
Studies in Larval Amphibian Habitat Partitioning
36
Studies in Larval Amphibian Habitat Partitioning W. RONALD HEYER SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY • NUMBER 242
Transcript of Studies in Larval Amphibian Habitat Partitioning

Studies in Larval AmphibianHabitat Partitioning
W. RONALD HEYER
SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY • NUMBER 242

SERIES PUBLICATIONS OF THE SMITHSONIAN INSTITUTION
Emphasis upon publication as a means of "diffusing knowledge" was expressedby the first Secretary of the Smithsonian. In his formal plan for the Institution, JosephHenry outlined a program that included the following statement: "It is proposed topublish a series of reports, giving an account of the new discoveries in science, andof the changes made from year to year in all branches of knowledge." This themeof basic research has been adhered to through the years by thousands of titles issuedin series publications under the Smithsonian imprint, commencing with SmithsonianContributions to Knowledge in 1848 and continuing with the following active series:
Smithsonian Contributions to AnthropologySmithsonian Contributions to Astrophysics
Smithsonian Contributions to BotanySmithsonian Contributions to the Earth SciencesSmithsonian Contributions to the Marine Sciences
Smithsonian Contributions to PaleobiologySmithsonian Contributions to Zoo/ogySmithsonian Studies in Air and Space
Smithsonian Studies in History and Technology
In these series, the Institution publishes small papers and full-scale monographsthat report the research and collections of its various museums and bureaux or ofprofessional colleagues in the world cf science and scholarship. The publications aredistributed by mailing lists to libraries, universities, and similar institutions throughoutthe world.
Papers or monographs submitted for series publication are received by theSmithsonian Institution Press, subject to its own review for format and style, onlythrough departments of the various Smithsonian museums or bureaux, where themanuscripts are given substantive review. Press requirements for manuscript and artpreparation are outlined on the inside back cover.
S. Dillon RipleySecretarySmithsonian Institution

S M I T H S O N I A N C O N T R I B U T I O N S T O Z O O L O G Y • N U M B E R 242
Studies in Larval AmphibianHabitat Partitioning
W. Ronald Heyer
SMITHSONIAN INSTITUTION PRESSCity of Washington
1976

A B S T R A C T
Heyer, W. Ronald. Studies in Larval Amphibian Habitat Partitioning. Smith-sonian Contributions to Zoology, number 242, 27 pages, 3 figures, 14 tables,1976.—Habitat partitioning by larval amphibians was studied in two environ-ments. A two-year study was undertaken at the Chesapeake Bay Center for En-vironmental Studies and a comparative dry and wet season study was completedat Barro Colorado Island, Panama.
At the Chesapeake site, partial year to year differences in temporal larvalhabitat partitioning combined with different year to year population sizesindicate: (1) different habitat utilization from one year to the next by the larvae,and (2) different biological interactions involving predator-prey and species as-sociations from year to year.
The stream bed pond habitat on Barro Colorado Island is effectively par-titioned by the tadpole community on the basis of space and time. Predationis an organizing factor in the composition of the tadpole community, inter-specific competition apparently is not.
This study concludes that competition among tadpoles does not normallyoccur in nature. Bursts of primary productivity provide superabundant foodresources. The habitat partitioning found in tadpole communities reflects thetemporal and microhabitat differences associated with the evolutionary lifehistory of each of the species occurring in any pond. Tadpole communities areadditive collections of different species, not integrated, interactive superorgani-zations.
OFFICIAL PUBLICATION DATE is handstamped in a limited number of initial copies and is recordedin the Institution's annual report, Smithsonian Year. SERIES COVER DESIGN: The coral Montastreacavernosa (Linnaeus).
Library of Congress Cataloging in Publication DataHeyer, W RonaldStudies in larval amphibian habitat partitioning.(Smithsonian contributions to zoology ; no. 242)Bibliography: p.Supt. of Docs, no.: SI 1.27:2421. Tadpoles. 2. Habitat partitioning. 3. Amphibians—Ecology. I. Title. II. Series: Smith-
sonian Institution. Smithsonian contributions to zoology ; no. 242.QL1.S54 no. 242 [QL668.E2] 591'.08s [597'.8'0439] 76-608161

Contents
Page
Introduction 1Acknowledgments 1
Chesapeake Bay Area 2Methods and Materials 2Temporal Succession 2Microhabitat Partitioning 8Microhabitat Metrics 10Discussion 13
Barro Colorado Island 15Methods and Materials 15Results 15Habitat Partitioning 17Discussion 17
Other Studies of Larval Habitat Partitioning 18Tadpole Community Organization 19The Generalized Anuran Life Cycle and Its Evolutionary Maintenance . 22Selection, Population Control, and the Anuran Life Cycle 24Evolutionary Modifications of the Generalized Anuran Life Cycle 25Literature Cited 26
in


Studies in Larval AmphibianHabitat Partitioning
W. Ronald Heyer
Introduction
The single goal of this study was to learn moreabout how habitats are partitioned by larval am-phibians. Data were gathered from two very differ-ent sites, a temperate pond and a series of tropicalstream ponds.
The study of habitat partitioning by larval am-phibians has been previously neglected. There arefewer than a half dozen descriptive studies andonly 4 quantitative studies (cited in Heyer, 1973).The temperate pond (present study) was followedfor two years to study year to year variability inhabitat partitioning. The tropical stream ponds,located on Barro Colorado Island, were sampledonce during the dry season and once during thewet season to determine the effects of seasonality onhabitat partitioning by stream pool larval amphib-ians.
The studies at the two sites are described sep-arately because the methods and analyses of dataare different. Patterns of habitat partitioning evi-denced at the sites are then compared. Any pat-terns of habitat partitioning common to these twoenvironmental extremes should be common tolarval amphibia in other environments.
It is necessary to define larval amphibian habitatpartitioning as used in this study. The habitat con-sists of one or more ponds found at a specific site.
W. Ronald Heyer, Department of Vertebrate Zoology, Na-tional Museum of Natural History, Smithsonian Institution,Washington, D.C. 20560.
The habitat contains the resources utilized bydifferent species of larval amphibians; different lar-val species utilize the habitat resources in differentways. The patterns of habitat usage produced bydifferent larval species are manifested as a par-titioning of the habitat resources. Larvae may par-tition the habitat in terms of space, time, or food(among the more obvious categories). Examples ofpartitioning include: (1) where the habitat consistsof a series of ponds at a single site, larvae of aparticular species may occur in only a few of theavailable ponds; (2) within a pond, larvae of asingle species may be found only in certain micro-habitats or they may utilize only certain kinds offood; (3) two species may use the same pond atdifferent times as larvae. The studies reportedherein reveal both space and time habitat par-titioning by larval amphibians.
ACKNOWLEDGMENTS.—Roy W. McDiarmid, Uni-versity of South Florida, Richard J. Wassersug,University of Chicago, and George R. Zug, Smith-sonian Institution, carefully read the entire manu-script. Linda R. Maxson, University of Illinois, readthe section discussing the amphibian genetic sys-tem. Comments from the reviewers considerablyimproved the paper, but the reviewers do notnecessarily agree with all of the conclusions or dis-cussion.
The following persons supplied specific infor-mation: Ronald I. Crombie and James F. Lynch,both of the Smithsonian Institution, William F.
1

SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
Pyburn, University of Texas at Arlington, andKentwood Wells, University of Connecticut.
The Environmental Sciences Program of theSmithsonian Institution supported the field workin Panama.
Elena, Laura, and Miriam Heyer participated inthe field work both in Panama and in the Saturdayexcursions to the Chesapeake site. Without theirhelp and cooperation, the study would not havematerialized.
Chesapeake Bay Area
METHODS AND MATERIALS.—Larval samples weretaken once a week from 9 March to 29 June 1974and from 22 March to 28 June 1975. A sample for15 June 1974 was not obtained due to illness. Anadditional sample of 26 July 1975 was taken todetermine whether any drastic changes occurredafter June, the arbitrarily determined end of thesampling period. In both years, sampling precededthe onset of tadpole occurrence. Samples weretaken from a large, shallow pond surrounded byhardwood forest at the Smithsonian's ChesapeakeBay Center for Environmental Studies near An-napolis, Maryland. The common trees in theforest are beeches, hickories, oaks, and tulip trees.Three microhabitats were sampled by dipnet eachsampling period. The dipnet had a triangular open-ing. For the surface samples, the base of the tri-angle was placed just under the surface of thewater. The surface film and about the top 5 cmof water were swept at the grassy edge of the pondwhere the water was about 30-40 cm deep. Thenet skimmed the top of the grass. The midwatersample was taken farther from the shore, in about60-70 cm water, in grassy and open water areas.The net opening was completely immersed, thenthe net was propelled through the water. Openwater areas were emphasized in the midwater sam-ple. The bottom sample was taken in the samegeneral area as the midwater samples, but thegrassier areas were emphasized. For the bottomsample, the net was bumped along the bottom. Thelinear distances sampled for the sweep types werevariable, but comparable. Generally, at least a25 m linear sweep was made for each sample. Ifthe net had an abundance of tadpoles, no furthersampling was done in that microhabitat. If few or
no tadpoles were collected, additional sweeps weremade. The same microhabitats were sampled eachweek in the same way both years. The surface,midwater, and bottom sweeps sampled three moreor less discrete areas of the pond. The results wereof course only a reflection of the dynamics of thepopulations of the entire pond.
The tadpoles of each sweep type were formalinpreserved in separate containers. In the laboratorythe samples were sorted and identified to specieslevel. Salamanders were measured by vernier cali-pers, snout-vent and total lengths measured to thenearest 0.5 mm. Tadpoles were staged (Gosner,1960). For very large samples (unless all were hatch-lings), 50 individuals were staged and the totalsweep sample for that species was extrapolated fromthe staged subset. The 1974 sample is representedby 1182 individuals, the 1975 sample by 2755 in-dividuals. All specimens (total study) are in thecollections of the Division of Reptiles and Am-phibians, National Museum of Natural History.
TEMPORAL SUCCESSION.—1974 Pattern: Amby-stoma maculatum egg masses were first seen on 16March, but may have been present (and overlooked)previously. Hatchlings were not collected until 4May (Figure 1). The previous week, 27 April, anempty salamander jelly case was found in the pond,but no larvae were collected in the sweeps. Manydead egg masses were found in April and May. Atthe end of June, some of the larvae were meta-morphic size.
Ambystoma opacum lays its eggs in the fall. Lar-vae were present in the pond when samplingstarted on 9 March. The population demonstrateda synchronous growth pattern (Figure 1). The lastlarvae, of metamorphic size, were collected on 18May.
Hyla crucifer hatchlings were first collected on13 April (Figure 1). Hatchlings continued to becollected through 4 May, indicating a probablebreeding season of about a month for H. cruciferin 1974. Metamorphic larvae were first collectedon 25 May. Only prometamorphic larvae were col-lected on 8 June. All larvae apparently completedmetamorphosis prior to 22 June (no sample avail-able for 15 June). The time spreads of hatchlingappearance and metamorphosis are comparable.
Hyla chrysoscelis (not versicolor, James Lynch,pers. comm.) were first collected on 1 June (Figure

