
The distribution of mutagenic activity in red, rose and white wines
8
Mutation Research, 223 (1989) 205-212 205 Elsevier MTR 01406 The distribution of mutagenic activity in red, rose and white wines Tai Nguyen, Larry Fluss, Roberta Madej, Charles Ginther and Terrance Leighton Department of Microbiology and Immunology, University of California, Berkeley, CA 94720 (U.S.A.) (Received 22 July 1988) (Revision received 5 December 1988) (Accepted 22 December 1988) Keywords: Wine; Ames test, wine Summary Using a modified Salmonella typhimurium TA98 Ames-test system, more than 150 red, white and rose wines were analyzed for direct-acting and microsomal enzyme-enhanced mutagenic activity. The following conclusions were reached from analysis of this wine mutagenicity data base. White and rose wines, as well as grape juices, exhibited little or no detectable direct-acting or microsomal enzyme-enhanced mutagenic activity. However, red wine samples contained highly variable amounts of mutagens, ranging from undetectable to levels 30-fold above the sensitivity limit of the assay system. The variations in red wine mutagenicity were unrelated to grape variety, vintage, aging methods or production region. Hence, individual winery production practices must represent the most significant contribution to the variations observed. Damage to DNA by environmental agents is inferred to be a highly significant factor in de- termining the incidence of human cancer (Hiatt et al., 1977; Ames, 1979; Doll and Peto, 1981; Peto and Schneiderman, 1981; Ames, 1986). Epidemio- logical evidence suggests that no more than 30% of human cancers can be accounted for by the effects of tobacco (Knudson, 1977; Wynder and Gori, 1977; Higginson and Muir, 1979; Doll and Peto, 1981; Peto and Schneiderman, 1981). Hence, other environmental factors, such as diet, alcohol and beverage consumption, etc., are presumed to be of primary importance in the etiology of hu- Correspondence: Dr. Terrance Leighton, Department of Mi- crobiology and Immunology, University of California, Berke- ley, CA 94720 (U.S.A.) (415) 642-3033. man cancer (Ames, 1979; Doll and Peto, 1981; Peto and Schneiderman, 1981; Ames, 1986). The development of rapid bacterial mutagene- sis assays by Ames and coworkers, and others (Ames et al., 1975; Hollstein et al., 1979), has added enormously to our knowledge of which naturally occurring components ingested by hu- mans possess genotoxic activity (Ames, 1979, 1986; Miller et al., 1979; Brown, 1980; Hirono, 1981, 1987; Hoey et al., 1981). In an investigation of the mutagenicity of various natural food and beverage products, Ames and coworkers (Tamura et al., 1980) reported that two red wines possessed di- rect-acting mutagenic activity, while two white wines exhibited no mutagenic activity. Stavric and colleagues (1983) also reported detecting muta- genic activity in several red wines. Given that nearly 9 billion gallons of wine are produced and consumed annually (Amerine et al., 0165-1218/89/$03.50 © 1989 Elsevier Science Publishers B.V. (Biomedical Division)
-
Upload
tai-nguyen -
Category
Documents
-
view
212 -
download
0
Transcript of The distribution of mutagenic activity in red, rose and white wines

Mutation Research, 223 (1989) 205-212 205 Elsevier
MTR 01406
The distribution of mutagenic activity in red, rose and white wines
Tai Nguyen, Larry Fluss, Roberta Madej, Charles Ginther and Terrance Leighton Department of Microbiology and Immunology, University of California, Berkeley, CA 94720 (U.S.A.)
(Received 22 July 1988) (Revision received 5 December 1988)
(Accepted 22 December 1988)
Keywords: Wine; Ames test, wine
Summary
Using a modified Salmonella typhimurium TA98 Ames-test system, more than 150 red, white and rose wines were analyzed for direct-acting and microsomal enzyme-enhanced mutagenic activity. The following conclusions were reached from analysis of this wine mutagenicity data base. White and rose wines, as well as grape juices, exhibited little or no detectable direct-acting or microsomal enzyme-enhanced mutagenic activity. However, red wine samples contained highly variable amounts of mutagens, ranging from undetectable to levels 30-fold above the sensitivity limit of the assay system. The variations in red wine mutagenicity were unrelated to grape variety, vintage, aging methods or production region. Hence, individual winery production practices must represent the most significant contribution to the variations observed.
Damage to DNA by environmental agents is inferred to be a highly significant factor in de- termining the incidence of human cancer (Hiatt et al., 1977; Ames, 1979; Doll and Peto, 1981; Peto and Schneiderman, 1981; Ames, 1986). Epidemio- logical evidence suggests that no more than 30% of human cancers can be accounted for by the effects of tobacco (Knudson, 1977; Wynder and Gori, 1977; Higginson and Muir, 1979; Doll and Peto, 1981; Peto and Schneiderman, 1981). Hence, other environmental factors, such as diet, alcohol and beverage consumption, etc., are presumed to be of primary importance in the etiology of hu-
Correspondence: Dr. Terrance Leighton, Department of Mi- crobiology and Immunology, University of California, Berke- ley, CA 94720 (U.S.A.) (415) 642-3033.
man cancer (Ames, 1979; Doll and Peto, 1981; Peto and Schneiderman, 1981; Ames, 1986).
