
Strong Anticancer Potential of Nano-triterpenoid from Phytolacca decandra against A549...
11
- RESEARCH ARTICLE - Strong Anticancer Potential of Nano-triterpenoid from Phytolacca decandra against A549 Adenocarcinoma via a Ca 2D -dependent Mitochondrial Apoptotic Pathway Q14 Jayeeta Das, Sreemanti Das, Avijit Paul, Asmita Samadder, Anisur Rahman Khuda-Bukhsh* Cytogenetics and Molecular Biology Laboratory, Department of Zoology, University of Kalyani, Kalyani 741235, India Available online --- Received: Mar 14, 2013 Revised: Jul 17, 2013 Accepted: Jul 23, 2013 KEYWORDS A549 lung adenoma cells; apoptosis; betulinic acid; intracellular calcium content; Phytolacca decandra; PLGA nanoparticles Abstract We isolated a triterpenoid from an ethanolic extract of Phytolacca decandra and na- noencapsulated it with biodegradable nontoxic polymers of poly(lactide-co-glycolide) to examine if the nanoform of this hitherto unexplored betulinic-acid derivative (NdBA) could produce a stronger anticancer effect by rendering better drug bioavailability and targeted delivery than the nonencapsulated betulinic-acid derivative (dBA). The nano- particles were characterized with the help of physicochemical and morphological studies involving dynamic light scattering and atomic force microscopy. A549 cancer cells were exposed to NdBA and dBA at the IC 50 doses of 50 mg/mL and 100 mg/mL, respectively. Mitochondrial dysfunction-mediated apoptosis was determined by exam- ining the changes in the intracellular calcium content, the reactive oxygen species accumulation, the cytochrome c release, the upregulation of Bcl-2-associated-X protein (Bax) and caspase 3, the downregulation of B cell lymphoma 2, and the mitochondrial membrane potential (DJ m ) depolarization. Apoptosis was also verified by acridine or- ange staining observed under fluorescence microscopy and annexin V-fluorescein iso- thiocyanate/propidium iodide staining through flow cytometric studies. The levels of intracellular adenosine triphosphate/adenosine diphosphate ratio decreased, and the * Corresponding author. Cytogenetics and Molecular Biology Laboratory, Department of Zoology, University of Kalyani, Kalyani 741235, India. Q2 E-mail: [email protected], [email protected] (A.R. Khuda-Bukhsh). + MODEL 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69 70 71 72 73 74 75 76 77 78 79 80 81 82 83 84 85 86 87 88 89 90 91 92 93 94 95 96 97 98 99 100 101 102 103 104 105 106 107 108 109 110 111 112 113 114 115 116 117 118 119 120 121 122 123 124 JAMS135_proof ■ 19 August 2013 ■ 1/11 Please cite this article in press as: Das J, et al., Strong Anticancer Potential of Nano-triterpenoid from Phytolacca decandra against A549 Adenocarcinoma via a Ca 2þ -dependent Mitochondrial Apoptotic Pathway, Journal of Acupuncture and Meridian Studies (2013), http:// dx.doi.org/10.1016/j.jams.2013.07.009 Copyright ª 2013, International Pharmacopuncture Institute pISSN 2005-2901 eISSN 2093-8152 http://dx.doi.org/10.1016/j.jams.2013.07.009 Available online at www.sciencedirect.com Journal of Acupuncture and Meridian Studies journal homepage: www.jams-kpi.com J Acupunct Meridian Stud 2013;--(-):--e--
-
Upload
anisur-rahman -
Category
Documents
-
view
212 -
download
0
Transcript of Strong Anticancer Potential of Nano-triterpenoid from Phytolacca decandra against A549...

Q14
+ MODEL
123456789
1011121314151617181920212223242526272829303132333435363738394041424344454647484950515253545556575859606162
63646566676869
JAMS135_proof ■ 19 August 2013 ■ 1/11
Available online at www.sciencedirect.com
Journal of Acupuncture and Meridian Studies
journa l homepage: www. jams-kp i .com
J Acupunct Meridian Stud 2013;--(-):--e--
70717273
- RESEARCH ART ICLE - 747576777879808182838485Strong Anticancer Potential of Nano-triterpenoidfrom Phytolacca decandra against A549Adenocarcinoma via a Ca2D-dependent MitochondrialApoptotic Pathway
8687888990
Jayeeta Das, Sreemanti Das, Avijit Paul, Asmita Samadder,Anisur Rahman Khuda-Bukhsh*
91929394959697
Cytogenetics and Molecular Biology Laboratory, Department of Zoology, University of Kalyani, Kalyani 741235,India
Available online - - -
9899
100101102103104105
Received: Mar 14, 2013Revised: Jul 17, 2013Accepted: Jul 23, 2013
KEYWORDSA549 lung adenoma
*
106107108109110111112
PAd
CopIht
cells;apoptosis;betulinic acid;intracellular calcium
content;Phytolacca decandra;PLGA nanoparticles
Corresponding author. CytogeneticIndia.E-mail: [email protected], k
113114115
lease cite this article in press as: Dadenocarcinoma via a Ca2þ-dependex.doi.org/10.1016/j.jams.2013.07.0
pyright ª 2013, International PharmSSN 2005-2901 eISSN 2093-8152tp://dx.doi.org/10.1016/j.jams.201
AbstractWe isolated a triterpenoid from an ethanolic extract of Phytolacca decandra and na-noencapsulated it with biodegradable nontoxic polymers of poly(lactide-co-glycolide)to examine if the nanoform of this hitherto unexplored betulinic-acid derivative (NdBA)could produce a stronger anticancer effect by rendering better drug bioavailability andtargeted delivery than the nonencapsulated betulinic-acid derivative (dBA). The nano-particles were characterized with the help of physicochemical and morphologicalstudies involving dynamic light scattering and atomic force microscopy. A549 cancercells were exposed to NdBA and dBA at the IC50 doses of 50 mg/mL and 100 mg/mL,respectively. Mitochondrial dysfunction-mediated apoptosis was determined by exam-ining the changes in the intracellular calcium content, the reactive oxygen speciesaccumulation, the cytochrome c release, the upregulation of Bcl-2-associated-X protein(Bax) and caspase 3, the downregulation of B cell lymphoma 2, and the mitochondrialmembrane potential (DJm) depolarization. Apoptosis was also verified by acridine or-ange staining observed under fluorescence microscopy and annexin V-fluorescein iso-thiocyanate/propidium iodide staining through flow cytometric studies. The levels ofintracellular adenosine triphosphate/adenosine diphosphate ratio decreased, and the
s and Molecular Biology Laboratory, Department of Zoology, University of Kalyani, Kalyani 741235,
Q2
[email protected] (A.R. Khuda-Bukhsh).
