
ThalassemicDNA-ContainingRedBloodCellsAreunder OxidativeStress
6
Hindawi Publishing Corporation Anemia Volume 2012, Article ID 943974, 5 pages doi:10.1155/2012/943974 Research Article Thalassemic DNA-Containing Red Blood Cells Are under Oxidative Stress Mutaz Dana, Eugenia Prus, and Eitan Fibach Department of Hematology, Hadassah-Hebrew University Medical Center, Ein-Kerem, P.O. Box 12000, Jerusalem 91120, Israel Correspondence should be addressed to Eitan Fibach, fi[email protected] Received 12 September 2011; Revised 1 December 2011; Accepted 4 December 2011 Academic Editor: Maria Cappellini Copyright © 2012 Mutaz Dana et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. We studied the nature of enucleated RBCs containing DNA remnants, Howell-Jolly (HJ) RBCs and reticulocytes (retics), that are characteristically present in the circulation of thalassemic patients, especially after splenectomy. Using flow cytometry methodol- ogy, we measured oxidative status parameters of these cells in patients with β-thalassemia. In each patient studied, these cells had higher content of reactive oxygen species and exposed phosphatidylserine compared with their DNA-free counterparts. These re- sults suggest that oxidative stress in thalassemic developing erythroid precursors might, through DNA-breakage, generate HJ-retics and HJ-RBCs and that oxidative stress-induced externalization of phosphatidylserine is involved in the removal of these cells from the circulation by the spleen, a mechanism similar to that of the removal of senescent RBCs. 1. Introduction The development of red blood cells (RBCs) from their pro- genitors in the bone marrow includes the process of enuclea- tion in which the final stages of nucleated erythroid precur- sors (orthochromatic normoblasts) expel their nuclei to gen- erate enucleated reticulocytes (retics), which leave the mar- row and mature into RBCs [1]. Normally, nucleated RBCs (normoblasts) are undetectable in the circulation, but in some hematological pathologies (e.g., thalassemia and sickle cell disease) they can be found in large numbers [1]. These diseases are also characterized by mature RBCs and retics that contain DNA remnants, that are called Howell-Jolly (HJ) bodies [1]. The frequency of these cells, which is very low, has been quantified using a flow cytometry technique [2–4]. The spleen is the major site of the reticuloendothelial system where senescent RBCs at the end of their life-span are removed by erythrophagocytosis [5]. It also removes from the circulation normoblasts and HJ-cells; thus, in thalassemia and sickle cell disease, the number of these cells in the patients’ circulation increases considerably following sple- nectomy [1]. The removal of senescent RBCs has been attributed to various mechanisms [5], including exposure (externaliza- tion) of phosphatidylserine (PS) on their surface [6]. The macrophages of the reticuloendothelial system carry surface receptors that specifically bind PS, by which they internalize senescent RBCs [7]. The mechanism by which normoblasts and HJ-cells are removed from the circulation is unknown. We have previously shown that in hemolytic anemias, including thalassemia and sickle cell disease, RBCs are under oxidative stress [8], and they generate more reactive oxygen species (ROS) and contain less reduced glutathione than normal RBCs, which results in membrane changes such as lipid peroxidation and externalization of PS. Using flow cytometry, in the present study we show that HJ-RBCs and retics are under oxidative stress and carry exposed PS, which may present the trigger for their phago- cytosis by macrophage and removal in the spleen. 2. Materials and Methods Blood Samples. Peripheral blood (PB) samples were obtained from normal donors and splenectomized and nonsplenec- tomized patients with β-thalassemia intermedia and major. The samples were obtained from the counting vials after all diagnostic laboratory tests were completed. The research was approved by the Hadassah-Hebrew University Medical Centre Human Experimentation Review Board. The patients’
Transcript of ThalassemicDNA-ContainingRedBloodCellsAreunder OxidativeStress

Hindawi Publishing CorporationAnemiaVolume 2012, Article ID 943974, 5 pagesdoi:10.1155/2012/943974
Research Article
Thalassemic DNA-Containing Red Blood Cells Are underOxidative Stress
Mutaz Dana, Eugenia Prus, and Eitan Fibach
Department of Hematology, Hadassah-Hebrew University Medical Center, Ein-Kerem, P.O. Box 12000, Jerusalem 91120, Israel
Correspondence should be addressed to Eitan Fibach, [email protected]
Received 12 September 2011; Revised 1 December 2011; Accepted 4 December 2011
Academic Editor: Maria Cappellini
Copyright © 2012 Mutaz Dana et al. This is an open access article distributed under the Creative Commons Attribution License,which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
We studied the nature of enucleated RBCs containing DNA remnants, Howell-Jolly (HJ) RBCs and reticulocytes (retics), that arecharacteristically present in the circulation of thalassemic patients, especially after splenectomy. Using flow cytometry methodol-ogy, we measured oxidative status parameters of these cells in patients with β-thalassemia. In each patient studied, these cells hadhigher content of reactive oxygen species and exposed phosphatidylserine compared with their DNA-free counterparts. These re-sults suggest that oxidative stress in thalassemic developing erythroid precursors might, through DNA-breakage, generate HJ-reticsand HJ-RBCs and that oxidative stress-induced externalization of phosphatidylserine is involved in the removal of these cells fromthe circulation by the spleen, a mechanism similar to that of the removal of senescent RBCs.