The development of rapid bacterial mutagene- sis assays by Ames and coworkers, and others (Ames et al., 1975; Hollstein et al., 1979), has added enormously to our knowledge of which naturally occurring components ingested by hu- mans possess genotoxic activity (Ames, 1979, 1986; Miller et al., 1979; Brown, 1980; Hirono, 1981, 1987; Hoey et al., 1981). In an investigation of the mutagenicity of various natural food and beverage products, Ames and coworkers (Tamura et al., 1980) reported that two red wines possessed di- rect-acting mutagenic activity, while two white wines exhibited no mutagenic activity. Stavric and colleagues (1983) also reported detecting muta- genic activity in several red wines.
Given that nearly 9 billion gallons of wine are produced and consumed annually (Amerine et al.,
0165-1218/89/$03.50 © 1989 Elsevier Science Publishers B.V. (Biomedical Division)

206
1980), at least one-third of which is red wine, it would seem important to understand in much greater detail the origin, distribution and molecu- lar identity of mutagens found in wine. To this end we have initiated a comprehensive survey of direct-acting and microsomal enzyme-enhanced frameshift mutagenic activity in red, white and rose wine types.
Materials and methods
Wine and grape juice samples. More than 150 commercially produced red, white and rose wines, as well as grape juices, were purchased or col- lected as winery samples. Representative wines from Australia, Bulgaria, France, Greece, Hunga- ry, Israel, Italy, Spain and the United States were selected. These samples included wines produced from the following grape varieties: Barbara, Ca- bernet Franc, Cabernet Sauvignon, Chardonnay, Concord, Corvina, Gamay, Kadarka, Merlot, Nebbiolo, Petite Sirah, Pinot Nob, Sangiovese, Tempranillo and Zinfandel. Vintages spanned a period of 20 years. When possible, varietal wines from different producers within the same region were examined.
Sample preparation and concentration. Meth- ods of wine or juice sample preparation and con- centration, developed by Ames and coworkers, were employed (Yamasaki and Ames, 1977; Tamura et al., 1980). 50 ml of wine or juice were passed through a 1.75 ml XAD-2 polystyrene bead column (Isolab Inc.) The column was eluted with 10 ml of acetone, and the acetone fraction dried under nitrogen at 60-70 ° C. The dried residue was suspended in 1 ml of dimethyl sulfoxide (DMSO, Kodak spectrograde) and assayed for mutagenic activity in the TA98 Ames Test System, using duplicate samples.
The efficiency of mutagen recovery by XAD-2 column chromatography was assessed by internal standard methods. Known amounts of quercetin (16.7 mg/1) were added to 12% ethanol solutions (pH 3.5), or to white wine samples which lacked intrinsic mutagenic activity. Quercetin is a direct-acting mutagen known to exist in the skin of red grapes and in red wines (Brown, 1980;
Brown and Dietrich, 1979; Nagao et al., 1981), and has similar chromatographic and mutagenic properties to wine mutagens. Quercetin levels were measured using UV spectrophotometric analysis of the XAD-2 column elutant. Quercetin also served as an internal mutagenicity standard for the calibration of Ames-test assays.
Standardization of the Ames-test system for the quantitation of wine mutagens. Establishing a mutagenicity data base for more than 150 wines required comparing Ames-test system measure- ments obtained over a period of several years. To facilitate such comparisons, the statistical detec- tion and reproducibility limits of this assay system were determined. Methods allowing improved re- producibility of the Ames-test system, by the use of actively growing exponential-phase cultures of known cell densities, were developed and imple- mented.
The mutagen levels in wine were detected using strains TA98, TA100 and TA102 in a modified Ames-test system. The strains, generously pro- vided by B.N. Ames, were grown and stored on ampicillin plates (Maron and Ames, 1983), or at - 8 0 ° C as liquid cultures to which 10% (v/v) DMSO has been added.
Ames and others (Maron and Ames, 1983; Goggelmann, 1981) have established that the via- bility of nutrient broth-grown test cultures de- creases when incubated for prolonged periods at 37 ° C. However, "overnight cultures" (grown ap- prox. 15 h) are routinely used in the Ames-test system. Since the postexponential phase of a bacterial culture is not a physiological steady-state, the response of such cultures to mutagens is not always reproducible (Grafe et al., 1981). Altering the growth regimen in the following manner pro- duced cultures that were in steady-state balanced exponential growth, and which responded repro- ducibly as a mutagenicity test system.
