
The Climacteric in Ripening Tomato - Plant Physiology · ripening tomato fruit ... tion rate at the...
6
Plant Physiol. (1971) 48, 235-240 The Climacteric in Ripening Tomato Fruit1 Received for publication February 2, 1971 DAVID J. CHALMERS2 AND KINGSLEY S. ROWAN Botany School, University of Melbourne, Parkville, Victoria, 3052, Australia ABSTRACT Phosphofructokinase is identified as the regulator reaction activated at the onset of the climacteric rise in respiration of the ripening tomato fruit (Lycopersicon esculentum Mill). The concentration of ATP in the fruit increases to a maximum value after the elimacteric peak of respiration is past. Orthophosphate is proposed as the most probable activator of phosphofructo- kinase in the ripening fruit. Fifteen hours after infiltrating tomato fruit with orthophos- phate, the rate of respiration increased and remained high un- til the end of the experiment, 45 hours after infiltration. In experiments where tomato plants were grown at various nutri- ent levels of P, the rate of respiration when fruit harvested at the mature-green stage reached the respiratory climacteric was correlated with the concentration of orthophosphate in the fruit at the end of the experiment. These results are consistent with the hypothesis that stimulation of phosphofructokinase through increasing concentration of orthophosphate in the cytoplasm of the fruit contributes to the elimacteric rise in respiration. Hess (18) has implicated the irreversible reactions of the EMP' pathway as sites of regulation of carbon flux in living cells, and many other workers have reported evidence for regu- lation by one or more of these reactions in a wide variety of tissues. In higher plants, Givan (16) has identified PFK as regulator in cultured cells of Acer pseudoplatanus subjected to anoxia, and Adams and Rowan (3) have reported sequential regulation of induced respiration in sliced carrot tissue of PK and PFK. During the climacteric rise in respiration of ripening fruit, G6P decreases in avocado (35), and FDP increases in banana (7). Both observations are consistent with the stimula- tion of PFK. The crossover theorem of Chance (14, 15) is used herein to identify PFK as the site of regulation during the climacteric rise in respiration of tomato fruits. Stimulation of PFK in a ripening tomato fruit could be due I D. J. C. was supported by the Victorian State Department of Agriculture. 2Present address: Scoresby Horticultural Research Station, P. 0. Box 174, Ferntree Gully, Victoria, 3156, Australia. 'Abbreviations: EMP: Embden-Meyerhof-Parnas; IG: immature green; MG: mature green; PB: prebreaker; B: breaker; HR: half- ripe; TR: table-ripe; CR: canning ripe; F6P: fructose-6-P; G6P: glucose-6-P; FDP: fructose- 1,6-diP; TP: glyceraldehyde-3-P + di- hydroxyacetone-P; PEP: phosphoenolpyruvate; PFK: phosphofruc- tokinase; PK: pyruvate kinase; PYR: pyruvate; IRGA: infrared gas analyzer; DNP: 2,4-dinitrophenol. to a decrease in the concentration of a negative effector, or the increase in concentration of a positive effector, an antago- nist of a negative effector, or of either substrate. In a study of this enzyme in pea seed, Kelly and Turner (20, 21) have iden- tified 10 effectors. In the first part of the work reported here no change in concentration of any metabolite appeared to fall in any of the above categories, and, since several workers have proposed that changes in permeability accompany the climac- teric (6, 29, 30, 33), we postulate that, at the climacteric, the diffusive resistance of the tonoplast decreases, allowing P,, a known positive effector of PFK, to diffuse down a concentra- tion gradient from vacuole to cytoplasm, thus stimulating PFK. Phosphatase activity, which increases during the climacteric in ripening apples (25), could be responsible for stimulation of PFK by increasing the level of P, in the cytoplasm, but our un- published work has failed to demonstrate a positive association between phosphatase activity and the climacteric in tomato fruit. Orthophosphate accounts for 70% of total P in the potato tuber (11, 22) and Spirodela oligorrhiza (9) of which approxi- mately 90% was in a nonmetabolic pool not freely available to the cytoplasm. In Spirodela, this pool is depleted by im- posing P deficiency, presumably by diffusion (10) from vacuole to cytoplasm. If diffusion of P, from a similar non-metabolic pool in tomato fruit cells is limited by impermeable mem- branes, any process increasing the permeability of the mem- branes could increase the concentration of P, in the cytoplasm, thus inducing the climacteric rise in respiration. Although in some experiments with banana slices the respiratory increase preceded the increase in efflux (33), the changes in permea- bility of tonoplast and plasmalemma need not coincide, and the concentration of cytoplasmic P, could rise before the in- creased efflux from the slices could be detected. Alternately, failure of a metabolic "pump" maintaining the concentration gradient, rather than an increase in permeability, would have the same effect. Invoking permeability changes of this type is restating the concept of decreasing "organization resistance" originally proposed by Blackman and Parija (12) as an ex- planation of the climacteric in ripening apples. In Spirodela (10), P deficiency greatly reduced the level of total P, and the proportion of Pi in the nonmetabolic pool without greatly affecting the level of organic phosphates. Therefore, tomatoes were grown under varying phosphate re- gimes to modify the amount of P1 in the nonmetabolic pool (vacuole) potentially available to stimulate respiration. Ac- cording to the hypothesis to be tested, the maximum respira- tion rate at the peak of the climacteric should be a function of the total concentration of Pi in the fruit, providing the P, concentration does not rise above the optimum for activation of PFK. This is quite distinct from correlating the increasing respiration rate before the climacteric maximum with other biochemical parameters, the approach used in previous studies by other workers. 235 www.plantphysiol.org on June 21, 2018 - Published by Downloaded from Copyright © 1971 American Society of Plant Biologists. All rights reserved.
