
Structure-dependent Ah receptor agonist activities of chlorinated biphenylenes
4
Structure-dependent Ah receptor agonist activities of chlorinated biphenylenes Shaheen Khan a , Alex Konstantinov b , Brock Chittim b , Alan McAlees b , Brian Yeo b , Stephen Safe a,c, * a Department of Veterinary Physiology and Pharmacology, Texas A&M University, College Station, TX 77843-4466, USA b Wellington Laboratories, 345 Southgate Dr. Guelph, Ont., Canada N1G3M5 c Institute of Biosciences and Technology, Texas A&M University System Health Science Center, 2121 W. Holcombe Blvd., Houston, TX 77030-3303, USA Received 20 February 2006; accepted 11 April 2006 Available online 3 May 2006 Abstract Polychlorinated biphenylenes (PCBP) have been identified as combustion by-products that bind the aryl hydrocarbon receptor (AhR) and exhibit 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD)-like activity. This study investigates the Ah-responsiveness of 2,3,6,7-tetrachlo- robiphenylene (2,3,6,7-CBP), 2,3,6-CBP, 2,3-CBP and 2-CBP in breast cancer cells. MCF-7 or ZR-75 cells were treated with different concentrations (1–100 nM) of the compounds alone to determine their activity as inducers of CYP1A1 protein expression or luciferase activity in cells transfected with a construct (pDRE 3 ) containing three tandem dioxin responsive elements (DREs) linked to a luciferase reporter gene. In both assays, the order of potency was 2,3,6,7-CBP > 2,3,6-CBP > 2,3-CBP 2-CBP, and 2,3,6,7-CBP and TCDD were equipotent. Similar results were also observed in an antiestrogenic assay in MCF-7 cells, confirming the high AhR agonist activity of 2,3,6,7-CBP in breast cancer cells. Ó 2006 Elsevier Ltd. All rights reserved. Keywords: Ah receptor; Polychlorinated biphenylenes; MCF-7; ZR-75 1. Introduction Polychlorinated aromatics are industrial chemicals and their by-products that have been identified as contaminants in almost every component of the ecosystem (Rappe, 1992; Rappe, 1993). Compounds such as the polychlorinated biphenyls (PCBs), dibenzo-p-dioxins (PCDDs), and dibenzofurans (PCDFs) are highly persistent in the envi- ronment, and their bioaccumulation in fish, wildlife and humans and potential adverse effects have been of concern to government and intergovernment regulatory agencies (Kutz et al., 1991; Rappe, 1992; Rappe, 1993; Alcock et al., 1998; van Leeuwen et al., 2000; Noren and Mei- ronyte, 2000; Schecter et al., 2001). PCBs, PCDDs and PCDFs invariably occur as complex mixtures in environ- mental samples, and structure–toxicity relationships have identified 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) as the most toxic of the polychlorinated aromatic compounds (Poland et al., 1979; Poland and Knutson, 1982; Safe, 1986; Goldstein and Safe, 1989). The toxic effects of TCDD have been extensively characterized and the mechanism of action involves binding to and subsequent activation of the intra- cellular aryl hydrocarbon receptor (AhR), which modu- lates expression of multiple genes that results in the downstream tissue-specific biochemical and toxic responses (Poland et al., 1979; Poland and Knutson, 1982; Safe, 1986). The AhR is activated by structurally-diverse phytochemi- cals and structurally-related halogenated aromatics (Poland et al., 1979; Poland and Knutson, 1982; Safe, 1986; Safe, 1990; Seidel et al., 2000; Denison and Nagy, 2003), and there 0887-2333/$ - see front matter Ó 2006 Elsevier Ltd. All rights reserved. doi:10.1016/j.tiv.2006.04.003 * Corresponding author. Address: Department of Veterinary Physiology and Pharmacology, Texas A&M University, College Station, TX 77843- 4466, USA. Tel.: +1 979 845 5988; fax: +1 979 862 4929. E-mail address: [email protected] (S. Safe). www.elsevier.com/locate/toxinvit Toxicology in Vitro 20 (2006) 1234–1237