NUMBER 242
1). The two larvae collected were both well beyondstage 25. Hyla chrysoscelis was not abundant in thepond; sampling error probably accounts for hatch-lings not being collected prior to 1 June. Collec-tions of only large larvae on 22 and 29 June sug-gest that the breeding period for H. chrysoscelisended in early June in 1974.
Rana clamitans were first heard calling on 11May. Larvae were first collected on 1 June. Larvaewere never abundant, but hatchlings were col-lected through the entire sampling period (Figure1).
Rana sylvatica larvae were first collected on 6April. Population growth appeared rather syn-chronous; metamorphic larvae were last collectedon 1 June (Figure 1).
Bufo americanus were collected only once, ashatchlings, on 20 April. Concerted efforts to re-collect the species within the sampling areas wereunsuccessful for the remainder of the samplingyear.
No Acris crepitans larvae were collected in 1974.Tadpoles could be seen easily only in the open
water areas of the pond. In 1974, from mid-Aprilto mid-May, Rana sylvatica larvae were predomi-nant; the population, estimated visually, appearedto be very large in the open water. After the R.sylvatica had metamorphosed, there appeared to belittle tadpole activity in the open water areas ofthe pond.
1975 Pattern: Ambystoma maculatum eggs werefound on the first sampling time of 22 March. Lar-vae were not collected until 26 April (Figure 2).Small larvae were collected over a long period, from26 April to 31 May, apparently reflecting differencesin hatching times of the various egg masses. Larvaewere present throughout the remainder of thesampling period (Figure 2). Only one egg masswas found in which a noticeable percentage of eggswas inviable.
Very few Ambystoma opacum larvae were col-lected in 1975. Sampling errors apparently accountfor lack of specimens from the sampling dates of29 March through 19 April. Larvae were last col-lected on 26 April (Figure 2).
Hyla crucifer egg masses were first observed on22 March. Hatchlings were collected from 12 Aprilto 31 May, an 8 week period (Figure 2). Meta-morphs were first collected on 31 May and last
collected on 28 June. Judging by the staging graphsand relative numbers (Figure 2), the populationhad completed, or was in the process of, metamor-phosis by 28 June.
Hyla chrysoscelis larvae were first collected on31 May. The staging graphs (Figure 2) suggest thatthe breeding period was short and the tadpolegrowth was synchronous through 28 June.
Overwintered Rana clamitans larvae were col-lected on the first sampling date, 22 March (Figure2). Calling males were first heard on 10 May, andthe first spring hatched larvae were collected on10 May as well. Early developmental stages of theoverwintering larvae were much larger than thesame stages of the spring hatched larvae. Larvaewere present throughout the entire sampling pe-riod.
Rana sylvatica eggs were observed on 22 March.Hatchlings were first collected on 19 April andlast collected on 10 May. Late prometamorphiclarvae were first collected on 24 May and last col-lected on 28 June (Figure 2).
On 3 May, a calling male of Acris crepitans wascaptured for identification. Calling males wereheard on sampling dates from 3 May through 21June. Larvae were never abundant and were firstcollected on 21 June (Figure 2).
On 26 July, a sample was taken to determinewhether the sampling time span concluding withthe end of June was adequate to describe the es-sential tadpole interactions. Larvae of three specieswere collected: Acris crepitans (6 individuals), Hylachrysoscelis (2), and Rana clamitans (18). Hatch-lings were collected only of R. clamitans. Thissample indicated that while larvae were still in thepond, most of the larval activity had concludedbefore the end of June. No additional species wererepresented, nor were the species present repre-sented by large numbers.
The sample date of 5 April is notable becauseno larvae were collected, although they must havebeen present in the pond (Figure 2). The tempera-ture was cold at the sampling time. The air tem-perature was 0.4° C, the water temperature 3.4°C, and there was a film of ice on the water. Thecold temperature may have caused the larvae, as abehavioral response, to leave the sites they normallyinhabited when temperatures were more favorable.
In the open areas of the pond, the predominant

t!27
APR
IL
5 20
AP
RIL
5
13 A
PR
ILo
00
00
r °
r5
6 A
PR
IL
005
30 M
AR
CH
5
23 M
AR
CH
5 005 00
5 16
MA
RC
Ho
9
5 9
MAR
CH
500
°

NUMBER 242
100
AMBYST. AMBYST. HYLAMACUL. OPACUM CRUCIFER
HYLA RANA RANACHRYSOSCELIS CLAMITANS SYLVATICA
. 0
. 0
- 0
. 0
. 2
1 1. 7
I.. 15
.1. 2
-
-
•
-
•
1
•
-
0
0
0
0
1
2
1' 1 8
L
;3
1
•
-1
•
•
•
•
•
-I
-
-
33
33 j
116 :i
7 !
i3
10
0
0
45 25 45 25 45 25 45
opacum, left to right, < 15 mm, 16-22 mm, 23-29 mm, > 30 mm. Horizontal axes for tad-poles = Gosner (1960) stages. Dashed vertical lines = metamorphosis. Number within eachgraph = total number of individuals per sample.)

SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
species, estimated visually, was Bnfo americanus. two years produces some striking similarities andComparison of the 1974 and 1975 Patterns: A striking contrasts (Figure 3).
comparison of the patterns observed during the Each of three species repeated in 1975 the tem-
lOOiI •
o
I(VI
lOOi
Oac
2a><M 0
100
100
tra.
100
_Jaca.< •
100
AMBYST. AMBYST. HYLA
MACUL. OPACUM CRUCIFER
100
I100
I2 0
. 3
IV
36
50
292
427
514
HYLA RANA
CHRYSOSCELIS CLAMITANSRANA
SYLVATICA
25
k
45 25 45 25
18
19
1L45 25
BUFO ACRISAMERICANUS CREPITANS
82
142
130
145 25 45 25 45
NUMBER 242
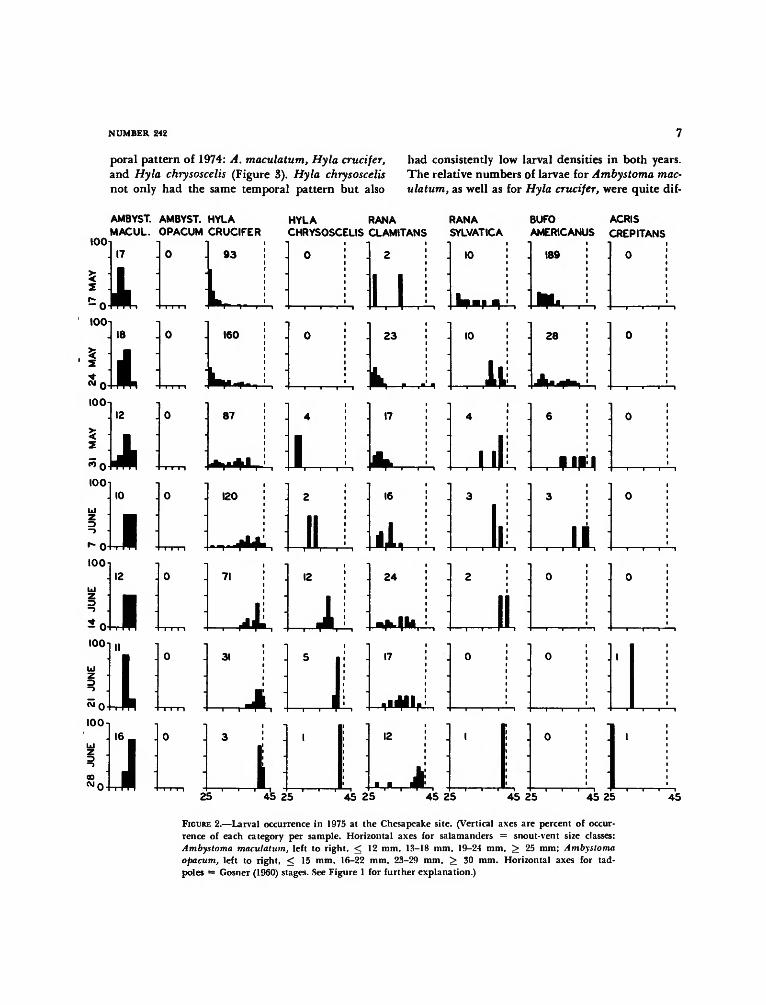
poral pattern of 1974: A. maculatum, Hyla crucifer,and Hyla chrysoscelis (Figure 3). Hyla chrysoscelisnot only had the same temporal pattern but also
had consistently low larval densities in both years.The relative numbers of larvae for Ambystoma mac-ulatum, as well as for Hyla crucifer, were quite dif-
100
AMBYST. AMBYST. HYLA HYLA RANA RANA BUFO ACRISMACUL. OPACUM CRUCIFER CHRYSOSCELIS CLAMITANS SYLVATICA AMERICANUS CREPITANS
10
10
iv
189
28
Jl
25 45 25 45 25 45 25 45 25 45 25 ^
FIGURE 2.—Larval occurrence in 1975 at the Chesapeake site. (Vertical axes are percent of occur-rence of each category per sample. Horizontal axes for salamanders = snout-vent size classes:Ambystoma maculatum, left to right, < 12 mm, 13-18 mm, 19-24 mm, > 25 mm; Ambystomaopacum, left to right, < 15 mm, 16-22 mm, 23-29 mm, > 30 mm. Horizontal axes for tad-poles = Gosner (1960) stages. See Figure 1 for further explanation.)
45

SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
A. MACULATUM
A. OPACUMH. CRUCIFERH. CHRYSOSCEUSR. CLAMITANSR. SYLVATICA
B. AMERICANUSA. CREPITANS
• • • • • • i
MARCH APRIL MAY JUNE
FIGURE 3.—Presence of larvae in the Chesapeake site pond for 1974 and 1975. (Upper line foreach species = 1974, lower line = 1975. Dates supplied for which no larvae were collecteddue to presumed sampling error—see text. Dashed lines = probable presence of larvae priorto initiation of sampling in 1975—see text.)
ferent in 1975 from what they had been in 1974.Although data were not collected on egg masses,visual observation indicated that comparable num-bers of A. maculatum egg masses were laid in 1974and 1975. Egg mortality, high in 1974 and low in1975, probably accounts for the different densities oflarval population in the two years. The patterns ofHyla crucifer are similar at the onset of both yearsbut different at the end. The data presented inFigures 1 and 2 suggest that the extended breedingseason of Hyla crucifer in 1975 may account for thelarger larval population in that year.
The remaining species show striking temporaland population density differences when the datafor the two years are compared. Of these, the mostprofound differences are found in Rana clamitans,individuals of which overwintered in 1974-75, butnot in 1973-74. The first appearance of spring-hatched larvae of R. clamitans also differs: 1 Junein 1974, 10 May in 1975.
In order to understand fully the observed simi-larities and differences of the patterns of temporal
succession and population density recorded in eachof the two years, climatic patterns will have to beexamined and analyzed in detail together with thelarval data. Long term sampling, over several years,not just two, is necessary to reveal relationships be-tween climate and larval populations. For this rea-son, these relationships are not examined in thispaper. Sampling at the Chesapeake site is plannedto continue for the next few years to address theserelationships.
MICROHABITAT PARTITIONING.—The numbers of
individuals of each species were totalled by type ofsweep (Tables 1 and 2) to determine the differencesin abundance for each sweep type.
In both 1974 and 1975, Ambystoma maculatumindividuals were not equally distributed among thethree sweep types, more individuals being found inthe surface sweeps than in any of the other twosweep types. Ambystoma opacum individuals werestatistically equally distributed among the threesweep types in both years (based on small-sizedsamples). Most Hyla crucifer larvae were collected
TABLE 1.—Distribution of individuals in microhabitats in 1974 (ns = not significant; • = sig-nificant at 5% level; • • = significant at or beyond 1% level)
Species Surface Midwater Bottom
Ambyatoma maculatum...Amby8toma opacumHyla cruciferHyla chry8OBceliaRana clamitansRana syIvaticaBufo americanus
116
452637
296
2347174
1210
42239
480
7.88*2.36 (ns)
575.81**12.54**3.88 (ns)
113.67**592.00**
NUMBER 242
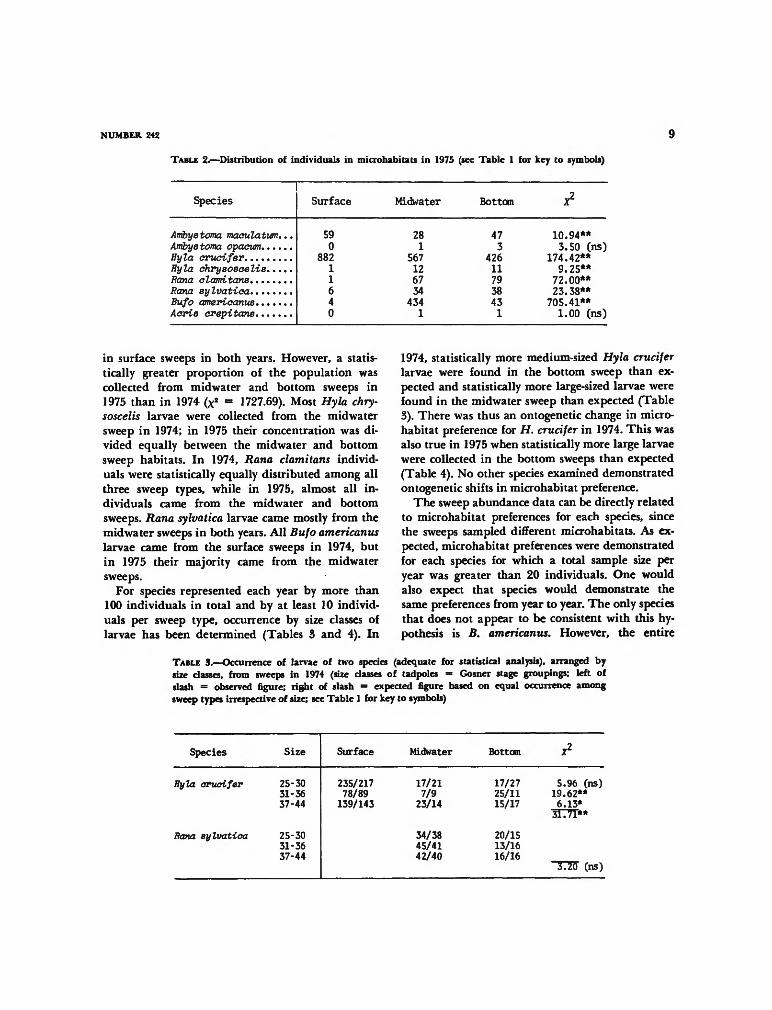
TABLE 2.—Distribution of individuals in microhabitats in 1975 (see Table 1 for key to symbols)
Species Surface Midwater Bottom x2
Ambyetoma maculatum...
Hyla crucifer
Rana olamitansRana sylvatioa
Acris crepitans
590
88211640
281
567126734
4341
473
426117938431
10.94**3.50 (ns)
174.42**9.25**
72.00**23.38**705.41**1.00 (ns)
in surface sweeps in both years. However, a statis-tically greater proportion of the population wascollected from midwater and bottom sweeps in1975 than in 1974 (x
2 = 1727.69). Most Hyla chry-soscelis larvae were collected from the midwatersweep in 1974; in 1975 their concentration was di-vided equally between the midwater and bottomsweep habitats. In 1974, Rana clamitans individ-uals were statistically equally distributed among allthree sweep types, while in 1975, almost all in-dividuals came from the midwater and bottomsweeps. Rana sylvatica larvae came mostly from themidwater sweeps in both years. All Bufo americanuslarvae came from the surface sweeps in 1974, butin 1975 their majority came from the midwatersweeps.
For species represented each year by more than100 individuals in total and by at least 10 individ-uals per sweep type, occurrence by size classes oflarvae has been determined (Tables 3 and 4). In
1974, statistically more medium-sized Hyla cruciferlarvae were found in the bottom sweep than ex-pected and statistically more large-sized larvae werefound in the midwater sweep than expected (Table3). There was thus an ontogenetic change in micro-habitat preference for H. crucifer in 1974. This wasalso true in 1975 when statistically more large larvaewere collected in the bottom sweeps than expected(Table 4). No other species examined demonstratedontogenetic shifts in microhabitat preference.
The sweep abundance data can be directly relatedto microhabitat preferences for each species, sincethe sweeps sampled different microhabitats. As ex-pected, microhabitat preferences were demonstratedfor each species for which a total sample size peryear was greater than 20 individuals. One wouldalso expect that species would demonstrate thesame preferences from year to year. The only speciesthat does not appear to be consistent with this hy-pothesis is B. americanus. However, the entire
TABLE 3.—Occurrence of larvae of two species (adequate for statistical analysis), arranged bysize classes, from sweeps in 1974 (size classes of tadpoles = Gosner stage groupings; left ofslash = observed figure; right of slash = expected figure based on equal occurrence amongsweep types irrespective of size; see Table 1 for key to symbols)
Species Size Surface Midwater Bottom
By la crucifer
Rana sylvatica
25-3031-3637-44
25-3031-3637-44
235/21778/89
139/143
17/217/9
23/14
34/3845/4142/40
17/2725/1115/17
20/1513/1616/16
5.96 (ns)19.62**6.13*
31771**
"7720" (ns)

10 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
TABLE 4.—Occurrence of larvae of five species (adequate for statistical analysis), arranged bysize classes, from sweeps in 1975 (size classes of salamanders in mm; see Table 1 for key tosymbols and Table 3 for further explanation)
Species
Ambystoma maculatwn
By la crucifer
Rana alamitans
Rana sylvatiaa
Bufo americanus
Size
3=1213-1819-24
^25
25-3031-3637-44
25-3031-3637-44
25-3037-44
25-3031-36
Surface
21/1511/1116/2011/13
740/73648/5594/90
Midwater
8/72/5
10/98/6
502/47031/3534/58
38/3517/2012/12
20/2210/7
368/37161/59
Bottom
6/1212/919/1610/10
324/35838/2764/44
39/4126/2314/14
30/286/9
36/323/5
10779" (ns)
5.43 (ns)5.82 (ns)
19.20**30.46**
""1720" (ns)
"XoT (ns)
~O9~ (ns)
1974 sample was collected in a single, surface sweepwhich passed through hatching egg strings. The1975 data doubtless demonstrate the larval micro-habitat preferences in contrast to the 1974 informa-tion, which may indicate nothing more than theegg deposition sites. Hyla crucifer, while demon-strating a preference for the surface sweep micro-habitat, showed a shift toward statistically moreindividuals using the midwater and bottom micro-habitats in 1975 than in 1974. This may be due todifferences in total population densities for the twoyears.
The ontogenetic change in microhabitat pref-erences for H. crucifer in 1974 and 1975 is interest-ing. The data for the two years combined suggestthat smaller larvae tend to be in shallow surfaceareas and as they grow, they move into deeper, moreopen parts of the pond.
MICROHABITAT METRICS.—Commonly used meas-ures of niche breadth and niche overlap can encom-pass the factors of temporal and spatial partitioningby the larvae when the measures are based on in-dividual sweeps (see Heyer, 1974, for a previousanalysis and justification).
Microhabitat Breadth: The formula used to de-termine microhabitat breadth is:
B, -^f'zlpij';iVt i
where 57 is the microhabitat breadth of species j ,Ns is the number of sweeps in which individualsof species ; were collected, Nt is the total number ofsweeps, and p j ; is the proportion of occurrences ofspecies ; in sweep i (modified from Levins, 1968).
Microhabitat breadths for 1974 and 1975 are pre-sented in Table 5. When the species are rankedwithin years by broadest microhabitat width to nar-rowest, only one species shows the same relativeposition. Hyla crucifer had the broadest microhab-itat width both years. Relative to H. crucifer, H.chrysoscelis had narrower microhabitat widths bothyears but stood in varying relationship to the otherspecies. The other congeneric pairs, involving Am-bystoma and Rana show reversals of relative rankof microhabitat widths for 1974 and for 1975.
There are no straightforward, powerful, statisticaltests to apply to measures based on proportions todetermine statistical differences from one year to the

NUMBER 242 11
TABLE 5.—Microhabitat breadths (largest/smallest = largest value divided bysmallest to determine magnitude of difference from one year to next;see text)
Species 1974 1975 largest/smallest
Ambystoma maculatum,Arriby8toma opacum....Hy la eruciferBy la ehrysoaoelis...Rana olamitansRana sylvaticaBufo amerieanusAcris arepitans
0.01791.35186.11091.13070.32935.42280.01560
0.97110.0194
13.55490.01812.17500.20350.58500.0003
54.269.72.2
62.56.6
26.637.5
next. For this reason, the larger value is simply di-vided by the smaller value to determine the orderof magnitude of difference from one year to thenext. Biologically, there was but a single speciesthat had the same microhabitat breadth for bothyears: Acris crepitans essentially was absent bothyears. I interpret the differences between micro-habitat widths measured in 1974 and in 1975 to bebiologically significant for all of the other species.
Microhabitat Overlap: The formula used to de-termine microhabitat overlap (Horn, 1966) is:
ajk = 2Y,pijpik
where jk = overlap of species / and k, and pif isthe proportion of occurrence of species ; in sweep i.Values range from 0 (no overlap) to 1 (completeoverlap). Larval salamanders and larval anuransare in different trophic levels. Tadpoles are pri-marily herbivorous, while salamander larvae arecarnivorous and sometimes eat tadpoles. Co-occur-rences in sweeps (which the overlap measure repre-sents in a single value) thus give information onpotential predator-prey and competition relation-ships between salamander larvae and tadpoles. Highoverlap values are interpreted as the potential (butnot necessarily the existence) for predation or com-petition to occur. Low overlap values are inter-preted as indicating that predation or competitionis not important.
Overlap values for all comparisons involvingsalamanders are presented in Table 6. The verylow overlap value for 1975 between the two speciesof Ambystoma and no overlap in 1974 confirmsprevious conclusions (Worthington, 1968; 1969) that
larval Ambystoma maculatum and A. opacum essen-tially do not co-occur in the same ponds.
In examining the sweep data for possiblepredator-prey relationships between the salamandersand tadpoles, the data for all salamanders (in addi-tion to being compared on a species-species over-lap with tadpoles) were combined, on the assump-tion that both species would be potential tadpolepredators and it would make no difference to tad-pole species x whether it was preyed upon by eithersalamander species a or b. It is clear from the re-sults (Table 6) that Ambystoma opacum potentiallyis not an effective tadpole predator in the studypond. Almost all potential salamander predationis accounted for by Ambystoma maculatum. Twoadditional generalizations may be drawn from theoverlap values. First, potential salamander preda-tion, on all species of tadpoles, was much greaterin 1975 than in 1974. The differences involved sug-gest that salamander predation on tadpoles prob-ably did not have much of an effect in 1974, whilesalamander predation on tadpoles in 1975 couldhave been a strong natural selective force. Second,predictions can be made about which species oftadpoles should most likely be affected by salaman-der predation. In 1974, only Rana sylvatica shouldhave had any noticeable predation by salamanderlarvae. In 1975, Rana clamitans and Rana sylvaticashould have been preyed on extensively bysalamander larvae. Such predictions are besttreated cautiously until more is known about feed-ing habits in larval salamanders. For example, thepredator-prey size relationships are unknown, butthe Rana tadpoles become quite large and mayescape predation after reaching a certain size.