-
Upload
vuongtuong -
Category
Documents
-
view
218 -
download
1
Transcript of The Climacteric in Ripening Tomato - Plant Physiology · ripening tomato fruit ... tion rate at the...

Plant Physiol. (1971) 48, 235-240
The Climacteric in Ripening Tomato Fruit1Received for publication February 2, 1971
DAVID J. CHALMERS2 AND KINGSLEY S. ROWANBotany School, University of Melbourne, Parkville, Victoria, 3052, Australia
ABSTRACT
Phosphofructokinase is identified as the regulator reactionactivated at the onset of the climacteric rise in respiration of theripening tomato fruit (Lycopersicon esculentum Mill). Theconcentration of ATP in the fruit increases to a maximum valueafter the elimacteric peak of respiration is past. Orthophosphateis proposed as the most probable activator of phosphofructo-kinase in the ripening fruit.
Fifteen hours after infiltrating tomato fruit with orthophos-phate, the rate of respiration increased and remained high un-til the end of the experiment, 45 hours after infiltration. Inexperiments where tomato plants were grown at various nutri-ent levels of P, the rate of respiration when fruit harvested atthe mature-green stage reached the respiratory climacteric wascorrelated with the concentration of orthophosphate in thefruit at the end of the experiment. These results are consistentwith the hypothesis that stimulation of phosphofructokinasethrough increasing concentration of orthophosphate in thecytoplasm of the fruit contributes to the elimacteric rise inrespiration.
Hess (18) has implicated the irreversible reactions of theEMP' pathway as sites of regulation of carbon flux in livingcells, and many other workers have reported evidence for regu-lation by one or more of these reactions in a wide variety oftissues. In higher plants, Givan (16) has identified PFK asregulator in cultured cells of Acer pseudoplatanus subjectedto anoxia, and Adams and Rowan (3) have reported sequentialregulation of induced respiration in sliced carrot tissue of PKand PFK. During the climacteric rise in respiration of ripeningfruit, G6P decreases in avocado (35), and FDP increases inbanana (7). Both observations are consistent with the stimula-tion of PFK. The crossover theorem of Chance (14, 15) isused herein to identify PFK as the site of regulation duringthe climacteric rise in respiration of tomato fruits.
Stimulation of PFK in a ripening tomato fruit could be due
I D. J. C. was supported by the Victorian State Department ofAgriculture.
2Present address: Scoresby Horticultural Research Station, P. 0.Box 174, Ferntree Gully, Victoria, 3156, Australia.
'Abbreviations: EMP: Embden-Meyerhof-Parnas; IG: immaturegreen; MG: mature green; PB: prebreaker; B: breaker; HR: half-ripe; TR: table-ripe; CR: canning ripe; F6P: fructose-6-P; G6P:glucose-6-P; FDP: fructose- 1,6-diP; TP: glyceraldehyde-3-P + di-hydroxyacetone-P; PEP: phosphoenolpyruvate; PFK: phosphofruc-tokinase; PK: pyruvate kinase; PYR: pyruvate; IRGA: infrared gasanalyzer; DNP: 2,4-dinitrophenol.
to a decrease in the concentration of a negative effector, orthe increase in concentration of a positive effector, an antago-nist of a negative effector, or of either substrate. In a study ofthis enzyme in pea seed, Kelly and Turner (20, 21) have iden-tified 10 effectors. In the first part of the work reported hereno change in concentration of any metabolite appeared to fallin any of the above categories, and, since several workers haveproposed that changes in permeability accompany the climac-teric (6, 29, 30, 33), we postulate that, at the climacteric, thediffusive resistance of the tonoplast decreases, allowing P,, aknown positive effector of PFK, to diffuse down a concentra-tion gradient from vacuole to cytoplasm, thus stimulating PFK.Phosphatase activity, which increases during the climacteric inripening apples (25), could be responsible for stimulation ofPFK by increasing the level of P, in the cytoplasm, but our un-published work has failed to demonstrate a positive associationbetween phosphatase activity and the climacteric in tomatofruit.
