
The Beauty and Utility of DNA Origami The Beauty and Utility of DNA Origami Pengfei Wang,1,2 Travis...
24
Review The Beauty and Utility of DNA Origami Pengfei Wang, 1,2 Travis A. Meyer, 1,2 Victor Pan, 1 Palash K. Dutta, 1 and Yonggang Ke 1, * This review focuses on fundamental advances in DNA origami design and its emerging applications. For convenience, applications are coarsely categorized into nanofabrication, biosensing, and drug delivery. Nonetheless, considering the highly interdisciplinary nature of DNA nanotechnology, many works in the field do not fall within clearly defined boundaries. INTRODUCTION TO DNA ORIGAMI Over the past three decades, DNA, the genetic information carrier in most living or- ganisms, has seen an ever-expanding role as a material for the construction of nano- scale objects. 1–3 One technique in particular, known as DNA origami, has opened up the ability for researchers to design arbitrarily shaped complex three-dimensional (3D) nanostructures. Origami refers to the art of folding and sculpting a flat sheet of paper into arbitrarily shaped objects. DNA origami is a process of molecular self-folding: a long single-stranded DNA (scaffold), typically M13 phage genomic DNA (7,000 bp), is folded into prescribed objects by hundreds of short synthetic DNA oligonucleotides, typically 20–60 bp long, which are designed to be comple- mentary to different parts of the scaffold DNA (Figure 1A). The synthetic DNA strands crosslink spatially distant segments of a scaffold together, hence the term ‘‘staples.’’ For a comprehensive elaboration of DNA origami design principles, one can refer to this review article. 4 In the first report of DNA origami in 2006 by Paul Rothemund, 5 planar, arbitrarily shaped, two-dimensional (2D) objects with length scales around 100 nm were constructed (Figure 1B). Topologically, these 2D DNA origamis contained DNA double helices bundled together within a 2D plane by ‘‘crossovers,’’ where DNA strands crossed between neighboring DNA he- lices. Simple 3D hollow containers—a tetrahedron 6 (Figure 1C) and a cube 7 (Fig- ure 1D)—were constructed in 2009 via the folding of 2D planar DNA origami sheets. The milestone advancement in constructing 3D DNA origami structures was intro- duced by Shih and coworkers in 2009, 8 when they bundled DNA helices into a hon- eycomb lattice to construct solid 3D objects (Figure 1E). They developed a design platform called caDNAno, 9 which drastically simplifies the design process to enable a general route to the rapid manufacture of sophisticated shapes. The Shih group later demonstrated square 10 and hybrid lattice 11 design concepts. Subsequent work enabled the introduction of curvature by simply adding or deleting bases be- tween adjacent DNA helical crossovers (Figure 1F). 12 In a departure from the tradi- tional rigid lattice model, a study published in 2011 by Yan and coworkers used concentric DNA rings to define the surface feature of 3D objects, inducing curvature through the manipulation of strand crossover positions (Figure 1G). 13 The above DNA origami design strategies all pack DNA helixes in parallel to construct objects. In 2013, the Yan group developed a wire-frame design strategy that uses DNA four-arm junctions as vertices within a network of DNA helices to generate DNA gridiron 2D and 3D objects (Figure 1H). 14 They subsequently reported another method of creating more complex objects with a variety of multi-arm The Bigger Picture Functions arise from structures. At the nanoscale, precise organization of biological and non-biological materials in three- dimensional (3D) space holds great promise for a vast array of applications in the fields of structural biology, biophysics, sustainable energy, photonics, electronics, medicine, etc. Researchers are in constant pursuit of more potent methods that offer improved precision and higher resolution. Bottom-up structural nucleic acid (DNA and RNA) nanotechnology, specifically DNA origami, has emerged as a technique with unprecedented capability for the self-assembly of rationally designed nanostructures. DNA origami nanostructures are fully addressable with nanometer precision, which makes them a robust platform for the precise organization of other functional materials toward applications in a variety of fields. Chem 2, 359–382, March 9, 2017 ª 2017 Elsevier Inc. 359
Transcript of The Beauty and Utility of DNA Origami The Beauty and Utility of DNA Origami Pengfei Wang,1,2 Travis...

Review
The Beauty and Utility of DNA OrigamiPengfei Wang,1,2 Travis A. Meyer,1,2 Victor Pan,1 Palash K. Dutta,1 and Yonggang Ke1,*
This review focuses on fundamental advances in DNA origami design and its
emerging applications. For convenience, applications are coarsely categorized
into nanofabrication, biosensing, and drug delivery. Nonetheless, considering
the highly interdisciplinary nature of DNA nanotechnology, many works in the
field do not fall within clearly defined boundaries.
INTRODUCTION TO DNA ORIGAMI
Over the past three decades, DNA, the genetic information carrier in most living or-ganisms, has seen an ever-expanding role as a material for the construction of nano-scale objects.1–3 One technique in particular, known as DNA origami, has opened upthe ability for researchers to design arbitrarily shaped complex three-dimensional(3D) nanostructures. Origami refers to the art of folding and sculpting a flat sheetof paper into arbitrarily shaped objects. DNA origami is a process of molecularself-folding: a long single-stranded DNA (scaffold), typically M13 phage genomicDNA (!7,000 bp), is folded into prescribed objects by hundreds of short syntheticDNA oligonucleotides, typically 20–60 bp long, which are designed to be comple-mentary to different parts of the scaffold DNA (Figure 1A). The synthetic DNAstrands crosslink spatially distant segments of a scaffold together, hence the term‘‘staples.’’ For a comprehensive elaboration of DNA origami design principles,one can refer to this review article.4 In the first report of DNA origami in 2006 byPaul Rothemund,5 planar, arbitrarily shaped, two-dimensional (2D) objects withlength scales around 100 nm were constructed (Figure 1B). Topologically, these2D DNA origamis contained DNA double helices bundled together within a 2Dplane by ‘‘crossovers,’’ where DNA strands crossed between neighboring DNA he-lices. Simple 3D hollow containers—a tetrahedron6 (Figure 1C) and a cube7 (Fig-ure 1D)—were constructed in 2009 via the folding of 2D planar DNA origami sheets.
The milestone advancement in constructing 3D DNA origami structures was intro-duced by Shih and coworkers in 2009,8 when they bundled DNA helices into a hon-eycomb lattice to construct solid 3D objects (Figure 1E). They developed a designplatform called caDNAno,9 which drastically simplifies the design process to enablea general route to the rapid manufacture of sophisticated shapes. The Shih grouplater demonstrated square10 and hybrid lattice11 design concepts. Subsequentwork enabled the introduction of curvature by simply adding or deleting bases be-tween adjacent DNA helical crossovers (Figure 1F).12 In a departure from the tradi-tional rigid lattice model, a study published in 2011 by Yan and coworkers usedconcentric DNA rings to define the surface feature of 3D objects, inducing curvaturethrough the manipulation of strand crossover positions (Figure 1G).13
The above DNA origami design strategies all pack DNA helixes in parallel toconstruct objects. In 2013, the Yan group developed a wire-frame design strategythat uses DNA four-arm junctions as vertices within a network of DNA helices togenerateDNAgridiron 2Dand 3Dobjects (Figure 1H).14 They subsequently reportedanother method of creating more complex objects with a variety of multi-arm
The Bigger Picture
Functions arise from structures. Atthe nanoscale, preciseorganization of biological andnon-biological materials in three-dimensional (3D) space holdsgreat promise for a vast array ofapplications in the fields ofstructural biology, biophysics,sustainable energy, photonics,electronics, medicine, etc.Researchers are in constantpursuit of more potent methodsthat offer improved precision andhigher resolution. Bottom-upstructural nucleic acid (DNA andRNA) nanotechnology,specifically DNA origami, hasemerged as a technique withunprecedented capability for theself-assembly of rationallydesigned nanostructures. DNAorigami nanostructures are fullyaddressable with nanometerprecision, which makes them arobust platform for the preciseorganization of other functionalmaterials toward applications in avariety of fields.
Chem 2, 359–382, March 9, 2017 ª 2017 Elsevier Inc. 359

B C D E F
G H I J K
Q SR
L M N PO
T
Scaffold
Staples
+
Square LatticeHoneycomb Lattice
2D Origami 3D Origami with Helices Bundled on Lattices 3D Wireframe Origami A
Figure 1. Overview of DNA Origami Design Strategies and Structures
(A) DNA origami design strategies for constructing 2D and 3D objects.
(B) DNA origami smiley face. Reprinted by permission from Macmillan Publishers Ltd: Nature
(Rothemund5), copyright 2006.
(C and D) A hollow tetrahedron (C) and cube (D) formed from the folding of flat DNA origami
sheets. (C) Reprinted with permission from Ke et al.6 Copyright 2009 American Chemical Society.
(D) Reprinted by permission from Macmillan Publishers Ltd: Nature (Andersen et al.7), copyright
2009.
(E) A slotted cross constructed from honeycomb DNA lattice. Reprinted by permission from
Macmillan Publishers Ltd: Nature (Douglas et al.8), copyright 2009.
(F) A curved 6-helix bundle spiral-like object. From Dietz et al.12 Reprinted with permission from
AAAS.
(G) A nanoflask built from concentric rings of DNA helixes. From Han et al.13 Reprinted with
permission from AAAS.
(H) A 3D gridiron structure based on DNA four-arm junctions. From Han et al.14 Reprinted with
permission from AAAS.
(I) A wireframe flower-and-bird pattern. Reprinted by permission from Macmillan Publishers Ltd:
Nature Nanotechnology (Zhang et al.15), copyright 2015.
(J) A wireframe icosahedron. From Veneziano et al.16 Reprinted with permission from AAAS.
1Wallance H. Coulter Department of BiomedicalEngineering, Georgia Institute of Technologyand Emory University, Atlanta, GA 30322, USA
2Co-first author
*Correspondence: [email protected]
http://dx.doi.org/10.1016/j.chempr.2017.02.009
360 Chem 2, 359–382, March 9, 2017

junctions.15 Controlling the length of the connecting loops can tune the angle be-tween arms (Figure 1I). On the basis of this strategy, Bathe and colleagues developeda design algorithm called DAEDALUS to fully automate the design process, whichenabled them to create numerous polyhedral objects (Figure 1J).16 Hogberg and col-leagues reported an alternative wire-frame strategy that renders designed objectsinto triangulated meshes.17 To optimize the scaffold routing, they developed an al-gorithm (vHelix, based on Autodesk Maya). 3D objects such as the Stanford bunnywere fabricated from this method (Figure 1K). The wire-frame method has demon-strated its outstanding design power over creating symmetric and asymmetric ob-jects, which has largely enriched the design toolbox of DNA origami. In addition,these wire-frame objects have greater promise in biomedical applications becausethey have shown enhanced stability under physiological conditions,16,17 largely asa result of their wire-frame nature. Helices of conventional DNAorigami are generallydensely packed in parallel and thus largely depend on cations (e.g., Mg2+ and Na+),whichphysiological fluids lack, to neutralize the negative charge repulsion amonghe-lices during and after folding. Wire-frame origami structures typically have only oneor two helixes aligned, requiring less shielding and thus facilitating enhanced resis-tance to cation depletion under physiological conditions.
Expanding the size of DNA origami structures is desired for a variety of applications.One method is to use longer scaffold DNA, as demonstrated by several reports.24,25
Nonetheless, this requires the usage of a larger number of staple strands with uniquesequences and thus increases the cost. In addition, this method is not suitable forfabricating larger structures on the micron scale. A more feasible strategy is the hi-erarchical assembly of DNA origami tiles into discrete superstructures or 1D, 2D, and3D lattice structures through sticky-end cohesion or base-pair stacking. NumerousDNA origami discrete superstructures have been realized, as illustrated in Figures1L–1P.8,12,18–20 Micrometer-sized DNA origami lattice structures have also beenassembled through polymerization of DNA origami tiles either in aqueous solution(Figures 1Q–1S)20–22,26 or on a solid substrate (Figure 1T).23,27–29
RNA molecules largely resemble DNA both structurally and chemically but arebiologically more versatile. For instance, RNA has some advantages over DNA,
Figure 1. Continued
(K) A wireframe rabbit. Reprinted by permission from Macmillan Publishers Ltd: Nature (Benson
et al.17), copyright 2015.
(L) An icosahedron assembled from three origami units. Reprinted by permission from Macmillan
Publishers Ltd: Nature (Douglas et al.8), copyright 2009.
(M) A 12-tooth gear assembled from four origami units. From Dietz et al.12 Reprinted with
permission from AAAS.
(N) A cookie-like superstructure with nine square origami nanostructures assembled along a
preformed frame. Reprinted with permission from Zhao et al.18 Copyright 2011 American Chemical
Society.
(O) A hexagonal prism assembled from 12 DNA origami tripods. From Iinuma et al.19 Reprinted with
permission from AAAS.
(P) A robot assembled from three origami precursors. From Gerling et al.20 Reprinted with
permission from AAAS.
(Q) A 2D lattice assembled from a DNA origami cross motif. Reprinted with permission from Liu
et al.21 Copyright 2011 Wiley-VCH.
(R) A honeycomb 2D lattice assembled from a DNA origami hexagon motif. Reprinted with
permission from Wang et al.22 Copyright 2016 American Chemical Society.
(S) Dynamic 1D and 2D lattices. From Dietz et al.20 Reprinted with permission from AAAS.
(T) A 2D lattice of a triangle motif assembled on a mica substrate. Reprinted with permission from
Aghebat Rafat et al.23 Copyright 2011 Wiley-VCH.
Scale bars, 20 nm (C–F, K, and P), 50 nm (B, H, J, M, and S), 75 nm (G), 100 nm (I, L, N, O, and R inset),
1 mm (Q), 300 nm (Q, inset), 500 nm (R), and 200 nm (T).
Chem 2, 359–382, March 9, 2017 361