Cells inoculated into 10 ml of Oxoid nutrient broth No. 2 were grown overnight at 37 °C with shaking at 350 rpm. These cultures contained 5.8 X 10 9 cells/ml as measured by plate counts of diluted cultures. 1.9 ml of the overnight culture was added to 8.1 ml of fresh, prewarmed broth (to yield a Klett reading of 100; green filter, 540 nm), and returned to shaking incubation at 37 ° C. Cell

growth was followed for approx. 1 h until the culture had attained 180 Klett units (1.5 × 109 cells/ml). This growth procedure produced a physiologically homogeneous bacterial population well suited to the reproducible detection of wine mutagens.
Mutagenicity assays were performed as follows. 0.1 ml of the tester strain grown to 180 Klett units was distributed into preincubation tubes contain- ing 0.5 ml sodium phosphate buffer (0.2 M, pH 7.4), and variable amounts of quercetin standard (0, 2.5, 5.0 or 7.5 /~g/plate) or concentrated wine extract (15 or 30 #l/plate) . The mixtures were incubated at 37 °C in a stationary waterbath for 20 min. 2 ml of histidine limited soft agar (0.6% agar, 0.6% NaC1, 0.05 mM L-histidine, and 0.05 mM biotin) was then added to each tube, mixed, and immediately poured onto VB-minimal glucose plates (Maron and Ames, 1983). When measuring the effect of rat-liver microsomal enzymes on the activation of wine mutagens, 0.5 ml of buffer was replaced with 0.5 ml of standard $9 mixture (0.1 ml rat-liver $9 (Aroclor-induced), 8/~moles MgC12, 33 /xmoles KCI, 5 /~moles glucose 6-phosphate, and 4 /~moles NADP + in 100 /xmoles pH 7.4 sodium phosphate buffer). All assays were per- formed in duplicate in each experiment. After the plates were incubated for 48 h at 37 o C, the num- ber of his + revertants/plate was counted.
In each experiment a standard curve relating the number of revertants per plate to concentra- tion of quercetin added to that plate (duplicate samples of 0, 2.5, 5.0 and 7.5/~g quercetin/plate, with or without $9) was prepared. The average number of revertants on the control plates con- taining no quercetin was subtracted from the numbers of revertants on the quercetin-containing plates. To determine the activity of the con- centrated wine samples in quercetin equivalents (mg/1), the numbers of revertants/plate produced by duplicate wine samples were determined. The number of spontaneous revertants on the control plates was subtracted from each determination, and the resulting number of revertants was com- pared to the standard curve. Use of a quercetin standard curve allowed reproducible and accurate calculation of a normalized level of mutagenicity (quercetin mutagenic equivalents in mg/ l ) in wine samples. Quercetin standards were prepared by
207
dissolving 5.6 mg quercetin dihydrate (Sigma or Kodak) in 10 ml of DMSO.
In other techniques associated with the Ames- test system, (1) media preparation, (2) TA98 stor- age, (3) phenotype verification, and (4) $9 pre- paration and storage, the suggestions of Maron and Ames (1983) were followed.
Results and discussion
Quantitation of wine mutagen isolation and con- centration methods. UV spectrophotometric anal- ysis of the eluate from 1.75 ml XAD-2 columns established that > 95% of a quercetin standard (16.7 mg/1) was recovered from a 12% ethanol solution (pH 3.5), and from a 1979 Santa Barbara Chardonnay which contained no intrinsic muta- genic activity. Mutagenic activity equivalent to 16.4 mg quercetin/1 was recovered from both sample preparations.
Ames-test system spontaneous reversion rates. The spontaneous reversion levels of TA98 in as- says containing no microsomal enzymes, and in assays containing S9-1iver homogenate, were quantitated in 23 different experiments over a period of one year. In the absence of activating enzymes, the background reversion rate for the modified Ames-tester system was 41 (standard deviation + 8) revertants per plate. The number rose when tester plates contained $9 liver extract, to 56 (standard deviation +10) revertants per plate.
Standardizing the Ames-test system response to quercetin. Known quantities of quercetin (2.5, 5.0 and 7.5/~g) were tested for mutagenic activity in the Ames-test system, without addition of S9- liver homogenate, in 23 separate experiments over a period of 1 year. After the number of sponta- neous revertants on control plates were subtracted from each quercetin test value, the net number of revertants per plate, lacking $9 and containing varying concentration of quercetin, were: 2.5 /tg quercetin, 64 + 14; 5.0 /xg, 144 + 24; and 7.5 /~g, 213 + 29. The effect of $9 on the mutagenic activ- ity of known quantities of quercetin was measured in 13 separate experiments over a 10-month period.