Orthophosphate accounts for 70% of total P in the potatotuber (11, 22) and Spirodela oligorrhiza (9) of which approxi-mately 90% was in a nonmetabolic pool not freely availableto the cytoplasm. In Spirodela, this pool is depleted by im-posing P deficiency, presumably by diffusion (10) from vacuoleto cytoplasm. If diffusion of P, from a similar non-metabolicpool in tomato fruit cells is limited by impermeable mem-branes, any process increasing the permeability of the mem-branes could increase the concentration of P, in the cytoplasm,thus inducing the climacteric rise in respiration. Although insome experiments with banana slices the respiratory increasepreceded the increase in efflux (33), the changes in permea-bility of tonoplast and plasmalemma need not coincide, andthe concentration of cytoplasmic P, could rise before the in-creased efflux from the slices could be detected. Alternately,failure of a metabolic "pump" maintaining the concentrationgradient, rather than an increase in permeability, would havethe same effect. Invoking permeability changes of this type isrestating the concept of decreasing "organization resistance"originally proposed by Blackman and Parija (12) as an ex-planation of the climacteric in ripening apples.
In Spirodela (10), P deficiency greatly reduced the level oftotal P, and the proportion of Pi in the nonmetabolic poolwithout greatly affecting the level of organic phosphates.Therefore, tomatoes were grown under varying phosphate re-gimes to modify the amount of P1 in the nonmetabolic pool(vacuole) potentially available to stimulate respiration. Ac-cording to the hypothesis to be tested, the maximum respira-tion rate at the peak of the climacteric should be a functionof the total concentration of Pi in the fruit, providing the P,concentration does not rise above the optimum for activationof PFK. This is quite distinct from correlating the increasingrespiration rate before the climacteric maximum with otherbiochemical parameters, the approach used in previous studiesby other workers.
235
www.plantphysiol.orgon June 21, 2018 - Published by Downloaded from Copyright © 1971 American Society of Plant Biologists. All rights reserved.

CHALMERS AND ROWAN
MATERIALS AND METHODS
Plant Material. Fruits for extraction and assay of inter-mediates (Lycopersicon esculentum Mill) were grown at theScoresby Horticultural Research Station, Victoria (cu. "V10")or under glass at the Botany School, Melbourne University(cu. "Grosse Lisse"). Harvested fruits were graded into ripe-ness classes essentially by the method described by Rowan etal. (28). Since in experiment 1 grading into only five classes(MG, B, HR, TR, and CR) was inadequate, in experiment 2,an IG class, three grades of PB (PB1, PB2 and PB3), threegrades of B (B1, B2 and B3) and two grades of HR (HR1 andHR.) were selected in addition to MG and TR (CR was notselected in this experiment). Fruits containing varying con-centrations of P, (cu. "Grosse Lisse") were obtained by grow-ing in sand culture using the following solutions (19): solutionA: KNO3 (0.404 g per liter), Ca(NO3)2 (0.656 g per liter),MgSO4 (0.368 g per liter); solution B: NaH,PO (0.208 g perliter). Eight plants were used; five received 0.5 liter per dayof both solutions A and B for 6 weeks, and then 1 liter perday until the initial fruit set. The remaining three plants re-
ceived 0.5 liter per day of both solutions A and B for 6 weeksand thereafter 1 liter of solution A and 0.5, 0.25, and 0.125liter respectively of solution B daily. After the initial fruit set,three of the five plants receiving full nutrients were reducedto the lower levels of solution B. When necessary, supplemen-tary water was applied, and at weekly intervals nutrients wereleached from the sand culture by exhaustive watering. Fruitswere harvested at the mature green stage. Phosphofructokinasewas extracted from immature fruit (1-10 g; cultivar "GrosseLisse") harvested from plants grown under glass at the BotanySchool.
Respiration Measurements. The rate of respiration was cal-culated by passing CO1-free air at a monitored rate throughdarkened respiration chambers and measuring the concentra-tion of CO1 in the effluent airstream by a colorimetric method(13) or by using an IRGA (Grubb Parsons model SB2 or3074). Appropriate calibration curves were constructed bypreparing standard gas mixtures with a gas-mixing pump (H.Wosthoff, type 27-3). The respiration rate of single fruit was
followed except in experiment1 (Fig. 2) where samples con-taining eight fruits were used. The respiration rates of fruitsfrom which glycolytic intermediates and ATP were extracted(Figs. 1 and 2) was followed continuously for 48 hr beforeextraction. In contrast, fruits used in studying the effect ofendogenous P, level on the climacteric maximum were har-vested at the mature green stage, and the respiration rate was
followed continuously while ripening in the respirometer.Infiltration Experiments. Fruits used were grown at Geral-
ton, West Australia (cu. "Geralton Smooth Skin") and boughtfrom local wholesalers. Experimental solutions (0.3 ml) wereinfiltrated into mature green fruits (100 g) by the method ofSpencer(31).
Extraction and Assay of Intermediates. Acid-soluble ex-
tracts were prepared by blending 100 g of tissue in 80 or 100ml HCIO4 (10%, v/v) for 2 min, filtering the blend through apad of dry "Hyflo Supercel" over Whatman No. 541 (41H)paper and adjusting the pH to 8 with 8 N KOH; the precipitateof KC104 was removed by sedimentation overnight (26). Alloperations were performed in a cold room at1 C. Extractsused for determination of glycolytic intermediates were de-colorized by shaking with activated charcoal for1 min beforeplacing in a freezing cabinet at -15 C. The water content was
determined in replicate samples of tissue.Glycolytic intermediates were determined enzymatically (8).