-
Upload
shaheen-khan -
Category
Documents
-
view
214 -
download
1
Transcript of Structure-dependent Ah receptor agonist activities of chlorinated biphenylenes

www.elsevier.com/locate/toxinvit
Toxicology in Vitro 20 (2006) 1234–1237
Structure-dependent Ah receptor agonist activitiesof chlorinated biphenylenes
Shaheen Khan a, Alex Konstantinov b, Brock Chittim b, Alan McAlees b,Brian Yeo b, Stephen Safe a,c,*
a Department of Veterinary Physiology and Pharmacology, Texas A&M University, College Station, TX 77843-4466, USAb Wellington Laboratories, 345 Southgate Dr. Guelph, Ont., Canada N1G3M5
c Institute of Biosciences and Technology, Texas A&M University System Health Science Center, 2121 W. Holcombe Blvd., Houston, TX 77030-3303, USA
Received 20 February 2006; accepted 11 April 2006Available online 3 May 2006
Abstract
Polychlorinated biphenylenes (PCBP) have been identified as combustion by-products that bind the aryl hydrocarbon receptor (AhR)and exhibit 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD)-like activity. This study investigates the Ah-responsiveness of 2,3,6,7-tetrachlo-robiphenylene (2,3,6,7-CBP), 2,3,6-CBP, 2,3-CBP and 2-CBP in breast cancer cells. MCF-7 or ZR-75 cells were treated with differentconcentrations (1–100 nM) of the compounds alone to determine their activity as inducers of CYP1A1 protein expression or luciferaseactivity in cells transfected with a construct (pDRE3) containing three tandem dioxin responsive elements (DREs) linked to a luciferasereporter gene. In both assays, the order of potency was 2,3,6,7-CBP > 2,3,6-CBP > 2,3-CBP � 2-CBP, and 2,3,6,7-CBP and TCDD wereequipotent. Similar results were also observed in an antiestrogenic assay in MCF-7 cells, confirming the high AhR agonist activity of2,3,6,7-CBP in breast cancer cells.� 2006 Elsevier Ltd. All rights reserved.
Keywords: Ah receptor; Polychlorinated biphenylenes; MCF-7; ZR-75
1. Introduction
Polychlorinated aromatics are industrial chemicals andtheir by-products that have been identified as contaminantsin almost every component of the ecosystem (Rappe, 1992;Rappe, 1993). Compounds such as the polychlorinatedbiphenyls (PCBs), dibenzo-p-dioxins (PCDDs), anddibenzofurans (PCDFs) are highly persistent in the envi-ronment, and their bioaccumulation in fish, wildlife andhumans and potential adverse effects have been of concernto government and intergovernment regulatory agencies(Kutz et al., 1991; Rappe, 1992; Rappe, 1993; Alcocket al., 1998; van Leeuwen et al., 2000; Noren and Mei-
0887-2333/$ - see front matter � 2006 Elsevier Ltd. All rights reserved.
doi:10.1016/j.tiv.2006.04.003
* Corresponding author. Address: Department of Veterinary Physiologyand Pharmacology, Texas A&M University, College Station, TX 77843-4466, USA. Tel.: +1 979 845 5988; fax: +1 979 862 4929.
E-mail address: [email protected] (S. Safe).
ronyte, 2000; Schecter et al., 2001). PCBs, PCDDs andPCDFs invariably occur as complex mixtures in environ-mental samples, and structure–toxicity relationships haveidentified 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) asthe most toxic of the polychlorinated aromatic compounds(Poland et al., 1979; Poland and Knutson, 1982; Safe, 1986;Goldstein and Safe, 1989). The toxic effects of TCDD havebeen extensively characterized and the mechanism of actioninvolves binding to and subsequent activation of the intra-cellular aryl hydrocarbon receptor (AhR), which modu-lates expression of multiple genes that results in thedownstream tissue-specific biochemical and toxic responses(Poland et al., 1979; Poland and Knutson, 1982; Safe,1986).