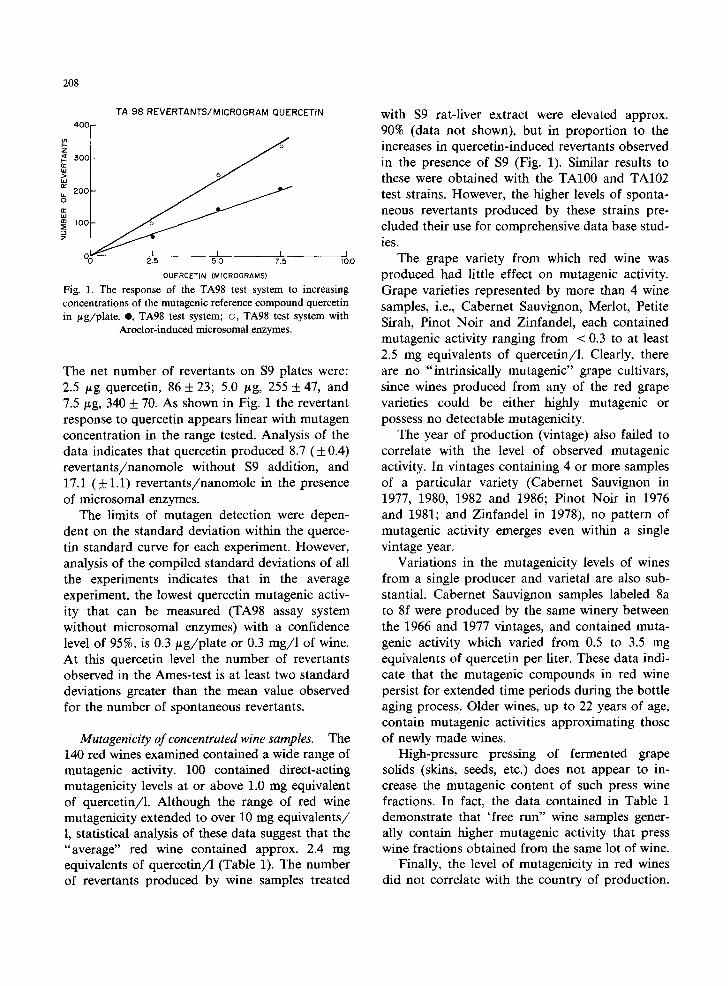
208
4 0 0
i 500 ~: 200
m I00
z
T A 9 8 R E V E R T A N T S / M I C R O G R A M Q U E R C E T I N
_ _ 1 I I I 0~" 2.5 5.0 7.5 I0,0
QUERCETIN (MICROGRAMS)
Fig. l. The response of the TA98 test system to increasing concentrations of the mutagenic reference compound quercetin in /~g/plate. @, TA98 test system; o, TA98 test system with
Aroclor-induced microsomal enzymes.
The net number of revertants on $9 plates were: 2.5 /~g quercetin, 86 + 23; 5.0 /~g, 255 + 47, and 7.5/Lg, 340 + 70. As shown in Fig. 1 the revertant response to quercetin appears linear with mutagen concentration in the range tested. Analysis of the data indicates that quercetin produced 8.7 ( + 0.4) revertants/nanomole without $9 addition, and 17.1 ( _ 1.1) revertants/nanomole in the presence of microsomal enzymes.
The limits of mutagen detection were depen- dent on the standard deviation within the querce- tin standard curve for each experiment. However, analysis of the compiled standard deviations of all the experiments indicates that in the average experiment, the lowest quercetin mutagenic activ- ity that can be measured (TA98 assay system without microsomal enzymes) with a confidence level of 95%, is 0.3/~g/plate or 0.3 mg/1 of wine. At this quercetin level the number of revertants observed in the Ames-test is at least two standard deviations greater than the mean value observed for the number of spontaneous revertants.
Mutagenicity of concentrated wine samples. The 140 red wines examined contained a wide range of mutagenic activity. 100 contained direct-acting mutagenicity levels at or above 1.0 mg equivalent of quercetin/1. Although the range of red wine mutagenicity extended to over 10 mg equivalents/ I, statistical analysis of these data suggest that the "average" red wine contained approx. 2.4 mg equivalents of quercetin/1 (Table 1). The number of revertants produced by wine samples treated
with $9 rat-liver extract were elevated approx. 90% (data not shown), but in proportion to the increases in quercetin-induced revertants observed in the presence of $9 (Fig. 1). Similar results to these were obtained with the TA100 and TA102 test strains. However, the higher levels of sponta- neous revertants produced by these strains pre- cluded their use for comprehensive data base stud- ies.
The grape variety from which red wine was produced had little effect on mutagenic activity. Grape varieties represented by more than 4 wine samples, i.e., Cabernet Sauvignon, Merlot, Petite Sirah, Pinot Noir and Zinfandel, each contained mutagenic activity ranging from < 0.3 to at least 2.5 mg equivalents of quercetin/1. Clearly, there are no "intrinsically mutagenic" grape cultivars, since wines produced from any of the red grape varieties could be either highly mutagenic or possess no detectable mutagenicity.