including a greater potential to be cloned and expressed in large quantities, as wellas functional capabilities such as scaffolding proteins, detecting ligands, andreleasing small interfering RNAs. RNA nanotechnology has grown in parallel alongwith the development of DNA nanotechnology.30,31 Nonetheless, the realizationof RNA origami had not been reported until 2013, when the Sugiyama32 andMao33,34 groups independently demonstrated the folding of a long single-strandedRNA (ssRNA, !1,000 bp) into prescribed objects by using DNA staples (Figure 2A,left). The ssRNA scaffold was in vitro transcribed from a T7-promoter-containingdouble-stranded DNA (dsDNA) template by T7 RNA polymerase. Objects wereassembled according to a conventional thermal annealing protocol. In 2014, Su-giyama and colleagues35 made pure RNA origami objects by using RNA staples tofold the RNA scaffold (Figure 2A, right). These RNA staples were also producedby in vitro transcription. In the same year, Andersen and colleagues36 reportedthe work of ssRNA origami (Figure 2B), which can self-fold into predesigned tilestructures. Angle-defined kissing loops were incorporated into the tiles to allow theirfurther hierarchical assembly into larger lattice structures, such as hexagonal 2D
A
PlasmidPCR
T7 Promoter
dsDNA Templates
ssRNA Scaffold
DNA Staples
RNA-DNA Hybrid Origami RNA Origami
T7 RNA Polymerase
RNA Staples
Annealing Annealing
BssRNA Origami
Co-transcriptional Assembly
T7 RNA Polymerase
T7 RNA Polymerase
4H-AO Tile
Hexagonal Lattice Assembled from 2H-AE Tile
dsDNA Template
Figure 2. Overview of RNA Origami
(A) RNA-DNA hybrid origami (left) and RNA origami (right) objects assembled via folding of a
ssRNA scaffold by short DNA or RNA staples. (Left) Reproduced from Wang et al.33 with permission
from the Royal Society of Chemistry. (Right) Reprinted with permission from Endo et al.35 Copyright
2014 Wiley-VCH.
(B) Co-transcriptional folding of ssRNA origami into tiles and further hierarchical assembly into
2D lattices. From Geary et al.36 Reprinted with permission from AAAS.
Scale bars, 100 nm.
362 Chem 2, 359–382, March 9, 2017

lattices. These ssRNAmolecules were produced by in vitro transcription with T7 RNApolymerase. In addition to using conventional thermal annealing, this work demon-strated that these RNA architectures can self-fold and assemble isothermally duringthe transcriptional process. Although RNA assembly principles are the same as thosefor DNA, which rely primarily on Watson-Crick base pairing, RNA assembly has itsown unique characteristics: (1) RNA-DNA and RNA-RNA duplexes have 11 bp perhelical turn, (2) abundant secondary and tertiary structures exist within RNA strands,and (3) RNA is more vulnerable to nuclease degradation. These aforementionedpoints need to be taken into account in the design and preparation of RNA origamiarchitectures.
In addition to DNA origami, single-stranded DNA (ssDNA) bricks developed by Yinand colleagues are another versatile toolset for constructing well-defined, arbitrarilyshaped DNA structures.37,38 Both strategies have their own strengths and limita-tions, and thus people should choose according to their specific applications:
(1) DNA origami typically produces higher yields of correct nanostructures. Theuse of a single scaffold and excess staples minimizes the possibility of missingstrands, which can occur in techniques such as DNA bricks, where precise stoi-chiometry among strands is required.
(2) DNA origami objects can have enhanced stability under working conditionsbecause the continuous scaffold DNA routes through and holds the wholeobject.
(3) DNA bricks possess higher modularity than DNA origami. A set of brickscould serve as a canvas to allow for the building of a large number of differentobjects, whereas a unique set of strands is generally necessary for building asingle origami object.
(4) DNA bricks offer a better choice for controlling the sequence and function-ality of DNA strands. For instance, a 4H 3 4T rectangle composed ofL-DNA of random sequence has demonstrated high nuclease resistance.38
DNA ORIGAMI FOR NANOFABRICATION
The DNA origami technique has been used extensively for the fabrication of nanoscaleconstructs, including many that are unobtainable (or extremely difficult to obtain) withother approaches. At the macroscopic scale, top-down fabrication of materialsis used to create products with specific structures and functions. Unfortunately, thissame capability has been difficult to achieve on the nanoscale, given that bottom-upself-assembly methods suffer from limited scope as a result of thermodynamic con-straints, and top-down methods are time consuming and costly. DNA origami helpsto bridge this fabrication gap, because the ability to self-assemble arbitrary shapeswith full addressability allows for the fabrication of nanostructures with well-definedform and the capacity to precisely arrange heteroelements. DNA origami has beenused as template structures for the organization of both synthetic nanomaterial compo-nents and biomolecules, as molds for nanoparticle synthesis, as masks for nanolithog-raphy, and as scaffolds for artificial enzyme cascades, etc. The devices fabricated bythese techniques have been utilized for many applications, including nanoplasmonics,nanophotonics, biosensing, and drug delivery. The last two applications are separatelyreviewed and discussed after the nanofabrication section.
Nanofabrication with Synthetic Nanomaterials
A wide variety of nanomaterials can interface with DNA through a range of conjuga-tion techniques, which have been well summarized in a previous review.39 By far the
Chem 2, 359–382, March 9, 2017 363

most commonmaterial used for nanofabrication with DNA origami is gold, althoughthere have been some reports on the use of silver.40 Gold nanoparticles (AuNPs) andgold nanorods (AuNRs) have very interesting optical properties that can be modu-lated through inter-particle interactions. Furthermore, AuNPs can be easily incorpo-rated into DNA nanostructures through the facile conjugation of thiolated-oligonu-cleotides with the gold surface. There are a vast number of examples in the literatureof nanofabrication using AuNPs, but only a few representative studies will be high-lighted here. Recent work by Oleg Gang’s group has demonstrated the ability forDNA origami nanostructures to program the fabrication of well-defined 2D networksof AuNPs.41 The authors used a cross-shaped origami tile with a hole in the center forcapturing a single AuNP. Theymodified the four edges of the tile with unique ssDNAextensions to allow for a wide range of anisotropic and specific connections betweendifferent tiles in a lock-and-key mechanism (Figure 3A). The authors used this systemto fabricate well-defined AuNP clusters and chains, as well as large 2D arrays. Thesame group has also used 3D polyhedral DNA origami nanostructures to constructwell-defined 3D AuNP crystal structures.51,52 Other groups have explored similarAuNP arrays, including the use of hexagonal tiles22 and ‘‘nano-flowers’’53 for con-structing linear chains and hexagonal and square lattices. In addition to organizingnanoparticles into well-defined patterns, DNA origami has also been utilized toselectively functionalize the surface of nanoparticles. For instance, Wang and col-leagues54 used a DNA origami clamp to encapsulate a AuNR, creating an address-able surface that facilitated the anchoring of AuNPs at selected positions to formAuNR-AuNP clusters of prescribed configurations. Fan and colleagues55 transferreda molecular pattern on DNA origami onto AuNPs by removing the origami templatewhile leaving oligonucleotides at selected locations on the surface of AuNPs, whichserved as docking sites for the attachment of other particles. Prior to this work, asimilar strategy was demonstrated by Sleiman and colleagues,56 who used non-origami DNA polyhedral structures.
The prevailing rationale for using DNA nanostructures to organize AuNPs is the abil-ity to use interactions between the nanoparticles as a means of manipulating light.This was elegantly demonstrated by Liedl and colleagues, who used a 24-helixbundle DNA origami rod to pattern the self-assembly of AuNPs into helices witheither a right-handed or left-handed pitch (Figure 3B).42 These nanostructuresexhibited pronounced circular dichroism effects in the visible range of the electro-magnetic spectrum, and these effects changed on the basis of the helicity of theorganized AuNPs. Subsequently, the same group demonstrated switchable circulardichroism by using a similar DNA origami rod-based architecture, where the orien-tation of the superstructure in reference to incident light could be controlled.57
Other groups have similarly used DNA origami to control the plasmonic behaviorof AuNRs, including a AuNR capable of walking along a DNA origami track,58
stacked arrangements of offset AuNRs,59 chiral AuNP rings,60 and static61 and dy-namic62 AuNR dimers.
The fabrication of devices where AuNPs interact with other optically active materialshas also been accomplished with DNA origami. In their pioneering work, Tinnefeldand colleagues constructed a zepto-liter electromagnetic ‘‘hotspot’’ by organizing100 nm AuNP dimers on a 12-helix bundle DNA origami pillar (Figure 3C).43 Whenthe authors placeda fluorescent dye in thegapbetween theAuNPs, theydemonstratedfluorescence enhancements of up to 117-fold. By using this system, the authors wereable to monitor DNA hybridization and Holliday junction conformational dynamics atthe single-molecule level. Other groups have conducted similar studies, including dis-tance-dependent quenching and enhancement of fluorescent-dye-AuNP systems on a
364 Chem 2, 359–382, March 9, 2017

X12 X12--+
=
57nm
10nm
34nm16nm
Right-Handed HelixLeft-Handed HelixA
5 nm
-3 nm
Cou
nts
/10m
s
1000
Time (s)
2000
2 4
CD
(mde
g)
50
-50
-25
25
0
400 500 600 700Wavelength (nm)
94nm
1000
2000
50
2 40
---
B C
D F
G H I
No NP
Dimer
E
HF Vapor (High Moisture)
HF Vapor (Low Moisture)
SiO2Si
J
1.0
0.5
0.0
1.5
2.0
Number Of Donors31 2 4 5 6
Donor (Cy3)Acceptor (Cy5)
G
D
g
S
Figure 3. DNA Origami for Nanofabrication with Synthetic Nanomaterials
(A) 2D AuNP arrays assembled by DNA origami. Reprinted by permission from Macmillan
Publishers Ltd: Nature Chemistry (Liu et al.41), copyright 2016.
(B) Chiral AuNP helixes assembled on DNA origami templates and subsequent interactions with
polarized light. Reprinted by permission from Macmillan Publishers Ltd: Nature (Kuzyk et al.42),
copyright 2012.
(C) DNA-origami-directed nanoantenna with two AuNPs forming a hotspot that increased the
photon count rate nearly 50 times. From Acuna et al.43 Reprinted with permission from AAAS.
(D) AuNP and quantum dot assembled on a triangular DNA origami. Reprinted with permission
from Samanta et al.44 Copyright 2014 American Chemical Society.
(E) DNA-origami-based light-harvesting system (left) and its antenna-effect dependence on the
number of donors (right). Reprinted with permission from Hemmig et al.45 Copyright 2016
American Chemical Society.
(F) DNA-origami-templated single-walled carbon nanotube (SWNT) cross-junction (inset) and
DNA-templated SWNT connection to electrodes. Reprinted by permission from Macmillan
Publishers Ltd: Nature Nanotechnology (Maune et al.46), copyright 2010.
(G) Casting of a silver nanoparticle within a DNA origami mold. From Sun et al.47 Reprinted with
permission from AAAS.
(H) Size-controlled liposomes templated by a DNA origami ring. Reprinted by permission from
Macmillan Publishers Ltd: Nature Chemistry (Yang et al.48), copyright 2016.
(I) Hydrofluoric acid (HF)-vapor-mediated negative (top) and positive (bottom) pattern transfer
from DNA origami to SiO2 substrate. Reprinted with permission from Surwade et al.49 Copyright
2011 American Chemical Society.
(J) AFM image of triangular DNA origami with three Cy5 dyes (top right) and AFM images with
corresponding epifluorescence images of origami structures in a photonic crystal cavity (PCC) (left).
Wide-field fluorescence microscopy images show optical emission of dye-modified DNA
nanostructures in lithography-patterned PCCs to create an image of van Gogh’s The Starry Night
(bottom right). Reprinted by permission from Macmillan Publishers Ltd: Nature (Gopinath et al.50),
copyright 2016.
Scale bars, 200 nm (A), 20 nm (B and G), 100 nm (F and I), 5 nm (F, inset), 50 nm (H), 250 nm (J, left),
and 50 nm (J, right).
Chem 2, 359–382, March 9, 2017 365

