
Temporal regulation and overlap organization of two Caulobacter flagellar genes
13
J. Mol. Biol. (1989) 205. 7 l-83 Temporal Regulation and Overlap Organization of Two Caulobacter Flagellar Genes Jeffrey B. Kaplan?, Andrew Dingwall, Ruth Bryan Robert Champer and Lucille Shapirot Department of Microbiology, College of Physicians and Surgeons of Columbia University 701 West 168th Street New York, ,VY 10032, U.S.A. (Received 26 April 1988, and in revised form 17 August 1988) The biogenesis of the bacterial flagellum and chemotaxis apparatus in both Esch,erichia coli and Caulobacter crescentus requires the ordered expression of over 40 genes whose expression is controlled by a trans-acting regulatory hierarchy. In C. crescentus, additional control mechanisms ensure that the transcription of these genes is initiated at the correct time in the cell cycle. We demonstrate here that two flagellar genes, jlaE and $aY, whose products function in trans to modulate the level of transcription of other flagellar genes, are t.hemselves temporally controlled. DNA sequence analysis of the 3413 base-pairs encompassing the flaE and $a Y coding sequences and the 5’ regulatory region showed that $aE encodes a protein of 16,000 M, and $a Y a protein of 17,000 1M,. Evidence that ,flaE and $a Y are transcribed as a polycistronic message includes (1) the polar effect of Tn5 insertions; (2) deletion analysis showing that the jlaE promoter is essential for complementation of both JEaE and flay alleles; and (3) nuclease S1 assays showing protection of a transcript spanning both genes. The transcript start site in front of$aE was determined and the - 10 region conforms to the E. coli sigma 28 promoter consensus sequence. Nuclease S, analysis also revealed a protected fragment whose size was consistent with a t’ranscript initiating in vivo at a consensus “nif’ promoter sequence in front of the $a Y gene. The entire promoter region and an upstream consensus sequence that might, be a regulatory element for the jla Y gene lies within the carboxyl-terminal coding sequence of the&E gene. 1. Introduction Caulobacter crescentus bears a single, polar flagellum that is synthesized and assembled at a defined time in the cell cycle (for a review, see Shapiro, 1985). Like other bacterial flagella, it has a complex architecture. It consists of at least three subassemblies that include a transmembrane basal body composed of a rod threaded through several rings (Stallmeyer et al., 1985), a helical hook structure (Wagenknecht et aE., 1981), and a filament with an ordered alignment of three different flagellin subunits (Johnson et al., 1979; Koyasu et al., 1981; Weissborn et al., 1982; Driks & Bryan, unpublished results). Over 40 genes are involved in the biogenesis and function of this subcellular organelle and the accompanying chemotaxis 7 Present address: Lederle Laboratories, Pearl River. NY 10965. U.S.A. 1 Author to whom all correspondence should be sent. apparatus (Ely et al., 1984, 1986). In both C. crescentus and Escherichia coli ) the genes involved in flagellar biogenesis and the chemotactic response are organized in a trans.acting regulatory hierarchy that appears to contribute to tjhe ordered assembly of the flagellar structure and the chemo- taxis apparatus (Komeda, 1982, 1986; Champer et al.. 1985, 1987; Chen et al., 1986). Superimposed on this regulatory cascade in C. crescentus are addi- tional mechanisms that control timing of gene expression (Bryan et al., 1987). Mechanisms are also operative that orchestrate localized placement of flagellar and chemotaxis gene products at. one pole of the cell (Gomes & Shapiro, 1984; Nat>han et al., 1986; Loewy et aZ., 1987). Flagellar genes involved in the synthesis of the basal body (Hahnenberger & Shapiro, 1987, 1988), hook (Ohta et aZ., 1985; Chen et al., 1986) and the flagellins (Minnich & Newton, 1987: Loewy et al., 1987; Champer et al., 1987) have been shown to be turned on at specific times in the cell cycle. The 71 (HJZ%~P83ti~X9/01007 I-1 3 $cwH)/O 0 1989 Aractrmic I’rrss Limit&
Transcript of Temporal regulation and overlap organization of two Caulobacter flagellar genes

J. Mol. Biol. (1989) 205. 7 l-83
Temporal Regulation and Overlap Organization of Two Caulobacter Flagellar Genes
Jeffrey B. Kaplan?, Andrew Dingwall, Ruth Bryan Robert Champer and Lucille Shapirot
Department of Microbiology, College of Physicians and Surgeons of Columbia University
701 West 168th Street New York, ,VY 10032, U.S.A.
(Received 26 April 1988, and in revised form 17 August 1988)
The biogenesis of the bacterial flagellum and chemotaxis apparatus in both Esch,erichia coli and Caulobacter crescentus requires the ordered expression of over 40 genes whose expression is controlled by a trans-acting regulatory hierarchy. In C. crescentus, additional control mechanisms ensure that the transcription of these genes is initiated at the correct time in the cell cycle. We demonstrate here that two flagellar genes, jlaE and $aY, whose products function in trans to modulate the level of transcription of other flagellar genes, are t.hemselves temporally controlled. DNA sequence analysis of the 3413 base-pairs encompassing the flaE and $a Y coding sequences and the 5’ regulatory region showed that $aE encodes a protein of 16,000 M, and $a Y a protein of 17,000 1M,. Evidence that ,flaE and $a Y are transcribed as a polycistronic message includes (1) the polar effect of Tn5 insertions; (2) deletion analysis showing that the jlaE promoter is essential for complementation of both JEaE and flay alleles; and (3) nuclease S1 assays showing protection of a transcript spanning both genes. The transcript start site in front of$aE was determined and the - 10 region conforms to the E. coli sigma 28 promoter consensus sequence. Nuclease S, analysis also revealed a protected fragment whose size was consistent with a t’ranscript initiating in vivo at a consensus “nif’ promoter sequence in front of the $a Y gene. The entire promoter region and an upstream consensus sequence that might, be a regulatory element for the jla Y gene lies within the carboxyl-terminal coding sequence of the&E gene.
1. Introduction
Caulobacter crescentus bears a single, polar flagellum that is synthesized and assembled at a defined time in the cell cycle (for a review, see Shapiro, 1985). Like other bacterial flagella, it has a complex architecture. It consists of at least three subassemblies that include a transmembrane basal body composed of a rod threaded through several rings (Stallmeyer et al., 1985), a helical hook structure (Wagenknecht et aE., 1981), and a filament with an ordered alignment of three different flagellin subunits (Johnson et al., 1979; Koyasu et al., 1981; Weissborn et al., 1982; Driks & Bryan, unpublished results). Over 40 genes are involved in the biogenesis and function of this subcellular organelle and the accompanying chemotaxis
7 Present address: Lederle Laboratories, Pearl River. NY 10965. U.S.A.
1 Author to whom all correspondence should be sent.
apparatus (Ely et al., 1984, 1986). In both C. crescentus and Escherichia coli ) the genes involved in flagellar biogenesis and the chemotactic response are organized in a trans.acting regulatory hierarchy that appears to contribute to tjhe ordered assembly of the flagellar structure and the chemo- taxis apparatus (Komeda, 1982, 1986; Champer et al.. 1985, 1987; Chen et al., 1986). Superimposed on this regulatory cascade in C. crescentus are addi- tional mechanisms that control timing of gene expression (Bryan et al., 1987). Mechanisms are also operative that orchestrate localized placement of flagellar and chemotaxis gene products at. one pole of the cell (Gomes & Shapiro, 1984; Nat>han et al., 1986; Loewy et aZ., 1987).