1. Introduction
The development of red blood cells (RBCs) from their pro-genitors in the bone marrow includes the process of enuclea-tion in which the final stages of nucleated erythroid precur-sors (orthochromatic normoblasts) expel their nuclei to gen-erate enucleated reticulocytes (retics), which leave the mar-row and mature into RBCs [1]. Normally, nucleated RBCs(normoblasts) are undetectable in the circulation, but insome hematological pathologies (e.g., thalassemia and sicklecell disease) they can be found in large numbers [1]. Thesediseases are also characterized by mature RBCs and reticsthat contain DNA remnants, that are called Howell-Jolly (HJ)bodies [1]. The frequency of these cells, which is very low, hasbeen quantified using a flow cytometry technique [2–4].
The spleen is the major site of the reticuloendothelialsystem where senescent RBCs at the end of their life-span areremoved by erythrophagocytosis [5]. It also removes fromthe circulation normoblasts and HJ-cells; thus, in thalassemiaand sickle cell disease, the number of these cells in thepatients’ circulation increases considerably following sple-nectomy [1].
The removal of senescent RBCs has been attributed tovarious mechanisms [5], including exposure (externaliza-tion) of phosphatidylserine (PS) on their surface [6]. The
macrophages of the reticuloendothelial system carry surfacereceptors that specifically bind PS, by which they internalizesenescent RBCs [7]. The mechanism by which normoblastsand HJ-cells are removed from the circulation is unknown.
We have previously shown that in hemolytic anemias,including thalassemia and sickle cell disease, RBCs are underoxidative stress [8], and they generate more reactive oxygenspecies (ROS) and contain less reduced glutathione thannormal RBCs, which results in membrane changes such aslipid peroxidation and externalization of PS.
Using flow cytometry, in the present study we show thatHJ-RBCs and retics are under oxidative stress and carryexposed PS, which may present the trigger for their phago-cytosis by macrophage and removal in the spleen.
2. Materials and Methods
Blood Samples. Peripheral blood (PB) samples were obtainedfrom normal donors and splenectomized and nonsplenec-tomized patients with β-thalassemia intermedia and major.The samples were obtained from the counting vials afterall diagnostic laboratory tests were completed. The researchwas approved by the Hadassah-Hebrew University MedicalCentre Human Experimentation Review Board. The patients’

2 Anemia
mutations and some relevant clinical parameters (e.g., trans-fusion and chelation therapy, splenectomy) were previouslysummarized [9]. In polytransfused patients, blood sampleswere obtained before transfusion, that is, at least 3 weeksfollowing the previous transfusion. Informed consent wasobtained in all cases.
Flow Cytometry Measurements of Oxidative Stress Markers.Cells were stained for transferrin-receptor by incubating with5 μL of APC-conjugated antibodies (Ab) to CD71 at 4◦C for30 minutes. The sample was washed and then divided intotwo aliquots: one aliquot was stained for ROS with 2′-7′-dichlorofluorescin diacetate (DCFH, Sigma, St, Louis, MO),at final concentration of 0.1 mM, at 37◦C for 15 minutes,then washed three times with Ca++- and Mg++-free Dul-becco’s phosphate-buffered-saline (PBS) (Biological Indus-tries, Beit-HaEmek, Israel). A stock solution of 20 mM DCFwas prepared in methanol (Bio Lab, Jerusalem, Israel). Theother aliquot was stained for external phosphatidylserine(PS), by suspending the cells in 100 μL of calcium buffer((10 mM HEPES, 140 mM NaCl and 2.5 mM CaCl2 (pH 7.4))and 2 μL of FITC-conjugated Annexin-V (IQ Products, Gro-ningen, The Netherlands). After 15 minutes at room tem-perature, in the dark, the cells were washed three times withcalcium buffer and resuspended in 0.5 mL of the same buffer.