ATP was estimated using luciferinluciferase (32) as describedby Bergmeyer (8), measuring the light emitted at 562 nm with
an Aminco-Bowman spectrophotofluorometer coupled with anHitachi model QPD 73 potentiometric recorder. Orthophos-phate was measured by the colorimetric method of Allen (4).
Extraction and Assay of PFK Activity at Various Pi Con-centrations. One hundred grams of immature tomatoes wereblended with a Braun juice extractor from which the tissuehomogenate flowed rapidly into a beaker containing 50 ml ofextracting medium. The extracting medium (20 mm tris buffer,pH 7.5) contained sodium metabisulfite (10 mM) to prevent theformation of tannins (5) and EDTA (2 mM). The extract wasfiltered through "Hyflo Supercel" on Whatman No. 541 (41H)filter paper. The fraction precipitating between 33 and 45%(w/v) saturated (NH4)2S0, was collected by centrifugation at10,OOOg for 30 min. The precipitate was redissolved in a smallvolume of 20 mm tris buffer, pH 7.5 and was used in the PFKassay.The reaction was carried out at 25 C in the presence of
aldolase, triose phosphate isomerase, and glycerol-i-phosphatedehydrogenase in 50 mm tris, pH 7.5. Fructose-6-P was gen-erated from G6P by phosphoglucose isomerase or F6P freeof FDP was used. ATP and magnesium (final concen-tration 5 mM) were also added to the reaction cuvet. The con-centration of NADH, between 2.5 and 5 fkM in the reactioncuvet, was standardized spectrophotometrically. The rate ofNADH oxidation was measured with an Aminco-Bowmanspectrophotofluorometer coupled to an Hitachi (series QPD73) recorder. The excitation wavelength was 340 nm andemission was measured at 460 nm. Fluorescence was propor-tional to the concentration of NADH for the concentrationrange used. The effect of phosphate concentration on a crudepreparation of PFK was investigated at two F6P levels. Phos-phate (Na2HPO4) was added to the assay mixture in the cuvetbefore starting the reaction with PFK extract.
RESULTS
Changes in EMP Intermediates and ATP. Figure 1 showsthe changes in concentration of EMP intermediates in fruitsfrom IG to TR stages and shows that the respiratory climac-teric begins at stage PB1 and continues to the "breaker" stage(Bi-BB3). The concentrations of F6P and G6P decrease betweenstages MG to PB1, while FDP increases sharply; all otherintermediates measured increase during this interval. The con-centration of pyruvate increases. Both sets of changes occurduring the climacteric increase in rate of respiration. Ortho-phosphate increases from the start of the experiment until thePB3 stage. Similar changes occurred in experiment1 but thestage at which they occurred could not be localized so ac-curately.
Figure 2 shows the concentration of ATP in fruits fromMG to CR in experiment1 using combined samples; the in-crease in ATP concentration occurs one stage later than theonset of the climacteric rise, and the climacteric peak at theHR stage also precedes the maximum concentration of ATP.ATP concentration and rate of respiration both increase byapproximately 30%.The Effect of P1 Concentration on the Activity of PFK. The
optimalPi concentration lies between1 and 5 mm at both 0.8and 3 mm F6P, butPi above 30 mm inhibits with 3 mm F6P.Near the optimal P, concentration, the effect of F6P concen-tration is greatly reduced (Fig. 3).
The Effect of Infiltration of P1 on the Rate of Respiration ofPreclimacteric Tomato Fruits. Respiration decreased rapidlyafter infiltration of water, followed by a small rise whichslowly returned to the initial rate after about 40 hr (Fig. 4a).Infiltration with1 mm DNP caused a similar decrease and re-turn to the initial rate as with water, but the rate of respiration
236 Plant Physiol. Vol. 48, 1971
www.plantphysiol.orgon June 21, 2018 - Published by Downloaded from Copyright © 1971 American Society of Plant Biologists. All rights reserved.

Plant Physiol. Vol. 48, 1971 THE CLIMACTERIC IN TOMATO FRUIT
then increased further to a maximum after about 20 hr, fol-lowed by a decline for the remainder of the experiment (Fig.4b). Return to the initial respiration rate was less rapid (4 hr)after infiltration with 0.7 M KCl, but the subsequent doublingof the respiration rate was extremely rapid, being completewithin a further 3-hr period (Fig. 4c). Immediately after thepeak was reached, the respiration rate declined to the initialrate after approximately 40 hr. After infiltration of 0.7 Mphosphate buffer, pH 7.5, the rate of respiration remainedbelow the initial level for 8 hr (Fig. 4d), then increased toapproximately 50% above the initial rate. After reaching a
C _-X 40C: L.L.0 :))
o O-Z ",'30
L-0X 0 20tr E
1 0
8
6a.
u 4
2
* * .M
ot~~~~t
0~~~~~
? T i I I I
A DP
_ 7 8 D=6P
36 _
32 _
28 _-
CL24(D
020
16
12
8
so 6
(L4
2
300
200
100
Pyruvate~~~~~ ~~~PE P
I I
Pi
I II I
I.G. MG. PEl PEB2 PB.3 RE B2 B3 H.R1 HR2 T R.Stage of Maturity
I 60a)
-55LL0
- , 50
a)Q)cr 0 4500)
3 6-5
O -o 6.00
5-50
0k 5.Q
1 0
0-8
0-6 iC-L 1o tom
0 4
0-2
1 6
1-4 x
121 2
08 ULa-06
0-4
4
_-
_-
_-
ATP
M.G. B. H.R. T.R. C.R.Stage of Maturity
FIG. 2. Respiratory rate at 20 C and concentration of ATP iniato fruits harvested at five stages of maturity (experiment 1).
