l e s p a t i a l escapes from herbivory: how do t h e s e a f f e c t t h e e v o l u t i o n o f h e r b i v o r e r e s i s t a n c e i n t r o ~ i c a l m a r i n e communities? Oecologia ( ~ e r l . ) 64:396- 407.
Hay, M.E . 1984b. Pa t t e rns o f f i s h and urchin graz ing on Caribbean cora l r e e f s : a r e p r e v i o u s r e s u l t s t y p i c a l ? E C O ~ ogy 65: 446-454.
Hay, M . E . 1985 . S p a t i a l p a t t e r n s o f herbivore impact and t h e i r importance i n maintaining a lga l spec i e s r i chnes s . Proc, F i f t h I n t . Coral Reef Congr., T a h i t i . 4:29-34.
Hay, M. E . 1986. Assoc ia t iona l p l an t d e f e n s e s and t h e m a i n t e n a n c e o f spec i e s d i v e r s i t y : t u rn ing competi tors i n t o accarnpl i c e s . Am. Nat. 128:617- 641.
Hay, M.E . , R .R . Lee, R . A . Guieb, and M . M . B e n n e t t . 1986. Food p r e f e r e n c e and chemotaxis i n t h e sea urchin Arbacia
punctul a t a (Lamarck) Phil i pp i . J . Exp. Mar. Biol . Ecol. 96:147-153.
Hay, M . E . , J .E. Duffy, C . A . P f i s t e r , and W . Feni ca l . 1987 . Chemical defense aga ins t d i f f e r e n t marine herb ivores : a r e amphi pods i n s e c t equ iva l en t s? Ecology 68:1567-1580.
Hay, M . E . , P . E . Renaud, and W . Fenica l . 1988 . Large mobile versus small sedentary herbivores and t h e i r r e s i s t ance t o seaweed chemical defenses . Oecol ogi a 1 . . 75:264-252.
Hi 1 debrand, S. F . , and L . E. Cab1 e . 1930. Development and 1 i f e h i s t o r y o f four teen t e l eostean f i s h e s a t Beaufort , N . C . Bul l . U.S. Bur. F ish . 46:383- 488.
H i l d e b r a n d , S . F . , and W . C . S c h r o e d e r . 1928. Fishes o f Chesapeake Bay. Bull . U.S. Bur. Fish. 43: l-366.
Hixon, M.A., and W . N . Bros tof f . 1983. Damsel f i s h as keystone spec i e s i n r eve r se : in te rmedia te d i s tu rbance and d i v e r s i t y of r ee f a lgae . Science 220:511-513.
Hodson, R.G., J.O. Hackman, and C . R . Bennett. 1981. Food h a b i t s o f young s p o t s i n nursery a r eas of t h e Cape Fear River e s tua ry , North Carol i na. Trans. Am. Fish. Soc. 110:495-501.
Hoffman, S.G., and D . R . Robertson. 1983. Fo rag ing and r e p r o d u c t i o n o f two Car ibbean r e e f t o a d f i s h e s ( B a t r a c h o i d i d a e ) . B u l l . Mar. S c i . 33:919-927.
H o l l i n g , C.S. 1973. R e s i l i e n c e and s t a b i l i t y o f e c o l o g i c a l s y s t e m s . Annu. Rev. Ecol. Sys t . 4 : l -23 .
Hoss, D . E . 1974. Energy requirements o f a p o p u l a t i o n o f p i n f i s h Lasodon rhomboides (Linnaeus) . Ecology 55:848- 855.
Hoyt , W.D. 1920. Marine a l g a e o f Beaufort , N . C . and ad j acen t reg ions . Bul l . Bur. Fish. Wash. 36:367-556.
Humm, H.J. 1952. Notes on t h e marine a lgae o f F lo r ida . I . The i n t e r t i d a l

rocks a t marineland. Stud. Fla. S t a t e Rhodophyta. The U n i v e r s i t y o f North Univ. 7:17-23. Carol ina Press , Chapel H i l l . 206pp.
Humm, H.J. 1969. Dis t r i bu t ion of mar ine a lgae along t h e A t l a n t i c c o a s t of North America. Phycol ogi a 7: 43-53.
Huntsman, G . R . , and C.S. Manooch, 11 I . 1978. Coastal pe lag ic and reef f i s h e s in t h e South A t l a n t i c Bight. Pages 97- 106 in H . Clepper ed. Proceedings second annual marine r e c r e a t i o n a l f i s h e r y symposium, Sport Fish. I n s t . , Washington D . C .
Hurme, A . K . 1979. Rubbl e-mound s t r u c t u r e s a s a r t i f i c i a l r e e f s . P roc . Coastal S t r u c t . 79:1042-1051.
Jackson, J.B.C. 1977. Competition on marine, hard s u b s t r a t a : t h e a d a p t i v e s ign i f i cance of sol i t a r y and c o l o n i a l s t r a t e g i e s . Am. Nat. 111:743-767.
J o h n s g a r d , P . A . 1981. The p l o v e r s , sandpipers , and sn ipes of t h e wor ld . Universi ty of Nebraska Press , Li ncol n. 541 pp.
Johnson, G.F., and L . A . de Wit. 1978. E c o l o g i c a l e f f e c t s o f an a r t i f i c i a l i s l a n d , Rincon Is1 and , Punta G o r d a , Ca l i fo rn i a . U.S. Army Corps Eng. C E R C Misc. Rep. No. 78-3:l-108.
Jones, C.P., and A.J. Mehta. 1978. Ponce de Leon I n l e t . Glossary of In1 e t s Report No. 6 . Dep. Coastal Ocean. Eng . , Univ. F l a . , Fla . Sea Grant Col l .
Jones, G.P . 1984a. Population ecology o f t h e t e m p e r a t e r e e f f i s h P s e u d o l a b r u s c e l i d o t u s Bl och and Schnei d e r ( P i s c e s : L a b r i d a e ) . I . F a c t o r s in f luenc ing recru i tment . J . Exp. Mar. Biol . Ecol . 75: 257-276.
Jones, G . P . 1984b. Population ecology o f t h e temperate r ee f f i s h Pseudo1 a b r u s c e l i d o t u s Bloch and S c h n e i d e r ( P i s c e s : L a b r i d a e ) . 11. F a c t o r s in f luenc ing adu l t d e n s i t y . J . Exp. Mar. Biol . Ecol . 75:277-303.
