
The Cdk inhibitor p27 in human cancer: prognostic potential and relevance to anticancer therapy
15
p27 (also known as KIP1, and encoded by CDKN1B) is an atypical tumour suppressor that regulates G0 to S phase transitions by binding to and regulating the activity of cyclin-dependent kinases (Cdks) 1,2 . In G0 and early G1, p27 translation and protein stability are maximal and it binds and inhibits cyclin E–CDK2 (REF. 1,2). The progres- sive decrease in p27 during G1 allows cyclin E–CDK2 and cyclin A–CDK2 to activate the transcription of genes that are required for the G1–S transition and to partici- pate in the initiation of DNA replication 3 . D-type cyclin– Cdks are activated by mitogens to cause G0 exit and G1 pro- gression. p27 has a dual role in the regulation of cyclin D– CDK4 and cyclin D–CDK6 complexes. Under adverse conditions, including contact inhibition 4 and after UV irradiation, p27 can inhibit D-type cyclin–Cdks. In early G1 to mid-G1, p27 also promotes assembly and nuclear import of cyclin D1–Cdk complexes 1 . Tyrosine phospho- rylation of p27 leads to activation of assembled cyclin D–Cdks 4,5 (see below). Many signal transduction networks regulate p27, and it integrates diverse signals into a final decision between proliferation and cell-cycle exit. The Cdk-inhibitory activ- ity of p27 is controlled by its concentration, subcellular localization and phosphorylation status. The levels and activity of p27 increase in response to cell density 6 , dif- ferentiation signals 1,7 , following loss of adhesion to the extracellular matrix 8 , and in response to growth-inhibitory signalling by transforming growth factor β (TGFβ) or by the drug lovastatin 1,6 . In addition to its effects on the cell cycle, p27 also regulates cell migration at least in part independently from its ability to bind Cdks 8 . Although p27 is not a classic tumour suppressor like p53, as it is rarely mutated or deleted in human cancers 9 , it is fre- quently deregulated in cancer: p27 protein levels are reduced or the protein is mislocalized in most human cancers and this is associated with a poor prognosis. Oncogenic activation of receptor tyrosine kinases (RTK), phosphatidylinositol 3-kinase (PI3K), SRC, or Ras–mitogen activated protein kinase (MAPK) pathways cooperate to inactivate p27 or accelerate its proteolysis in human cancers. Here we review the mechanisms leading to aberrant p27 regulation in human tumours and cover the multivariate prognostic studies of p27 in common human epithelial malignancies: cancers of the lung, head and neck, colon, prostate, ovary and breast. The potential for p27 to serve as a predictor of response to chemotherapy, radiation and hormonal therapies is also discussed. p27 regulation in normal and malignant cells CDKN1B transcription. Whereas cell cycle-dependent changes in p27 are largely post-transcriptionally regu- lated, several transcription factors act on the CDKN1B promoter. Forkhead box class O family (FoxO) proteins activate CDKN1B transcription, in response to cytokines or promyelocytic leukaemia protein (PML) and nuclear Akt signalling 10–12 . Besides regulating CDKN1B tran- scription, FOXO4 inhibits Akt to promote nuclear p27 localization and decreases expression of COP9 subunit 5 (COPS5; also known as CSN5 and JAB1), a protein that mediates p27 degradation (see below) 13 . FOXM1, also increases p27 stability 14 . Thus, p27 regulation by FoxO proteins exceeds effects on transcription. *Braman Family Breast Cancer Institute, and Department of Biochemistry and Molecular Biology, University of Miami Miller School of Medicine, 1580 NW 10th Avenue, Miami, Florida 33136, USA. ‡ Division of Medical Biochemistry, Biocenter, Innsbruck Medical University, Fritz-Pregl-Str. 3, A-6020 Innsbruck, Austria. Correspondence to J.M.S. e-mail: [email protected] doi:10.1038/nrc2347 The Cdk inhibitor p27 in human cancer: prognostic potential and relevance to anticancer therapy Isabel M. Chu*, Ludger Hengst ‡ and Joyce M. Slingerland* Abstract | The cyclin-dependent kinase (Cdk) inhibitor p27 (also known as KIP1) regulates cell proliferation, cell motility and apoptosis. Interestingly, the protein can exert both positive and negative functions on these processes. Diverse post-translational modifications determine the physiological role of p27. Phosphorylation regulates p27 binding to and inhibition of cyclin–Cdk complexes, its localization and its ubiquitin-mediated proteolysis. In cancers, p27 is inactivated through impaired synthesis, accelerated degradation and by mislocalization. Moreover, studies in several tumour types indicate that p27 expression levels have both prognostic and therapeutic implications. NATURE REVIEWS | CANCER VOLUME 8 | APRIL 2008 | 253 REVIEWS © 2008 Nature Publishing Group
Transcript of The Cdk inhibitor p27 in human cancer: prognostic potential and relevance to anticancer therapy

p27 (also known as KIP1, and encoded by CDKN1B) is an atypical tumour suppressor that regulates G0 to S phase transitions by binding to and regulating the activity of cyclin-dependent kinases (Cdks)1,2. In G0 and early G1, p27 translation and protein stability are maximal and it binds and inhibits cyclin E–CDK2 (Ref. 1,2). The progres-sive decrease in p27 during G1 allows cyclin E–CDK2 and cyclin A–CDK2 to activate the transcription of genes that are required for the G1–S transition and to partici-pate in the initiation of DNA replication3. D-type cyclin– Cdks are activated by mitogens to cause G0 exit and G1 pro-gression. p27 has a dual role in the regulation of cyclin D– CDK4 and cyclin D–CDK6 complexes. Under adverse conditions, including contact inhibition4 and after UV irradiation, p27 can inhibit D-type cyclin–Cdks. In early G1 to mid-G1, p27 also promotes assembly and nuclear import of cyclin D1–Cdk complexes1. Tyrosine phospho-rylation of p27 leads to activation of assembled cyclin D–Cdks4,5 (see below).
Many signal transduction networks regulate p27, and it integrates diverse signals into a final decision between proliferation and cell-cycle exit. The Cdk-inhibitory activ-ity of p27 is controlled by its concentration, subcellular localization and phosphorylation status. The levels and activity of p27 increase in response to cell density6, dif-ferentiation signals1,7, following loss of adhesion to the extracellular matrix8, and in response to growth-inhibitory signalling by transforming growth factor β (TGFβ) or by the drug lovastatin1,6. In addition to its effects on the cell cycle, p27 also regulates cell migration at least in part independently from its ability to bind Cdks8. Although
p27 is not a classic tumour suppressor like p53, as it is rarely mutated or deleted in human cancers9, it is fre-quently deregulated in cancer: p27 protein levels are reduced or the protein is mislocalized in most human cancers and this is associated with a poor prognosis.
Oncogenic activation of receptor tyrosine kinases (RTK), phosphatidylinositol 3-kinase (PI3K), SRC, or Ras–mitogen activated protein kinase (MAPK) pathways cooperate to inactivate p27 or accelerate its proteolysis in human cancers. Here we review the mechanisms leading to aberrant p27 regulation in human tumours and cover the multivariate prognostic studies of p27 in common human epithelial malignancies: cancers of the lung, head and neck, colon, prostate, ovary and breast. The potential for p27 to serve as a predictor of response to chemotherapy, radiation and hormonal therapies is also discussed.
p27 regulation in normal and malignant cellsCDKN1B transcription. Whereas cell cycle-dependent changes in p27 are largely post-transcriptionally regu-lated, several transcription factors act on the CDKN1B promoter. Forkhead box class O family (FoxO) proteins activate CDKN1B transcription, in response to cytokines or promyelocytic leukaemia protein (PML) and nuclear Akt signalling10–12. Besides regulating CDKN1B tran-scription, FOXO4 inhibits Akt to promote nuclear p27 localization and decreases expression of COP9 subunit 5 (COPS5; also known as CSN5 and JAB1), a protein that mediates p27 degradation (see below)13. FOXM1, also increases p27 stability14. Thus, p27 regulation by FoxO proteins exceeds effects on transcription.
*Braman Family Breast Cancer Institute, and Department of Biochemistry and Molecular Biology, University of Miami Miller School of Medicine, 1580 NW 10th Avenue, Miami, Florida 33136, USA. ‡Division of Medical Biochemistry, Biocenter, Innsbruck Medical University, Fritz-Pregl-Str. 3, A-6020 Innsbruck, Austria. Correspondence to J.M.S. e-mail: [email protected]:10.1038/nrc2347
The Cdk inhibitor p27 in human cancer: prognostic potential and relevance to anticancer therapyIsabel M. Chu*, Ludger Hengst‡ and Joyce M. Slingerland*
Abstract | The cyclin-dependent kinase (Cdk) inhibitor p27 (also known as KIP1) regulates cell proliferation, cell motility and apoptosis. Interestingly, the protein can exert both positive and negative functions on these processes. Diverse post-translational modifications determine the physiological role of p27. Phosphorylation regulates p27 binding to and inhibition of cyclin–Cdk complexes, its localization and its ubiquitin-mediated proteolysis. In cancers, p27 is inactivated through impaired synthesis, accelerated degradation and by mislocalization. Moreover, studies in several tumour types indicate that p27 expression levels have both prognostic and therapeutic implications.
NATURE REVIEWS | cancer VOLUME 8 | APRIL 2008 | 253
REVIEWS
© 2008 Nature Publishing Group

Menin, encoded by the MEN1 (multiple endo-crine neoplasia) tumour suppressor gene, maintains CDKN1B expression in differentiated pancreatic islet cells15. Whereas mature Men1 mutant mice develop insulinomas with reduced p27, young animals express p27 at normal levels in hyperplastic and dysplastic islets, indicating that loss of p27 in these insulinomas requires additional molecular events16. Interestingly, a germline frameshift mutation in CDKN1B dramatically reduced p27 expression and caused multiple endocrine neoplasia (MENX)17 in rats. Germline mutations in CDKN1B also exist in some human patients with the MEN1 phenotype17,18.
Although MYC represses the CDKN1B promoter19, its effects on p27 are predominantly post-transcriptional. MYC induces expression of proteins, including cyclin D1, that sequester and inhibit p27 (Ref. 1). MYC also induces expression of Cullin 1 (CUL1)20 and cyclin-dependent kinase subunit 1B (CKS1B)21, which are involved in p27 proteolysis (see below).
The E2F1 transcription factor, which is itself activated by cyclin–CDK2, activates the CDKN1B promoter, lead-ing to feedback inhibition of E2F1 action22. Finally, tran-scription factors SP1 and NFY23, HES1 (Ref. 24) and the vitamin D3 receptor25 all regulate p27 expression and might modulate p27 levels during development.
p27 translation. CDKN1B mRNA translation is maximal in quiescence and early G1 and decreases after mitogen stimulation26 as cells progress toward S-phase6,27,28 (fIG. 1). Cell cycle-dependent translation of p27 is mediated by the 5′ UTR of CDKN1B28, which contains an upstream open-reading frame and an internal ribosome entry site (IRES). The IRES maintains CDKN1B translation
during quiescence when most cellular cap-dependent translation is reduced29,30. Defective IRES-mediated CDKN1B translation due to deficiency in the pseudo-uridine synthase gene in individuals with congenital X-linked dyskeratosis may contribute to their increased susceptibility to cancer31.
Several proteins regulate CDKN1B translation. The polypyrimidine tract-binding protein, PTB (also known as PTBP1), binds the CDKN1B IRES to increase IRES-dependent translation. Small interfering RNA-mediated PTB depletion shortens G1 phase and reduces p27 levels32. By contrast, Elav (Drosophila embryonic lethal, abnormal vision-like 1) family members, ELAVL1 (also known as Hu antigen R) and ELAVL4 (Hu antigen D), bind the CDKN1B IRES30,33 to inhibit translation30. ELAVL1 depletion increases CDKN1B translation, leading to G1 arrest30. Overexpression of ELAVL1 in ovarian34,35 and colon cancer cells36 might account for the reduced p27 levels observed in a subset of these can-cers. Rho37 and CDK4 (Ref. 38) can both impair CDKN1B translation through its 3′ UTR.
Translation of CDKN1B can also be downregulated by microRNAs (miRNAs), which also act on the 3′ UTR (reviewed in Ref. 39). This highly conserved mechanism regulating p27 protein expression is found in flies and humans: in Drosophila melanogaster, inac-tivation of Dicer 1, the double-stranded RNase III iso-zyme that is essential for miRNA biogenesis (but not small interfering RNAs), delayed the G1–S transition of germline stem cells through increased translation of the p27 homologue, dacapo. Using independent experimental approaches, downregulation of human p27 by miRNAs was recently demonstrated in an elegant screen for p27 regulatory miRNAs, in cells for which the cell cycle was inhibited by Dicer inactiva-tion, and by a bioinformatics screen. Two polycistronic miRNAs sharing the same seed sequence, miR-221 and miR-222 (miR-221/222), bind to the CDKN1B 3′ UTR and inhibit its translation39. Downregulation of high endogenous levels of miR-221/222 in different tumour cells increased p27 expression and inhibited tumour cell proliferation39. Dead end 1 (DND1) pre-vents the access of miR221/222 to the p27 transcript40, and can protect the p27 transcript from miRNA-medi-ated downregulation. Interestingly, increased miR-221/222 polycistron expression correlates with poor prognosis in glioblastoma and in pancreatic carcino-mas (Ref. 39). These miRNAs are also overexpressed in prostate39,41, colon and stomach cancer42, thyroid papillary carcinoma43 and in chronic lymphocytic leukaemia44. Thus, miRNA-mediated inhibition of p27 translation emerges as a novel mechanism that can contribute to the reduced levels of p27 frequently observed in human cancers.
p27 proteolysis. p27 proteolysis rises dramatically dur-ing G1 phase progression, and the p27 half-life falls five- to eightfold during G0 to S phase45. Multiple dif-ferent mechanisms regulate ubiquitin-dependent p27 proteolysis. Constitutive activation of a number of these degradation pathways contributes to oncogenesis.
At a glance
• p27 (also known as KIP1, and encoded by CDKN1B) is regulated by multiple signal transduction pathways in normal and malignant cells.
• CDKN1B transcription can be regulated by the FoxO family and genetic defects that reduce CDKN1B transcription can predispose to multiple endocrine neoplasia (MEN)-like phenotypes.
• CDKN1B 5′UTR mediates its cell cycle-dependent translation and several proteins can bind the CDKN1B IRES to modulate its translation.
• micro-RNA-mediated inhibition of p27 translation emerges as a novel mechanism that can reduce p27 in some human cancers.
• p27 proteolysis is initiated by several different mechanisms.