0-5 1 5 10 50
Pi Concentration (mM)
3 FIG. 3. The effect of Pi concentration on PFK activity at 0.8CL mm F6P (M) and 3 mMF6P (M).
2peak 15 hr after infiltration, the rate declined but leveled off20% higher than the initial rate for the remainder of the ex-
periment. A similar result was obtained when this experimentwas repeated.The Relation between P, Concentration and the Respiration
Rate at the Climacteric Maimum. The P, concentration andthe respiratory rate at the climacteric were highly correlated(F = 19.34, p = 0.1%). Regression line A (Fig. 5) is calcu-lated from the data obtained directly from the experiment. Re-gression line B includes additional data using similar plantsgrown in soil with optimum nutrients.
DISCUSSION
FIG. 1. Respiratory rate at 20 C and concentration of inter-mediates of the EMP pathway in tomato fruits harvested at elevenstages of maturity (experiment 2).
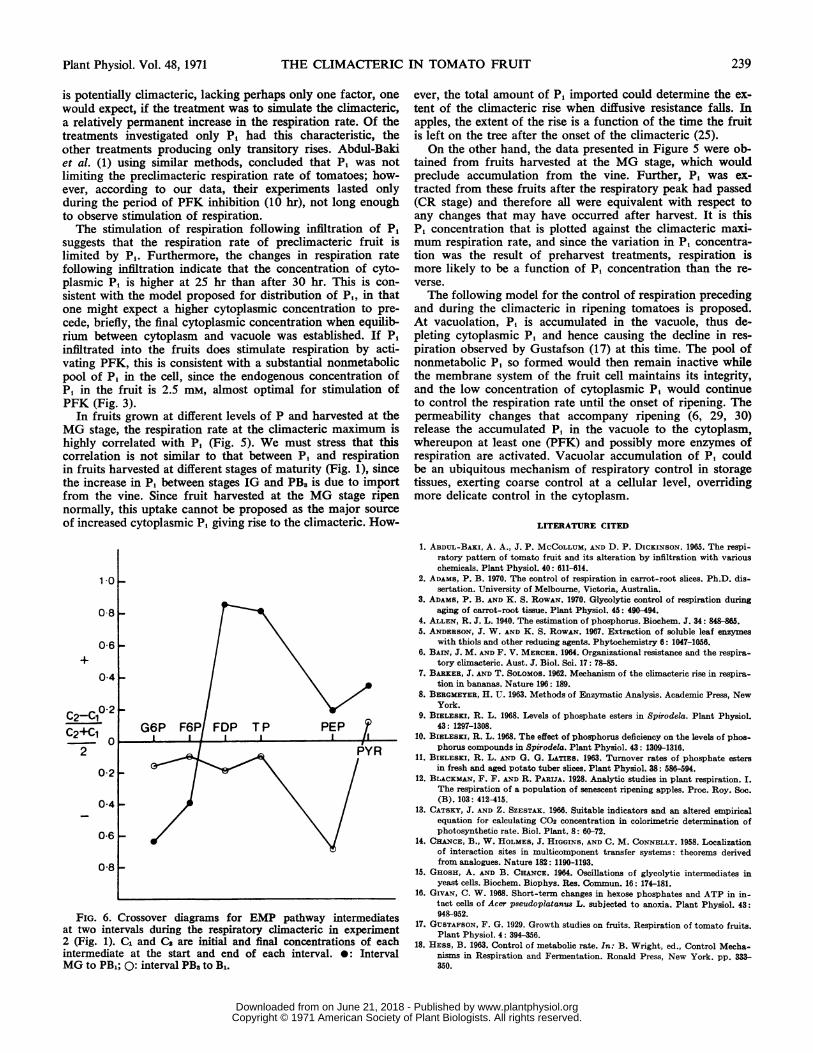
Figure 1 shows that the climacteric begins between the MGand PB1 stages. The commencement of the respiratory increaseis accompanied by a crossover at PFK (Fig. 6). Between the
237
.00U
.L
a,
E0
00-au0
II
I
www.plantphysiol.orgon June 21, 2018 - Published by Downloaded from Copyright © 1971 American Society of Plant Biologists. All rights reserved.

CHALMERS AND ROWAN Plant Physiol. Vol. 48, 1971
a water
201 1w. * 000.. ** *e o00 0 --
10
b40 .0* DNP40 .~~~~~0000
* 0030 ...-e*eeo0. ..
