
Antiviral Effects of a Transgenic RNA-Dependent RNA- Polymerase

Plant Physiol. (1968) 43. 405-410
Template Requirement of Maize RNA Polymerase'Ernest R. Stout2 and Rusty J. Mans
Department of Botany, University of Maryland, College Park, Maryland
Receivecl December 28, 1967.
A bstract. Maize RNA polymerase utilizes heated deoxyribonucleic acid more effectivelythan native deoxyribonucleic acid as a template for ribonucleic acid synthesis. A ribenucleicacid-deoxyribonucleic acid hybrid accumulates in the presence of 'heated deoxyribonucleic acid.The amount of product formed with either native or heat-denatured deoxyribonucleic acid doesnot exceed the amount of deoxyribonucleic acid added as template.
The itemplate requirement of several RNA poly-merases ('Nucleoside-Htriphosphate: RNA nudleotidyltransferase, EC 2.7.7.6), isolated and purified frommicroorganism's (5, 7, 11, 41 ) anid mamtmals ( 13, 32),is best satisfied with native DNA. Heat denatured,calf thymus DNA actually inhibits the polymeraseisolated from bovine lymiphosarcoma tissue (12).In characterizing the partially purified enzyme frommaize seedllings we have shown that, lat relativelyhigh levels, native or denatured DNA from severalspecies satisfies the template requirement of themaize enzyme (42). The data presented heredemonstrate thait at rate limiting concentrations,denatured DNA from maize seed1lings or calfthym'us is 8 times more effective than native DNAin satisfying the template requirement. Further-more, an RNA-DNA hybrid, analogous to thatformed by Azotobacter vinelandii RNA polymerase(44), accutmulates u'pon incuLbation of the enzymewith 'sub'stra'te and denatured DNA. The hybridis not found w'hen native DNA is used as template.
Materials and Methods
Enzyme Preparationt and Assay. Soluble RNApolymerase was isolated f'rom 5-day-old maize seed-lings as described previously (42). The am'moniumsullfate precipitate was desalited on Sephadex G-50ra,ther than by di,a)lysis prior to column chroma-tography on diethylaminoethyl-(DEAE) cellu'lose.The enzyme had a specific activity of 1870. Spe-cific activity is def'ined as rnLmoles of AMP incor-porated in 10 minutes at 300 per milligram prote'in.
1 This investigation was supported in part by theUnited States Atomic Energy Commission (ReportNumber NYO-3536-8), and is Scientific Article No.A1400, Contribution No. 4009 of the M\aryland Agricul-tural Experimenit Station (Departmnent of Botany). Dr.Stou't was supported on a United States Public HealthService Postdoctoral Fellowship No. 5-F2-CA-28, 208-02from the National Cancer Institute.
2 Present address: Biology Department, VirginiaPolytechnic Institute, Blacksburg, Virginia 2-106'1.
RNA polymerase activiity was assayed as theincorporation of a-32P-labeled ribonucleotides fromribonucleoside triphosphates inito acid-insolulbleproduct at 300. A standard reaction mixture con-tained 40 wmoles tri's-Cl '('pH 8.4), 0.5 jxmoles eachof ATP, GTP, and UTP, 0.45 ,umoles a-32P-CTP(specific activity 17.4 mc/mmole), 5 ,umoles mag-nesium acetate, 5 1kmoles dith'iothreitol, 20 ,umoles(NH4)2S04, 50 jg DNA, and 50 jug of Fraction IVenzyme (42) in 0.2 ml. Samples were removedduring incubation and acid-insoluible radioactivitywas determined by the method of Boillum (2) asmodified by Mans and Novelli (26). Products wereisolated a,fteir the addiition of carrier yeaist RNAand HCl04. Base analyses were performed onDEAE-celtlulose paper 'by the method of Jacobson(20) as des-crilbed previously (42).
Biological Materials and Reagents. Calf thymusDNA and synthetic poilydeoxyribonucleotides weregenerously provided by F. J. Bollum. Maize DNAwas prepared from 5-day-old seedlings by themethod of Bolton (3), including treatment withRNase. A,ll other materials were pu.rchased fromcommercia4l sources. Maize grain (Zea mDays L.,WF9 X Bear 38) from Bear Hybrid Seed Company,Decatur, Illinois was germiinaited and the seedlingsutilized for all preparations. Uniform germinationand preparations of reproducible activities havebeen obtained with this grain for -more than 5 years.Pancreatic RNase (polyribonucileotide 2-oligonuoleo-tide-transferase, 2.7.7.16) 5 X crystallized fromGeneral Biochemicals, w;as heated for 10 minutesat 800 to inactivate DNalse. Pancreatic DNase (de-oxyribonuicleate oligonuic,leotide-hydro!lase, 3.1.4.5)electrophoretically pu.rified and firee of RN'ase waspurchased from Worthington Biochemical. Corpora-tion. The unlabeled rilbonuoleotides were obtainedfrom the following companies: P-L Biochemicals,Generall Biochemical's and Schwarz BioResearch,Incorporated. Labeled ribonucleotides were pur-chased, exclusively, from Schwarz BiooResearch,Incorporated.
