
Supporting Information - PNAS€¦ · Supporting Information Kim and McGinnis...
9
Supporting Information Kim and McGinnis 10.1073/pnas.1016386108 SI Materials and Methods Plasmid Constructs and Biochemical Analyses. The GST-tagged grh- WT construct was generated by subcloning Drosophila grh-N cDNA into pGEX-4T3 vector in frame with GST. grh deletion mutants (N1, N2, and N3) were subcloned into the pGEX-4T3 after PCR amplification from the grh-N cDNA. grh mutants (S88A, S91A, 2A, 2E, and Pan A) in the pGEX-4T3 were cre- ated by PCR-based site-directed mutagenesis (Stratagene). These GST-tagged GRH constructs were expressed in BL21 Escherichia coli and then purified by using GST-agarose column (Clontech). The purified proteins were subjected to in vitro ERK kinase assays. The ERK kinase assay was performed in a reaction solution containing 8 mU of purified mouse ERK2 (Upstate Biotechnology), 20 mM Hepes at pH 7.2, 10 mM MgCl 2 , 3 mM β-mercaptoethanol, 0.1 mM ATP, 0.12 mg/mL BSA, and 0.2 μCi/ μL γ-[ 32 P]ATP at 30 °C for 30 min. Phosphorylated protein was detected by autoradiography, and total substrate was shown by Coomassie blue staining. Relative phosphorylation was calcu- lated in the following way: The relative mass of GRH proteins in a band was determined by measuring band intensities of inverted Coomassie blue-stained gel images whose backgrounds were set as a black reference. Relative protein quantities were divided by their molecular weight to derive the relative moles of protein in a band. Band intensities obtained from autoradiography were divided by the relative molar amount to determine the relative phosphorylation of molecules. Phosphorylation of full-length GRH was standardized as 1.0 in every experiment. The averages obtained from three independent experiments are presented as a bar graph, and the SDs are shown by error bars. The grh WT, grh 2A, grh PanA, and grh 2E sequences were subcloned into pcDNA 3.1(-)/Myc-His A vector, and expressed in rabbit reticulocyte lysates (Promega). The expressed proteins were subjected to EMSA assay as described (1). The primers used for generating the PCR fragments or site-directed muta- genesis are described in “Primer Sequences” below. For quan- tification of the amount of radioactive probe bound by GRH proteins, the relative amount of GRH–DNA shifted complex in a band was determined by measuring nonsaturated band in- tensities of EMSA gel images. Drosophila Genetics and Cuticular Preparation. grh-WT, grh-2A, grh- PanA, and grh-2E cDNA sequences were subcloned into the pUAST-attB vector and targeted to the identical 22A3 insertion site in the VK-37 strain via the phiC31-integrase system (2). Transgenic UAS-GRH WT, UAS-GRH 2A, UAS-GRH PanA, and UAS-GRH 2E in grh IM homozygotes were activated with e22c-GAL4 by using the UAS-GAL4 system, which allows en- dogenous-level GRH expression in late embryonic epidermal cells. However, at early embryonic stages, before stage 11, transgenic GRH protein levels driven by e22c-GAL4 were much lower than endogenous GRH. e22c-driven transgenic GRH ex- pression was not sufficient to rescue the lethality of grh IM mu- tants, which is probably due, in part, to the lower levels of transgenic GRH in early embryos, as well as the e22c driver not restoring the CNS expression of GRH (3). To analyze cuticle phenotypes, Drosophila embryos were collected on standard apple juice agar plates at 25 °C for 6 h, and then aged for an additional 30 h, before cuticular preparation (4). The grh IM allele (5) and e22c-GAL4 line (6) were obtained from the Bloomington Drosophila Stock Center, and en-GAL4 was a gift from E Bier (University of California at San Diego). The Moe-GFP (sGMCA) line, which carries a constitutively expressed actin binding domain of moesin fused to GFP (7), was a gift from TH Millard (University of Manchester, Manchester, UK) and used to visualize and follow reepithelialization after epidermal wounding. Generation of GRH Antisera and Immunostaining. The DNA encod- ing C-terminal fragment of GRH-N protein (residues 603–1063) was subcloned into pGEX-4T-3 and transformed into BL21 E. coli strain for protein induction. The induced protein was purified by using GST-agarose column (Clontech), and then the purified protein was sent to Pocono Rabbit Farm and Lab to generate anti-GRH sera from a guinea pig. The crude sera from the guinea pig were sub- jected to affinity purification by PVDF-immobilized GST-tagged GRH-N (8). Anti-Fasciclin 3, anti-Coracle, and anti–DE-cadherin antibodies were obtained from the Developmental Studies Hy- bridoma Bank. Immunostaining was done as described (9), using anti-GRH (1:200), anti-Fasciclin 3 (7G10, concentrate, 1:400; ref. 10), anti-Coracle (C615.16, concentrate, 1:200; ref. 11), and anti– DE-cadherin (DCAD2, 1:20, concentrate; ref. 12) antibodies as primary antibodies, and Invitrogen Alexa-Fluor secondary anti- bodies (1:400). Embryos were mounted in Prolong Gold antifade reagent (Invitrogen). All images of immunostained stage 16 embryos were collected with a Leica SP2 laser-scanning upright confocal microscope. Gain and offset were set to nonsaturated fluorescence levels with identical instrumental settings for each independent ex- periment. Images were compiled from confocal optical sections scanned at 0.6- to 0.8-μm thickness, and the maximum projection images are presented in Fig. 2, Fig. S3 and Fig. S4. Quantification of protein expression level was done by measuring fluorescence in- tensity of the average projection images of the nonsaturated stacks. The averages of fluorescence intensities from more than three em- bryos were normalized to the control value of grh IM/+ heterozygous embryos, and presented as