116117118119120121122123124
s J, et al., Strong Anticancer Potential of Nano-triterpenoid from Phytolacca decandra against A549nt Mitochondrial Apoptotic Pathway, Journal of Acupuncture and Meridian Studies (2013), http://09
acopuncture Institute
3.07.009

2 J. Das et al.
+ MODEL
125126127128129130131132133134135136137138139140141142143144145146147148149150151152153154155156157158159160161162163164165166167168169170171172173174175176177178179180181182183184185186
187
JAMS135_proof ■ 19 August 2013 ■ 2/11
188189190191192
Please cite this article in press as: DaAdenocarcinoma via a Ca2þ-dependedx.doi.org/10.1016/j.jams.2013.07.0
ATPase activity increased more strikingly in A549 cells exposed to NdBA than in A549cells exposed to dBA. Overall results showed that both drugs directly target the mito-chondrial oxidative phosphorylation system, with NdBA having a stronger effect, indi-cating NdBA to be a better candidate for the development of an anticancer drug foruse against lung adenocarcinomas.
193
194 1951961971981992002012022032042052062072082092102112122132142152162172182192202212222232242252262272282292302312322332342352362372382392402412422432442452462472481. Introduction
Cancer is one of the most dreaded diseases worldwide.Lung cancer is a common cancer type, but still the pa-tients continue to have a poor prognosis. As cancer cellsare endowed with the property of uncontrolled prolifer-ation and growth, search for agents that can triggerapoptosis or programmed cell death in tumor cells hasbecome a major goal in anticancer drug discovery [1]. Inresponse to apoptotic stimuli triggered by anticancerdrugs, several proteins (such as cytochrome c) arereleased from the mitochondrial intermembrane spaceinto the cytoplasm that mediates the activation of cas-pase cascades, which in turn ultimately promotesapoptosis. Calcium is a key regulator of cell survivalbecause cytosolic free calcium concentration is a majorregulatory factor for large number of cellular processessuch as muscle contraction, secretion, cell differentia-tion, and apoptosis. However, sustained elevation ofintracellular calcium plays a role in cell death as well [2].The proapoptotic effects of calcium are mediated by adiverse range of calcium-sensitive factors that are com-partmentalized in various intracellular organelles,including endoplasmic reticulum (ER) and mitochondria[3]. Excessive calcium load to the mitochondria mayinduce apoptosis by impairing mitochondrial function, forexample, generation of reactive oxygen species (ROS) bydamaged respiratory chain [4]. Calcium accumulation hasalso been reported to stimulate mitochondrial ROS pro-duction in cells [5,6]. Mitochondrial calcium uptake alsotriggers significant membrane depolarization, resulting ina decrease in the mitochondrial membrane potential [3].
Many plant-derived compounds are known to havecurative potentials against cancer [7e9]. The World HealthOrganization estimated that approximately 80% of world’sinhabitants rely mostly on traditional medicines for theirprimary healthcare [10]. The root extract of Phytolaccadecandra, a plant commonly known as “Chui-Xu Shang-Lu”in China, has been used for centuries as a traditional folkmedicine for the treatment of tumors, edema, bronchitis,and abscesses [11]. Recently, we studied the anticancerpotentials of the ethanolic root extract of P decandra andthe nanoencapsulated form of the whole extract [8]. Yet,no work had earlier been carried out on any of the majoringredients contained in the crude ethanolic extract of PDor the nanoencapsulated form of any such individualingredient. However, the extract of a congeneric species,Phytolacca americana, has been reported to contain tri-terpene saponins (phytolacca saponin), phytolaccagenicacid, phytolaccagenin, etc. [12]. We isolated one ofthe major ingredients of PD, namely, 1-isopropenyl-5a,5b,8,8,11a-pentamethyl-1,2,3,4,5,5a,6,7,7a,8,11,11a,
s J, et al., Strong Anticancer Potent Mitochondrial Apoptotic Pathw09
11b,12,13,13a,13b-octadecahydro cyclopenta[a]chrysene-3a-carboxylic acid, which is presumably a derivative ofbetulinic acid (dBA) and which had not been tested for itsanticancer potential. We therefore became interested tostudy the possible anticancer efficacy of dBA and alsowanted to ascertain if the poly(lactide-co-glycolide) (PLGA)nanoencapsulation of dBA (NdBA) could accentuate itsanticancer potential.
Therefore, the aims of the present study were todetermine: (1) if both dBA and NdBA induced apoptosis inhuman lung adenocarcinoma (A549) cells in vitro, whichoften defy any such chemotherapeutic venture; (2) if theactive ingredient and its nanoencapsulated form showedany apoptotic ability, and whether NdBA had a strongereffect than dBA; (3) if the apoptotic phenomenon wasassociated with change in calcium content; and finally, (4)if both dBA and NdBA modulated the intracellular calciumcontent, to ascertain the possible role of Ca 2þ ion in themitochondrial apoptotic pathway.