All experimenits reported here were conduticted
405
Dow
nloaded from https://academ
ic.oup.com/plphys/article/43/3/405/6093852 by guest on 12 February 2022
PLANT PIIYSIOILOGY
wiith a single preparation of ithe appropriate DNAand indiv,idually 'frozen 'ajliqtuots of this preparationto avoid change in DNA template activity owingto storage, salt efifeots or pH (21). Simi,larly,individually frozen a,liquots of 1 RNA polymerasepreparatioin wvere ui.tilized. However, several dif-ferent 'preparations of DNA and RNA polymeraseexhibit all of the activities reported here.
Experimental Results
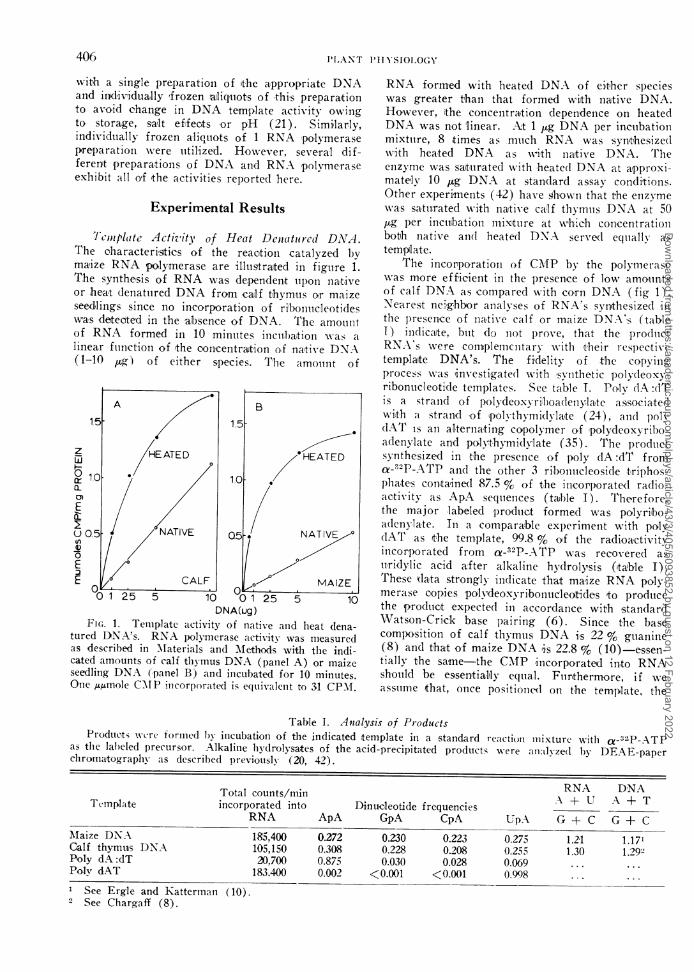
Temnplate Activity of Heat Denatured DNA.The characteritices of the reaction catalyzed byma,ize RNA polylmerase are illustraited in figuire 1.
T.he synthe,sis of RNA was dependent tll)Ol naitiveor hea,t clenattired DNA from cailf thymus or maizeseedlings since no incorporation of rilbonticleotideswvas detected in the 'absence of DNA. The amounttof RNA formed in 10 minltites inculbation was alinea.r fuinctioni of 'the concentration of native DNA(1-10 tug) of either spe,cies. T'he amoulnt of
A B
~~~~~~~1.5
EATED HEATED
0
E
U 0.5; NNATIVE 05
0
E 0
E CALF MAIZE~~~~~0
0 1 25 5 10 0 1 25 5 10DNA(ug)
FIG. 1. Temiiplate activity of native anid heat dena-tured DNA's. RNA polymerase activity was measuredats described in 'Materials and Methods with tle indi-cated amounts of calf thymus DNA (paniel A) or maizeseedling DNA (panel B) and incubated for 10 minutes.Onle ,u,mole CVIP incorporated is equivalent to 31 CPMI.
RNA formed with heated DNA of eit,her specieswas greater than that formed with native DNA.Howeve,r, ithe co-ncentration dependence on heatedDNA was not linear. Xkt 1 1ug DNA per inctubationmixture, 8 times as much RNA was synthesizedwith heated DNA as wvith naitive DNA. Theenzyme was saturated with heated DNA at a^pproxi°mately 10 ,Fg DNA at standard assay conditions.Other experiments (42) have shown that the enzymewas sattirated with native calf thymus DNA at 50,ag per incubation miixture at which con,centrationbo.th native and( heate(d DNA served equally- astemplate.