bar graphs in Fig. 2G and Fig. S5. The SEs are shown by error bars. P values were calculated by using one-way ANOVA analysis. Epidermal Wounding and Imaging of Embryos. Embryos were col- lected for 2 h on standard apple juice agar plates at 25 °C and then aged for an additional 15 h. The washed embryos were dechorionated in 50% bleach and then washed with distilled water. The dechorionated embryos were transferred onto slides with double-sided sticky tape, dehydrated 5–10 min in a desiccation chamber, and covered in hal- ocarbon oil (2:1 ratio of 700:27 halocarbon oil) or PBS. Then, the stage 16–17 embryos were laterally wounded with a microinjection needle or top-wounded with a thin tungsten needle. The wounded live embryos were placed in a humid chamber for 4–6 h at room temperature (≈22 °C) for induction of epidermal wound reporters and then visualized by using an epifluorescence-equipped Axioplan 2 imaging system (Carl Zeiss MicroImaging) to determine the wound response. Images of top-wounded embryos were obtained with a Leica SP2 laser-scanning upright confocal microscope, choosing representative embryos as described (1). Epidermal Barrier Permeability Assay. Stage 17 embryos aged after 2-h embryo collection at 22 °C were dechorionated in 50% bleach for 2 min and washed with distilled water for 1 min in a 100-μm cell strainer (BD Falcon). The embryos in the cell strainer were placed in isopropanol solution and soaked for 30 s and then dried for 1 min. The isopropanol-treated embryos were trans- ferred into heptane and 0.3% 1-butanol solution for 90 s and soaked in heptane for 15 s, and then dried for 90 s. The embryos were washed with Shields and Sang M3 insect media (M3 media) (Sigma-Aldrich) and immersed in 0.1% Rhodamine B dye in M3 Kim and McGinnis www.pnas.org/cgi/content/short/1016386108 1 of 9
Transcript of Supporting Information - PNAS€¦ · Supporting Information Kim and McGinnis...

Supporting InformationKim and McGinnis 10.1073/pnas.1016386108SI Materials and MethodsPlasmid Constructs and Biochemical Analyses. The GST-tagged grh-WT construct was generated by subcloning Drosophila grh-NcDNA into pGEX-4T3 vector in frame with GST. grh deletionmutants (N1, N2, and N3) were subcloned into the pGEX-4T3after PCR amplification from the grh-N cDNA. grh mutants(S88A, S91A, 2A, 2E, and Pan A) in the pGEX-4T3 were cre-ated by PCR-based site-directed mutagenesis (Stratagene).These GST-tagged GRH constructs were expressed in BL21
Escherichia coli and then purified by using GST-agarose column(Clontech). The purified proteins were subjected to in vitro ERKkinase assays. The ERK kinase assay was performed in a reactionsolution containing 8 mU of purified mouse ERK2 (UpstateBiotechnology), 20 mM Hepes at pH 7.2, 10 mM MgCl2, 3 mMβ-mercaptoethanol, 0.1 mM ATP, 0.12 mg/mL BSA, and 0.2 μCi/μL γ-[32P]ATP at 30 °C for 30 min. Phosphorylated protein wasdetected by autoradiography, and total substrate was shown byCoomassie blue staining. Relative phosphorylation was calcu-lated in the following way: The relative mass of GRH proteins ina band was determined by measuring band intensities of invertedCoomassie blue-stained gel images whose backgrounds were setas a black reference. Relative protein quantities were divided bytheir molecular weight to derive the relative moles of protein ina band. Band intensities obtained from autoradiography weredivided by the relative molar amount to determine the relativephosphorylation of molecules. Phosphorylation of full-lengthGRH was standardized as 1.0 in every experiment. The averagesobtained from three independent experiments are presented asa bar graph, and the SDs are shown by error bars.The grh WT, grh 2A, grh PanA, and grh 2E sequences were
subcloned into pcDNA 3.1(-)/Myc-His A vector, and expressedin rabbit reticulocyte lysates (Promega). The expressed proteinswere subjected to EMSA assay as described (1). The primersused for generating the PCR fragments or site-directed muta-genesis are described in “Primer Sequences” below. For quan-tification of the amount of radioactive probe bound by GRHproteins, the relative amount of GRH–DNA shifted complex ina band was determined by measuring nonsaturated band in-tensities of EMSA gel images.
Drosophila Genetics and Cuticular Preparation. grh-WT, grh-2A, grh-PanA, and grh-2E cDNA sequences were subcloned into thepUAST-attB vector and targeted to the identical 22A3 insertionsite in the VK-37 strain via the phiC31-integrase system (2).Transgenic UAS-GRH WT, UAS-GRH 2A, UAS-GRH PanA,and UAS-GRH 2E in grhIM homozygotes were activated withe22c-GAL4 by using the UAS-GAL4 system, which allows en-dogenous-level GRH expression in late embryonic epidermalcells. However, at early embryonic stages, before stage 11,transgenic GRH protein levels driven by e22c-GAL4 were muchlower than endogenous GRH. e22c-driven transgenic GRH ex-pression was not sufficient to rescue the lethality of grhIM mu-tants, which is probably due, in part, to the lower levels oftransgenic GRH in early embryos, as well as the e22c driver notrestoring the CNS expression of GRH (3).To analyze cuticle phenotypes,Drosophila embryoswere collected
on standard apple juice agar plates at 25 °C for 6 h, and then aged foran additional 30 h, before cuticular preparation (4). The grhIM allele(5) and e22c-GAL4 line (6) were obtained from the BloomingtonDrosophila Stock Center, and en-GAL4 was a gift from E Bier(University of California at San Diego). The Moe-GFP (sGMCA)line, which carries a constitutively expressed actin binding domain
ofmoesin fused toGFP (7), was a gift fromTHMillard (Universityof Manchester, Manchester, UK) and used to visualize and followreepithelialization after epidermal wounding.