2. Materials and methods
2.1. Reagents
All chemicals and reagents used in this study were pur-chased from Sigma Chemical Co. (St. Louis, MO, USA).
2.2. Cell culture
Human nonsmall cell lung carcinoma A549 cells were pro-cured from the National Center for Cell Science (Pune,India). The cells were cultured at 5 � 105 cells/mL in Dul-becco’s modified Eagle’s medium supplemented with 10%fetal bovine serum and 1% PSN antibiotic at 37 �C, 5% CO2
for our experimental purpose.
2.3. Source of P decandra and identification of itsmajor bioactive component
The ethanolic root extract of P decandra used as homeo-pathic mother tincture was manufactured and supplied byBoiron Laboratories (Lyon, France). The ethanolic extractwas evaporated at 60 �C (to remove alcohol from theextract). The yield of the extract was about 52%; it wasisolated using the standard column chromatographymethod over silica gel using a mixture of chloroform andhexane (9:1) and was sent for mass spectroscopy, Fouriertransform infrared, and nuclear magnetic resonance ana-lyses [8].
ntial of Nano-triterpenoid from Phytolacca decandra against A549ay, Journal of Acupuncture and Meridian Studies (2013), http://

Q3
Q4
Q5
Q6
Q7
Q1 Anticancer potential of Phytolacca decandra 3
+ MODEL
249250251252253254255256257258259260261262263264265266267268269270271272273274275276277278279280281282283284285286287288289290291292293294295296297298299300301302303304305306307308309310
311312313314315316317318319320321322323324325326327328329330331332333334335336337338339340341342343344345346347348349350351352353354355356357358359360361362363364365366367368369370371372
JAMS135_proof ■ 19 August 2013 ■ 3/11
2.4. Preparation of blank nanoparticles and dBA-loaded nanoparticles
For the PLGAencapsulation of dBA, the solvent displacementtechnique [13] was deployed under optimal conditions,which has been described elsewhere [8]. Following the samemethod, blank nanoparticles were prepared without theaddition of dBA, and tested for its possible apoptotic effecton A549. These blank nanomaterials failed to demonstrateany apoptotic effect on A549, for which only dBA-loadednanoparticles were considered for further studies.
2.5. Atomic force microscopic studies of NdBA
Samples for atomic force microscopic (AFM) imaging wereprepared by dropping the NdBA suspension on a freshlycleaned mica sheet and allowing it to dry in air. Theobservation was recorded through AFM (Veeco di CP-11)imaging in amplitude and tapping modes.
2.6. Particle size determination by dynamic lightscattering method
Determination of the average size and distribution of thepolymeric micelles were conducted using a dynamic lightscattering (DLS) machine (Malvern Instrument, South-borough, UK) [8].
2.7. Cell proliferation assay
The viability of A549 cells was assessed by the MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide)assay [14] after exposure to either dBA or NdBA. Cells wereseeded in 96-well plates at a density of 106 cells/mL andwere incubated with different concentrations of the drug(25 mg/mL, 50 mg/mL, 75 mg/mL, and 100 mg/mL) for 24hours. The intracellular formazan crystals formed weresolubilized with dimethyl sulfoxide, and the absorbance ofthe solution was measured at 595 nm by using a microplatereader (Thermo-Scientific, Multiskan ELISA, USA). The cellsurvival rate was calculated as percentage of MTT inhibitionby adopting the following formula: percentage ofsurvival Z (mean experimental absorbance/mean controlabsorbance) � 100%.
2.8. Surface morphological studies of A549 cells byAFM
Control A549 cells and drug treated cells were first fixed ina mixture of 2% paraformaldehyde and 2.5% glutaraldehydein 0.1 M phosphate buffer for 8 hours at 4 �C, then dehy-drated in increasing grades of ethanol (50e100%), andfinally dried and photographed by AFM (Veeco di CP-11).
2.9. Measurement of intracellular Ca2D content
Changes in [Ca2þ]i were assessed using the Ca2þ-sensitivefluorescent indicator dye Fura 2-AM (Santa Cruz). The A549cells were washed with calcium free buffer [120 mM NaCl,5 mM KCl, 1 mM MgSO4, 0.96 mM NaH2PO4, 0.2% glucose,
Please cite this article in press as: Das J, et al., Strong Anticancer PoteAdenocarcinoma via a Ca2þ-dependent Mitochondrial Apoptotic Pathwdx.doi.org/10.1016/j.jams.2013.07.009
0.1% bovine serum albumin (BSA), 20 mM HEPES, pH Z 7.2]and loaded with 6mM of Fura 2-AM at 37 �C for 30 minutes.Cells were divided into two parts: one was analyzed in afluorimeter (PerkinElmer, USA) and the other part wasexamined under a fluorescent microscope (Leica,Germany).
2.10. Determination of mitochondrial membranepotential (Jm)
Changes in the mitochondrial membrane potential of con-trol A549 cells and drug-treated groups were determinedusing a fluorescent probe, Rhodamine 123, according to thestandard protocol [15]. Jm was assessed using flowcytometry (FACS caliber, BD Bioscience) and fluorescencemicroscopy (Leica).
2.11. Assessment of mitochondrial adenosinetriphosphate/adenosine diphosphate ratio andATPase activity
The ratio of adenosine triphosphate/adenosine diphos-phate (ATP/ADP) was measured using the Enzylight ADP/ATP ratio bioluminescence assay kit (ELDT 100Q9) andassessed in a luminometer (Thermo-Scientific Varier Skan)[16]. ATPase enzyme activity was determined by the stan-dard procedure [17] using phenol red indicator.