Flagellar genes involved in the synthesis of the basal body (Hahnenberger & Shapiro, 1987, 1988), hook (Ohta et aZ., 1985; Chen et al., 1986) and the flagellins (Minnich & Newton, 1987: Loewy et al., 1987; Champer et al., 1987) have been shown to be turned on at specific times in the cell cycle. The
71 (HJZ%~P83ti~X9/01007 I-1 3 $cwH)/O 0 1989 Aractrmic I’rrss Limit&

72 J. B. Kaplan et al.
control of this temporal expression occurs at the transcriptional level. Two flagellar genes, JEaE and jla Y, are part of a gene cluster containing genes encoding three flagellins (Fig. 1). The $aE and jZa Y gene products function in trans to modulate the level of expression of the flagellin genes and several genes encoding chemotaxis functions (Bryan et al., 1984, 1987; Champer et al., 1985, 1987). Evidence is presented here that the $aE and jlaY genes are themselves temporally controlled. The jZaE and JEaY genes are organized in an operon and the polycistronic jl.aE Y message is first detected in the early predivisional cell. The transcription start site at the JlaE promoter was precisely mapped using a DNA sequence ladder, and a consensus sigma 28 promoter was found at the - 10 region. Nuclease S, protection assays also revealed an additional protected fragment whose size was consistent with a transcript intitiating at the beginning of the JaY gene. Analysis of the complete 3413 base-pair (bpt) nucleotide sequence comprising both genes and their 5’ regulatory region revealed a “nif ‘-type promoter sequence (Ausubel, 1984) at the beginning of the jlaY gene. In addition to this flay “nif’ promoter sequence, a 13 bp consensus sequence that is also found 30 to 100 bp upstream from five other flagellar genes, was found 66 bp upstream from the $a Y start site. The entire Jla Y upstream regulatory region and promoter sequence lies within the sequence encoding the 30 carboxyl-terminal amino acid residues of flaE. From analysis of the nucleo- tide sequence, examination of codon usage, and the G +C content of possible reading frames, it seems probable that flaE encodes a 16,000 M, protein and jZaY encodes a 70,000 M, protein. Translational fusions using an aphA reporter gene were con- structed to confirm the predicted reading frames. Extensive secondary structure in the region of overlap between the two genes suggests t’hat they may require complex regulation to perform their role in the trans-acting flagellar hierarchy (Champer et al., 1987).
Table 1 Bacterial strains
Strain Genotype Reference
A. C. crescentus (‘Ulfj Wild-type Poindexter (1964) (‘B1.W Wild-type (syuchronizable) Evinger & Agabian
(1977) SC1062 &El78 : : Tn5 prod. str-104 Purucker r/ a/.
(1982) SC’1133 ,flaY184 : : Tn5 cysB, str-142 Pururker ~1 ccl.
(1982) SC1121 ~IaYl83 : : Tn5 cysB. str-142 Purucker d al.
(1982) AE8001 jaY801 : : Tnj-L-1332 C:hamper rt d
(1987) s(‘274 flaYl29 [Tsl lap rt al. ( 1984)
13. E. coli HIS101 rK. mU, rprA 13, sull, leuB6, (‘old Spring Harbor
13 l-, proA 2, lacZ4. SmR Laboratory
21 Ci/mmol; Schwarz mixture) was from Schwarz-Mann. [y-J’P]ATP (> 3000 Ci/mmol) was obtained from Amersham Corporation. Staphylococcus A cells were purchased from Bethesda Research Laboratory. RNA used for nuclease S, assays was extracted with phenol at 65”C, as described (Amemiya et al., 1980).
(c) DNA manipulations
Conditions for restriction endonuclease digestions werp those recommended by commercial suppliers. Ligations were performed at DNA concentrations of 20 to 50 pg/ml in 10 mM-Tris HCl (pH ‘7.5). 10 mM-Mgcl,, 10 mM-2- mercaptoethanol, and 05 mM-ATP for 2 to 12 h at 12°C. Deletions in bacteriophage Ml3 DNA were generated as described by Dale et al. (1985). Other cloning methods were carried out as described by Maniatis et al. (1982).
(d) Construction of pRBl0
2. Materials and Methods
(a) Bacterial strains
The bacterial strains used in this study are listed in Table 1. C. crescentus strains were grown in peptone yeast extract medium or M2 minimal glucose medium at 30°C (Contreras et al.. 1978). E. coli strains were grown in LB medium at 37°C.
(b) Materials
Restriction enzymes were obtained from Kew England Biolabs and Boehringer-Mannheim. Calf intestine alkaline phosphatase, phage T4 polynucleotide kinase and nuclease S, were purchased from Boehringer-Mannheim. 14C-labeled reconstituted protein hydrolysate (RPH.
pRBl0. a plasmid containing the jZaE and $aY genes but with the JIUE promoter region deleted (Fig. 1). was
constructed in several steps from pRB3. pRB3 is a pRK290 plasmid that contains a 10 kb EcoRI insert extending from the EcoRI site injgK (see Fig. l(a)) to the next EcoRI site 10 kb downstream (Fig. l(a)). This insert encodes a portion of jlgK, and the entire jlgJ, &E and jZaY genes (Fig. 1; Bryan et al., 1987). pRB3 was partially digested with EcoRT and ligated with EcoRI-digested. phosphatase-treated pBR322. The resulting plasmid contained both pRK290 and pBR322 and the pRB3 insert. This plasmid was then digested with Hind111 and religated to form an insert that extends from the EcoRI site in jlgK to the HindITI site to the right of JlaY (Fig. l(a)). The resulting plasmid pRB9 (Fig. 1 (c)) was digested with BcZI and religated to yield a plasmid that contains a small piece of pRK290. a portion of jZaE beginning at the BcZI site, the entire $a Y gene, and most of pBR322. This plasmid was then digested with Hind111 and ligated into pRK404 (Dittn et al.. 1985) t’o yield pRBl0 (Fig. l(c)).
(P) Z)XA sequence determination und anulysie
t Abbreviations used: bp. base-pair(s); kb, lo3 bases or The source of DXA for sequencing determinations of base-pairs; RPH. reconst,ituted protein hydrolysate: the C. crescentus $aE Y operon was plasmid pBR3 (Bryan NPT. neomycin phosphotransferase. et al., 1987). This plasmid contains a 10.8 kb EcoRI

Organization of Two Caulobacter Flagellar Genes 73
Flogellins
(a)
sc14 Sk3 A .A ,ol, flaE - flay ak’es (cl Kx2 1133 1121 8001 274
pRB9 %--A H - ----
u - + t + + t Bc
pRBl0 ” I H I - --mnd.
Figure 1. (a) Physical and genetic map of a C. crescentus flagellar gene cluster showing 3 flagellin encoding genes, $gL, jZgK and jZgJ, and the 2 regulatory genes, jZaE and flay (Bryan et al., 1987). The open arrows represent the protein coding regions and direction of transcription. The numbers below the flagellin genes indicate the size (in x lo3 M,) of the encoded proteins. The line below represents the physical map of an 11.9 kb fragment containing these genes. The vertical lines represent the locations of restriction sites (H, WindIII; R! EcoRI; S, SalI; Bc, BcZI; B, BarnHI; X, XhoI). There are additional EcoRI, SalI, BarnHI, XhoI, Hind111 and BcZI sites outside the central SaZI-HindIII fragment (see Purucker et al., 1982; Minnich & Newton, 1987). (b) Enlarged map of the 3413 bp SalI-HindIII fragment encodingJEaE and fla Y. The open boxes represent the gene coding regions and the broken lines above the map represent the start site, extent, and the direction of transcription. The triangles below indicate the locations of 3 Tn5 insertion mutations (SC1062, SC1 133 and SC1 121; Bryan et al., 1984, 1987), and one Tn5-VB32 insertion mutation (AE8001; Champer et al., 1987). The location of a temperature-sensitive (Ts) mutant in flu Y (SC274) was mapped to the left of t,he XhoI site by its inability to form motile recombinants in the presence of pRB4. Motile recombinants were found in the presence of pRB6 (unpublished results). (c) Clones used for complementation analysis. pRK290 (Ditta et al., 1980; Bryan et al., 1987) is the vector for pRB2 and pRB4. pRB6 and pRB9 contain both pRK292 and most of pBR325 (Bryan et al., 1987). The open regions in pRB4 and pRB6 represent deleted regions. The triangle in pRB2 shows the location of the Tn5 insert (from SClO62). pRBl0 cloned into pRK404 (Ditta et al., 1985) and its construction is described in Materials and Methods. Complementation of the JEa Y and the jlaE alleles was measured by observing motility in the light microscope and by determining whether large diffuse colonies form on 0.3% (w/v) semi-solid agar, as described (Bryan et al., 1987).
