
The Auditory System - UConn Health · The topotopic organization is present along the auditory...
26
The human auditory system Auditory cortex Medial geniculate body (Thalamus) Inferior colliculus Lateral lemniscus Superior olive Spiral ganglion Cochlea Ventral cochlear nucleus Dorsal cochlear nucleus (Adapted from Bear et al)
Transcript of The Auditory System - UConn Health · The topotopic organization is present along the auditory...

The human auditory system
Auditory cortex
Medial geniculate body (Thalamus)
Inferior colliculus
Lateral lemniscus
Superior olive
Spiral ganglion
Cochlea
Ventral cochlear nucleus
Dorsal cochlear nucleus
(Adapted from Bear et al)
Moderador
Notas de la presentación
The human auditory system This slide outlines the auditory pathway in humans. The transverse sections in the main part of the figure are taken at the levels indicated by the numbers on the small brain diagram to the left of the slide. The pathway (from one ear only) is outlined showing the key processing centres in the auditory system at different levels of the brainstem, thalamus and cortex. The thalamus receives its main projection from the inferior colliculus (see previous lecture). The auditory region of the thalamus is called the medial geniculate body (MGB) and this in turn is the source of the main projection to the auditory cortex. The auditory cortex is located in the temporal lobe of the brain. In humans the primary auditory cortex (the first cortical region that receives the main thalamic input) is known as A1 and corresponds to Heschl’s gyrus. Other subdivisions of the auditory cortex are found in front and behind this region including the planum temporale or temporal plane. El sistema auditivo humano Esta diapositiva ilustra un esquema de la vía auditiva en humanos. Las secciones coronales en la parte principal de la diapositiva se han tomado de los niveles indicados con números en el diagrama sagital pequeño del cerebro situado en la parte izquierda de la diapositiva. El esquema de la vía (originada de un solo oído) muestra los centros de procesamiento claves o principales del sistema auditivo a nivel del tronco del encéfalo, tálamo y corteza. El núcleo talámico auditivo es el cuerpo geniculado medial (CGM), que recibe sus proyecciones ascendentes principalmente desde el colículo inferior (ver lecciones previas) y a su vez envia sus proyecciones a la corteza auditiva. La corteza auditiva está lozalizada en el lóbulo temporal del cerebro. En humanos la corteza auditiva primaria (la primera región que recibe impulsos desde el tálamo) se conoce como A1 y la primera circunvolución transversa (de Heschl). Por delante y detrás de la corteza A1 podemos encontrar regiones como las del plano temporal.

The topotopic organization is present along the auditory system
Moderador
Notas de la presentación
The topotopic organization is present along the auditory system As discussed in previous lectures, a tonotopic representation of sound frequency is a key feature in the organisation of the auditory pathway as shown here for the cat. A particular region of the cochlea (lower left) is represented topographically (as shown in red) in each structure of the auditory pathway as shown in the other examples which show some of the centres in the auditory pathway. This is also the case in the medial geniculate body (top right), the primary auditory cortex (A1, top left) and in the neighbouring anterior auditory field immediately anterior to A1. En todos las niveles de la sistema auditivo existe una organización tonotópica Como se ha visto anteriormente, existe una representación tonotópica de la frecuencias del sonido y ello es una característica fundamental en la organización de la vía auditiva, como se ilustra en esta imagen que es un esquema de la vía auditiva del gato. Una zona concreta de la cóclea (parte inferior izquierda) está representada topográficamente (marcado en rojo) en cada una de las estructuras o núcleos de la vía auditiva. Lógicamente esto ocurre también en la corteza auditiva primaria (parte superior izquierda) y en los campos auditivos anteriores adyacentes a la corteza auditiva primaria.

The organisation and function of the auditory thalamus
Moderador
Notas de la presentación
The organisation and function of the auditory thalamus The thalamus is an important centre that is sometimes termed the gateway to the cortex. Recent studies have demonstrated that the auditory thalamus also makes important non cortical connections. Organización y función del tálamo auditivo El tálamo es un centro importante que a veces se llama la puerta de entrada a la corteza. Los estudios recientes han demostrado que el tálamo auditivo también tiene conexiones importantes no corticales.

From Bear Connors and Paradiso: Neuroscience, Exploring the brain
Moderador
Notas de la presentación
The thalamus is a collection of nuclei in the centre of the brain through which all sensory information (except the olfactory system) passes on route to the cortex. The slide shows a midsaggital section through the human brain to reveal the medial (inner surface). The thalamus is labelled and it can be seen in the context of other brain structures. This region of the brain, together with the hypothalamus below it, constitute the diencephalon. ______________- El tálamo es un grupo de núcleos localizado en el centro del cerebro a través del cual pasa y hace sinapsis toda la información sensorial (excepto la del sistema olfativo) en la ruta a la corteza. Esta diapositiva muestra una sección sagital a través del plano medial del cerebro humano que permite visualizar la superficie interna del cerebro. Podemos ver el tálamo en relación con otras estructuras del cerebro. Esta región del cerebro, junto al hipotálamo localizado debajo de él, constituye el diencéfalo.