• Tyrosine (Tyr) phosphorylation of p27 by Abl and Src family kinases reduces p27–CDK2 inhibition and transforms p27 from inhibitor to substrate of cyclin–CDK2 complexes.
• p27 phosphorylation at Thr157 and Thr198 by members of the AGC kinase family promotes assembly of p27–cyclin D–CDK4/6, but catalytic activation requires tyrosine phosphorylation.
• Cytoplasmic mislocalization of p27 is activated by AGC family kinases and contributes to RHOA inhibition and increased cell motility in cancers.
• p27 levels are reduced in the most common and lethal human epithelial cancers and this is associated with poor patient outcome.
• Restoration of p27 levels and/or nuclear localization may predict response to molecular therapies that target EGFR and IGFR families, MAP2K (also known as MEK), BCR-ABL and SRC.
R E V I E W S
254 | APRIL 2008 | VOLUME 8 www.nature.com/reviews/cancer
© 2008 Nature Publishing Group

Nature Reviews | Cancer
G0/G1 S phase
KPC mediated degradation
p27 translation
Thr187 phosphorylation
p27 protein level
Restriction point
Nuclear export
SCFSKP2-mediated degradation
Thr187-phosphorylation dependent p27 proteolysis. Best characterized and probably most potent is the polyubiquitylation of p27 by the SCFSKP2 (S-phase kinase associated protein 1 (SKP1)/Cullin/F-Box protein: S-phase kinase associated protein 2 (SKP2)) complex, E3 ubiquitin ligase45. The SCFSKP2 RING-type ubiquitin ligase consists of SKP1, CUL1, the ring finger protein RBX1 (also known as ROC1), the F-box pro-tein SKP2 and the accessory protein CKS1B (reviewed in Refs 45,46). Cyclin–CDK2-bound, threonine 187 (Thr187)-phosphorylated p27 binds to the phosphate-binding site of CKS1B, to SKP2 and to the CKS1–SKP2 interface. Cyclin A–CDK2 binding further stimulates p27 recruitment to SCFSKP2, because cyclin A binds SKP2 and CDK2 binds CKS1B.
RBX1 can be replaced in the SCFSKP2 complex by Ro52 (also known as tripartite motif-containing pro-tein 21 (TRIM21)). Ro52 is part of another SCF-like complex that ubiquitylates Thr187-phosphorylated p27. Importantly, Ro52 depletion causes a p27-dependent G1 arrest of HeLa cells47, highlighting a requirement for Ro52–SCF SKP2 in p27 proteolysis in these cells.
Tyrosine phosphorylation regulates Cdk inhibition by p27 and p27 proteolysis. Efficient ubiquitylation by SCFSKP2 requires prior p27 phosphorylation at Thr187 by active cyclin E- or cyclin A–CDK2. Although free active cyclin– CDK2 can phosphorylate Cdk-bound p27 at Thr187 efficiently in vitro, p27-bound CDK2 is catalytically inac-tive48. Recent data indicate that tyrosine phosphorylation of p27 by Abl and Src family kinases initiates the transi-tion of p27 from inhibitor of cyclin E–CDK2 in G0 to substrate of cyclin–CDK2 in G1 (Refs 5,49).
There are three tyrosines in p27 at residues 74, 88 and 89; all within its Cdk-inhibitory domain. Tyrosine 88 (Tyr88) is highly conserved among all Cip and Kip fam-ily Cdk inhibitors, including human p21 (also known as WAF1) and p57 proteins, whereas tyrosine 74 (Tyr74) is specific for p27-type inhibitors. Crystal structure analysis shows that Tyr88 is part of a 310-helix in p27 that inserts into the CDK2 catalytic cleft and displaces ATP50. Tyr74 forms hydrophobic interactions with CDK2 (fIG. 2).
Overexpression of ABL or SRC increases detection of Thr187-phosphorylated p27 and decreases p27 levels and stability. ABL and LYN kinases primarily phos-phorylate Tyr88 in vitro, whereas SRC phosphorylates both Tyr88 and Tyr74. SRC activation and SRC-p27 binding precede loss of p27 during G1 progression. SRC-phosphorylated p27 has reduced steady state binding to cyclin E–CDK2 potentially due to loss of the interaction of Tyr74 phosphorylated p27 with sites on the N-lobe of CDK2. Although Tyr88 phospho-rylation only weakly reduces p27 affinity for CDK2, NMR structure analysis indicates that the inhibitory 310 helix of CDK2-bound Tyr88 phosphorylated p27 is ejected from the catalytic cleft of cyclin–CDK2 (Refs 5,49), opening up the ATP binding pocket of the kinase. Thus, phosphorylation of p27 at Tyr88 would allow p27-bound cyclin–CDK2 to phosphorylate p27 at Thr187 to activate SCFSKP2-mediated p27 proteolysis during the G1–S transition5,49 (fIGs 2, 3).
Thr187-phosphorylation independent p27 proteolysis. SKP2-mediated p27 proteolysis can also occur in G1 independently of Thr187 phosphorylation51, indicating that an additional SKP2-dependent ubiquitin ligase com-plex exists that may not require CKS1B for p27 recruit-ment. Recently a complex of SKP2, Cullin 4A (CUL4A), damaged-DNA binding protein 1 (DDB1) and the COP9 signalosome was shown to target p27 for proteolysis52. That this complex regulates p27 proteolysis is supported by the observation that another COP9 signalosome sub-unit, COPS5, also binds to p27 independently of Thr187 and accelerates its proteolysis53.
Export of p27 in early G1 also allows degradation of cytoplasmic p27 by a SKP2- and Thr187-independent pathway45. In early G1, p27 phosphorylation at serine 10 (Ser10) by kinase-interacting stathmin (KIS)54 or the minibrain-related kinase, MIRK (or dual tyrosine phosphorylation related kinase 1, DYRK1)55 increases p27 binding, through its nuclear export signal (NES), to CRM1 (or exportin 1) to promote nuclear export of p27 (Refs 56–58). The export of p27 requires loss of cyclin–CDK2 binding, as CRM1 binds p27 in its CDK2 interaction site. Cytoplasmic p27 can be ubiquitylated by the ubiquitin ligase Kip1 ubiquitylation-promoting complex (KPC)45 (fIG. 3).
In quiescent G0 cells, another proteolytic mechanism regulates p27 stability59. The degradation of p27 in G0 might involve cyclin–Cdk binding or binding of another protein to its Cdk-binding motif. Phosphorylation of threonine 198 (Thr198) of p27 by kinases, including AMP-activated protein kinase (AMPK), seems to stabi-lize p27 in G0 (Refs 60,61) and thus Thr198 might play
Figure 1 |Schematicmodelofp27levelsandregulationduringG0/G1toSphase.During G1 progression, p27 (also known as KIP1) translation falls. Nuclear export and Kip1 ubiquitylation-promoting complex (KPC)-mediated degradation increase in early G1. Threonine 187 (Thr187) phosphorylation increases in late G1 and targets p27 for SCFSKP2 (S-phase kinase associated protein 1 (SKP1)/Cullin/F-Box protein: S-phase kinase associated protein 2 (SKP2))-mediated proteolysis.
R E V I E W S
NATURE REVIEWS | cancer VOLUME 8 | APRIL 2008 | 255
© 2008 Nature Publishing Group

Nature Reviews | Cancer
a
b
SRC/ABL
p27
L83
p27
Cdk2 N-lobeCdk2 N-lobe
Cdk2 C-lobeCdk2 C-lobe
F80
V64
L134
L83
Y88 pY88
L134
F80
F82F82
V64
Y88
SRC/ABL
p27 p27
L25
pY74
L67
V79
Cdk2 C-lobe Cdk2 C-lobe
Cdk2 N-lobeCdk2 N-lobe
V30
L67
L25
Y74
V79V30
a part in this pathway. Genetic studies using p27S10A knock-in mice also indicate that Ser10 phosphorylation is necessary for p27 stability in G0 (Refs 59,62).
Activation of p27 proteolysis in human cancersOncogenic signalling accelerates p27 proteolysis. Many human cancers show increased Src family levels or activ-ity usually due to oncogenic RTK activation. Epidermal growth factor receptor (EGFR) and ERBB2 both bind SRC to catalyse mutual kinase activation63. In epithelial cells, overexpression of EGFR family members activates p27 proteolysis and Ras-driven p27 proteolysis might involve both Rho- and MAPK-dependent mechanisms7. The ERBB2 gene is amplified and EGFR is overexpressed in up to 30% of primary breast cancers and both are also overexpressed in other epithelial malignancies64,65. In human breast cancer, both ERBB2 amplification66,67 and SRC activation49 are statistically correlated with reduced levels of p27.
The identification of pathways that drive p27 proteoly-sis has opened new avenues for therapeutic intervention in cancer. MAP2K (also known as MEK) inhibition with U0126 (Refs 68,69), inhibition of ERBB2 by trastuzumab (also known as Herceptin)70, of EGFR by gefitinib71 and
of both by lapatinib72, can all reverse p27 proteolysis mediated by overexpression of EGFR-family members. Moreover, combining SRC inhibitors with antioestrogen therapies might delay or reverse resistance to hormone therapy in breast cancer49 (see below).
Whereas RTK and SRC activation are major media-tors of p27 loss in human epithelial cancers, activa-tion of ABL, LYN, LCK and FYN would promote p27 proteolysis in lymphoma and other haematopoietic malignancies73,74. Levels of p27 are reduced in BCR-ABL transformed chronic myelogenous leukaemia (CML) cells75. Inhibition of BCR-ABL kinase in CML cells by imatinib inhibits Tyr88 phosphorylation of p27, restores p27 expression and inhibits cell proliferation5. Interestingly, imatinib resistance can emerge due to overexpression of LYN76, which cannot be inactivated by imatinib5, indicating a potential role for dual Abl and Src family inhibitors to prevent CML progression.
SKP2 overexpression reduces p27 levels in a subset of human tumours. Mutational inactivation of the phos-phatase and tensin homologue (PTEN) tumour suppres-sor gene in primary human cancers77 contributes to loss of p27. PTEN loss increases SKP2 expression7 and lack
Figure 2 |Modellingeffectsoftyrosinephosphorylationonp27onp27–cDK2.a | Hydrophobic interactions between p27Tyr74 and leucine 67 (Leu67) and valine 79 (Val79) in the N-lobe of cyclin-dependent kinase 2 (CDK2) are disrupted by tyrosine (Tyr) phosphorylation at p27Tyr74 and would reduce p27–CDK2 affinity. b | p27 phosphorylation at Tyr88 disrupts hydrophobic interactions between Tyr88 in the inhibitory 310 helix of p27 with phenylalanine 80 (Phe80), Phe82, Leu83 and Leu134 in the catalytic cleft of CDK2. This would eject p27 from the ATP binding site of CDK2, to enable partially active p27-bound cyclin–CDK2 to phosphorylate p27 at threonine 187 (Thr187) and promote SCFSKP2-mediated p27 proteolysis. Figure courtesy of Amjad Farooq. SCFSKP2, S-phase kinase associated protein 1 (SKP1)/Cullin/F-Box protein: S-phase kinase associated protein 2 (SKP2).
R E V I E W S
256 | APRIL 2008 | VOLUME 8 www.nature.com/reviews/cancer
© 2008 Nature Publishing Group
RanGTP CRM1
Nature Reviews | Cancer
Cyclin E–CDK2 phosphorylates T187
SCFSKP2Degradation
PI3K
PDK1
AGC kinases
Nuclear import blocked
p27 losesinhibitory action
SRC/ABL
SRC/ABL
Degradation by KPC ubiquitin ligase
Degradation
p27P
KPC1KPC2
p27P
P
KISDYRK
Nucleus
Cytoplasm
p27 dissociates from CDK2
ROCKRHOA
p27CDK4/6
Cyclin D–CDK assembly
P
Cyclin D P
P
Activated kinase
p27P S10
p27P
P
p27 binds and inhibits cyclin E–CDK2
T157
T198
Y74Y88P P
CDK2
Cyclin Ep27
PT187
P PCDK2
Cyclin Ep27CDK2
Cyclin Ep27
p27CDK4/6P
P PCyclin D
p27CDK4/6
P PCyclin D
CDK4/6
Cyclin D
Activated kinase
CDK2
Cyclin E
Pp27
P
P
G0/G1 S phase
P Y74 and Y88
P S10 P
P PT157
T198
T187
of PTEN staining correlates with reduced p27 protein levels in several human cancers78. Increased SKP2 levels have been observed in human lymphomas79,80 and in primary colon81,82, breast83, head and neck84–86, prostate87, lung88 and gastric89 cancers. Although SCFSKP2 has many substrates, including the Cdk inhibitors p21 and p57, the RB1-related protein p130, MYC, ORC1 and CDT1, deletion of Cdkn1b was sufficient to rescue the Skp2–/– mouse phenotype90,91. This suggests that p27 is the key substrate of SKP2 ubiquitin ligases in vivo. Although high SKP2 levels are invariably correlated with reduced p27, in most tumours loss of p27 occurs without increased SKP2. Thus, reduced p27 arises from mechanisms other than SKP2 overexpression in most human cancers.
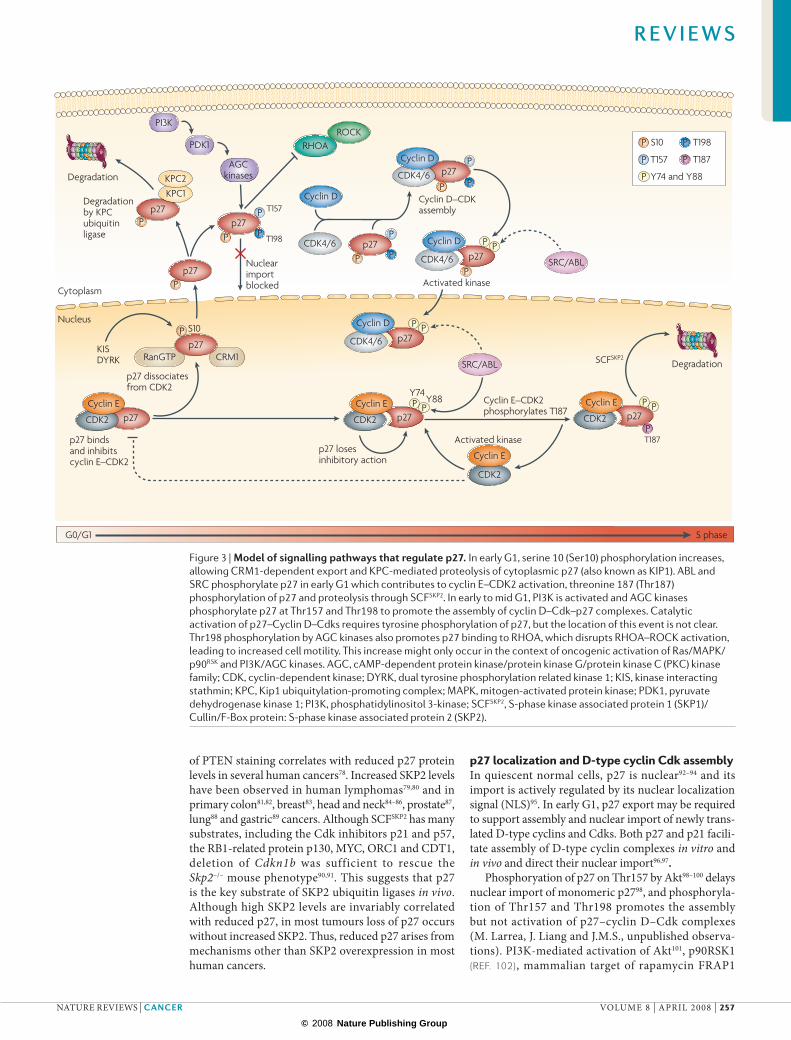
p27 localization and D-type cyclin Cdk assemblyIn quiescent normal cells, p27 is nuclear92–94 and its import is actively regulated by its nuclear localization signal (NLS)95. In early G1, p27 export may be required to support assembly and nuclear import of newly trans-lated D-type cyclins and Cdks. Both p27 and p21 facili-tate assembly of D-type cyclin complexes in vitro and in vivo and direct their nuclear import96,97.