Kapraun, D.F. 1980a. An i l l u s t r a t e d guide t o t h e benthic marine a lgae o f c o a s t a l North C a r o l i n a Vol. I :
Kapraun, D . F . 1980b. F l o r i s t i c a f f i n i t i e s of North Carol i na inshore b e n t h i c mar ine a l g a e . Phyco log ia 19:245-252.
Kapraun, D . F . 1984. An i l l u s t r a t e d guide t o t he benth ic marine a lgae of coas t a l North Carol ina Vol . I I : Chl orophyta and Phaeophyta. Bi bl . Phycol . 58: 1 - 172.
Kapraun, D.F., and F.W. Zechman. 1982. Seasonal i t y and v e r t i c a l zonation of benth ic marine algae on a North Carol ina c o a s t a l j e t t y . B u l l . Mar. S c i . 32:702-714.
Karlson, R . H . 1978. Predat ion and space u t i l i z a t i o n p a t t e r n s i n a mar ine epi faunal community. J . Exp. Mar. Biol . Ecol . 31:225-239.
Kastendiek, J . 1982. Competi tor-mediated coexis tence : i n t e r a c t i o n s among t h r e e s p e c i e s o f b e n t h i c mac roa lgae . J . Exp. Mar. Biol . Ecol. 62:201-210.
Keough, M.J . , and H . C h e r n o f f . 1987. Di s ~ e r s a l and D O D U ~ a t i on v a r i a t i o n i n t he ' bryozoan fjuqul a ner i t i na. Ecology 68:199-210.
Kerr, G . A . 1976. Indian River Coastal zone s tudy inventory. Annu. Rep. Harbor Branch Consortium Indian River Coastal Zone Study. 2: l -105.
K i e s l i c h , J.M. 1981. T i d a l i n l e t response t o j e t t y cons t ruc t ion . GITI Report 19. U.S. Army Coastal Eng. Res. Cent. Sp r ing f i e ld , Va.
Kilby, J.D. 1955. The f i s h e s of two Gulf c o a s t a l marsh a r e a s of F l o r i d a . Tulane Stud. Zool. 2:173-247.
Kjelson, M.A. , D.S. Pe t e r s , G.W. Thayer, and G . N . Johnson. 1975. The general feeding ecology of pos t l a rva l f i s h e s i n t h e Newport River e s tua ry . Fish. Bull . 73:137-144.
Knott, D . M . , R . F . Van Dolah, and D . R . Calder . 1984. Ecological e f f e c t s o f r u b b l e w e i r j e t t y c o n s t r u c t i o n a t Murrell s I n l e t , South Carol i n a . Vol . 2:

Changes i n macrobenth ic cornmun.ities o f sandy beach and nearshore environments. U.S Army Corps Eng. Tech. Rep. EL-84-4, Coasta l Eng. Res. Cent., Waterw. Exp. Stn., Vicksburg, Miss. 99 pp.
Knutson, P.L., and K. F i n k e l s t e i n . 1987. Environmental cons i de ra t i ons f o r dune s t a b i l i z a t i o n p r o j e c t s . U.S. Army Engineer Waterways Experiment S ta t i on , Tech. Rep. EL-87-2. 39 pp.
Kobyl i n s k i , G. J., and P.F. Sheridan. 1979. D i s t r i b u t i o n , abundance, f eed ing and l ong - t e rm f l u c t u a t i o n s of spot , Leiostomus xanthurus, and croaker , M i c r o ~ o s o n i a s undulatus, i n Apala- ch ic01 a Bay, F l o r i d a , 1972-1977. Con t r i b . Mar. Sc i . 22:149-161.
Lawrence, J.M., and P.M. Sammarco. 1982. E f f e c t s o f f e e d i n g on t h e environment: Echinoidea. Pages 499-519 i n M. Jangoux and J.M. Lawrence, eds. Echinoderm n u t r i t i o n . A.A. Balkema Press, Rotterdam, The Nether1 ands.
Legget t , W.C. 1977. Dens i t y dependence, dens i t y independence and r e c r u i tment i n t h e American shad (Alosa sap id iss ima) p o p u l a t i o n o f t h e Connect icut R i ve r . Pages 3-17 i n W. van Winkle, ed. Proceedings o f t h e conference on assess ing t h e e f f e c t s o f power -p lan t induced m o r t a l i t y on f i s h popu la t ions . Pergamon Press. New York, N.Y.
Lewis, S.M., and B. Kensley. 1982. Notes o f t h e eco logy and behav io r o f Pseudam~h i tho ides i n c u r v a r i a ( J u s t ) (Crustacea, Amphipoda, Amphithoidea). J. Nat. H i s t . 16:267-274.
L i n d q u i s t , D.G., and R.M. D i l l aman. 1986. T r o p h i c m o r p h o l o g y o f f o u r w e s t e r n A t l a n t i c b l e n n i e s (P isces :B lenn i idae) . Copeia 1986:207-213.
L i n d q u i s t , D.G., M.V. Ogburn, W.B. S t a n l e y , H.L. Troutman, and S.M. P e r e i r a , 1985. F i s h u t i l i z a t i o n p a t t e r n s on rubble-mound j e t t i e s i n North C a r o l i n a . B u l l . Mar . S c i . 37~244-251 .
L i nk , G . W , , Jr. 1980. Age, growth, r ep roduc t i on , feeding, and e c o l o g i c a l observa t ions on t h e t h r e e spec ies o f
C e n t r o p r i s t i s ( P i s c e s : S e r r a n i d a e ) i n N o r t h C a r o l i n a w a t e r s . Ph.D. D i s s e r t a t i o n . U n i v e r s i t y o f N o r t h Ca ro l i na a t Chapel W i l l . 277pp.
Lippson, A.J., and R.L. Lippson. 1984. L i f e i n t h e Chesapeake Bay. Johns Hopkins Un iv . Press, Ba l t imo re . 229 pp.
L i t a k e r , W., C.S. Duke, B.E. Kenney, and J. Ramus. ( 1 9 8 7 ) . S h o r t - t e r m e n v i r o n m e n t a l v a r i a b i 1 i t y and phy top lank ton abundances i n a sha l low t i d a l e s t u a r y . P a r t 1: w i n t e r and summer. Mar. B i o l 96:115-121
Lubchenco, J. 1978. P l a n t s p e c i e s d i v e r s i t y i n a m a r i n e i n t e r t i d a l communi t y : importance o f h e r b i v o r e food p r e f e r e n c e and a1 g a l c o m p e t i t i v e a b i l i t i e s . Am. Nat. 112:23-39.