Medial geniculate body
Moderador
Notas de la presentación
The thalamus is here seen on the left in transverse section taken at the level of the inset drawing (upper left) The drawing on the right shows the shape of the thalamus, and the position of the auditory subdivision of the thalamus (the medial geniculate body) is marked. _____________ Aquí podemos ver el tálamo en la sección transversal de la izquierda tomada en el nivel 2 del dibujo insertado en la parte la izquierda superior. El dibujo a la derecha muestra la forma del tálamo, y la localización de la subdivisión auditiva del mismo (el cuerpo geniculado medial) que está marcada con un círculo.



MGV
MGD
MGM
Thalamic reticular nucleus
IC (central)
IC (dorsal cortex)
IC (external cortex)
Superior Colliculus
VNLL
DNLL
Cochlear nucleus
Spinal cord
La corteza
Po
V
D
M L
Moderador
Notas de la presentación
The medial geniculate body (MGB) can be divided into three subdivisions based on cytoarchitecture the inputs it receives from other brain structures and the targets of its outputs The three divisions of the MGB are represented schematically in this slide: the ventral nucleus (MGV) dorsal nucleus (MGD) and the medial nucleus (MGM). Other thalamic nuclei which also have an auditory component are the posterior group (Po) and the reticular nucleus of the thalamus. These divisions have also been subdivided but for simplicity they are not discussed here. The ventral nucleus receives its input from central nucleus of the inferior colliculus and is therefore part of the core, lemniscal ascending pathway of the auditory system. The main input is from the ipsilateral IC although there is also a smaller contralateral projection. The dorsal and medial nuclei receive inputs predominantly from the cortices of the inferior colliculus, while the medial nucleus also receives inputs from the superior colliculus, ventral and dorsal nuclei of the lateral lemniscus (VNLL and DNLL) the cochlear nucleus complex, the trigeminal nuclei, vestibular system and spinal cord. Thus the MGM integrates inputs from diverse sources and systems. All MGB divisions also receive descending connections from the auditory cortex. ____________________- El cuerpo geniculado medial (MGB) se puede dividir en tres subdivisiones en función de su citoarquitectura, las entradas aferentes que recibe de otras estructuras del cerebro y de las dianas de sus salidas eferentes. En esta diapositiva se representan esquemáticamente las tres divisiones del MGB : la división o núcleo ventral (MGV), la división o núcleo dorsal (MGD) y la división o el núcleo medial (MGM). Otros núcleos talámicosc que también tienen un componente auditivo son el grupo posterior (Po) y el núcleo reticular del tálamo. Estas otros componentes talámicos también poseen subdivisiones adicionales pero por brevedad y para simplificar no las trataremos aquí. El núcleo ventral recibe sus aferencias del núcleo central del colículo inferior y por lo tanto forma parte de la vía ascendente lemniscal ‘principal’ del sistema auditivo. La entrada principal procede del IC ipsilateral aunque hay también una proyección contralateral más pequeña. Los núcleos dorsales e medial reciben sus aferencias predominantes desde las cortezas del colículo inferior, mientras que el núcleo medial también recibe entradas del colículo superior, asi como desde los núcleos ventral y dorsal del lemniscus lateral (VNLL y DNLL), de los núcleos cocleares, los núcleos del trigeminal, el sistema vestibular y la médula espinal. Así pues, el MGM integra aferencias de sistemas diversos y vías. Todas las divisiones del MGB también reciben conexiones descendentes de la corteza auditiva.

MGV
MGD
MGM
Thalamic reticular nucleus
Cortex: core
Cortex: belt & (core)
Cortex: core & belt
Lateral Amygdala
Outputs of the subdivisions of the medial geniculate body
Moderador
Notas de la presentación
Outputs of the subdivisions of the medial geniculate body The subdivisions of the MGB also differ in the organisation of their outputs. In the macaque monkey the predominant output of MGV, consistent with it being the main lemniscal component of the pathway, is to the core regions of the auditory cortex (see later for definition), while the MGD projects mainly to the belt regions of the cortex (although it has a weaker projection to the core). In contrast, the MGM subdivision which receives inputs from a wide range of brainstem nuclei projects widely to the auditory cortex targeting both the core and belt regions. In addition, in the rat and guinea pig (and probably in other species) the medial subdivision sends a projection to the lateral amygdala, part of the limbic system - a region of the brain concerned with emotions, and to the frontal and somatic cortices. It is suggested that this constitutes part of the pathway which governs our emotional responses to sounds. The finding of a direct projection from the cochlear nucleus to the medial subdivision is evidence for a pathway that give acoustic information fast access to the emotional centres of the brain (see later) All three subdivisions make connections via axon collaterals with the inhibitory thalamic reticular nucleus, although the role of this nucleus is not clear. Proyecciones del cuerpo geniculado medial Las subdivisiones del MGB también se diferencian por la organización de sus proyecciones eferentes. En el mono macaco, la proyección predominante del MGV, es hacia las regiones principales de la corteza auditiva (véase más adelante para su definición), lo cual apoya que forma parte del componente lemniscal principal de la vía auditiva, mientras que el MGD proyecta principalmente a las regiones adyacentes la corteza primaria (aunque tiene una proyección más débil a la parte principal). En cambio, la subdivisión MGM que recibe una amplia gama de proyecciones desde distintos núcleos del tronco del encéfalo, también proyecta extensamente a la corteza auditiva principal y adyacente. Además, en la rata y el cobayo, (y probablemente en la otras especies) la subdivisión MGM envía una proyección a la amigdala lateral, que forma parte del sistema límbico (una región del cerebro relacionada con las emociones) y a las cortezas frontales y somáticas. Se ha sugerido que el MGM forma parte de la vía que gobierna nuestras respuestas emocionales a los sonidos. Recientemente se ha encontrado una proyección directa desde los núcleos cocleares a la subdivisión MGM lo cual es una evidencia clara de una vía de transmisión de información acústica rápida a los centros emocionales del cerebro (véase más adelante) Las tres subdivisiones del MGB tienen conexiones a través de colaterales con el núcleo reticular talámico que es inhibitorio, aunque su papel funcional no está claro.