The incorporation of CI\IP by the poly-merasewas more efficient in the presen'ce of low am,ountsof calf DNA as compared with co,rn DNA (fig 1).Nearest neighbor anialyses of RNA's synthesized illthe presen,ce of native calf or maize DNA's (tableI) indicaite, bult do not prove, that the p)ro(ldlc'tRNA's were com'plemniitary with their respectivetemplaite DNA's. The fidelity of the copyilngIprocess was 'investigated with svnlthetic polydeoxy-ribonucleotide templates. See table I. Poly dA :dTis a straind of polydeoxYril)oadleny,la,te associate(lwith a strand of 'polythymidyla'te (24), aind poly(IAT is an alternaiting copolymer of polydeoxyribo-adenylate and polvthymidvlate (35). The productsynthesized in the presence of poly dA :dT froma-"2P-ATP and the other 3 ribonutcleoside triphos-p)hate's conitained 87.5 % of the incorporated radio-activity as ApA sequeniices (tab)le I). Therefore,the major labeled prodtlct formed was polyribo-adlenylate. In a comparable experiment with polyflAT as t,he tem,plate, 99.8 % o'f the radioactivityincorporated from ac-32P-ATP was recovered as
uridylic acid after alkaline hydrolysis (itable I).These data strongly indicate th'at ma'ize RNA poly-merase copies poWNdeoxyribonucledtides to producethe p}rodtlut ex-pected in accordance with standardWatson-Crick base pairing (6). Since the basecomposition of calf thymtis DNA i,s 22 % guaniine(8) and that of maize DNA 'i,s 22.8 % (10)-essen-tially the same-the CAMP incorporated into RNAshouild be essenltialily eqtal. Furnthermore, if we
assulme that, once positiotned oln the template, the
Table 1. Analysis of ProductsProduct; were ftormied by inicubation of the indicated template in a standard reaction imiixture with a-32P-ATPas the lal)eled precursor. Alkaline hydrolysates of the acid-precipitated products were analyze(d by DEAE-paperchromatography as described previously (20, 42).
Template
Maize DNACalf thymtus DNAPoly dA:dTPolv dAT
Total counts/minincorporated into
RNA ApA
185,400 0.272105,150 0.30820,700 0.875
183.400 0.002
Dinucleotide frequenciesGpA CpA0.2300.2280.030
<0.001
0.2230.2080.028
<0.001
RNA DNAA + U A+T
UpA G + C G+ C
0.2750.2550.0690.998
1.21 1.1711.30 1.292
I See Ergle and Katterman (10).2 See Chargaff (8).
406
Dow
nloaded from https://academ
ic.oup.com/plphys/article/43/3/405/6093852 by guest on 12 February 2022

STOUT AND AMANS-TEMP\'LATE REQIJlRErMENTS OF POLYMERASE
Table II. Dentatur-ationi of DNVA
HvperchromicityDNNA DNasel Heat2
Calf tlivmnus 30.8 17.0Mlaize seedliing 30.0 8.3
' 42 ,ug calf thymmus DNA or 46 pg maize seedlingDNA was incubated for 15 minutes with 100 pgRNase free DNase in 0J1 M sodium acetate (pH 5)xvith 15 ,mmoles magnesium acetate at 300 in a finalvolume of 3.0 ml. The change in absorbancy at 260nm was followed with time (23).
2 A stoppered tube containing 42 pkg calf thymus DNAor 46,ag maize seedling DNA in 1 ml SSC (28) atpH 7.4 was placed in boiling water for 10 minutesand then pluniged into crushed ice. The change inabsorbancv at 260 iim was measured at room temper-ature.
rate of polymerization for each of the ribonuc,leo-tide precuirso.rs i!s the same, we caniniot account forthe more efficient utilization o,f DNA observed forone species over that of anoither sipecies.
The differen,ce in template activity of the 2DNA's is correlated with a difference in responseof the DNA'is ito heat denaturation. Data in tableII show thalt denaturation of DNA of either speciesby trea'tment with deoxyribonuclease (23) reisultedin 30 % hyperchromi,city at 260 nm. However,heating the DNA of either species in a boiling waterbath followed by rapid cooling (the treatment tusedto prepare the theated DNA's uised in the experi-men.ts -of figure 1) resutillted in twice the hyper-chromicity (43) of callf DNA as seen with maize
5 ~ NATIVENE
00 10 20 30 0 10 20 30INCUBATION TIME (minutes)
FIG. 2. Time course of RNA synthesis with nativeand heat denatured DNA's. RNA polymerase activitywas measured as described under Materialis and IMethods,vith 1 pg of calf thymus DNA (panel A) or 1 pgmaize seedling DNA (panel B). One /qu,mole CMIP in-corporated is equivalent to 31 CPM1.