0
D 20 _ *-
cc 10_CO)
-oco
O C KCIcc'1 30-*-00Ir ~~~~0 000
.r0000 .0
10~~~~(D 0 ....*@@@-................. 0
.*d
KNaHPO30-4.00 000000000..00.
20@00 *o.ooo0
10
10 20 30 4G 50
Time (hours)
FIG. 4. The effect of treatment by vacuum infiltration on therate of respiration (26 C) of mature-green tomato fruits. Treat-ments (all 0.3 ml per fruit): a: Water; b: 1 mM DNP; c: 0.7M KC1; d: 0.7 M KNaHPO4, pH 7.5.
stages of PB. and B1, a crossover occurs as PK, while the rateof respiration is still increasing. Thus the increase in respira-tion is due in part to the sequential activation of glycolysis,initiated by the activation of PFK, and the reasons for in-volving P, in this stimulation have already been discussed. Theincrease in ATP (Fig. 2) appears to be too late to stimulatePFK. A similar increase in ATP occurs after the onset of theclimacteric in fruits of other species (27, 28, 34, 35). Pyruvatekinase extracted from plants is activated by monovalent cationsand Mg2" (23) but does not readily respond to effectors activeon the animal enzyme (FDP, ATP, and P,) (2). Again, in-creasing concentration of K+ or Mg2" in the cytoplasm throughincreasing permeability of the tonoplast could activate this re-action, but would lead to simultaneous rather than sequentialactivation of PFK and PK. A more likely explanation is thatPK is stimulated by ADP produced in the PFK reaction.The climacteric does not have the properties of a finely
regulated respiratory flux because the changes in levels ofmetabolites are from one steady state to another rather thancontinuous fluctuation (Fig. 1), and therefore it is reasonableto propose that it is the effect of release of a large pool of asingle effector such as P1. Results of the inifitration experi-ments, while not conclusive, support this contention. The ini-tial decrease in respiration after infiltration, also observed byAbdul-Baki et al. (1) and Spencer (31), is probably due toevacuation of CO2 from the vascular tissue and intercellular
spaces of the fruit. Recovery from this decrease in respirationvaries with the treatment, being rapid with water and DNP(3 hr), slower with KCI (4 hr), and requiring 12 hr after P,treatment. The treatment with 1 mm DNP is similar to thatused by Spencer (31) and Abdul-Baki et al. (1), and produceda similar response in respiration in this experiment. The lagfollowing infiltration of phosphate buffer is probably due to P,rather than cations, since recovery after KCI infiltration is morerapid. Salt at 0.2 M inhibits respiration (24) and, since 0.7 M
KCI was infiltrated, we would expect some inhibition of respi-ration until the KCl solution was diluted by diffusion within thefruit. The cation concentration in the phosphate buffer was
somewhat higher, but, in addition, Pi higher than 30 mm in-hibits PFK (Fig. 3) and thus the infiltrated P, would take longerthan KCl to be diluted bAlow inhibitory concentration. The in-crease in respiration following KCI infiltration could be "salt"respiration or stimulation of pyruvate kinase by K+.
In these infiltration experiments, the most important evi-dence that P, may be limiting respiration in preclimactericfruit comes not from the rise in respiration following the ini-tial inhibition, but rather from the observation that the respira-tion rate of the P,-treated fruit does not return or begin toreturn to the basal rate after the rise, while it does so in allother treatments. If it is assumed that the mature green fruit
0-11.-
c-
0)
a)
-
0
a)
.G
x
cc
)
cc
E
E
0
a)
0
cc
E
301
25
Ba
a a
*0* 0
0
0 //. -
0 .
0 0
0
0
0
0 0
0
3
Pi Concentration(mM)4
FIG. 5. The correlation between rate of respiration (20 C) oftomato fruits at the climacteric maximum with the concentrationof total P1 in the fruits at the end of the experiment. 0: Fruitsfrom plants grown for the experiment at various levels of P. Re-gression line A is calculated from these observations only. (F =
19.34, p = 0.1%). *: Fruits from plants grown in later experi-ments at an optimum level of P. Regression line B is calculatedfrom all observations.
238
www.plantphysiol.orgon June 21, 2018 - Published by Downloaded from Copyright © 1971 American Society of Plant Biologists. All rights reserved.
THE CLIMACTERIC IN TOMATO FRUIT
is potentially climacteric, lacking perhaps only one factor, onewould expect, if the treatment was to simulate the climacteric,a relatively permanent increase in the respiration rate. Of thetreatments investigated only P1 had this characteristic, theother treatments producing only transitory rises. Abdul-Bakiet al. (1) using similar methods, concluded that P, was notlimiting the preclimacteric respiration rate of tomatoes; how-ever, according to our data, their experiments lasted onlyduring the period of PFK inhibition (10 hr), not long enoughto observe stimulation of respiration.The stimulation of respiration following infiltration of P,
suggests that the respiration rate of preclimacteric fruit islimited by P1. Furthermore, the changes in respiration ratefollowing infiltration indicate that the concentration of cyto-plasmic P, is higher at 25 hr than after 30 hr. This is con-sistent with the model proposed for distribution of P,, in thatone might expect a higher cytoplasmic concentration to pre-cede, briefly, the final cytoplasmic concentration when equilib-rium between cytoplasm and vacuole was established. If P,infiltrated into the fruits does stimulate respiration by acti-vating PFK, this is consistent with a substantial nonmetabolicpool of P, in the cell, since the endogenous concentration ofP, in the fruit is 2.5 mm, almost optimal for stimulation ofPFK (Fig. 3).