Ascending Auditory pathways
IC
CORTEX
MGB
IT AI AII
M V D
LC DCIC-CEN
SC - TegmentumFigure 110.6
Moderador
Notas de la presentación
The main ascending outputs of the inferior colliculus are shown. The largest single ascending output is the projection from the central nucleus of the inferior colliculus (IC-CEN) to the ventral division (V) of the medial geniculate body (MGB) in the diencephalon. As mentioned in the last slide, this projection consists of both glutamatergic and GABAergic axons. The dorsal cortex (DC) projects primarily to the dorsal division (D) of the medial geniculate body that also receives inputs from the superior colliculus (SC) and midbrain tegmentum. The lateral cortex projects primarily to the medial division (M) of the medial geniculate body that also receives inputs from the central nucleus and dorsal cortex. The pathways from the IC-CEN, LC, and DC to the MGB all follow the brachium of the inferior colliculus (see slide 3). The central nucleus and dorsal cortex project to the IC on the contralateral side. The central nucleus also projects to the medial geniculate body on the contralateral side. Aquí se muestran Las proyecciones ascendentes principales del colliculus inferior. La única proyección más grande ascendente es la proyección que se origina en el núcleo central del colliculus inferior (IC-CEN) y termina en la división ventral (V) del cuerpo geniculado medial (MGB) en el diencéfalo. Como deciamos en la diapositiva anterior, esta proyección esta hecha a base de glutamato y de axones GABAergicos. La corteza dorsal (DC) proyecta principalmente a la división dorsal (D) del MGB que recibe también las entradas del colliculus superior (SC) y desde el tegmentum (en el mesencéfalo). La corteza lateral proyecta principalmente a la división medial (M) del MGB que recibe también las proyecciones desde del núcleo central y corteza dorsal del IC. Todos los axones del IC-CEN, de LC, y de DC al MGB lo hacen a través del brachium del colliculus inferior (ver diapositiva 3). El núcleo central y la corteza dorsal proyectan al IC del lado de contralateral. El núcleo central proyecta también al MGB del lado contralateral. From: Oliver DL (1997) Anatomy of the central auditory nervous system. Part XI. Chapter 110. In: Encyclopedia of Acoustics (Crocker MJ, ed), pp 1381-1388. New York: John Wiley & Sons, Inc.

GABA IC projections to MGB
• GABA in the neuron (green)
• Retrograde transport of red latex microspheres (red beads)
Moderador
Notas de la presentación
GABA IC projections to MGB This figure shows both excitatory and inhibitory outputs from the IC and their effects on the medial geniculate body in the thalamus. A-B (right)show the GABA neurons in the IC (filled circles) and the glutamatergic neurons (open squares and triangles). IC neurons that project to the medial geniculate body in the thalamus are shown in red in the image on the left. They are labeled by retrograde transport with latex microspheres. The GABA-ergic IC neuron is labeled green with an antibody to GABA. The white overlay shows a neuron from the MGB filled in a brain slice after recording and the recording of the neurons response after stimulation of the brachium of the inferior colliculus. The GABAergic input influences the early and late parts of the response, while the main excitation follows the early GABA response. Thus, GABA from the IC regulates the beginning of the excitatory response in the MGB. Proyección GABAergica al MGB Esta figura muestra las proyecciones excitantorias e inhibitorias del IC y sus efectos en el cuerpo geniculado medial en el tálamo. A-B ( derecha) muestra las neuronas GABAergicas en el IC (círculos rellenos) y las neuronas de glutamatergicas (los cuadrados abiertos y los triángulos). Las neuronas de IC que proyectan al cuerpo geniculado medial en el tálamo se muestran en rojo en la imagen en la izquierda. Estas neuronas se pueden visualizar porque se han marcada por el transporte retrógrado con microesferas de látex tras una inyección en el MGB. Además, las neuronas GABAergicas del IC se marcan en verde con un anticuerpo para GABA. La neurona blanca muestra una neurona del MGB rellena en una rodaja de cerebro después de registrar y el trazo de su registro después de estimular del brachium del colliculus inferior. La proyección GABAérgica influye las porciones temprana y tardia de la respuesta, mientras la excitación principal sigue la respuesta temprana de GABA. Así pues la proyección de l GABA del IC regula el inicio de la respuesta excitante en el MGB. From: Peruzzi D, Bartlett E, Smith PH, Oliver DL (1997) A monosynaptic GABAergic input from the inferior colliculus to the medial geniculate body in rat. J Neurosci 17:3766-3777.