12 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
TABLE 6.—Microhabitat overlaps for comparisons involving salamander larvae (see Table 5for further explanation)
Pai r comparison
All salamanders - Hyla aruciferAll salamanders - Hyla chrysoscelisAll salamanders - Rana alamitansAll salamanders - Rana sylvatioaAll salamanders - Bufo ameriaanusAll salamanders - Aoria crepitans
Amby8toma maaulatum - Ambystoma opaaum
Ambystoma maculatum - Hyla aruciferAmbystoma maaulatum - Hyla ahrysoscelis....
Ambystoma maaulatum - Rana aylvatioa
Amby8toma maaulatum - Acris arepitansAmbystoma opaaum - Hyla aruaiferAmbystoma opaaum - Hyla dhrysosaelis
Ambystoma opaaum - Rana sylvatiaa
Ambystoma opaaum - Aerie orepitans
1974
0.05830.06030.03850.11110.00040
0
0.06250.09760.06120.1163000.0085000.02240.00070
1975
0.18790.11060.29200.25740.15650.0168
0.0126
0.19380.11490.26830.26940.16190.01680.000300.01960.02320.00120
largest/smallest
3.21.87.62.3
391.2
"
-
3.11.24.42.3
--
28.3--
1.01.7
~
Walters (1975) demonstrated that Ambystoma lar-vae have definite prey preferences among tadpoleswhen offered a choice; however, she did not in-clude aquatic insects in her experiments. Larvalsalamanders may prefer insects over tadpoles, evento the extent that tadpoles are not eaten, althoughavailable. Anderson and Graham (1967) demon-strated that vertical migration and stratificationoccur in larval Ambystoma. They found that duringthe evening the larval salamanders leave the sitesthey have occupied during the day and begin ac-tively feeding. If most feeding is done at dusk, thenthe overlap values reported here have little or nobearing on the predator-prey relationships of larvalsalamanders and tadpoles, since all sweeps weretaken during the day. Walters (1975) reported di-urnal foraging in Ambystoma larvae, but had nodata to indicate whether most of the foraging takesplace during the day or at dusk.
All sweeps were taken when tadpoles were active,so the overlap values represent potential competi-tion when tadpole species-tadpole species compari-sons are made. All of the tadpoles in this studyhave the same gross feeding morphology. Thus,competition may be potentially occurring among all
individuals of all species. For this reason, overlapshave been calculated for each species comparedwith all other tadpoles in addition to species-species overlap comparisons (Table 7). There is noexperimental evidence with which to relate overlapvalues to potential competition among and withinspecies. Does a value of 0.1062 mean that competi-tion is strong enough so that selection will operateto reduce the overlap? For purposes of this paper,the following common-sense hypothesis will be usedto evaluate the overlap values. The range of valuesfrom about 0.1 to 0.25 represents the degree of po-tential competition that the process of natural selec-tion would operate to reduce the microhabitatoverlap. Values below 0.1 indicate potential com-petition is not significant, and values above 0.25indicate potential competition is significant.
Each of three species had roughly the sameamount of overlap with all other tadpoles in 1975that it had in 1974: Hyla chrysoscelis, Rana syl-vatica, and Acris crepitans. Of these, potential com-petition was a marginal factor only for Ranasylvatica, and that for only 1974. For each of theother three species, the degree of overlap with allother tadpole species was different in 1975 from

NUMBER 242 IS
TABLE 7.—Microhabitat overlaps for comparisons involving only tadpoles (see Table 5 forfurther explanation)
Pair comparison 1974 1975 largest/smallest
Hyla crucifer - All other tadpoles. . . .Hyla chry808celi8 - All other tadpolesEana alcanitans - All other tadpoles.. .Rana aylvatica - All other tadpoles.. .Bufo americanus - All other tadpoles..Aaris crepitans - All other tadpoles..
Hyla aruaifer -Hyla chrysoscelieHyla crucifer - Rana clamitansHyla aruaifer - Rana sylvaticaHyla aruaifer - Bufo americanusHyla aruaifer r Aarie orepitansHyla chry8O8celi8 - Rana clamitans....Hyla chrysoscelis - Rana eylvatiaa....Hyla chrysosaelis - Bufo americanus...Hyla chrysoecelie - Aaris arepitane...Rana clamitans - Rana sylvaticaRana clamitans - Bufo americanus-.Rana clamitans - Acris arepitansRana sylvatiaa - Bufo americanusRana sylvatica - Aaris arepitansBufo americanus - Aaris arepitane
0.15310.05640.03950.10620.00960
0.03050.02040.13850.0146000.0336000.018900000
0.32250.04380.15190.06790.09670.0076
0.07040.16650.11370.12150.00740.09800.04360.02150.05850.22310.10600.01890.16990.02320
2.11.33.81.610.1-
2.38.21.28.3--1.3--
11.8----
what it had been in 1974. For Hyla crucifer, po-tential competition was important both years butmore significant in 1975. For Rana clamitans, po-tential competition was a factor only in 1975, as italso was (marginally) for Bufo americanus. At thecommunity level it would appear that larvae ofHyla crucifer are the only ones that would experi-ence potential competition from the rest of thetadpole community on a year to year basis.
Although feeding morphologies of all tadpolesunder consideration are similar, interspecific compe-tition might be most important between certainspecies pairs. Each of the following species pairsshowed the same overlap in 1974 that they did in1975 (Table 7): Hyla crucifer-Rana sylvatica, Hylacrucifer-Acris erepitans, Hyla chrysoscelis—Ranasylvatica, and Bufo americanus-Acris erepitans.Among these, only the Hyla crucifer-Rana sylvaticapair shows an overlap value that indicates poten-tial competition as a factor. All of the other speciespair overlap values are very different for 1974 ascompared to those for 1975. For the following pairspotential competition is not a factor in either year:Hyla crucifer-Hyla chrysoscelis, Hyla chrysoscelis-
Bufo americanus, Hyla chrysoscelis-Acris erepitans,Rana clamitans-Acris erepitans, and Rana sylvatica-Acris erepitans. For the following pairs, potentialcompetition is significant for only one of the twoyears: Hyla crucifer-Rana clamitans, Hyla crucifer-Bufo americanus, Hyla chrysoscelis-Rana clamitans,Rana clamitans-Rana sylvatica, Rana clamitans-Bufo americanus, Rana sylvatica-Bufo americanus.Thus, based on the two consecutive years of thisstudy, 8/15 of the total species-species overlapsprobably do not have competitive interactions,while in only 1/15 of the species-species overlapsis competition potentially a year to year factor.
DISCUSSION.—The aquatic larval community ap-pears to have distinct temporal patterns each year.Some of the species show the same temporal pat-terns each year and others show different temporalpatterns each year.
Each species of larvae has essentially the samemicrohabitat preference for the two years.
The analyzed microhabit breadths reflect tem-poral and spatial partitioning with population den-sities, thereby illustrating the comparative habitatutilization each year. The overall picture is that

14 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
the total habitat utilization was very different ineach of the two years studied and would likely bedifferent every year.
The microhabitat overlap analyses allow insightsinto the biological interactions occurring in thepond. Overlaps of salamander larvae and tadpolesgive indications of predator-prey relationships,while overlaps of species in the same trophic levelgive insights into potential competition.
Salamander larvae can prey heavily on tadpolesin nature (Anderson, 1968). Even if most prey capture is done at twilight in microhabitats other thanthose occupied by the salamanders in daylight, theoverlap data presented herein exemplify the kindsof relationships that occur between predators andtadpole prey. The overlap data indicate that thetadpole populations in 1974 were in very differentpredator environments from those in 1975.
The lack of competition between Ambystomamaculatum and A. opacum in Maryland due totemporal partitioning has been documented and dis-cussed previously (Worthington, 1968; 1969). Therole of competition among tadpoles has not been aswell documented. Wilbur and Collins (1973) andBrockelman (1969) demonstrated that intraspecificcompetition among tadpoles may occur at high ex-perimental densities. Brockelman (1969) found noevidence for intraspecific competition in naturalpond habitats. De Benedictis (1974) was unable todemonstrate conclusively that interspecific competi-tion occurs in field simulated experiments. An anal-ysis of the available literature suggests that whilecompetition is a theoretical possibility among tad-poles, the densities found in nature fall well belowthe densities required to demonstrate competitionexperimentally. For example, comparative figuresextrapolated from Wilbur and Collins (1973) andTurnipseed and Altig (1975) indicate a density of 1tadpole per 192 cm3 of water as being necessary fortadpole competition to occur (based on the resultsof 300 larvae density runs in Wilbur and Collin'sexperimental pens), while the greatest recordedtadpole population density found in nature wasidentified in a pond edge in Mississippi as 1 tadpoleper 285 cm3 water (conservative estimate, using 5cm water depth to Turnipseed and Altigs' density/area data).
In order to demonstrate the occurrence of actualcompetition among tadpoles, it would be necessary
to determine a limiting factor in the environment.Tadpoles are not territorial; many species freelyform association groups and/or schools (Wassersug,1973). Thus, it appears unlikely that space wouldbe a limiting factor. The pond has abundant sub-merged vegetation and a deep organic silt bottom;all indications are that it is a productive pond andthat food would not be limiting. The tadpolespecies-species overlap values indicate poten-tial competition, not the existence of competi-tion. Most of the overlap values are low, indicatingthat there is little potential competition withinmost of the tadpole community. The highest con-sistent values involve Hyla crucifer (Table 7). Hylacrucifer has a small tadpole, and although the num-bers of individuals may be high, the biomass maynot be appreciable, especially compared to Ranaclamitans and R. sylvatica. The overall results ofthis study indicate that if interspecific competitionis occurring, it most likely is occurring betweenHyla crucifer and Rana sylvatica. Until experimentsare performed to determine the density levels re-quired to demonstrate competition between thesetwo species, the most reasonable assumption is thatcompetition is not important for these populationsat the study pond. The only other overlap valuereally indicative of potential competition is that ofRana clamitans-Rana sylvatica for 1975 (Table 7).While individuals of both species were occupyingthe same microhabitats, the total number of indi-viduals collected were not great: 148 R. clamitansand 79 R. sylvatica. These numbers came from areasonable volume of habitat sampled. I concludethat the total densities involved are of such a mag-nitude that competition was not a factor for Ranaclamitans and Rana sylvatica in 1975. In the ab-sence of any demonstrated limiting factor, the over-lap analysis provides no evidence that competitionamong tadpoles was occurring in the pond in eitheryear. Since the search for evidence of competitionproduced such generally negative results, it appearsthat competition was not likely limiting populationgrowth in any of the tadpole species in the studypond. What the overlap values do demonstrate isthat the temporal and spatial relationships withinthe biological larval community were quite differ-ent in 1974 from what they were in 1975.
The conclusions drawn with respect to habitatpartitioning for the study pond are as follows. An-

NUMBER 242 15
nual differences in larval temporal habitat parti-tioning combined with different annual populationdensities result in: (1) differential habitat utilizationeach year by the larvae, and (2) different biologicalinteractions involving predator-prey and speciesassociations each year. Generally low microhabitatoverlap values for tadpole species indicate that com-petition was not regulating the tadpole community,but that larval spatial and temporal distributionseffectively partitioned the habitats. Overall, eachlarval species was in a different environment withrespect to natural selection in 1974 from what itexperienced in 1975.
Barro Colorado Island
The stream habitats at Barro Colorado Island(BCI) consist of semi-isolated or completely iso-lated stream bed pools. Sampling was done once inthe dry season (11 February to 5 March 1975) andonce in the wet season (1-20 July 1975) to deter-mine gross temporal patterns of tadpole occurrence.Sampling procedures were designed to determinehow fish (predators) and tadpoles spatially occupythe habitat.
METHODS AND MATERIALS.—Each stream bed poolwas measured by length, breadth, and greatestdepth. The measured pool was then sampled by alarge or small dipnet appropriate to the size ofthe pond. Presence or absence of fish was noted.Fish were not collected, so the assumption that thefish were all predators probably overestimates thereal situation. In the dry season, the presence orabsence of tadpoles by species was noted. In a fewsamples the individuals were preserved for confirma-tion of identifications. In the wet season, all tad-poles collected were preserved. Efforts were madeto collect all individuals in the small ponds. In thelaboratory, all specimens were identified, counted,and staged (Gosner, 1960).
RESULTS.—Few larvae were collected in anysingle pond; for this reason, only the presence orabsence of larvae was noted in the dry season. Thewet season sampling, in which quantitative countswere taken, probably reflects the same quantitativerelationships as the dry season sampling. Duringthe wet season the greatest number of tadpoles col-lected in a pond was 18, the average number oftadpoles in a pond that had tadpoles was 3.6.
Ninety-seven ponds were sampled in the dry sea-son, 80 were sampled in the wet season (Table 8).The ponds were larger in the wet season (x 2 =
21.17, P<.01) as might be expected. Fish occurred ina total of 63 ponds, tadpoles occurred in a total of47 ponds. Fish and tadpoles co-occurred in only 1pond.
TABLE 8.—Size classes of ponds (volume in m*) sampled byseasons at Barro Colorado Island
Pond size
1 - 9....0.1 - 0.9..0.01 - 0.09.0.001 - 0.009
Dry season Wet season
2225221
643247
Distribution of fish among pond size classes ispresented in Table 9. Fish occurred in the largerponds. This aspect of habitat preference was evi-dent only in the dry season when there were moresmall ponds available to choose from. During thewet season, when ponds were larger, the fish dis-tribution among size classes of ponds was random.
The distribution of all tadpoles among pond sizeclasses is presented in Table 10. Tadpoles occurredin a random sample of the available pond sizes inboth the dry and wet seasons.
In the dry season, tadpoles of Bufo typhonius,Colostethus nubicola, and Smilisca sila were col-lected from the stream bed ponds. Bufo typhoniuswere collected in only one pond where they werecompleting metamorphosis. The sampling periodincluded the very end of the reproductive seasonof Bufo typhotiius. The Colostethus nubicola andSmilisca sila larvae were at all stages of development since the sampling period was during thebreeding season of these species. Although samplesizes are not large enough for statistical analysis,the raw data clearly suggests that Colostethusnubicola occurred randomly in the available pondsduring the dry season when small ponds were moreabundant (Table 11). The same applies to Smiliscasila larvae (Table 12). Colostethus nubicola tad-poles were found in 6 ponds where Smilisca silatadpoles did not occur, S. sila tadpoles occurred in7 ponds were C. nubicola did not occur, and C.nubicola and 5. sila co-occurred in 8 ponds. The