Lubchenco, J. 1980. A l ga l zona t i on i n t h e New Eng land r o c k y i n t e r t i d a l commun i ty : an e x p e r i m e n t a l a n a l y s i s . Ecol ogy 61 :333-344.
Lubchenco, J., and S.D. Gaines. 1981. A u n i f i e d app roach t o m a r i n e - p l a n t h e r b i v o r e i n t e r a c t i o n s . I . Popu la t ions and communi t i e s , Annu. Rev. Ecol . Sys t . 12:405-437.
Lunz, G.R. 1943. The y i e l d o f c e r t a i n o y s t e r l ands i n South Ca ro l i na . Am. M i d l . Nat. 30:806-808.
Manooch, C.S., 11. 1984. F i she rman ' s g u i d e t o f i s h e s o f t h e s o u t h e a s t e r n U n i t e d S t a t e s . N o r t h C a r o l i n a S t a t e Museum. Rale igh. 362 pp.
Ma r t i n , A.L. 1966. Feeding and d i g e s t i o n i n two i n t e r t i d a l uammar ids: Marinosammarus obtusatus and p i r l o t i . J. ZOO^. 148:515-552.
Mathieson, A.C., and C.J. Dawes. 1975. Seasona l s t u d i e s o f F l o r i d a sub1 i t t o r a l mar ine a lgae. B u l l . Mar. Sc i . 25:46-65.
M a t u r o , F . J. 1959 . Seasona l d i s t r i b u t i o n and s e t t l i n g r a t e s o f e s t u a r i n e bryozoa. Ecology 40: 116-127.
McCfoskey, L.R. 1970. The dynamics o f t h e community assoc ia ted w i t h a mar ine

s c l e r a c t i n i an c o r a l . I n t . Rev. Gesamten. Hydrobioi . 5 5 ~ 1 3 - 8 1 .
McDougall, K . D . 1943. S e s s i l e marine i n v e r t e b r a t e s o f B e a u f o r t , North Carol ina . Ecol . Monogr. 13 ~322-374.
Menge, B . A . 1976. Organizat ion o f t h e New England rocky i n t e r t i d a l community: r o l e o f p r e d a t i o n , c o m p e t i t i o n and env i ronmenta l he terogenei ty . Ecol . Monogr. 46:355- 393.
Menge, B.A., and J .P. Sutherland. 1976. Species d i v e r s i t y g rad ien t s : syn thes i s of t h e r o l e s of predat ion , competi t ion, and temporal he terogenei ty . Am. Nat . 110:351-359.
Mercer, L.P. 1984a. A b io log ica l and f i s h e r i e s profi 1 e of spo t t ed s e a t r o u t , Cvnoscion nebulosus. Spec. Sci . Rep. No. 40, Pro j . SF-13, NMFS.
Mercer, L . P . 19846. A b io log ica l and f i s h e r i e s p r o f i l e o f t h e r e d drum, Sciaenoas oce l l a t u s . Special Sci . Rep. No. 41, Pro j . SF-13, NMFS.
Merriner, J.V. 1975. Food h a b i t s o f t h e weakfish, Cvnosci on r eaa l i s , in North C a r o l i n a w a t e r s . Chesapeake S c i . 16: 74-76.
Mi l l e r , G . C . , and W.J. Richards. 1979. Reef f i s h h a b i t a t , faunal assemblages, and f a c t o r s determining d i s t r i b u t i o n i n t h e South A t l a n t i c Bight . Proc. Gulf Caribb. F ish . I n s t . 32:114-130.
Montgomery, W . L . 1977. Die t and gut morphology i n f i s h e s with spec i a1 reference t o t h e monkeyface prickleback Cebidichthvs v io l aceus (St ichae idae : Blennio ide i ) . Copeia 1977:178-182.
Montgomery, W . L . 1980. The impact o f non-se lec t ive grazing by the g i a n t blue damselfi sh Microspathodon dorsa l i s on a l g a l communi t ies i n t h e Gulf o f Ca l i fo rn i a , Mexico. Bull. Mar. Sc i . 30: 290-303.
Montgomery, W.L. , and S.D. Gerking. 1980. Marine macroalgae a s food f o r f i s h e s : an evalua t ion of po ten t i a l food q u a l i t y . Environ. Biol . Fish. 5:143-153.
Mook, D.H. 1976. S tudies on fou l ing inve r t eb ra t e s i n t h e Indian River . 1: seasonal i t y of se t t lement . Bull . Mar. S c i . 26:610-615.
Mook, D.H. 1980. Seasonal v a r i a t i o n i n spec i e s composition of r ecen t ly s e t t l e d f o u l i n g communit ies a l o n g an env i ronmen ta l g r a d i e n t i n t h e I n d i a n R i v e r Lagoon, F l o r i d a . E s t u a r i n e Coastal Mar. S c i . 11:573-581.
Mook, D.H. 1981. Effec ts o f d is turbance and i n i t i a l s e t t l e m e n t on f o u l i n g communi t y s t r u c t u r e . Ecology 62:5221 526.
Mook, D.H . 1983a. Indian River fou l ing organisms, a review. Fla. Sc i . 46:162- 167.
Mook, D.H. 1983b. Responses of common fou l ing organisms i n the Indian River, F l o r i d a , t o v a r i o u s p r e d a t i o n and d i s t u r b a n c e i n t e n s i t i e s . E s t u a r i e s 6:372-379.
Munroe, T.A., and R.A. Lotspeich. 1979. Some 1 i f e h i s t o r y a s p e c t s o f t h e seaboard goby (Gobiosoma qinsburqi ) i n Rhode Is1 and. Es tuar ies 2:22-27.
Naughton, S.P. , and C.H. Saloman. 1981. Stomach contents of juveni les o f king mackere l (Scomberomorus c a v a l l a ) and S p a n i s h mackerel (S. macul a t u s ) . Northeast Gulf Sci . 5:71-74.