2.12. Microscopic analysis of intracellular ROS
Mitochondrial ROS was detected using the fluorescentprobe 20,70-dichlorofluorescein diacetate (DCFH-DA). DCFH-DA is cell-permeable and is cleaved by nonspecific ester-ases and oxidized by peroxides produced in the cells toform fluorescent 20,70-dichlorofluorescin (DCF). The in-tensity of DCF fluorescence is proportional to the amount ofperoxide produced in the cells. In brief, cells of differenttreatment groups were incubated for 30 minutes withDCFH-DA (5 mg/mL) at 37 �C in the dark [18] and examinedwith a fluorescence microscope (Leica, Germany).
2.13. Acridine Orange staining of apoptosis
Acridine Orange is a metachromatic dye that differentiallystains nucleic acids. Chromatin condensation is an earlyevent of apoptosis, and the condensed chromatin is muchmore sensitive to DNA denaturation than normal chromatin.The extent of apoptosis in drug A549 cells was determinedby acridine orange staining (1 mg/mL) preincubated withRNase A (1 mg/mL), and the extent of fluorescence wasexamined with a fluorescence microscope (Leica) [19].
2.14. Estimation of apoptotic% by flow cytometry
In order to evaluate apoptosis and necrosis, externalizationof phosphatidylserine during apoptosis and leakage fromnecrotic cells was observed by annexin V-FITC/propidiumiodide (PI) dual staining using the standard protocol [19].Each determination was based on a mean fluorescence in-tensity of 10,000 events.
ntial of Nano-triterpenoid from Phytolacca decandra against A549ay, Journal of Acupuncture and Meridian Studies (2013), http://

Q8
Q9
Q10
11
4 J. Das et al.
+ MODEL
373374375376377378379380381382383384385386387388389390391392393394395396397398399400401402403404405406407408409410411412413414415416417418419420421422423424425426427428429430431432433434
435436437438439440441442443444445446447448449450451452453454455456457458459460461462463464465466467468469470471472473474475476477478479480481482483484485486487488489490491492493494495496
JAMS135_proof ■ 19 August 2013 ■ 4/11
2.15. Immunoblot
2.15.1. Preparation of cytosolic fractionControl and drug-treated A549 cells were collected,washed with ice-cold phosphate-buffered saline, resus-pended in buffer A [10 mmol/L TriseMOPS pH 7.4,200 mmol/L sucrose, 1 mmol/L ethylene glycol tetraaceticacid (EGTA), 1 mg/mL BSA, 1 mmol/L dithiothreitol], andlysed by homogenization. The suspension was centrifugedat 1000 � g for 10 minutes at 4 �C. The resulting superna-tant was centrifuged at 10,000 � g for 15 minutes to pelletmitochondria. Cytosolic fraction (supernatant) was ob-tained by centrifugation at 12,000 � g for 1 hour at 4 �C.
2.15.2. Isolation of mitochondria and preparation ofmitochondrial fractionMitochondria were isolated by harvesting A549 cells (ofdifferent groups) with trypsin/EDTA, centrifuged at 300 � gfor 5 minutes, washed in phosphate-buffered saline, andresuspended in buffer A (10 mmol/L TriseMOPS pH 7.4,200 mmol/L sucrose, 1 mmol/L EGTA, 1 mg/mL BSA,1 mmol/L dithiothreitol). Cells were lysed with a homoge-nizer (40 strokes), and the suspension was centrifuged at2500 � g for 10 minutes at 4 �C. The supernatant was thencentrifuged at 10,000 � g for 10 minutes at 4 �C. Theresulting pellet was washed and resuspended in buffer B(10 mmol/L TriseMOPS pH 7.4, 250 mmol/L sucrose,1 mmol/L EGTA, 1 mg/mL BSA and 1 mmol/L phenyl-methanesulfonyl fluoride).
The immunoblot analysis of both cytosolic and mito-chondrial fractionswas conducted by using anticytochrome cand caspase3 antibodies (Santa Cruz Biotechnology). Alka-line phosphatase-conjugated secondary antibodies (SigmaChemical Co.) [20] were used for this purpose. For quanti-tative analysis of each band, we determined the band in-tensity using the Total Lab software (Ultra Lum, USA).
2.16. RNA extraction and quantitative reversetranscriptase-polymerase chain reaction analysis
Total RNA from A549 cells was extracted using Trizol re-agent according to the manufacturer’s instructions. cDNAwas reverse transcribed by reverse transcriptase and cDNAwas used as a template for polymerase chain reaction (PCR)amplification with the aid of Taq polymerase as per thestandard practice [21]. Synthetic oligonucleotide primersfor reverse transcription (RT)-PCR were procured fromBioserve Biotech (Hyderabad, India). The sequences of theforward and reverse primers used for specific amplificationsare as follows: G3PDH (50-ATGGGGAAGGTGAA-GGTCGG-30
and 50-GGATGCTAAGCAGTTGGT-30); Bcl2 (50-TACCGTCGT-GACTTCGCAGAG-30 and 50-GGCAGGCTGAGCAGGGTCTT-30);Bax (50-CGGCGAATTGGAGATGAACTG-30 and 50-GCAAAGTA-GAAGAGGGCAACC-30).
2.17. In vivo assessment of relative absorption ofdBA and NdBA in various tissues
To determine the relative absorption potential of NdBA anddBA, in various tissues, both drugs were orally administeredto mice for 7 days at a dose of 10 mg/kg body weight. At the
Please cite this article in press as: Das J, et al., Strong Anticancer PoteAdenocarcinoma via a Ca2þ-dependent Mitochondrial Apoptotic Pathwdx.doi.org/10.1016/j.jams.2013.07.009
end of the treatment period, the mice were sacrificed, andbrain, lung, liver, and kidney tissues were dissected out,homogenized in 2 mL ice-cold phosphate-buffered saline,and diluted 100 times. The resulting diluted homogenateswere analyzed in a Spectrophotometer (Shimadzu, UVevi-sible Spectrophotometer) at a wavelength of 479 nm (lmax).Data were normalized with negative control (tissue fromsaline-treated mice only).