Representation of sound frequency in the ventral MGB and lateral posterior group (PO)
Imig and Morel 1985
Moderador
Notas de la presentación
Sound frequency is represented tonotopically in the ventral subdivision. The left panel shows the best frequencies of recordings made at points along three electrode tracks crossing the ventral division as seen in the transverse plane. When data from several such experiments covering different parts of the ventral subdivision are reconstructed its frequency organisation is found to be highly complex as the model on the right shows. The representation of high middle and low frequencies are identified by the hatching on the interlocking shapes. The frequency representation in the posterior group is also shown. Neurons in the ventral division are most well tuned to frequency (although some are broadly tuned and have more than one peak) and their latency of firing in response to sound stimuli is short. Many have onset rather than sustained responses to pure tones. About 90% of the neurons are responsive to binaural stimuli, showing either EE or EI properties as reported at more peripheral levels of the auditory pathway, e.g.,SOC. In contrast, the dorsal subdivision (MGD) is non tonotopic and many of the neurons respond best to complex stimuli and respond poorly to pure tones. Long response latencies are also characteristic of many neurons in this the dorsal subdivision. Neurons in the deeper regions of the dorsal subdivision however tend to respond most like those in the ventral division. The medial division (MGM) is tonotopically organised, albeit more weakly than MGV. Neurons with narrow or broad tuning to pure tones are intermixed. _______________________ La frecuencia del sonido se representa tonotopicamente en la subdivisión ventral. El panel izquierdo muestra las mejores frecuencias de los registros realizados en puntos a lo largo de tres tractos de electrodos que cruzan la división ventral según se puede observar en el plano transversal. Cuando los datos de varios de estos experimentos que cubren diversas partes de la subdivisión ventral se reconstruyen podemos observar que su organización tonotopica es altamente compleja como se puede ver en el modelo que se muestra a la derecha. La representación de frecuencias altas, medias y bajas se identifica por formas con distintas tramas que se interdigitizan. También se muestra la representación de la frecuencia en el grupo posterior. Las neuronas de la división ventral están mejor sintonizadas a la frecuencia (aunque algunos se sintonizan con respuestas muy amplias en frecuencia o con más de un pico) y su latencia a los estímulos de los sonidos es corta. Hay muchas más neuronas que poseen respuestas de inicio que respuestas sostenidas a los tonos puros. El 90% de las neuronas responden a los estímulos binauriculares, mostrando características EE o EI según lo expuesto niveles más periféricos de la vía auditivo, como por ejemplo en el SOC. En cambio, la subdivisión dorsal (MGD) no posee una organización tonotópica y muchas de las neuronas responden mejor a estímulos complejos que a tonos puros. Muchas neuronas en esta subdivisión dorsal poseen latencias largas. Sin embargo, las neuronas en las regiones más profundas de la subdivisión dorsal tienden a responder como las den la división ventral. La división medial (MGM) se organiza tonotopicamente, no obstante está organización es menos clara que en MGV. Las neuronas con una sintonización fina y amplia a los tonos puros se entremezclan.

Anderson et al 2006 E J Neurosci
Latency of firing to clicks in different divisions of MGB
Moderador
Notas de la presentación
Latency of firing to clicks in different divisions of MGB The median latency of firing of neurons varies across the different divisions of the MGB. The shortest latencies are shown by neurons of MGM. Some neurons in MGM phase-lock to click-rates as high as 1000 Hz, a higher rate than is seen in the other subdivisions. Both these findings are consistent with the notion that MGM receives a direct input from an earlier stage in the auditory pathway than the other two divisions. It seems likely that these properties reflect the direct input that MGM receives from the cochlear nucleus. Latencia de la descarga en respuesta a clicks en diversas divisiones de MGB La latencia media de la descarga varía entre las diversas divisiones del MGB. Las neuronas de MGM tienen la latencia más cortas. Algunas neuronas de MGM muestran sincronía de fases a clicks presentados con una frecuencia de hasta 1000 Hz, una tasa más alta de lo que se puede ver en las otras subdivisiones. Estos resultados confirman que MGM recibe una entrada directa de un núcleo auditivo inferior o anterior al colículo inferior a diferencia de las otras dos divisiones. Parece probablemente que estas características reflejan la proyección directa que recibe el MGM del núcleo coclear.

Conditioned stimulus -tone
Auditory cortex
Amygdala
Lateral
CentralMGVMGM
Defence responses
Hypothalamus / pituitry
Autonomic nervous system
Behaviour
Circuits for auditory fear conditioning
Moderador
Notas de la presentación
Circuits for auditory fear conditioning This fast route to MGM could be important in the role it plays in the emotional responses to sound via its outputs to the limbic system. The MGM is known to be important for mediating fear condition to sounds. This behaviour is abolished by destruction of MGM. Cells in the MGM also increase their firing to an auditory conditioned stimulus following fear conditioning to that stimulus and show long term potentiation. MGM neurons also show plasticity whereby the neuron’s frequency selectivity adjusts to that of the conditioned stimulus. Experiments in rats where signalling mechanisms in the MGM were blocked supports the idea that activity in connections from the auditory thalamus contribute to fear memory formation in the lateral amygdala. The slide shows the circuits believed to underlie such behaviour. _____________________ Circuiteria del condicionamiento al miedo mediado por sonido La vía rápida al MGM podría jugar un papel muy importante para las respuestas emocionales al sonido a través de la proyección del MGM al sistema límbico. Sabemos que El MGM es importante para el condicionamiento al miedo mediado por sonidos. Este comportamiento desaparece con la destrucción de MGM. Las células en el MGM también aumentan su descarga ante un estímulo condicionado auditivo después de un condicionamiento al miedo y demuestran una potenciación a largo plazo. Las neuronas de MGM también demuestran plasticidad por el que la selectividad de la frecuencia de una neurona se ajusta a la del estímulo condicionado. Los experimentos en las ratas donde se bloquean los mecanismos de señalización en el MGM apoyan la idea de que las conexiones del tálamo auditivo contribuyen a la formación de la memoria del miedo en la amigdala lateral. La diapositiva muestra los circuitos involucrados en este tipo de comportamiento.