Generation of GRH Antisera and Immunostaining. The DNA encod-ingC-terminal fragmentofGRH-Nprotein(residues603–1063)wassubcloned intopGEX-4T-3 and transformed intoBL21E.coli strainfor protein induction. The induced protein was purified by usingGST-agarose column (Clontech), and then the purified protein wassent to Pocono Rabbit Farm and Lab to generate anti-GRH serafrom a guinea pig. The crude sera from the guinea pig were sub-jected to affinity purification by PVDF-immobilized GST-taggedGRH-N (8). Anti-Fasciclin 3, anti-Coracle, and anti–DE-cadherinantibodies were obtained from the Developmental Studies Hy-bridoma Bank. Immunostaining was done as described (9), usinganti-GRH (1:200), anti-Fasciclin 3 (7G10, concentrate, 1:400; ref.10), anti-Coracle (C615.16, concentrate, 1:200; ref. 11), and anti–DE-cadherin (DCAD2, 1:20, concentrate; ref. 12) antibodies asprimary antibodies, and Invitrogen Alexa-Fluor secondary anti-bodies (1:400). Embryos were mounted in Prolong Gold antifadereagent (Invitrogen).All images of immunostained stage 16 embryoswere collected with a Leica SP2 laser-scanning upright confocalmicroscope. Gain and offset were set to nonsaturated fluorescencelevels with identical instrumental settings for each independent ex-periment. Images were compiled from confocal optical sectionsscanned at 0.6- to 0.8-μm thickness, and the maximum projectionimages are presented in Fig. 2, Fig. S3 and Fig. S4. Quantification ofprotein expression level was done by measuring fluorescence in-tensity of the average projection images of the nonsaturated stacks.The averages of fluorescence intensities from more than three em-bryos were normalized to the control value of grhIM/+ heterozygousembryos, andpresented as bar graphs inFig. 2GandFig. S5.TheSEsare shown by error bars. P values were calculated by using one-wayANOVA analysis.
Epidermal Wounding and Imaging of Embryos. Embryos were col-lectedfor2honstandardapplejuiceagarplatesat25°Candthenagedfor an additional 15 h. The washed embryos were dechorionated in50%bleachand thenwashedwithdistilledwater.Thedechorionatedembryos were transferred onto slides with double-sided sticky tape,dehydrated 5–10 min in a desiccation chamber, and covered in hal-ocarbon oil (2:1 ratio of 700:27 halocarbon oil) or PBS. Then, thestage 16–17 embryos were laterally wounded with a microinjectionneedle or top-wounded with a thin tungsten needle. The woundedlive embryos were placed in a humid chamber for 4–6 h at roomtemperature (≈22 °C) for induction of epidermal wound reportersand then visualized by using an epifluorescence-equipped Axioplan2 imaging system (Carl Zeiss MicroImaging) to determine thewound response. Images of top-wounded embryos were obtainedwith a Leica SP2 laser-scanning upright confocal microscope,choosing representative embryos as described (1).
Epidermal Barrier Permeability Assay. Stage 17 embryos aged after2-h embryo collection at 22 °C were dechorionated in 50% bleachfor 2 min and washed with distilled water for 1 min in a 100-μmcell strainer (BD Falcon). The embryos in the cell strainer wereplaced in isopropanol solution and soaked for 30 s and thendried for 1 min. The isopropanol-treated embryos were trans-ferred into heptane and 0.3% 1-butanol solution for 90 s andsoaked in heptane for 15 s, and then dried for 90 s. The embryoswere washed with Shields and Sang M3 insect media (M3 media)(Sigma-Aldrich) and immersed in 0.1% Rhodamine B dye in M3
Kim and McGinnis www.pnas.org/cgi/content/short/1016386108 1 of 9

media for 5 min. Thereafter, treated embryos were washed withM3 media for 2 min. The entire procedure was done on cellstrainers. The dye-immersed embryos were mounted in halo-carbon oil (2:1 ratio of 700:27 halocarbon oil). The fluorescenceimages were immediately taken under epifluorescence-equippedAxioplan 2 imaging system (Carl Zeiss MicroImaging) or LeicaDM 2500 microscope.For epidermal barrier permeability assay after wounding, the
stage 16–17 embryos aged after 2-h embryo collection at 22 °Cwere dechorionated, aligned on slides with double-sided stickytape, dehydrated 5–10 min in a desiccation chamber, and coveredwith halocarbon oil (2:1 ratio of 700:27 halocarbon oil). Theembryos were laterally wounded with a microinjection needleapparatus. The wounded live embryos were placed in a humidchamber for 16 h at room temperature (≈22 °C). The embryoswere removed from the slide by using heptanes and transferredthem to a 100-μm cell strainer. After washing them with M3media, they were immersed in 0.025% Rhodamine B dye in M3media for 2 min and then washed with M3 media for 2 min. Thedye-immersed embryos were mounted in halocarbon oil, andfluorescence images were immediately obtained with epifluor-escence-equipped Leica DM 2500 microscope. The percentile ofpermeable embryos was counted from each group, and the con-trol value from grhIM/+ heterozygotes was subtracted from valuesof all groups. Then, the percentile was recalculated and presentedas a bar graph in Fig. 5G. P values were calculated by using one-way ANOVA analysis.