3. Results
3.1. Mass spectroscopic studies of ethanolic extractof Phytolacca decandra
The mass spectra value of the component with 80% occur-rence was m/z Z 438.35 (Mþ), suggesting a triterpenoidgroup, presumably a derivative of betulinic acid with themolecular formula C30H46O2 (Fig. 1A), which was used asdBA.
3.2. Characterization of nanoparticles of NdBA
The surface morphology of NdBA was spherical in shape,with a smooth surface, but without any pinholes or cracks.Fig. 1B and C show two- and three-dimensional images,respectively, by atomic force microscopy. The meandiameter of nanoparticles was found to be 110 � 0.5 nm,with a PDI Qof 0.254 and a zeta potential of e17.6 � 0.24 mV.DLS data corroborated well with the AFM result (Fig. 1D).The two-dimensional fast Fourier transformation image(Fig. 1E) in AFM also showed the uniform spatial frequencyof the topographic planar signal of NdBA.
3.3. Cytotoxicity assay
The cell viability of A549 cells was examined after treat-ment of dBA and NdBA at 24 hours. A dose-dependentcytotoxicity was obtained. Although no apparent cytotoxiceffect on cell viability was observed at lower doses, the IC50
value of both dBA and NdBA was found to be 50 mg/mL(Fig. 2A).
3.4. Morphology of A549 cells
A549 cells exposed to dBA and NdBA at doses of 50 mg/mLand 100 mg/mL, respectively, showed at different fre-quencies typical characteristics of apoptosis such as cellblebbing, cytoplasmic shrinkage and detachment from eachother, floating in the medium, distortion, and nuclearcondensation (Fig. 2B).
3.5. Intracellular [Ca2D] content
To verify whether a calcium signal was involved in theapoptosis pathway, the Ca2þ-sensitive dye Fura 2-AM wasused to investigate whether exposure to dBA and NdBAwould induce [Ca2þ]i in A549 cells. The cells displayed anincrease in fluorescence intensity after exposure to 50 mg/mL and 100 mg/mL of both dBA and NdBA in comparison tothe control (Fig. 2C and D).
ntial of Nano-triterpenoid from Phytolacca decandra against A549ay, Journal of Acupuncture and Meridian Studies (2013), http://
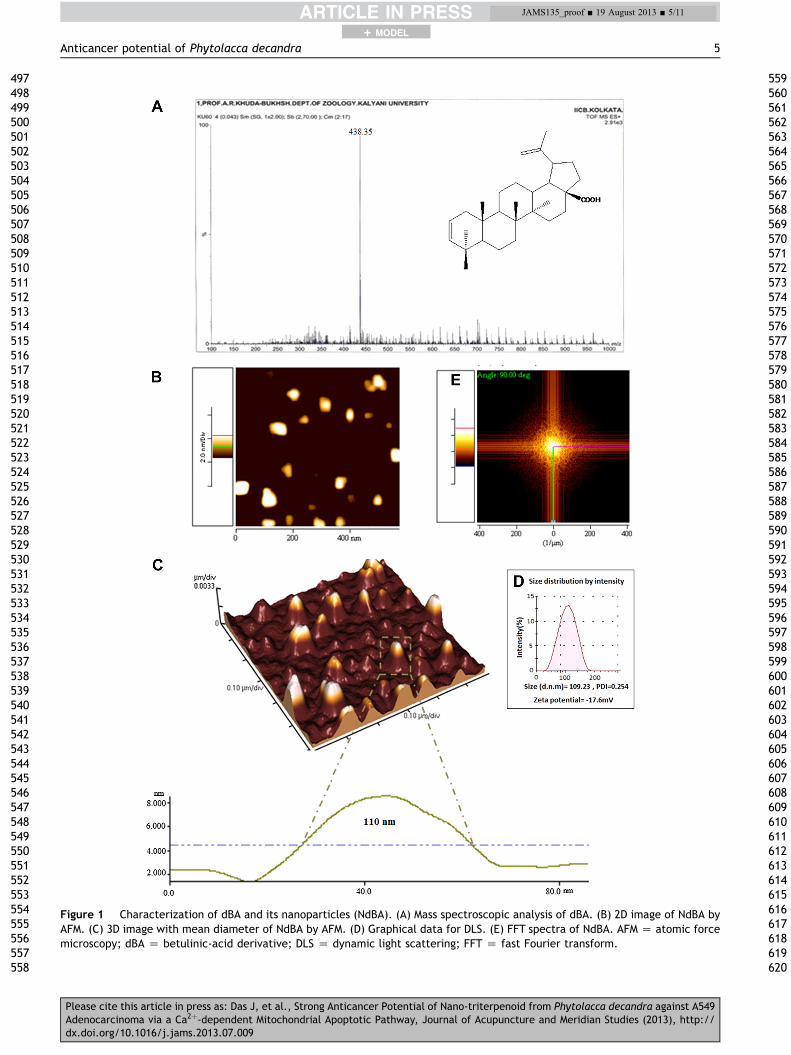
Figure 1 Characterization of dBA and its nanoparticles (NdBA). (A) Mass spectroscopic analysis of dBA. (B) 2D image of NdBA byAFM. (C) 3D image with mean diameter of NdBA by AFM. (D) Graphical data for DLS. (E) FFT spectra of NdBA. AFM Z atomic forcemicroscopy; dBA Z betulinic-acid derivative; DLS Z dynamic light scattering; FFT Z fast Fourier transform.