Phosphoryation of p27 on Thr157 by Akt98–100 delays nuclear import of monomeric p2798, and phosphoryla-tion of Thr157 and Thr198 promotes the assembly but not activation of p27–cyclin D–Cdk complexes (M. Larrea, J. Liang and J.M.S., unpublished observa-tions). PI3K-mediated activation of Akt101, p90RSK1 (Ref. 102), mammalian target of rapamycin FRAP1
Figure 3 |Modelofsignallingpathwaysthatregulatep27.In early G1, serine 10 (Ser10) phosphorylation increases, allowing CRM1-dependent export and KPC-mediated proteolysis of cytoplasmic p27 (also known as KIP1). ABL and SRC phosphorylate p27 in early G1 which contributes to cyclin E–CDK2 activation, threonine 187 (Thr187) phosphorylation of p27 and proteolysis through SCFSKP2. In early to mid G1, PI3K is activated and AGC kinases phosphorylate p27 at Thr157 and Thr198 to promote the assembly of cyclin D–Cdk–p27 complexes. Catalytic activation of p27–Cyclin D–Cdks requires tyrosine phosphorylation of p27, but the location of this event is not clear. Thr198 phosphorylation by AGC kinases also promotes p27 binding to RHOA, which disrupts RHOA–ROCK activation, leading to increased cell motility. This increase might only occur in the context of oncogenic activation of Ras/MAPK/p90RSK and PI3K/AGC kinases. AGC, cAMP-dependent protein kinase/protein kinase G/protein kinase C (PKC) kinase family; CDK, cyclin-dependent kinase; DYRK, dual tyrosine phosphorylation related kinase 1; KIS, kinase interacting stathmin; KPC, Kip1 ubiquitylation-promoting complex; MAPK, mitogen-activated protein kinase; PDK1, pyruvate dehydrogenase kinase 1; PI3K, phosphatidylinositol 3-kinase; SCFSKP2, S-phase kinase associated protein 1 (SKP1)/Cullin/F-Box protein: S-phase kinase associated protein 2 (SKP2).
R E V I E W S
NATURE REVIEWS | cancer VOLUME 8 | APRIL 2008 | 257
© 2008 Nature Publishing Group

Nature Reviews | Cancer
Larynx
Oesophageal
Lung
Oral cancer
Bladder
Prostate
GastricHepatoma
Melanoma
Endometrial
Colon
Lymphoma
Astrocytoma
Breast
Glioma
Thyroid
Ovary
Leukaemia
(or mTOR), and serum and glucocorticoid inducible kinase (SGK) may all contribute to p27 phosphoryla-tion at Thr157 and Thr198 and facilitate p27–cyclin D– Cdk assembly in different cell types (F. Hong, M. Larrea, J. Liang and J.M.S., unpublished observa-tions). Additional phosphorylation events may regulate p27–cyclin D–Cdk import, and tyrosine phosphoryla-tion of p27 allows the activation of this complex4,5 (fIG. 3). Of note, tyrosine dephosphorylation following granulocyte colony stimulating factor (GCSF) treat-ment led to redistribution of p27 from CDK4 to CDK2 (Ref. 103).
Active p27–cyclin D–CDK4/6 complexes accumulate in the cytoplasm of many transformed cells104–106. This may be driven by MAP2K107 or by Ras108 and Akt98,109,110. Oncogenic activation of PI3K- and MEK-dependent kinases in cancer cells may promote cytoplasmic accu-mulation of p27 observed by immunostaining in human breast, ovarian, colon, prostate, oesophageal and thyroid cancers98,109–113. It remains mechanistically unclear why oncogenic activation of receptor tyrosine kinases drive PI3K-mediated cytoplasmic mislocalization of p27 in some cancers and p27 proteolysis in others.
Cytoplasmic p27, migration and metastasisThe interaction of p27 with proteins that regulate cytoskeleton dynamics is, at least in part, independent of its Cdk inhibitory action. Depending on the cellular context or the experimental approach, p27 can inhibit or promote cell motility8. In fibrosarcoma cells and normal fibroblasts, p27 inhibits migration by binding to stathmin, a microtubule-destabilizing protein114, and impairs stathmin binding to tubulin, causing increased microtubule polymerization114.
By contrast, in hepatocellular carcinoma cells, cyto-plasmic p27 co-localizes with actin fibres to stimulate cell migration through action on GTPase Rac106,115. p27 null mouse embryonic fibroblasts (MEFs) show decreased cell motility, increased actin stress fibres and focal adhesions, and increased RHOA activity. In Cdkn1b–/– MEFs, overexpressed p27 binds and inhibits RHOA to reduce cell motility8. In MCF-7 breast cancer cells, overexpression of NLS defective p27 increased cytoplasmic p27, reduced RHOA activity and increased cell motility116 (fIG. 3). Overexpression of a p27 mutant localizing to the cytoplasm increased melanoma cell motility and metastasis in vivo117. When a p27 mutant lacking stable interaction with Cdks (p27CK–) was intro-duced into p27 null mice, the p27CK– mutant localized to both nucleus and cytoplasm and these animals showed an expansion of tissue progenitor cells and developed tumours in multiple organs118. Thus, cytoplasmic p27 may have oncogenic function(s) — independent of its ability to regulate Cdks — that influence cell migration through effects on RHOA and stathmin.
Data increasingly indicate that p27 integrates mecha-nisms regulating both cell proliferation and migration. In addition to its positive regulation of Cdks by assem-bly and import of D-type cyclin–Cdks, the potential for cytoplasmic p27 to augment cell motility, cell migration, invasion and metastasis may explain why p27 is rarely entirely lost in human malignancies.
Prognostic studies of p27 in human cancersAlthough reduced p27 has been observed in cancers of the upper gastrointestinal tract, skin and in haemato-poietic malignancies, glioma and sarcoma9, the fol-lowing review is limited to multivariate analyses of p27 in human epithelial malignancies that account for the greatest mortality in humans (fIG. 4).
Loss of p27 occurs early in tumour progression. Quiescent normal epithelia of breast, prostate, ovary, lung and other sites express high levels of nuclear p27. In colonic epithelia, nuclear p27 levels are reduced in proliferating crypt cells, and the basal proliferative cells of the oral epidermis show reduced p27 whereas terminally differ-entiated cells in more superficial cell layers express high levels of p27 (Ref. 119). p27 is reduced in premalignant and non-invasive cancerous lesions, including ductal carcinoma in situ (DCIS) of the breast92,120–122. In one study, high levels of p27 were observed in 85% of cells in atypical ductal hyperplasia, but only in 40% of DCIS and 34% of invasive breast cancer cells compared with over 95% of normal duct epithelial cells123. Reduced p27 has been observed in oral dysplasia119,124 and is prog-nostic for subsequent development of oral squamous carcinoma (P = 0.02)124. ‘Normal’ oral mucosa adjacent to tumour tissue can also show reduced p27 (Ref. 125). p27 is reduced in benign prostatic hypertrophy126–128, a hyperplastic premalignant prostatic neoplasm, and p27 levels in ovarian tumours of low malignant potential are reduced compared with normal epithelia but higher than those in aggressive ovarian cancers129,130. The utility of p27 immunohistochemistry in serial biopsies to predict
Figure 4 | p27expressionlevelsandcancer.A majority of human malignancies in many organ sites ( see figure), show reduced p27 protein levels.
R E V I E W S
258 | APRIL 2008 | VOLUME 8 www.nature.com/reviews/cancer
© 2008 Nature Publishing Group

Multivariate analysisThe observation and analysis of more than one statistical variable at a time.
Univariate analysisThe observation and analysis of one statistical variable at a time.
results of chemoprevention trials in oral dysplasia and benign prostatic hypertrophy warrants investigation. A rise in p27 may be an early indicator of efficacy of the intervention and reversion to normal differentiation.
Lung cancer. Multivariate analysis of p27 in non-small cell lung cancers (NSCLC) include seven small (n = 62–149) retrospective studies (Supplementary information S1 (table)), all of which showed significance on univariate
analysis131–137. Five show that reduced p27 levels are of independent prognostic significance with 1.82–5.94 fold increased risk of disease recurrence or death131,132,134,136,137. The two studies that failed to show multivariate signifi-cance showed a trend to poor outcome with reduced p27 (Refs 133,135). Although Ras overexpression reduces p27 levels in fibroblasts7, reduced p27 did not correlate statis-tically with Ras mutation138 or overexpression133, but did correlate with reduced cancer cell differentiation (high tumour grade) in two studies135,138, and with increased proliferation (indicated by Ki67 staining138; TABLe 1). Protein extracts from primary lung cancers with low p27 levels show high p27 proteolytic activity139.
EGFR is overexpressed in up to 62% of NSCLC140, and activating EGFR mutations have been shown to predict response to EGFR blockade by gefitinib141,142. EGFR activation leads to p27 proteolysis in cell lines71; thus therapeutic EGFR blockade should increase p27 in treated cancers. Whether reduced tumour p27 levels at diagnosis or a post-treatment increase in p27 predicts response to pure EGFR blockers such as gefitinib and tipifarnib in NSCLC warrants further investigation.
Head and neck cancer. Nine out of thirteen small, retro-spective multivariate studies in head and neck cancer show reduced p27 is associated with adverse outcome125,143–154 (Supplementary information S1 (table)). In contrast to other cancers, none of the ten head and neck cancer stud-ies shows a correlation between p27 levels and the degree of tumour differentiation or grade (TABLe 1). Reduced p27 correlates strongly with advanced disease stage in eight out of ten studies and with increased tumour size. Because head and neck cancers often become chemo- and radia-tion resistant, the use of p27 as a predictor of response to chemoprevention for dysplastic lesions warrants further investigation.
Colorectal cancer. Nine out of thirteen retrospective mul-tivariate analyses of colorectal cancers (n = 80–418) show reduced p27 is associated with a 1.43–11 fold increase in relative risk of disease recurrence or death82,155–166 (TABLe
2). Three of the four remaining studies showed a trend towards significance between low p27 protein levels and poor disease outcome161,164,165. Old tumour blocks might have reduced p27 staining quality164. Sarli et al. found that changes in p27 protein levels are most prognostic in patients with stage II disease166, whereas reduced p27 was most significant in stage III cancers in another study162.
Most colorectal cancer studies scored p27 based on the percentage of tumour nuclei that stained posi-tive for p27. Some, however, scored both p27 staining intensity and the percentage of positive nuclei162,165, with varying cut-off values used to define low p27 protein levels. Other studies reported on both cytoplasmic and nuclear staining. Cytoplasmic p27 staining, observed in 19–35% of colorectal cancers162,165,167, correlated with high nuclear p27 scores165,167 and with improved outcome in one study165. A correlation between reduced nuclear p27 and increased proliferation observed in advanced stage, metastatic colon cancer168 was not confirmed in earlier stage disease157,160,167. Reduced nuclear p27 did
Table 1 | Correlations between p27 and other prognostic factors
n LnM Stage Grade Prolif.* Size refsLung149 0.892 0.805 – – – 131133 NS 0.087 0.008 – 0.0423 13869 – 0.97 – – – 132207 1.0 0.7 0.7 0.7 – 13398 NS – 0.0005 – – 135136 NS 0.0225 NS – – 13462 – – – 0.004 – 137Head and neck109 0.0076 0.0008 – – 0.0023 144132 0.318 0.018 0.298 – – 125102 0.036 0.0003 0.35 <0.0001 <0.0001 151106 0.24 0.169 0.182 0.961 – 14977 0.51 0.031 0.34 – – 143128 0.21 – 0.61 – – 14581 <0.05 <0.05 NS – – 150137 NS – NS – – 154
94 0.02 0.02 NS – – 14660 0.46 0.027 – – 0.06 147Colorectal40 0.2859 0.2698 0.0007 NS 0.0292 167124 – 0.021 0.02 – – 156
66 – – – 0.0001 – 16889 – 0.19 0.13 0.123 – 157171 0.0012 0.0005 NS – NS 15880 0.13 – 0.77 0.47 0.17 16067‡ – 0.473 0.008 – 0.967 162 49§ – 0.044 0.24 – 0.393 162 104 0.008 0.001 – – – 161 168 – NS 0.022 – 0.876 16480 – 0.014 0.05 – – 82108 – NS <0.01 – – 166418 – NS NS – – 165Ovarian80 – <0.001 <0.001 <0.001 – 130165 – NS NS – – 18199 – NS NS NS – 18066 – NS NS – – 178103 0.002 – 0.003 – – 129185 – 0.543 0.73 – – 179150 – 0.008 0.27 – – 113
*Prolif., proliferation; indicated by Ki67 staining. ‡Stage 2. §Stage 3. LNM, lymph node metatstases; n, number of tumours; NS, not significant; ‘–’, not reported.
R E V I E W S
NATURE REVIEWS | cancer VOLUME 8 | APRIL 2008 | 259
© 2008 Nature Publishing Group

not correlate with increased tumour stage or size, or with the presence of lymph node metastases (TABLe 1). Although several different antibodies have been used, a number of studies that failed to show a statistical correlation between reduced nuclear p27 and outcome all used the Dako SX53G8 anti-p27 antibody161,163,164,166,169.