Epidermal Wound Closure Assay. Embryos were collected for 1 h onstandard apple juice agar plates at 22 °C and then aged for anadditional 23 h at 22 °C. The washed embryos were dechorio-nated and then washed with distilled water. The dechorionatedembryos were transferred onto slides with double-sided stickytape, dehydrated 5–10 min in a desiccation chamber, and coveredin halocarbon oil (2:1 ratio of 700:27 halocarbon oil). Then, thestage 16–17 embryos were laterally wounded with a microinjec-tion needle. The wounded live embryos were placed in a humidchamber for 8 h at room temperature (≈22 °C) for epidermalwound closure. For imaging of the embryos, the wounded em-bryos were loosened from the tape with dissecting needle (FisherScientific; no. 08–964-A) and then transferred onto new slidewith double-sided sticky tape. To immobilize the embryos, 1-phenoxy-2-propanol (Sigma-Aldrich; no. 484423) diluted 1:50with distilled water was used. Subsequently, wound sites of theimmobilized embryos were reoriented to visualize embryonicwounds from a top view and then mounted in halocarbon oil (2:1ratio of 700:27 halocarbon oil). All images of live wounded em-bryos at stage 17 were collected with a Leica SP2 laser-scanningupright confocal microscope. The images were compiled fromconfocal optical sections, and the maximum projection imagesare presented in Fig. S9.
Primer Sequences. The following primers were used for indicatedpurposes.
(i) Subcloning full-length GRH into pGEX-4T35′-GCGGAATTCCTCCACATCCACCGCCACAA-3′and 5′-CGCCTCGAGCTATACAGCCTCCTTGTCGTA-CA-3′
(ii) Subcloning WT and mutant GRH into pUAST-attB5′-GCGGAATTCATGTCCACATCCACCGCC-3′and 5′-CGCGCGGCCGCCTACTGATTGGGCAGCTC-C-3′
(iii) Subcloning WT and mutant GRH into pcDNA 3.1(-)/Myc-His A5′-GCGCTCGAGGCCACCATGTCCACATCCAC-3′and 5′-CGCAAGCTTCTGATTGGGCAGCTCCGT-3′
(iv) Subcloning deletion mutant GRH into pGEX-4T3(a) GRH-N15′-GCGGAATTCCTCCACATCCACCGCCACAA-3′and 5′-CGCCTCGAGCTAATTGGTGCTGGTGTCCA-CCA-3′(b) GRH-N25′-GCGGAATTCCTCCACATCCACCGCCACAA-3′and 5′-CGCCTCGAGCTAACCCCCGCCATCCGCTG-GCA-3′(c) GRH-N35′-GCGGAATTCCTCCACATCCACCGCCACAA-3′and 5′-CGCCTCGAGCTACAGTGGTGAACTGTTT-GTGTT-3′(d) GRH-C (for GRH antibody generation)5′-GCG GAA TTC CAA TGG ATC CAC GCG ATC -G-3′ and 5′-CGC CTC GAG CTA TAC AGC CTC CTTGTC GTA CA-3′
(v) Site-directed mutagenesis of GRH(a) GRH S54A5′-AGCGACGCATCCCTAGCGCCCATCCAACAAG-GC-3′ and 5′-GCCTTGTTGGATGGGCGCTAGGGA-TGCGTCGCT-3′(b) GRH S72A5′-GGTAACACAAACAGTGCACCACTGGCGCCCA-AC-3′ and 5′-GTTGGGCGCCAGTGGTGCACTGTT-TGTGTTACC-3′(c) GRH S88A5′-ACAACAATGCACCGAGCACCGGACGCACCG-CAG-3′ and 5′-CTGCGGTGCGTCCGGTGCTCGGT-GCATTGTTGT-3′(d) GRH S91A5′-CACCGATCACCGGACGCACCGCAGCCAGAAT-TG-3′ and 5′-CAATTCTGGCTGCGGTGCGTCCGG-TGATCGGTG-3′(e) GRH T125A5′-GGCTGTATTTATATACGAAGCGCCCAAGGTG-GTGATGCCAG-3′ and 5′-CTGGCATCACCACCTT-GGGCGCTTCGTATATAAATACAGCC-3′(f) GRH S259A5′-GCGGTATATCAGACAGCACCATTGCCGCTAG-AC-3′ and 5′-GTCTAGCGGCAATGGTGCTGTCTG-ATATACCGC-3′(g) GRH T423A5′-GCTCAATGGCCAGACCGCACCTATCGATGTC-TCTG-3′ and 5′-CAGAGACATCGATAGGTGCGGT-CTGGCCATTGAGC-3′(h) GRH S591A5′-CCATGGCAGCCAGAACGCTCCCACCACATCG-CTGG-3′ and 5′-CCAGCGATGTGGTGGGAGCGTT-CTGGCTGTTATGG-3′(i) GRH S639A5′-CTTCCGATATCACCTGGAGGCCCCCATCAGT-TCATCGCAG-3′ and 5′-CTGCGATGAACTGATGG-GGGCCTCCAGGTGATATCGGAAG-3′(j) GRH S693A5′-ATGTTCCGCGAGGAGAAGGCTCCCGAGGAT-GAGATCAA-3′ and 5′-TTGATCTCATCCTCGGGA-GCCTTCTCCTCGCGGAACAT-3′(k) GRH S855A5′-AAGCCGCCGGTGCTATTCGCGCCCGCCGAGG-ACATG-3′ and 5′-CATGTCCTCGGCGGGCGCGAA-TAGCACCGGCGGCTT-3′(l) GRH S902A5′-GGCCATGAGACTGACGCGCCGGACCTGAAG-GGG-3′ and 5′-CCCCTTCAGGTCCGGCGCGTCAG-TCTCATGGCC-3′