The AhR is activated by structurally-diverse phytochemi-cals and structurally-related halogenated aromatics (Polandet al., 1979; Poland and Knutson, 1982; Safe, 1986; Safe,1990; Seidel et al., 2000; Denison and Nagy, 2003), and there

S. Khan et al. / Toxicology in Vitro 20 (2006) 1234–1237 1235
has been extensive research on the toxicology of TCDD-likehalogenated aromatics due to their potential adverse effectson humans and wildlife. Polychlorinated biphenylenes(PCBPs) have been identified as combustion products ofPCBs (Rappe et al., 1982). Studies by Poland and coworkers(1979) showed that 2,3,6,7-tetrachlorobiphenylene (CBP)exhibited AhR-dependent potencies similar to that ofTCDD. This study now reports the structure–activity rela-tionships among four synthetic chlorinated biphenylenesusing induction of CYP1A1 or CYP1A1-dependent activi-ties and their antiestrogenic responses in MCF-7 breastcancer cells as endpoints. The results show that 2,3,6,7-CBP and 2,3,6-CPB significantly induced CYP1A1, whereasthe 2,3-dichloro- and 2-chloro isomers were inactive. More-over, a similar structure–activity relationship was alsoobserved for the antiestrogenic effects of the CBPs, and thisis another well-characterized AhR-mediated response (Safeand Wormke, 2003). These studies confirm the potent AhRagonist activity of 2,3,6,7-CBP.
2. Materials and methods
2.1. Preparation of chlorinated biphenylenes
Chlorinated biphenylenes were synthesized essentiallyusing the method previously described by Baker andcoworkers (Baker et al., 1958). The structures and purity(>98%) were confirmed by 1H NMR and GC–MS. Meltingpoints were determined on a Kofler hot-stage, and areuncorrected. Proton NMR spectra were recorded fordichloromethane-d2 solutions on a 400 MHz Bruker spec-trometer (University of Guelph, Guelph, Canada).Observed couplings are reported in Hz. High resolutiongas chromatography/low resolution mass spectrometryanalyses were conducted on a Shimadzu GCMS-QP2010using a J&W 15 m DB-5HT column (0.25 mm ID, 0.1 lMfilm). The injections were done in splitless mode, with theinjector temperature at 300 �C and a helium carrier gas flowof 1.50 ml/min. The following temperature program wasused: initial oven temperature: 140 �C; hold of 5 min; rampat 10 �C/min to 325 �C; hold 20 min. Full scan spectra (50–1000 amu) were obtained. Ions with relative intensitiesgreater than 15% are reported along with the chemical shiftdata for the proton NMR of the four CBP congeners.
2.2. 2-Chlorobiphenylene
m.p. 70–71 �C (lit. 67.5–68.5 �C); m/z (rel. intensity) 188(M + 2, 33%), 186 (M+, 100%), 151 (48%), 150 (30%), 93(16%), 75 (25%); NMR: d 6.57 (1H, dd, J 7.2 & 0.8, H4),6.63 (1H, dd, J 1.2 & 0.8, H1), 6.68 (2H, m, H6 & H7),6.73 (1H, dd, J 7.2 & 1.2, H3), 6.79 (2H, m, H5 & H8).
2.3. 2,3-Dichlorobiphenylene
m.p. 136–138 �C; m/z (rel. intensity) 222 (M + 2, 65%),220 (M + 1, 100%), 185 (21%), 150 (47%), 110 (16%), 75
(21%); NMR: d 6.71 (2H, s, H1 & H4), 6.74 (2H, m, H6& H7), 6.85 (2H, m, H5 & H8).
2.4. 2,3,6-Trichlorobiphenylene
m.p. 160–161 �C; m/z 258 (M + 4, 30%), 256 (M + 2,95%), 254 (M + 1, 100%), 219 (15%), 184 (39%), 92(23%); NMR: d 6.66 (1H, d, J 8.2, H5), 6.73 (1H, d, J
1.2, H8), 6.75 (1H, s, H4), 6.76 (1H, s, H1), 6.84 (1H, dd,J 8.2 & 1.2, H6).
2.5. 2,3,6,7-Tetrachlorobiphenylene
Decomposition >200 �C; m/z 292 (M + 4, 47%), 290(M + 2, 100%), 288 (M + 1, 81%), 220 (21%), 218 (33%),110 (16%), 109 (25%); NMR: d 6.81 (4H, s, H1,H4,H5,H8).
TCDD was synthesized in the laboratory by the conden-sation of 4,5-dichlorocatechol plus 2,4,5-trichloronitroben-zene and purified as described (Mason et al., 1986) to give>99% purity as determined by chromatographic analysis.