triangular DNA origami tile63 and ‘‘nano-antennas’’ for hotspot-induced enhancementof surface-enhanced Raman spectroscopy.64 AuNP-quantum-dot heteroelement de-vices have also been constructed for studying the distance-dependent fluorescencequenching between the two elements (Figure 3D).44 Similar work from the Liedl groupinvestigated the fluorescence lifetime of AuNP-quantum-dot pairs on a DNA origamiframe.65
The ability to control the transfer of energy on the nanoscale is another application inwhich precise fabrication is required. For example, arrangements of small-moleculefluorescent dyes have been fabricated for light-harvesting applications using DNAnanostructure templates. In one study, Tinnefeld, Chin, and colleagues were ableto drastically enhance the Forster resonance energy transfer (FRET)-induced emis-sion from an acceptor dye by varying the number and placement of donor dyes.45
When a ring-based structure (mimicking structural motifs found in photosyntheticbacteria) was used, the fluorescence intensity of the acceptor dye increased 2-foldin relation to the emission following direct excitation (Figure 3E). The ability to pre-cisely control the geometry of these energy-transfer pathways by using DNA origamican be used to engineer more efficient light-harvesting systems for applications suchas solar cells. The control of electronmovement has been demonstrated with carbonnanotube arrangements fabricated with DNA origami. Han and colleagues wereable to demonstrate that the construction of cross-junctions between two differentDNA-functionalized carbon nanotube populations (semi-conductor and metal)led to the generation of field-effect transistor behavior (Figure 3F).46 Gothelf andcolleagues66 recently used a DNA origami template to direct the routing of individ-ual polymers, which can serve as nanoscale wires with polymers that are suitablyconductive.
Although DNA origami nanostructures are primarily used in nanofabrication as ascaffold for the attachment of various elements, it has also been used as a scaffoldfor the synthesis of nanoparticles. In their pioneering work, Yin and colleaguesused hollow DNA origami constructs as a mold to facilitate the growth of both silverand gold nanoparticles into shapes defined by the DNA structure.47 They were ableto synthesize nanoparticles that differed in both size and shape by enclosing a smallAuNP seed inside a structurally rigid DNA origami frame, such that further growth ofthe nanoparticle was constrained by the DNA nanostructure (Figure 3G). In a similarapproach, Lin and colleagues designed DNA origami rings that could template thegrowth of liposomes.48 With lipid-modified DNA strands incorporated on the DNAframe as nucleation sites, the self-assembly of further lipids into a bilayer structureled to the formation of liposomes with a diameter defined by the DNA origamiring (Figure 3H). This allowed the authors to assemble highly monodisperse lipo-somes in four different sizes by using a variety of different lipid species. Otherstudies on the interaction between DNA nanostructures and lipids have been previ-ously reviewed.67
DNA origami structures have been used as masks for the construction of nanoscalefeatures via lithography, given that the size of the features created in this manner aremuch smaller than those created by other lithographic techniques. Yin, Liu, and col-leagues found that the presence of a silicon dioxide substrate altered the etchingrate of the substrate in the presence of hydrofluoric acid vapor (Figure 3I).49 In alow-moisture environment, the DNA nanostructures acted as a protective mask toprevent substrate etching, leading to a positive-tone pattern transfer. However, ina high-moisture environment, substrate etching was accelerated in the presenceof DNA, leading to negative-tone pattern transfer. This technique was improved
366 Chem 2, 359–382, March 9, 2017

by Tiron and colleagues, who created patterns with <10 nm resolution,68 and tech-niques involvingDNAmetallization have been used to transfer patterns to substratesother than silicon dioxide.69 DNA origami nanostructures have also been used inconjunction with traditionally fabricated lithographic substrates for the creation ofhybrid devices with enhanced functionality. Rothemund and colleagues were ableto guide the self-assembly of fluorescent-dye-modified DNA origami structures in-side lithographic-patterned photonic crystal cavities.50 The use of the DNA nano-structures allowed the authors to precisely control the number and location ofdyes within each cavity to create a system in which the intensity of cavity emissioncould be digitally varied (Figure 3J).
Nanofabrication with Biomolecules
In biology, the spatial and temporal arrangement of biomolecules into well-definedcomplexes leads to enhanced activity or new functionality. The ability to mimic thesecomplexes through nanofabrication with DNA origami is a useful tool for better un-derstanding native multi-protein systems in an effort to construct more efficientenzymatic nanoreactors, conduct single-molecule biophysical studies, or achievesuper-resolution optical imaging.
Advances in the ability to functionalize DNA nanostructures with proteins have enabledthe synthesis of a wide variety of nanofabricated biomolecule systems; for more infor-mation, we direct readers to an excellent review on the subject.70 Although simplerDNA nanostructures had been previously used for the assembly of multi-enzyme sys-tems, Yan and colleagueswere the first to useDNAorigami to scaffold amore complex,three-protein system.71 Using a 2D rectangular DNAorigami tile, the authors were ableto investigate the effects of protein distance on a bi-enzymatic process using glucoseoxidase and horseradish peroxidase. The authors found that placing a non-enzymatic‘‘bridge’’ proteinbetween the twoenzymescould increase the reaction rateeven further(Figure 4A). The authors attributed this effect to the more effective ‘‘shuttling’’ of reac-tion intermediates between the hydration shells of the two enzymes. Subsequentstudies by the same group increased the complexity of the design by utilizing a flexiblessDNA linkermodifiedwith cofactorNAD+ to control thepreference for two competingenzymatic reactions.79 3D DNA origami nanostructures have also been used for modu-lating and studying enzymatic behavior, such as using a ‘‘nanocage’’ to encapsulatemultiple enzymes to improve reaction efficiency.80
DNA origami nanostructures have also been used for controlling and studying non-enzymatic proteins. Shih, Reck-Peterson, and colleagues used a 12-helix bundleDNA origami tube as a platform for organizing the motor proteins kinesin anddynein.72 Using single-molecule fluorescence techniques to track the DNA nano-structure, the authors were able to study the interplay between motor proteinswith opposite-polarity movement (Figure 4B). Hogberg, Teixeira, and colleaguesalso used a tubular DNA origami structure to pattern proteins, wherein theyfocused on understanding how the spatial organization of ligands affects recep-tor-mediated cell responses.73 By controlling the distance between two Ephrin-A2 molecules, the authors were able to show how ligand placement alters receptorEphA2 phosphorylation and subsequent cell invasion (Figure 4C). Shih, Bellot, andcolleagues created a trapezoidal ‘‘nanoactuator’’ in which the activity of a splitenhanced green fluorescent protein (EGFP) could be controlled allosterically.74
The incorporation of additional DNA oligonucleotides could force the trapezoidinto either ‘‘closed’’ or ‘‘open’’ conformations, regulating the distance betweenthe EGFP halves and leading to increased fluorescence only in the closed confor-mation (Figure 4D).
Chem 2, 359–382, March 9, 2017 367

Time (min)0
1.0
0.6
10
β-Gal BridgeNTV BridgeNo Bridge
0.8
0.4
0.2
0.05 15 20
A
GOx HRPBridge
Hydration Layer
Nter-EGFP
Cter-EGFP
C DB
NA168
NA42
NA168Locking Strands
NA42Locking Strands
NA-UL
3 Dynein: 4 Kinesin
2.5
1.5
2.0
1.0
0.5
0.0
NCONC10
0NC40
NC
100
Tim
e
Rela
tive
Sign
al P
er C
ell
NC-Empty
E F
G H
0
1 70 nm
Imagewith P1*
Washing
Imagewith P2*
n
Overlayof n cycles
Design Schamatic
Single Particle Class Average
A Representative Single Molecule Image
Overlay of Scheamtic with Image
5 nm
5 nm
Exploring Base Stacking Forces on Single Molecule Level
M. jannaschii TBP
SSV T6 Promotor
Low FRET
High FRET
0 pN 2.5 pN 4.1 pN 6.2 pN 9.3 pN 11.4 pNN=15,224 N=11,489 N=10,631 N=10,432 N=11,968 N=12,733
0.4 0.8 0.4 0.8 0.4 0.8 0.4 0.8 0.4 0.8 0.4 0.8Nor
mal
ized
co
unts
1
Force (pN)0 5 10 15 20
P bent
0.1
0.5
FRET Efficiency E
Figure 4. DNA Origami for Nanofabrication with Biomolecules
(A) Assembly of a GOx-HRP cascade on DNA origami with a protein bridge in between to facilitate
H2O2 diffusion through a connected hydration layer and increased reaction rates. Reprinted with
permission from Fu et al.71 Copyright 2012 American Chemical Society.
(B) Single-molecule fluorescent measurements of the movement of DNA origami nanostructures
attached to opposite-polarity motor proteins. From Derr et al.72 Reprinted with permission from
AAAS.
(C) DNA origami nanostructure modified with Ephrin-A2 molecules (left) and relative
phosphorylation activity from structures carrying different ligand organizations (right). Scale
bars, 20 nm. Reprinted by permission from Macmillan Publishers Ltd: Nature Methods (Shaw
et al.73), copyright 2014.
(D) DNA-nanodevice-based eGFP complementation, a method for regulating biomolecular
interactions. Reprinted by permission from Macmillan Publishers Ltd: Nature Communications (Ke
et al.74), copyright 2016.
(E) DNA-origami-based single-molecule study of base-stacking forces. From Kilchherr et al.75
Reprinted with permission from AAAS.
(F) DNA-origami-based single-molecule force clamp for measuring DNA bending induced by
TATA-binding protein via changes in FRET intensity. From Nickels et al.76 Reprinted with
permission from AAAS.
(G) DNA origami rectangle serves as a substrate for studying transient binding of dye-labeled
oligonucleotides for achieving multiplexing super-resolution imaging via DNA-PAINT. Scale bar,
25 nm. Reprinted by permission from Macmillan Publishers Ltd: Nature Methods (Jungmann
et al.77), copyright 2014.
(H) Optical imaging of densely packed clusters on a DNA origami substrate with 5 nm resolution by
DNA-PAINT. Scale bars, 10 nm. Reprinted by permission from Macmillan Publishers Ltd: Nature
Nanotechnology (Dai et al.78), copyright 2016.
368 Chem 2, 359–382, March 9, 2017

The nature of DNA origami structures makes them excellent substrates for conductingdelicate single-molecule biophysical studies. Dietz and colleagues75 recently utilized aDNA origami structure to study the forces and lifetimes of DNA base-pair stacking in-teractions with optical tweezers (Figure 4E). Two DNA origami beams were attachedto micrometer-sized beads, which were trapped by optical tweezers. The two beamswere tethered together by a 68 bp ssDNA. An array of blunt-end DNA duplexes withvaried numbers protruded from each beam. The authors could examine the stackingforces and lifetimes between the arrays of blunt ends on either arm by tracking theforce-extension and constant-distance data with optical tweezers. They found thatstacking forces dependedon the number, orientation, and typeof base, in good agree-ment with previous studies. Another report by Liedl, Tinnefeld, and colleagues76 em-ployed DNA origami as a nanoscopic force clamp to investigate known biophysicalmodels (e.g., Holliday junction and TATA-binding protein) on a single-molecule basis(Figure 4F). This nanoscopic force clamp is based on measuring the efficiency ofFRET at distances correlated to varying entropic forces acting on the system. The au-thors controlled the entropic force by varying the contour length of the ssDNA springtethered to the origami substrate. Utilizing this system, they observed that the TATA-binding protein induced Holliday-junction conformer transitions and DNA duplexbending.
Another utilization of DNA origami structures lies in super-resolution microscopystudies based on the DNA-PAINT81 technology, a variation of point accumulationfor imaging in nanoscale topography. DNA-PAINT imaging is carried out withdiffusing, dye-labeled short oligonucleotides that interact transiently with the sam-ple. DNA origami could serve as a perfect substrate for studying and improving theDNA-PAINT technology before its broad application in imaging biological samples,where the Yin group has been the pioneer in this exciting field. They have largelydemonstrated the capabilities of DNA-PAINT for multiplex super-resolution imag-ing (Figure 4G),77 quantitative super-resolution imaging (qPAINT),82 and super-res-olution imaging of biomolecules in densely packed clusters with 5 nm resolution(Figure 4H),78 all benchmarked on DNA origami substrates.
Challenges and Perspectives
Onemajor constraint on the use of DNAorigami for fabrication at the nanoscale is thesize limitations—the overall dimensions of these nanostructures are limited by thelength of the scaffold strand. Given that ssDNA scaffolds significantly longer thanthe traditionally used M13 genome are not only technically challenging to acquirebut also mechanically fragile, it seems unlikely that the fabrication of large-scale as-semblies can be achieved with traditional DNA origami. One potential option is touse DNA origami nanostructures as individual building blocks for the formation oflarger superstructures.19 Another limitation is the relatively static nature of thesenanostructures; the ability to fabricate dynamic materials with programmable move-ment at the macro-level adds a level of utility that cannot easily be translated to thenanoscale. A considerable amount of work has gone into developing dynamic DNAmachines, highlighted in a previous review,83 particularly through the use of DNA-driven strand-displacement reactions. However, further advances in the ability tocontrol the movement of DNA origami structures would open up a range of new util-ities for nanofabricated devices. In particular, dynamic mechanisms that occur morequickly, are more complex, or can be achieved without the introduction of new DNAspecies are needed. Stimuli-responsive alternative nucleic acid structures, such asi-motifs, aptamers, and G-quadruplexes, potentially offer methods for designing‘‘smart’’ dynamic nanostructures. Work in this field has been highlighted in severalexcellent reviews.84,85
Chem 2, 359–382, March 9, 2017 369