Kim and McGinnis www.pnas.org/cgi/content/short/1016386108 2 of 9

(m) GRH S909A5′-GACCTGAAGGGGGCCGCACCGTTCCTGCTCC-AC-3′ and 5′-GTGGAGCAGGAACGGTGCGGCCC-CCTTCAGGTC-3′(n) GRH T920A5′-GGCCAGAAGGTGGCCGCGCCGACGCTCAAG-TTC-3′ and 5′-GAACTTGAGCGTCGGCGCGGCCA-CCTTCTGGCC-3′(o) GRH T950A5′-AACATGTTGACCAGCGCACCCCTGACCGACT-TT-3′ and 5′-AAAGTCGGTCAGGGGTGCGCTGG-TCAACATGTT-3′(p) GRH T965A5′-GAAGCGCGGCAGGATGGCGCCGCCGACCTC-GGAA-3′ and 5′-TTCCGAGGTCGGCGGCGCCATC-CTGCCGCGCTTC-3′
(q) GRH T985A5′-GAACGAGGAGGTGTATGCACCGTTGCACGTG-GTGC-3′ and 5′-GCACCACGTGCAACGGTGCATA-CACCTCCTCGTTC-3′(r) GRH S88E5′-CACAACAATGCACCGAGAACCGGACTCACCG-CAGC-3′ and 5′-GCTGCGGTGAGTCCGGTTCTCG-GTGCATTGTTGTG-3′(s) GRH S91E5′-CACCGAGAACCGGACGAACCGCAGCCAGAA-TTG-3′ and 5′-CAATTCTGGCTGCGGTTCGTCCGG-TTCTCGGTG-3′
1. Pearson JC, Juarez MT, Kim M, Drivenes Ø, McGinnis W (2009) Multiple transcriptionfactor codes activate epidermal wound-response genes in Drosophila. Proc Natl AcadSci USA 106:2224–2229.
2. Venken KJ, He Y, Hoskins RA, Bellen HJ (2006) P[acman]: A BAC transgenic platformfor targeted insertion of large DNA fragments in D. melanogaster. Science 314:1747–1751.
3. Uv AE, Harrison EJ, Bray SJ (1997) Tissue-specific splicing and functions of theDrosophila transcription factor Grainyhead. Mol Cell Biol 17:6727–6735.
4. Bergson C, McGinnis W (1990) An autoregulatory enhancer element of the Drosophilahomeotic gene Deformed. EMBO J 9:4287–4297.
5. Ostrowski S, Dierick HA, Bejsovec A (2002) Genetic control of cuticle formation duringembryonic development of Drosophila melanogaster. Genetics 161:171–182.
6. Wood W, et al. (2002) Wound healing recapitulates morphogenesis in Drosophilaembryos. Nat Cell Biol 4:907–912.
7. Kiehart DP, Galbraith CG, Edwards KA, Rickoll WL, Montague RA (2000) Multipleforces contribute to cell sheet morphogenesis for dorsal closure in Drosophila. J CellBiol 149:471–490.
8. Koelle MR, Horvitz HR (1996) EGL-10 regulates G protein signaling in the C. elegansnervous system and shares a conserved domain with many mammalian proteins. Cell84:115–125.
9. Mace KA, Pearson JC, McGinnis W (2005) An epidermal barrier wound repair pathwayin Drosophila is mediated by grainy head. Science 308:381–385.
10. Patel NH, Snow PM, Goodman CS (1987) Characterization and cloning of fasciclin III: Aglycoprotein expressed on a subset of neurons and axon pathways in Drosophila. Cell48:975–988.
11. Fehon RG, Dawson IA, Artavanis-Tsakonas S (1994) A Drosophila homologue ofmembrane-skeleton protein 4.1 is associated with septate junctions and is encoded bythe coracle gene. Development 120:545–557.
12. Oda H, Uemura T, Harada Y, Iwai Y, Takeichi M (1994) A Drosophila homolog ofcadherin associated with armadillo and essential for embryonic cell-cell adhesion. DevBiol 165:716–726.
WT
S91A
S88A WT
PanA2A
A B
0%
20%
40%
60%
80%
100%
0%
20%
40%
60%
80%
100%
Rel
ativ
e Ph
osph
oryl
atio
n
Rel
ativ
e Ph
osph
oryl
atio
n
Fig. S1. Relative phosphorylation of GRH by ERK quantified by densitometry. (A) As shown in Fig. 1D, GRH S91A mutant displays a strong reduction in the invitro phosphorylation by ERK compared with GRH WT. The bar graph indicates the mean of four independent experiments, and error bars indicate SDs. (B) Asshown in Fig. 1E, GRH Pan Ala (PanA) mutation abolishes ERK-dependent phosphorylation compared with GRH WT and GRH S88A/S91A (2A) mutant. The bargraph indicates the mean of three independent experiments, and error bars indicate SDs.