16 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
TABLE 9.—Fish occurrence among size classes of ponds (volume in m1) sampled by seasons atBarro Colorado Island (for x5 determination, sample sizes are grouped so that expected values> 5; expected values are determined by multiplying percent occurrence of all fish in all pondsby number of ponds sampled in each size category; see Table 1 for key to symbols)
Pond sizeDry season Wet season Total
Observed Expected Observed Expected Observed Expected
1 - 90.1 - 0 .9 . . ,0.01 - 0.09.0.001 - 0.009,
11728
2
11126in
= 9.55** X2 =
41
2.00 (ns)
32434
2
2232710
8.37**
TABLE 10.—Tadpole occurrence among size classes of ponds sampled by seasons at BarroColorado Island (for x2 determination, sample sizes are grouped so that the expected values> 5; expected values are determined by multiplying percent occurrence of all tadpoles in allponds by number of ponds sampled in each size category)
10nn
Pond
101 -001 -
size
90.90.090.009
DryObserved
12108
x2
seasonExpected
05115
= 2.69 (ns)
WetObserved
111122
x2
seasonExpected
21482
= 2.60 (ns)
TotalObserved
2132210
x2 -
Expected
217207
2.33 (ns)
TABLE 11.—Occurrence of Colostethus nubicola tadpoles among size classes of ponds sampledby seasons at Barro Colorado Island (for x2 determination, sample sizes are grouped so thatthe expected values > 5; expected values are determined by multiplying percent of occur-rence of all C. nubicola larvae in all ponds by number of ponds sampled in each size category)
Pond sizeDryi
Observed
0284
seasonExpected
0383
WetObserved
0382
seasonExpected
1741
TotalObserved
05
166
Expected
110124
1 - 90.1 - 0.9..0.01 - 0.09.0.001 - 0.009
X2 8.12** X2 5.52*
two species were randomly associated with eachother (x2 = .28,P>.05).
Colostethus nubicola, Centrolenella fleischmanni,and Physalaemus pustulosus larvae were collectedin the stream ponds in the wet season. All stages oflarvae were present indicating the sampling pe-riod was within the breeding season of all three
species. Physalaemus pustulosus larvae were col-lected in 6 ponds, in 1 pond it was the only speciescollected, in 1 pond Physalaemus pustulosus co-occurred with Colostethus nubicola, in 3 pondsPhysalaemus pustulosus co-occurred with Centro-lenella fleischmanni, and in 1 pond Physalaemuspustulosus co-occurred with Centrolenella fleisch-

NUMBER 242 17
TABLE 12.—Occurrence of Smilisca sila larvaeamong size classes of ponds sampled in one sea-son at Barro Colorado Island (expected valuesare determined by multiplying percent occur-rence of all S. sila larvae in all ponds by num-ber of ponds sampled in each size category)
Pond size
1 - 90.1 - 0.90.01 - 0.090.001 - 0.009
Dry seasonObserved Expected
1 02 37 85 3
manni and Colostethus nubicola. In the wet season,Colostethus nubicola demonstrated a greater occur-rence in smaller ponds than expected (Table 11).This preference was evident only in the wet season,when there were fewer small ponds than in the dryseason. Over all, Colostethus nubicola tadpoles wereassociated with smaller ponds (Table 11). Cen-trolenella fleischmanni tadpoles were distributedrandomly among the pond sizes (Table 13). Coloste-thus nubicola tadpoles were found in 10 pondswhere Centrolenella fleischmanni tadpoles werenot found, Centrolenella fleischmanni tadpoles werefound in 12 ponds where Colostethus nubicola tad-poles were not found, and both Centrolenellafleischmanni and Colostethus nubicola tadpoleswere found in 3 ponds. The two species tend tomutually exclude each other, but the statisticalevidence for this is not strong (x2 = 5.36, . 1 >P>.05):
TABLE 13.—Occurrence of Centrolenella fleisch-manni larvae among size classes of ponds sam-pled in one season at Barro Colorado Island(for x1 determination, sample sizes are groupedso that the expected values > 5; expected valuesdetermined by multiplying percent occurrenceof all C. fleischmanni larvae in all ponds by num-ber of ponds sampled in each size category)
Pond
10.10.010.001
size
- 9.- 0.- 0.- 0.
909009
WetObserved
1950
seasonExpected
1841
- .11 (ns)
HABITAT PARTITIONING.—In terms of habitat par-titioning, the habitat is considered as the entire col-lection of stream bed ponds.
Temporal Partitioning: The habitat is clearly par-titioned temporally over a seasonal cycle. Only onespecies, Colostethus nubicola, was present as tad-poles in the two seasonal sampling periods.
Spatial Partitioning: Space partitioning is evidentin the total pond system. Of all the ponds with tad-poles present, 68% had but a single species. Wheremore than one species occurred in a pond, I foundno evidence for microhabitat partitioning withinthe pond. All species were found in the leaf litterthat had accumulated in the pond bottoms.
The stream bed tadpole community was effec-tively partitioning the available habitat on thebasis of time and space.
DISCUSSION.—While the effective habitat parti-tioning evidenced in the BCI stream tadpoles seemsto be the product of selective forces acting previ-ously upon the species making up the community,the possible roles of predation and competitioninvite speculation.
Fish predation in the tropics recently has beenhypothesized to be a major organizing factor intadpole communities (Heyer, McDiarmid, andWeigmann, 1975). Two observations, in addition tothe data already presented, are pertinent to thequestion of possible fish predation as a factor inthe BCI study. The single pond sampled in whichtadpoles and fish co-occurred had a small surfacearea, but was a deep crevice pond, in the bottom ofwhich had accumulated a deep layer of leaf litter.The single Centrolenella fleischmanni larva col-lected in this pond was in the deep leaf litter. Oneof the streams sampled had a weir type dam con-structed on it with a pond behind it. Data from thislarge pond (7 x 6 x 1 m) have not been includedherein previously, since the pond is not part of thenatural stream bed habitat. In the dry season, fishwere in the pond and no tadpoles were collectedalthough Physalaemus foam nests were often ob-served on the periphery of the pond. In earlyMarch, the pond was completely drained, and theaccumulated silt shoveled out. In the followingJuly, in numerous observations, no fish were seenbut a dense tadpole community was present con-sisting of Agalychnis callidryas and Physalaemuspustulosus. The data presented herein are consist-

18 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
ent with the following conclusions. Fish are effec-tive predators on tadpoles, completely eliminatingthem from certain pond habitats. The tadpoleshave two patterns of occurrence in response to thefish: (1) Colostethus nubicola demonstrates a hab-itat preference, using ponds smaller than thoseusually occupied by fish; (2) the remaining speciesapparently exploit the patchy occurrence of fish inthe total pond habitat system. That is, tadpoles aredeposited in enough ponds so that some of suchponds will be fish free. For such an accommodationto be effective, either a single egg clutch wouldhave to be partitioned among several ponds or theindividuals would have to have multiple clutchesduring their reproductive life. Relative to theanuran species identified in the BCI study, the onlyclutch deposition patterns available are for Centro-lenella fleischmanni, in which the male has beenshown to fertilize several clutches (data for femalesunknown, McDiarmid, 1975) and Physalaemuspustulosus, in which the female can produce up tofour clutches a year with as little as six weeks be-tween clutches (Davidson and Hough, 1969).
The very low numbers of tadpoles collected inthe individual ponds strongly suggests that compe-tition is not operating. The existence of manyponds with neither fish nor tadpoles, which ap-peared similiar to the ponds containing tadpolessuggests that there was no competition for spaceas a limiting factor by itself. It is very difficult, how-ever, to rule out competition (at least intraspecific)for a combination of space and food. In contrast tothe many temperate zone temporary ponds that arehighly productive, many tropical streams have ex-tremely low productivity (Fittkau, et. al., 1975). Nodata are available for the productivity of thestreams on BCI, but the productivity is probablylow, judging by the clearness of the water. Produc-tivity may be higher during the dry season, whentapir feces remain in the ponds rather than beingcontinually flushed from them. Interspecific compe-tition for food resources is not a likely factor, sincethe species which most often co-occur with eachother have very different feeding morphologies.Colostethus nubicola larvae have large funnelmouths; the larvae often feed on the surface film.The species that co-occur with Colostethus in thesame ponds, Smilisca sila in the dry season andCentrolenella fleischmanni in the wet season, are
substrate feeders with scraping and chewing mouth-parts. Each pond, however, may well have limitedresources. The tadpoles are certainly capable ofmoving from pond to pond during rains when theponds are interconnected. The low numbers ofindividuals in each pond may represent movementof individuals reflecting competition. The total dis-tribution pattern of larvae would then be a com-bination of the pond site where eggs were depositedand a radiation to adjoining ponds.
The data and analysis from the BCI study allowthe following general conclusions to be drawn. Thestream bed pond habitat system is effectively parti-tioned by the tadpole community on the basis ofspace and time. Predation is an organizing factor inthe composition of the tadpole community, inter-specific competition apparently is not.
Other Studies of Larval Habitat Partitioning
Larval salamander population dynamics havebeen the focus of study by several workers, mostnotably Anderson and his coworkers (1967, 1968,1971), Shoop (1974), Wilbur (1972), Wilbur andCollins (1973), and Worthington (1968, 1969). Mostof these studies have dealt with predation, compe-tition, survival, and temporal organization of larvalsalamander communities. Wilbur (1972), combinedexperimental manipulations and field observationsto demonstrate that predation and competitionwere important factors in the functioning and or-ganization of the larval salamander communities ofMichigan.
I am aware of only four studies in which quan-titative data were taken to deal with questions con-cerning habitat partitioning by anuran larvae.Dixon and Heyer (1968) followed a pond in westernMexico for two weeks. Since all microhabitats sam-pled were combined, no information was availableon possible spatial partitioning within the pond.The data strongly indicated temporal partitioningof the habitat by the larvae.
I previously reported on habitat partitioning ofthe temporary pond environment by the tadpolecommunity for a seasonal tropical site in north-eastern Thailand (1973, 1974). Quantitative sweepswere taken from a series of ponds to determine thetemporal partitioning patterns and a series of mi-crohabitat sweeps were taken from a single pond to