DNA (table II). It is Inot known how much ofthe difference in hyperchromici'ty between the 2heated DNA's is a function of the separation ofthe 2 strands (29), ithe random reassociation ofportions of the separated strands on cooling (9)and weakening oif the internucleotide stackingforces (1). However, the greater susceptability ofthe calf DNA to heat denatuiratioin does correlatewith greater template activity of the calf DNA(,fig 1). These data suggest that -the RNA poly-merase from maize requires single-stranded DNAas temipla'te. The activity seen with high levels ofnative DNA (42) may represen't a response of theenzyme to ,ow lev-els of contaminatiing denaturedDNA or to a slow denaturation of native DNAduring the incubation period.
Rate and Amnouint of RNA Synthesized. Thereaction rates of maize RNA polymerase on heatedDNA's are significantly faster than on nativeDNA's. The reaction slowed appreciably in 20minutes in the presence of heated maize DNA(fig 2B), whereas, the rate of CMP incorporationremaiined constant throughout the 30 minutte incuba-tion in the presence of naitive maize DNA. Essen-tially the same restult was obtained wilth calf DNA(ifig 2A). Hiowever, incorporation of CMP con-
tinued, butit at a reduced ra,te, throughouit the ilncuI-bation with heated calf DNA. Calctl.ating fromthe total radioactivity incorporated, the specificactivity o'f the a-32P-CTP laibeiled prectursor ulsed,and t.he base composiition of 'the product formed inthis incuibationi (fiig 2A and table I); 0.41 ,ug ofRNA was synthesized in the presence of 1 -g heatedcalf DNA in 30 miniutes. U'poIn longer incubationi,the amount of RNA synithesized did not exceed theamotlilnt of DNA templaite added. The slowing ofthe reaction rate, as the concentration of the produictapproached that of the heated telmiplate, suiggestedthat an RNA-DNA hybrid (4, 14, 17, 33, 36, 38) wasaccumulating. The formation of an RNA-DNAhybrid would also explain the faillture of pancreaticRNase to completely inhi'bit maize RNA polymerasewxhen inc'luded in the reaction mixtutre or 'to com-
pletely digest the acid-insoluible product (27) sinceRNA-DNA hybrids are resistant to pancreaticRNase ( 18).
Formation of RNTA-DNA Hybrid. Evidencewas sought for the accumuflation of aIn RNase re-
sistant hybrid as a product of the maize RNApolymerase re.action. -More -than 90 % of the RNAsynthesized with native ca,lf or maize DNA was
suscepitilble to digestion with pancreatic RNase(table III). However, in the presence of heatdenattu-red DNA's more 'than half (73.2 % with calfand 58.6 % wi.th maize) of the radioacti-ve productformed in 45 minuttes was resistant to RNase di-gestion. These ressults are analogous with those ofWarner et al. (44) who demonstrated 'the formationof RNA-DNA hybrid with single-stranlded DNAand purified RNA polymerase from A-otobactervinelandii. Early in the inculbation of the maize
407
Dow
nloaded from https://academ
ic.oup.com/plphys/article/43/3/405/6093852 by guest on 12 February 2022

PLANT PHYSIOLOGY
Table IIJ. Hfvbrid Foriatiol1Standard 0.2 m.l reaction mixtures containing 5 ,ug
of the indicated DNA were incubated at 300 for 45minutes. 50 ,umoles of sodium acetate (pH 5), 5/hmolles EDTA and 20 gg of DNa.se-free RNase wereadded and the incubation continued for an additional30 minutes. Changes in acid-ins-oluble radioactivitywere followed throughout the first and second incuba-tion periods. The data reported wvere calculated as:Total insoluble 32p rem-naining (2nd ilncubation)
X 100Tota.l 32p incorporated (Ist incubation)
Template DNA
Calf thynmutsM\a,ize see(llinig
RNase resistant productNaltive Denatured
8.14.2
E
(0~oU0.5zo
0C,)ED
73.258.6
enzyme with denatuiredl DNA, mlore than 90 % ofthe product was RNase resistant.