Neal W.J., W.C. Blakeney, J r . , O . H . Pi lkey, Jr. , and O.H. Pilkey, Sr . 1984. Living with t h e South Carolina shore. Duke U n i v e r s i t y P r e s s , Durham, North Carolina. 205 pp.
Ne? son, B . V . , and R.R . Vance. 1979. Die1 f o r a g i n g p a t t e r n s o f t h e s e a u r c h i n C e n t r o s t e ~ h a n u s coronatus a s a p reda to r avoidance s t r a t e g y , Mar. BS 01. 51 : 251 - 258.
Nel son, W . G . 1979. Experimental s t u d i e s o f s e l e c t i v e p r e d a t i o n on amphi pods: consequences f o r amphi pod d i s t r i b u t i o n and abundance, J. Exp. Mar. B i 01 . Ecol . 38: 225-245.

Ne lson , W.G. 1980a. The b i o l o g y o f ee l g rass (Zos te ra mar ine L , ) amphi pods. Crustaceana 39:59-89.
Nelson, W.G. 1980b. A comparat ive s t udy o f amphipods i n seagrasses f rom F l o r i d a t o Nova S c o t i a . B u l l . Mar. Sc i . 30:80- 89.
Nelson, W.G. 1981. Experimental s t u d i e s o f decapod and f i s h p r e d a t i o n on seagrass macrobenthos. Mar. B i o l . Prog. Ser. 5:141-149.
N i c o t r i , M.E. 1977 . The i m p a c t o f c r u s t a c e a n h e r b i v o r e s on c u l t u r e d seaweed p o p u l a t i o n s . A q u a c u l t u r e 12:127-136.
N i c o t r i , M.E. 1980. Fac to rs i n v o l v e d i n h e r b i v o r e f ood p re fe rence . J. Exp. Mar. B i o l . Ecol . 42:13-26.
Norton, T.A., and M.R. Benson. 1983. E c o l o g i c a l i n t e r a c t i o n s be tween t h e brown seaweed Sarqassum muticum and i t s assoc ia ted fauna. Mar. B i o l . 75: 169- 177.
Nummedal, D., G.F. Oe r t e l , D.K. Hubbard, and A.C. Hine. 1977. T i d a l i n l e t v a r i a b ~ l i t y - - C a p e H a t t e r a s t o Cape Canaveral . Coas ta l sediments '77. p roc . F i f t h Symp. WPCO D i v . Am. Soc. C iv . Eng. 1977:543-562.
Odum, W.E., and E. J. H e a l d . 1972 . T r o p h i c a n a l y s i s o f an e s t u a r i n e mangrove commun i t y . B u l l Mar . S c i . 22:671-738.
Bgburn, M . V . 1984. Feeding eco logy and t h e r o l e o f a lgae i n t h e d i e t o f t h e sheepshead Archosarqus probatocephalus (P isces: Spar idae) on two No r t h C a r o l i n a j e t t i e s . M.S. Thes is . U n i v e r s i t y o f No r t h C a r o l i n a a t Wi lmington. 68 pp.
Ogden, J.S., and P.S. Lobel . 1978. The r o l e o f he rb i vo rous f i shes and u r c h i n s i n c o r a l r e e f communit ies. Envi ron . B i o l . F ish . 3:49-63.
O l l a , B.L., A.J. Bejda, and A.D. M a r t i n . 1974. D a i l y a c t i v i t y , movements, feedins, and seasonal occurrence i n t h e t a u t o g , Taut.asq o n i t i s . F i s h . B u l l . 72:27-35,
Ortega, S. 1981. Environmental s t r e s s , compe t i t i on and dominance o f Crassos t rea v i r q i n i c a near Beau fo r t , No r t h Ca ro l i na , USA. Mar. B i o l . 62:47-56.
Overs t ree t , R.M., and R.W. Heard. 1978a. Food o f t h e r e d drum, Sciaenops o c e l l a tus f rom M i s s i s s i p p i Sound. G u l f Res. Rep. 6:131-135.
Overs t ree t , R.M., and R.W. Heard. 1978b. Food con ten t s o f s i x commercial f i s h e s f rom M i s s i s s i p p i Sound. G u l f . Res. Rep. 7:137-149.
Overs t ree t , R.M., and R.W. Heard. 1982. Food con ten t s o f s i x commercial f i s h e s f rom M i s s i s s i p p i Sound. G u l f Res. Rept. 7: 137-149.
Paine, R.T. 1966. Food web comp lex i t y and s p e c i e s d i v e r s i t y . Am. N a t . 100:65-75.
Paine, R.T. 1974. I n t e r t i d a l community s t r u c t u r e : exper imenta l s t u d i e s on t h e r e l a t i o n s h i p be tween a d o m i n a n t compe t i t o r and i t s p r i n c i p a l p reda to r . Oecolog ia ( B e r l . ) 15:93-120.
Paine, R.T. 1980. Food webs: l i n kage , i n t e r a c t i o n s t r e n g t h and commun i ty i n f r a s t r u c t u r e . J. Anim. Ecol . 29: 667-685.
Parchure, T.M. 1982. S t . Marys en t rance . Glossary o f I n l e t s Report 11. Dep. Coasta l Ocean. Eng., Univ . F l a. F l a. Sea Grant Col 1 . 46 pp.
Parker, R.O., Jr., R.B. Stone, and C.C . Buchanan. 1979. A r t i f i c i a l r e e f s o f f M u r r e l l s I n l e t , South Carol i na . Mar. F i sh . Rev. 1979:12-24.
Paul, V . J . , M.E. Hay, J.E. Du f f y , W . Fen ica l , and K. Gustafson (1987). Chemical defense i n t h e seaweed Ochtodes s e c u n d i r a m e a ( M o n t a g n e ) H o w e (Rhodophyta) : e f f e c t s o f i t s monoter- penoid components upon d i v e r s e c o r a l r e e f he rb i vo res . J. Exp. Mar. B i o l . Ecol . 114:249-260.
Pearse , J .S. , and A.H. H i n e s . 1979. Expansion o f a c e n t r a l C a l i f o r n i a k e l p f o r e s t f o l l o w i n g t h e mass m o r t a l i t y of sea u r ch i ns . Mar. B i o l . 51:83-91.

Pearson, J .C. 1929. Natural h i s t o r y and c o n s e r v a t i o n o f r e d f i s h and o t h e r commercial sc iaenids on the Texas coas t . Bull . U.S. Bur. Fish. 44:129-214.