Prostate cancer. Seven out of nine multivariate analy-ses, involving 1,464 patients, show reduced nuclear p27 is an independent predictor of decreased time from prostatectomy to biochemical disease recurrence (increasing prostate specific antigen (PSA) levels), with a hazard ratio (HR) for low p27 levels of between two to
Table 2 | p27 as a prognostic factor
n %lowp27
Score cut-off(%)
DFS(univariate)*
OS(univariate)* DFS(multivariate)‡
OS(multivariate)‡ MedianFU(yrs)
refs
Colorectal149 10 0, <50%, >50% 0 ND P = 0.0014 ND 2.9; P = 0.003 ND 155124 14 0, <50, >50% 0 P<0.01 P<0.01 2.21 (1.30–3.77);
P = 0.0032.53 (1.32–4.86); P = 0.005
4.6 156
89 33 <24, 25–49, 50–74, >75%
<50 ND P = 0.0069 ND 2.38 (1.22–4.76); P = 0.01
ND 157
171 49 <46.8, >46.8% <46.8 median
P = 0.0025 P<0.0005 ND 2.38 (1.18–4.54); P = 0.0146
5.6 158
156 67 0, <25 ,25– 50, >50%
<25 P = 0.06 P = 0.02 2 (0.9–5); P = 0.08 3.33 (1.25–10); P = 0.01
3.9 159
80 46 <30, >30% <30 ND P = 0.0003 ND 4.032 (1.32–12.35); P = 0.015
ND 160
86 54 <20%, >20% <20 P = 0.006 26.555 P = 0.001 (DSS)
20.85 (18.8–22.9); P = 0.001
11.16 (9.10–13.22); P = 0.001 (DSS)
3 161
206§ 30 Intensity and <50, >50%
<50; SI<0.7
ND P = 0.152 ND 1.428 (1.00–2.5); P = 0.058
4.7 162
49|| 30 Intensity and <50, >50%
<50; SI<0.7
ND P = 0.014 ND 3.2 (1.3–7.7); P = 0.01
4.7 162
103 27 <10, >10% <10 P = 0.14 P = 0.6 ND ND 5 163168 46 <50, >50% <50 ND P = 0.68 ND 1.15 (0.77–1.71);
P = 0.48883.9 164
80 55 <50%, >50% <50 ND 4.81 (2.28–10.10); P = 0.001
ND 3.70 (1.49–8.85); P = 0.005
ND 82
418¶ 48 <10,10-20,>20% <10 ND P = 0.1815 (DSS) ND ND ND 165418# 81 <10, mod>10,
intense>10%<10 ND P = 0.0359 (DSS) ND 1.46 (0.99–2.18);
P = 0.059 (DSS)ND 165
108 31 <50, >50% <50 P = 0.072 P = 0.069 ND ND 4.5 166Prostate113 20 <25, 25–50,
50–75, >75%<25 P = 0.0037 ND 2.08 (1.01–4.27);
P = 0.047ND ND 128
86§ 16 <30, >30% <30 2.20 (0.91–5.36); P = 0.082
ND 3.04 (1.21–7.64); P = 0.0184
ND 3.3 170
67** ND ND <30 ND ND 4.99 (1.52–16.39); P = 0.0081
ND ND 170
96 13 <10, 11–50, >50%
<10, 11–50, >50
P = 0.004 P = 0.01 ND P = 0.04 9.5 171
138 28 <50, >50% <50 P = 0.3 ND NS ND 5 (mean) 17592 38 <50, >50% <50 P<0.0001 P<0.01 (DSS) 4.14 (2.00–8.54);
P<0.0013.26 (1.02–10.35); P = 0.045 (DSS)
9.4 126
86 23 <10, >10% <10 P = 0.0006 P = 0.046 P = 0.0012 P = 0.051 5.9 174161 ND <45, >45% <45 2.41 (1.23–4.7);
P = 0.01ND 2.44 (1.2–4.96);
P = 0.014ND 3.5 172
76 66 <45, >45% <45 4.40 (1.28–15.13); P = 0.019
ND 5.15 (1.41–18.83); P = 0.013
ND 3.6 177
162 69 <40, >40% <40 P = 0.499 ND 1.116 (0.651–1.914); P = 0.690
ND 5.5 87
463 ND SI 0,1,2,3,4 <2 or >3 vs 2–3
2.27 (1.40–3.68); P = 0.0009
ND 2.28 (1.32–3.92); P = 0.0029
ND 7 173
*The univariate and ‡multivariate analyses show hazard ratio (HR); 95% confidence interval (CI) (where calculated) and P value. §All tumours. ||Stage 3 only. ¶Nuclear p27 scored. #Cytoplasmic p27 scored. **T2–3 tumours analysed. DFS, disease free survival; DSS, disease specific survival; FU, follow-up; int, intermediate; mod, moderate; n, number of tumours; ND, no data; NS, not significant; OS, overall survival; SI, scoring index.
R E V I E W S
260 | APRIL 2008 | VOLUME 8 www.nature.com/reviews/cancer
© 2008 Nature Publishing Group

Ki67A monoclonal antibody that marks the late s phase of the cell cycle. This is frequently used to mark proliferating cells in tissues or cell suspensions.
Gleason grading systemThe ‘gold standard’ for grading prostate cancer, used by pathologists worldwide. This system involves assessing both the predominant and secondary pattern of gland formation within a prostate sample. The sample is scored to create a Gleason ‘sum’, ranging from 2 to 10, with the highest number indicating the most aggressive cancer. Patients with a Gleason sum of less than 6 typically respond well to therapy, whereas patients with a Gleason sum greater than 7 usually have poor outcomes.
five126,128,170–174 (TABLe 2). Two studies that failed to show independent prognostic significance for p27 used a higher percentage (40 and 50%, respectively) of p27 positive nuclei as cut offs87,175.
Despite the considerable heterogeneity within pros-tate tumours, Thomas et al. found that p27 scores in areas of highest Gleason grade (poorest differentiation) in pre-operative core biopsies correlate well with p27 levels in resected tumours in 44 matched specimens176.
The prognostic value of p27 may be greatest in organ-confined disease. The HR increased from 2.4 in a cohort of 161 patients with prostate cancer, to 5.15 in patients with organ confined disease (n = 76)177. One retrospec-tive study (TABLe 2) (n = 463) found cytoplasmic p27 was prognostic for poor disease free survival (DFS) (HR 2.18 (1.36–3.49), P = 0.0012)173. This study used image analysis and a digitalized scoring system that may not be readily reproducible in community hospital settings. Reduced nuclear p27 correlated strongly with a high Gleason score in most studies (TABLe 3).
Ovarian cancer. All of six relatively small retrospec-tive multivariate analyses, including 757 cancer sam-ples, show reduced p27 to be prognostic on univariate analysis and of these, four show independent prog-nostic value for p27, with HR for mortality of 2.3–3.7 (Refs 113,129,130,178–181) (Supplementary information S1 (table)). In one study, although p27 was not prognos-tic in the entire cohort, it was significant in advanced stages III and IV130 (TABLe 1). Other groups show reduced p27 predicts poor outcome after chemotherapy130,180.
Breast cancer. p27 has been most extensively studied in breast cancer, and 11 out of 17 multivariate analy-ses show reduced p27 is of independent prognostic significance66,92–94,182–193 (C. Catzavelos and J.M.S, per-sonal communication). Most studies that failed to show independent prognostic value were highly significant on univariate analysis66,182–186. In total, almost five thou-sand cancer samples analysed with 5–17 years median follow-up, indicate that reduced tumour p27 increases risk of relapse or death by 1.3–4 fold (TABLe 4). In a ret-rospective analysis of p27 in 2,031 cancer samples from a randomized clinical trial, reduced p27 was an independ-ent prognostic factor for reduced overall survival (HR 1.31, confidence interval (CI) 1.05–1.64)187. Another
study of 1,015 prospectively accrued node-negative breast cancers showed reduced p27 to be independently prognostic, (HR 1.53, CI 1.05–2.23) (C. Catzavelos and J.M.S, personal communication). In 500 patients with premenopausal hormone receptor positive breast cancer, low p27 correlated strongly with poor survival (HR 3.45, CI 1.72–6.67)192.
Reduced p27 was usually correlated statistically with increased proliferation, high tumour grade and with oes-trogen receptor (ER) and progesterone receptor (PR) negative status (TABLe 5). Lobular carcinomas tend to have higher p27 (Refs 182,194). Reduced p27 correlates with lower cyclin D1 (Refs 182,183) and with ERBB2 amplification66,67 (C. Catzavelos and J.M.S, personal communication), and may have higher prognostic value in earlier stage disease. W. Foulkes et al., reported an independent HR 1.96 for low p27 in 230 cancers that increased to HR 4.76 in the earlier stage, node negative subgroup (P = 0.03 in both)193.
p27 as a predictor of treatment responseAlthough several factors, such as tumour differentiation, or grade, size and stage, are prognostic for poor patient outcome, few factors specifically predict response to particular anticancer treatments. Well known predictive factors in breast cancer include amplification of ERBB2, which predicts response to trastuzumab 195 and ER which predicts response to the antioestrogen tamoxifen196. Factors that predict response to therapy are arguably more important than prognostic factors because they influence treatment decisions. However, further stud-ies are required to determine if changes in p27 levels, localization or other modifications can predict treatment responses. For example, a study of radiation after sur-gery for large, aggressive oesophageal cancers showed reduced p27 correlated with improved outcome143. This may reflect activation of p27 proteolysis in rapidly grow-ing cancers that have increased radiation-induced cell death. By contrast, following radiation used with curative intent in early cervical197 or laryngeal153 cancers, high p27 levels predicted improved outcome. In these early stage tumours, which express high levels of p27, the reduced metastatic potential associated with high p27 might outweigh the potential influence of proliferation rate on the immediate outcome of radiotherapy. The potential predictive role of p27 for radiation responsiveness can vary significantly among tumour types and warrants further study in large randomized prospective trials.
So far, only one large prospective randomized trial in colon cancer unequivocally indicates that p27 predicts response to specific chemotherapeutic agents (BOX 1). Further analysis of p27 in large prospective randomized trials will be required to identify the predictive value of p27 for chemotherapy responses in different contexts. For early stage disease, high p27 may continue to predict improved overall survival. For cell cycle active agents in more advanced highly proliferative cancers, reduced p27 might predict response to chemotherapy but ultimately, patient demise. Our ability to identify cells within the cancer population that have regenerative potential may inform the design of future prognostic and predictive
Table 3 | Correlations of p27 with other prognostic factors in prostate cancer
n LnM Stage Grade SVI PSa refs113 NS – 0.032 0.0002 0.054 12886 – 0.818 0.041 – 0.319 17096 – 0.32 0.004 – 0.36 171138 0.03 – 0.002 0.007 0.2 17544 – 0.028 0.0001 – 0.69 17692 0.02 0.02 0.01 – – 126
86 0.21 0.3 0.8 – 0.9 174162 – 0.5 0.58 – 0.57 87640 – – – – 0.0087 173LNM, lymph node metatstases; NS, not significant; PSA, prostate-specific antigen; SVI, seminal vesicle involvement; P values are shown in the table.
R E V I E W S
NATURE REVIEWS | cancer VOLUME 8 | APRIL 2008 | 261
© 2008 Nature Publishing Group

studies. There is early evidence that chemotherapy can promote expansion of human tumour stem cells in vivo207. The predictive potential of p27 may be most clear when analysis is restricted to this tumour cell fraction and with dynamic analysis of samples before and after treatment.
Hormonal therapy for breast cancer. Two studies of p27 in randomized trial populations suggest that high p27 may predict response to hormonal therapy. Porter et al. analysed 2,031 patients from the South Western Oncology Group (SWOG) S9313 trial who were rand-omized to receive adjuvant doxycycline and cyclophos-phamide, with adjuvant tamoxifen given for hormone receptor positive cancers. They found that p27 was a stronger independent prognostic factor in hormone receptor positive tumours (HR 1.42 (1.05–1.94)) than
in the entire group (HR 1.31 (1.05–1.94))187. In another trial that randomized 512 premenopausal women with hormone receptor positive cancers to receive either tamoxifen with ovarian suppression or chemotherapy with no subsequent hormonal therapy, reduced p27 was most strongly predictive of failure of adjuvant hormonal therapy (HR 6.25 (1.19–20), P = 0.003)192.
A retrospective analysis of p27 in over 600 hor-mone receptor positive, node negative breast cancers showed high p27 predicted improved outcome follow-ing adjuvant treatment with tamoxifen alone, (HR 2.23 (1.09–4.56) P = 0.03), but p27 levels did not discriminate outcome in untreated hormone receptor positive disease (C. Catzavelos and J.M.S, personal communication). Taken together, data from these three studies support fur-ther investigation of the predictive role of p27 for response to tamoxifen and other antioestrogen treatments.
Table 4 | p27 as a prognostic factor in breast cancer
n %lowp27
Score cut-off(%)
DFS(univariate)*
OS(univariate)*
DFS(multivariate)‡
OS(multivariate)‡
MedianFU(yrs)
refs
168 56 <25, 25–49, 50–75, >75%
<50 2.26 P = 0.0072 P = 0.05 2.13 (1.14–3.98); P = 0.0170
ND 6 92
246 63 Low/int/hi <Int/hi ND 2.9 (1.7–4.9); P<0.001
ND 2.7 (1.3–6.0); P = 0.01
5.2 93
202 50 <50, >50% <50 ND P = 0.00233 ND 3.40 (1.12–10.31); P = 0.0306
5.5 94
189 56 1–3 intensity, 1–4 % nuclei
score ≤5 P = 0.0025 1.99 (1.34–2.95); P = 0.0005
ND 1.38 (0.89–2.14); P = 0.144
16.7 183
181 69 <50, >50% <50 P = 0.0001 P = 0.0012 2.44 (1.92–3.13); P = 0.042
2.71 (2.22–3.97); P = 0.049
5 188
102 41 <50, >50% <50 P = 0.0015 P = 0.0124 ND 2.97 P = 0.0218 ND 189
202 63 <50, >50% <50 5.9 (2.1–16.7); P = 0.0008
P = 0.001 3.9 (1.4–11.1); P = 0.01
6.4 P = 0.01 6.4 190
512 66 <50, >50% <50 P = 0.41 ND NS ND 9.6 184216 50 Range
0–93.6%<62.4 median
ND P = 0.0012 ND 4.06 (1.04–11.92); P<0.05
4.7 191
197 58 <50,>50% <50 P = 0.017 (5yr); P = 0.268 (17yr)
P = 0.013 (5yr); P = 0.67 (17yr)
ND NS 17 185
512 19 <50, >50% <50 1.72 (1.12–2.63); P = 0.01
2.86 (1.45–8.05); P = 0.002
1.89 (1.22–2.94); P = 0.004
3.45 (1.72–6.67); P<0.001
5.5 192
198 (node –)
43 <50, >50% <50 1.21 (0.83–1.78); P = 0.32
1.33 (0.84–2.11); P = 0.23
NS NS 13 66
263 (node +)
44 <50, >50% <50 0.99 (0.74–1.32); P = 0.92
0.96 (0.69–1.32); P = 0.78
ND ND 13 66
830 ND H-score 0,1,2,3; range 0–300
H-score <200
P = 0.45 P = 0.0002 ND P = 0.778 8.7 182
175 50 <10, 10–50, >50%
<50 ND P = 0.045 ND 1.57 (0.98–3.27); P = 0.073
4.9 186
230 61 ND ND ND 2.32 (1.28–4.35); P = 0.006
ND 1.96 (1.05–3.57); P = 0.03
9.3 193
2,031 46 SI 1–7 <median 1.31(1.1–1.57); P = 0.0024
1.5 (1.21–1.86); P = 0.0002
1.18 (0.98–1.42) 1.31 (1.05–1.64) 7 187
1,015 41 <25, 25–49, 50–75, >75%
<25 2.27 (1.6–3.23); P = 0.0001
ND 1.53 (1.05–2.23); P = 0.03
ND 9 C. Catzavelos§
*The univariate and ‡multivariate analyses show hazard ratio (HR); 95% confidence interval (CI) (where calculated) and P value. §C. Catzavelos et al., personal communication. DFS, disease free survival; DSS, disease specific survival; FU, follow-up; hi, high; int, intermediate; n, number of tumours; ND, no data; NS, not significant; OS, overall survival; SI, scoring index.