2.6. Cells and chemicals
MCF-7 and ZR-75 cells were obtained from AmericanType Culture Collection (ATCC, Manassas, VA) and werecultured in DME-F12 (Sigma) media supplemented with5% fetal bovine serum (FBS). DMEM/F12 [Dulbecco’smodified Eagles medium (DMEM) Ham F-12 (DMEM/F-12)] and antibiotic/antimycotic solution and 17b-estradiol(E2) were purchased from Sigma (St. Louis, MO). FBSwas obtained from JRH Biosciences (Lenexa, KS). Lucifer-ase and b-galactosidase enzyme assay systems were obtainedfrom Promega Corp. (Madison, WI). Sp1 and CYP1A1 anti-bodies were obtained from Santa Cruz Biotechnology, Inc.(Santa Cruz, CA). TCDD was made in this laboratory.
2.7. Western blot analysis
MCF-7 cells were seeded into six-well plates in phenolred-free medium DMEM/F-12 supplemented with 2.5%charcoal-stripped FBS. After 18 h, cells were treated withdifferent compounds for 12 h and then harvested in lysisbuffer (500 mM sodium chloride (NaCl), 50 mM HEPES(pH 7.5), 10% glycerol, 1.5 mM magnesium chloride(MgCl2), 1% Triton X-100, 1 mM EGTA) on ice for30 min followed by centrifugation at 13,000 rpm for15 min. Protease inhibitor cocktail (Sigma) was added tolysis buffer at 1:100 dilution. Fifty micrograms of proteinfrom each treatment group were loaded on 10% SDS poly-acrylamide gel electrophoresis (SDS-PAGE), and trans-ferred to polyvinylidene difluoride (PVDF) membrane.Membranes were blocked in Blotto (5% milk, Tris-bufferedsaline, and 0.05% Tween20) and probed with primary anti-bodies CYP1A1 H-270 (1:500) and Sp1 PEP2 (1:1,000) for2 h and then probed with anti-Rabbit secondary antibodyfor 1 h. Bands were visualized using the ECL detection sys-tem (NEN) and scanned using a Sharp JX-330 scanner.
0
1500
2000
1000
500Rel
ativ
eL u
c if e
r as e
a ct iv
ity
10nM
1nM
100
nM
10nM
1nM
100
nM
10nM
100
nM
DM
SO
DM
SO
DM
SO
DM
SO
DM
SO
1Mμ μ μ
1M
1M
10nM
100
nM
10nM
100
nM
TCDD 2,3,6,7-CBP 2,3,6-CBP 2,3-CBP 2-CBP
* * *
*
*
**
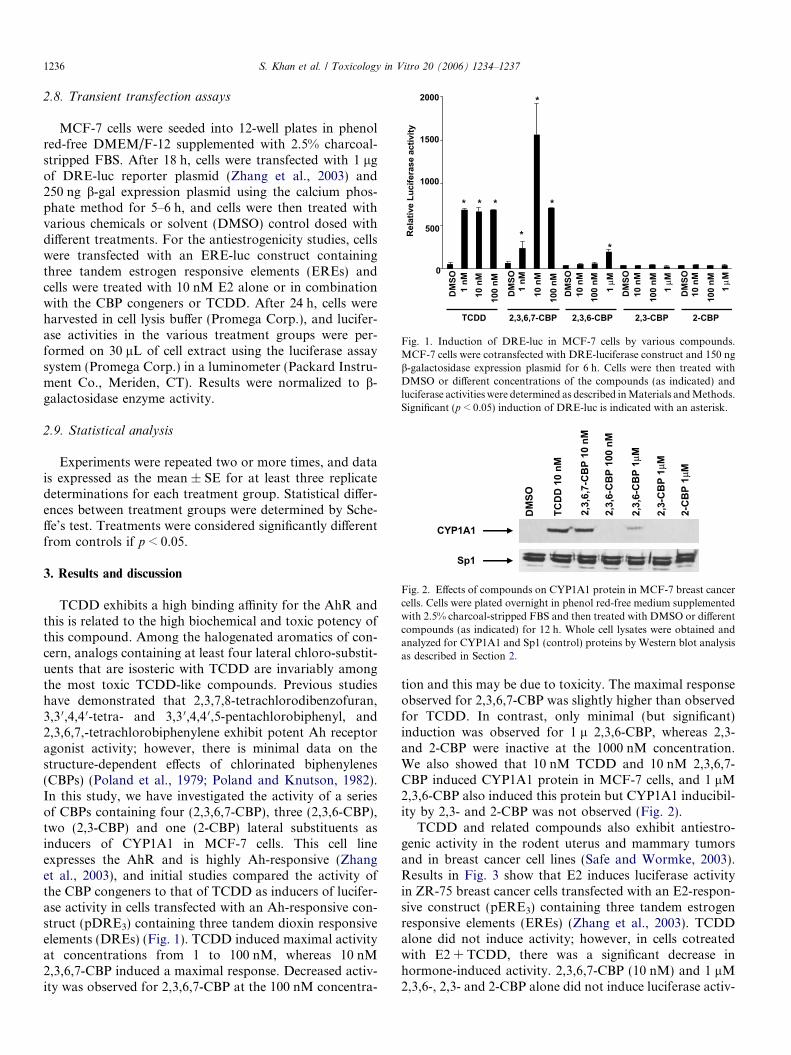