The auditory cortex
Moderador
Notas de la presentación
The auditory cortex This section explains the basic organisation of auditory cortex in humans, and in number of other species that have been used in experimental studies. A key finding is the topographic mapping of sound frequency on the cortical surface. La corteza auditiva Esta sección explica la organización básica de la corteza auditiva en seres humanos, y en gran número de otras especies que se han utilizado en estudios experimentales. Un hallazgo muy importnate es el mapeo topográfico de la frecuencia del sonido en la superficie cortical.

Primary auditory cortex: Heschl’s gyrus
Location of auditory cortex in human brain
Planum Temporale
Moderador
Notas de la presentación
Location of auditory cortex in human brain On the left is a human brain with the temporal lobe outlined by the red dotted line. The primary auditory cortex is on the superior surface of the temporal lobe which runs parallel with the Sylvian fissure and is normally hidden from view by the overlying cortex of the frontal and parietal lobes. When this is removed by making a cut through the brain at the angle of the blue dotted line the upper surface of the temporal lobe is revealed. The primary auditory cortex is to be found in the first Heschl’s gyrus which runs transversely across the temporal lobe. Behind Heschl’s gyrus is the planum temporale. Until recently this was believed to be principally involved in the processing of speech and was reported as being larger in the left hemisphere than in the right. More recent studies suggest that that its function is more complex and that it plays a more generic role in sound processing not just speech processing. Localización de la corteza auditiva en el cerebro humano En la imagen de la izquierda podemos observar el lóbulo temporal delineado en rojo con una línea discontinua. La corteza auditiva esta en la superficie superior del lóbulo temporal que es paralelo al surco de Silvio, el cual esta normalmente oculto por los lóbulos de cortezas frontal y parietal que la cubren por encima. Si disecamos y quitamos estos lóbulos frontal y parietal, haciendo un corte a través del cerebro siguiendo el plano que muestra la linea de puntos azul, podemos ver claramente la parte superior de la superficie del lóbulo temporal. La corteza auditiva primaria humana se sitúa en la primera circunvolución transversa (de Heschl) en el opérculo temporal del surco lateral (de Silvio), y corresponde al área 41 de Brodmann. Por detrás de la primera circunvolución transversa (de Heschl) se encuentra el planum temporale. Hasta hace muy poco se pensaba que esta área estaba involucrada en el procesamiento del habla y se decía que era mayor en el hemisferio izquierdo que en el derecho. Sin embargo estudios más recientes han demostrado que su función es mucho más compleja y juego un papel mucho más genérico en el procesamiento del sonido y no solamente en el procesamiento del habla.