R E V I E W S
262 | APRIL 2008 | VOLUME 8 www.nature.com/reviews/cancer
© 2008 Nature Publishing Group

p27 is required for G1 arrest by tamoxifen or oes-trogen deprivation in ER positive breast cancer lines198. In breast cell lines, SRC phosphorylates p27 to reduce its inhibitory function and ERBB2 (Refs 69,199), SRC49 and MAP2K (MEK)68 overexpression all accelerate p27 proteolysis. In ER positive, tamoxifen-resistant lines, the EGFR and ERBB2 inhibitor lapatinib, and SRC and MAP2K (MEK) inhibitors can all restore the antipro-liferative effects of tamoxifen49,68,72. Reduced p27 levels were strongly correlated with activated SRC in primary
breast cancers49. Preclinical investigations and early trials are underway to test the potential for SRC inhibi-tors to cooperate with antioestrogens in treatment of ER and/or PR positive breast cancer.
Predicting response to targeted therapiesAs noted earlier, RTK activation can accelerate p27 pro-teolysis through SRC and can also impair p27 function through PI3K activation of Akt, p90, SGK and mTOR. Several clinical trials have shown the efficacy of antibodies or small molecule inhibitors of EGFR family members in lung, colon and breast cancers65. mTOR inhibitors have shown promise in a number of human tumours. Reduced levels and/or mislocalization of p27 in pre-treatment biopsies may indicate the potential to respond to these treatments and an increase in p27 levels or a shift from cytoplasmic to nuclear localization on post-treatment biopsy may predict response to these novel targeted thera-pies. Furthermore, tyrosine phosphorylation of p27 may correlate with responsiveness to tyrosine kinase inhibitors. As we come to understand the complexity of molecular pathways regulating p27, it will be possible to design com-bination therapies with the potential to restore p27 stability and function to induce growth arrest in human cancers.
Technical hurdles for clinical applicationThere are few reliable molecular prognostic factors and even fewer markers that have predictive value for treat-ment response in human cancers200. For broad clinical application a prognostic factor must show reproduc-ible independent prognostic significance, be relatively inexpensive and be technically easy to use in commu-nity hospitals. Although there are many retrospective studies of p27, further validation of this marker in the context of prospective clinical trials with uniform treat-ment conditions is required. Although many technical challenges need to be resolved before p27 could be
Table 5 | Correlation of p27 levels with other prognostic factors in breast cancer
n LnM Stage Grade Proliferation Size er Pr erBB2 refs168 0.060 – 0.001 NS – 0.001 0.001 – 9268 0.0001 0.001 0.028 0.133 0.194 0.349 0.667 – 186189 NS – <0.0001 0.0004 0.016 <0.0001 – – 18377 – – p<0.05 – – <0.01 <0.01 – 195181 0.0001 – 0.002 – – 0.171 – – 188102 0.685 – – – 0.121 0.006 – – 189202 0.24 – <0.0001 – 0.002 <0.0001 – – 190197 0.1 0.0008 0.004 0.02 0.04 0.001 0.02 – 185216 NS NS – NS NS <0.0001 – – 191198 – – 0.001 0.77 1 0.09 0.25 0.04 66 (node –)263 – – 0.01 0.007 1 0.22 0.22 0.27 66 (node+)830 – 0.571 <0.001 <0.001 0.005 <0.001 – – 182512 0.5 – 0.15 – 0.4 – – – 192175 – – – 0.028 – – – – 18682 >0.1 – <0.01 – >0.5 >0.05 – – 206
2,031 – – – – – <0.001 <0.001 – 1871,015 – – 0.001 – 0.001 0.001 0.001 0.03 C. Catzavelos**C. Catzavelos et al., personal communication. ER, oestrogen receptor; LNM, lymph node metastasis; n, number of tumours; PR, progesterone receptor NS, not significant; ‘–’, not reported. Numbers in the table are P values.
Box 1 | Predictive potential of p27 for response to chemotherapy
In retrospective analyses, reduced p27 (also known as KIP1) correlated with reduced survival after platinum based chemotherapy for non-small cell lung cancer202 and ovarian cancers130,180. Reduced p27 also correlated with poor outcome after adjuvant chemotherapy for breast cancer123. By contrast, Psyrri et al. reported that in advanced stage III and IV ovarian cancers, reduced p27 predicted an improved response to adjuvant paclitaxel/platinum after debulking surgery, hazard ratio 3.22 (1.7–6.22) P<0.001 (Ref. 113). This may reflect the efficacy of cycle active chemotherapy for tumours with high proliferation and low p27.
A number of analyses stratified chemo-treated patients according to low and high p27 scores and assessed outcome. Benefit from chemotherapy was observed only in cancers with low p27. A relatively small colorectal study showed benefit from adjuvant 5 fluorouracil (5FU) prodrug was limited to cancers with reduced p27 scores203. By contrast, analysis of 500 stage III colorectal cancers from the prospective randomized CALGB trial of adjuvant 5FU plus leucovorin versus 5FU, leucovorin plus irinotecan (Saltz regimen) showed reduced p27 (scores of <30%) predicted longer time to recurrence after both adjuvant chemotherapy regimens, but correlated with poor patient survival (M. Loda, M. Bertagnolli, R. Schilsky, personal communication).
In one retrospective breast cancer study, benefit from adjuvant chemotherapy was restricted to tumours with low p27. With high p27, survival was similar in chemo-treated and untreated groups188. In the prospective International Breast Cancer Surgery Group trial V of 461 early stage breast cancers randomized to no adjuvant therapy, one or six cycles of adjuvant CMF (cyclophosphamide, methotrexate and fluorouracil), benefit from chemotherapy was most notable for cancers with low p27 levels, with a trend to greater significance in node negative versus positive patients66.
R E V I E W S
NATURE REVIEWS | cancer VOLUME 8 | APRIL 2008 | 263
© 2008 Nature Publishing Group

1. Sherr, C. J. & Roberts, J. M. CDK inhibitors: positive and negative regulators of G1-phase progression. Genes Dev. 13, 1501–1512 (1999).
2. Hengst, L. & Reed, S. I. Inhibitors of the Cip/Kip family. Curr. Top. Microbiol. Immunol. 227, 25–41 (1998)
3. Nigg, E. A. Targets of cyclin-dependent protein kinases. Curr. Opin. Cell Biol. 199, 187–193 (1993).
4. James, M., Ray, A., Leznova, D. & Blain, S. W. Differential modification of p27Kip1 controls its cyclin D–cdk4 inhibitory activity. Mol. Cell Biol.(2007).Showed that p27 in active cyclin D1–CDK4 complexes is phosphorylated on tyrosine whereas p27 in catalytically inactive cyclin D1–CDK4 is not.
5. Grimmler, M. et al. Cdk-inhibitory activity and stability of p27Kip1 are directly regulated by oncogenic tyrosine kinases. Cell 128, 269–280 (2007).
6. Hengst, L. & Reed, S. I. Translational control of p27Kip1 accumulation during the cell cycle. Science 271, 1861–1864 (1996).This paper demonstrated that CDKN1B transcript levels vary little, but translation is maximal in quiescence and falls during G1 progression.
7. Liang, J. & Slingerland, J. M. Multiple roles of the PI3K/PKB (Akt) pathway in cell cycle progression. Cell Cycle 2, 339–345 (2003).
8. Besson, A., Assoian, R. K. & Roberts, J. M. Regulation of the cytoskeleton: an oncogenic function for CDK inhibitors? Nature Rev. Cancer 4, 948–955 (2004).An important review of primary work by these authors and others showing that cytoplasmic p27 has effects on cell motility that are, at least in part, independent of its Cdk regulatory functions.
9. Slingerland, J. & Pagano, M. Regulation of the cdk inhibitor p27 and its deregulation in cancer. J. Cell Physiol. 183, 10–17 (2000).
10. Medema, R. H., Kops, G. J., Bos, J. L. & Burgering, B. M. AFX-like Forkhead transcription factors mediate cell-cycle regulation by Ras and PKB through p27kip1. Nature 404, 782–787 (2000).
11. Dijkers, P. F. et al. Forkhead transcription factor FKHR-L1 modulates cytokine-dependent transcriptional
regulation of p27KIP1. Mol. Cell. Biol. 20, 9138–9148 (2000).References 10 and 11 were the first to show that p27 transcription is opposed by Akt action on forkhead transcription factors.
12. Trotman, L. C. et al. Identification of a tumour suppressor network opposing nuclear Akt function. Nature 441, 523–527 (2006).
13. Yang, H. L., Zhao, R. Y., Yang, H. Y. & Lee, M. H. Constitutively active FOXO4 inhibits Akt activity, regulates p27 Kip1 stability, and suppresses HER2-mediated tumorigenicity. Oncogene 24, 1924–1935 (2005).
14. Wang, X. H. et al. Increased hepatic forkhead box M1B (FoxM1B) levels in old-aged mice stimulated liver regeneration through diminished p27(KiP1) protein levels and increased Cdc25B expression. J. Biol. Chem. 277, 44310–44316 (2002).
15. Karnik, S. K. et al. Menin regulates pancreatic islet growth by promoting histone methylation and expression of genes encoding p27(Kip1) and p18(INK4c). Proc. Natl Acad. Sci. USA 102, 14659–14664 (2005).
16. Fontaniere, S., Casse, H., Bertolino, P. & Zhang, C. X. Analysis of p27Kip1 expression in insulinomas developed in pancreatic β-cell specific Men1 mutant mice. Familial Cancer 5, 49–54 (2006).
17. Pellegata, N. S. et al. Germ-line mutations in p27Kip1 cause a multiple endocrine neoplasia syndrome in rats and humans. Proc. Natl Acad. Sci. USA 103, 15558–15563 (2006).
18. Georgitsi, M. et al. Mutation analysis of aryl hydrocarbon receptor interacting protein (AIP) gene in colorectal, breast, and prostate cancers. Br. J. Cancer 96, 352–356 (2007).
19. Yang, W. et al. Repression of transcription of the p27Kip1 cyclin-dependent kinase inhibitor gene by c-Myc. Oncogene 20, 1688–1702 (2001).
20. O’Hagan, R. C. et al. Myc-enhanced expression of Cul1 promotes ubiquitin-dependent proteolysis and cell cycle progression. Genes Dev. 14, 2185–2191 (2000).
21. Keller, U. B. et al. Myc targets Cks1 to provoke the suppression of p27Kip1, proliferation and lymphomagenesis. EMBO J. 26, 2562–2574 (2007).
22. Wang, C. G. et al. Activation of p27Kip1 expression by E2F1 — a negative feedback mechanism. J. Biol. Chem. 280, 12339–12343 (2005).
23. Inoue, T., Kamiyama, J. & Sakai, T. Sp1 and NF-Y synergistically mediate the effect of vitamin D-3 in the p27Kip1 gene promoter that lacks vitamin D response elements. J. Biol. Chem. 274, 32309–32317 (1999).
24. Murata, K. et al. Hes1 directly controls cell proliferation through the transcriptional repression of p27Kip1. Mol. Cell. Biol. 25, 4262–4271 (2005).
25. Huang, Y. C., Chen, J. Y. & Hung., W. C. Vitamin D-3 receptor/Sp1 complex is required for the induction of p27Kip1 expression by vitamin D-3. Oncogene 23, 4856–4861 (2004).
26. Agrawal, D. et al. Repression of p27kip1 synthesis by platelet-derived growth factor in BALB/c 3T3 cells. Mol. Cell Biol. 16, 4327–4336 (1996).
27. Millard, S. S. et al. Enhanced ribosomal association of p27Kip1 mRNA is a mechanism contributing to accumulation during growth arrest. J. Biol. Chem. 272, 7093–7098 (1997).
28. Gopfert, U., Kullmann, M. & Hengst, L. Cell cycle-dependent translation of p27 involves a responsive element in its 5′-UTR that overlaps with a uORF. Hum. Mol. Genet. 12, 1767–1779 (2003).
29. Miskimins, W. K., Wang, G., Hawkinson, M. & Miskimins, R. Control of cyclin-dependent kinase inhibitor p27 expression by cap-independent translation. Mol. Cell. Biol. 21, 4960–4967 (2001).
30. Kullmann, M., Gopfert, U., Siewe, B. & Hengst, L. ELAV/Hu proteins inhibit p27 translation via an IRES element in the p27 5′ UTR. Genes Dev. 16, 3087–3099 (2002).Showed ELAV /Hu proteins bind an IRES in the 5′ UTR to impair p27 translation. Overexpression of ELAV in cancers might reduce p27 translation and contribute to development or progression of certain cancers.
broadly applied in clinical evaluation of cancers (BOX 2), similar significant hurdles were overcome in the course of integrating ER and ERBB2 assays into the routine evaluation of newly diagnosed breast cancer.
ConclusionsThere have been major advances in gene and protein expression profiling of human cancers, and techniques to assay multi-gene expression have important prog-nostic and predictive potential201. Despite the challenges of establishing a consensus for optimal immuno-histochemical staining and scoring techniques, p27 has important potential as a prognostic factor and warrants further validation in prospective randomized trials. p27 immunohistochemistry might ultimately assist in the
identification of patients who may benefit from specific molecular targeted therapies. More importantly, the study of p27 in human cancer lines and tumours has informed our understanding of key signal transduction pathways and their roles in the regulation of normal and malignant cell proliferation and opened new avenues for targeted therapeutic intervention.