Fig. 1. Induction of DRE-luc in MCF-7 cells by various compounds.MCF-7 cells were cotransfected with DRE-luciferase construct and 150 ngb-galactosidase expression plasmid for 6 h. Cells were then treated withDMSO or different concentrations of the compounds (as indicated) andluciferase activities were determined as described in Materials and Methods.Significant (p < 0.05) induction of DRE-luc is indicated with an asterisk.
DM
SO
TCD
D 1
0 nM
2,3,
6,7-
CB
P 10
nM
2,3,
6-C
BP
100
nM
2,3,
6-C
BP
1M
2,3-
CB
P 1μ μ
μ M
2-C
BP
1M
CYP1A1
Sp1
Fig. 2. Effects of compounds on CYP1A1 protein in MCF-7 breast cancercells. Cells were plated overnight in phenol red-free medium supplementedwith 2.5% charcoal-stripped FBS and then treated with DMSO or differentcompounds (as indicated) for 12 h. Whole cell lysates were obtained andanalyzed for CYP1A1 and Sp1 (control) proteins by Western blot analysisas described in Section 2.
1236 S. Khan et al. / Toxicology in Vitro 20 (2006) 1234–1237
2.8. Transient transfection assays
MCF-7 cells were seeded into 12-well plates in phenolred-free DMEM/F-12 supplemented with 2.5% charcoal-stripped FBS. After 18 h, cells were transfected with 1 lgof DRE-luc reporter plasmid (Zhang et al., 2003) and250 ng b-gal expression plasmid using the calcium phos-phate method for 5–6 h, and cells were then treated withvarious chemicals or solvent (DMSO) control dosed withdifferent treatments. For the antiestrogenicity studies, cellswere transfected with an ERE-luc construct containingthree tandem estrogen responsive elements (EREs) andcells were treated with 10 nM E2 alone or in combinationwith the CBP congeners or TCDD. After 24 h, cells wereharvested in cell lysis buffer (Promega Corp.), and lucifer-ase activities in the various treatment groups were per-formed on 30 lL of cell extract using the luciferase assaysystem (Promega Corp.) in a luminometer (Packard Instru-ment Co., Meriden, CT). Results were normalized to b-galactosidase enzyme activity.
2.9. Statistical analysis
Experiments were repeated two or more times, and datais expressed as the mean ± SE for at least three replicatedeterminations for each treatment group. Statistical differ-ences between treatment groups were determined by Sche-ffe’s test. Treatments were considered significantly differentfrom controls if p < 0.05.