The auditory cortex in the cat brain
Moderador
Notas de la presentación
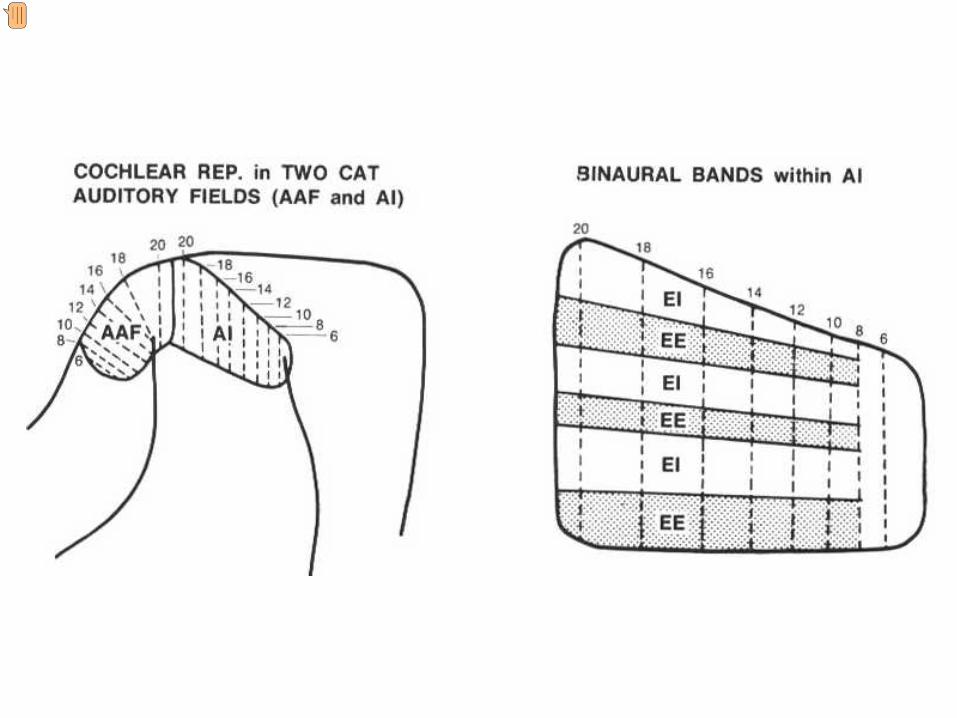
The auditory cortex in the cat brain The auditory cortex of the cat is mostly on the surface of the brain and therefore more accessible than in primates. There are several cortical regions in the auditory cortex that respond to sound. Sound frequency is mapped topographically onto these areas represented here according to the response of the basilar membrane. A = APEX of the cochlea (activated by low frequencies) and B = BASE activated by high frequencies. The direction of increasing sound frequency is represented by the red arrows. The frequency mapping in each area is a mirror image of that in adjacent regions The anterior auditory field seen in previous and next slides is here labelled SSF Corteza auditiva del gato La corteza auditiva del gato se localiza sobre la superficie del cerebro y por tanto es accesible más facilmente que en los primates. En gato hay varias regiones corticales que responsen al sonido. La frecuenia del sonido está representada topográficamente en estas áreas siguiendo el patrón que se genera en la membrana basilar de la cóclea. A = Apex of the cóclea (activada por frecuencias bajas) and B = base de la cóclea, activada por frecuencias altas. La progresión del incremento de frecuencias se indica por la flechas rojas. La organización tonotópica de cada región es una imagen especular de la región adyacente. El campo auditivo anterior que observamos aqui, y en diapositivas siguientes se refiere como SSF. .
Moderador
Notas de la presentación
On the right are AI and the anterior auditory field (AAF) in cat (see previous slide) showing the tonotopic frequency representation. The dotted lines represent iso frequency laminae which run orthogonal to the tonotopic axis. Frequency remains approximately constant along the dotted lines. The picture on the left shows an expanded representation of AI. Running across the iso frequency laminae are bands of cortex representing different patterns of binaural interaction. EE (excitation/excitation) represents areas where stimulation of both ears activates the neurons, and EI (excitation/ inhibition) to areas where stimulation of the contralateral ear gives excitation and stimulation of the ear ipsilateral to the cortex gives an inhibitory response. This organisation relates to the localisation of sounds in the horizontal plane which depends of comparing the properties of the sound in one ear with the other. A la izquierda se muestra el area AI y el campo auditivo anterior (AAF) en gato (ver diapositiva anterior) y sus correspodientes organizaciones tonotópicas.Las líneas de puntos representan láminas de isofrecuencia que están orientadas perpendicularmente al eje tonotópico. La frecuencia permanece constante a lo largo de las línes discontinuas. ____________________ A la derecha está el área AI y el campo auditivo anterior (AAF) en el gato (véase la diapositiva anterior) que muestra la representación tonotopica de la frecuencia. Las líneas punteadas representan los laminae de isofrecuencia que están orientadas perpendicularmente al eje tonotopico. La frecuencia permanece aproximadamente constante a lo largo de las líneas punteadas. La imagen de la derecha muestra una representación aumentada de A1 Existen bandas que están orientadas de manera perpendicular a las láminas de isofrecuencia, que representan distintos patrones de interacción binauricular. EE (excitación/excitación) representa areas donde la estimulación por los dos oidos activa a las neuronas y EI (excitación/inhibición) áreas donde la estimulación contralateral proporciona excitación y la estimulación ipsilateral a la corteza da lugar a respuestas inhibitorias. Esta organización se refiere a la localización del sonido en el plano horizontal, lo cual depende de la comparación de las propiedades del sonido de un oído con las del otro.