Anticancer potential of Phytolacca decandra 5
+ MODEL
497498499500501502503504505506507508509510511512513514515516517518519520521522523524525526527528529530531532533534535536537538539540541542543544545546547548549550551552553554555556557558
559560561562563564565566567568569570571572573574575576577578579580581582583584585586587588589590591592593594595596597598599600601602603604605606607608609610611612613614615616617618619620
JAMS135_proof ■ 19 August 2013 ■ 5/11
Please cite this article in press as: Das J, et al., Strong Anticancer Potential of Nano-triterpenoid from Phytolacca decandra against A549Adenocarcinoma via a Ca2þ-dependent Mitochondrial Apoptotic Pathway, Journal of Acupuncture and Meridian Studies (2013), http://dx.doi.org/10.1016/j.jams.2013.07.009

Figure 2 Assessment of cell viability and measurement of [Ca2þ]i content. (A) Graphical representation of MTT assay todetermine IC50 value of dBA and NdBA. (B) Morphological studies in A549 cells by AFM after administration of dBA and NdBA after 24hours of treatment. (C) Microscopic study of Fura 2-AM staining. (D) Fluorometric assessment of A549 cells. (a) Control A549; (b)50 mg/mL dBA; (c) 100 mg/mL dBA; (d) 50 mg/mL NdBA; (e) 100 mg/mL NdBA. AFM Z atomic force microscopy; dBA Z betulinic-acidderivative; MTT Z 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide; NdBA Z nanoform of dBA.
6 J. Das et al.
+ MODEL
621622623624625626627628629630631632633634635636637638639640641642643644645646647648649650651652653654655656657658659660661662663664665666667668669670671672673674675676677678679680681682
683684685686687688689690691692693694695696697698699700701702703704705706707708709710711712713714715716717718719720721722723724725726727728729730731732733734735736737738739740741742743744
JAMS135_proof ■ 19 August 2013 ■ 6/11
Please cite this article in press as: Das J, et al., Strong Anticancer Potential of Nano-triterpenoid from Phytolacca decandra against A549Adenocarcinoma via a Ca2þ-dependent Mitochondrial Apoptotic Pathway, Journal of Acupuncture and Meridian Studies (2013), http://dx.doi.org/10.1016/j.jams.2013.07.009

Figure 3 Fluorescence microscopic and flow cytometric analyses of mitochondrial membrane potential (DJm), ATP/ADP ratio,ATPase activity and generation of ROS. (A) Microscopic images of A549 cells for (DJm). (B) Flow cytometric analyses of (DJm). (C)Graphical representation showing measurement of ATP/ADP ratio. (D) Graphical representation showing ATPase activity. (E)Microscopic image of generation of ROS. (a) Control A549; (b) 50 mg/mL dBA; (c) 100 mg/mL dBA; (d) 50 mg/mL NdBA; (e) 100 mg/mLNdBA. ATP/ADP ratio Z adenosine triphosphate/adenosine diphosphate ratio; dBA Z betulinic-acid derivative; NdBA Z nanoformof dBA; ROS Z reactive oxygen species.
Anticancer potential of Phytolacca decandra 7
+ MODEL
745746747748749750751752753754755756757758759760761762763764765766767768769770771772773774775776777778779780781782783784785786787788789790791792793794795796797798799800801802803804805806
807808809810811812813814815816817818819820821822823824825826827828829830831832833834835836837838839840841842843844845846847848849850851852853854855856857858859860861862863864865866867868
JAMS135_proof ■ 19 August 2013 ■ 7/11
3.6. Change in mitochondrial membrane potential(DJm)
Compared with the control, dBA- and NdBA-treated cellsshowed decrease in Rhodamine123 fluorescence intensitywith the increase in dBA and NdBA concentrations after 24hours of incubation (Fig. 3A), with relatively more effectivemitochondrial membrane depolarization in NdBA-treatedA549 cells. A similar change was observed in dBA- and NdBA-treated cellswhenalso analyzedflowcytometrically (Fig. 3B).
3.7. ATP/ADP ratio and ATPse activity
The ATP/ADP ratio was found to have decreased in A549cells with an increasing dose of dBA- and NdBA-treated cells
Please cite this article in press as: Das J, et al., Strong Anticancer PoteAdenocarcinoma via a Ca2þ-dependent Mitochondrial Apoptotic Pathwdx.doi.org/10.1016/j.jams.2013.07.009
(Fig. 3C). Subsequently, a significant increase in the level ofATPase activity was observed in dBA- and NdBA-treatedA549 cells (Fig. 3D).
3.8. Generation of ROS
Generation of ROS was least in control A549 cells.Compared to control, a significantly greater quantity of ROSwas generated when cells were treated with dBA and NdBA(Fig. 3E).
3.9. Fluorescence study by acridine orange staining
The control A549 cells showed green fluorescence, indi-cating them to be viable, whereas dBA- and NdBA-treated
ntial of Nano-triterpenoid from Phytolacca decandra against A549ay, Journal of Acupuncture and Meridian Studies (2013), http://

8 J. Das et al.
+ MODEL
869870871872873874875876877878879880881882883884885886887888889890891892893894895896897898899900901902903904905906907908909910911912913914915916917918919920921922923924925926927928929930
931932933934935936937938939940941942943944945946
JAMS135_proof ■ 19 August 2013 ■ 8/11
cells (100 mg/mL) showed red fluorescence with condensedchromatin, indicating that they were undergoing apoptosis(Fig. 4A).
3.10. Estimation of apoptosis %
Data demonstrated that both dBA and NdBA stimulatedapoptosis (Annexin-positive cells) or necrosis (PI-positivecells) in a dose-dependent manner. dBA increased theproportion of early apoptotic cells from 0.5 � 0.42% (con-trol A549 group) to 12.61 � 2.87% and 25.28 � 5.38% in50 mg/mL and 100 mg/mL, respectively. A similar result wasalso obtained in NdBA-treated group, where the % of earlyapoptotic cells was increased to 26.81 � 3.69% and28.57 � 4.28% in 50 mg/mL and 100 mg/mL treatmentgroups, respectively (Fig. 4B, aee).