3. Results and discussion
TCDD exhibits a high binding affinity for the AhR andthis is related to the high biochemical and toxic potency ofthis compound. Among the halogenated aromatics of con-cern, analogs containing at least four lateral chloro-substit-uents that are isosteric with TCDD are invariably amongthe most toxic TCDD-like compounds. Previous studieshave demonstrated that 2,3,7,8-tetrachlorodibenzofuran,3,3 0,4,4 0-tetra- and 3,3 0,4,4 0,5-pentachlorobiphenyl, and2,3,6,7,-tetrachlorobiphenylene exhibit potent Ah receptoragonist activity; however, there is minimal data on thestructure-dependent effects of chlorinated biphenylenes(CBPs) (Poland et al., 1979; Poland and Knutson, 1982).In this study, we have investigated the activity of a seriesof CBPs containing four (2,3,6,7-CBP), three (2,3,6-CBP),two (2,3-CBP) and one (2-CBP) lateral substituents asinducers of CYP1A1 in MCF-7 cells. This cell lineexpresses the AhR and is highly Ah-responsive (Zhanget al., 2003), and initial studies compared the activity ofthe CBP congeners to that of TCDD as inducers of lucifer-ase activity in cells transfected with an Ah-responsive con-struct (pDRE3) containing three tandem dioxin responsiveelements (DREs) (Fig. 1). TCDD induced maximal activityat concentrations from 1 to 100 nM, whereas 10 nM2,3,6,7-CBP induced a maximal response. Decreased activ-ity was observed for 2,3,6,7-CBP at the 100 nM concentra-
tion and this may be due to toxicity. The maximal responseobserved for 2,3,6,7-CBP was slightly higher than observedfor TCDD. In contrast, only minimal (but significant)induction was observed for 1 l 2,3,6-CBP, whereas 2,3-and 2-CBP were inactive at the 1000 nM concentration.We also showed that 10 nM TCDD and 10 nM 2,3,6,7-CBP induced CYP1A1 protein in MCF-7 cells, and 1 lM2,3,6-CBP also induced this protein but CYP1A1 inducibil-ity by 2,3- and 2-CBP was not observed (Fig. 2).
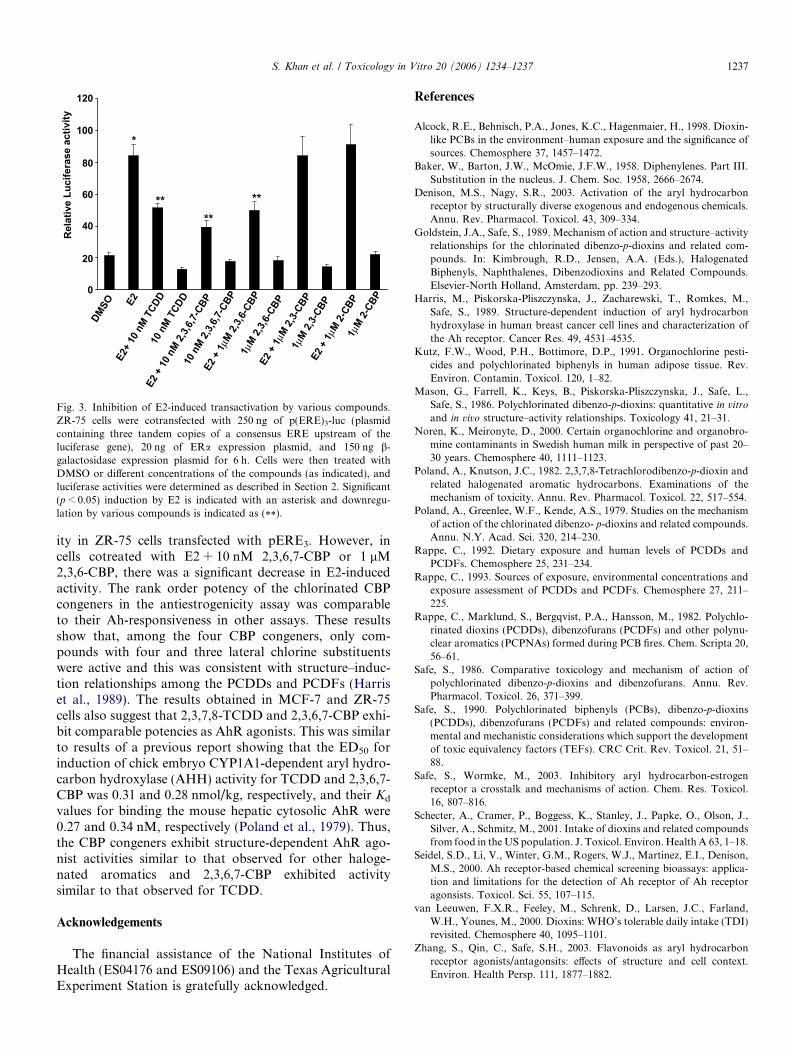
TCDD and related compounds also exhibit antiestro-genic activity in the rodent uterus and mammary tumorsand in breast cancer cell lines (Safe and Wormke, 2003).Results in Fig. 3 show that E2 induces luciferase activityin ZR-75 breast cancer cells transfected with an E2-respon-sive construct (pERE3) containing three tandem estrogenresponsive elements (EREs) (Zhang et al., 2003). TCDDalone did not induce activity; however, in cells cotreatedwith E2 + TCDD, there was a significant decrease inhormone-induced activity. 2,3,6,7-CBP (10 nM) and 1 lM2,3,6-, 2,3- and 2-CBP alone did not induce luciferase activ-
0
20
40
60
80
100
120
Rel
ativ
e Lu
cife
rase
act
ivity
E2 +
10 n
M2,3
,6,7-
CBP
10 n
M2,3
,6,7-
CBP
E2 +
1μM 2,
3,6-C
BP1μ
M 2,3,6