restriction fragment that spans the region from the internal EcoRI site in jZgK (Fig. l(a)) out to the EcoRI site distal to the end of the flay gene in a pRK290 vector. Plasmids pRB2 (Bryan et al., 1987) and pSP3 (Purucker et al., 1982) were used as a source of restriction fragments for DNA sequence analysis of the insertion site of jZaE178: : Tn5 (SC1062) (Figs l(a) and 2(c)). DNA sequence analysis of the insertion site ofJla Y : : Tn5-VB32 (AE8001: Champer et al., 1987) was performed by subcloning a 900 bp PatI fragment containing the &Y/Tn5-VB32 junction int*o the P&I site of M13mp9 (Fig. 2(c)).
All DNA sequences were determined completely on both strands of the DNA. In addition. all restriction sites were overlapped by sequence determination from different DNA fragments. Thus, no sequence information was missed as a result of the loss of a very small restriction fragment. DNA sequences were determined using the dideoxy nucleotide chain termination method of Sanger et al. (1977) with the modifications for G+C-rich DNA described by Kaplan et al. (1985). The polyacryl- amide/urea gel electrophoresis system described by Sanger & Coulson (1978) was employed. DNA sequence analysis was facilitated by computer programs (Queen & Korn, 1980; J. Kaplan. unpublished results).
(f) Xuclease S, protection studies
Nuclease S1 mapping of the juE Y mRNA transcrip- tion start site was performed as described by Berk &
Sharp (1977). The probe for mapping the start site was a 408 bp PvuII~TupI fragment (bp 328 to 736; Fig. 3) 5’- end-labeled at the TuqI end with [y-3ZP]ATI’ (Fig. 4). C. crescentus RNA hybridization and nuclease S, diges- tion were as described by Loewy et al. (1987). Protected DNA fragments were resolved on So/b (w/v) polyacryl- amide sequencing gels containing 7 M-urea.
A DNA sequencing ladder for mapping the precise 5’ end of the JlaEY mRNA transcript was generated as follows, First, the 408 bp PvuTT-TapI fragment used as a probe was cloned into the &‘maI/AccI sites of M13mp8. This phage was used as a template for standard dideoxy nucleotide sequencing reactions, except t,hat the chain elongation reactions were terminated after 3 min by the addition of EDTA to a final concentration of 50 mM. Next, each of the 4 sequencing reactions was precipitated with ethanol and digested with TugI. Finally, the restriction endonuclease digestions were precipitated with ethanol, dried under vacuum and resuspended in 15 ~1 of deionized formamide containing 0.01 y/, (w/v) xylene cyanol. A small portion of each reaction was loaded adjacent to the S, nuclease-protected products. Each of these 4 dideoxy nucleotide sequencing reactions revealed a set of nested fragments originating at the same 7’aqI site that was labeled in the probe.
(8) CeZZ synchronization
Derivatives of C. crescentus CB15N were grown in M2 minimal glucose medium and synchronized following

74 J. B. Kaplan et al
centrifugation in a Ludox gradient as described (Evinger & Agabian, 1977; Mansour et al.. 1980). Swarmer cells were washed, diluted into fresh M2 medium, and allowed to proceed through the cell cycle at 30°C. At various stages in the cell cycle, as indicated in the legend to Fig. 4, portions were pulse-labeled with 15 &i of [14C]recon- stituted protein hydrolysate (RPH; 21 Ci/mmol: Schwarz mixture) for 20 min at 30°C. Alternatively. unlabeled portions were taken from synchronous cultures and the cells were collected by centrifugation, washed twice with M2 medium, and used for the extraction of RNA.
(h) Immunoprecipitation
The synthesis of the flagellins or neomycin phospho- transferase II (NPT II) was measured by immuno- precipitation of [ “C]RPH-labeled cell extracts with rabbit antisera against flagellin (Shapiro et al., 1982) or NPT II (Champer et al., 1985). Labeled cells were lysed and immunoprecipitated as described (Shapiro et al., 1982). Samples were analyzed by SDS/polyacrylamide gel electrophoresis and proteins were visualized by autoradiography.
3. Results
(a) DNA sequence of the flaEY gene cluster
The organization of the JEaEY gene cluster and the upstream flagellin genes, jigL, flgK and JlgJ, is shown in Figure l(a). The transcription of each of
the flagellin genes is initiated from its own promoter (Loewy et a2.. 1987; Minnich &, Newton. 1987). In order to determine if the jZaE Y genes encode a polycistronic message and to define the regulatory elements upstream and within this gene cluster, t,hr DNA sequence of a 3413 bp SalI-Hind111 fragment containing the jlaE Y gene cluster along with the 5’ and 3’-flanking regions was analyzed. Figure 2 shows the strategy used to determine the DNA sequence. The sequence of both strands was determined and all restriction sites were overlapped at least’ once. The complete DNA sequence and predicted amino acid sequence of the jlaEI’ gene cluster along with a portion of t,he upstjream ,flgJ gene is presented in Figure 3.
Analysis of the jaE Y DNA sequence revealed a t,otal of 19 different’ open reading frames (in both directions) that begin with an ATG or GTG start codon, terminate with a TAA, TGA or TAG stop codon and are capable of encoding polypeptides of at least 50 residues. Each of these open reading frames was further analyzed with respect to base composition. C. crescentus DNA is composed of 67 o/o G+C (Wood et al., 1976). It has been shown that in species with genomes of high G+C cont,ent. protein coding regions exhibit. a periodicity of base usage, being moderately G-t-C-rich in the first codon positJions, low in the second codon positions and very high in third codon positions (Kaplan et
fW (a) (b)
k-525 bp+-4~+j~&------ 2640 bp 4383 bp-l
HrtdIlI 15 bp I
SdI 7 __Jr---
BunHI I
I -_-..-e
Pstr
Htn-3 I II 11 I I
/!&I r--
NueI II I I I I II
Mu1 I I I r-
Xtld _----I
SWUI I IO II III I I Il7lllll 1
Hpolr --III I F-KC3 II UI I I I Ulll I II I I I I IH
TagI n-- lnlm I I I II I I
HmPI J III~UI~IIO II 0 I II ill I II l 0 I II I II I I w 111 I !
Figure 2. DNA sequencing strategy, (a) Schematic diagram of the $aEY gene cluster. The open arrows on top indicate the flaE and fEa Y coding regions and direction of transcription. The sizes of the flanking and coding regions are indicated below. (b) Maps of Ml3 subclones. The vertical lines represent the locations of restriction sites (indicated on the left) used to generate fragments for DNA sequence analysis. The arrows indicate the extent and direction of sequence determined from each cloned fragment. The broken lines indicate the extent of sequence determined from phage T4 polymerase-generated deletions. (c) Method used to determine the junction sequences of 2 different Tnj insertion mutations. The hatched boxes represent the insert,ion elements and the thin lines represent the chromosomal sequences. The arrows indicate the extent and direction of sequence determined from restriction sites (indicated above the boxes) near the insertion sites.