In fruits grown at different levels of P and harvested at theMG stage, the respiration rate at the climacteric maximum ishighly correlated with P, (Fig. 5). We must stress that thiscorrelation is not similar to that between P1 and respirationin fruits harvested at different stages of maturity (Fig. 1), sincethe increase in P, between stages IG and PB3 is due to importfrom the vine. Since fruit harvested at the MG stage ripennormally, this uptake cannot be proposed as the major sourceof increased cytoplasmic P, giving rise to the climacteric. How-
101o-
08k
06
0-4p-
C2-Cl02
C2+C1 0
2
0 21
0-4I
061
G6P F6P FDP T P PEP
0-8
FIG. 6. Crossover diagrams for EMP pathway intermediatesat two intervals during the respiratory climacteric in experiment2 (Fig. 1). C1 and C2 are initial and final concentrations of eachintermediate at the start and end of each interval. *: IntervalMG to PB1; 0: interval PB3 to B1.
ever, the total amount of P1 imported could determine the ex-tent of the climacteric rise when diffusive resistance falls. Inapples, the extent of the rise is a function of the time the fruitis left on the tree after the onset of the climacteric (25).On the other hand, the data presented in Figure 5 were ob-
tained from fruits harvested at the MG stage, which wouldpreclude accumulation from the vine. Further, P, was ex-tracted from these fruits after the respiratory peak had passed(CR stage) and therefore all were equivalent with respect toany changes that may have occurred after harvest. It is thisP1 concentration that is plotted against the climacteric maxi-mum respiration rate, and since the variation in P, concentra-tion was the result of preharvest treatments, respiration ismore likely to be a function of P1 concentration than the re-verse.The following model for the control of respiration preceding
and during the climacteric in ripening tomatoes is proposed.At vacuolation, P, is accumulated in the vacuole, thus de-pleting cytoplasmic P, and hence causing the decline in res-piration observed by Gustafson (17) at this time. The pool ofnonmetabolic P, so formed would then remain inactive whilethe membrane system of the fruit cell maintains its integrity,and the low concentration of cytoplasmic P, would continueto control the respiration rate until the onset of ripening. Thepermeability changes that accompany ripening (6, 29, 30)release the accumulated P, in the vacuole to the cytoplasm,whereupon at least one (PFK) and possibly more enzymes ofrespiration are activated. Vacuolar accumulation of P, couldbe an ubiquitous mechanism of respiratory control in storagetissues, exerting coarse control at a cellular level, overridingmore delicate control in the cytoplasm.
LITERATURE CITED
1. ABDuJL-BAI, A. A., J. P. MCCOLLUM, AND D. P. DICKINsON. 1965. The respi-ratory pattern of tomato fruit and its alteration by infiltration with variouschemicals. Plant Physiol. 40: 611-614.
2. ADAMS, P. B. 1970. The control of respiration in carrot-root slices. Ph.D. dis-sertation. University of Melbourne, Victoria, Australia.
3. ADAMS, P. B. AND K. S. RowAN. 1970. Glycolytic control of respiration duringaging of carrot-root tissue. Plant Physiol. 45: 490-494.
4. ALLEN, R. J. L. 1940. The estimation of phosphorus. Biochem. J. 34: 848-85.5. ANDERSON, J. W. AND K. S. RowAN. 1967. Extraction of soluble leaf enzymes
with thiols and other reducing agents. Phytochemistry 6: 1047-1056.6. BArN, J. M. AND F. V. MERCER. 1964. Organizational resistance and the respira-
tory climacteric. Aust. J. Biol. Sci. 17: 78-85.7. BARKER, J. AND T. SOLOMOS. 1962. Mechanism of the dlimacteric rise in respira-
tion in bananas. Nature 196: 189.8. BERGMEYER, H. U. 1963. Methods of Enzymatic Analysis. Academic Press, New
York.9. BIELEsKI, R. L. 1968. Levels of phosphate esters in Spirodela. Plant Physiol.
43: 1297-1308.10. BIELEsKi, R. L. 1968. The effect of phosphorus deficiency on the levels of phos-
phorus compounds in Spirodela. Plant Physiol. 43: 1309-1316.11. BIELESKI, R. L. AND G. G. LATIES. 1963. Turnover rates of phosphate esters
in fresh and aged potato tuber slices. Plant Physiol. 38: 586594.12. BLACKMAN, F. F. AND R. PARIJA. 1928. Analytic studies in plant respiration. I.
The respiration of a population of senescent ripening apples. Proc. Roy. Soc.(B). 103: 412-415.
13. CATSKY, J. AND Z. SZESTAR. 1966. Suitable indicators and an altered empiricalequation for calculating CO2 concentration in colorimetric determination ofphotosynthetic rate. Biol. Plant. 8: 60-72.