The year of production (vintage) also failed to correlate with the level of observed mutagenic activity. In vintages containing 4 or more samples of a particular variety (Cabernet Sauvignon in 1977, 1980, 1982 and 1986; Pinot Noir in 1976 and 1981; and Zinfandel in 1978), no pattern of mutagenic activity emerges even within a single vintage year.
Variations in the mutagenicity levels of wines from a single producer and varietal are also sub- stantial. Cabernet Sauvignon samples labeled 8a to 8f were produced by the same winery between the 1966 and 1977 vintages, and contained muta- genic activity which varied from 0.5 to 3.5 mg equivalents of quercetin per liter. These data indi- cate that the mutagenic compounds in red wine persist for extended time periods during the bottle aging process. Older wines, up to 22 years of age, contain mutagenic activities approximating those of newly made wines.
High-pressure pressing of fermented grape solids (skins, seeds, etc.) does not appear to in- crease the mutagenic content of such press wine fractions. In fact, the data contained in Table 1 demonstrate that 'free run" wine samples gener- ally contain higher mutagenic activity that press wine fractions obtained from the same lot of wine.
Finally, the level of mutagenicity in red wines did not correlate with the country of production.
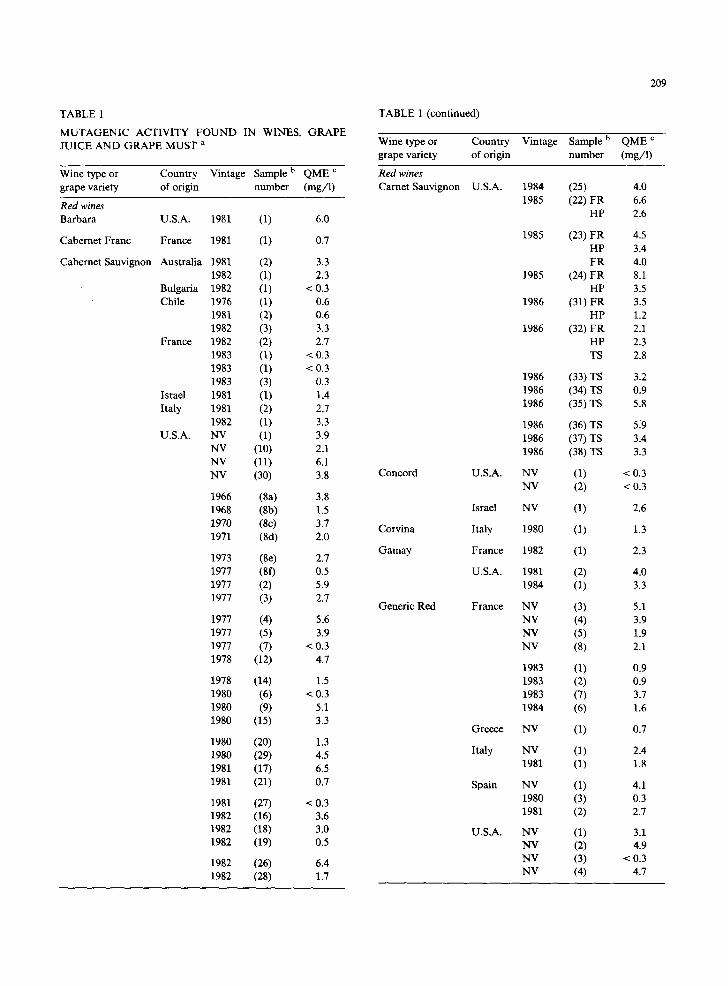
TABLE 1
M U T A G E N I C ACTIVITY F O U N D IN WINES, G R A P E JUICE AND GRAPE MUST a
Wine type or Country Vintage Sample b QME c grape variety of origin number (mg/1)
Red wines Barbara U.S.A.
Cabernet Franc France
Cabemet Sauvignon Australia
Bulgaria Chile
France
Israel Italy
U.S.A.
1981 (1) 6.0
1981 (1) 0.7
1981 (2) 3.3 1982 (1) 2.3 1982 (1) < 0.3 1976 (1) 0,6
1 9 8 1 (2) 0,6 1982 (3) 3.3 1982 (2) 2,7 1983 (1) < 0.3 1983 (1) < 0.3 1983 (3) 0.3 1981 (1) 1.4 1981 (2) 2.7 1982 (1) 3.3 NV (1) 3.9 NV (10) 2.1 NV (11) 6.1 NV (30) 3.8
1966 (8a) 3.8 1968 (8b) 1.5 1970 (8c) 3.7 1971 (8d) 2.0
1973 (8e) 2.7 1977 (8f) 0.5 1977 (2) 5.9 1977 (3) 2.7
1977 (4) 5.6 1977 (5) 3.9 1977 (7) < 0.3 1978 (12) 4.7
1978 (14) 1.5 1980 (6) < 0.3 1980 (9) 5.1 1980 (15) 3.3
1980 (20) 1.3 1980 (29) 4.5 1981 (17) 6.5 1981 (21) 0.7
1981 (27) < 0.3 1982 (16) 3.6 1982 (18) 3.0 1982 (19) 0.5
1982 (26) 6.4 1982 (28) 1.7
TABLE 1 (continued)
Wine type or Country Vintage Sample b QME c grape variety of origin number (mg/1)
Red wines Carnet Sauvignon U.S.A.