Pea r son , T.G., C.S. Br imley , and H . H . Br imley . 1942. B i r d s o f North Carolina. 2nd ed. Bynum P r i n t i n g Co., Raleigh. 434 pp.
Peckol , P. 1982. Seasonal occurrence and reproduction of some marine a lgae o f t h e Continental Shel f , North Carol ina . Bot. Mar. 25:185-190.
P e c k o l , P . , and R . B . S e a r l e s . 1983. Effec ts of s easona l i t y and d i s tu rbance on t h e p o p u l a t i o n development i n a Carol ina Continental Shel f community. Bull . Mar. Sci . 33:67-86.
Peterson, C . H . , and N . M . Peterson. 1979. The ecology of i n t e r t i d a l f l a t s o f North Carol i na: a community prof i 1 e U . S. Fish and Wild l i fe Serv ice , O f f i c e of Biological Serv ices . FWS/OBS-79/39. 73 PP-
P f i s t e r , C .A. 1987. Associat ional p l a n t r e f u g e s : c o n v e r g e n t p a t t e r n s i n t e r r e s t r i a l and marine communities a r e t h e r e s u l t o f d i f f e r i n g mechanisms. M.S. T h e s i s . U n i v e r s i t y o f North Carolina a t Chapel H i l l . 39 pp.
Pi lkey, O.H.,Jr. , O.H. P i lkey ,Sr . , and R. Turner. 1975. How t o l i v e wi th an i s l and . A handbook t o Bogue Banks, North C a r o l i n a . North C a r o l i n a Department o f Na tu ra l and Economic Resources, Raleigh. 119 pp.
i l key , O . H . , J r . , W.J. Neal, O.H.Pilkey, S r . , and S.R. Riggs . 1980. From C u r r i t u c k t o Ca labash . L i v i n g w i t h North Carol i na ' s b a r r i e r i s l ands . Duke U n i v e r s i t y P r e s s , Durham, North Carolina. 245 pp.
P i l k e y , O . H . , J r . , D . C . Sharma, H . R . Wanless, L.J. Doyle, O.H. Pi1 key, Sr . W.J. Neal , and B . L . Gruver . 1984. L iv ing w i t h t h e Eas t F l o r i d a s h o r e . Duke U n i v e r s i t y P r e s s , Durham, North Carol ina. 259 pp.
Ramus, J . , and M. Venable. 1987. Temporal ammoni um pa tchiness and growth
r a t e i n :odium and Ulva (Ulvophyceae). J . Phycol 23: 518-523.
Randall, J E 1967. Food h a b i t a t s o f r ee f f i s h e s of the West Indies . S tudies i n ~ r o p i c a l Oceanography (Miami) . 5:665-847.
Randall, J . E . , and W.D. Hartman. 1968. S p o n g e - f e e d i n g f i s h e s o f t h e West Indies . Mar. Biol . 1:216-225.
Rauck, G . , and 3 .5 . Zi j l s t r a . 1978, On t h e n u r s e r y aspec t of the Wadden Sea f o r some c o m m e r c i a l f i s h s p e c i e s and p o s s i b l e long- term changes. Rapp. P. V . Reun. Cons. I n t . Explor. Mer. 172:266- 275.
Reed, D . C . , and M.S. Fos ter . 1984. The e f f e c t s o f canopy s h a d i n g on a l g a l r e c r u i t m e n t and growth in a g i a n t kelp f o r e s t . Ecology 65:937-948.
Reid, G.K., A. I n g l i s , and H . D . Hoese. 1956. Summer foods of some f i s h spec ie s i n East Bay, Texas. Southwest Nat. 1: 100-104.
Richards, S.W. 1976. Age, growth, and food o f b l u e f i sh ( Pomatomus sa l t a t r i x ) from e a s t - c e n t r a l Long Is1 and sound from J u l y t h r o u g h November 1975. T r a n s . Amer. F i s h . Soc. 105:523-525.
R i c h a r d s , S . W , 1979. I c h t h y o p l a n k t o n abundance and d i v e r s i t y i n t he e a s t e r n Gulf o f Mexico. Report t o t he Bureau of Land Management under C o n t r a c t No. AA550-CT7-28, June 1979. 546 pp.
R i c h a r d s o n , J . P . 1978. E f f e c t s o f e n v i r o n m e n t a l f a c t o r s on t h e l i f e h i s t o r i e s and s e a s o n a l i t y o f some inshore b e n t h i c marine a lgae i n North C a r o l i n a . P h . D . D i s s e r t a t i o n . U n i v e r s i t y o f North Carolina a t Chapel H i l l . 141 pp.
Richardson, J . P. 2979. Overwintering s f Dic tvota dichotoma (Phaeophyceae) near i t s n o r t h e r n d i s t r i b u t i o n l i m i t on t h e e a s t Coas t of North America. J. Phycoj . 15:22-26.
Richardson, 3 , P. 1981. Pe r s i s t ence and d e v e l o p m e n t o f Dasya bai % I ouv iana (Gemel j n ) Montagne (Rhodophyceae,

Dasyaceae) i n N o r t h Caro l i na. Phycol ogia 20:385-391.
Richardson, J.P. 1982. L i f e h i s t o r y o f B r v o ~ s i s plumosa (Hudson) Agardh (Chl orophyceae) i n North Carol ina, USA. Bot. Mar. 25:177-183.
Robins, C.R., G.C. Ray, and J. Douglas. 1986. A f i e l d guide t o A t l a n t i c coast f ishes o f North America. Peterson F i e l d Guide S e r i e s No.32. Houghton M i f f l i n Co., Boston. 354 pp.
Schwartz, F.J., and B.W. Dutcher. 1963. Age, growth, and food o f the oys ter t o a d f i sh n e a r Sol omans, Mary l and. Trans. Amer. F ish . Soc. 92:170-173.
Scot t , S.L., L.M. Swinson, M.B. Dickinson, and C.H. Howell. 1983. F i e l d guide t o t h e b i r d s o f North America. Nat. Geo. Soc., Washington, DC. 463 pp.
Searles, R.B. 1981. Seaweeds from Gray's Reef , Georg ia . N o r t h e a s t Gul f S c i . 5:45-48.