Organization of Two Caulobacter Flagellar Genes 75
Figure 3. DNA and predicted amino acid sequences of the $aEY gene cluster. The sequence shown includes the last 9 codons of$gJ. a 659 bp intercistronic region, the 450 bpflaE coding region. a 15 bp intercistronic region, the 2040 bp ~‘Zfla Y coding region and 383 bp of 3’.flanking region. The sequence of the sense strand is given in the 5’ to 3’ orientation. The nucleotides are numbered below the sequence. The last digit of the number appears directly below the nucleotide. The sequence from bp 1 to 167, which contains a portion of the 5’ end of&J, was reported by Gill & Agabian (1983). The DKA sequence from bp 162 to 3574 was determined in this st,udy. These 2 sequences were overlapped and the sequence of bp 1 t.o 167 was confirmed by sequencing leftward from the BcZI site at bp 259. The locations of restriction sites discussed in the text are boxed, as are the$uE and $aY start and stop codons. Dyad symmetries are indicated by, horizontal arrows above the sequence. The locations of the major transcription start sites are indicated by + 1. The locations of 2 different Tn5 insertion mutations, from strains SC1062 and AE8001, are also shown. The 13 bp consensus sequences are underlined.
al., 1985; Crawford & Eberly, 1986). Only two of Several additional lines of evidence suggest that the 19 open reading frames exhibited this pattern of these two open reading frames represent the flaE base usage (Table 2), suggesting that they in fact and flay protein coding regions. First! the pattern c-orrespond to expressed genes. of codon usage (Table 3) correlates highly with the

76 J. B. Kaplan et al.
Table 2 Rase usage in flaE and flay codons
flaE jlaY CaulobacW (G +c contentc)
Pseudomon&
1st base 61 61 66 67 2nd base 51 51 48 45 3rd base 84 83 X6 86 Bverage 65 65 67 66 Total codons 149 679 3209 3932
“Average G + C content of 11 different C. crescentus genes including trpF, trpB, trpA and usg (Ross & Winkler, 1988); flgJ (Gill & Agabian, 1983); flaD (Hahnenberger & Shapiro, 1988); j&J, $aJ, jlaH and jlah’ (Mullin & Newton, personal communication); and aodA (Steinman, personal communication).
bAverage G+C content of 9 different, Pswdomonas aeruginosa chromosomal and plasmid-encoded genes (taken from Crawford & Eberly (1986)).
‘The numbers are VO G +C.
codon usage patterns and amino acid composition plasmids carrying deletions in the cloned $aEY of 11 other sequenced C. crescentus genes (see region suggest that, the coding region extends to the footnote a to Table 2). Second, complementation XhoI site at bp 3081, and that the internal SalI tests of chromosomal mutants in JlaEY with sites at bp 1793 are within the coding sequence
Table 3 Codon usage in flaE and flay
jlaE jiaY All
other’ jiaE 411
otherd
Ala GCY’ GCG GCT GCA
Asn AA( .4,4T
Asp GA( GAT
Arg ax CGG CGT (‘GA AGC AG A
Cps TG(’ TGT
Gin CAG CAA
Glu GAG GAA
Gly GGC GGT GGG GG.4
His CAC CAT
lie AT(’ ATT AT.4
11 29 348 3 26 121 I 4 28 1 1 12
5 19 96 0 6 21
4 37 162 2 12 44
3 14 97 2 16 27 1 6 PO 3 5 8 0 2 5 0 1 1
0 4 9 1 0 3
5 18 103 0 8 15
6 15 86 1 5 53
11 55 225 0 12 25 4 12 23 1 5 12 2 2 3 0 3 8
5 14 127 1 2 6 0 0 0
Leu CTQ CT<’ TTG CTT (‘TA TTA
Lps AAG AA.4
Met ATG
Phe TT(’ TTT
Pro CCG VW C‘(‘T (:(‘A
Ser TC(’ TV; A(:( TCT TCA AGT
Thr AW 4CG ACT AC.4
Trp TGG
TW T8( TAT
Tal GT(’ GTG OTT GTA
7 6 2
0
3
1
,- ;I
4 17 103 3 I1 26 6 19 24 0 d 9 0 1 4 0 4 3 6 44 1 .i7 4 23 73 1 0 8 0 4 0 2 6 I9
28 2
12
14 2
11 8 5 0
1-l 6
175 46 13 IO 2 0
I IO 9
I 62
71 13
98 38 29
3
26 20
136 xx “3
3
“The number in this column indicates the total number of times a rodon appears in 3209 codons from 11 different (‘. crescentus genes as described in footnote a to Tahlr 2.

Organization of Two Caulobacter Flagellar Genes 77
(Fig. l(c)). Third, genetic analysis of Tn5 insertion mutations in this region (Bryan et al., 1987) indicates that the direction of transcription of $aE and jIaY is the same as that of other genes in the cluster (Minnich & Newton, 1987; Loewy et al., 1987). Taken together, these data suggest that the
jIaE and flay translational phases shown in Figure 3 are correct.
These results were confirmed by constructing gene fusions linking either the JlaE or $a Y gene to a.phA (the Kn’ (kanamycin resistant) gene from transposon Tn501; Vakulenko et al., 1987) in all three reading phases (Kaplan, Bryan & Loewy, unpublished results). One set of gene fusions linked the JlaE gene at the HpaII site (bp 867 to 870; Fig. 3) to uphA, and another set linked the $aY gene at the BamHI site (bp 1592 to 1597; Fig. 3) to aphA. These gene fusions were cloned into pRK291 (a plasmid capable of replicating in C. crescentus), mated into C. crescentus, and screened for their ability to confer a Kn’ phenotype. Only those gene fusions linking the genes in the predicted reading phase shown in Figure 3 resulted in a Kn’ phenotype, confirming the reading phase of the jlaE and ,&la Y genes.
Examination of the JlaE Y DNA sequence shows that JlaE and fray are indeed closely linked, being separated by only 15 bp of DNA. There are, however, 659 bp of DNA between the end of JlgJ and the beginning of jlaE (bp 28 to 686; Fig. 3). Analysis of this intercistronic sequence failed to reveal any significant open reading frames that
conform to patterns of base usage and codon usage seen in other C. crescentus genes. However, at - 140 bp from the start of transcription of the jlaE gene a 13 bp consensus sequence is present (Fig. 3) that may play a role in the regulation of fEaE and $aY gene expression. The 13 bp consensus sequence is also found between - 35 and - 110 of at least five other flagellar genes (Table 4). Our initial analysis of transcription from plasmids carrying portions of the upstream region and an adjacent reporter gene suggests that the sequences upstream from the PvuII site at bp -328 are not required for flaE expression (Kaplan, Loewy & Bryan, unpublished results).
The DNA sequence data presented in Figure 3 predict the primary structure of the FlaE and Flay proteins. The jlaE gene encodes a protein having 149 amino acid residues with a predicted molecular mass of 15,854 daltons. The $u Y gene encodes a 679 residue protein with a predicted molecular mass of 70,068 daltons. The distribution of amino acid residues in the predicted FlaE and Flay proteins is not significantly different from the average distribu- tion of residues in 11 other C. crescentus proteins derived from DNA sequence data (see footnote b in Table 2), providing further evidence that the predicted jZaE and jlaY translational phases represent expressed genes.
The precise site of insertion of two different Tn5 transposons in the jZaE Y region, JlaE: : Tn5 (SC1062; Purucker et al., 1982) and flay: : Tn5-VB32 (AE8001; Champer et al., 1987),
Table 4 The 5’ regulatory regions of tla genes
13.mer consensus Sigma 28 promoters
- 109 AGGCTTAGGCCGT
- 140 T(:GTTT(‘CGT(‘CC jr,
-66 AGGTTTC’C(‘GCTT
-54 A(:(:(‘TOTG(:(I(‘GT
-35 AGGTTTAGGTCGT
f --.