14. CHANCE, B., W. HOLMES, J. HIGGINs, AND C. M. CONNELLY. 1958. Localizationof interaction sites in multicomponent transfer systems: theorems derivedfrom analogues. Nature 182: 1190-1193.
15. GHOSa, A. AND B. CHANCE. 1964. Oscillations of glycolytic intermediates inyeast cells. Biochem. Biophys. Res. Commun. 16: 174-181.
16. GIvAN, C. W. 1968. Short-term changes in hexose phosphates and ATP in in-tact cells of Acer pseudoplatanus L. subjected to anoxia. Plant Physiol. 43:948-952.
17. GUSTAFSON, F. G. 1929. Growth studies on fruits. Respiration of tomato fruits.Plant Physiol. 4: 394-356.
18. HEss, B. 1963. Control of metabolic rate. In: B. Wright, ed., Control Mecha-niSmS in Respiration and Fermentation. Ronald Press, New York. pp. 333-350.
-Pla.nt Physiol. Vol. 48, 1971 239
IPYR
www.plantphysiol.orgon June 21, 2018 - Published by Downloaded from Copyright © 1971 American Society of Plant Biologists. All rights reserved.

240 CHALMERS
19. HEWITT, E. J. 1966. Sand and water culture metlhods used in the study of plantnutrition. Commonw. Bur. Hort. Plant. Crops Tech. Commun. No. 22 (re-vised), 1-574.
20. KELLY, G. J. AND J. F. TURNER. 1969. The regulation of pea seed phospho-fructokinase by phosphoenolpyru-ate. Biochem. J. 115: 481-487.
21. KELLY, G. J. ANSD J. F. TURNER. 1970. The regulation of pea-seed phosphofruc-tokinase by 6-phosphogluconate, 3-phosphoglycerate, 2-phosphoglycerateand phosphoenolpyruvate. Biochim. Biophys. Acta 208: 360-367.
22. LOUGHMAN, B. C. 1960. Uptake and utilization of phosphate associatedl withrespiratory changes in potato tuber slices. Plant Physiol. 35: 418-424.
23. MILLER, G. AN-D H. J. EVANS. 1957. The influence of salts on py'ruvate kinasefrom tissues of higher plants. Plant Physiol. 32: 346-354.
24. PORATH. E. AND A. POLJAKOFF-MAYBER. 1968. The effect of salinity in thegrowtth medium on carbohydrate metabolism of pea root tips. Plant CellPhysiol. 9: 195-203.
25. RHODES, MI. J. C. AND L. S. C. WOOLTORTON. 1967. The respiratory climactericin apple fruits. The action of hydrolytic enzymes in peel tissue during theclimacteric period in fruit detached from the tree. Phytochemistry 6: 1-12.
26. ROWAAN-, K. S. 1958. Phosphorylated compounds in plants. II. The estimationof hexose phosphates and adenosine pyrophosphates in plant tissues by thenmethod of Slater. J. Exp. Bot. 9: 436-445.
27. Row.A.N, K. S., W. B. McGLASSON, AND H. K. PRATT. 1969. Changes in adeno-
ALND ROWAN Plant Physiol. Vol. 48, 1971
sine pyrophosphates in cantaloupe fruit ripening normally and after treat-ment with ethylene. J. Exp. Bot. 20: 145-155.
28. ROWAN, K. S., H. K. PRATT, AN-D R. N. ROBERTSON. 1958. The relatioiiship ofhigh energy phosphorus content, protein synthesis and the climacteric rise inrespiration in ripening avocado and tomato fruits. Aust. J. Biol. Sci. 11: 320-335.
29. SACHER, J. A. 1962. Relations between changes in membrane permeability andthe climacteric in bananas and avocadlo. Nature 195: 577-578.
30. SACHER, J. A. 1966. Permeability characteristics and amino acid incorporationduring senescence (ripening) of banana tissue. Plant Physiol. 41: 701-708.
31. SPENCER, M. S. 1959. Ethylene nmetabolism in ripening tomato fruit. Can. J.Biochem. Physiol. 37: 53-59.
32. STREHLER, B. L. AND J. R. TOTTER. 1952. Firefly luminescence in the study ofenergy transport mechanism. I. Substrate and enzyme determinations. Arch.Biochem. Biophys. 40: 2841.
33. WADE, N. L. 1970. Physiology of banana ripening: A study of the effects ofethylene and kinetin, of cell permeability, and of changes in RNA anid pro-tein content. Ph.D. dissertation. University of Sydney, N.S.W., Australia.
34. YOUNnG, R. E. AND J. B. BIALE. 1967. Phosphorylation in avocado fruit slicesin relation to the respiratory climacteric. Plant Physiol. 42: 1357-1362.
35. YOUNG, R. E., C. S. POPPER, AND J. B. BIALE. 1962. Specific activity of plhos-phate esters of avocado fruit tissue as determined by neutron activatiOln.Plant Physiol. 37: xxxiii.
www.plantphysiol.orgon June 21, 2018 - Published by Downloaded from Copyright © 1971 American Society of Plant Biologists. All rights reserved.


