Note added in proofA recent report identifies Pirh2 as a ubiquitin ligase for p27 (Ref. 208). In addition, p27 binds and inhibits casein kinase 2-α′ (CK2-α′) in differentiated cardiomyocytes, in which angiotensin II induces p27 proteolysis through CK2-α′-dependent p27 phosphorylation on Ser83 and Thr187 (Ref. 209).
Box 2 | Technical hurdles to the application of p27 immunohistochemistry in the clinic
For broad application in the clinic, quality assurance issues for standardization of staining and scoring of p27 need to be resolved. Many studies indicate that p27 can be reliably evaluated in tissue microarrays166,173,182,186,187,204. One breast-cancer study showed 96% concordance between scores from core biopsies and surgical specimens186. Monoclonal p27 antibodies from Transduction Laboratories, Neomarkers, and Novocastra have not been compared head to head in any large studies. There is a need to establish uniform p27 scoring systems and cut-off levels for low and high p27 scores for different tumour types. Whereas most prognostic analyses have scored p27 based on the percentage of tumour nuclei positive, using various cut-offs ranging from 0–50%, others score both percentage of positive cells and intensity of staining (TABLes 2,4). Most studies have not considered or scored cytoplasmic p27. In oesophageal205, breast98 and prostate cancers173, cytoplasmic p27 was associated with poor outcome, whereas in colon cancer, cytoplasmic p27 was associated with high nuclear p27 (Ref. 165) and improved outcome. A scoring system that incorporates localization, percentage of positive cells and intensity may be important. In one small breast cancer study, high nuclear p27 levels correlated with the best patient outcome, and low nuclear levels with cytoplasmic p27 identified the worst patient outcome98.
R E V I E W S
264 | APRIL 2008 | VOLUME 8 www.nature.com/reviews/cancer
© 2008 Nature Publishing Group

31. Yoon, A. et al. Impaired control of IRES-mediated translation in X-linked dyskeratosis congenita. Science 312, 902–906 (2006).
32. Cho, S. C., Kim, J. H., Back, S. H. & Jang, S. K. Polypyrimidine tract-binding protein enhances the internal ribosomal entry site-dependent translation of p27Kip1 mRNA and modulates transition from G1 to S phase. Mol. Cell. Biol. 25, 1283–1297 (2005).
33. Millard, S. S., Vidal, A., Markus, M. & Koff, A. A U-rich element in the 5′ untranslated region is necessary for the translation of p27 mRNA. Mol. Cell Biol. 20, 5947–5959 (2000).
34. Erkinheimo, T. L. et al. Cytoplasmic HuR expression correlates with poor outcome and with cyclooxygenase 2 expression in serous ovarian carcinoma. Cancer Res. 63, 7591–7594 (2003).
35. Denkert, C. et al. Overexpression of the embryonic-lethal abnormal vision-like protein HuR in ovarian carcinoma is a prognostic factor and is associated with increased cyclooxygenase 2 expression. Cancer Res. 64, 189–195 (2004).
36. de Silanes, I. L. et al. Role of the RNA-binding protein HuR in colon carcinogenesis. Oncogene 22, 7146–7154 (2003).
37. Vidal, A., Millard, S. S., Miller, J. P. & Koff, A. Rho activity can alter the translation of p27 mRNA and is important for RasV12-induced transformation in a manner dependent on p27 status. J. Biol. Chem. 277, 16433–16440 (2002).
38. Gonzalez, T. et al. Inhibition of Cdk4 activity enhances translation of p27kip1 in quiescent Rb-negative cells. J. Biol. Chem. 278, 12688–12695 (2003).
39. le Sage, C., Nagel, R. & Agami, R. Diverse ways to control p27Kip1 function: miRNAs come into play. Cell Cycle 6, 2742–2749 (2007).Important recent review of work by these authors and others showing that p27 translation is reduced by miRNA-221/222 and might contribute to p27 loss in human cancer.
40. Kedde, M. et al. RNA-binding protein Dnd1 inhibits microRNA access to target mRNA. Cell 131, 1273–1286 (2007).
41. Galardi, S. et al. miR-221 and miR-222 expression affects the proliferation potential of human prostate carcinoma cell lines by targeting p27Kip1. J. Biol. Chem. 282, 23716–23724 (2007).
42. Volinia, S. et al. A microRNA expression signature of human solid tumors defines cancer gene targets. Proc. Natl Acad. Sci. USA103, 2257–2261 (2006).
43. Visone, R. et al. MicroRNAs (miR)-221 and miR-222, both overexpressed in human thyroid papillary carcinomas, regulate p27Kip1 protein levels and cell cycle. Endocr. Relat. Cancer 14, 791–798 (2007).
44. Calin, G. A. et al. A MicroRNA signature associated with prognosis and progression in chronic lymphocytic leukemia. N. Engl. J. Med. 353, 1793–1801 (2005).References 42–44 and others within reference 38 demonstrate overexpression of miRNA-221/222 in human neoplasia.
45. Nakayama, K. I. & Nakayama, K. Ubiquitin ligases: cell-cycle control and cancer. Nature Rev. Cancer 6, 369–381 (2006).Important review of primary work by these authors and others that covers proteolytic mechanisms governing p27 degradation.
46. Bloom, J. & Pagano, M. Deregulated degradation of the cdk inhibitor p27 and malignant transformation. Semin. Cancer Biol. 13, 41–47 (2003).Important conceptual review of p27 proteolysis by the SCFSKP2 ubiquitin ligase pathway.
47. Sabile, A. et al. Regulation of p27 degradation and S-phase progression by Ro52 RING finger protein. Mol. Cell. Biol. 26, 5994–6004 (2006).
48. Pavletich, N. P. Mechanisms of cyclin-dependent kinase regulation: structures of Cdks, their cyclin activators, and Cip and INK4 inhibitors. J. Mol. Biol. 287, 821–828 (1999).
49. Chu, I. et al. p27 phosphorylation by Src regulates inhibition of cyclin E–Cdk2. Cell 128, 281–294 (2007).References 5 and 49 first showed that tyrosine phosphorylation of p27 has effects on Cdk binding and promotes SCFSKP2-mediated p27 proteolysis.
50. Russo, A. A., Jeffrey, P. D., Patten, A. K., Massague, J. & Pavletich, N. P. Crystal structure of the p27Kip1 cyclin-dependent-kinase inhibitor bound to the cyclin A-Cdk2 complex. Nature 382, 325–331 (1996).Solved the crystal structure of the N-terminal fragment of p27 bound to cyclin A–CDK2.
51. Malek et al. A mouse knock-in model exposes sequential proteolytic pathways that regulate p27Kip1 in G1 and S phase. Nature 413, 323–327 (2001)
52. Bondar, T. et al. Cu14A and DDB1 associate with Skp2 to target p27Kip1 for proteolysis involving the COP9 signalosome. Mol. Cell. Biol. 26, 2531–2539 (2006).
53. Tomoda, K., Kubota, Y. & Kato, J. Degradation of the cyclin-dependent-kinase inhibitor p27Kip1 is instigated by Jab1. Nature 398, 160–165 (1999).References 52 & 53 indicate that p27 proteolysis might be linked to COP9 signalosome function.
54. Boehm, M. et al. A growth factor-dependent nuclear kinase phosphorylates p27(Kip1) and regulates cell cycle progression. EMBO J. 21, 3390–3401 (2002).
55. Deng, X., Mercer, S. E., Shah, S., Ewton, D. Z. & Friedman, E. The cyclin-dependent kinase inhibitor p27Kip1 is stabilized in G0 by Mirk/dyrk1B kinase. J. Biol. Chem. 279, 22498–22504 (2004).
56. Rodier, G. et al. p27 cytoplasmic localization is regulated by phosphorylation on Ser10 and is not a prerequisite for its proteolysis. EMBO J. 20, 6672–6682 (2001).
57. Ishida, N. et al. Phosphorylation of p27Kip1on serine 10 is required for its binding to CRM1 and nuclear export. J. Biol. Chem. 277, 14355–14358 (2002).
58. Connor, M. K. et al. CRM1/Ran-mediated nuclear export of p27Kip1 involves a nuclear export signal and links p27 export and proteolysis. Mol. Biol. Cell 14, 201–213 (2003).References 56–58 demonstrate that p27 is exported from the nucleus in a CRM1-dependent manner and show serine 10 phosphorylation promotes nuclear export.
59. Besson, A. et al. A pathway in quiescent cells that controls p27Kip1 stability, subcellular localization, and tumor suppression. Genes Dev. 20, 47–64 (2006).This paper showed that serine 10 phosphorylation regulates p27 stability in G0.
60. Liang, J. et al. The energy sensing LKB-1–AMPK pathway regulates p27kip1 phosphorylation mediating the decision to enter autophagy or apoptosis. Nature Cell Biol. 9, 218–224 (2007).
61. Kossatz, U. et al. C-terminal phosphorylation controls the stability and function of p27kip1. EMBO J. 25, 5159–5170 (2006).
62. Kotake, Y., Nakayama, K., Ishida, N. & Nakayama, K. I. Role of serine 10 phosphorylation in p27 stabilization revealed by analysis of p27 knock-in mice harboring a serine 10 mutation. J. Biol. Chem. 280, 1095–1102 (2005).This paper showed that serine 10 phosphorylation regulates p27 stability in G0.
63. Ishizawar, R. & Parsons, S. J. c-Src and cooperating partners in human cancer. Cancer Cell 6, 209–214 (2004).
64. Pegram, M. D., Pauletti, G. & Slamon, D. J. HER-2/neu as a predictive marker of response to breast cancer therapy. Breast Cancer Res. Treat. 52, 65–77 (1998).
65. Arteaga, C. L. & Baselga, J. Clinical trial design and end points for epidermal growth factor receptor-targeted therapies: implications for drug development and practice. Clin. Cancer Res. 9, 1579–1589 (2003).
66. Spataro, V. J. et al. Decreased immunoreactivity for p27 protein in patients with early-stage breast carcinoma is correlated with HER-2/neu overexpression and with benefit from one course of perioperative chemotherapy in patients with negative lymph node status: results from International Breast Cancer Study Group Trial, V. Cancer 97, 1591–1600 (2003).
67. Newman, L. et al. Correlation of p27 protein expression with HER-2/neu expression in breast cancer. Mol. Carcinog. 30, 169–175 (2001).
68. Donovan, J. C., Milic, A. & Slingerland, J. M. Constitutive MEK/MAPK activation leads to p27Kip1 deregulation and antiestrogen resistance in human breast cancer cells. J. Biol. Chem. 276, 40888–40895 (2001).
69. Yang, H. Y., Zhou, B. P., Hung., M. C. & Lee, M. H. Oncogenic signals of HER-2/neu in regulating the stability of the cyclin-dependent kinase inhibitor p27. J. Biol. Chem. 275, 24735–24739 (2000).This paper demonstrated that p27 proteolysis is activated by overexpression of Her2.
70. Nahta, R., Takahashi, T., Ueno, N. T., Hung., M. C. & Esteva, F. J. P27kip1 down-regulation is associated with trastuzumab resistance in breast cancer cells. Cancer Res. 64, 3981–3986 (2004).
71. Busse, D. et al. Reversible G1 arrest induced by inhibition of the epidermal growth factor receptor tyrosine kinase requires up-regulation of p27KIP1 independent of MAPK activity. J. Biol. Chem. 275, 6987–6995 (2000).
72. Chu, I., Blackwell, K., Chen, S. & Slingerland, J. The dual ErbB1/ErbB2 inhibitor, lapatinib (GW572016), cooperates with tamoxifen to inhibit both cell proliferation- and estrogen-dependent gene expression in antiestrogen-resistant breast cancer. Cancer Res. 65, 18–25 (2005).
73. Moller, M. B., Skjodt, K., Mortensen, L. S. & Pedersen, N. T. Clinical significance of cyclin-dependent kinase inhibitor p27Kip1 expression and proliferation in non-Hodgkin’s lymphoma: independent prognostic value of p27Kip1. Br. J. Haematol. 105, 730–736 (1999).
74. Tsihlias, J., Kapusta, L. & Slingerland, J. The prognostic significance of altered cyclin-dependent kinase inhibitors in human cancer. Annu. Rev. Med. 50, 401–423 (1999).
75. Andreu, E. J. et al. BCR-ABL induces the expression of Skp2 through the PI3K pathway to promote p27Kip1 degradation and proliferation of chronic myelogenous leukemia cells. Cancer Res. 65, 3264–3272 (2005).
76. Donato, N. J. et al. BCR–ABL independence and LYN kinase overexpression in chronic myelogenous leukemia cells selected for resistance to STI571. Blood 101, 690–698 (2003).
77. Vivanco, I. & Sawyers, C. L. The phosphatidylinositol 3-kinase AKT pathway in human cancer. Nature Rev. Cancer 2, 489–501 (2002).
78. Tsutsui, S. et al. Inactivation of PTEN is associated with a low p27Kip1 protein expression in breast carcinoma. Cancer 104, 2048–2053 (2005).
79. Latres, E. et al. Role of the F-box protein Skp2 in lymphomagenesis. Proc. Natl Acad. Sci. USA 98, 2515–2520 (2001).First demonstration that SKP2 is overexpressed in human lymphoma and that SKP2 overexpression promotes lymphoma progression in a mouse model in vivo.
80. Chiarle, R. et al. Increased proteasome degradation of cyclin-dependent kinase inhibitor p27 is associated with a decreased overall survival in mantle cell lymphoma. Blood 95, 619–626 (2000).
81. Hershko, D. et al. Inverse relation between levels of p27Kip1 and of its ubiquitin ligase subunit Skp2 in colorectal carcinomas. Cancer 91, 1745–1751 (2001).
82. Shapira, M. et al. The prognostic impact of the ubiquitin ligase subunits Skp2 and Cks1 in colorectal carcinoma. Cancer 103, 1336–1346 (2005).
83. Signoretti, S. et al. Oncogenic role of the ubiquitin ligase subunit Skp2 in human breast cancer. J. Clin. Invest. 110, 633–641 (2002).
84. Shintani, S. et al. Skp2 and Jab1 expression are associated with inverse expression of p27KIP1 and poor prognosis in oral squamous cell carcinomas. Oncology 65, 355–362 (2003).
85. Kudo, Y. et al. High expression of S-phase kinase-interacting protein 2, human F-box protein, correlates with poor prognosis in oral squamous cell carcinomas. Cancer Res. 61, 7044–7047 (2001).