Caudal Rostral
Moderador
Notas de la presentación
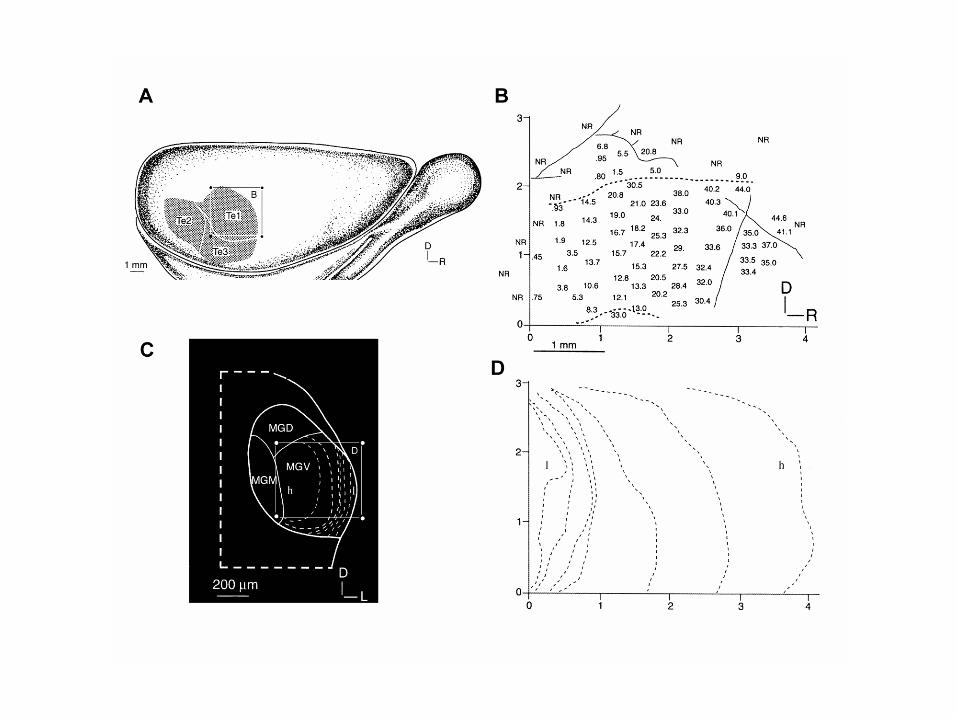
As in humans, the auditory cortex in the macaque monkey is covered by the parietal cortex. The primary auditory cortex (A1) is, as in the cat, surrounded by other areas. All these areas show a tonotopic mapping as indicated by the letters l and h standing for high and low frequency respectively. Note that in A1 high frequencies are represented caudally and low frequencies rostrally, and that again like the cat tonotopic maps in neighbouring areas are mirror images of one another. ___________ Al igual que ocurre en los humanos, la corteza auditiva del mono macaco está cubierta por la corteza parietal. La corteza auditiva primaria (AI) está rodeada por áreas secundarias como en el gato. Todas estas áreas muestran una representación tonotópica como se indica por las letras l y h que se refieren a bajas y altas frecuencias respectivamente. Obsérvese que en AI las frecuencias altas (h) se localizan en la parte caudal y las bajas (l) en la parte rostral. Los mapas tonotópicos de regiones adyacentes son especulares tal como vimos para el gato.

Multiple tonotopic maps in the cortex
Rostral Caudal
Moderador
Notas de la presentación
Multiple tonotopic maps in the cortex The prevailing model for the primate auditory cortex is now a core and belt model, in which core primary areas (shown here in blue) are surrounded by a belt of other areas (dark grey). The core areas are called AI, R and RT while the surrounding belt areas are designated medial or lateral (the letters M or L) with further letters to indicate rostral (R) anterior (A), medial (M) and caudal (C). Mapas tonotópicos múltiples en la corteza auditiva El modelo aceptado de la corteza auditiva primaria de primates incluye una zona central y áreas adyacentes, donde la zona central (core, en azul) y está rodeada de las zonas adyacentes (belt, gris oscuro). La zona central incluye las áreas AI, R y RT, mientras que las zonas adyacentes se denominan lateral o medial (L y M) con letras adicionales que indical rostral (R), anterior (A), medial (M) y caudal (C).

Properties of auditory cortical areas
Moderador
Notas de la presentación
Properties of auditory cortical areas The existence of several distinct areas in auditory cortex is well established. Recent studies have proposed that not only these different areas are specialised for the processing of specific stimulus properties, but also a still controversial hypothesis that there are two distinct processing streams or pathways in the cortex. Características de áreas corticales auditivas La corteza auditiva tiene varias áreas distintas bien delimitadas. Estudios recientes han propuesto que estas diversas áreas no sólo están especializadas para procesar de las características específicas del estímulo, sino también han propuesto una hipótesis, aún polémica, según la cual que hay dos corrientes o flujos de procesamiento distintos en la corteza.