Additional evidence in suipport of the acctlnlti1a-tion of an RNA-DNA hybrid in the presence ofheated DNA was obtaine(d by suicrose densityg-radient centrifugation of the plro(llict. The acid-insolulble, radioactive procduict wvas di,stributedthrouigho-ut the lower two-thir(ds of the ttibe witha major com,ponent having a sedimentation coeffi-cient of approximately 14 to 16S (fig 3A). Thedistribution of radioactivitv correlated with thedlistribuition oif the template DNA on the gradien'tas indicated by abso:rbancy measuirements at 260 nm.Essen,tially all the added: DNA was accounted forby the absorbancy in fractions 1 throutgh 11. Thetotal uiltraviole't absorbancy recovered from thegradielnt was 2.2-fold that calculated from theamotunt of DNA added to the gradient. Therefore,the major portion of the absorbancy exhibited infractions 14 through 16 Nvas ascribed to tinincor-porated ribonucleotides not removed from theproduct by dia,lysis. (The absorbancy (lata repre-sent acid-soltuble as well as acid-insoluble materialis.)After the product was heated for 10 mintuteis at100° and *then centrifuiged in the suicrose gradient,all the radioactive product NN'as conlcentrated in theuipper one-third of the ttube peaking near the posi-tion of a 4 to 6S component (fig 3B). Similarly,the ultraviolet absorbing mnaterial of tihe heatedproduct was concentrated in the upper one-third ofthe ttibe. All of the ultraviolet absorbaincy observedin fractionls I throuigh 11 of the uinheated prodtict(fig 3A) wa(s accotinted for as the increa!se inabsorbancy in fraction,s 12 throulgh 15 of the heatedproduct (,fig 3B). The changes in the sedimenta-tioIn behavior and senisitivity to RNase of the 9P-labheled product after heating is consistanit wi,th thebehavior of an RNA-DNA hyNbrid (44). Proof ofhybrid formation will require physical and chemicalcharacterization of the product isollated from anRNA polymerase incuibationi miixture containiingheated DNA.
LLiiDz
9
rI)bz
0U
A
BOTTOM
B
TOPTOP BOTTOMFRACTION NUMBER
FIG. 3. Effect of heating on sedimentation of RNApolynmerase prodIuct. A reactionl imiixtutre. 6-fol(d largerthani standard, conitaining 300 ug of heat denature(dcalf thvi-mu:s D.NA was incubated at 300 for 30 minutts.The reaction was stopped by dilutioni to 0.1 M with coldsodiumii acetate (pH 5) an(d to 0.4 % sodium dlodecy lsulfate. The mlixtulre was dialyzed for 6 houirs againstcold. flowxing 0.01 \i sodium acetate (,pH 5) and thellaigainst runnllilng distilled water for 18 houirs at 230. Thedial-zedl solut.ion was adjutsted to 0.1 X SSC (28)(pH 5). ail(l approxinmately one-half (80,632 CPMI) waslayered over a 2.5 to 15 % sucrose gradiemit in 0.03 Mrtris-Cl (pH 7.3) and; 0.1 \i NaCl (panel A). Theremainiing portion of thle dialyzed product (64,656 CPM\I)was heatedl in a boiling water bath for 10 minutes,chilled in ice andl theni lavered as ab)ove ( panel B).After centrifugation at 39,000 rpmi for 3.5 hours inthe SW 39 rotor at 20, fractions w-ere collected bvdropping from the bottom of the tubes. The fractioiusx-ere anal ized for aci-idnsoluble radioactivitx anld tot-1labsorbance at 260 nim. Ninety-nine percent of theradioactive products w ere recovered fromii thil gra(lilt s.
Discussion
The reaction catalyzed by the R-NA po)olymeraseisola,ted from maize differs in several resl)ects fromthe reactioni catalyzed by the microbial enzymes.A) Tl'he plant enzyme uises (lenatllre( I)NA moreefficiently than native DNA \vhereas .he microbialenzymes are more efficient ott natve 1)N.A .
408
Dow
nloaded from https://academ
ic.oup.com/plphys/article/43/3/405/6093852 by guest on 12 February 2022

STOUT AND AIANS-TE'MPLATE REQUIREMENTS OF POLYMIERASE
11,41). In its temiplate requiirement the maizeRNA polymerase more nearly resembles the DNApolymerases isolated from calf thymtis (2) andbacteria (34) than the microbial RNA polymerases.B) The inakility of ithe plianit enzyme to utilizeRNA or synthetic oligoribonucleotide templates(42) is in contrast with their utiilization by micro-bial enzymes (16, 22, 30, 40). C) The formationof homopolymers in the presence of he!ated DNAis catalyzed by the microbial enzymes (6,40),wherea,s the formation of a homopolymer is notdetected with the maize polymerase. D) RNApolymerases isolated froim microbial cells (6, 19, 31,39, 45) are fo-und associaited with a DNA-rich frac-tion of celil homogen;ates. The bulk of the RNApollymerase from maize seedlings is fouln(d in theDNA-poor, soluble portion of tissuie homogenates(27).