Concord U.S.A.
Israel
Corvina Italy
Gamay France
U.S.A.
Generic Red France
Greece
Italy
Spain
U.S.A.
1984 (25) 4.0 1985 (22) FR 6.6
HP 2.6
1985 (23) FR 4.5 HP 3.4 FR 4.0
1985 (24) FR 8.1 HP 3.5
1986 (31) FR 3.5 HP 1.2
1986 (32) FR 2.1 HP 2.3 TS 2.8
1986 (33) TS 3.2 1986 (34) TS 0.9 1986 (35) TS 5.8
1986 (36) TS 5~9 1986 (37) TS 3.4 1986 (38) TS 33
NV (1) < 0.3 NV (2) < 0,3
NV (I) 2.6
1980 (1) 1.3
1982 (1) 2.3
1981 (2) 4.0 1984 (1) 3.3
NV (3) 5.1 NV (4) 3.9 NV (5) 1.9 NV (8) 2.1
1983 (1) 0.9 1983 (2) 0.9 1983 (7) 3.7 1984 (6) 1.6
NV (1) 0.7
NV (1) 2.4 1981 (1) 1.8
NV (1) 4.1 1980 (3) 0.3 1981 (2) 2.7
NV (1) 3.1 NV (2) 4.9 NV (3) < 0.3 NV (4) 4.7
209
210
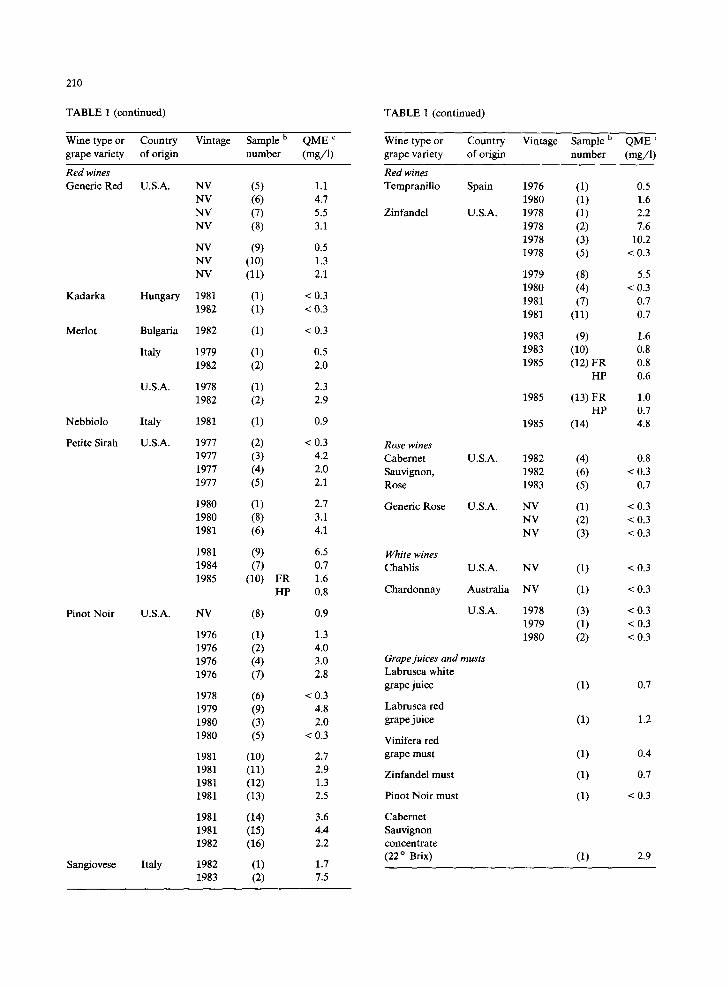
TABLE 1 (continued)
Wine type or Country Vintage Sample b QME ¢ grape variety of origin number (rag/l)
Red wines Generic Red U.S.A.
Kadarka Hungary
Merlot Bulgaria
Italy
U.S.A.
Nebbiolo Italy
Petite Sirah U.S.A.