For every assay, 2 μL of propidium iodide (PI, Mallinck-rodt Chemical Works, St. Louis, MO), dissolved in 0.1%sodium citrate, was added before analysis. Cells stained withanti-CD71 Ab alone, cells stained with anti-CD71 Ab andannexin-V, or cells stained with anti-CD71 Ab and DCF wereused as controls to set the compensation levels. Followingtreatment as indicated above, the cells were analyzed witha Fluorescence Activated Cell Sorter (FACS-calibur, Becton-Dickinson, Immunofluorometry systems, Mountain View,CA). Instrument calibration and settings were performedusing CaliBRITE-3 beads (Becton-Dickinson). The cells werepassed at a rate of ∼1,000 per second, using saline as thesheath fluid. A 488 nm argon laser beam was used for excita-tion. Threshold was set on forward light scatter (FSC) toexclude platelets and cell debris. Gates were set on RBCs,HJ-RBCs, retics, HJ-retics, normoblasts, and WBCs. Cellslabeled with DCF and annexin-V were detected by the FL-1PMT, and cells labeled with APC-conjugated anti-CD-71 Aband PI were detected by the FL-4 and FL-2 PMT, respectively.All PMTs were set on log amplification. The Mean Fluo-rescence Intensities (MFIs) and the percentages of positivecells were calculated using the FACS-equipped CellQuestsoftware (Becton-Dickinson). The results are expressed as theaverage ± standard deviation (SD) and compared using thetwo-sample Student’s t-test for differences in means.
3. Results and Discussion
PB cells were simultaneously stained with an anti-CD71 Aband PI, and either DCF or annexin-V. The anti-CD71 Abmarks the transferrin receptor, and PI the nucleic acid con-tent. To evaluate the contribution of RNA (particularly in
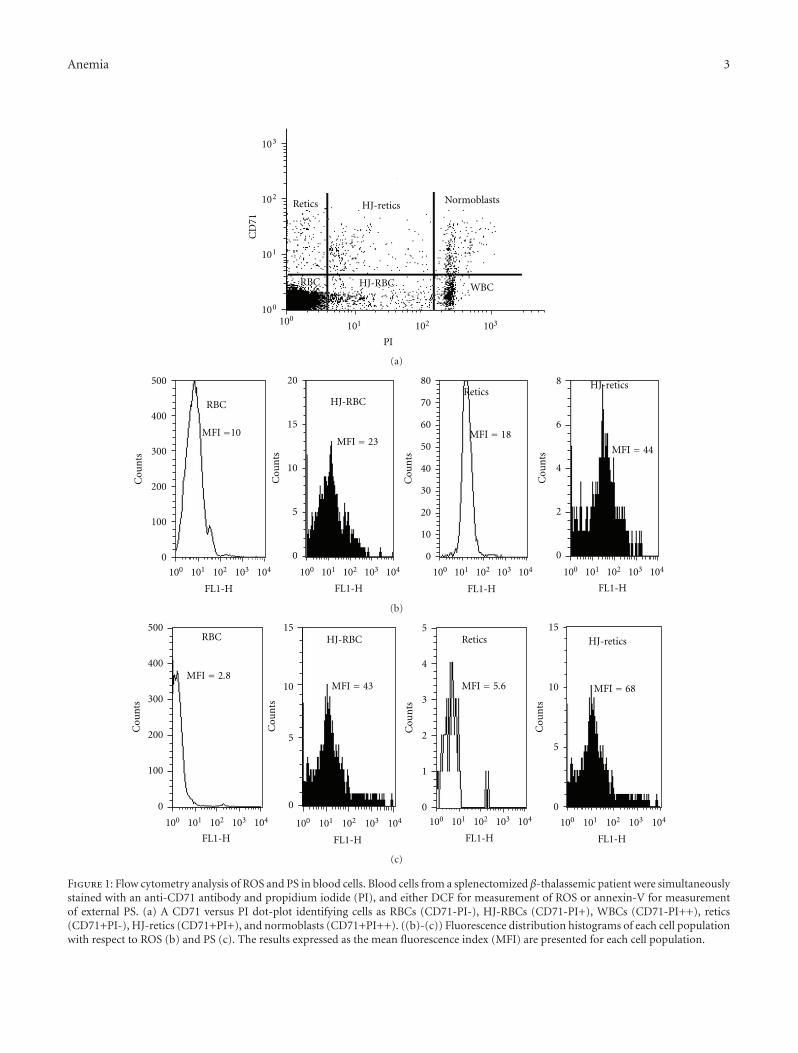
retics which contain small amounts of residual RNA) to thePI staining, PB cells were stained with PI in the presence orabsence of RNase (0.4 mg/mL, Invitrogen, Carlsbad, CA). Nodifference was noted in the pattern of PI staining betweenthese samples. The staining procedure identified cells asRBCs (CD71-PI-), HJ-RBCs (CD71-PI+), WBCs (CD71-PI++), retics (CD71+PI-), HJ-retics (CD71+PI+), and nor-moblasts (CD71+PI++). Figure 1(a) shows a flow cytometrydot-plot (PI versus CD71) analysis of a blood sample derivedfrom a representative splenectomized β-thalassemic patient,indicating the various cell populations. The fluorescencedistribution histograms of each cell population with respectto DCF-fluorescence, indicating generation of ROS, and an-nexin V-fluorescence, indicating exposed PS, with theirMFIs, are shown in Figures 1(b) and 1(c), respectively. Theresults indicate higher ROS and PS in retics than in matureRBCs, and, more critically, in HJ-cells compared with theirnon-HJ counterparts: in the experiment presented in Figure1(b), showing ROS results, the MFI of HJ-RBCs was 2.3-foldhigher than that of RBCs, and the MFI of HJ-retics was 2.4-fold higher than retics. In Figure 1(c), showing PS results, theMFI of HJ-RBCs was 15.3-fold higher than that of RBCs, andthe MFI of HJ-retics was 12.1-fold higher than retics.