-99 .~(;(:C‘(“r(‘l‘AA(‘GT
-
-10 GCCGAAAATC’T E. coli tsra
(‘GGCAAAATCGjgJb
C’CGCTTAATCT &EC (‘CGCAAAATCT consensus
“nif’ promoters 24 -13
TGTCCAG N, TGTC jZuY’
AGGCCCA N, TTGC jZbN*
TGGCCCG N, TTC,C Hook P,, TGGCCCA N, TTGC fig,!,’ TGGCCCA N, TTG(’ &K’
CGGCCGA S, ATGC ~. flaIP
TGGC’CC A
Y TTGC consensus Gi4
a Frederikse & Shapiro (unpublished results). ’ Gill & Agabian (1983). ’ This study. d Dingwall. Hahnenberger & Shapiro (unpublished results) e (‘hen et al. (1986). ’ The DN.4 sequenw is not available. B Minnich 6 Newton (1987): Mullin el nl. (1987). h Hahnenbwger Cys Shapiro (1988).

78 J. B. Kaplan et al.
were determined by subcloning restriction frag- ments containing the right or left junction regions into Ml3 for DNA sequence analysis. The strategy used to determine the junction sequences is shown in Figure 2(c). Both the right and left junctions of the insertions in SC1062 were determined, and a duplication of the 9 bp sequence 5’-CGGCCTTGA-3 (bp 1066 to 1074; Fig. 3) was found at either end. This sequence lies within the predicted jaE coding region and provides further evidence that this coding region represents the flaE gene. The left junction of the Tn5-VB32 insertion mutation in AE8001 was determined as described in Materials and Methods. This element was found to be inserted in the predicted flay coding region after bp 2748 (Fig. 3). The sequence at the right junction of AE8001 was not determined.
(b) Transcriptional regulation and complementation analysis of the
flaEY gene cluster
In order to define the functional unit or units containing the JlaE and flay genes, portions of these genes, shown in Figure l(c), were cloned into broad host range plasmids capable of replicating in C. crescentus. These plasmids were then used to complementation test variousJlaE and $a Y mutant strains (Fig. l(b) and (c)). The insert in plasmid pRB9 contains intact jIaE and flay genes, and pRB9 was able to complement all jZaE and $aY mutant strains. A plasmid, pRB2, containing flaE : : Tn5 from SC1062 and an intact flay gene, was unable to complement several chromosomal mutations infla Y (Fig. l(c)). This polar effect of the flaE : : Tn5 insertion suggested that JlaE and $a Y might form a polycistronic transcriptional unit. Furthermore, a plasmid, pRB10, with a deletion of the 5’ regulatory region in front of&E and a small portion of the 5’ coding sequence of $aE, but with an intact ja Y gene, was unable to complement jZa Y alleles. Conversely, plasmids pRB4 and pRB6, which contain deletions in $a Y, were unable to complement SC1062, indicating that the expression of the chromosomal copy of $a Y is blocked by the presence of jZaE : : Tn5 in cis.
To determine if a transcript spanning both genes could be detected and to measure the time of expression of the JlaE and $a Y genes as a function of the cell cycle, nuclease S, protection assays were done using a 1436 bp SalI-BamHI fragment labeled at the 5’ end (Fig. 4(a)). The large fragment was chosen because it extends beyond theJlaE gene into flay (Fig. 4(a)). The size of the RNA fragment protected by hybridization to the SalI-BamHI fragment should be 930 bases if the two genes are read together to form a polycistronic message. A protected fragment of that size was observed (Fig. 4(b)). Using portions of RNA from cells at various times in the cell cycle, the 930 bp protected fragment was first detected at 0.5 division unit, but increased in abundance at O-66 and 0.83 division
unit. Therefore, the transcript originating from the flaE gene extends into the flay gene and is temporally controlled. The SalI-BamHI fragment also protected two smaller RNA fragments of approximately 470 bases. The presence of these protected fragments at 470 bp suggests that either there is an internal promoter site that can allow initiation of a flay transcript, or that the smaller RNAs are processed fragments from the poly- cistronic message. The larger RNA fragment in the doublet appears at the same time in the cell cycle as does the 930 bp protected fragment? whereas t,he smaller fragment in the doublet firpt appears earlier, at 0.25 division unit. The start of these transcripts coincides with a region in front of the $aY gene that contains a “nif’ promoter consensus sequence and an upstream 13 bp consensus sequence that is present in front of several $a genes (Fig. 5). Both the promoter sequence and the 13 bp consensus sequence lie within the 3’ coding sequence of t,he adjacent jlaE gene.
Many C. crescentus genes lack an E. c&like Shine-Dalgarno sequence immediately upstream from the translation start site (Ross & Winkler, 1988; I>. Mullin $ A. Newton, personal communication). However, the Shine-Dalgarno-like sequence 5’-AAGGGT-3’, in which four out of five nucleotides could theoretically form pairs with the 3’ end of 16 S rRNA from C. crescentus (Feingold et al., 1985), lies upstream from the flaE start codon (bp 681 to 686; Fig. 3). No apparent Shine- Dalgarno-like sequences are found preceding the $aY translation start site (Fig. 5). Instead, as is often the case in C. crescentus, the protein coding region is immediately preceded by a region capable of forming a complex secondary structure (Fig. 5). This region could form any one of several stable secondary structures with an average calculated free energy of formation (AG) of -28 kcallmol (1 cal=4.184 J). Two 10 bp and three 7 bp regions capable of forming stem and loop structures are indicated in Figure 5(a) and one possible RNA configuration is shown in Figure 5(b). The locat’ion of the “nif’ promoter consensus sequence, the jlaE translation stop signal and the flay translation start, are shown in the DNA sequence and then indicated again in the mRNA transcript in Figure 5. A very similar secondary structure configuratiorl. with alternate modes of folding, exists in the overlap region between the basal bodr $a11 and flblV genes (Dingwall & Shapiro. unpublished results).
In order to determine the precise t,ransoription start site in front of the first gene of the cluster. jaE, nuclease S, experiments were performed using a 5’-end-labeled probe isolated from t,his region. The results of this experiment are shown in Figure 4(c). When a 408 bp PvuII-TagJ fragment labeled at the 5’ end of the non-coding strand (Fig. 4(a)) was hybridized to total cellular RNA from P. crescentus strain CB15N, a doublet approximately 85 bp in length was detected following treatment, with nuclease S, (Fig. 4(c)). By running a DNA
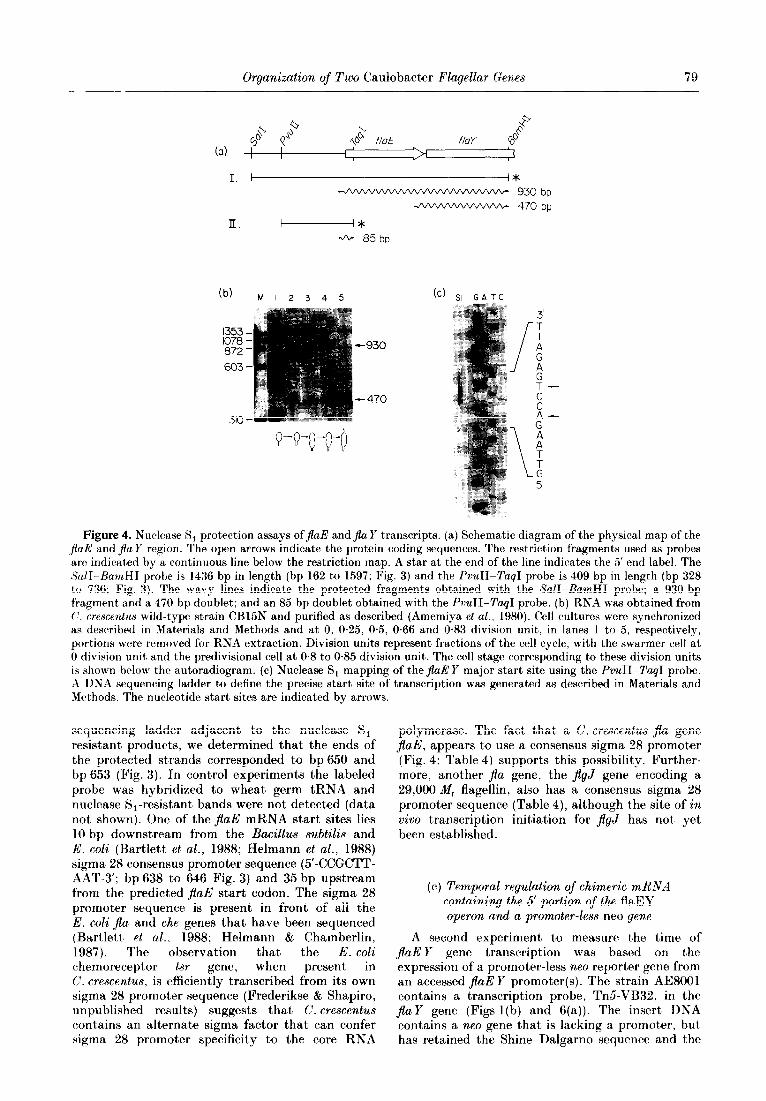
Organization of Two Caulobacter Flagellar Genes 79
-JVVVVVVV\n/V\A/\r\/ 930bp - 470bp
(b) M I 2 3 4 5 (cl SI GATC
1% 1 872- -930
603-