These differences in template requirement be-tween -the plant and microbial enzymes are in accordwith the suggestion (25) that the enzymatic ap-parattis required for RNA synthesis in cells havingtheir temiplate material (DNA) in an organizedchromosome (e.g., fliowering plants) (Iiffers fromthat in cells in which the DNA is not integratedinito the classical chromosomal configuration (bac-teria). The more effective reversal of aictinomycinD inhi'bikt'ion of the RNA polyme-rase from He/acoldls by heat denatured DNA compare(l with nativeDNA or RNA (15), also fits into this pattern.However, data from stfill another tisstue havingorganized chromosomes is not consistant with thisnotion. Furth and Ho (12) 'have demonstratedthat healt denattured DNA inhi'bits RNA synthesisby the soluble polymerase isolated from bovinelymphosarcoma cells. Experimental verif cation ofthe hypothe.sis put forth 'by Lin et ail. (25) mustawait a more extensive survey of the temiplatereqtlirements of RNA polymerases isolated fromdiverse biological species.
The inability of the maize polymerase to cata-lytica,lly utilize either denatured or naltivre templiatessuiggests that the transcription process in this plantrequires additional enzym,ic activiities (37). De-naituration of t'he native template seems to be therate limiting process. We suggest that a template-denaturing activity may be required for t'he cata-lytic use of DNA in the synthesis of plant RNA.This activity woulld prepare the native template fortranscription by locally opening or weakening theforces holding the 2 stranids of DNA together.RNA synthesis by the polymerase wo-uld then pro-ceed alonng 'the DNA strand( by way of a transienthybrid of a few nucleotfides. Release of the productfrom the template strand may also involve anenzymic process. This fiinal step in RNA syn-thesis may be catalyzed by the postulated tem,plate-denatuiring activity or by sitill another enzymicprotein. The results presented in this paper suiggestthat these postuilated activities may have been lostin purifying the "soluble' enzyme from the DNA-
poor fraction of the maize seedling homogenates(42).
Acknowledgment
The atuthors thank Dr. H. D. Gauclh and Dr. D. T.Morgan for their suggestions in the preparation of thismanuscript.
Literature Cited
Subsequent to completion of the experimental wvorkreported here and during the preparation of this manu-script, Karkas and Chargaff (1967) reported similarobservations on the higher template aotivity of dena-tured DNA's with E. coli RNA polymerase. In theirpaper they suggest that hybrid may accumulate in thepresence of low levels of denatured DNA. See Proc.Natl. Acad. Sci. 58: 1645-51.
1. APPLEQUIST, J. 1961. Estimation of base pairinigin nucleic acids from hypochromism. J. Am.Chem. Soc. 83: 3158-59.
2. BOLLUM, F. J. 1959. Thermail conversion of non-priming deoxyribonucleic acid to primer. J. Biol.Chem. 234: 2733-34.
3. BOLTON, E. T., R. J. BRITTEN, D. B. COWIE, R. B.ROBERTS, P. SZAFRANSKI, AND M. J. WARING.1965. Interaction of nucleic acids. Plant nucleicacids. In: Carnegie Institution Year Book 64.W'ashington, D. C. p 314.
4. BONNER, J., R. C. HUANG, AND N. MAHESHWARI.1961. The physical state of newly synthesizedRNA. Proc. Natl. Acad. Sci. 47: 1548-54.
5. BURMA, D. P., H. KROGER, S. OCHOA, R. C. WVAR-NER, AND J. D. WEILL. 1961. Further studieson deoxyribonucleic acid-dependent enzymatic syn-thesiis of ribonucleic acid. Proc. Nat]. Acad. Sci.47: 749-52.
6. CHAMBERLIN, M1., R. L. BALDWIN, AND P. BERG.1963. An enzymaticailly synthesized RNA of al-ternating base sequence: physical 'and chemicalcharacterization. J. AMol. Biol. 7: 334-39.
7. CHAMBERLIN, M. AND P. BERG. 1962. Deoxyribo-nucleic acid-directed synthesis of ribonucleic acidby an enzyme from Escherichia coli. Proc. Nat].Acad. Sci. 48: 81-94.
8. CH1ARGAFF. E. 1959. Isolationi and compositioni ofthe deoxvpentose nucleic acids and of the corre-sponding nucleoproteins. In: The Nucleic Acids.I. E. Chargaff and J. N. Davidson, eds. Aca-demic Press, New York. p 354.
9. DoTY, P., H. BOEDTKER, J. R. FRESCO, R. HASSEL-KORN, AND M. LITT. 1959. Seconidary structurein ribonucleic acids. Proc. Natl. Acad. Sci. 45:482-99.