DNA ORIGAMI FOR BIOSENSING
The characteristics of DNA origami nanostructures, and the associated benefits,offer potential advantages for use in sensing applications. Biosensing is a criticallyimportant function in a diverse set of fields ranging from medicine to food scienceto military defense. Biosensors are used for studying genetic mutations, detectingpathogenic microorganisms, and monitoring air quality, etc. The nanometeraddressability of DNA origami allows for the precise placement of various sensingelements in complex configurations, and the bottom-up self-assembly methodmakes production significantly simpler than comparable top-down approaches.DNA origami nanostructures are especially useful for single-molecule biosensing,which facilitates ultra-low detection limits as well as the ability to directly monitordynamic processes. The large profile of DNA nanostructures, particularly 2D de-signs, facilitates immobilization onto surfaces such as mica or glass, and the abilityto place sensing elements in asymmetric patterns simplifies target identification.DNA origami sensors are also very promising for multiplexed assays for many ofthe same reasons listed previously. In a broad sense, DNA origami constructs usedfor biosensing can be divided into two categories: binding-based devices andnanopores.
Binding-Based Biosensors
DNA origami nanostructures that fall within this category utilize a specific molecularinteraction (nucleic acid hybridization, protein-ligand binding, etc.) to alter theshape or profile of the sensor. These conformational changes can be observed viasingle-molecule techniques such as atomic force microscopy (AFM), transmissionelectron microscopy (TEM), or optical tweezers for identification of the target. Giventhe high specificity and well-understood behavior of Watson-Crick base pairing, it isno surprise that DNA origami nanostructures have been used for detecting specificnucleic acid sequences with considerable success. In their pioneering work, Yan andcolleagues were the first to demonstrate that DNA origami could be used as a plat-form for single-molecule mRNA detection.86 The authors used a rectangular 2Dnanostructure with sequences complementary to three different genes (Rag1,Myc, and Actb) extending out from the surface. When the target mRNA specieswere present, they bound to these ‘‘capture’’ probes and formed a dsDNA V-shapedprotrusion, which could be visualized by AFM. By using pre-formed DNA-dumbbellextensions arranged in asymmetric patterns to provide a ‘‘barcode,’’ the authorscould discriminate between different nanostructures containing probes for eachgene, thus providing multiplexed assaying (Figure 5A). Using this approach, the au-thors were able to get semiquantitative estimates of target concentration and wereable to demonstrate picomolar sensitivity. In a similar work, Fan, He, and colleaguesused an asymmetric ‘‘Map of China’’ 2D origami nanostructure, which obviates theneed for additional indexing agents for target identification during multiplexed as-says.94 They utilized biotinylated DNA oligomers complementary to the targetsequence, which enabled the use of streptavidin-biotin binding to provide a clearerAFM signal. However, this approach also increases cost by using modified oligonu-cleotides for each target.
Along with detecting specific genes, DNA origami biosensors have also been devel-oped to detect single-nucleotide polymorphisms (SNPs). In subsequent work, Fan,He, and colleagues used their Map of China structure to discriminate between twoDNA sequences containing a single-base mismatch.95 In this instance, the authorsutilized a kinetically controlled toe-hold-mediated strand-displacement process inwhich a perfectly matched target sequence displaced a biotinylated oligomer from
370 Chem 2, 359–382, March 9, 2017

Without Targets
WithTargets
Control β-actinC-mycRag-1
Locked Tiles
Target
Unlocked Tiles
50 n
m 0 nM PDGF
25 nM PDGF
20 secExt
ensi
on
Time
+ SA
SA
Curr
ent/
nA
0
5
10
0 5 10 15 2520Time/s
Level 1
Level 2
Level 1
Level 2
ssDNAOverhang
Level 1
Level 2
Prey
AT-Prey
CGCG-Prey
5s
0.1 nA
5s
0.1 nA
Baseline
HP Backward
HP Forward
1 2 3 4
1 2 3 4
100 ms10 p
A
500 ms
50 p
A
500 µs
50 p
A
A B
C D
E F
HG
Figure 5. DNA Origami for Biosensing
(A) Detection of specific mRNA sequences after hybridization to a rectangular DNA origami
template. From Ke et al.86 Reprinted with permission from AAAS.
(B) Detection of SNPs with a kinetically controlled toe-hold-mediated strand-displacement
reaction. Reprinted with permission from Subramanian et al.87 Copyright 2011 American Chemical
Society.
(C) Protein detection using DNA origami pliers. Binding of target molecules leads to a
conformational change from X-shaped to parallel-shaped structures. Scale bars, 300 nm. Reprinted
by permission from Macmillan Publishers Ltd: Nature Communications (Kuzuya et al.88), copyright
2011.
(D) Mechanochemical sensing of PDGF with a chain-linked DNA nanostructure. Binding of target
molecules leads to increases in structure flexibility and the distance between optical beads.
Reprinted with permission from Koirala et al.89 Copyright 2014 Wiley-VCH.
(E) Funnel-like origami structure with a tail to guide the correct insertion of origami into nanopores
and a current-versus-time graph of origami insertion into a solid-state nanopore. Reprinted with
permission from Bell et al.90 Copyright 2012 American Chemical Society.
(F) Origami nanoplate on a nanocapillary with overhanging predator strands (top left) capturing
prey strands (bottom left). Current traces show that CGCG prey strands (top right) are retained for a
significantly longer time than AT prey strands (bottom right). Reprinted with permission from
Hernandez-Ainsa et al.91 Copyright 2013 American Chemical Society.
(G) Origami nanopore and current traces showing the translocation of a single DNA hairpin (HP).
From Langecker et al.92 Reprinted with permission from AAAS.
(H) Large origami nanopore and current traces showing the translocation of dsDNA. Reprinted by
permission from Macmillan Publishers Ltd: Nature Communications (Krishnan et al.93), copyright
2016.
Chem 2, 359–382, March 9, 2017 371

the nanostructure, leading to a loss of streptavidin binding. When a mismatchedtarget was used, the strand-displacement reaction stalled at the mismatched base,and the biotinylated strand was not removed. Seeman and colleagues extendedthis work by using a similar kinetically controlled strand-displacement reactionto displace incumbent ‘‘signal strands,’’ leading to a double-stranded to single-stranded conformational change that could be visualized by AFM.87 The authorspatterned the signal strands to form the characters A, C, G, and T within the samenanostructure, such that a specific mutation in the target sequence would lead todisplacement of specific signal strands and the subsequent disappearance of thecorresponding character (Figure 5B).
3D DNA nanostructures have been used for sensing specific DNA sequences by us-ing a conformational change of the entire structure as the signal output. Komiyamaand colleagues used a ‘‘plier’’-shaped nanostructure that transitioned between threedistinct parallel, anti-parallel, and X-shaped conformations upon target binding,88
whereas Shih, Bellot, and colleagues used a trapezoidal structure in which bindingof the target sequence changed the angle between two arms.74
Sensors for protein biomarkers have also been fabricated with DNA origami.Although the interface between DNA nanostructures and any arbitrary nucleicacid target is clear, the sensing of proteins is currently limited to a select numberof targets, typically proteins that interact directly with DNA (enzymes, binding ele-ments, proteins with aptamer targets, etc.) or with a small number of oligonucleotidemodifications (biotin, fluorescein, etc.). Komiyama and colleagues were the first touse a DNA origami nanostructure to detect protein species by using their plier-shaped system.88 Binding of a target molecule to detection elements on botharms brings the two arms together, leading to a conformational transition betweenthe energetically favorable X shape to the more unfavorable linear-parallel confor-mation (Figure 5C). This transition was monitored at the single-molecule level byAFM, as well as bulk measurements from FRET-fluorescence microscopy. Althoughthe initial targets used for this study have limited downstream applicability (strepta-vidin and anti-fluorescein IgG bind to biotin and fluorescein, respectively), it is easyto envision this system adapting to more biomedically relevant species through theaddition of new detection elements. The authors also used this system to detectother species, including specific metal ions (sodium, potassium, and silver), ATP,and pH changes.96
Subsequent work by other groups extended the use of DNA origami nanostructuresfor protein biosensing to other target molecules. In 2014, Sugiyama, Mao, and col-leagues used a mechanochemical-sensing system in which the presence of a targetanalyte led to a change in the mechanical properties of a DNA origami nanostruc-ture, which could be detected with optical tweezers.89 The authors designed aseven-tiled, 2D structure with fixed linkages between tiles formed on one edgeand a dsDNA ‘‘locking’’ duplex linkage on the other edge. If the ‘‘locking’’ strandswere displaced, the flexibility of the tiled-structure drastically increased, and thischange could be observed as a stepwise increase in distance with optical tweezers(Figure 5D). The authors demonstrated that this approach could be used for thesensing of platelet-derived growth factor (PDGF) if the locking strands were formedfrom a PDGF aptamer—in the presence of PDGF, the locking duplex dissociated infavor of the more energetically stable aptamer-protein interaction. Using this tech-nique, the authors found that concentrations as low as 10 pM could be detectedwithin 10 min. By using two different locking strands within the same tiled structure,they also demonstrated multiplexed assaying.
372 Chem 2, 359–382, March 9, 2017

Other examples of protein-based biosensing using DNA origami nanostructuresinclude the detection of restriction enzymes with the trapezoidal nanoactuatordeveloped by Shih, Bellot, and colleagues74 and the use of aptamers on 2D DNAorigami templates for detecting the malaria biomarker Plasmodium falciparumlactate dehydrogenase (PfLDH)97 and thrombin.98
Additionally, DNA origami nanostructures have been used for detecting and study-ing dynamic processes of biomolecules at the single-molecule level. This work, pio-neered by Endo and Sugiyama and discussed in detail in an excellent review, useshigh-speed AFM imaging within a structurally rigid DNA origami frame to monitorchanges in various substrates, including alternative nucleic acid conformationssuch as G-quadruplexes and i-motifs, as well as protein activities such as DNAmethylation and repair.99
Biosensors Using DNA Origami Nanopores
Nanopores have been used extensively in biosensing applications. This technologyis capable of sensing molecules while foregoing the need for chemical modificationsor amplification of the analyte, making it attractive as a versatile method for molec-ular detection.100,101 Application of a voltage across a thin membrane containing asingle pore creates an electrophoretic force that causes molecules to translocatethrough the pore. These molecules displace the conducting ions in the pore,reducing conductivity and causing a measurable drop in current. Although the ma-jority of applications use silicon-based membranes (solid-state nanopores), there isalso interest in studying and using biological nanopores.
When solid-statemembranes areused, producingnanoporesof smaller (<10nm)diam-eter requires time-consuming electron-beam lithography techniques, and preciselycontrolling the placement of binding sites is challenging.100 Thus, DNA origami hasbeen explored as a tool for offering greater geometrical and chemical control ofsolid-state nanopores. Recent research has reported the use of DNAorigami structuresin conjunction with solid-state nanopores for the formation of nanopore-DNA origamihybrids. When subjected to an electrical current, the negatively charged DNA experi-ences an electrophoretic force toward the positive pole of the nanopore, leading a sin-gle origami to settle over the solid-state pore to form a nanopore-DNA origami com-plex. To demonstrate this concept, Keyser and colleagues used a funnel-shapedorigami bearing a long tail (Figure 5E), which they threaded through the pore to allowthe funnel to insert in the correct orientation.90 The DNAorigami structures themselvescan contain a pore with prescribed dimensions bearing various ligands or other modi-fications, so any molecules translocating through the nanopore-DNA origami complexcould be subject to interactions with the nanostructure pores’ chemical modifications.One example of this has been demonstrated by the Keyser group, who added ‘‘pred-ator’’ strands to the origami pore (Figure 5F).91 These strands bind the target ‘‘prey’’strands, increasing their translocation time and creating a more pronounced currentdrop than non-specific molecules.
Separately from these solid-state nanopores, biological nanopores exist in abun-dance in all living organisms and are necessary for the intra- and extracellular traf-ficking of ions and molecules.102 Their important role in facilitating this transporthas made them popular topics of study for understanding how cells move ionsand biological material. Researchers have extensively studied nanopore proteins,such as a-hemolysin, and are now interested in designing and fabricating syntheticbiological nanopores. Given the need to engineer these nanopores with preciselydesigned dimensions and characteristics, DNA origami has arisen as the technology
Chem 2, 359–382, March 9, 2017 373