Figure 4 Apoptosis%, immunoblot and RT-PCR. (A) Microscopic imapoptosis. (C) Immunoblot of mitochondrial cyt C, cytosolic cyt CIn vivo assessment of effective absorption of dBA and NdBA in varidBA; (d) 50 mg/mL NdBA; (e) 100 mg/mL NdBA. Ln1 Z control A549;NdBA; Ln5 Z 100 mg/mL NdBA. ap < 0.05, bp < 0.01, cp < 0.001 veacid derivative; NdBA Z nanoform of dBA; RT-PCR Z reverse tran
Please cite this article in press as: Das J, et al., Strong Anticancer PoteAdenocarcinoma via a Ca2þ-dependent Mitochondrial Apoptotic Pathwdx.doi.org/10.1016/j.jams.2013.07.009
3.11. Immunoblot and RT-PCR studies
The expressions of protein were analyzed using the immu-noblot technique (Fig. 4C, aee) and mRNA expression levelsby RT-PCR (Fig. 4D, aee) in different treated cells.
The expression level of cytochrome c in the mito-chondrial fraction was found to be down-regulated withincreasing doses of dBA and NdBA, whereas the expres-sion level of cytochrome c in cytosolic fraction was foundto be up-regulated with increasing doses of dBA andNdBA.
The mRNA expression of Bcl-2-associated-X protein (Bax)was up-regulated in a dose-dependent manner, whereasthat of Bcl2 was down-regulated in dBA- and NdBA-treatedcells. The expression of caspase 3 was found to be up-regulated with increasing doses of dBA and NdBA.
age of acridine orange staining. (B) Flow cytometric analysis of, caspase 3 keeping b-actin as acting. (D) RT-PCR analysis. (E)ous tissues. (a) Control A549; (b) 50 mg/mL dBA; (c) 100 mg/mLLn2 Z 50 mg/mL dBA; Ln3 Z 100 mg/mL dBA; Ln4 Z 50 mg/mLrsus control A549 cells for statistical analysis. dBA Z betulinic-scription-polymerase chain reaction.
947948949950951952953954955956957958959960961962963964965966967968969970971972973974975976977978979980981982983984985986987988989990991992
ntial of Nano-triterpenoid from Phytolacca decandra against A549ay, Journal of Acupuncture and Meridian Studies (2013), http://
Anticancer potential of Phytolacca decandra 9
+ MODEL
993994995996997998999
1000100110021003100410051006100710081009101010111012101310141015101610171018101910201021102210231024102510261027102810291030103110321033103410351036103710381039104010411042104310441045104610471048104910501051105210531054
10551056105710581059106010611062
JAMS135_proof ■ 19 August 2013 ■ 9/11
3.12. Tissue distribution assay of PLGAnanoparticles
The data revealed that most of the particles were detectedin the liver (45.4% NdBA and 40.9% dBA), followed by kidney(16.6% NdBA and 10.98% dBA), lung (14.67% NdBA and 8.31%dBA), and brain (3.88% NdBA and 0% dBA) in all 7 days(Fig. 4E) of the study.
Figure 5 Schematic representation of se
Please cite this article in press as: Das J, et al., Strong Anticancer PoteAdenocarcinoma via a Ca2þ-dependent Mitochondrial Apoptotic Pathwdx.doi.org/10.1016/j.jams.2013.07.009
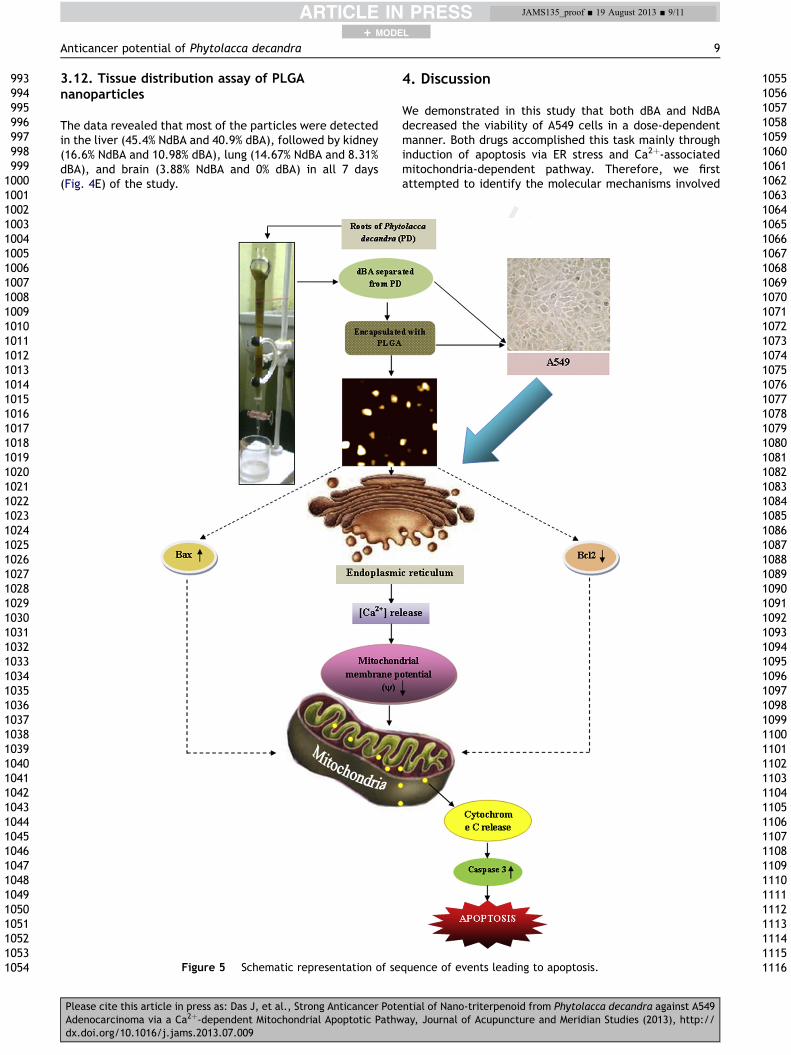
4. Discussion
We demonstrated in this study that both dBA and NdBAdecreased the viability of A549 cells in a dose-dependentmanner. Both drugs accomplished this task mainly throughinduction of apoptosis via ER stress and Ca2þ-associatedmitochondria-dependent pathway. Therefore, we firstattempted to identify the molecular mechanisms involved
quence of events leading to apoptosis.