10. ERGLE, D. R. AN-D F. R. H. KATTER-MAN. 1961.Deoxyribonucleic acid of cotton. Plant Physiol.36: 811-15.
1 1. Fox, C. F. AND S. B. WREISS. 1964. Enizymiaticsynthesis of RNA. II. Properties of the cleoxy-ribonucleic acid-primed reaction with Alicro-coccusbysodeiktictus ribonucleic acid polymerase. J. Biol.Chem. 239: 175-85.
12. FURTH, J. J. AND P. Ho. 1965. The enzymiaticsynthesis of ribon ucleic acid in animal tissule. T.
409
Dow
nloaded from https://academ
ic.oup.com/plphys/article/43/3/405/6093852 by guest on 12 February 2022

PLANT PHYSIOLOGY
The deox-vrilbonueleic acid-directed synthesis ofribonucleic acid as catal3 zed by an enzyme ob-tained from bovine lvm)phosarcotmia tissue. J. Biol.Chem. 240: 2602-11.
13. FURTH, J. J. AND P. Loi. 1964. Thiernmal im1ac-tivation of the primer in DNA depetndent syn-thesis of RNA in animilal tisstie. Science 145:161-62.
14. GEIDIUSCHEK, E. P., T. N\AKAMIOTO, AND S. B.W\EISS. 1961. The e!1zymatic synthesis of RNA:Complenmentary interaction with DN A. Proc.
Nati. Acad. Sci. 47: 1405-15.15. GOLDBERG, I. H. AND M. RABINOW\ITZ. 1962. Ac-
tinomwCin D inhibitioni of deoxvribonucleic acid-dependent svnithesis of ribontucleic acid. Science134: 315-16.
16. GOLDBERG, I. H. 'M. RABINOWX ITZ. AND E. REICH.1962. Basis of actinomvcin actioni. I. DNAbinding and inhibition of R'NA polynmerase syn-
thetic reactionms bv ctin omxciii. Proc. Natl.Acad. Sci. 48: 2094-2101.
17. HALL, B. D. AND S. SPIEGELMAN. 1961. Sequencecomplementarity of T2-DNA and T2-specificRN\A. Proc. 'Natl. Acad. Sci. 47: 137-46.
18. HAYASHIf, M. AND S. SPIEGELrMAN. 1961. The se-
lective svnthesis of informationcal RNA in bac-teria. Proc. Natl. Acad. Sci. 47: 1564-80.
19. HURNW-ITZ, J., A. BRESLER. AN-D R. DIRI-NGER. 1960.The enzymatic incorporationl of ribonucleotidesinto polyribonucleotides and the effect of DNA.Biochem. Biophys. Res. Comtmun. 3: 15 19.
20. JACOBSON. K. B. 1962. Ribonucleotides of RNA:Separation b1 chromatography oil sheets of di-ethylvlalmitnoetllSlcellulose. Science 138: 515-16.
21 KARKAS, J. D. AND E. CIIARGAFF. 1966. TemiplatefuInctioIns in the enzymatic formation of polyri-bonucleotides. I. Integrity of the DNA template.Proc. Nat]. Acad. Sci. 56: 664-71.
22. KRAKOW, J. S. A.ND S. OCHOA. 1963. Ribonucleicacid polymerase of .Azvotobactcr vinelaidii. I.
Priminig by polyribonucleotides. Proc. Natl. Acad.Sci. 49: 88-94.
23. KuINITZ, 'M. 1950. Crystalline desoxvribonuclease.I. Isolation anid general properties. Spectropho-tomletric method for the measurement of desoxv-ri'bonuclease activity. J. Gen. Physiol. 33: 349-62.
24. LEE-HUAItG, S. AND L. F. CAVALIERT. 1961. Poly-riboonucleotides as temnplates for polydeoxyribonu-cleotides. Proc. Natl. Acad. Sci. 50: 1116-22.
25. LUN, H. J., J. D. KARKAS, AND E. CHARGAFF. 1966.Tem,plate functions in the enzymatic formation ofpolyribonucleotides. II. 'Metaphase chromosomes
as templates in enzytlmatic sy-nthesis of ribonucleicaci(l. Proc. Natl. Acad. Sci. 56: 954-Q9.
26. M\IANS, R. J. AND G. D. NOVELLI. 1961. M\easure-menit of the inlcorporation of radioactive amino,acics into proteini by a fillter-paper (lisk method.Archl. Biochemii. Biophys. 94: 48-53.
27. MIANS, R. J. AND G. D. NOVELLI. 1964. Ribonu-cleotide incorporation by a soluble enzyme frommaize. Biochim. Biophys. Acta 91: 186-88.
28. 'MAR-MUR. J. AND D. LANE. 1960. Strand separa-tioIn and specific recombinationl in deoxyribonu-cleic acids: biological studies. Proc. Natl. Acad.Sci. 46: 453-61.