NV (5) NV (6) NV (7) NV (8)
NV (9) NV (10) NV (11)
1981 (1) 1982 (1)
1982 (1)
1979 (1) 1982 (2)
1978 (1) 1982 (2)
1981 (1)
1977 (2) 1977 (3) 1977 (4) 1977 (5)
1980 (1) 1980 (8) 1981 (6)
1981 (9) 1984 (7) 1985 (10)
Pinot Noir U.S.A. NV (8)
1976 (1) 1976 (2) 1976 (4) 1976 (7)
1978 (6) 1979 (9) 1980 (3) 1980 (5)
1981 (10) 1981 (11) 1981 (12) 1981 (13)
1981 (14) 1981 (15) 1982 (16)
Sangiovese Italy 1982 (1) 1983 (2)
FR HP
1.1 4.7 5.5 3.1
0.5 1.3 2.1
< 0.3 < 0.3
< 0.3
0.5 2.0
2.3 2.9
0.9
< 0.3 4.2 2.0 2.1
2.7 3.1 4.1
6.5 0.7 1.6 0.8
0.9
1.3 4.0 3.0 2.8
< 0.3 4.8 2.0
< 0.3
2.7 2.9 1.3 2.5
3.6 4.4 2.2
1.7 7.5
TABLE 1 (continued)
Wine type or Country Vintage Sample b QME c grape variety of origin number (mg/1)
Red wines Tempranillo Spain
Zinfandel U.S.A.
1976 (1) 0.5 1980 (1) 1.6 1978 (1) 2.2 1978 (2) 7.6 1978 (3) 10.2 1978 (5) < 0.3
1979 (8) 5.5 1980 (4) < 0.3 1981 (7) 0.7 1981 (11) 0.7
1983 (9) 1.6 1983 (10) 0.8 1985 (12) FR 0.8
HP 0.6
1985 (13) FR 1.0 HP 0.7
1985 (14) 4.8
White wines Chablis U.S.A. NV (1) < 0.3
Chardonnay Australia NV (1) < 0.3
U.S.A. 1978 (3) < 0.3 1979 (1) < 0.3 1980 (2) < 0.3
Grape juices and musts Labrusca white grape juice (1) 0.7
Labrusca red grape juice (1) 1.2
Vinifera red grape must (1) 0.4
Zinfandel must (1) 0.7
Pinot Noir must (1) < 0.3
Cabemet Sauvignon concentrate (22 ° Brix) (1) 2.9
Rose wines Cabernet U.S.A. 1982 (4) 0.8 Sauvignon, 1982 (6) < 0.3 Rose 1983 (5) 0.7
Generic Rose U.S.A. NV (1) < 0.3 NV (2) < 0.3 NV (3) < 0.3

Table 1 (continued)
a Abbreviations and codes: NV, non-vintage; TS, winery tank sample; FR, wine prepared from free run juice; HP, wine prepared from hard press juice; FW, final wine.
b Individual wine samples were identified by random sample numbers during mutagen measurements.
c QME, quercetin mutagenic equivalent (mg/l), was de- termined by comparing the number of TA98 revertants pro- duced in response to 15 or 30 /xl of concentrated wine sample to the number of TA98 revertants produced in re- sponse to 0, 2.5, 5.0 and 7.5 /~g of quercetin (the standard curve prepared for each experiment). The derived inter- polated values are reported as QME.
Each country represented by more than 3 wines produced wines varying widely in mutagenicity.
The white wines and rose wines, as well as the unfermented grape juices, contained low or unde- tectable levels of mutagenicity. Since at least 3 of the Chardonnay wines which were tested were aged in oak barrels, but exhibited no mutagenic- ity, it is unlikely that aging red wine in oak barrels affects their mutagenicity levels.
Individual winery production methods must be primarily responsible for the range of mutagenic activities observed in red wine. Surprisingly, the grape must from which red wine is produced contains relatively low levels of mutagenicity, even when the resulting wine exhibits significant muta- genicity. Hence it would appear that mutagenic compounds are produced during the red wine fermentation and aging processes.
These studies establish that a wide range of red wine products contain substantial levels of direct- acting mutagenic activity. Further studies are in progress to identify the compound(s) responsible for this mutagenic effect and to understand the production practices which affect the formation and/or removal of these mutagens. Parallel inves- tigations are being carried out to extend these studies to include other wine types to which the human population is exposed.
Acknowledgements
This research was supported by grants from the National Institutes of Health (CA36890) and the University of California Cancer Research Coordi- nating Committee. We are grateful to Dr. Bruce Ames for helpful advice and discussions.
211
References
Amerine, M.A., W.H. Berg, R.E. Kunkee, C.S. Ough and V.L. Singleton and A.D. Webb (1980) Technology of Wine Making, 4th edn., AVI Publishing Company, Inc., West- port, CN.
Ames Bruce N. (1979) Identifying environmental chemicals causing mutations and cancer, Science, 204, 587-593.
Ames, Bruce N. (1986) Food constituents as a source of mutagens, carcinogens, and anticarcinogens, in: I. Knudsen (Ed.), Genetic Toxicology of the Diet, Liss, New York, pp. 3-32.