Figure 4. Nuclease S, protection assays of&E andJla Y transcripts. (a) Schematic diagram of the physical map of the $aE: and &Z Y region. The open arrows indicate the protein coding sequences. The restriction fragments used as probes are indicated by a continuous line below the restriction map. A star at the end of the line indicates the 5’ end label. The SW-HamHI probe is 1436 bp in length (bp 162 to 1597; Fig. 3) and the PvuTI-Tag1 probe is 409 bp in length (bp 328 to 736: Fig. 3). The wavy lines indicate the protected fragments obtained with the SalI-BarnHI probe; a 930 bp fragment and a 470 bp doublet; and an 8.5 bp doublet obtained with the PvuII-TaqI probe. (b) RNA was obtained from (‘. crescentus wild-type strain CB15N and purified as described (Amemiya et al., 1980). Cell cultures were synchronized as described in Materials and Methods and at 0, 0.25, 0.5, 0.66 and 0.83 division unit, in lanes 1 t*o 5, respectively, portions were removed for RNA extraction. Division units represent fractions of the cell cycle, with the swarmer cell at 0 division unit and the predivisional cell at 0.8 to 0.85 division unit. The cell stage corresponding to these division units is shown below the autoradiogram. (c) Nuclease S, mapping of the jlaE Y major start site using the PvuII-Tap-1 probe. A DNA sequencing ladder to define the precise start site of transcription was generated as described in Materials and Methods. The nucikotide start sites are indicated by arrows
sequencing ladder adjacent to the nuclease S,- resistant products, we determined that the ends of the protected st,rands corresponded to bp 650 and bp 653 (Fig. 3). In control experiments the labeled probe was hybridized to wheat germ tRNA and nuclease S1-resistant bands were not detected (data not shown). One of the jlaE mRNA start sites lies 10 bp downstream from the Bacillus subtilis and E. coli (Bartlett et al., 1988; Helmann et al., 1988) sigma 28 consensus promoter sequence (5’-CCGCTT- AAT-3’; bp 638 to 646 Fig. 3) and 35 bp upstream from the predicted jlaE start codon. The sigma 28 promoter sequence is present in front of all the E. coli $a and the genes that have been sequenced (Bartlett et al.. 1988; Helmann & Chamberlin, 1987). The observation that the E. coli chemoreceptor tsr gene, when present in C. crescentus, is efficiently transcribed from its own sigma 28 promoter sequence (Frederikse & Shapiro, unpublished results) suggests that C. crescentus contains an alternate sigma factor that can confer sigma 28 promoter specificity to the core RNA
polymerase. The fact that a C. crescentus jla gene JlaE, appears to use a consensus sigma 28 promoter (Fig. 4; Table 4) supports this possibility. Further- more, another $a gene, the jlgJ gene encoding a 29,000 M, flagellin, also has a consensus sigma 28 promoter sequence (Table 4), although the site of in vivo transcription initiation for JlgJ has not yet been established.
(c) Temporal regulation of chimeric mRNA containing the 5’ portion of the flaEY operon and a promoter-less neo gene
A second experiment to measure the time of flaE Y gene transcription was based on the expression of a promoter-less neo reporter gene from an accessed $aE Y promoter(s). The strain AE8001 contains a transcription probe, Tn5-VB32, in the Jla Y gene (Figs 1 (b) and 6(a)). The insert DNA contains a neo gene that is lacking a promoter, but has retained the Shine-Dalgarno sequence and the

80 J. B. Kaplan et al.
Tn5-
(a) 1062 1100 t
C~TTTCCcGcTTfiCCIG5CGGCCTTG~c*~GGGGcTcGGcG*G*Tc~GcG~T~~~
“13-mer” -24
10 7 7 7 10 - -- e
CTAGACC@TCGC CAGCGACACIOGGCMMGTC (+l)
stop start
(b)
C-G A-U
1100 1115
Figure 5. (a) Nucleotide sequence of the overlap region between the jla,E and the jZa Y genes. The jlaE stop codon and jIa Y start codon are boxed. A “ 13.mer” consensus sequence and a “nif’ promoter sequence, both found upstream from several other C. crescentusja genes, are also boxed. as is a possible + 1 mRNA start site for the $aY transcript. The site of insertion of Tn5 in SC1062 is indicated by a vertical arrow at bp 1100. Inverted repeat regions are indicated with horizontal arrows and numbers. (b) One of several potential secondary structures in the mRNA derived from the sequence shown above
AUG translation start site (Bellofatto et al., 1984). A short sequence between the end of the inter- rupted $a Y gene and the beginning of the neo gene contains translation stop signals in all three reading frames, ensuring the synthesis of a chimeric mRNA but not a protein fusion. The precise site of insertion of Tn5-VB32 in the jIaY gene was determined by DNA sequence analysis, as described above (section (a)). In order to determine the time of expression of the chimeric mRNA as a function of the cell cycle, swarmer cells were obtained from strain AE8001 and allowed to proceed synchron- ously through one cell division cycle (Fig. 6(b)). At 0, 0.25, 0.5 and 0.8 division unit, portions of cells were pulse-labeled with i4C-labeled amino acids, cell extracts were divided and immunoprecipitated with either antibody to NPT II (the product of the
neo gene) or with anti-flagellin antibody (Fig. 6(b)). NPT II was first detected at 0.5 division unit and was fully expressed at 0.8 division unit. Flagellin synthesis detected in portions of the same cell fractions showed that the NPT II first appeared before the synthesis of the 25,000 and 27,000 M, flagellins, but coincident’ with the 29,000 M, flagel- lin. In the AE8001 strain, t,he 29,000 M, flagellin is over-expressed. The level of synthesis of the 25,000 M, and 27,500 n/l, flagellins is relatively low in AE8001 because the product of the $a Y gene is needed in trans for the normal level of expression of these flagellin genes (Purucker et al.. 1982; Champer et al., 1987). Comparison of the time of joEI transcription from the beginning of the operon (Fig. 4(b)) and the time of expression of the chimeric jiaE Y-neo gene shows them t’o be approximately

Organization of Two Caulobacter Flagellar Genes 81
(a) Tn5VB32 fhE
PzmRNA I
NPTII
(b) I 2 3 4 I23 4
Figure 6. (a) The cell cycle expression of the promoter-less NPT II reporter gene (Tn&VB32) inserted into the chromosomal $u Y gene in strain AESOOl is shown diagrammatically. The synthesis of NPT II and flagellins as a control was measured by immunoprecipitation of cell extracts obtained from synchronized populations pulse-labeled with “C-labeled amino acids using anti-NPT II antibody or anti-flagellin antibody as described (Champer et al., 1987). (b) The temporal expression of NPT II and flagellins in C. crescentus AE8001. The precise insertion site of TnSVB32 in $a Y of strain AE8001 is shown in Fig. 3. The disruption of the chromosomal fla Y gene by Tn5-VB32 causes down- regulation of expression of the flagellin gene expression (Champer et al., 1987). In (b) the lanes show immunoprecipitated protein from the specific stages of the cell cycle shown diagrammatically at the bottom of the Figure. The generation time in this experiment was 180 min. Lane 1, swarmer cell extracts (0 division unit); lane 2 st(alked cell extracts (0.25 division unit); lane 3 elongated stalked cell extracts (0.5 division unit); lane 4, predivisional cell extracts (0.8 division unit). K = M, x 103.
the same, suggesting that the chimeric gene is being read from the jlaE promoter.