Figure 2(a) depicts the frequency of HJ-RBCs in the PB ofnormal donors and in thalassemic patients. The results showno HJ-RBCs in normal donors and much higher frequencyof HJ-RBCs in splenectomized patients compared with non-splenectomized patients. Figures 2(b)-2(c), which summa-rize the average ROS generation and percentage of PS-expos-ing cells, show that both parameters were significantly higherin HJ-RBCs versus RBCs and in HJ-retics versus retics. Theresults also show that both parameters are higher in cellsfrom splenectomized versus nonsplenectomized patients,suggesting that the spleen removes the most damaged cells.
Although the process of nuclear expulsion from develop-ing RBC precursors has been studied extensively [10, 11], thereasons for nuclear remnants (HJ-bodies) leftover in enucle-ated retics and RBCs in certain diseases have not been studiedbefore. We now report that in β-thalassemia the generationof ROS and the externalization of PS, both parameters ofoxidative stress, are elevated in HJ-retics and HJ-RBCs com-pared with their no-HJ-containing counterparts. ROS maybe the cause of HJ formation. They are known to cause DNAbreaks [12] that may generate micronuclei in various celltypes [13], including lymphocytes and neutrophils. The oc-currence of micronuclei has been used as a biomarker forcytogenetic damage [14, 15]. These micronuclei are equiva-lent to the HJ bodies in RBCs. The mechanism of HJ bodies’formation must occur prior to nuclear expulsion. We havepreviously demonstrated that thalassemic erythroid precur-sors, including orthochromatic normoblasts, are at higheroxidative status than their normal counterparts [9]. It mightbe hypothesized that DNA/nuclear breaks induced by oxida-tive stress might result in incomplete expulsion of the nuclearmaterial, resulting in nuclear remnants which remain inretics and mature RBCs.
Several studies [16], including our own [9], indicatedthat ROS stimulate PS externalization on RBCs. Exposed
Anemia 3
100
101
102
103
100101 102 103
Retics HJ-retics Normoblasts
RBC HJ-RBC WBC
CD
71
PI
(a)
RBC HJ-RBCRetics
HJ-retics
MFI =10
0
100
200
300
400
500
100 101 102 103 104
FL1-H
0
5
10
15
20
0
10
20
30
40
50
60
70
80
0
2
4
6
8
100 101 102 103 104 100 101 102 103 104 100 101 102 103 104
FL1-H FL1-H FL1-H
Cou
nts
Cou
nts
Cou
nts
Cou
nts
MFI = 23MFI = 18
MFI = 44
(b)
0
100
200
300
400
500
0
5
10
15
0
1
2
3
4
5
0
5
10
15
Cou
nts
100 101 102 103 104
FL1-H
RBC HJ-RBC Retics HJ-retics
100 101 102 103 104
FL1-H
100 101 102 103 104
FL1-H
100 101 102 103 104
FL1-H
Cou
nts
Cou
nts
Cou
nts
MFI = 2.8MFI = 43 MFI = 5.6 MFI = 68
(c)
Figure 1: Flow cytometry analysis of ROS and PS in blood cells. Blood cells from a splenectomized β-thalassemic patient were simultaneouslystained with an anti-CD71 antibody and propidium iodide (PI), and either DCF for measurement of ROS or annexin-V for measurementof external PS. (a) A CD71 versus PI dot-plot identifying cells as RBCs (CD71-PI-), HJ-RBCs (CD71-PI+), WBCs (CD71-PI++), retics(CD71+PI-), HJ-retics (CD71+PI+), and normoblasts (CD71+PI++). ((b)-(c)) Fluorescence distribution histograms of each cell populationwith respect to ROS (b) and PS (c). The results expressed as the mean fluorescence index (MFI) are presented for each cell population.