NUMBER 242 19
determine spatial partitioning within a pond. Gutcontents were analyzed to determine any obviouspartitioning patterns of the available food resources.The study concluded that the available pond hab-itats were partitioned by the tadpole communityon the basis of time and space, not food.
Calef (1973a) reported on the population dy-namics of a single anuran species system in a lakein British Columbia. The study is of interest asit is the only reported study of tadpole populationdynamics that covered two years (consecutive) andthe study represents an extreme situation in termsof anuran diversity. Calef (1973a) found similar re-sults in both years: almost identical numbers of eggmasses were laid in both years and survivorship wasessentially the same both years. Food was not a lim-iting resource; the tadpoles grew at the maximumrate set by the aquatic habitat temperature. Thetadpole populations were regulated by predation(primarily salamander).
In an unpublished study, Wiest (1974), reportedon a series of ponds in Brazos County, Texas. Wiestquantitatively sampled the larvae of a series ofponds and reported the individual results fromfour of them. He did not sample for microhabitatdifferences within ponds. Each of the four pondshad different combinations of species of larvae.Within each pond a temporal succession of specieswas demonstrated.
The results of all studies that have examined theentire anuran larval communities are similar andconsistent with the following conclusions. Habitatsare partitioned by space and time, not food. Preda-tion, not competition, has a strong role in control-ling and organizing the tadpole communities.
Tadpole Community Organization
A tadpole community is defined as an assemblageof anuran larval species at a single site. Two ex-treme levels of organization are possible: (1) a com-munity with no interspecific interactions betweenor among any of the component populations, or (2)a community with either symbiotic relationshipsthat tie the community together, or competitionthat partitions the community into componentparts. With this spectrum of tadpole communitiesin mind, I have examined the Chesapeake and BCIsystems to see what type of community organizationthey most closely resemble.
The three predominant species collected at BCIare all stream breeding frogs: Centrolenella fleisch-manni, Colostethus nubicola, and Smilisca sila. Theevolutionary history of each of the three species isdistinct. All members of the genus Centrolenellafor which larvae are known have rheocolous larvaewith long, thin tails, bodies flattened dorsoventrally,and a strong body and tail musculature capable ofcontinued, powerful flipping movements. The mor-phology is typical of larvae adapted to life in run-ning water. Mouthparts consist of a scraping andchewing apparatus. Breeding activities for all spe-cies that have been studied coincide with the onsetof the rainy season in those areas with seasonal rain-fall patterns. Centrolenella fleischmanni occurs fromsouthern Mexico to Panama, westward to Ecuador,and eastward to Surinam (Starrett and Savage,1973). Colostethus nubicola larvae resemble Centro-lenella larvae in body and tail shape, strong mus-culature, and flipping movements when out ofwater. Colostethus nubicola larvae differ fromCentrolenella larvae in that the mouthparts form apronounced funnel, adapted for feeding on thesurface film. Colostethus nubicola extends fromCosta Rica through Panama into western Colombia(Savage, 1968). The tadpole of Smilisca sila has atypical pond morphology in contrast to the streammorphologies of the other two species. The bodyand tail are weakly muscled, the body is globoseand the tail relatively short but broad, with highfins. The primitive reproductive pattern for thegenus Smilisca involves pond larvae (Duellman andTrueb, 1966). Smilisca sila has become adapted tobreeding in quiet stream bed pools. Such poolspredictably exist only during the dry season, whichis the breeding season for Smilisca sila. The mouth-part morphology is the typical scraping and chewingkind. They are known only from Costa Rica andPanama (Duellman, 1970).
In each of the seasons (dry and wet) there are twoco-occurring species in the community composed ofthe three predominant species. In the dry season,the surface film feeding C. nubicola larvae coexistwith the substrate processing S. sila larvae. In thewet season, the surface film feeding C. nubicolalarvae coexist with the substrate processing C.fleischmanni larvae. The community organizationof these three species can be explained in two verydifferent ways. First, it may be that competition for

20 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
resources was the source of natural selective forceswhich led to the morphological divergence ofmouthparts of Colostethus and the temporal parti-tioning of the substrate feeding Centrolenella andSmilisca larvae. The evolutionary result would thusbe a competition free, compartmentalized streambed larval community. The second explanation isthat the three species occur as larvae in a com-munity in the way they do because there happen tooccur in the same area three species of frogs thathave stream breeding adaptations. The organizationof the community is due to the way the componentscoexist, no further evolutionary adjustments beingnecessary. I favor the last explanation for the fol-lowing reasons. (1) The associations of the species aslarvae appear to be due to similar choices of pondhabitats by the three species. If the associationswere biologically mediated (e.g., competition) onewould not expect the random associations found inthis study. (2) The geographic distributions of thespecies argue against long term, species wide inter-specific interactions. Divergence in morphology ortemporal use of the habitat would almost have tobe a one sided divergence involving only Smiliscasila, as it is the only species which throughout itsentire range would consistently encounter the othertwo species. (3) The partitioning of the habitat bythe stream tadpole community can be better ex-plained in terms of other features of the life his-tories of the species involved. The reason that C.nubicola larvae have funnel mouthpart streamadapted larvae is likely that the species evolved inan environment characterized by streams lackingstanding algal growths or deteritus and the foodresources were concentrated in the surface film. Thereason that Smilisca sila breeds during the dry sea-son is likely that the ancestral stock, having a typi-
cal pond life history, became isolated in an areawhere the only quiet ponds in the environmentwere in the stream beds during the dry season. Thereason that Centrolenella fleischmanni breeds in therainy season is likely because the exposed eggclutches that are placed on undersides of leavesover streams are prone to desiccation during the dryseason and rains frequently are needed to insurethat the hatchlings reach an appropriate streamhabitat.
The same kinds of arguments can be made forthe Chesapeake tadpole community. Table 14 pre-sents the major species wide characteristics of breed-ing habitats and seasons for those species comprisingthe tadpole community at the Chesapeake study site(data from Conant, 1975; R. I. Crombie and G. R.Zug, pers. comms.). Most of the species have uniquecombinations of breeding seasons and types ofponds primarily used for breeding. These data sug-gest that each species has a unique reproductivepattern reflecting the conditions in which thatspecies occurred at the time of its genetic isolation.If interspecific competition had been the majorevolutionary force in the habitat partitioning be-tween congeners, their segregation should remainapparent today, although they would no longer bein competition. The essentially complete partition-ing of larvae of Hyla crucifer from Hyla chryso-scelis and Rana clamitans from Rana sylvaticacould be interpreted as competition having oc-curred between congeneric pairs historically, withnatural selection operating to reduce larval inter-specific competition. The present almost completehabitat partitioning within the species pairs wouldbe the result of natural selection operating to re-duce competition. The alternate explanation oflarval habitat partitioning within congeneric
TABLE 14.—Major characteristics of breeding habitats and seasons for species comprising thetadpole community at the Chesapeake site (C = common, r = rare)
Species
Aavie crepitans....Bufo americanua....Hyla aruaiferBy la chry808celie..Rana clamitansRana ayIvatica
Breeding season Types of breeding pondsEarly Late Temporary Permanent Sluggish
wooded open wooded open streams
C r C CC C C r rC C C r r
C r r C rC r C C
C C r

NUMBER 242 21
pairs seems more plausable. The ancestral stockof Hyla crucifer likely was isolated in and becameadapted to an environment in which the only avail-able breeding ponds were temporary ones whichappeared in the spring. The ancestral stockof Hyla chrysoscelis was likely isolated in andbecame adapted to an environment in which theonly available breeding ponds were permanent onesand in addition, the breeding season was associatedwith late spring and summer warm rains. Similarly,the ancestral stock of Rana sylvatica was likely iso-lated in and became adapted to an environmentin which the only available breeding ponds weretemporary wooded ponds appearing in the spring.The ancestral stock of Rana clamitans was likelyisolated and became adapted to an environment inwhich the only breeding sites were streams andopen, permanent ponds and also the breeding sea-son was associated with late spring and summerwarm rains. Since the early adaptive histories of theseveral species, the geographic ranges have ex-panded and contracted according to major climato-logical changes so that the species ranges nowbroadly overlap. Each pond in the species rangewill have (more or less) some of the major char-acteristics associated with the primary adaptivepond type of each species. Thus, certain woodedpermanent ponds will contain the essential fea-tures of the temporary wooded ponds to whichRana sylvatica is primarily adapted. When suchponds are encountered, they will be utilized byR. sylvatica in the spring, while the same pond mayalso be used by Rana clamitans in the late springand early summer. The habitat partitioning of thelarvae of these two species is thus most likely not aresult of biological interactions at the larval stages,but rather a passive reflection of different adaptivereproductive patterns of the species involved. Asimilar observation may be made for the entirecommunity of frogs utilizing the same ponds. Ifbiological interactions at the larval stage wereimportant in the control and organization of thetadpole community, the interactions would have tobe reasonably predictable for natural selection tobe effective. The Chesapeake community organiza-tion argues against this. Each component speciespresent as larvae was in an essentially different se-lective environment in each of the two yearsstudied.
Although the role of competition among tadpoleshas implicitly (e.g., Wiest, 1974) or explicitly(Wilbur and Collins, 1973) been accorded majorimportance in the control and organization of tad-pole communities, this present study concludesthat interspecific competition among tadpoles isnot important in the control or organization of tad-pole communities. Further circumstantial evidencearguing against significant tadpole competition innature is relevant to this conclusion. If interspecifictadpole competition had occurred historically,selection should have operated to displace the com-peting species either with respect to space, time, orfeeding morphologies. If this were true, a predictioncould be stated that tadpoles with the same feedingmorphologies feeding on the same foods within thesame type microhabitat should not occur in thesame pond at the same time. Data are availablefrom two pond studies which do not support thisprediction. For tadpoles with scraping and chewingmouthparts, the best test is to look for co-occurring,similar size larvae belonging to the same genus.Wiest (1974) did not sample for microhabitat dif-ferences, but one of the ponds he reported hadonly one microhabitat. This was pond D, which wasa uniform, shallow grassy pond in a heavily grazedpasture. The same larval stages of Pseudacris clarkiand P. streckeri, which have the same mouthparts,occurred together in this pond. The beakless mid-water feeding microhylid genera at Sakaerat,Thailand, all have morphologically similar larvae,of the same size, and all feed on the same food inthe same way (Heyer, 1973). Three ponds therethat had a single uniform microhabitat were sam-pled. For all practical purposes, the ponds weredirt aquaria. There was no standing vegetation inthe water. If competition had been important his-torically, no two species of any of the midwaterfeeding genera should have occurred together inthese ponds, however, the following pairs did occurtogether, refuting the importance of the potentialfor interspecific tadpole competition in nature:Glyphoglossus molossus and Microhyla pulchra,Glyphoglossus molossus and Kaloula mediolincata,Kaloula pulchra and Microhyla pulchra (Heyer,1973). In another pond which was sampled bymicrohabitats, Glyphoglossus molossus and Micro-hyla ornata larvae occurred in the same sweeps.
Interspecific behavioral adjustments appear to be

22 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
rare among tadpoles, indicating that generally,interspecific competition among tadpoles is evolu-tionarily unimportant. In field experiments,Agalychnis larvae occupied the water column andthe pond floor when it was the only species present,but when Hyla rosenbergi, a bottom dwelling tad-pole species, was in the same pond, the Agalychnislarvae all occupied the water column (McDiarmid,pers. comm.). Two partially exclusive explanationsmay account for the type of behavioral shift evi-denced by the Agalychnis larvae. First, there iscompetition for space when Agalychnis is presentwith bottom dwelling tadpoles which may occasiona behavioral response by Agalychnis: partitioningthe habitat to avoid competition. Second, the re-sponse by Agalychnis may be purely density depend-ent. Individual Agalychnis larvae may occupy thebottom if there are less than a given number oflarvae already present on the bottom (includingother Agalychnis individuals). If there are manytadpoles on the bottom, at least some of theAgalychnis larvae will occupy the water column.If all the bottom larvae belong to another species,the results would appear to be habitat partitioningin response to another species, not just other indi-vidual tadpoles, but with so little data, it is impos-sible to choose among the alternatives. Larvalbehavioral shifts in response to other species appearto be rare, however, and they appear to be part ofthe total adaptive set of the species that evidencesthe behavioral switch. Thus the response is com-pletely one sided; there are no accompanying be-havioral shifts in other members of the samecommunity. My prediction is that behavioralchanges in response to other tadpoles will be foundin relatively specialized larvae, such as midwateradapted Agalychnis, such behavioral flexibilityallowing greater exploitation of the habitat. Thus,when no other bottom dwelling tadpoles are pres-ent, Agalychnis larvae will utilize the bottom, eventhough their adaptive morphology is specializedfor midwater feeding (Starrett, 1973).
Another factor that would refute larval compe-tition as the origin of temporal partitioning bytadpoles is the following. At the point in the specia-tion process where reproductive isolation is beingreinforced by selection, one premating isolatingmechanism that can undergo character displacementis temporal partitioning of the breeding season. If
selection operates to displace the breeding seasonof two closely related taxa, the larvae would alsodemonstrate temporal habitat partitioning.
Finally, a theoretical model based on a com-munity analagous to a tadpole community predictsthat several phytoplankton cropping species maycoexist without competitive exclusion (Smith, et.al., 1975).
The available evidence indicates that communi-ties of tadpoles represent nothing more than acollection of species co-occurring as larvae. A tad-pole community is not an integrated whole; bio-logical interactions within the community are notimportant evolutionarily; there is no natural selec-tion operating directly to integrate or partition thecommunity. Similar conclusions have been reportedrecently in other aquatic communities, involvingpredatory fish (Mendelson, 1975) and dragonflylarvae (Benke and Benke, 1975).
The Generalized Anuran Life Cycleand Its Evolutionary Maintenance
Wassersug (1975), from a morphological analysisof tadpole adaptive types, proposed that the tad-pole stage evolved to exploit the bursts of primaryproductivity characteristic of temporary ponds. Thetemporary food resources are exploitable becausethe tadpole stage is temporary, coinciding with thebursts of primary productivity.
Heyer, McDiarmid, and Weigmann (1975) pro-posed a model that from an ecological viewpointpresents reasons why the generalized anuran lifehistory should be maintained in temporary pondsduring the larval stage. As long as there are tem-porary ponds with bursts of primary produc-tivity, selection will favor the maintenance ofthe generalized anuran life cycle which includes theherbivorous tadpole stage. The total environmentfor which the tadpole stage is adapted is markedby extreme variability and unpredictability. Theenvironment is unpredictable from a climatic-physical aspect. Ponds may be present in a locationin one year and absent the next (Wiest, 1974;Collins, 1975). The biological environment variesin the same pond from one year to the next(Chesapeake data). For any species found in aseries of ponds, the biological environment variesfrom pond to pond (Heyer, 1973; Wiest, 1974;BCI data).