4. Discussion
The large number of genes required for the biogenesis of bacterial flagella are subject to complex control mechanisms that order the time of expression and modulate the amount of gene product. Two C. crescentus flagellar genes, jlaE and jla Y, function in trans to modulate the level of expression of the genes encoding the flagellins fjlgL, jgK and jZgJ) and those encoding chemotaxis functions (Bryan et al., 1984, 1987; Champer et al., 1987). Physical and genetic analyses have shown that theJlaE and $aY genes are part of a cluster of ,$a genes that includes jlgL, JlgK and JlgJ (Purucker et al., 1982; Milhausen et al., 1982; Gill & Agabian, 1983; Bryan et al., 1984, 1987; Loewy et al., 1987; Minnich & Newton, 1987). These flagellin genes lie 3’ and adjacent t*o $aE and jla Y. Each flagellin gene is expressed from its own promoter and is temporally controlled at the transcriptional level (Loewy et al., 1987; Minnich & Newton, 1987). We report here the complete nucleotide sequence of the jlaE Y gene cluster and demonstrate that these genes are under temporal control. The two genes are read as a polycistronic message, but an internal promoter site appears to allow the separate transcription of the $aY gene. However, comple- ment’ation analysis of insertions and deletions in $a?3 and flay show a clear requirement for the expression of both genes in t,he cis orientation.
(a) Regulation of flaEY expression
Nuclease S, mapping of in vivo synthesized RNA transcripts was used to identify the JlaE and $a Y promoters. The polar effect of a Tn5 insertion in $aE on the contiguous $a Y gene and the inability of a clone of JlaE Y with the flaE promoter region deleted to complement mutant alleles of JlaY suggested that both genes might be transcribed on a single, polycistronic mRNA. The nuclease S1 protection experiments confirmed that a transcript originating upstream from flaE and extending through the intercistronic region between flaE and jlaY and into the flay coding sequence is synthesized in viva. The 5’ end of the transcript initiating upstream from jlaE was mapped to an A and a T residue shown in Figure 3. The promoter in the -10 region conforms to a consensus sigma 28 sequence (Table 4) used by jla and the genes in E. coli (Helmann & Chamberlin, 1987).
Two shorter nuclease S, -protected fragments, that either initiate just upstream from the JlaY coding sequence, or are processed products from the polycistronic message, were also detected by nuclease S, assays. The consensus “nif promoter sequence found in front of the JlaY gene would generate a transcript the size of the smaller protected fragments. The “nif’ gene promoter consensus sequence of Klebsiella pneumoniae is similar to several C. crescentus flagellar gene promoters (Table 4; Ausubel, 1984; Mullin et al., 1987). Although the precise 5’ end of the jIaY protected fragments was not mapped, this promoter sequence and a 13 bp consensus sequence (Table 4)

82 J. B. Kaplan et al.
at -66 bp from the putative transcription start sites lie within the 3’ coding sequence of JlaE (Fig. 5). Negative results in complementation tests of various chromosomal jz%Y mutants with a plasmid, pRB2, carrying JEaE : : Tn5 and an intact flay gene, showed that JlaE must be expressed in cis to allow complementation of $aY (Fig. l(c)). The cis-act’ing requirement of$aE was confirmed by the inability of plasmids bearing jaE Y genes with small deletions in $aY to complement SC1062, which contains JlaE : : Tn5 in the chromosome. The Tn5 in SC1062 (and pRB2) is insetted near the 3’ end of jaE, between the 13 bp consensus sequence and the “nif” promoter (Fig. 5). The disruption of the continuity of the 13 bp regulatory element and the “nif” promoter might’ account for the inability of pRB2 to complement jZa Y mutants (Fig. 1 (c)).
The region of overlap between the two genes has the potential to encode a mRNA with extensive secondary structure (Fig. 5). We are currently constructing mutations in the region of overlap in order to understand what function, if any, this internal promoter, upstream consensus sequence, and potential for secondary structure might have in controlling the differential expression of jlaE and JlaY. We have recently found that flaD and jbN, two C. crescentus genes which probably encode basal body proteins, are organized in the same overlap orientation as are t,he ,flaE and $a Y genes (Dingwall, unpublished results).
(b) Temporal order of$ageLlar gene expression
The bacterial flagellum is a complex, multi- component structure that is built on the surface of the cell with only its innermost component, the M-ring, embedded in the cytoplasmic membrane. In C. crescentus, the flagellum is assembled from the most cell-proximal portion, the inner rings of the basal body, followed by the rod and outer rings of the basal body, the hook, and then the cell-distal filament (Hahnenberger & Shapiro, 1987). In both C. crescentus and E. coli the genes involved in the biogenesis of the flagellum are controlled by a trans- acting regulatory hierarchy, with the first proteins assembled encoded by the genes nearest to the top of the hierarchy, and the last assembled, the flagellin components of the filament, at the bottom of the hierarchy (Komeda, 1982, 1986; Champer et al., 1985, 1987). In C. crescentus, the order of the flagellar genes in the regulatory hierarchy and the order of assembly of their encoded proteins reflects their time of expression as a function of the cell cycle. For example, the jlaD basal body gene is expressed before the JlaK hook gene (Hahnenberger & Shapiro, 1988), which is expressed before the flagellin genes (Ohta et al., 1985; Loewy et al., 1987). It therefore appears as if the temporal control of synthesis of the flagellar components contributes to the mechanism used by the cell to build this complex structure on the cell surface.
The jlaE and jla Y genes have been placed in the middle of the trans-acting hierarchy (Champer et al.,
1987). Although it is not known if the proteins encoded by these genes are struct’ural components of the flagellum, it is known that muta,nts defective in $aE and flaY fail to assemble a functional flagellum, and that normal levels of the flagellins and the chemotaxis proteins are not made. JEaE and jiaY function in trans to modulate the level of transcript’ion of Qa and the genes lower in t,hc regulatory hierarchy. We have shown here that $aE and JlaY are transcribed on a polycist,ronic message and that the appearance of this mRNA is temporally controlled. Using bot,h nuclease S, protection assays (Fig. 4) and expression of a ~PO reporter gene from a chimeric mR,NA (Fig. (i), WC’ found that the flaE Y genes are first expressed somewhat earlier than the flagellin genes a,nd continue to be expressed coincident with them. Because t,wo smaller RNA fragments were also detected by the nuclease S, assavs, and the fragments (which are very similar in sjze) map to a sequence in front of the $a Y gene that’ conforms to a “nif’ promoter consensus sequence, the possibility exists that under some conditions $0, I may be transcribed independently of $aE. The larger of the two protected fragments was det)ected coincident with the time of appearance of the jaE Y polycistronic message, whereas the smaller of the two was expressed earlier in t’he cell cyc*le (Fig. 4). The time of expression of the chimerich flaE Y-nro mRNA, as measured by immunoprecipitation of NPT II. was the same as that of the polycistronic mRSA and the larger of the doublet’ RNAs. The lack of appearance of t,he protected $air:Y poly- cistronic fragment at earlier times in the cell cvcle argues against) processing of that’ transcaript to $eld the $a Y mRNA unless the rate of processing varies during the coell cycle.