4 Anemia
0
0.1
0.2
0.3
0.4
Normal
HJ-
RB
C fr
equ
ency
Non splenectomized Splenectomizedthalassemia thalassemia
(a)
HJ-RBC
0
20
40
60
80ROS generation
Normal
RBC Retics
Splenectomized thalassemic patients
Non splenectomized thalassemic patients
HJ-retics
0
20
40
60
80
RBC Retics
RBC Retics
HJ-RBC HJ-retics
0
20
40
60
80
RBC ReticsHJ-RBC HJ-retics
DC
F-fl
uor
esce
nce
(M
FI)
DC
F-fl
uor
esce
nce
(M
FI)
DC
F-fl
uor
esce
nce
(M
FI)
(b)
PS exposure
0
20
40
60
80Normal
RBC Retics
Non splenectomized thalassemic patients
Splenectomized thalassemic patients
0
20
40
60
80
RBC ReticsHJ-RBC HJ-retics
0
20
40
60
80
RBC ReticsHJ-RBC HJ-retics
An
nex
in V
(%
)A
nn
exin
V (
%)
An
nex
in V
(%
)
HJ-RBC HJ-reticsRBC Retics
(c)
Figure 2: The frequency of HJ-cells and their oxidative status in normal donors and thalassemic patients. Cells obtained from the bloodof normal donors and splenectomized and nonsplenectomized thalassemic patients (N = 6 in each group) were stained and analyzed as inlegends to Figure 1(a). (a) The frequency of HJ-RBCs. (b) ROS generation. (c) PS exposure. The results are expressed as the percentage inthe RBC population (a), the average ± S.D of the mean DCF-fluorescence index (MFI) for ROS (b) and the percentage of cells positivelystained with annexin-V for PS (c).
PS was suggested, in addition to other mechanisms such asreduced expression of CD47 [17] and binding of autologousimmunoglobulins and opsonins [18, 19] to signal erythroph-agocytosis and removal of senescent RBCs from the circula-tion. To our knowledge, the signals for phagocytosis and re-moval of peripheral blood normoblasts or HJ-cells have notbeen studied. Our findings of enhanced exposure of PS onHJ-cells might suggest that exposed PS might participate in
the removal of such cells by the spleen, although other signalscannot be ruled out.
In conclusion, the results of the present study suggest thatoxidative stress in developing erythroid precursors mightgenerate HJ-retics and HJ-RBCs and that oxidative stress-induced externalization of PS might be involved in theirremoval from the circulation by the spleen, a mechanismsimilar to that of the removal of aging (senescent) RBCs.

Anemia 5
References
[1] J. Jandl, Blood—Textbook of Hematology, Little, Brown andCompany, Boston, Mass, USA, 1996.
[2] S. D. Dertinger, Y. Chen, R. K. Miller et al., “MicronucleatedCD71-positive reticulocytes: a blood-based endpoint of cyto-genetic damage in humans,” Mutation Research, vol. 542, no.1-2, pp. 77–87, 2003.
[3] T. Offer, A. Bhagat, A. Lal et al., “Measuring chromosomebreaks in patients with thalassemia,” Annals of the New YorkAcademy of Sciences, vol. 1054, pp. 439–444, 2005.
[4] V. L. Harrod, T. A. Howard, S. A. Zimmerman, S. D. Dertinger,and R. E. Ware, “Quantitative analysis of Howell-Jolly bodiesin children with sickle cell disease,” Experimental Hematology,vol. 35, no. 2, pp. 179–183, 2007.