NUMBER 242 23
One evolutionary response designed to exploitunpredictable environments is to maximize bothreproductive effort and genetic variability simul-taneously. The amphibian genetic system appearsto be well suited to maximize genetic variability inits reproductive effort. Polyploidy is not known tooccur in higher vertebrates (parthenogenetic lizardsexcluded), possibly because the higher vertebrateis a complexly integrated machine that can notfunction if the genetic controls are changed to anygreat degree. Having an extra chromosome set maybe too much of a genetic change to allow the highervertebrate cell to function properly. Polyploidy isknown for lower vertebrates, including frogs, andcould be interpreted to mean that the amphibiancell can tolerate considerable genetic changes andstill function. (This argument does not imply thatmammals are more highly evolved than frogs. Itdoes imply that frogs have more evolutionarymechanisms available than mammals.) Wilson,Sarich, and Maxson (1974) have shown that closelyrelated species of frogs have much larger geneticdifferences than similar pairs of mammal species.They assumed constant rates of protein evolutionand concluded the frog species are much olderthan the mammal species. Another interpretationis that many mutations in albumin structure maybe deleterious in the homeostatic mammalian sys-tem although such can be tolerated in the less wellregulated amphibian system. These inferences sug-gest that compared to the mammalian genetic sys-tem, the anuran genetic system is preadapted to,and continually selected for, maintenance of largeamounts of variability. One of the reasons largeclutch size may be adaptive in frogs is that a widevariety of genotypes can be expressed in everyreproductive effort. Some of the genotypes likelywould be adaptive in the situation in which anyegg clutch was laid, but due to the unpredictabilityof the pond environments, there is no way fornatural selection to operate to select a genotypethat will always work best. Natural selection wouldoperate to select those individuals that maximizedgenetic variability in each reproductive effort. It isnot necessary to postulate an increased mutationrate; selection would maintain, rather than reduce,the variability present.
There are no available direct data on theamounts of genetic variation present in a single
large frog egg clutch, but there is circumstantialevidence to suggest that the amount of variation isconsiderable. Many frog egg clutches brought infrom the field and raised under laboratory condi-tions experience heavy mortality, particularly atthree stages: gastrulation, hatching, and metamor-phosis. These three times represent periods whendifferent parts of the genotype are activated for thefirst time. Death may be due to defective geno-types. High genetic variability would account forthis high rate of genetic death as many of thecombinations are lethal in some egg clutches. Thus,it would appear that the number of genotypes ismaximized within a single clutch. The larger theclutch, the more possible genotypes can be exposedand selected in the pond where the clutch is laid.
In addition to the maximizing of individualclutch size, breeding adults should produce multipleclutches in order to ensure reproductive success inan unpredictable environment. Crump (1974) foundthis to be true for the single gravid female recap-ture she reported: Hyla rhodopepla is a small,pond breeding hylid which produced 2 clutches in31 days. This is the only known data on tropicalpond breeding frogs other than that for Physalae-mus previously discussed (Davidson and Hough,1969). To achieve maximum reproductive effort,frogs in the temperate zones could either be longlived, producing one clutch a year, or if shortlived, produce multiple clutches in one year. Somespecies of temperate frogs are known to be longlived, reproducing over several seasons (Collins,1975). Rana clamitans combines long life and theproduction of multiple clutches within a repro-ductive season (literature summarized in Collins,1975). There are no data on number of clutcheslaid per season by small species; in Michigan,most individuals of small frog species have onlyone reproductive season, but some individualsbreed over at least two reproductive seasons(Collins, 1975). Thus frogs exploiting temporaryponds with large clutch size and multiple clutcheswould have selective advantages over frogs withsmaller clutch sizes and/or single clutches. Saltheand Duellman (1973) demonstrated that, of all thefrogs they studied, pond breeding frogs have eggclutches with the most eggs.
The generalized anuran life cycle involving large,multiple clutches with essentially random popula-

24 SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY
tion control at the larval level should occasionallyresult in great densities of frogs when, due to therandom character of tadpole population control,there is little or no tadpole population reduction.Such outbreaks of frogs have been reported innewspapers and at least once in scientific journals(Scott and Starrett, 1974).
In some ways, the situation Calef (1973a) de-scribed is not consistent with the conclusionsreached in this paper (see "Other Studies of LarvalHabit Partitioning" above). The larval populationdynamics were very similar in the two consecutiveyears studied by him. At the northern distributionallimit of frog occurrence, one would not expect tofind a system where larval population structuresare the same from one year to the next. If the larvalpopulations are as stable as the two year studyindicates, the simplicity of the system may wellaccount for this stability. There is one species offrog which breeds in permanent water in BritishColumbia. The predatory salamanders are perma-nent residents of the lake in which these frogsbreed. Calef (pers. comm.) found salamanders inall ponds and lakes where Rana aurora bred. Thesystem is apparently in a steady state and predicta-ble; ponds are effectively permanent, predator andprey interaction always occur between the samepredator and the same prey. Under these pre-dictable conditions, Calef (1973a) concluded thatthe biological interactions (predator-prey) controland stabilize the community structure and popula-tion density of Rana aurora larvae. Calef's studyadds support to the hypothesis that modificationsof the anuran generalized life cycle only occur inrelatively predictable environments (see "Evolu-tionary Modifications of the Generalized AuranLife Cycle" below). His study also indicates thatnot all predictable environments are limited totropical wet forests.
Selection, Population Control,and the Anuran Life Cycle
The data, analyses, and conclusions presentedhere are summarized in terms of features of the lifecycle where natural selection and population con-trol likely are most effective.
Calling Sites: Competition for calling sites ismost evident in those species with long breedingseasons. In those species for which calling sites havebeen a limiting factor, territoriality has been a com-mon evolutionary response (for example: Ranaclamitans, Wells, pers. comm.; dendrobatids,Duellman, 1966, Silverstone, 1975; centrolenids,McDiarmid and Adler, 1974). Territoriality invol-ving male calling sites can be an important popu-lation control mechanism.
Egg Deposition Sites: Egg deposition sites maybe the same as calling sites (e.g. centrolenids,McDiarmid, 1975) or within the defended territorybut not identical with calling sites (e.g. dendro-batids, Crump, 1972; Silverstone, 1973). Territori-ality may have arisen in response to limitedavailability of egg deposition sites. Some non-territorial frogs have specific egg deposition sites,the availability of which may control the popula-tion size of the species (e.g. some Phyllomedusa laytheir eggs on leaves of particular species of plants,Pyburn, pers. comm.).
Clutch Size and Number: As discussed above,selection should favor large clutch size and multipleclutches in frogs with generalized life cycles. Amost effective population control at this level isclimatic uncertainty, such as ponds drying up oregg masses freezing. Egg predation can be importantwithin ponds (Walters, 1975) but unpredictableamong ponds or from year to year. In these situa-tions, selection will favor large, multiple clutches.
Larvae: Certain kinds of predator-prey inter-actions can have an evolutionary effect on type ofpond choice by species of frogs. As argued else-where (Heyer, McDiarmid, and Weigmann, 1975),for selection to operate, the predator must elimi-nate, not reduce, tadpole populations. Predation byfish (Heyer, McDiarmid, and Weighmann, 1975)and turtles (Heyer and Muedeking, in prep.) hasbeen identified as a factor that has likely pro-duced evolutionary change in terms of choice ofponds; the smaller, temporary ponds, free of fishand turtles being favored. The relationship betweenpond size and occurrence of (either, but not both)fish and Colostethus larvae in the BCI stream pondsis consistent with this hypothesis.
Salamander and invertebrate predators within

NUMBER 242 25
temporary ponds may effectively reduce populationsizes (Calef, 1973a; Walters, 1975), but the level ofpredation usually is variable among ponds or fromyear to year. In these situations, selection will favormaximizing variability of reproductive efforts; thatis, large, multiple clutches will be favored.
Adults: Direct data are not available, but itseems likely that all effective population controloccurs in the stages already listed. Competition forresources (other than calling or egg deposition sites)is probably not important. Scott and Starrett (1974)reported on an unusual situation where the numberof adult Agalychnis spurrelli was several timesgreater in one area than normally encountered.This suggests that adult frog populations are nor-mally well below the carrying capacity of theenvironment. Calef (1973a) suggested that if survi-vorship from juvenile to adult Rana aurora weresimilar to the rate of larval survival, the adultpopulation would be sufficient to maintain thepopulation, but he found adult populations to be3 to 6 times as large as those needed to maintainthe population (Calef, 1973b). Thus, for Ranaaurora, most population reduction is occurring inthe larval stage.
Evolutionary Modifications of theGeneralized Anuran Life Cycle
Temporary ponds that are characterized by burstsof primary productivity occur in seasonal environ-ments. The appearance and duration of such pondsis highly variable. By contrast, environments withlittle seasonality do not have bursts of primaryproductivity in their ponds, the ponds are longlived, and the biological communities more pre-dictable and stable. In relatively aseasonal, stableenvironments, selection 'should operate to finelyadapt the organism to the environment. Therewould not be a selective advantage to maximizinggenetic variability within a clutch. In fact, selectionshould operate to reduce genetic variability to theoptimum genotype(s) for the stable environmentin which the species occurs. Large clutch size andmultiple clutches would not have selective advan-tages per se. Evolution could operate on otherparameters involving smaller clutch sizes that wouldadapt the life cycle to stable environments, such asparental care, brooding, and increase of yolk peregg, which is a prerequisite for direct developmentin most frogs (Salthe and Duellman, 1973; also seeHeyer, 1975).