The results present a paradox. Genes that are VO- transcribed and function genetically as a cist,ron contain an internal promoter sequence that appears to be functional. Jf transcription from t,he $a Y promoter within%aE is real, it must in some way be insufficient for complete $a Y expression. E:xperi- ments are in progress to see whether the $a 1’ promoter is, in fact. read independently of the expression of ,flaE.
We thank Sandra Reuter for performing t’he car11 synchronies; Scott Siege for help with t,he c*omputer work: and Austin Newton. David Mullin, Malcolm Winkler and Howard Steinman for providing us wit)h sequence da& prior to publication. This investigation was support,ed b:, U.S. I’ublic Health Service grants GM11301 and GM32506 from the Xational Institutes of Health. J.B.K. was a Xational Institutes of Health postdoctoral ftllow and A.D. received predoctoral support, from National Tnst,itutes of Health training grant GM07491-07.
References
Amemiya, K., E&boy, 13. & Shapiro, I,. (1980). Virology, 104, 109-l 16.
Ausubel, F. M. (1984). (tell, 37, 5 -6.

Organization of Two Caulobacter Flagellar Genes 83
Bartlett, D. H., Frantz, B. B. & Matsumura, P. (1988). J. Bacterial. 170, 1575-1581.
Bellofatto, V., Shapiro, L. & Hodgson, D. (1984). Proc. Nut. Acad. sci., U.S.A. 81, 1035-1039.
Berk, J. A. & Sharp, P. A. (1977). Cell, 12, 721-732. Bryan, R., Purucker, M., Gomes, 6. L., Alexander, W. &
Shapiro, L. (1984). Proc. Nut. Acad. Sci., U.S.A. 81, 1341-1345.
Bryan, R., Champer, R., Gomes, S. L., Ely, B. & Shapiro, I,. (1987). Mol. Gen. Genet. 206, 3033306.
Champer, R., Bryan, R., Gomes, S. L., Purucker, M. & Shapiro, L. (1985). Cold Spring Harbor Symp. Quant. Biol. 50, 831~-840.
Champer, R., Dingwall, A. & Shapiro, L. (1987). J. Mol. Biol. 194, 71--80.
Chen, L.-S., Mullin, D. & Newton, A. (1986). Proc. Nut. Acad. Sci., U.S.A. 83, 2860-2864.
Contreras, I., Shapiro, 1,. & Henry, S. (1978). J. Bacterial 135, 1130-l 136.
Crawford. P. & Eberly, L. (1986). Mol. Biol. Evol. 3. 436-448.
Dale, R. M. K., McClure, B. A. & Houchins, J. P. (1985). Plasmid, 13, 31-40.
Ely, B., Gerardot, C. J., Fleming, D. L., Gomes, S. L.. Frederikse. P. & Shapiro. 1~. (1986). Genetics, 114, 717-730.
Ditta, G., Stanfield, S., Corbin, D. & Helinski, 1). R. (1980). Proc. Nat. Acad. Sci., U.S.A. 77, 7347-7351.
Ditta, G., Schmidhauser, T.. Yakobson, E., Lu, P., Liang, X.-W., Finlay, D. R., Guiney, D. & Helinski, D. R. (1985). Plasmid, 13, 149-153.
Ely, B., Croft, R. H. & Gerardot, C. J. (1984). Genetics, 108, 523-532.
Evinger, M. & Agabian, K. (1977). J. Bacterial. 123, 294-30 1.
Feingold, J., Bellofatto, V., Shapiro, L. & Amemiya, K. (1985). J. Bucteriol. 163, 155-166.
Gill, P. & Agabian, N. (1983). J. Biol. Chem. 258, 7395- 7401.
Gomes, S. L. & Shapiro, L. (1984). 1. Mol. Biol. 178, 551-568.
Hahnenberger. K. & Shapiro. L. (1987). J. Mol. Biol 194, 91-103.
Hahnenberger. K. & Shapiro. L. (1988). J. Bucteriol. 170,4119-4124.
Helmann. J. D. & Chamberlin, M. J. (1987). Proc. Nat. Acad. Sci., U.S.A. 84, 6422-6424.
Helmann, tJ. D., Marquez, L. M. & Chamberlin, M. J. (1988). J. Bacterial. 170, 1568-1574.
Johnson, R., Walsh, J., Ely, B. & Shapiro, L. (1979). J. Bucteriol. 138, 984-989.
Kaplan, J. B., Merkel, W. K. $ Nichols, B. P. (1985). J. Mol. Biol. 183, 327-340.
Komeda, Y. (1982). J. Bacterial. 150, 16-26. Komeda, Y. (1986). J. Bacterial. 168, 1315-1318. Koyasu, S., Asada, M., Fukuda. A. & Okada, Y. (1981).
J. Mol. Biol. 153, 471-475. Loewy, Z. G.? Bryan, R. A., Reuter, S. & Shapiro, L.
(1987). Genes Develop. 1, 626-635. Maniatis, T., Fritsch, E. F. & Sambrook, ,J. (1982).
Editors of Molecular Cloning: A Laboratory Munual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.
Mansour, J., Henry, S. & Shapiro, L. (1980). J. Bacterial. 141, 262-269.
Milhausen, M., Gill, P. R., Parker, G. & Agabian, x. (1982). Proc. Nut. Acad. Sci., C.S.A. 79, 6847-6851.
Minnich, S. A. & Newton, A. (1987). Proc. Nat. Acad. Sci., L’.S.A. 84, 1142-l 146.
Mullin, D., Minnich, S., Chen, L.-S. & Pu’ewton, A. (1987). J. Mol. Biol. 195, 939-943.
Nathan, P., Gomes, S. L., Hahnenberger. K., Zu’ewton, A, & Shapiro, L. (1986). J. Mol. Biol. 191, 433-440.
Ohta, N., Chen. L.-S., Swanson, E. & Newton. A. (1985). J. Mol. Biol. 186, 107-115.
Poindexter, J. S. (1964). Bacterial. Rev. 28. 231-295. Purucker, M., Bryan, R., Amemiya. K.,. Ely, B. &
Shapiro, L. (1982). Proc. Nat. Acad. Bci., l’.S.A. 79, 6797-6801.
Queen, C. L. & Korn. L. J. (1980). Methods Enzymol. 65, 595-609.
Ross, C. & Winkler, M. (1988). J. Bacterial. 170, 757-768. Sanger, F. & Coulson, A. R. (1978). FEBS Letters, 87,
107-l 10. Sanger, F., Nicklen, 8. & Coulson, A. R. (1977). Proc. Nut.
Acad. Sci., U.S.A. 74, 5463-5467. Shapiro, L. (1985). Annu. Rev. Cell Biol. 1, 173--207. Shapiro, L., Mansour, J., Shaw, P. & Henry, S. (1982).
J. Mol. Biol. 59, 303-322. Stallmeyer, M. J. B., DeRosier, D.. Aizawa, S. I.,
Macnab, R. M., Hahnenberger, K. & Shapiro, L. (1985). Biophys. J. 47, 48a.
Vakulenko. S., Miklos, K., Horvath, B. & Simonesits, A. (1987). Nucl. Acids Res. 15, 8111-8115.
Wagenknecht, T., DeRosier, D., Shapiro, L. & Weissborn, A. (1981). J. Mol. Biol. 151, 439-465.
Weissborn. A., Steinman, H. M. & Shapiro, L. (1982). J. Biol. Chem. 257, 2066-2074.
Wood, N., Rake, ,4. & Shapiro, L. (1976). ,J. Bacterial. 126, 1305-1315.
Edited by I. Holland

















![Torque Generated by Flagellar Motorof Escherichia - DAMTP · TorqueGenerated bythe Flagellar Motorof Escherichiacoil ... TES,N-tris[hydroxymethyl]methyl-2-aminoethanesulfonic acid.](https://static.fdocuments.us/doc/165x107/5c90c4f509d3f2c8148bd888/torque-generated-by-flagellar-motorof-escherichia-torquegenerated-bythe-flagellar.jpg)