Ames, B.N., J. McCann and E. Yamasaki (1975) Methods for detecting carcinogens and mutagens with the Salmonella/ mammalian-microsome mutagenicity test, Mutation Res., 31, 347-363.
Brown, J.P. (1980) Review of the genetic effects of naturally occurring flavonoids, anthraquinones and related com- pounds, Mutation Res., 75, 243-277.
Brown, J.P., and P.S. Dietrich (1970) Mutagenicity of plant flavonols in the Salmonella/mammalian microsome test: activation of flavonol glycosidases from rat cecal bacteria and other sources, Mutation Res., 66, 233-240.
Doll, R., and R. Peto (1981) The Causes of Cancer: Qualitative estimates of avoidable risks of cancer in the United States today, J. Natl. Cancer Inst., 66, 1192-1265.
Goggelmann, W. (1981) Culture conditions and influence of the number of bacteria on the number of revertants, in: A. Kappas (Ed.), Progress in Mutation Research, Vol. 2, Else- vier, Amsterdam, pp. 173-178.
Grafe, A., I.E. Mattern and M. Green (1981) A European collaborative study of the Ames assay, I. Results and general interpretation, Mutation Res., 85, 391-410.
Hiatt, H.H., J.D. Watson and J.A. Winsten (Eds.) (1977) Origins of Human Cancer, Cold Spring Harbor Labora- tory, Cold Spring Harbor, New York.
Higginson, J., and C.S. Muir (1979) Environmental carcinogen- esis: misconceptions and limitations to cancer control, J. Natl. Cancer Inst., 63, 1291-1298.
Hirono, I. (1981) Natural carcinogenic products of plant origin, CRC Crit. Rev. Toxicol., 8, 235-277.
Hirono, I. (Ed.) (1987) Naturally occurring carcinogens of plant origin, in: Bioactive Molecules, VoL 2, Kodansha, Tokyo and Elsevier, Amsterdam.
Hoey, J., C. Montveruay and R. Lambert (1981) Wine and tobacco: risk factors for gastric cancer in France, Am. J. Epidemiol., 113, 668-674.
Hollstein, M., J. McCann, F.A. Angelosanto and W.W. Nichols (1979) Short-term tests for carcinogens and mutagens, Mu- tation Res., 65, 133-226.
Knudson, A.G. (1977) Genetic predisposition to cancer, in: H.H. Hiatt, J.D. Watson and J.A. Winsten (Eds.), Origins of Human Cancer, Vol. 4, Cold Spring Harbor Laboratory, Cold Spring Harbor, New York, pp. 45-52.
Maron, D.M., and B.N. Ames (1983) Revised methods for the Salmonella mutagenicity test, Mutation Res., 113, 173-215.
Miller, E.C., J.A. Miller, I. Hirono, T. Suglmura and S. Takayama (Eds.) (1979) Naturally Occurring Carcino-

212
gens-Mutagens and Modulators of Carcinogenesis, Japan Scientific Societies Press, Tokyo and University Park Press, Baltimore.
Nagao, M., N. Morita, T. Yahagi, M. Shimizu, M. Fukuoka, K. Yoshihira, S. Natori, T. Fujino and T. Sugimura (1981) Mutagenicities of 61 flavonoids and 11 related compounds, Environ. Mutagen., 3, 401-419.
Peto, R., and M. Schneiderman (Eds.) (1981) Banbury Report 9. Quantification of Occupational Cancer, Cold Spring Harbor Laboratory, Cold Spring Harbor, New York.
Stavric, B., D.R. Stolz and R. Klassen (1983) Toxicants in foods with emphasis on mutagens in beverages, fruits and vegetables, Toxicon, Suppl. 3, 409-412.
Tamura, G., C. Gold, A. Ferro-Luzzi and B.N. Ames (1980) Fecalase: A model for activation of dietary glycosides to mutagens by intestinal flora, Proc. Natl. Acad. Sci. (U.S.A.), 77, 4961-4965.
Wynder, E.L., and G.B. Gori (1977) Contribution of the en- vironment to cancer incidence: an epidemiologic exercise, J. Natl. Cancer Inst., 58, 825-832.
Yamasaki, E., and B.N. Ames (1977) Concentration of muta- gens from urine by adsorption with the non-polar resin XAD-2: Cigarette smokers have mutagenic urine, Proc. Natl. Acad. Sci. (U.S.A.), 74, 3555-3559.










![INDEX []...INDEX SOMMELIER’S SELE TION & ROSE WINES 1 WINES BY THE GLASS 2 SPARKLING WINES 3 WHITE WINE USA 5 FRANCE, Burgundy, Bordeaux, Loire & Rhône 6 FRANCE, Languedoc, Alsace,](https://static.fdocuments.us/doc/165x107/5e8e262b4f3fa8295b239ef9/index-index-sommelieras-sele-tion-rose-wines-1-wines-by-the-glass.jpg)