Literature Cited
Anderson, J. D.1968. A Comparison of the Food Habits of Ambystoma
macrodactylum sigillatum, Ambystoma macrodacty-lum croceum, and Ambystoma tigrinum californ-iense. Herpetologica, 24:273-284.
Anderson, J. D., and R. E. Graham1967. Vertical Migration and Stratification of Larval
Ambystoma. Copeia, 1967:371-374.Anderson, J. D., D. D. Hassinger, and G. H. Dalrymple
1971. Natural Mortality of Eggs and Larvae of Ambystomat. tigrinum. Ecology, 52:1107-1112.
Benke, A. C, and S. S. Benke1975. Comparative Dynamics and Life Histories of Co-
existing Dragonfly Populations. Ecology, 56:302-317.Brockelman, W. Y.
1969. An Analysis of Density Effects and Predation inBufo americanus Tadpoles. Ecology, 50:632-644.
Calef, G. W.1973a. Natural Mortality of Tadpoles in a Population of
Rana aurora. Ecology, 54:741-758.1973b. Spatial Distribution and "Effective" Breeding Pop-
ulation of Red-legged Frogs (Rana aurora) inMarion Lake, British Columbia. The CanadianField-Naturalist, 87:279-284.
Collins, J. P.1975. A Comparative Study of the Life History Strategies
in a Community of Frogs, ix + 148 pages. Ph.D.dissertation, University of Michigan.
Conant, R.1975. A Field Guide to Reptiles and Amphibians of East-
ern and Central North America. Second edition,xii + 429 pages. Boston: Houghton Mifflin Co.
Crump, M. L.1972. Territoriality and Mating Behavior in Dendrobates
granuliferus (Anura: Dendrobatidae). Herpetologica,28:195-198.
1974. Reproductive Strategies in a Tropical Anuran Com-munity. University of Kansas Museum of NaturalHistory Miscellaneous Publications, 61:1-68.
Davidson, E. H., and B. R. Hough1969. Synchronous Oogenesis in Engystomops pustulosus,
a Neotropic Anuran Suitable for Laboratory Studies:Localization in the Embryo of RNA Synthesized atthe Lampbrush Stage. Journal of ExperimentalZoology, 172:25-48.
DeBenedictus, P. A.1974. Interspecific Competition Between Tadpoles of Rana
pipiens and Rana sylvatica: An Experimental FieldStudy. Ecological Monographs, 44:129-151.
Dixon, J. R , and W. R. Heyer1968. Anuran Succession in a Temporary Pond in Colima,
Mexico. Bulletin of the Southern California Acad-emy of Sciences, 67:129-137.
Duellman, W. E.1966. Aggressive Behavior in Dendrobatid Frogs. Herpeto-
logica, 22:217-221.1970. The Hylid Frogs of Middle America. University of
Kansas Museum of Natural History Monograph,1:1-753.
Duellman, W. E., and L. Trueb1966. Neotropical Hylid Frogs, Genus Smilisca. University
of Kansas Museum of Natural History Publications,17:281-375.
Fittkau, E. J., U. Irmler, W. J. Junk, F. Reiss, and G. W.Schmidt
1975. Productivity, Biomass, and Population Dynamics inAmazonian Water Bodies. Pages 289-311 in F. B.Golley and E. Medina, editors, Tropical EcologicalSystems: Trends in Terrestrial and Aquatic Re-search, xii + 398 pages. New York: Springer-Verlag.
Gosner, K. L.1960. A Simplified Table for Staging Anuran Embryos
and Larvae with Notes on Identification. Herpeto-logica, 16:183-190.
Heyer, W. R.1973. Ecological Interactions of Frog Larvae at a Seasonal
Tropical Location in Thailand. Journal of Her-petology, 7:337-361.
1974. Niche Measurements of Frog Larvae from a Sea-sonal Tropical Location in Thailand. Ecology,55:651-656.
1975. A Preliminary Analysis of the Intergcneric Relation-ships of the Frog Family Leptodactylidae. Smith-sonian Contributions to Zoology, 199:1-55.
Heyer, W. R., R. W. McDiarmid, and D. L. Weigmann1975. Tadpoles, Predation and Pond Habitats in the
Tropics. Biotropica, 7:100-111.
Heyer, W. R., and M. H. MuedekingIn prep. Notes on Tadpoles as Prey for Naiads and Tur-
tles.Horn, H. S.
1966. The Measurement of "Overlap" in ComparativeEcological Studies. American Naturalist, 100:419-424.
Levins, R.1968. Evolution in Changing Environments: Some The-
26

NUMBER 242 27
oretical Explorations. Princeton University Mono-graphs in Population Biology, 2:1-120.
McDiarmid, R. W.1975. Glass Frog Romance along a Tropical Stream.
Terra, 13:14-18.McDiarmid, R.TV., and K. Adler
1974. Notes on Territorial and Vocal Behavior of Neo-tropical Frogs on the Genus Centrolenella. Her-petologica, 30:75-78.
Mendelson, J.1975. Feeding Relationships among Species of Notropis
(Pisces: Cyprinidae) in a Wisconsin Stream. Eco-logical Monographs, 45:199-230.
Salthe, S. N., and W. E. Duellman1973. Quantitative Constraints Associated with Reproduc-
tive Mode in Anurans. Pages 229-249 in J. L. Vial,editor, Evolutionary Biology of the Anurans: Con-temporary Research on Major Problems, xii + 470pages. Columbia: University of Missouri Press.
Savage, J. M.1968. The Dendrobatid Frogs of Central America. Copeia,
1968:745-776.Scott, N. J., Jr., and A. Starrett
1974. An Unusual Breeding Aggregation of Frogs, withNotes on the Ecology of Agalychnis spurrelli (Anura:Hylidae). Bulletin of the Southern California Acad-emy of Sciences, 73:86-94.
Shoop, C. R.1974. Yearly Variation in Larval Survival of Ambystoma
maculatum. Ecology, 55:440-444.Silverstone, P. A.
1973. Observations on the Behavior and Ecology of aColombian Poison-arrow Frog, the K6koe-pa (Den-drobates histrionicus Berthold). Herpetologica,29:295-301.
1975. Frogs Worthy of a Maiden's Kiss. Terra, 13:19-21.Smith, O. L., H. H. Shugart, R. V. O'Neill, R. S. Booth, andD. C. McNaught
1975. Resource Competition and an Analytical Model ofZooplankton Feeding on Phytoplankton. The Amer-ican Naturalist, 109:571-591.
Starrett, P. H.1973. Evolutionary Patterns in Larval Morphology. Pages
251-271 in J. L. Vial, editor, Evolutionary Biologyof the Anurans: Contemporary Research on MajorProblems, xii + 470 pages. Columbia: University ofMissouri Press.
Starrett, P. H., and J. M. Savage1973. The Systematic Status and Distribution of Costa
Rican Glassfrogs, Genus Centrolenella (Family Cen-trolenidae), with Description of a New Species.Bulletin of the Southern California Academy ofSciences, 72:57-78.
Turnipseed, G., and R. Altig1975. Population Density and Age Structure of Three
Species of Hylid Tadpoles. Journal of Herpetology,9:287-291.
Walters, B.1975. Studies of Interspecific Predation within an Amphib-
ian Community. Journal of Herpetology, 9:267-279.Wassersug, R. J.
1973. Aspects of Social Behavior in Anuran Larvae. Pages273-297 in J. L. Vial, editor, Evolutionary Biologyof the Anurans: Contemporary Research on MajorProblems, xii + 470 pages. Columbia: University ofMissouri Press.
1975. The Adaptive Significance of the Tadpole Stagewith Comments on the Maintenance of Complexlife Cycles in Anurans. American Zoologist, 15:405-417.
Wiest, J. A., Jr.1974. Anuran Succession at Temporary Ponds in a Post
Oak Savannah Region of Texas. 177 pages. M.S.thesis, Texas A 8c M University.
Wilbur, H. M.1972. Competition, Predation, and the Structure of the
Ambystoma-Rana sylvatica Community. Ecology,53:3-21.
Wilbur, W. H., and J. P. Collins1973. Ecological Aspects of Amphibian Metamorphosis.
Science, 182:1305-1314.Wilson, A. C, V. M. Sarich, and L. R. Maxson
1974. The Importance of Gene Rearrangement in Evolu-tion: Evidence from Studies on Rates of Chro-mosomal, Protein, and Anatomical Evolution.Proceedings of the National Academy of Sciences,71:3028-3030.
Worthington, R. D.1968. Observations on the Relative Sizes of Three Species
of Salamander Larvae in a Maryland Pond. Her-petologica, 24:242-246.
1969. Additional Observations on Sympatric Species ofSalamander Larvae in a Maryland Pond. Herpeto-logica, 25:227-229.


REQUIREMENTS FOR SMITHSONIAN SERIES PUBLICATION
Manuscripts intended for series publication receive substantive review within theiroriginating Smithsonian museums or offices and are submitted to the Smithsonian InstitutionPress with approval of the appropriate museum authority on Form SI-36. Requests forspecial treatment—use of color, foldouts, casebound covers, etc.—require, on the sameform, the added approval of designated committees or museum directors.
Review of manuscripts and art by the Press for requirements of series format and style,completeness and clarity of copy, and arrangement of all material, as outlined below, willgovern, within the judgment of the Press, acceptance or rejection of the manuscripts and art.
Copy must be typewritten, double-spaced, on one side of standard white bond paper,with 1V4" margins, submitted as ribbon copy (not carbon or xerox), in loose sheets (notstapled or bound), and accompanied by original art. Minimum acceptable length is 30 pages.
Front matter (preceding the text) should include: title page with only title and authorand no other information, abstract page with author/title/series/etc., following the establish-ed format, table of contents with indents reflecting the heads and structure of the paper.
First page of text should carry the title and author at the top of the page and an unnum-bered footnote at the bottom consisting of author's name and professional mailing address.
Center heads of whatever level should be typed with initial caps of major words, withextra space above and below the head, but with no other preparation (such as all caps orunderline). Run-in paragraph heads should use period/dashes or colons as necessary.
Tabulations within text (lists of data, often in parallel columns) can be typed on the textpage where they occur, but they should not contain rules or formal, numbered table heads.
Formal tables (numbered, with table heads, boxheads, stubs, rules) should be sub-mitted as camera copy, but the author must contact the series section of the Press for edito-rial attention and preparation assistance before final typing of this matter.
Taxonomic keys in natural history papers should use the alined-couplet form in thezoology and paleobiology series and the multi-level indent form in the botany series. Ifcross-referencing is required between key and text, do not include page references within thekey, but number the keyed-out taxa with their corresponding heads in the text.
Synonymy in the zoology and paleobiology series must use the short form (taxon,author, yearpage), with a full reference at the end of the paper under "Literature Cited."For the botany series, the long form (taxon, author, abbreviated journal or book title, volume,page, year, with no reference in the "Literature Cited") is optional.
Footnotes, when few in number, whether annotative or bibliographic, should be typedat the bottom of the text page on which the reference occurs. Extensive notes must appear atthe end of the text in a notes section. If bibliographic footnotes are required, use the shortform (author/brief title/page) with the full reference in the bibliography.
Text-reference system (author/year/page within the text, with the full reference in a"Literature Cited" at the end of the text) must be used in place of bibliographic footnotes inall scientific series and is strongly recommended in the history and technology series:"(Jones, 1910:122)" or ".. . . Jones (1910:122)."
Bibliography, depending upon use, is termed "References," "Selected References," or"Literature Cited." Spell out book, journal, and article titles, using initial caps in all majorwords. For capitalization of titles in foreign languages, follow the national practice of eachlanguage. Underline (for italics) book and journal titles. Use the colon-parentheses systemfor volume/number/page citations: "10(2):5-9." For alinement and arrangement ofelements, follow the format of the series for which the manuscript is intended.
Legends for illustrations must not be attached to the art nor included within the text butmust be submitted at the end of the manuscript—with as many legends typed, double-spaced, to a page as convenient.
Illustrations must not be included within the manuscript but must be submitted sepa-rately as original art (not copies). All illustrations (photographs, line drawings, maps, etc.)can be intermixed throughout the printed text. They should be termed Figures and shouldbe numbered consecutively. If several "figures" are treated as components of a single largerfigure, they should be designated by lowercase italic letters (underlined in copy) on the illus-tration, in the legend, and in text references: "Figure 9JD." If illustrations are intended to beprinted separately on coated stock following the text, they should be termed Plates and anycomponents should be lettered as in figures: "Plate 9b_." Keys to any symbols within anillustration should appear on the art and not in the legend.
A few points of style: (1) Do not use periods after such abbreviations as "mm, ft,yds, USNM, NNE, AM, BC." (2) Use hyphens in spelled-out fractions: "two-thirds." (3)Spell out numbers "one" through "nine" in expository text, but use numerals in all othercases if possible. (4) Use the metric system of measurement, where possible, instead ofthe English system. (5) Use the decimal system, where possible, in place of fractions.(6) Use day/month/year sequence for dates: "9 April 1976." (7) For months in tabular list-ings or data sections, use three-letter abbreviations with no periods: "Jan, Mar, Jun," etc.
Arrange and paginate sequentially EVERY sheet of manuscript—including ALL frontmatter and ALL legends, etc., at the back of the text—in the following order: (1) title page,(2) abstract, (3) table of contents, (4) foreword and/or preface, (5) text, (6) appendixes,(7) notes, (8) glossary, (9) bibliography, (10) index, (11) legends.



