Kim and McGinnis www.pnas.org/cgi/content/short/1016386108 3 of 9

GRHDdc
- WT 2A PanA 2E - WT 2A PanA 2Ewt wt wt wt wt mut mut mut mut mut
GRH
1.0 1.4 1.3 1.0 0 0 0 0 00
NS
NS
Fig. S2. Mutations of ERK phosphorylation sites in GRH do not hamper binding toDdc enhancer site. The DNA binding activities of GRHWT, GRH 2A, GRH PanA,andGRH S88E/S91E(2E) proteins (expressed in a rabbit reticulocyte lysate system, SIMaterials andMethods) were analyzed by electrophoretic mobility shift assaysusing 32P-labeled oligonucleotide probe (wt) containing a consensus GRH binding site from the Ddc epidermal wound-response enhancer. Probes with mutantGRH sites (mut) were used as binding specificity controls. Arrowhead indicates GRH–DNA shifted complexes. The intensities of GRH–DNA complexes werequantified, and calculated relative value is indicated below each corresponding band. Unbound probe is displayed in the bottom of the gel, and all data weretaken from the same gel. NS indicates nonspecific shifted probe complexes and represents an oligonucleotide binding activity from the reticulocyte lysate.
Kim and McGinnis www.pnas.org/cgi/content/short/1016386108 4 of 9
DC
AD
2G
RH
grhIM hrg+/ IM: e22c>GRHWT
grhIM: e22c>GRHPanAgrhIM: e22c>GRH2A
DC
AD
2G
RH
grhIM: e22c
C’
B C
E
E’
F
F’
grhIM: e22c>GRH2E
D’
A’
D
A
B’
25 m
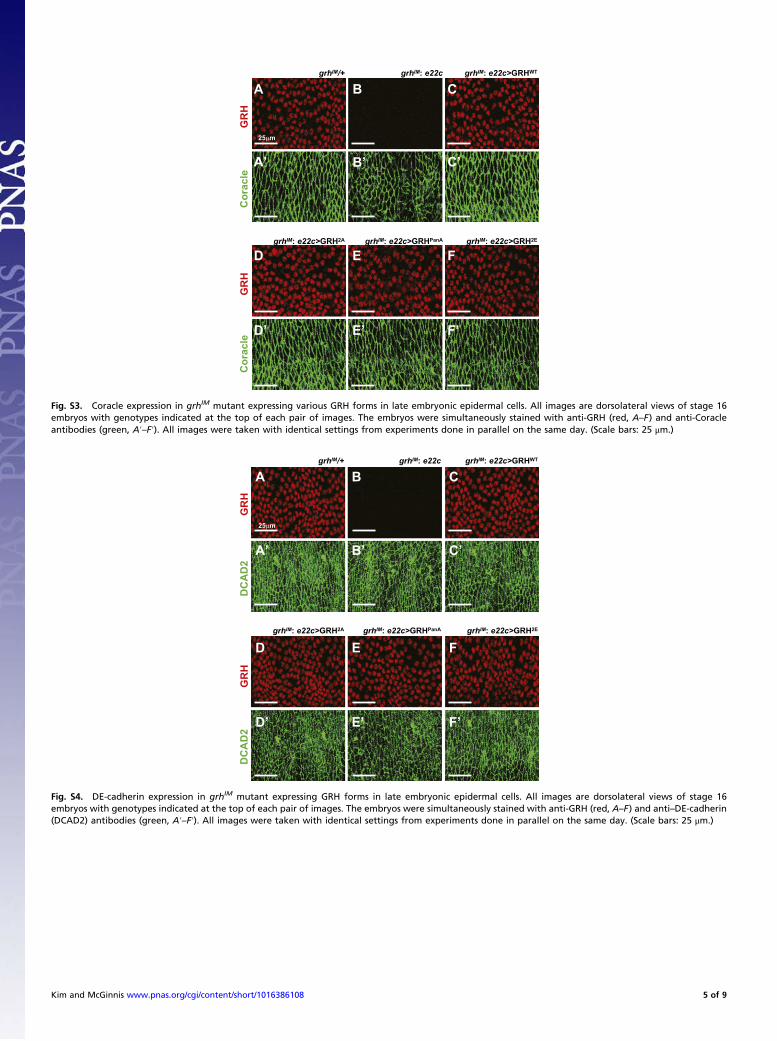
Fig. S4. DE-cadherin expression in grhIM mutant expressing GRH forms in late embryonic epidermal cells. All images are dorsolateral views of stage 16embryos with genotypes indicated at the top of each pair of images. The embryos were simultaneously stained with anti-GRH (red, A–F) and anti–DE-cadherin(DCAD2) antibodies (green, A′–F′). All images were taken with identical settings from experiments done in parallel on the same day. (Scale bars: 25 μm.)
Cor
acle
GR
H
grhIM hrg+/ IM: e22c>GRHWT
Cor
acle
GR
H
grhIM: e22c
A’ B’ C’
A B C
D E
D’ E’
F
F’
grhIM: e22c>GRH2A grhIM: e22c>GRHPanA grhIM: e22c>GRH2E
25 m
Fig. S3. Coracle expression in grhIM mutant expressing various GRH forms in late embryonic epidermal cells. All images are dorsolateral views of stage 16embryos with genotypes indicated at the top of each pair of images. The embryos were simultaneously stained with anti-GRH (red, A–F) and anti-Coracleantibodies (green, A′–F′). All images were taken with identical settings from experiments done in parallel on the same day. (Scale bars: 25 μm.)