-CBP
E2 +
1μM 2,
3-CB
P1μ
M 2,3-
CBP
1μM 2-
CBP
E2 +
1μM 2-
CBP
DMSO E2
E2+
10 n
MTC
DD10
nM
TCDD
*
****
**
Fig. 3. Inhibition of E2-induced transactivation by various compounds.ZR-75 cells were cotransfected with 250 ng of p(ERE)3-luc (plasmidcontaining three tandem copies of a consensus ERE upstream of theluciferase gene), 20 ng of ERa expression plasmid, and 150 ng b-galactosidase expression plasmid for 6 h. Cells were then treated withDMSO or different concentrations of the compounds (as indicated), andluciferase activities were determined as described in Section 2. Significant(p < 0.05) induction by E2 is indicated with an asterisk and downregu-lation by various compounds is indicated as (**).
S. Khan et al. / Toxicology in Vitro 20 (2006) 1234–1237 1237
ity in ZR-75 cells transfected with pERE3. However, incells cotreated with E2 + 10 nM 2,3,6,7-CBP or 1 lM2,3,6-CBP, there was a significant decrease in E2-inducedactivity. The rank order potency of the chlorinated CBPcongeners in the antiestrogenicity assay was comparableto their Ah-responsiveness in other assays. These resultsshow that, among the four CBP congeners, only com-pounds with four and three lateral chlorine substituentswere active and this was consistent with structure–induc-tion relationships among the PCDDs and PCDFs (Harriset al., 1989). The results obtained in MCF-7 and ZR-75cells also suggest that 2,3,7,8-TCDD and 2,3,6,7-CBP exhi-bit comparable potencies as AhR agonists. This was similarto results of a previous report showing that the ED50 forinduction of chick embryo CYP1A1-dependent aryl hydro-carbon hydroxylase (AHH) activity for TCDD and 2,3,6,7-CBP was 0.31 and 0.28 nmol/kg, respectively, and their Kd
values for binding the mouse hepatic cytosolic AhR were0.27 and 0.34 nM, respectively (Poland et al., 1979). Thus,the CBP congeners exhibit structure-dependent AhR ago-nist activities similar to that observed for other haloge-nated aromatics and 2,3,6,7-CBP exhibited activitysimilar to that observed for TCDD.
Acknowledgements
The financial assistance of the National Institutes ofHealth (ES04176 and ES09106) and the Texas AgriculturalExperiment Station is gratefully acknowledged.
References
Alcock, R.E., Behnisch, P.A., Jones, K.C., Hagenmaier, H., 1998. Dioxin-like PCBs in the environment–human exposure and the significance ofsources. Chemosphere 37, 1457–1472.
Baker, W., Barton, J.W., McOmie, J.F.W., 1958. Diphenylenes. Part III.Substitution in the nucleus. J. Chem. Soc. 1958, 2666–2674.
Denison, M.S., Nagy, S.R., 2003. Activation of the aryl hydrocarbonreceptor by structurally diverse exogenous and endogenous chemicals.Annu. Rev. Pharmacol. Toxicol. 43, 309–334.