of choice. With the customizability, versatility, and biological compatibility of DNA,nanopores fabricated from DNA are allowing researchers to better study existingbiological pores as well as utilize these pores for various technical applications.
The first biological nanopores fabricated with DNA origami were ‘‘nanobarrels,’’which contain a channel that can act as the pore. The seminal work in this fieldwas by Dietz, Simmel, and colleagues, who incorporated a cap bearing 26 hydro-phobic moieties between the barrel and a protruding stem (Figure 5G).92 These hy-drophobic moieties allowed the structure to thermodynamically favor incorporationinto lipid bilayers, effectively spanning the vesicle membrane to form an open pore.This work successfully demonstrated that functioning synthetic biological nanoporescould be designed with precise geometrical dimensions.
Although Dietz, Simmel, and colleagues and other groups have demonstrated DNAnanopore integration into lipid bilayers, spontaneous insertion remains highly incon-sistent. Subsequent work by the same group addressed this issue by using anew design featuring a larger moiety-bearing platform with a protruding stem ofincreased pore diameter.93 This design allows for the attachment of 57 hydrophobicmoieties, enabling the biological pore to spontaneously insert into lipid mem-branes. In addition, these pores demonstrate increased physical and electrically sta-bility and allow for the passage of larger analytes, increasing the potential of origaminanopores for use in versatile sensing applications (Figure 5H).
Challenges and Perspectives
Despite the advances made in biosensing using DNAorigami, particularly at the single-molecule level, the technology still has a longway togobefore it can live up to its consid-erablepotential. Forexample, the current useofDNAorigami inbiosensingapplicationsis limitedby the scopeof available analytes. Thedevelopment of newmethods for incor-porating a wider variety of biomolecules into DNAorigami nanostructures, including ni-trilotriacetate conjugation for the incorporation of His-tagged proteins103 andmethodsfor introducing single-stranded oligonucleotides onto specific locations on proteins,104
is already underway. For binding-based devices, the current techniques used for moni-toringanalyte-sensor interactions are limitedby their relatively low throughput and tech-nically challenging and expensive nature. AlthoughAFM, TEM, andoptical tweezers areexcellent tools for single-molecule studies in research lab settings, new outputs areneededbeforeDNAorigami sensors becomepractical for real-world applications.Mag-netic susceptibility and surface-enhanced Raman scattering are a couple of strategiesthat could be incorporated into DNA origami nanostructures to provide alternativesensing outputs. For DNA origami nanopores, a major challenge lies in the high ionicpermeability of the origami structures. DNA origami nanostructures only a few layersthick leak ions readily, reducing the contrast between background current and a trans-location event.105 To increase detection sensitivity, a method to reduce ion leakagemust be developed. Progress in the field of DNA structures as biological nanoporeshas advanced quickly, such that bilayer integration and transport regulation havebeen achieved in just a few years. However, there have yet to be any reported studiesintroducing selectivity to these nanopores to allow for the detection of specific species.Additional modifications to the biological nanopore geometry or chemistry are neededin order for biosensing using this technology to have significant technical applications.
DNA ORIGAMI FOR DRUG DELIVERY
The field of nanomedicine is founded upon the development of nanotechnology,especially the capability of fabricating nanoscale objects to be utilized as delivery
374 Chem 2, 359–382, March 9, 2017

vehicles for a variety of therapeutic cargos, including small molecules, proteins, nu-cleic acids, etc. DNA nanostructures have some essential characteristics of an effec-tive, customizable delivery system: (1) good biocompatibility, (2) full addressability,and (3) control over size, shape, and surface chemistry. In particular, the address-ability and control of nanostructure design are particular strengths that are infeasiblewith many comparable nanoparticle delivery systems. Although we specifically focuson DNA origami technology, comprehensive reviews of other DNA nanostructuresfor drug-delivery applications are available elsewhere.106,107
Delivery of Small Molecules
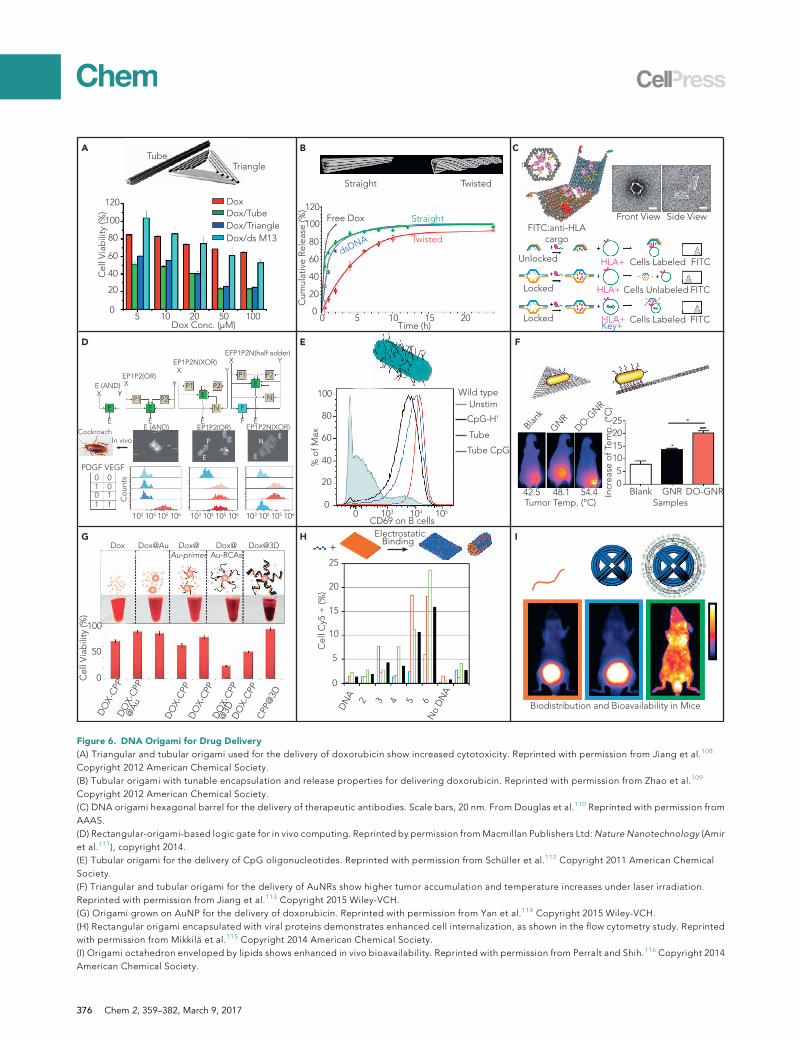
Anthracyclines (e.g., doxorubicin and daunorubicin) are a class of widely used anti-can-cer drugs that can intercalate into DNA duplexes and thus inhibit macromolecularbiosynthesis. DNA origami nanostructures have served as potent delivery vehicles fordoxorubicin or daunorubicin, given that a large number of DNA bases are availablefor non-covalent loading of these drugs. The first study was reported in 2012 byDing, Yan, and colleagues (Figure 6A), who utilized tubular and triangular DNAorigamistructures to deliver doxorubicin to induce cytotoxicity in doxorubicin-resistant cancercells.108 TheDNAorigami carriers led tomore cellular accumulation of doxorubicin andcytotoxicity than free doxorubicin. In 2014, the same group used DNA origami struc-tures to deliver doxorubicin for in vivo cancer therapy.117 Relative to controls, doxoru-bicin-loaded DNA origami showed significantly inhibited tumor growth. Hogberg andcoworkers demonstrated that adjusting the global twisting of DNA origami structurescan tune the encapsulation efficiency and release rate of doxorubicin (Figure 6B).109
They delivered doxorubicin into three different breast cancer cell lines by using DNAorigami nanostructures and found that cytotoxicity was higher and the cellular elimina-tion rate was lower than for free doxorubicin. In another study, Castro and colleaguesdesigned a rod-like DNA origami structure to load and deliver daunorubicin into adrug-resistant leukemia model with significantly enhanced cytotoxicity.118
Delivery of Proteins and Nucleic Acids
Proteins and nucleic acids have long served as functional materials for disease diag-nosis and therapy. Unlike the intercalating drugs, proteins and nucleic acids typicallyneed to be loaded through Watson-Crick base pairing between capturing strands onthe DNA structures and linking strands conjugated to proteins and nucleic acid cargos.In one study, Church’s group loaded antibodies into a DNA origami barrel nanostruc-ture whose transition between open and closed states was controlled via aptamer-based ‘‘locks’’ (Figure 6C).110 Upon recognition of cell-surface receptor ‘‘keys’’ on thecell membrane, the barrel was opened to release Fab antibody fragments that bindto human CD33 and CDw328 to inhibit growth of leukemia cells. The logic-gated lockswith specific recognition of cell membrane markers were designed to enhance target-ing specificity. Beyond this in vitro computing, Bachelet and colleagues took DNAorigami robot-based computing to in vivo models. They designed a series of DNA-origami-based logic gates that could conduct computational work in a cockroach ani-mal model (Figure 6D). The authors envisioned that their DNA origami robot could beused for simultaneous delivery of multiple therapeutics in a responsive manner,although this was not explicitly demonstrated in the work cited here.111
The loading of functional nucleic acids onto DNA origami structures is relativelystraightforward given the predictable Watson-Crick base pairing, such that tradi-tional nucleic acid hybridization can be utilized. Synthetic oligonucleotides contain-ing unmethylated cytosine-phosphate-guanosine (CpG)motifs are a potent activatorof innate and acquired immune responses and act primarily through Toll-like re-ceptor 9. The first report of CpG delivery using DNA origami structures was in
Chem 2, 359–382, March 9, 2017 375
Dox@Au-primer
A B C
D
Dox Conc. (µM)5 10 20 50 100
0
20
40
60
80
100
120
Cel
l Via
bilit
y (%
)
DoxDox/TubeDox/TriangleDox/ds M13
Unlocked
Locked
Locked
FITC:anti-HLAcargo
Cells Labeled
Cells Unlabeled
Cells Labeled FITC
FITC
FITCHLA+
HLA+
HLA+Key+
Front View Side View
TubeTriangle
Blank
GNR
DO-GNR
Blank GNR DO-GNRIncr
ease
of T
emp.
(°C
)
05
10152025
42.5 48.1 54.4
% o
f Max
0
20
40
60
80
100
0 103 104 105
CD69 on B cells
Unstim
CpG-H’
Tube
Tube CpG
Wild type
Dox Dox@Au Dox@Au-RCAs
Dox@3D
50
100
Cel
l Via
bilit
y (%
)
DO
X-CP
PD
OX-
CPP
@Au
DO
X-CP
PD
OX-
CPP
DO
X-CP
P@
3DD
OX-
CPP
CPP@
3D
E
E (AND)X
E
E
E
EE (AND) EP1P2(OR)
EEP1P2N(XOR)
EP1P2(OR)X
P1 P2
EP1P2N(XOR)X
EP1 P2
N
EP1 P2
NF
F E
YY
YX Y
EFP1P2N(half adder)
EN
E
P
EPDGF VEGF
0 01 00 11 1
103 104 105 106 103 104 105 106 103 104 105 106
Cou
nts
CockroachIn vivo
0
10
5
15
20
25
Cel
l Cy5
+ (%
)
DN
A 2 3 4 5 6N
o D
NA
Tumor Temp. (°C) Samples
Biodistribution and Bioavailability in Mice
HG
5 10 15 2000
20
40
60
80
100
120
Time (h)
Cum
ulat
ive
Rele
ase
(%)
Straight Twisted
Twisted
Straight
dsDNA
Free Dox
F
I+
ElectrostaticBinding
0
Figure 6. DNA Origami for Drug Delivery
(A) Triangular and tubular origami used for the delivery of doxorubicin show increased cytotoxicity. Reprinted with permission from Jiang et al.108
Copyright 2012 American Chemical Society.
(B) Tubular origami with tunable encapsulation and release properties for delivering doxorubicin. Reprinted with permission from Zhao et al.109
Copyright 2012 American Chemical Society.
(C) DNA origami hexagonal barrel for the delivery of therapeutic antibodies. Scale bars, 20 nm. From Douglas et al.110 Reprinted with permission from
AAAS.
(D) Rectangular-origami-based logic gate for in vivo computing. Reprinted by permission fromMacmillan Publishers Ltd:Nature Nanotechnology (Amir
et al.111), copyright 2014.
(E) Tubular origami for the delivery of CpG oligonucleotides. Reprinted with permission from Schuller et al.112 Copyright 2011 American Chemical
Society.
(F) Triangular and tubular origami for the delivery of AuNRs show higher tumor accumulation and temperature increases under laser irradiation.
Reprinted with permission from Jiang et al.113 Copyright 2015 Wiley-VCH.
(G) Origami grown on AuNP for the delivery of doxorubicin. Reprinted with permission from Yan et al.114 Copyright 2015 Wiley-VCH.
(H) Rectangular origami encapsulated with viral proteins demonstrates enhanced cell internalization, as shown in the flow cytometry study. Reprinted
with permission from Mikkila et al.115 Copyright 2014 American Chemical Society.
(I) Origami octahedron enveloped by lipids shows enhanced in vivo bioavailability. Reprinted with permission from Perralt and Shih.116 Copyright 2014
American Chemical Society.
376 Chem 2, 359–382, March 9, 2017