86. Gstaiger, M. et al. Skp2 is oncogenic and overexpressed in human cancers. Proc. Natl Acad. Sci. USA 98, 5043–5048 (2001).First demonstrated overexpression and oncogenicity of SKP2 in human epithelial cancer.
87. Drobnjak, M. et al. Altered expression of p27 and Skp2 proteins in prostate cancer of African-American patients. Clin. Cancer Res. 9, 2613–2619 (2003).
88. Yokoi, S. et al. A novel target gene, SKP2, within the 5p13 amplicon that is frequently detected in small cell lung cancers. Am. J. Pathol. 161, 207–216 (2002).
89. Masuda, T. A. et al. Clinical and biological significance of S-phase kinase-associated protein 2 (Skp2) gene expression in gastric carcinoma: modulation of malignant phenotype by Skp2 overexpression, possibly via p27 proteolysis. Cancer Res. 62, 3819–3825 (2002).
90. Nakayama, K. et al. Skp2-mediated degradation of p27 regulates progression into mitosis. Dev. Cell 6, 661–672 (2004).
91. Kossatz, U. et al. Skp2-dependent degradation of p27kip1 is essential for cell cycle progression. Genes Dev. 18, 2602–2607 (2004).
92. Catzavelos, C. et al. Decreased levels of the cell-cycle inhibitor p27Kip1 protein: prognostic implications in primary breast cancer. Nature Med. 3, 227–230 (1997).
R E V I E W S
NATURE REVIEWS | cancer VOLUME 8 | APRIL 2008 | 265
© 2008 Nature Publishing Group

93. Porter, P. L. et al. Expression of cell cycle regulators p27kip1 and cyclin E, alone and in combination, correlate with survival in young breast cancer patients. Nature Med. 3, 222–225 (1997).
94. Tan, P. et al. The cell cycle inhibitor p27 is an independent prognostic marker in small (T1a, b) invasive breast carcinomas. Cancer Res. 57, 1259–1263 (1997).References 92–94 were the first to demonstrate that p27 levels are frequently reduced in human breast cancer and that this is prognostic for poor patient outcome on multivariate analysis.
95. Zeng, Y., Hirano, K., Hirano, M., Nishimura, J. & Kanaide, H. Minimal requirements for the nuclear localization of p27Kip1, a cyclin-dependent kinase inhibitor. Biochem. Biophys. Res. Commun. 274, 37–42 (2000).
96. LaBaer, J. et al. New functional activities for the p21 family of CDK inhibitors. Genes Dev. 11, 847–862 (1997).
97. Cheng, M. et al. The p21Cip1 and p27Kip1 CDK ‘inhibitors’ are essential activators of cyclin D-dependent kinases in murine fibroblasts. EMBO J. 18, 1571–1583 (1999).References 96 and 97 provide biochemical and genetic evidence that p27 and p21 regulate the assembly of D-type cyclin–Cdk complexes.
98. Liang, J. et al. PKB/Akt phosphorylates p27, impairs nuclear import of p27 and opposes p27-mediated G1 arrest. Nature Med. 8, 1153–1160 (2002).This paper showed that Akt phosphorylated p27 to impair its nuclear import.
99. Shin, I., Rotty, J., Wu, F. Y. & Arteaga, C. L. Phosphorylation of p27Kip1 at Thr-157 interferes with its association with importin alpha during G1 and prevents nuclear re-entry. J. Biol. Chem. 280, 6055–6063 (2005).
100. Sekimoto, T., Fukumoto, M. & Yoneda, Y. 14-3-3 suppresses the nuclear localization of threonine 157-phosphorylated p27Kip1. EMBO J. 23, 1934–1942 (2004).
101. Fujita, N., Sato, S., Katayama, K. & Tsuruo, T. Akt-dependent phosphorylation of p27Kip1 promotes binding to 14-3-3 and cytoplasmic localization. J. Biol. Chem. 277, 28706–28713 (2002).
102. Fujita, N., Sato, S. & Tsuruo, T. Phosphorylation of p27Kip1 at threonine 198 by p90 ribosomal protein S6 kinases promotes its binding to 14-3-3 and cytoplasmic localization. J. Biol. Chem. 278, 49254–49260 (2003).
103. Kardinal, C. et al. Tyrosine phosphorylation modulates binding preference to cyclin-dependent kinases and subcellular localization of p27Kip1 in the acute promyelocytic leukemia cell line NB4. Blood 107, 1133–1140 (2006).
104. Soos, T. J. et al. Formation of p27–CDK complexes during the human mitotic cell cycle. Cell Growth Differ. 7, 135–146 (1996).
105. Zhang, H., Hannon, G. & Beach, D. p21-containing cyclin kinases exist in both active and inactive states. Genes Dev. 8, 1750–1758 (1994).
106. Nagahara, H. et al. Transduction of full-length TAT fusion proteins into mammalian cells: TAT–p27Kip1 induces cell migration. Nature Med. 4, 1449–1452 (1998).
107. Cheng, M., Sexl, V., Sherr, C. J. & Roussel, M. F. Assembly of cyclin D-dependent kinase and titration of p27Kip1 regulated by mitogen-activated protein kinase kinase (MEK1). Proc. Natl Acad. Sci. USA. 95, 1091–1096 (1998).
108. Liu, X. et al. Disruption of TGF-β growth inhibition by oncogenic ras is linked to p27Kip1 mislocalization. Oncogene 19, 5926–5935 (2000).
109. Viglietto, G. et al. Cytoplasmic relocalization and inhibition of the cyclin-dependent kinase inhibitor p27Kip1 by PKB/Akt-mediated phosphorylation in breast cancer. Nature Med. 8, 1136–1144 (2002).
110. Shin, I. et al. PKB/Akt mediates cell-cycle progression by phosphorylation of p27Kip1 at threonine 157 and modulation of its cellular localization. Nature Med. 8, 1145–1152 (2002).References 98, 109 and 110 were published together and all showed that cytoplasmic mislocalization of p27 in human breast cancer was associated with activation of Akt.
111. Viglietto, G. & Fusco, A. Understanding p27kip1 deregulation in cancer: down-regulation or mislocalization? Cell Cycle 1, 394–400 (2002).
112. Motti, M. L., De Marco, C., Califano, D., Fusco, A. & Viglietto, G. Akt-dependent T198 phosphorylation of cyclin-dependent kinase inhibitor p27kip1 in breast cancer. Cell Cycle 3, e89–e95 (2004).
113. Psyrri, A. et al. Subcellular localization and protein levels of cyclin-dependent kinase inhibitor p27 independently predict for survival in epithelial ovarian cancer. Clin. Cancer Res. 11, 8384–8390 (2005).
114. Baldassarre, G. et al. p27Kip1-stathmin interaction influences sarcoma cell migration and invasion. Cancer Cell 7, 51–63 (2005).
115. McAllister, S. S., Becker-Hapak, M., Pintucci, G., Pagano, M. & Dowdy, S. F. Novel p27kip1 C-terminal scatter domain mediates Rac-dependent cell migration independent of cell cycle arrest functions. Mol. Cell Biol. 23, 216–228 (2003).
116. Wu, F. Y. et al. Reduction of cytosolic p27Kip1 inhibits cancer cell motility, survival, and tumorigenicity. Cancer Res. 66, 2162–2172 (2006).
117. Denicourt, C., Saenz, C. C., Datnow, B., Cui, X. S. & Dowdy, S. F. Relocalized p27KiP1 tumor suppressor functions as a cytoplasmic metastatic oncogene in melanoma. Cancer Res. 67, 9238–9243 (2007).
118. Besson, A. et al. Discovery of an oncogenic activity in p27Kip1 that causes stem cell expansion and a multiple tumor phenotype. Genes Dev. 21, 1731–1746 (2007).Identified a novel pro-oncogenic action of p27 to promote expansion of cells with self-renewal capacity or tumour-initiating function.
119. Jordan, R., Bradley, G. & Slingerland, J. M. Reduced levels of the cell-cycle inhibitor p27KIP1 in epithelial dysplasia and carcinoma of the oral cavity. Am. J. Pathol. 152, 1–6 (1998).
120. De Paola, F. et al. p27/kip1 expression in normal epithelium, benign and neoplastic breast lesions. J. Pathol. 196, 26–31 (2002).
121. Moriya, T. et al. Immunohistochemical analysis of Ki-67, p53, p21, and p27 in benign and malignant apocrine lesions of the breast: its correlation to histologic findings in 43 cases. Mod. Pathol. 13, 13–18 (2000).
122. Oh, Y. L. et al. Expression of p21Waf1, p27Kip1 and cyclin D1 proteins in breast ductal carcinoma in situ: Relation with clinicopathologic characteristics and with p53 expression and estrogen receptor status. Path. Int. 51, 94–99 (2001).
123. Han, S. et al. Reduced expression of p27Kip1 protein is associated with poor clinical outcome of breast cancer patients treated with systemic chemotherapy and is linked to cell proliferation and differentiation. Breast Cancer Res. Treat. 55, 161–167 (1999).
124. Massarelli, E. et al. Loss of E-cadherin and p27 expression is associated with head and neck squamous tumorigenesis. Cancer 103, 952–959 (2005).
125. Pruneri, G. et al. Clinical relevance of expression of the CIP/KIP cell-cycle inhibitors p21 and p27 in laryngeal cancer. J. Clin. Oncol. 17, 3150–3159 (1999).
126. Vis, A. N. et al. Prognostic value of cell cycle proteins p27kip1 and MIB-1, and the cell adhesion protein CD44s in surgically treated patients with prostate cancer. J. Urol. 164, 2156–2161 (2000).
127. Cordon-Cardo, C. et al. Distinct altered patterns of p27KIP1 gene expression in benign prostatic hyperplasia and prostatic carcinoma. J. Natl Cancer Inst. 90, 1284–1291 (1998).
128. Tsihlias, J. et al. Loss of cyclin-dependent kinase inhibitor p27Kip1 is a novel prognostic factor in localized human prostate adenocarcinoma. Cancer Res. 58, 542–548 (1998).
129. Sui, L. et al. Implication of malignancy and prognosis of p27Kip1, Cyclin E, and Cdk2 expression in epithelial ovarian tumors. Gynecol. Oncol. 83, 56–63 (2001).
130. Korkolopoulou, P. et al. The combined evaluation of p27Kip1 and Ki-67 expression provides independent information on overall survival of ovarian carcinoma patients. Gynecol. Oncol. 85, 404–414 (2002).
131. Yatabe, Y. et al. p27Kip1 in human lung cancers: differential changes in small cell and non-small cell carcinomas. Cancer Res. 58, 1042–1047 (1998).
132. Ishihara, S. et al. The cyclin-dependent kinase inhibitor p27 as a prognostic factor in advanced non-small cell lung cancer: its immunohistochemical evaluation using biopsy specimens. Lung Cancer 26, 187–194 (1999).
133. Hommura, F. et al. Prognostic significance of p27KIP1 protein and ki-67 growth fraction in non-small cell lung cancers. Clin. Cancer Res. 6, 4073–4081 (2000).
134. Tsukamoto, S. et al. Reduced expression of cell-cycle regulator p27Kip1 correlates with a shortened survival in non-small cell lung cancer. Lung Cancer 34, 83–90 (2001).
135. Hayashi, H. et al. High cyclin E and low p27/Kip1 expressions are potentially poor prognostic factors in lung adenocarcinoma patients. Lung Cancer 34, 59–65 (2001).
136. Hirabayashi, H. et al. Prognostic significance of p27KIP1 expression in resected non-small cell lung cancers: analysis in combination with expressions of p16INK4A, pRB, and p53. J. Surg. Oncol. 81, 177–184 (2002).
137. Takahashi, S. et al. Relationship between post-operative recurrence and expression of cyclin E, p27, and Ki-67 in non-small cell lung cancer without lymph node metastases. Int. J. Clin. Oncol. 7, 349–355 (2002).
138. Catzavelos, C. et al. Reduced expression of the cell cycle inhibitor p27Kip1 in non-small cell lung carcinoma: a potential prognostic factor independent of ras. Cancer Res. 59, 684–688 (1999).
139. Esposito, V. et al. Prognostic role of the cyclin-dependent kinase inhibitor p27 in non-small cell lung cancer. Cancer Res. 57, 3381–3385 (1997).
140. Hirsch, F. R. et al. Epidermal growth factor receptor in non-small-cell lung carcinomas: Correlation between gene copy number and protein expression and impact on prognosis. J. Clin. Oncol. 21, 3798–3807 (2003).
141. Lynch, T. J. et al. Activating mutations in the epidermal growth factor receptor underlying responsiveness of non-small-cell lung cancer to gefitinib. N. Engl. J. Med. 350, 2129–2139 (2004).
142. Amann, J. et al. Aberrant epidermal growth factor receptor signaling and enhanced sensitivity to EGFR inhibitors in lung cancer. Cancer Res. 65, 226–235 (2005).
143. Anayama, T., Furihata, M., Ishikawa, T., Ohtsuki, Y. & Ogoshi, S. Positive correlation between p27Kip1 expression and progression of human esophageal squamous cell carcinoma. Int. J. Cancer 79, 439–443 (1998).
144. Fan, G. K. et al. Expression of protein p27 is associated with progression and prognosis in laryngeal cancer. Laryngoscope 109, 815–820 (1999).
145. Itami, A., Shimada, Y., Watanabe, G. & Imamura, M. Prognostic value of p27Kip1 and CyclinD1 expression in esophageal cancer. Oncology 57, 311–317 (1999).
146. Mineta, H. et al. Low p27 expression correlates with poor prognosis for patients with oral tongue squamous cell carcinoma. Cancer 85, 1011–1017 (1999).
147. Fujieda, S. et al. Expression of p27 is associated with Bax expression and spontaneous apoptosis in oral and oropharyngeal carcinoma. Int. J. Cancer 84, 315–320 (1999).
148. Venkatesan, T. K. et al. Prognostic significance of p27 expression in carcinoma of the oral cavity and oropharynx. Laryngoscope 109, 1329–1333 (1999).
149. Shamma, A. et al. Loss of p27KIP1 expression predicts poor prognosis in patients with esophageal squamous cell carcinoma. Oncology 58, 152–158 (2000).
150. Kagawa, Y., Yoshida, K., Hirai, T. & Toge, T. Significance of the expression of p27Kip1 in esophageal squamous cell carcinomas. Dis. Esophagus. 13, 179–184 (2000).
151. Tamura, N. et al. Cyclin-dependent kinase inhibitor p27 is related to cell proliferation and prognosis in laryngeal squamous cell carcinomas. J. Laryngol. Otol. 115, 400–406 (2001).