29. 'MESELSON, M. AND F. W\. STAhIL. 1958. The rep-
lication of DNA in Esclheriilichi coli. Proc. Natl.Acad. Sci. 44: 671-82.
30. NAKA-MOTO. T. AND) S. B. \VEISS. 1962. The blo-synthesis of RNA: Priminig by polyriboniucleo-tides. Proc. Natl. Acad. Sci. 48: 880-93.
31. OCHOA, S., D. P. BUR-MA, H. KROGER, AND J. D.\\'EILL. 1961. Deoxyribonucleic acid-dependentincorporation of tiucleotides froml inucleoside tri-plhosphlates into riboniuoleic acid. Proc. Natl. Acad.Sci. 47: 670-79.
32. RA-MUZ, 'M., J. DOLY, P. MANDEL, AND P. CHAM-BON. 1965. A soluble DNA-dependent RNApolynierase in nluclei of non-dividing animal cells.Bioclheimi. Biophy s. Res. CoI11mmun. 19: 114-20.
33. Rici, -\. 1960. A lhvbrid helix containing bothdeoxyribose and ribose polynucleotides and its re-
lation to the transfer of informnationi betxween thenucleic acids. Proc. Natl. Acad. Sci. 46: 1044-53.
34. RICHARDSON, C. C., C. L. SCHILDKRAUT, AND A.
KORNBERG. 1963. Studies onl the replicationi ofDN'A by R'NA pol!-iimerase. Cold Spring HarborSymp. Quiant. 13iol. XXVIII: 9-19.
35. SCHACHMAAN, H. H.. J. ADLER, C. M. RADDING, I.R. LEHM\AN. AND KORNBERG. 1960. EnIzymuaticsx! 1tl1eSis of deoxyribonucleic acid. VII. SynIthesis of a poolymer of dleoxx-adenylate and deoxy-
tlhvinidv late. J. Biol. Clheml. 235: 3242-49.36. ScHui.\iAN, H. 'M. A-ND D. -M. BONNER. 1962. A
natinralkl occu-rring DTX\-kRNiA conmplex fronmX(V'dnropo(r (1cv.isa. Proc. Natl. Acad. Sci. 48:53-63
37. SI'NGER, M. F. AND P. LEDER. 1966. MessenigerRNA: ain Cxvaltiation. Rev. Bioclhemi. 35:195-260.
38. SPIEGEIh\AN. S., I'). 1). HALL., AND R. STORCK.1961. The ocCIrriliec of natural DN -RNAcomiiplexes in (oili inlfected itlh T2. Proc.
Natl. Acad. Sci. 47- 113541.39. STEVENS, A. 1960. Inicorporationi of the adleniine
ribonticleotide itito RNA by cell fractionis fromE. col/i B. Bioclhemii. Biophls. Res. Coimmiiiuni. 3:92-96.
40. STFAEA\-S, A. 1964. Studies of the ribounucleic acidpolmnierase fronm Eschcr-ichia coli. II. Studies of11omopolymer f0rmiatioins. J. Biol. Chemii. 239:204-)9.
41. STEVENS, A. AND) J. HENRY. 1964. Studies of theribonucleic acid po'lymerase from Escherichia coli.I. Purificationi of the enzyne -and studies of RNAformation. J. Biol. Chemii. 239: 196-203.
42. STOUT. F. R. AND R. J. 'MANS. 1967. Partialpurificatioin anid properties of RN\A polymllerasefrom maize. Biochimn. Biophys. Acta 1134: 327-36.
43. SZYBAILSKT, WV. 1967. Effects of elevated tem-peratures oni DNA and oni some polynucleotides:Denatturationi, reniaturation and cleavage of glv-
cosidic an(c pIhosphate ester bonds. Chapt. IV.In: Thermiohiolog-. A. H. Rose, ed. AcademicPress, Inc., Lonidoni. p 73-122.
44. WARNER. R. C., H. H. SAAIUELS, 'M. T. ABBOTT,AND J. S. KARKONV. 1963. Ribonucleic acid poly-merase of A-Lotobactcr vinclandii. II. Forma-tioIn of D'NA-RNA hybrids xvith single-strandedDNAX as printer. Proc. Natl. Acad. Sci. 49:533-38.
45. WEISS, S. B. AND T. NAKAMOTO. 1961. Net syn-thesis of ribontucleic acid xith a microbial enizymerequirinig deoxx ribontucleic acid and four ribonu-cleosidle triphosphates. J. Biol. Chem. 236: PC19-20.
410
Dow
nloaded from https://academ
ic.oup.com/plphys/article/43/3/405/6093852 by guest on 12 February 2022


