Kim and McGinnis www.pnas.org/cgi/content/short/1016386108 5 of 9
00.20.40.60.8
11.21.41.6
00.20.40.60.8
11.21.4
grhIM : e22c
grhIM :
e22c>G
RH2E
grhIM :
e22c>G
RHPa
nA
grhIM :
e22c>G
RHW
T
grhIM :
e22c>G
RH2A
grhIM /+
grhIM : e22c
grhIM :
e22c>G
RH2E
grhIM :
e22c>G
RHPa
nA
grhIM :
e22c>G
RHW
T
grhIM :
e22c>G
RH2A
grhIM /+
A
B
*
ytisnetnIevitale
RytisnetnI
evitaleR
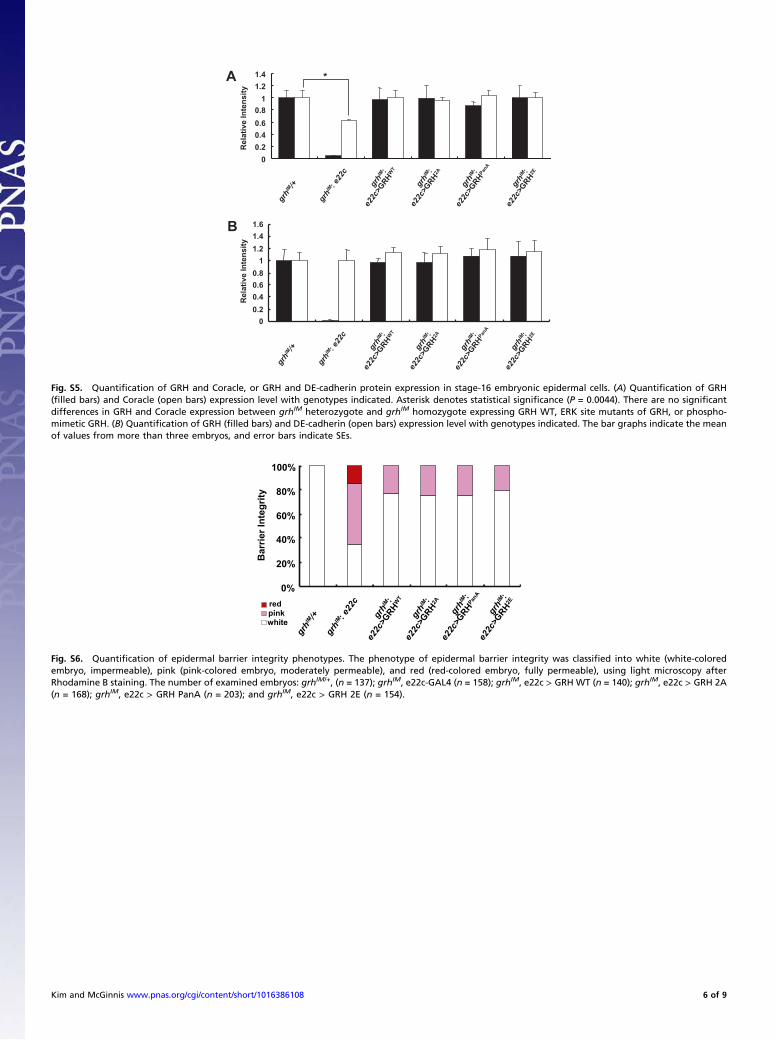
Fig. S5. Quantification of GRH and Coracle, or GRH and DE-cadherin protein expression in stage-16 embryonic epidermal cells. (A) Quantification of GRH(filled bars) and Coracle (open bars) expression level with genotypes indicated. Asterisk denotes statistical significance (P = 0.0044). There are no significantdifferences in GRH and Coracle expression between grhIM heterozygote and grhIM homozygote expressing GRH WT, ERK site mutants of GRH, or phospho-mimetic GRH. (B) Quantification of GRH (filled bars) and DE-cadherin (open bars) expression level with genotypes indicated. The bar graphs indicate the meanof values from more than three embryos, and error bars indicate SEs.
Bar
rier I
nteg
rity
grhIM : e22c
grhIM :
e22c>G
RH2A
grhIM :
e22c>G
RH2E
grhIM :
e22c>G
RHW
T
grhIM /+
redpinkwhite
grhIM :
e22c>G
RHPa
nA
0%
20%
40%
60%
80%
100%
Fig. S6. Quantification of epidermal barrier integrity phenotypes. The phenotype of epidermal barrier integrity was classified into white (white-coloredembryo, impermeable), pink (pink-colored embryo, moderately permeable), and red (red-colored embryo, fully permeable), using light microscopy afterRhodamine B staining. The number of examined embryos: grhIM/+, (n = 137); grhIM, e22c-GAL4 (n = 158); grhIM, e22c > GRH WT (n = 140); grhIM, e22c > GRH 2A(n = 168); grhIM, e22c > GRH PanA (n = 203); and grhIM, e22c > GRH 2E (n = 154).