2011 by Liedl and colleagues (Figure 6E).112 They designed a 30-helix tubularorigami structure with 62 CpG molecules decorated on the surface of the structure.CpG-carryingDNAorigami in spleen cells stimulated a significantly stronger immuneresponse than equal amounts of CpG delivered with Lipofectamine or without deliv-ery agents. Other functional nucleic acids, such as aptamers and small interferingRNAs, have been extensively used as cargo delivered by non-origami DNA struc-tures.106 Although, to the best of our knowledge, no study has used DNA origamito deliver these functional nucleic acids, we believe this is an area of active researchby several different groups.
Hybrid Systems with Multiple Functionalities
Another application of incorporating DNA origami structures with other materials isto achieve multiple functionalities. These materials include but are not limited toinorganic and organic nanomaterials, biomolecules, lipids, etc. Ding and colleaguesattached AuNRs onto triangular and tubular DNA origami structures to enable simul-taneous two-photon imaging and photothermal therapy after laser irradiation (Fig-ure 6F).113 In vitro cellular studies demonstrated that the DNA-origami-AuNR com-plex had significantly higher cell internalization efficiency and cytotoxicity thanAuNRs alone. In vivo, a higher accumulation of AuNRs at the tumor site and anincreased survival rate of mice bearing xenograft tumors were observed for the com-plex structure. Fan, Song, and colleagues tethered short DNA oligonucleotides ontoAuNPs, which they used as primers to initiate rolling circle amplification to generatea long, repetitive ssDNA protruding from the AuNP cores (Figure 6G).114 They usedthis long ssDNA as a scaffold to fabricate DNA origami structures around the AuNPcore. They also incorporated cell-penetrating peptides and doxorubicin into this 3Dsuperstructure to enhance cell internalization efficiency and to enable cytotoxicity,respectively. In vitro cellular experiments demonstrated that the as-fabricated 3Dsuperstructures had a high drug-loading capacity and enabled the efficient transportof signal reporters and drugs for cellular imaging and chemotherapy.
Improving DNA Origami’s Delivery Efficiency and Biostability
Although DNA origami structures can enter cells alone, their internalization effi-ciency and biostability need be further improved if they are to survive physiologicalenvironments. Inspired by viruses, several groups have found that the encapsulationof DNA origami nanostructures by other materials, such as viral proteins and lipids,can drastically increase cell internalization efficiency and biostability. Kostiainen andcolleagues used cowpea chlorotic mottle virus capsid proteins to coat and pack aplanar rectangular DNA origami into either a tubular or planar complex via electro-static interactions (Figure 6H).115 Compared with DNA origami alone, the encapsu-lated DNA origami showed a 13-fold increase in internalization in HEK293 cells.Confocal microscopy showed that the protein-encapsulated DNA origami was inclose proximity to the cell nucleus, suggesting that this complex could potentiallyhave nuclear uptake. Similarly, Shih and colleagues packaged a DNA origami octa-hedron into a PEGylated lipid bilayer to increase its in vitro and in vivo stability andbioavailability (Figure 6I).116 They demonstrated that the envelopment of DNAnanostructures into PEGylated lipids conferred protection against nuclease diges-tion, decreased immune activation by 2 orders of magnitude, and increased phar-macokinetic bioavailability by a factor of 17 in relation to that of non-encapsulatedDNA origami nanostructures.
Challenges and Perspectives
Cost is among the most challenging hurdles ahead of DNA origami’s practical applica-tions as a drug-delivery vehicle. Commercially available staple strands of a !7,000 bp
Chem 2, 359–382, March 9, 2017 377

origami structure at a synthesis scale of about 10 nmol typically cost several hundreddollars. Taking into account additional costs, such as the scaffoldDNA, oligonucleotidefunctionalization, and purification of origami, the actual cost per single DNA origamidesign would be even higher. Thus, economic productionmethods of scaffold and sta-ple DNA are urgently needed to lower this cost. In vivo production of DNA strands oreven entire origami structures could potentially solve this issue, given that simpleDNA tile structures have already been produced in vivo.119 In vitro and in vivo stabilityof DNA origami is another issue that needs to be addressed. Given the highly compactpacking of DNA duplexes in these nanostructures, high concentrations of cationic ions(e.g., Mg2+ and Na+) are required to neutralize the negative charge of the DNA back-bone and thus stabilize DNA origami structures. Typical physiological solutions (e.g.,PBS and medium) do not contain high enough concentrations of cationic ions to suffi-ciently stabilize the DNA origami structures. Many studies have been conducted totest the stability of DNA origami in solutions mimicking physiological conditions.They have found that less compact objects (e.g., wire-frame origami objects) aremore stable in cation-depleted solutions.16,17,120 In addition, proteins in the mediumcan nonspecifically coat the DNA origami structures to form a ‘‘protein corona’’ thatpotentially offers protection against structure dissociation and nuclease digestion.121
Further investigations are needed to fully elucidate the stability issue of DNA origamistructures. Although DNA origami structures have been experimentally demonstratedtoefficiently enter cells, the specific endocytosispathway remains tobeunveiled via sys-tematic mechanistic studies. Another challenging hurdle is that all studies have shownthat DNA origami structures are eventually transported to lysosomes for digestion.108
To enable efficient cargo release into the cytosol, lysosomal escape of DNAorigami ve-hiclesmight be necessary. Potential strategies includeusing targeting ligands to initiateuptake via a non-lysosomal pathway or conjugating functional molecules onto DNAorigami to facilitate lysosomal escape. A better understanding of the pharmacokineticsand pharmacodynamics of DNA origami structures in vivo also needs to be studiedbefore any clinical drug-delivery applications. Off-target issues are another challengethat needs to be addressed. The introduction of exogenous DNA has many potentialcomplications: activation of a potent immune response, sequence-specific interferencewith mRNAs or microRNAs leading to unwanted gene regulation, and long-term inte-gration into the genome. The chemical modification of the staple DNA strands, suchas via the introduction of modified phosphoramidites or post-synthetic modificationto render them biologically inert, could be one potential solution to these problems.
CONCLUDING REMARKS
In the 10 years since its inception, DNA origami has made remarkable strides towarda wide range of applications. In this review, we provide a relatively comprehensiveoverview of the advances in DNA origami technology and its broad applications inseparate sections on DNA origami design, nanofabrication, biosensing, and drugdelivery. In each section, we review and discuss representative works and concludewith challenges and perspectives from our point of view. Despite these manyexciting achievements, DNA-origami-based applications are still in their infancyand have a long way to go before they can be used for real-world problems. It isworth noting that the strengths and limitations of DNA origami relatively dependon specific applications. For instance, cost is one of the major concerns for drug de-livery, but it might not be as important for building an ultrasensitive biosensor plat-form that needs only trace amounts of DNA. Therefore, a case-by-case analysisshould be conducted when DNA origami is used for certain types of applications.And for some applications that do not need high structural complexity (e.g., intracel-lular RNA imaging), simple DNA tile structures could be utilized instead of DNA
378 Chem 2, 359–382, March 9, 2017

origami. Here, we summarize the major challenges that need to be addressed on ageneral basis before DNA origami can be applied broadly:
(1) Design: further simplified and automated design platforms need to be devel-oped, especially for researchers outside the DNA nanotechnology field.
(2) Cost: DNA is relatively expensive in comparison with other materials, sostraightforward and economic production methods are necessary.
(3) Scale up: the size of discrete origami structures is typically constrained within!100 nm because of the length of M13 scaffold DNA, and thus alternativestrategies need to be developed for size expansion.
(4) Chemical functionality: DNA is a relatively chemically inert biomolecule, andthus facile methods for adding a wide variety of functionalities needs furtherdevelopment. Current methods for introducing additional reactivity, throughthe introduction of alternative nucleotides during synthesis or post-syntheticmodifications, provide a good starting point but can be cost prohibitive.
(5) Stability: DNA is relatively vulnerable to damage caused by harsh fabricationenvironments or physiological conditions; thus, it requires enhanced stabilityto survive such environments.
(6) Immunogenicity: exogenous DNA has been shown to elicit potent immune re-sponses, and thus methods for minimizing these reactions are needed, pri-marily for in vivo applications.
(7) Defects: DNA origami structures contain assembly defects, which can hinderheteroelement or therapeutic incorporation. Optimizing structural designs(e.g., crossover pattern and staple length) and assembly conditions (e.g.,Mg2+ concentration and a thermal annealing protocol) can help to minimizestructural defects. Straightforward methods should also be developed toallow convenient examination of structural quality.
AUTHOR CONTRIBUTIONS
P.W. and T.A.M. investigated the literature and wrote and revised the manuscript.V.P. and P.K.D investigated the literature and wrote sections of the manuscript.Y.K. proposed the review topic, supervised the writing of the manuscript, and re-viewed the manuscript.
REFERENCES AND NOTES
1. Seeman, N.C. (2003). DNA in amaterial world.Nature 421, 427–431.
2. Stulz, E., and Clever, G.H., eds. (2015). DNA inSupramolecular Chemistry andNanotechnology (John Wiley and Sons, Inc.).
3. Stulz, E. (2012). DNA architectonics: towardsthe next generation of bio-inspired materials.Chemistry 18, 4456–4469.
4. Castro, C.E., Kilchherr, F., Kim, D.-N.,Shiao, E.L., Wauer, T., Wortmann, P., Bathe,M., and Dietz, H. (2011). A primer toscaffolded DNA origami. Nat. Methods 8,221–229.
5. Rothemund, P.W.K. (2006). Folding DNA tocreate nanoscale shapes and patterns. Nature440, 297–302.
6. Ke, Y., Sharma, J., Liu, M., Jahn, K., Liu, Y., andYan, H. (2009). Scaffolded DNA origami of aDNA tetrahedron molecular container. NanoLett. 9, 2445–2447.
7. Andersen, E.S., Dong, M., Nielsen, M.M.,Jahn, K., Subramani, R., Mamdouh,W., Golas,M.M., Sander, B., Stark, H., Oliveira, C.L., et al.(2009). Self-assembly of a nanoscale DNA boxwith a controllable lid. Nature 459, 73–76.
8. Douglas, S.M., Dietz, H., Liedl, T., Hogberg,B., Graf, F., and Shih, W.M. (2009). Self-assembly of DNA into nanoscale three-dimensional shapes. Nature 459, 414–418.
9. Douglas, S.M., Marblestone, A.H.,Teerapittayanon, S., Vazquez, A., Church,G.M., and Shih, W.M. (2009). Rapidprototyping of 3D DNA-origami shapes withcaDNAno. Nucleic Acids Res. 37, 5001–5006.
10. Ke, Y., Douglas, S.M., Liu, M., Sharma, J.,Cheng, A., Leung, A., Liu, Y., Shih, W.M., andYan, H. (2009). Multilayer DNA origamipacked on a square lattice. J. Am. Chem. Soc.131, 15903–15908.
11. Ke, Y., Voigt, N.V., Gothelf, K.V., and Shih,W.M. (2012). Multilayer DNA origami packed
on hexagonal and hybrid lattices. J. Am.Chem. Soc. 134, 1770–1774.
12. Dietz, H., Douglas, S.M., and Shih, W.M.(2009). Folding DNA into twisted and curvednanoscale shapes. Science 325, 725–730.
13. Han, D., Pal, S., Nangreave, J., Deng, Z., Liu,Y., and Yan, H. (2011). DNA origami withcomplex curvatures in three-dimensionalspace. Science 332, 342–346.
14. Han, D., Pal, S., Yang, Y., Jiang, S., Nangreave,J., Liu, Y., and Yan, H. (2013). DNA gridironnanostructures based on four-arm junctions.Science 339, 1412–1415.
15. Zhang, F., Jiang, S., Wu, S., Li, Y., Mao, C., Liu,Y., and Yan, H. (2015). Complex wireframeDNA origami nanostructures with multi-armjunction vertices. Nat. Nanotechnol. 10,779–784.
16. Veneziano, R., Ratanalert, S., Zhang, K.,Zhang, F., Yan, H., Chiu, W., and Bathe, M.(2016). Designer nanoscale DNA assemblies
Chem 2, 359–382, March 9, 2017 379