Roelofs, E.W. 1954. Food s tud ies of Searles, R.B. 1984. Seaweed biogeography young sc iaen id f i shes, M ic ro~oqon and o f the m id -A t lan t i c coast o f the Uni ted Leiostomus, from North Carol ina. Copeia States. Helgol . Meeresunters 38:259- 1954:151-153. 271.
Rosenberg, G., and H.W. Paer l . 1980. N i t r o g e n f i x a t i o n by b l uegreen a1 gae a s s o c i a t e d w i t h t h e s iphonous g reen seaweed Codi um d e c o r t i catum: e f f e c t s on ammonium uptake. Mar. B i o l . 61 : 151- 158.
Sale, P.F. 1980. The ecology o f f i shes on co ra l ree fs . Oceanogr. Mar. B i o l . Annu. Rev. 18:367-421.
Santel ices, B., and F.P. Ojeda. 1984. E f f e c t s o f canopy removal on t h e understory a lga l community s t r u c t u r e o f coastal f o r e s t s o f Macrocvsti s o v r i f e r a from southern South America. Mar. Ecol. Prog. Ser. 14: 165-173.
Schne ider , C.W. 1975. S p a t i a l and temporal d i s t r i b u t i o n o f benthic marine algae on t h e Cont inental She l f o f the Carol inas. Ph .D. Disse r ta t i on . Duke Univers i ty , Durham, North Carol ina. 196 PP.
Schne ider , C.W. 1976. S p a t i a l and temporal d i s t r i b u t i o n o f benth ic marine algae on t h e Cont inental She l f o f t he Carol inas. B u l l . Mar. Sci . 26:133-151.
Searles, R.B., M.H. Hommersand, and C.D. Amsler. 1984. The occurrence o f Codium f r a q i l e subs. tomentoso ides and C. t a v l o r i i i n North Carol ina. Bot. Mar. 27:185 187.
Searles, R.B., and C.W. Schneider. 1978. A check1 i s t and b i b l iography o f North Carol i na seaweeds. Bot. Mar. 21 :99- 108.
Searles, R.B., and C.W. Schneider. 1980. Biogeographic a f f i n i t i e s o f the shal low and deep water benth ic marine algae o f N o r t h C a r o l i n a . B u l l . Mar. S c i . 30:732-736.
Sears, J.R. 1971. Morphology, systematics and d e s c r i p t i v e ecology o f t h e sub1 i t t o r a l benthic marine algae o f southern Cape Cod and adjacent i s l ands . Ph.D. D i s s e r t a t i o n . U n i v e r s i t y o f Massachusetts, Amherst. 273 pp.
Sedbesry, G.R., and R.F. Van Dolah. 1984. Demersal f i s h assemblages a s s o c i a t e d w i t h hard bottom hab i ta t s i n t h e South A t l a n t i c B ight o f t he U.S.A. Environ. B i o l . Fishes 11:241-258.
Schneider, C.W., and R.B. Searles. 1979. Sher idan, P.F., and D.L. Trimm. 1983. Standing crop o f benth ic seaweeds on the Summer foods o f Texas coastal f i shes Caro l ina Cont inental She l f . Proc. I n t . r e l a t i v e t o age and h a b i t a t . Fish. Seaweed Symp . 9: 293-301. B u l l . 81 :643-647.
Schwartz, F.J. 1964. Fishes o f t he I s l e Simenstad, C.A., J.A. Estes , and K.W. of Wight and Assawoman Bays near Ocean Kenyon. 1978. Aleuts, sea o t t e r s , and City, Mary1 and. Chesapeake Sci . 5: 172- a1 t e r n a t e s t a b l e - s t a t e communi t ies . 193. Science 200:403-411.

Smith, S.M., J.G. Hoff, S.P. OrNeil , and M.P. Weinstein. 1984. Community and t rophic organization of nekton u t i l izing sha l low marsh h a b i t a t s , York River , Virginia. Fish. Bull. 82:455-467.
Sousa, W.P. 1979. Experimental i n v e s t i g a t i o n s o f d i s t u r b a n c e and e c o l o g i c a l s u c c e s s i o n i n a rocky i n t e r t i d a l a l g a l communi t y . Ecol . Monogr . 49: 227-254.
Spitsbergen, J.M. 1980. Seacoast 1 i f e . An ecological guide t o natural seashore communities in North Carol ina. Univ. N . C . Press, Chapel H i l l . 112 pp.
Springer, V . G . , and K.D. Woodburn. 1960. An ecological study of the f i s h e s of the Tampa Bay area. Fl a. Board Conserv. Mar. Res. Lab. Prof. Pap. Ser. 1. 104 pp.
Stefansson, U. , L.P. Atkinson, and D.F. Bumpus. 1971. Hydographi c proper t ies and c i rcu l a t ion of t h e North Carol ina shel f and slope waters. Deep Sea Res. 18:383-420.
Steimle, F.W., J r . , and L. Ogren. 1982. Food of f i s h co l l ec ted on a r t i f i c i a l r e e f s in the New York Bight and o f f Charleston, South Carolina. Mar. Fish. Rev. 44:49-52.
Steneck , R.S., and L . Wat l ing. 1982. Feeding capabil i t i e s and 1 imi ta t ions of he rb ivorous moll uscs : a f u n c t i o n a l group approach. Mar. Bi 01. 68: 299-319.
Stephenson, T.A., and A . S tephenson. 1952. Life between tidemarks in North America, Part 2. Northern Florida and the Carolinas. J. Ecol. 40:l-49.
Stephenson, T.A., and A. S tephenson. 1972. Life between tidemarks on rocky s h o r e s . W . H . Freeman and Co., San Francisco. 425 pp.
S t i c k n e y , R . R . , G.L. T a y l o r , and D . B . White. 1975. Food habi ts of f i v e s p e c i e s o f young s o u t h e a s t e r n United S ta tes es tuar ine Sciaenidae. Chesapeake Sci . 16:104-114.
Stokes, T . , and K. Shackleton. 1968. Birds of t h e At lan t i c Ocean. Macmillan Co., New York. 156 pp.
Stoner, A.M. 1980a. Feeding ecology o f Lasodon rhomboides (pi sces : Sparidae) : v a r i a t i o n and f u n c t i o n a l r e sponse . Fish. Bull. 78:337-352.
Stoner, A.M. 1980b. The r o l e of seagrass biomass in t h e organization of benthic macrofaunal assembl ages . Bu l l . Mar. Sci . 30: 537-551.