Kim and McGinnis www.pnas.org/cgi/content/short/1016386108 6 of 9
Wou
nd R
espo
nse
(msn
-WE1
)
grhIM/+ grhIM: e22c grhIM: e22c>GRHWT
grhIM: e22c>GRHPanAgrhIM: e22c>GRH2A
D E
F
A B C
50µm
0%
20%
40%
60%
80%
100%moderatestrong
grhIM : e22c
grhIM :
e22c>G
RH2A
grhIM :
e22c>G
RHPa
nA
grhIM :
e22c>G
RHW
T
grhIM /+
msn
-WE1
msn
-WE1
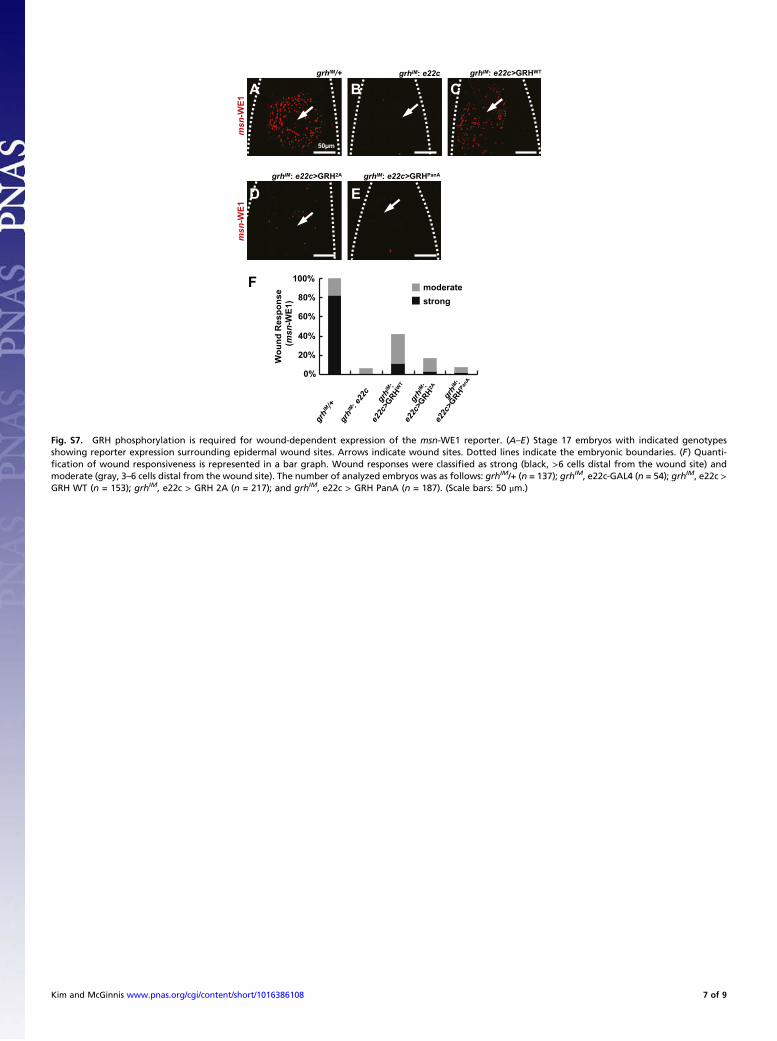
Fig. S7. GRH phosphorylation is required for wound-dependent expression of the msn-WE1 reporter. (A–E) Stage 17 embryos with indicated genotypesshowing reporter expression surrounding epidermal wound sites. Arrows indicate wound sites. Dotted lines indicate the embryonic boundaries. (F) Quanti-fication of wound responsiveness is represented in a bar graph. Wound responses were classified as strong (black, >6 cells distal from the wound site) andmoderate (gray, 3–6 cells distal from the wound site). The number of analyzed embryos was as follows: grhIM/+ (n = 137); grhIM, e22c-GAL4 (n = 54); grhIM, e22c >GRH WT (n = 153); grhIM, e22c > GRH 2A (n = 217); and grhIM, e22c > GRH PanA (n = 187). (Scale bars: 50 μm.)
Kim and McGinnis www.pnas.org/cgi/content/short/1016386108 7 of 9

en>G
RH
WT
en>G
RH
2Aen
>GR
HPa
nA
GRHDdc .47-GFP
25 m
Fig. S8. In unwounded embryos, Ddc induction upon GRH overexpression is not altered by mutation of ERK phosphorylation sites. Indicated WT or ERK sitemutants of GRH proteins were strongly overexpressed in posterior compartments of embryonic segments by using en-GAL4. The images all show lateral viewsof stage 16 embryos with the genotypes indicated at the left side of each pair of images. Left show Ddc 0.47-GFP reporter expression (green) in unwoundedembryos. Ddc 0.47-GFP fluorescent signals were observed in a subset of epidermal nuclei with en-GAL4 driven GRH protein. Right show the overexpression ofGRH proteins in engrailed stripes as visualized by anti-GRH immunofluorescent staining (red) in fixed stage-16 embryos. (Scale bars: 25 μm.)
Kim and McGinnis www.pnas.org/cgi/content/short/1016386108 8 of 9

A B
A’ B’
0%
20%
40%
60%
80%
100%
open wound
partially closed wound
closed wound
grhIM :
Moe-GFP
Moe-GFP
Moe-GFP grhIM: Moe-GFP
C
50µm
% o
f em
bryo
s
Fig. S9. GRH is not essential for epidermal wound closure at late embryonic stage. At 8 h after wounding, maximumprojection images of GFPfluorescence weretaken from control (A and A′) and grhIM homozygous (B and B′) embryo carrying Moe-GFP at stage 17. (A and B) Embryos with indicated genotypes showingpartially closed wound at 8 h after wounding. (A′ and B′) Embryos with indicated genotypes showing fully closed wound at 8 h after wounding. Arrows indicateepidermal wound sites. (C) Quantification of epidermal wound closure phenotype is shown in a bar graph. The extent of wound closure was classified as openwounds, partially closed wounds, and closed wounds in controls (n = 15) and grhIM homozygotes (n = 20) carrying Moe-GFP. (Scale bars: 50 μm.)
Kim and McGinnis www.pnas.org/cgi/content/short/1016386108 9 of 9


