programmed from the top down. Science352, 1534.
17. Benson, E., Mohammed, A., Gardell, J.,Masich, S., Czeizler, E., Orponen, P., andHogberg, B. (2015). DNA rendering ofpolyhedral meshes at the nanoscale. Nature523, 441–444.
18. Zhao, Z., Liu, Y., and Yan, H. (2011).Organizing DNA origami tiles into largerstructures using preformed scaffold frames.Nano Lett. 11, 2997–3002.
19. Iinuma, R., Ke, Y., Jungmann, R.,Schlichthaerle, T., Woehrstein, J.B., and Yin,P. (2014). Polyhedra self-assembled fromDNA tripods and characterized with 3D DNA-PAINT. Science 344, 65–69.
20. Gerling, T., Wagenbauer, K.F., Neuner, A.M.,and Dietz, H. (2015). Dynamic DNA devicesand assemblies formed by shape-complementary, non-base pairing 3Dcomponents. Science 347, 1446–1452.
21. Liu, W., Zhong, H., Wang, R., and Seeman,N.C. (2011). Crystalline two-dimensionalDNA-origami arrays. Angew. Chem. Int. Ed.50, 264–267.
22. Wang, P., Gaitanaros, S., Lee, S., Bathe, M.,Shih, W.M., and Ke, Y. (2016). ProgrammingSelf-Assembly of DNA Origami HoneycombTwo-Dimensional Lattices and PlasmonicMetamaterials. J. Am. Chem. Soc. 138, 7733–7740.
23. Aghebat Rafat, A., Pirzer, T., Scheible, M.B.,Kostina, A., and Simmel, F.C. (2014). Surface-assisted large-scale ordering of DNA origamitiles. Angew. Chem. Int. Ed. 53, 7665–7668.
24. Marchi, A.N., Saaem, I., Vogen, B.N., Brown,S., and LaBean, T.H. (2014). Toward largerDNA origami. Nano Lett. 14, 5740–5747.
25. Zhang, H., Chao, J., Pan, D., Liu, H., Huang,Q., and Fan, C. (2012). Folding super-sizedDNA origami with scaffold strands from long-range PCR. Chem. Commun. (Camb.) 48,6405–6407.
26. Woo, S., and Rothemund, P.W.K. (2011).Programmable molecular recognition basedon the geometry of DNA nanostructures. Nat.Chem. 3, 620–627.
27. Woo, S., and Rothemund, P.W.K. (2014). Self-assembly of two-dimensional DNA origamilattices using cation-controlled surfacediffusion. Nat. Commun. 5, 4889.
28. Kocabey, S., Kempter, S., List, J., Xing, Y., Bae,W., Schiffels, D., Shih,W.M., Simmel, F.C., andLiedl, T. (2015). Membrane-assisted growth ofDNA origami nanostructure arrays. ACSNano9, 3530–3539.
29. Suzuki, Y., Endo, M., and Sugiyama, H. (2015).Lipid-bilayer-assisted two-dimensional self-assembly of DNA origami nanostructures.Nat. Commun. 6, 8052.
30. Guo, P. (2010). The emerging field of RNAnanotechnology. Nat. Nanotechnol. 5,833–842.
31. Grabow, W.W., and Jaeger, L. (2014). RNAself-assembly and RNA nanotechnology. Acc.Chem. Res. 47, 1871–1880.
32. Endo, M., Yamamoto, S., Tatsumi, K., Emura,T., Hidaka, K., and Sugiyama, H. (2013). RNA-templated DNA origami structures. Chem.Commun. (Camb.) 49, 2879–2881.
33. Wang, P., Ko, S.H., Tian, C., Hao, C., andMao,C. (2013). RNA-DNA hybrid origami: foldingof a long RNA single strand into complexnanostructures using short DNA helperstrands. Chem. Commun. (Camb.) 49, 5462–5464.
34. Wang, P., Tian, C., Li, X., and Mao, C. (2014).Assembly of barcode-like nucleic acidnanostructures. Small 10, 3923–3926.
35. Endo, M., Takeuchi, Y., Emura, T., Hidaka, K.,and Sugiyama, H. (2014). Preparation ofchemically modified RNA origaminanostructures. Chemistry 20, 15330–15333.
36. Geary, C., Rothemund, P.W.K., and Andersen,E.S. (2014). RNA nanostructures. A single-stranded architecture for cotranscriptionalfolding of RNA nanostructures. Science 345,799–804.
37. Ke, Y., Ong, L.L., Shih, W.M., and Yin, P.(2012). Three-dimensional structures self-assembled from DNA bricks. Science 338,1177–1183.
38. Wei, B., Dai, M., and Yin, P. (2012). Complexshapes self-assembled from single-strandedDNA tiles. Nature 485, 623–626.
39. Samanta, A., and Medintz, I.L. (2016).Nanoparticles and DNA - a powerful andgrowing functional combination inbionanotechnology. Nanoscale 8, 9037–9095.
40. Pal, S., Deng, Z., Ding, B., Yan, H., and Liu, Y.(2010). DNA-origami-directed self-assemblyof discrete silver-nanoparticle architectures.Angew. Chem. Int. Ed. 49, 2700–2704.
41. Liu, W., Halverson, J., Tian, Y., Tkachenko,A.V., and Gang, O. (2016). Self-organizedarchitectures from assorted DNA-framednanoparticles. Nat. Chem. 8, 867–873.
42. Kuzyk, A., Schreiber, R., Fan, Z., Pardatscher,G., Roller, E.-M., Hogele, A., Simmel, F.C.,Govorov, A.O., and Liedl, T. (2012). DNA-based self-assembly of chiral plasmonicnanostructures with tailored optical response.Nature 483, 311–314.
43. Acuna, G.P., Moller, F.M., Holzmeister, P.,Beater, S., Lalkens, B., and Tinnefeld, P.(2012). Fluorescence enhancement at dockingsites of DNA-directed self-assemblednanoantennas. Science 338, 506–510.
44. Samanta, A., Zhou, Y., Zou, S., Yan, H., andLiu, Y. (2014). Fluorescence quenching ofquantum dots by gold nanoparticles: apotential long range spectroscopic ruler.Nano Lett. 14, 5052–5057.
45. Hemmig, E.A., Creatore, C., Wunsch, B.,Hecker, L., Mair, P., Parker, M.A., Emmott, S.,Tinnefeld, P., Keyser, U.F., and Chin, A.W.(2016). Programming Light-HarvestingEfficiency Using DNA Origami. Nano Lett. 16,2369–2374.
46. Maune, H.T., Han, S.-P., Barish, R.D.,Bockrath, M., Iii, W.A.G., Rothemund, P.W.K.,and Winfree, E. (2010). Self-assembly ofcarbon nanotubes into two-dimensional
geometries using DNA origami templates.Nat. Nanotechnol. 5, 61–66.
47. Sun, W., Boulais, E., Hakobyan, Y., Wang,W.L., Guan, A., Bathe, M., and Yin, P. (2014).Casting inorganic structures with DNAmolds.Science 346, 1258361.
48. Yang, Y., Wang, J., Shigematsu, H., Xu, W.,Shih, W.M., Rothman, J.E., and Lin, C. (2016).Self-assembly of size-controlled liposomes onDNA nanotemplates. Nat. Chem. 8, 476–483.
49. Surwade, S.P., Zhou, F., Wei, B., Sun, W.,Powell, A., O’Donnell, C., Yin, P., and Liu, H.(2013). Nanoscale growth and patterning ofinorganic oxides using DNA nanostructuretemplates. J. Am. Chem. Soc. 135, 6778–6781.
50. Gopinath, A., Miyazono, E., Faraon, A., andRothemund, P.W.K. (2016). Engineering andmapping nanocavity emission via precisionplacement of DNA origami. Nature 535,401–405.
51. Liu, W., Tagawa, M., Xin, H.L., Wang, T.,Emamy, H., Li, H., Yager, K.G., Starr, F.W.,Tkachenko, A.V., and Gang, O. (2016).Diamond family of nanoparticle superlattices.Science 351, 582–586.
52. Tian, Y., Wang, T., Liu, W., Xin, H.L., Li, H., Ke,Y., Shih, W.M., and Gang, O. (2015).Prescribed nanoparticle cluster architecturesand low-dimensional arrays built usingoctahedral DNA origami frames. Nat.Nanotechnol. 10, 637–644.
53. Schreiber, R., Santiago, I., Ardavan, A., andTurberfield, A.J. (2016). Ordering GoldNanoparticles with DNA OrigamiNanoflowers. ACS Nano 10, 7303–7306.
54. Shen, C., Lan, X., Lu, X., Meyer, T.A., Ni, W.,Ke, Y., and Wang, Q. (2016). Site-SpecificSurface Functionalization of Gold NanorodsUsing DNA Origami Clamps. J. Am. Chem.Soc. 138, 1764–1767.
55. Zhang, Y., Chao, J., Liu, H., Wang, F., Su, S.,Liu, B., Zhang, L., Shi, J., Wang, L., Huang, W.,et al. (2016). Transfer of Two-DimensionalOligonucleotide Patterns ontoStereocontrolled Plasmonic Nanostructuresthrough DNA-Origami-BasedNanoimprinting Lithography. Angew. Chem.Int. Ed. 55, 8036–8040.
56. Edwardson, T.G.W., Lau, K.L., Bousmail, D.,Serpell, C.J., and Sleiman, H.F. (2016).Transfer of molecular recognition informationfrom DNA nanostructures to goldnanoparticles. Nat. Chem. 8, 162–170.
57. Schreiber, R., Luong, N., Fan, Z., Kuzyk, A.,Nickels, P.C., Zhang, T., Smith, D.M., Yurke,B., Kuang, W., Govorov, A.O., and Liedl, T.(2013). Chiral plasmonic DNA nanostructureswith switchable circular dichroism. Nat.Commun. 4, 2948.
58. Zhou, C., Duan, X., and Liu, N. (2015). Aplasmonic nanorod that walks on DNAorigami. Nat. Commun. 6, 8102.
59. Lan, X., Lu, X., Shen, C., Ke, Y., Ni, W., andWang, Q. (2015). Au nanorod helicalsuperstructures with designed chirality. J. Am.Chem. Soc. 137, 457–462.
60. Urban, M.J., Dutta, P.K., Wang, P., Duan, X.,Shen, X., Ding, B., Ke, Y., and Liu, N. (2016).
380 Chem 2, 359–382, March 9, 2017