152. Kuo, M. Y. et al. Prognostic role of p27Kip1 expression in oral squamous cell carcinoma in Taiwan. Oral Oncol. 38, 172–178 (2002).
153. Korkmaz, H. et al. Prognostic significance of G1 cell-cycle inhibitors in early laryngeal cancer. Am. J. Otolaryngol. 26, 77–82 (2005).
154. Langer, R. et al. Prognostic significance of expression patterns of c-erbB-2, p53, p16INK4A, p27KIP1, cyclin D1 and epidermal growth factor receptor in oesophageal adenocarcinoma: a tissue microarray study. J. Clin. Pathol. 59, 631–634 (2006).
155. Loda, M. et al. Increased proteasome-dependent degradation of the cyclin-dependent kinase inhibitor p27 in aggressive colorectal carcinomas. Nature Med. 3, 231–234 (1997).Among the first studies to show loss of p27 protein expression is prognostic in human cancer. Showed that lysates from colon cancers with low p27 levels exhibit proteolytic activity toward recombinant p27 in vitro.
156. Belluco, C. et al. Absence of the cell cycle inhibitor p27Kip1 protein predicts poor outcome in patients with stage I–III colorectal cancer. Ann. Surg. Oncol. 6, 19–25 (1999).
R E V I E W S
266 | APRIL 2008 | VOLUME 8 www.nature.com/reviews/cancer
© 2008 Nature Publishing Group

157. Palmqvist, R., Stenling, R., Oberg, A. & Landberg, G. Prognostic significance of p27Kip1 expression in colorectal cancer: a clinico-pathological characterization. J. Pathol. 188, 18–23 (1999).
158. Tenjo, T. et al. Prognostic significance of p27kip1 protein expression and spontaneous apoptosis in patients with colorectal adenocarcinomas. Oncology 58, 45–51 (2000).
159. Rossi, H. A. et al. The prognostic value of invariant chain (Ii) and her-2/neu expression in curatively resected colorectal cancer. Cancer J. 8, 268–275 (2002).
160. Noguchi, T. et al. Prognostic significance of p27/kip1 and apoptosis in patients with colorectal carcinoma. Oncol. Rep. 10, 827–831 (2003).
161. Galizia, G. et al. Determination of molecular marker expression can predict clinical outcome in colon carcinomas. Clin. Cancer Res. 10, 3490–3499 (2004).
162. Manne, U. et al. Prognostic significance of p27(kip-1) expression in colorectal adenocarcinomas is associated with tumor stage. Clin. Cancer Res. 10, 1743–1752 (2004).
163. Rosati, G., Chiacchio, R., Reggiardo, G., De Sanctis, D. & Manzione, L. Thymidylate synthase expression, p53, bcl-2, Ki-67 and p27 in colorectal cancer: relationships with tumor recurrence and survival. Tumour Biol. 25, 258–263 (2004).
164. Wu, J. T. et al. Prognostic significance of DCC and p27Kip1 in colorectal cancer. Appl. Immunohistochem. Mol. Morphol. 13, 45–54 (2005).
165. Watson, N. F. et al. Cytoplasmic expression of p27kip1 is associated with a favourable prognosis in colorectal cancer patients. World J. Gastroenterol. 12, 6299–6304 (2006).
166. Sarli, L. et al. Loss of p27 expression and microsatellite instability in sporadic colorectal cancer. Surg. Oncol. 15, 97–106 (2006).
167. Ciaparrone, M. et al. Localization and expression of p27KIP1 in multistage colorectal carcinogenesis. Cancer Res. 58, 114–122 (1998).
168. Cheng, J. D., Werness, B. A., Babb, J. S. & Meropol, N. J. Paradoxical correlations of cyclin-dependent kinase inhibitors p21waf1/cip1 and p27kip1 in metastatic colorectal carcinoma. Clin. Cancer Res. 5, 1057–1062 (1999).
169. Galizia, G. et al. Prognostic value of p27, p53, and vascular endothelial growth factor in Dukes A and B colon cancer patients undergoing potentially curative surgery. Dis. Colon Rectum 47, 1904–1914 (2004).
170. Yang, R. M. et al. Low p27 expression predicts poor disease-free survival in patients with prostate cancer. J. Urol. 159, 941–945 (1998).
171. Cote, R. J. et al. Association of p27Kip1 levels with recurrence and survival in patients with stage C prostate carcinoma. J. Natl Cancer Inst. 90, 916–920 (1998).
172. Freedland, S. J. et al. Preoperative p27 status is an independent predictor of prostate specific antigen failure following radical prostatectomy. J. Urol. 169, 1325–1330 (2003).
173. Li, R. et al. Biological correlates of p27 compartmental expression in prostate cancer. J. Urol. 175, 528–532 (2006).
174. Kuczyk, M. A. et al. Predictive value of altered p27Kip1 and p21WAF/Cip1 protein expression for the clinical prognosis of patients with localized prostate cancer. Oncol. Rep. 8, 1401–1407 (2001).
175. Cheville, J. C. et al. Expression of p27kip1 in prostatic adenocarcinoma. Mod. Pathol. 11, 324–328 (1998).
176. Thomas, G. V. et al. Preoperative prostate needle biopsy p27 correlates with subsequent radical prostatectomy p27, Gleason grade and pathological stage. J. Urol. 164, 1987–1991 (2000).
177. Freedland, S. J. et al. Predicting biochemical recurrence after radical prostatectomy for patients with organ-confined disease using p27 expression. Urology 61, 1187–1192 (2003).
178. Newcomb, E. W. et al. Expression of the cell cycle inhibitor p27KIP1 is a new prognostic marker associated with survival in epithelial ovarian tumors. Am. J. Pathol. 154, 119–125 (1999).
179. Baekelandt, M., Holm, R., Trope, C. G., Nesland, J. M. & Kristensen, G. B. Lack of independent prognostic significance of p21 and p27 expression in advanced ovarian cancer: an immunohistochemical study. Clin. Cancer Res. 5, 2848–2853 (1999).
180. Masciullo, V. et al. p27Kip1 expression is associated with clinical outcome in advanced epithelial ovarian cancer: multivariate analysis. Clin. Cancer Res. 6, 4816–4822 (2000).
181. Schmider-Ross, A. et al. Cyclin-dependant kinase inhibitors CIP1 (p21) and KIP1 (p27) in ovarian cancer. J. Cancer Res. Clin. Oncol. 132, 163–170 (2006).
182. Barnes, A. et al. Expression of p27kip I in breast cancer and its prognostic significance. J. Pathology 201, 451–459 (2003).
183. Gillett, C. E., Smith, P., Peters, G., Lu, X. & Barnes, D. M. Cyclin-dependent kinase inhibitor p27Kip1 expression and interaction with other cell cycle-associated proteins in mammary carcinoma. J. Pathol. 187, 200–206 (1999).
184. Barbareschi, M. et al. p27kip1 expression in breast carcinomas: an immunohistochemical study on 512 patients with long-term follow-up. Int. J. Cancer 89, 236–241 (2000).
185. Leivonen, M., Nordling, S., Lundin, J., von Boguslawski, K. & Haglund, C. p27 expression correlates with short-term, but not with long-term prognosis in breast cancer. Breast Cancer Res. Treat. 67, 15–22 (2001).
186. Han, S. et al. Prognostic implication of cyclin E expression and its relationship with cyclin D1 and p27Kip1 expression on tissue microarrays of node negative breast cancer. J. Surg. Oncol. 83, 241–247 (2003).
187. Porter, P. L. et al. p27Kip1 and cyclin E expression and breast cancer survival after treatment with adjuvant chemotherapy. J. Natl Cancer Inst. 98, 1723–1731 (2006).An important paper confirming the prognostic potential of p27 and shows that p27 loss is most prognostic in ER-positive human breast cancers in retrospective analysis of samples from a prospective randomized clinical trial.
188. Wu, J. et al. Prognostic role of p27Kip1 and apoptosis in human breast cancer. Br. J. Cancer 79, 1572–1578 (1999).
189. Tsuchiya, A., Zhang, G. J. & Kanno, M. Prognostic impact of cyclin-dependent kinase inhibitor p27kip1 in node-positive breast cancer. J. Surg. Oncol. 70, 230–234 (1999).
190. Chappuis, P. O. et al. Germline BRCA1/2 mutations and p27Kip1 protein levels independently predict outcome after breast cancer. J. Clin. Oncol. 18, 4045–4052 (2000).
191. Nohara, T., Ryo, T., Iwamoto, S., Gon, G. & Tanigawa, N. Expression of cell-cycle regulator p27 is correlated to the prognosis and ER expression in breast carcinoma patients. Oncology 60, 94–100 (2001).
192. Pohl, G. et al. High p27Kip1 expression predicts superior relapse-free and overall survival for premenopausal women with early-stage breast cancer receiving adjuvant treatment with tamoxifen plus goserelin. J. Clin. Oncol. 21, 3594–3600 (2003).The first study showing the predictive potential of p27 in a prospective trial. This important study showed that reduced p27 levels correlate with poor outcome in multivariate analysis and reduced response to endocrine therapy in pre-menopausal women with breast cancer in a retrospective analysis of p27 in samples collected in a prospective clinical trial.
193. Foulkes, W. D. et al. The prognostic implication of the basal-like (cyclin E high/p27 low/p53+/glomeruloid-microvascular-proliferation+) phenotype of BRCA1-related breast cancer. Cancer Res. 64, 830–835 (2004).
194. Reed, W., Florems, V. A., Holm, R., Hannisdal, E. & Nesland, J. M. Elevated levels of p27, p21 and cyclin D1 correlate with positive oestrogen and progesterone receptor status in node-negative breast carcinoma patients. Virchows Arch. 435, 116–124 (1999).
195. Slamon, D. J. et al. Use of chemotherapy plus a monoclonal antibody against HER2 for metastatic breast cancer that overexpresses HER2. N. Engl. J. Med. 344, 783–792 (2001).
196. Osborne, C. K. Steroid hormone receptors in breast cancer management. Breast Cancer Res. Treat. 51, 227–238 (1998).
197. Oka, K., Suzuki, Y. & Nakano, T. Expression of p27 and p53 in cervical squamous cell carcinoma patients treated with radiotherapy alone: radiotherapeutic effect and prognosis. Cancer 88, 2766–2773 (2000).
198. Cariou, S. et al. Down-regulation of p21WAF1/CIP1 or p27Kip1 abrogates antiestrogen-mediated cell cycle arrest in human breast cancer cells. Proc. Natl Acad. Sci. USA 97, 9042–9046 (2000).This paper demonstrated that p27 and p21 are required for growth arrest by tamoxifen and other antioestrogens in breast cancer cells in culture.
199. Lane, H. A. et al. ErbB2 potentiates breast tumor proliferation through modulation of p27Kip1–CDK2 complex formation: receptor overexpression does not determine growth dependency. Mol. Cell Biol. 20, 3210–3223 (2000).
200. Hayes, D. F. Prognostic and predictive factors for breast cancer: translating technology to oncology. J. Clin. Oncol. 23, 1596–1597 (2005).
201. Van’t Veer, L. J., Paik, S. & Hayes, D. F. Gene expression profiling of breast cancer: a new tumor marker. J. Clin. Oncol. 23, 1631–1635 (2005).
202. Oshita, F. et al. Increased expression levels of cyclin-dependent kinase inhibitor p27 correlate with good responses to platinum-based chemotherapy in non-small cell lung cancer. Oncol. Rep. 7, 491–495 (2000).
203. Kobayashi, M., Shiraishi, T., Tonouchi, H., Miki, C. & Kusunoki, M. 5-FU improves p27-related poor prognosis in patients with Astler-Coller B2-C colorectal carcinoma. Oncol. Rep. 9, 29–33 (2002).
204. Prall, F., Ostwald, C., Nizze, H. & Barten, M. Expression profiling of colorectal carcinomas using tissue microarrays: cell cycle regulatory proteins p21, p27, and p53 as immunohistochemical prognostic markers in univariate and multivariate analysis. Appl. Immunohistochem. Mol. Morphol. 12, 111–121 (2004).
205. Singh, S. P. et al. Loss or altered subcellular localization of p27 in Barrett’s associated adenocarcinoma. Cancer Res. 58, 1730–1735 (1998).
206. Zeng, W. Q. et al. Relationships between levels of Skp2 and p27 in breast carcinomas and possible role of Skp2 as targeted therapy. Steroids 70, 770–774 (2005).
207. Chang, J. et al. Intrinsic resistance of tumorigenic breast cancer cells to chemotherapy J. Natl Cancer Inst. (in the press).
208. Hattori, T. et al. Pirh2 promotes ubiquitin-dependent degradation of the cyclin-dependent kinase inhibitor p27Kip1. Cancer Res. 67, 10789–10795 (2007).
209. Hauck, L. et al. Protein kinase CK2 links extracellular growth factor signaling with the control of p27Kip1 stability in the heart. Nature Med. 14, 315–324 (2008).
AcknowledgementsThis work was supported by National Cancer Institute grant 1R01CA105118-01 to J.M.S. and by Austrian Science Fund (FWF, Projects P-18873 and Special Research Program “Cell Proliferation and Cell Death in Tumours” Grant SFB021) to L.H. J.M.S. is supported by the Braman Family Breast Cancer Institute of the University of Miami Sylvester Comprehensive Cancer Center. The authors apologize for omissions in citation and coverage. Strict space and citation limits preculde inclu-sion of many important past and recent works. We thank A. Farooq for help with generating figure 2.
DATABASESEntrez Gene: http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?db=geneCDK2 | CDK4 | CDK6 | CDKN1B | COPS5 | CKS1B | CUL1 | cyclin A | cyclin D | cyclin E | dacapo | DDB1 | Dicer 1 | DND1 | E2F1 | EGFR | ELAVL1 | ELAVL4 | ERBB2 | FOXM1 | FOXO4 | HES1 | MEN1 | MYC | PML | PTBP1 | RBX1 | SKP1 | SKP2 | SP1 | TGFβ | TRIM21 | vitamin D3 receptor | WAF1National Cancer Institute: http://www.cancer.gov/breast cancer | colon cancer | head and neck cancer | lung cancer | ovarian cancer | prostate cancer | stomach cancer
FURTHER INFORMATIONDr. Ludger Hengst’s homepage: http://www.i-med.ac.at/imcbc/bc/Dr. Joyce M. Slingerland’s homepage: http://www.sylvester.org/reserach/KB/scientist.asp?id=j_slingerland
SUPPLEMENTARY INFORMATIONSee online article:S1 (table)
aLLLInKSareactIVeIntheOnLInePDF
R E V I E W S
NATURE REVIEWS | cancer VOLUME 8 | APRIL 2008 | 267
© 2008 Nature Publishing Group


