Cortical auditory areas in macaque monkey
after Rauschecker 1998
AL ML CL
CMA1R
Parabelt areas
Rostral Caudal
Lateral
Medial
Frequency
High High LowLow
Ban
dwid
th
Wid
eN
arro
w
Moderador
Notas de la presentación
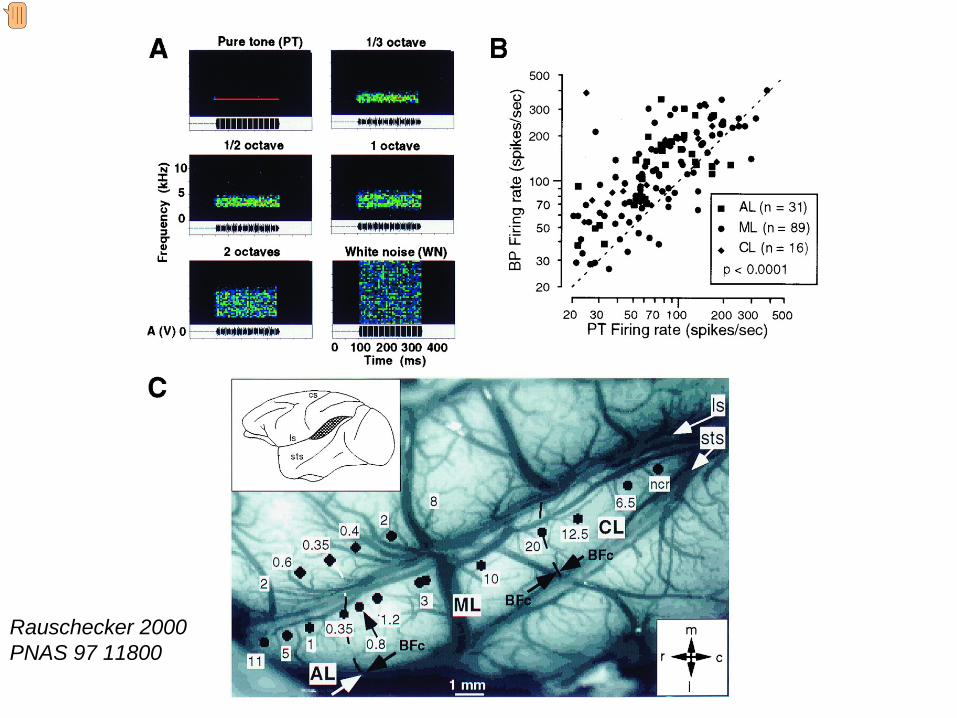
Cortical auditory areas in macaque monkey The slide shows a schematic representation of the superior temporal gyrus of the macaque. Two of the primary auditory areas (R and A1) are shown, surrounded by the belt areas (white). In turn surrounding the belt areas are a series of parabelt areas (yellow). Recent electrophysiological studies from Rauschecker’s laboratory have shown that certain response trends can be found in these different areas. Neurons in the core regions appear to respond best to sounds with narrow frequency bandwidths (eg pure tones), in contrast in the belt and parabelt regions, units respond most readily to more complex sounds with wider bandwidths as shown in the next slide. These neurons also responded well to frequency sweeps and monkey vocalisations. Rauschecker et al have also reported that neurons in the more rostral belt areas show a greater selectivity for particular types of monkey vocalisations, while the more caudal areas show greater selectivity for the location of the sound source. Áreas corticales auditivas en el mono macaco Esta diapositiva muesta una representación del gyrus temporal superior del macaco. Se muestran 2 áreas corticales primarias (R y A1, cuadros azules) rodeadas de las áreas adyacentes (cuadros blanco). Además existen áreas adyacentes perífericas (cuadros amarillos). Estudios electrofisiologicos recientes del laboratorio dirigido por el Prof. Rauschecker han demostrado que ciertas respuestas neuronales tienden a estas presentes de manera segregada en estas áreas diferentes. Neuronas en la zonal central o princila (core) responden mejor a sonidos con un ancho de banda en frecuencia muy estrecho (por ejemplo, tonos puros), mientras que la áreas adyacentes (belt) y periféricas (parabelt) responden mejor a sonidos complejos con anhos de banda más amplios como veremos en la diapositiva siguiente. Estas áreas también responden muy bien a cambios de frecuencia y a vocalizaciones del mono. Rauschecker y sus colaboradores también han demostrado que las áreas adyacentes (belt) más rostrales muestran una gran selectividad a vocalizaciones muy particulares del mono, mientras que las áreas adyacentes (belt) más caudales muestran mayor selectividad a a la localización de un foco sonoro.
Rauschecker 2000 PNAS 97 11800
Moderador
Notas de la presentación
Evidence that neurons in the belt areas prefer wider bandwidth sounds can be seen in this slide. shows the stimuli used in the experiment. These range in bandwidth between a pure tone and a white noise. For each cell recorded in the different belt areas, the firing rate to a pure tone (abscissa) is plotted against the firing for the wider bandwidth stimuli (ordinate). In all cases the points lie above the line of identity indicating that the response neuron responds more effectively to wider bandwidth sounds. the locations of the electrode penetrations and the best frequency at each site is mapped on the surface of the brain. Evidencias de que las neuronas de áreas adyacentes (belt) prefieren sonidos de banda ancha Muestra el tipo de estímulo empleado en estos experimentos, cuyo ancho de banda varía entre un de un tono puro y el de un ruido blanco. Para cada una de las neuronas registradas en las diferentes areas adyacentes se plotea la descarga neuronal para un tono puro (X, abscisa) versus la descarga de un estímulo de ancho de banda más amplia (Y, ordenada). En todos los casos, los puntos se localizan por encima de la línea de identidad lo que indica que las respuestas neuronales son más efectivas a sonidos de ancho de banda más ancho. muestra la localización de las penetraciones realizadas con los electrodos de registro y la representación de la mejor frecuencia según se ha mapeado sobre la superficie del cerebro.

The dorsal and ventral stream hypothesis
Posterior parietal cortex
Prefrontal cortex
What? Auditory cortex
Where?
D
V
Moderador
Notas de la presentación
Summary of the dorsal and ventral “what” and “where” stream hypothesis. Information about the identify of the stimulus (“what”) projects ventrally from the auditory cortex to the prefrontal cortex (blue arrow). The dorsal “where” stream projects dorsally to the posterior parietal cortex and to the prefrontal cortex. Recent studies on the processing of sounds (including speech) in humans using PET and fMRI provide further support for this notion (see later). Although the hypothesis appears attractive in outline there are experimental findings that do not fit with its predictions. Resumen de la hipótesis de flujos dorsal y ventral de “qué” y “donde”. La información sobre la identificación de un estímulo (“qué”) proyecta ventralmente desde la corteza auditiva a la corteza prefrontal (flecha azul). El flujo dorsal “donde” proyecta dorsalmente a la corteza parietal posterior y a la corteza prefrontal. Los estudios recientes sobre el procesamiento de sonidos (incluyendo el habla) en los seres humanos usando PET y fMRI proporcionan información adicional que sustenta esta teoría (véase más adelante). Aunque la hipótesis aparece atractiva en su conjunto, hay resultados experimentales que no confirman sus predicciones.


