Plasmonic Toroidal MetamoleculesAssembled by DNA Origami. J. Am. Chem.Soc. 138, 5495–5498.
61. Pal, S., Deng, Z., Wang, H., Zou, S., Liu, Y., andYan, H. (2011). DNA directed self-assembly ofanisotropic plasmonic nanostructures. J. Am.Chem. Soc. 133, 17606–17609.
62. Kuzyk, A., Schreiber, R., Zhang, H., Govorov,A.O., Liedl, T., and Liu, N. (2014).Reconfigurable 3D plasmonicmetamolecules. Nat. Mater. 13, 862–866.
63. Pal, S., Dutta, P., Wang, H.N., Deng, Z.T., Zou,S.L., Yan, H., and Liu, Y. (2013). QuantumEfficiency Modification of OrganicFluorophores Using Gold Nanoparticles onDNAOrigami Scaffolds. J. Phys. Chem. C 117,12735–12744.
64. Kuhler, P., Roller, E.-M., Schreiber, R., Liedl, T.,Lohmuller, T., and Feldmann, J. (2014).Plasmonic DNA-origami nanoantennas forsurface-enhanced Raman spectroscopy.Nano Lett. 14, 2914–2919.
65. Ko, S.H., Du, K., and Liddle, J.A. (2013).Quantum-dot fluorescence lifetimeengineering with DNA origami constructs.Angew. Chem. Int. Ed. 52, 1193–1197.
66. Knudsen, J.B., Liu, L., Bank Kodal, A.L.,Madsen, M., Li, Q., Song, J., Woehrstein, J.B.,Wickham, S.F.J., Strauss, M.T., Schueder, F.,et al. (2015). Routing of individual polymers indesigned patterns. Nat. Nanotechnol. 10,892–898.
67. Langecker, M., Arnaut, V., List, J., and Simmel,F.C. (2014). DNA nanostructures interactingwith lipid bilayer membranes. Acc. Chem.Res. 47, 1807–1815.
68. Diagne, C.T., Brun, C., Gasparutto, D., Baillin,X., and Tiron, R. (2016). DNAOrigamiMask forSub-Ten-Nanometer Lithography. ACS Nano10, 6458–6463.
69. Jin, Z., Sun, W., Ke, Y., Shih, C.J., Paulus,G.L.C., Hua Wang, Q., Mu, B., Yin, P., andStrano, M.S. (2013). Metallized DNAnanolithography for encoding andtransferring spatial information for graphenepatterning. Nat. Commun. 4, 1663.
70. Sacca, B., and Niemeyer, C.M. (2012). DNAorigami: the art of folding DNA. Angew.Chem. Int. Ed. 51, 58–66.
71. Fu, J., Liu, M., Liu, Y., Woodbury, N.W., andYan, H. (2012). Interenzyme substratediffusion for an enzyme cascade organized onspatially addressable DNA nanostructures.J. Am. Chem. Soc. 134, 5516–5519.
72. Derr, N.D., Goodman, B.S., Jungmann, R.,Leschziner, A.E., Shih, W.M., and Reck-Peterson, S.L. (2012). Tug-of-war in motorprotein ensembles revealed with aprogrammable DNA origami scaffold.Science 338, 662–665.
73. Shaw, A., Lundin, V., Petrova, E., Ford}os, F.,Benson, E., Al-Amin, A., Herland, A., Blokzijl,A., Hogberg, B., and Teixeira, A.I. (2014).Spatial control of membrane receptorfunction using ligand nanocalipers. Nat.Methods 11, 841–846.
74. Ke, Y., Meyer, T., Shih, W.M., and Bellot, G.(2016). Regulation at a distance of
biomolecular interactions using a DNAorigami nanoactuator. Nat. Commun. 7,10935.
75. Kilchherr, F., Wachauf, C., Pelz, B., Rief, M.,Zacharias, M., and Dietz, H. (2016). Single-molecule dissection of stacking forces inDNA. Science 353, 1116.
76. Nickels, P.C., Wunsch, B., Holzmeister, P.,Bae, W., Kneer, L.M., Grohmann, D.,Tinnefeld, P., and Liedl, T. (2016). Molecularforce spectroscopy with a DNA origami-based nanoscopic force clamp. Science 354,305–307.
77. Jungmann, R., Avendano, M.S., Woehrstein,J.B., Dai, M., Shih, W.M., and Yin, P. (2014).Multiplexed 3D cellular super-resolutionimaging with DNA-PAINT and Exchange-PAINT. Nat. Methods 11, 313–318.
78. Dai, M., Jungmann, R., and Yin, P. (2016).Optical imaging of individual biomolecules indensely packed clusters. Nat. Nanotechnol.11, 798–807.
79. Fu, J., Yang, Y.R., Johnson-Buck, A., Liu, M.,Liu, Y., Walter, N.G., Woodbury, N.W., andYan, H. (2014). Multi-enzyme complexes onDNA scaffolds capable of substratechannelling with an artificial swinging arm.Nat. Nanotechnol. 9, 531–536.
80. Zhao, Z., Fu, J., Dhakal, S., Johnson-Buck, A.,Liu, M., Zhang, T., Woodbury, N.W., Liu, Y.,Walter, N.G., and Yan, H. (2016). Nanocagedenzymes with enhanced catalytic activity andincreased stability against proteasedigestion. Nat. Commun. 7, 10619.
81. Jungmann, R., Steinhauer, C., Scheible, M.,Kuzyk, A., Tinnefeld, P., and Simmel, F.C.(2010). Single-molecule kinetics and super-resolution microscopy by fluorescenceimaging of transient binding on DNAorigami.Nano Lett. 10, 4756–4761.
82. Jungmann, R., Avendano, M.S., Dai, M.,Woehrstein, J.B., Agasti, S.S., Feiger, Z.,Rodal, A., and Yin, P. (2016). Quantitativesuper-resolution imaging with qPAINT. Nat.Methods 13, 439–442.
83. Song, C., Wang, Z.-G., and Ding, B. (2013).Smart nanomachines based on DNA self-assembly. Small 9, 2382–2392.
84. Dong, Y., Yang, Z., and Liu, D. (2014). DNAnanotechnology based on i-motif structures.Acc. Chem. Res. 47, 1853–1860.
85. Wang, F., Liu, X., and Willner, I. (2015). DNAswitches: from principles to applications.Angew. Chem. Int. Ed. 54, 1098–1129.
86. Ke, Y., Lindsay, S., Chang, Y., Liu, Y., and Yan,H. (2008). Self-assembled water-solublenucleic acid probe tiles for label-free RNAhybridization assays. Science 319, 180–183.
87. Subramanian, H.K.K., Chakraborty, B., Sha, R.,and Seeman, N.C. (2011). The label-freeunambiguous detection and symbolic displayof single nucleotide polymorphisms on DNAorigami. Nano Lett. 11, 910–913.
88. Kuzuya, A., Sakai, Y., Yamazaki, T., Xu, Y., andKomiyama, M. (2011). Nanomechanical DNAorigami ‘single-molecule beacons’ directlyimaged by atomic force microscopy. Nat.Commun. 2, 449.
89. Koirala, D., Shrestha, P., Emura, T., Hidaka,K., Mandal, S., Endo, M., Sugiyama, H., andMao, H. (2014). Single-moleculemechanochemical sensing using DNAorigami nanostructures. Angew. Chem. Int.Ed. 53, 8137–8141.
90. Bell, N.A.W., Engst, C.R., Ablay, M., Divitini,G., Ducati, C., Liedl, T., and Keyser, U.F.(2012). DNA origami nanopores. Nano Lett.12, 512–517.
91. Hernandez-Ainsa, S., Bell, N.A.W., Thacker,V.V., Gopfrich, K., Misiunas, K., Fuentes-Perez,M.E., Moreno-Herrero, F., and Keyser, U.F.(2013). DNA origami nanopores forcontrolling DNA translocation. ACS Nano 7,6024–6030.
92. Langecker, M., Arnaut, V., Martin, T.G., List, J.,Renner, S., Mayer, M., Dietz, H., and Simmel,F.C. (2012). Synthetic lipid membranechannels formed by designed DNAnanostructures. Science 338, 932–936.
93. Krishnan, S., Ziegler, D., Arnaut, V., Martin,T.G., Kapsner, K., Henneberg, K., Bausch,A.R., Dietz, H., and Simmel, F.C. (2016).Molecular transport through large-diameterDNA nanopores. Nat. Commun. 7, 12787.
94. Zhang, Z., Wang, Y., Fan, C., Li, C., Li, Y., Qian,L., Fu, Y., Shi, Y., Hu, J., and He, L. (2010).Asymmetric DNA origami for spatiallyaddressable and index-free solution-phaseDNA chips. Adv. Mater. 22, 2672–2675.
95. Zhang, Z., Zeng, D., Ma, H., Feng, G., Hu, J.,He, L., Li, C., and Fan, C. (2010). A DNA-Origami chip platform for label-free SNPgenotyping using toehold-mediated stranddisplacement. Small 6, 1854–1858.
96. Kuzuya, A., Watanabe, R., Yamanaka, Y.,Tamaki, T., Kaino, M., and Ohya, Y. (2014).Nanomechanical DNA origami pH sensors.Sensors (Basel) 14, 19329–19335.
97. Godonoga, M., Lin, T.-Y., Oshima, A.,Sumitomo, K., Tang, M.S.L., Cheung, Y.-W.,Kinghorn, A.B., Dirkzwager, R.M., Zhou, C.,Kuzuya, A., et al. (2016). A DNA aptamerrecognising a malaria protein biomarkercan function as part of a DNA origamiassembly. Sci. Rep. 6, 21266.
98. Rinker, S., Ke, Y., Liu, Y., Chhabra, R., and Yan,H. (2008). Self-assembled DNAnanostructures for distance-dependentmultivalent ligand-protein binding. Nat.Nanotechnol. 3, 418–422.
99. Endo, M., and Sugiyama, H. (2014). Single-molecule imaging of dynamic motions ofbiomolecules in DNA origaminanostructures using high-speed atomicforce microscopy. Acc. Chem. Res. 47,1645–1653.
100. Bell, N.A.W., and Keyser, U.F. (2014).Nanopores formed by DNA origami: a review.FEBS Lett. 588, 3564–3570.
101. Venkatesan, B.M., and Bashir, R. (2011).Nanopore sensors for nucleic acid analysis.Nat. Nanotechnol. 6, 615–624.
102. Dekker, C. (2007). Solid-state nanopores. Nat.Nanotechnol. 2, 209–215.
103. Shen, W., Zhong, H., Neff, D., and Norton,M.L. (2009). NTA directed protein
Chem 2, 359–382, March 9, 2017 381

nanopatterning on DNA Origaminanoconstructs. J. Am. Chem. Soc. 131, 6660–6661.
104. Rosen, C.B., Kodal, A.L.B., Nielsen, J.S.,Schaffert, D.H., Scavenius, C., Okholm, A.H.,Voigt, N.V., Enghild, J.J., Kjems, J., Tørring, T.,and Gothelf, K.V. (2014). Template-directedcovalent conjugation of DNA to nativeantibodies, transferrin and other metal-binding proteins. Nat. Chem. 6, 804–809.
105. Yoo, J., and Aksimentiev, A. (2013). In situstructure and dynamics of DNA origamidetermined through molecular dynamicssimulations. Proc. Natl. Acad. Sci. USA 110,20099–20104.
106. Li, J., Fan, C., Pei, H., Shi, J., and Huang, Q.(2013). Smart drug delivery nanocarriers withself-assembled DNA nanostructures. Adv.Mater. 25, 4386–4396.
107. Chen, Y.-J., Groves, B., Muscat, R.A., andSeelig, G. (2015). DNA nanotechnology fromthe test tube to the cell. Nat. Nanotechnol. 10,748–760.
108. Jiang, Q., Song, C., Nangreave, J., Liu, X., Lin,L., Qiu, D., Wang, Z.-G., Zou, G., Liang, X.,Yan, H., and Ding, B. (2012). DNA origami as acarrier for circumvention of drug resistance.J. Am. Chem. Soc. 134, 13396–13403.
109. Zhao, Y.-X., Shaw, A., Zeng, X., Benson, E.,Nystrom, A.M., and Hogberg, B. (2012). DNAorigami delivery system for cancer therapy
with tunable release properties. ACS Nano 6,8684–8691.
110. Douglas, S.M., Bachelet, I., and Church, G.M.(2012). A logic-gated nanorobot for targetedtransport of molecular payloads. Science 335,831–834.
111. Amir, Y., Ben-Ishay, E., Levner, D., Ittah, S.,Abu-Horowitz, A., and Bachelet, I. (2014).Universal computing by DNA origami robotsin a living animal. Nat. Nanotechnol. 9,353–357.
112. Schuller, V.J., Heidegger, S., Sandholzer, N.,Nickels, P.C., Suhartha, N.A., Endres, S.,Bourquin, C., and Liedl, T. (2011). Cellularimmunostimulation by CpG-sequence-coated DNA origami structures. ACS Nano 5,9696–9702.
113. Jiang, Q., Shi, Y., Zhang, Q., Li, N., Zhan, P.,Song, L., Dai, L., Tian, J., Du, Y., Cheng, Z., andDing, B. (2015). A Self-Assembled DNAOrigami-Gold Nanorod Complex for CancerTheranostics. Small 11, 5134–5141.
114. Yan, J., Hu, C., Wang, P., Zhao, B., Ouyang, X.,Zhou, J., Liu, R., He, D., Fan, C., and Song, S.(2015). Growth and origami folding of DNA onnanoparticles for high-efficiency moleculartransport in cellular imaging and drugdelivery. Angew. Chem. Int. Ed. 54, 2431–2435.
115. Mikkila, J., Eskelinen, A.-P., Niemela, E.H.,Linko, V., Frilander, M.J., Torma, P., andKostiainen, M.A. (2014). Virus-encapsulated
DNA origami nanostructures for cellulardelivery. Nano Lett. 14, 2196–2200.
116. Perrault, S.D., and Shih, W.M. (2014). Virus-inspired membrane encapsulation of DNAnanostructures to achieve in vivo stability.ACS Nano 8, 5132–5140.
117. Zhang, Q., Jiang, Q., Li, N., Dai, L., Liu, Q.,Song, L., Wang, J., Li, Y., Tian, J., Ding, B., andDu, Y. (2014). DNA origami as an in vivo drugdelivery vehicle for cancer therapy. ACS Nano8, 6633–6643.
118. Halley, P.D., Lucas, C.R., McWilliams, E.M.,Webber, M.J., Patton, R.A., Kural, C., Lucas,D.M., Byrd, J.C., and Castro, C.E. (2016).Daunorubicin-Loaded DNA OrigamiNanostructures Circumvent Drug-ResistanceMechanisms in a Leukemia Model. Small 12,308–320.
119. Elbaz, J., Yin, P., and Voigt, C.A. (2016).Genetic encoding of DNA nanostructures andtheir self-assembly in living bacteria. Nat.Commun. 7, 11179.
120. Hahn, J., Wickham, S.F.J., Shih, W.M., andPerrault, S.D. (2014). Addressing the instabilityof DNA nanostructures in tissue culture. ACSNano 8, 8765–8775.
121. Giljohann, D.A., Seferos, D.S., Patel, P.C.,Millstone, J.E., Rosi, N.L., and Mirkin, C.A.(2007). Oligonucleotide loading determinescellular uptake of DNA-modified goldnanoparticles. Nano Lett. 7, 3818–3821.
382 Chem 2, 359–382, March 9, 2017


