[5] D. Bratosin, J. Mazurier, J. P. Tissier et al., “Cellular and mol-ecular mechanisms of senescent erythrocyte phagocytosis bymacrophages. A review,” Biochimie, vol. 80, no. 2, pp. 173–195,1998.
[6] V. A. Fadok, D. L. Bratton, S. C. Frasch, M. L. Warner, and P.M. Henson, “The role of phosphatidylserine in recognition ofapoptotic cells by phagocytes,” Cell Death and Differentiation,vol. 5, no. 7, pp. 551–562, 1998.
[7] Z. Zhou, “New phosphatidylserine receptors: clearance ofapoptotic cells and more,” Developmental Cell, vol. 13, no. 6,pp. 759–760, 2007.
[8] J. Amer, A. Goldfarb, and E. Fibach, “Flow cytometric analysisof the oxidative status of normal and thalassemic red bloodcells,” Cytometry A, vol. 60, no. 1, pp. 73–80, 2004.
[9] I. Freikman, J. Amer, J. S. Cohen, I. Ringel, and E. Fibach,“Oxidative stress causes membrane phospholipid rearrange-ment and shedding from RBC membranes—an NMR study,”Biochimica et Biophysica Acta, vol. 1778, no. 10, pp. 2388–2394,2008.
[10] H. Yoshida, K. Kawane, M. Koike, Y. Mori, Y. Uchiyama, andS. Nagata, “Phosphatidylserine-dependent engulfment by ma-crophages of nuclei from erythroid precursor cells,” Nature,vol. 437, no. 7059, pp. 754–758, 2005.
[11] G. Keerthivasan, A. Wickrema, and J. D. Crispino, “Erythrob-last enucleation,” Stem Cells International, vol. 2011, Article ID139851, 9 pages, 2011.
[12] J. Cadet, T. Douki, and J. L. Ravanat, “Oxidatively generatedbase damage to cellular DNA,” Free Radical Biology and Med-icine, vol. 49, no. 1, pp. 9–21, 2010.
[13] P. Belloni, P. Latini, and F. Palitti, “Radiation-induced bystand-er effect in healthy G0 human lymphocytes: biological andclinical significance,” Mutation Research, vol. 713, no. 1-2, pp.32–38, 2011.
[14] D. F. Smith, J. T. MacGregor, R. A. Hiatt et al., “Micronucleatederythrocytes as an index of cytogenetic damages in humans:demographic and dietary factors associated with micro-nucleated erythrocytes in splenectomized subjects,” CancerResearch, vol. 50, no. 16, pp. 5049–5054, 1990.
[15] A. Vral, M. Fenech, and H. Thierens, “The micronucleus assayas a biological dosimeter of in vivo ionising radiation expo-sure,” Mutagenesis, vol. 26, no. 1, pp. 11–17, 2011.
[16] M. Foller, S. M. Huber, and F. Lang, “Erythrocyte pro-grammed cell death,” IUBMB Life, vol. 60, no. 10, pp. 661–668,2008.
[17] S. Khandelwal, N. van Rooijen, and R. K. Saxena, “Reducedexpression of CD47 during murine red blood cell (RBC)senescence and its role in RBC clearance from the circulation,”Transfusion, vol. 47, no. 9, pp. 1725–1732, 2007.
[18] U. Galili, I. Flechner, and E. A. Rachmilewitz, “A naturallyoccurring anti-alpha-galactosyl IgG recognizing senescenthuman red cells,” Progress in Clinical and Biological Research,vol. 195, pp. 263–278, 1985.
[19] A. Pantaleo, G. Giribaldi, F. Mannu, P. Arese, and F. Turrini,“Naturally occurring anti-band 3 antibodies and red blood cellremoval under physiological and pathological conditions,”Autoimmunity Reviews, vol. 7, no. 6, pp. 457–462, 2008.
Submit your manuscripts athttp://www.hindawi.com
Stem CellsInternational
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Disease Markers
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation http://www.hindawi.com Volume 2014
Immunology ResearchHindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Parkinson’s Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttp://www.hindawi.com






![RESEARCHARTICLE TheImpactofMyeloperoxidaseand ...€¦ · expectations frommedical science.Asubstantialbodyofliteraturehasproposedalinkbetween oxidative stressandpoor reproductiveoutcomes[1–3].Oxidativestress](https://static.fdocuments.us/doc/165x107/60509d378c4dd04cbf4c8cbd/researcharticle-theimpactofmyeloperoxidaseand-expectations-frommedical-scienceasubstantialbodyofliteraturehasproposedalinkbetween.jpg)