S t o n e r , A.W. 1980c. Abundance, r e p r o d u c t i v e seasona l i t y and h a b i t a t preferences of amphipod crustaceans i n s e a g r a s s meadows of Apalachea Bay, Florida. Cont. Mar. Sci . 23:63-77.
Stoner, A.W., and R.J. Livingston. 1984. Ontogene t i c p a t t e r n and f e e d i n g morphology in sympatric spar id f i s h e s ~.
from s e a g r a s s meadows .' Copei a 1984: 174-187.
Sutherland, J.P. 1974. Multiple s t a b l e points i n natural communities. Am. Nat. 108:859-873.
Sutherl and, J.P. 1977. Effect of S h i z o ~ o r e l i a removal i n the foul ing community a t Beaufort, N.C. Pages 155- 189 in B.C. Coull, ed. Ecology of marine benthos. University of South Carol ina Press, Col umbi a.
Sutherland, J.P. 1978. Functional r o l e s o f S h i z o a o r e l l a and S t v e l a i n t h e f o u l i n g community a t Beaufor t , North Carol ina. Ecology 59~257-264.
S u t h e r l and, J. P. 1981. The f o u l i ng community a t Beaufort, North Carol ina: a s t u d y i n s t a b i l i t y . Am. Nat. 118:499-519.
Sutherland, J.P., and R.H. Karlson. 1977. Development and s t a b i l i t y of the foul ing community a t Beaufort, North Carol i na. Ecol . Monogr. 47: 425-446.
Taylor, W.R. 1960. The marine algae of t h e e a s t e r n t r o p i c a l and s u b t r o p i c a l coas ts of the Americas. University of Michigan Press, Ann Arbor. 870 pp.
Tegner, M.J., and D.K. Dayton. 1977, Sea u rch in r e c r u i t m e n t p a t t e r n s and i m p l i c a t i o n s of commercial f i s h i n g . Science 196: 324-326.

Tegner , M.J., and P . K . Dayton. 1981. Population s t r u c t u r e , recru": tment and m o r t a l i t y o f two s e a u r c h i n s (Stronovlocentrotus f r anc i scanus and & p u r ~ u r a t u s ) in a kelp f o r e s t . Mar. Ecol . Prog . Ser . 6 : 223 - 228.
Thayer, G . W . , D. E. Hoss, M.A. Kjel son, W.F. H e t t l e r , J r . , and M . W . La Croix. 1974. Biomass o f zoop lank ton i n t h e Newport River e s tua ry and t h e inf luence of post1 arval f i s h e s . Chesapeake Sci . 15:9-16.
Thayer, G.W., S.M. Adams, and M . V . La Croix. 1975. S t ruc tu ra l and funct ional a s p e c t s o f a r e c e n t l y e s t a b l i s h e d ~ o s t e r a mar ina community. E s t u a r i n e Res. 1:517-540.
Thaye r , G . W . , W.J. Kenworthy, and M.S. Fonseca. 1984. The ecology o f ee lg ra s s meadows o f t h e A t l a n t i c c o a s t : a community p r o f i l e . U.S. Fish. Wildl. Serv. FWS/OBS-84/02. 147 pp.
Turner, C . H . , E . E . Ebert, and R. R . Given. 1969. Man-made reef ecology . Cal i f . Dep. Fish Game Fish Bull. 146:l-221.
Eng. Res. Cent . Waterw. Exp. S t n . , Vicksburg, Miss. 143 pp,
Van Dolah, R . F . , P.H. Wendt, C . A . Wenner, R.M. Martore, G . R . Sedberry, and C.A. Moore. 1986. Eco log ica l e f f e c t s o f r u b b l e w e i r j e t t y c o n s t r u c t i o n a t Murrell s In1 e t , South Carol i na. Vol ume 111: Community s t r u c t u r e and h a b i t a t u t i l i z a t i o n o f f i s h e s and c r a b s assoc ia ted with t h e j e t t i e s . U.S. Army Corps Eng. Tech. Rep. EL-84-4. Coastal Eng. Res. C e n t . , Waterw. Exp. S t n . , Vicksburg, Miss. 150 pp.
Van Dover, C . , and W . Kirby-Smith. 1979. F i e l d g u i d e t o common mar ine inve r t eb ra t e s o f Beaufort, NC. Pa r t 1. Gas t ropods , b i v a l v e s , amphi pods decapods , and echinoderms. Duke Universi ty Marine Laboratory, Beaufort. 78 PP-
Vine, P.J. 1974. The e f f e c t of a lga l grazing and aggressive behavior o f t h e f i s h e s Pomacentrus 1 i v i d u s and Acanthurus sohal on cora l r e e f ecology. Mar. Biol . 24:131-136.
U . S . Army Corps o f E n g i n e e r s . 1984. Wang, J . C . S . , and E . C . Raney. I971 Shore p r o t e c t i o n manual. 2 Vo l s . D i s t r i b u t i o n and f l u c t u a t i o n s i n t h e C o a s t a l Eng inee r ing Research C e n t e r , f i s h f a u n a o f t h e C h a r l o t t e Harbor Department o f t h e Army, Waterways e s t u a r y , F l o r i d a . C h a r l o t t e Harbor Experiment S t a t i o n , V icksburg , Estuarine Study, Mote Marine Laboratory, Mississippi . Sarasota , F lor ida . 56 pp.
U . S. Department of Commerce. 1987. Tide t a b l e s 1987. East Coast of North and South America. U.S. Department o f Commerce, Na t iona l Oceanic and Atmospher ic A d m i n i s t r a t i o n , Na t iona l Ocean Service . 289 pp.
Vance, R.R. , and R.J. Schmitt. 1979. The e f f e c t s o f t h e p r e d a t o r avo idance behav iou r o f t h e s e a u r c h i n Centrosteohanus coronatus on t h e breadth o f i t s d i e t . Oecologia (Ber l . ) 44:21- 25.
Van Dolah, R . F . , D.M. Knott, and D . R . Calder. 1984. Ecological e f f e c t s of r u b b l e w e i r j e t t y c o n s t r u c t i o n a t Murrel I s I n l e t , South Carol i na; Vol ume I : C o l o n i z a t i o n and community development on new j e t t i e s . U.S. Army Corps Eng. Tech. Rep, EL-84-4, Coastal
Whal i n , R . W . , F. E . Camf i e l d, N . E . Parker, R .A . Jachowski, and J.R. Weggel. 1984. Shore pro tec t ion manual. Vol s . 1 and 2. Dept . Army, Waterways Experiment S t a t i o n , Corps of Engineers, Coastal Engineering Research Center . U.S. Gov. P r i n t . Off . , Washington, DC. 597 PP .