Goldstein, J.A., Safe, S., 1989. Mechanism of action and structure–activityrelationships for the chlorinated dibenzo-p-dioxins and related com-pounds. In: Kimbrough, R.D., Jensen, A.A. (Eds.), HalogenatedBiphenyls, Naphthalenes, Dibenzodioxins and Related Compounds.Elsevier-North Holland, Amsterdam, pp. 239–293.
Harris, M., Piskorska-Pliszczynska, J., Zacharewski, T., Romkes, M.,Safe, S., 1989. Structure-dependent induction of aryl hydrocarbonhydroxylase in human breast cancer cell lines and characterization ofthe Ah receptor. Cancer Res. 49, 4531–4535.
Kutz, F.W., Wood, P.H., Bottimore, D.P., 1991. Organochlorine pesti-cides and polychlorinated biphenyls in human adipose tissue. Rev.Environ. Contamin. Toxicol. 120, 1–82.
Mason, G., Farrell, K., Keys, B., Piskorska-Pliszczynska, J., Safe, L.,Safe, S., 1986. Polychlorinated dibenzo-p-dioxins: quantitative in vitro
and in vivo structure–activity relationships. Toxicology 41, 21–31.Noren, K., Meironyte, D., 2000. Certain organochlorine and organobro-
mine contaminants in Swedish human milk in perspective of past 20–30 years. Chemosphere 40, 1111–1123.
Poland, A., Knutson, J.C., 1982. 2,3,7,8-Tetrachlorodibenzo-p-dioxin andrelated halogenated aromatic hydrocarbons. Examinations of themechanism of toxicity. Annu. Rev. Pharmacol. Toxicol. 22, 517–554.
Poland, A., Greenlee, W.F., Kende, A.S., 1979. Studies on the mechanismof action of the chlorinated dibenzo- p-dioxins and related compounds.Annu. N.Y. Acad. Sci. 320, 214–230.
Rappe, C., 1992. Dietary exposure and human levels of PCDDs andPCDFs. Chemosphere 25, 231–234.
Rappe, C., 1993. Sources of exposure, environmental concentrations andexposure assessment of PCDDs and PCDFs. Chemosphere 27, 211–225.
Rappe, C., Marklund, S., Bergqvist, P.A., Hansson, M., 1982. Polychlo-rinated dioxins (PCDDs), dibenzofurans (PCDFs) and other polynu-clear aromatics (PCPNAs) formed during PCB fires. Chem. Scripta 20,56–61.
Safe, S., 1986. Comparative toxicology and mechanism of action ofpolychlorinated dibenzo-p-dioxins and dibenzofurans. Annu. Rev.Pharmacol. Toxicol. 26, 371–399.
Safe, S., 1990. Polychlorinated biphenyls (PCBs), dibenzo-p-dioxins(PCDDs), dibenzofurans (PCDFs) and related compounds: environ-mental and mechanistic considerations which support the developmentof toxic equivalency factors (TEFs). CRC Crit. Rev. Toxicol. 21, 51–88.
Safe, S., Wormke, M., 2003. Inhibitory aryl hydrocarbon-estrogenreceptor a crosstalk and mechanisms of action. Chem. Res. Toxicol.16, 807–816.
Schecter, A., Cramer, P., Boggess, K., Stanley, J., Papke, O., Olson, J.,Silver, A., Schmitz, M., 2001. Intake of dioxins and related compoundsfrom food in the US population. J. Toxicol. Environ. Health A 63, 1–18.
Seidel, S.D., Li, V., Winter, G.M., Rogers, W.J., Martinez, E.I., Denison,M.S., 2000. Ah receptor-based chemical screening bioassays: applica-tion and limitations for the detection of Ah receptor of Ah receptoragonsists. Toxicol. Sci. 55, 107–115.
van Leeuwen, F.X.R., Feeley, M., Schrenk, D., Larsen, J.C., Farland,W.H., Younes, M., 2000. Dioxins: WHO’s tolerable daily intake (TDI)revisited. Chemosphere 40, 1095–1101.
Zhang, S., Qin, C., Safe, S.H., 2003. Flavonoids as aryl hydrocarbonreceptor agonists/antagonsits: effects of structure and cell context.Environ. Health Persp. 111, 1877–1882.


