Williams, D. M . 1980. Dynamics of t h e pomacent r id community on smal l p a t c h r e e f s in One Tree Lagoon (Great Ba r r i e r Reef). Bull . Mar. Sc i . 30:159-170.
i l l a m s , . M . , and P.F. Sa l e . 1981. S p a t i a l and tempora l p a t t e r n s of r e c r u i t m e n t o f j u v e n i l e c o r a l r e e f f i s h e s t o coral h a b i t a t s within "One Tree Lagoon", Great Ba r r i e r Reef. Mar. Biol . 65:245-253.

Williams, L.G. 1948. Seasonal alterations of indr.ine floras at Cape Lookout, North Carolina. Am. J , Bot. 35:682-695.
Williams, L . 6 . 1949. Marine algal ecology at Cape Lookout, North Carolina. Bull. Furman Univ. 31:l-21.
Wiseman, D.R. 1978. Benthic marine algae. Pages 23-36 in R.G. Zingmark, ed. An annotated checklist of the biota of the coastal zone of South Carolina. University of South Carolina Press, Col umbi a.
Wiseman, D.R., and C.W. Schneider. 1976. Investigations of the marine algae of South Carolina. I. New records of Rhodophyta. Rhodora 78:516-524.
Wells, H.W. 1961. The fauna of oyster beds, with special reference to the salinity factor. Ecol. Monogr. 31:239- 266.
Wells, H.W., and I.E. Gray. 1960. Some oceanic subtidal oyster populations. Nautilus 73:139-146.
Wells, H.W., M.J. Wells, and I.E. Gray. 1960. Marine sponges of North Carolina. J. Eiisha Mitchell Scf. Soc. 76:200- 245.
Wells, H.W., M.J. Wells, and 1-E. Gray. 1964. Ecology of sponges in Hatteras Harbor, North Carolina. Ecology 45:752-767.
Wenner, C.A., W.A. Roumillat, and C.W. Waltz. 1986. Contributions to the life
hi story o f black seabass, Centro~ristis stri ata, off the southeastern United States. Fish. Btrll . 84:723-741.
Wethey, D.S. 1983. Geographic limits and local zonation: the barnacles Seniibal anus (6alanus) and Chthamalus in .- New England. Biol . Bull. 165:330-341.
Wethey, D.S. 1984. Sun and shade mediate comneti t i nq in the barnacles Chthamal us and Semi bal anus: a field experiment. Biol. 6ull. 167:176-185.
Wood, 1. 1968. Physiological and ecological aspects of prey selection by the marine gastropod Urosal ~ i n x cinerea (Prosobranchi a: Muricidae) . Ma1 acologi a 6:267-320.
Young, D.K., M.A. Buzas, and M.W. Young. 1976. Species densities of macrobenthos associated with seagrass: a field experimental study of predation. J . Mar. Res. 34:577-592.
Young, D.K., and M.W. Young. 1978. Regulation of species densities of seagrass associated macrobenthos: evidence from field experiments in the Indian River estuary, Florida. J. Mar. Res. 36:569-593.
Zimmerman, R., R. Gibson, and J. Harrington. 1979. Herbivory and detri tivory among gammaridean amphi pods from a Florida seagrass community. Mar. Biol. 54:41-47.
Zingmark, R.G., ed. 1978. An annotated checklist of the biota of the coastal zone of South Carolina. Univ. South Carol ina Press, Col umbia. 364 pp.

110272 -101
REPORT DOGUMENTATtON ' '. 2. 3 Rs~tplsnt s Accession No
PAGE I Biological Report 85(7.20) 1 -- . - ----- --- 4. T~t le end Subtttle -I--?% Report Date i The Ecology of Rubble Structures of The South Atlantic Bight: A Community Profile
7. Author(*)
Mark E. Hays and John P. Sutherlandb -- - -- --------- 9- Authors' addresses : 'University of North Carolina at Chapel Hill b~uke University Institute of Marine Sciences Marine Laboratory Morehead City, NC 28557 Beaufort, NC 28516
/ September, 1988 6.
8. Performing Organiretion Rapt. Wo.
10. PmlectlTask/Worli Unit No.
11. ContractfC) or Grant(C) No.
IC) I - -- I__------ - _._I -____^._/
12. Sponronng Or..nltallon Name and Addmss (G)
1 I U.S. Department of the Interior Fish and Wildlife Service National Wetlands Research Center Washington, DC 20240
13. Typ. 01 Report a Period 12-4
-.-~ 14.
I 16. Abrtrbct (Limit: 2 0 words) I This community profile provides an introduction to the ecology of the communities living on and around rubble structures in the South Atlantic Bight (Cape Hatteras to Cape Canaveral). The most prominent rubble structures in the bight are jetties built at the entrances to major harbors. After an initial discussion of the various kinds of rubble structures and physical factors that affect the organisms associated with them, the major portion of the text is devoted to the ecology of rubble structure habitats. Community composition, distribution, seasonality, and the recruitment patterns of the major groups of organisms are described. The major physical and biological factors affecting the organization of intertidal, sunlit subtidal, and shaded subtidal communities are presented and the potential effects of complex interactions in structuring these communities are evaluated. The profile concludes with a general review of the effects of rubble structures on nearshore sediment dynamics and shore1 ine evolution.
South Atlantic Bight hard substrate communities South Carolina jetties fouling communities Georgi a groins encrusting organisms northern Florida rubble structures North Carolina
Algae anemones barnacles fish
I& Avr&labtftry StLEwmaoe
Unlimited distribution , 19. Sscur*ty Clarr (Thrl Report) 21. No of Pager
i Unclassified 1 i x t 6 7 _i_____ I--- -- -----
8 20. Swurtty C l r s ~ (Thls PagcJ 1 9. Prlcs I I Unclassified 1
( S w ANSt-LJ9.18) OCTtONAL CORM 272 (4-37) (Formarty NYlS-35) Department ot Commanze


