106310641065106610671068106910701071107210731074107510761077107810791080108110821083108410851086108710881089109010911092109310941095109610971098109911001101110211031104110511061107110811091110111111121113111411151116
ntial of Nano-triterpenoid from Phytolacca decandra against A549ay, Journal of Acupuncture and Meridian Studies (2013), http://

12
10 J. Das et al.
+ MODEL
11171118111911201121112211231124112511261127112811291130113111321133113411351136113711381139114011411142114311441145114611471148114911501151115211531154115511561157115811591160116111621163116411651166116711681169117011711172117311741175117611771178
11791180118111821183118411851186118711881189119011911192119311941195119611971198119912001201120212031204120512061207120812091210121112121213121412151216121712181219122012211222122312241225122612271228122912301231123212331234123512361237123812391240
JAMS135_proof ■ 19 August 2013 ■ 10/11
in dBA- and NdBA-induced apoptosis. Our main focus wasthen shifted toward understanding the possible role of al-terations in Ca2þ homeostasis induced by both dBA andNdBA. Such an approach seems to have not been made byany other workers earlier in respect of these drugs. It isknown that the apoptotic process is preceded by thecollapse of the mitochondrial potential, which could betriggered by multiple stimuli such as changes in Ca2þ, ox-ygen radicals, swelling of the matrix and rupture of theouter membrane with ensuing changes in the permeabilityof the outer mitochondrial membrane, and release ofapoptogenic factors. Results of the present study indicatedthat the intracellular calcium ([Ca2þ]i) concentrationincreased significantly after the administration of both dBAand NdBA. Pretreatment with EGTA (Ca2þ chelator) did notprevent the increase of [Ca2þ]i, which indicated that thecalcium was from an intracellular store and not from anextracellular store. Ca2þ release from the ER is known to beapoptogenic [22], which ultimately leads to the loss of[Ca2þ]i homeostasis, and associated with cellularapoptosisda common mechanism of cell death [23]. In thisregard, results of our studies are in conformity with thoseof Yang et al [24] and Gao et al [25], indicating that loss ofCa2þ from ER lumen plays an important role in exerting thecytotoxic effects of dBA and NdBA, presumably by inducingacute ER stress.
dBA and NdBA treatments also caused breakdown of theoxidative phosphorylation system by ATP/ADP ratio deple-tion and dissipation of DJm and elevation of ROS, indi-cating that dBA could target mitochondria; however, it wasnoted that NdBA acted more effectively compared to dBA.Disruption of DJm leads to mitochondrial permeabilizationthat commonly accompanies or precedes the release ofcytochrome c into the cytoplasm, leading to the formationof an active apoptosome complex and the activation ofcaspase cascade, which then activates the executionercaspase3 to orchestrate apoptosis [26].
Our result showed that intracellular ROS levels weresignificantly increased in a dose-dependent manner afteradministration of dBA and NdBA. However, the relationshipbetween intracellular calcium and oxidative stress may becomplex, because oxidative stress initiates changes in cal-cium concentrations in cells, which may subsequentlyactivate the production of ROS.
Marked chromatin condensation and apoptotic bodyformation in dBA- and NdBA-treated cells were alsoobserved in cells after the administration of the highestdose (100 mg/mL). The annexin V-FITC/PI assay revealedthat early and late apoptotic percentages after NdBAtreatments (50 mg/mL and 100 mg/mL) were significantlyhigher than those of dBA-treated ones. The result suggeststhat there is a possibility that both dBA and NdBA promotedthe levels of ER stress associated proteins and increased theexpression of Bax and also inhibited the levels of Bcl2,resulting in disruption of the mitochondrial membrane po-tential and subsequent release of cytochrome c frommitochondria to cytosol. This was followed by caspase3activation that finally resulted in apoptosis of A549 cells. Aschematic representation of the possible sequence ofevents is summarized in Fig. 5.
In order to examine if the nanoencapsulated ingredientcould readily enter different organ systems of living animals
Please cite this article in press as: Das J, et al., Strong Anticancer PoteAdenocarcinoma via a Ca2þ-dependent Mitochondrial Apoptotic Pathwdx.doi.org/10.1016/j.jams.2013.07.009
in vivo faster than the dBA, the relative distribution of thedrugs was assessed in different organ systems of mice,having a close resemblance to the human system, forextrapolation. The result confirmed that NdBA enteredfaster than dBA inside all tissues, including the lung, andcould also cross the bloodebrain barrier. This qualifiesNdBA to be a better candidate and a more potent anti-cancer drug for lung cancer, as it provided a better efficacyin drug delivery. This information can be of considerablehelp in future drug development and in the therapeuticmanagement of human lung cancer. However, prior to whenit can be recommended for practical use, more animalexperiments are necessary to confirm if it has a similarefficacy in other mammalian models.
Acknowledgments
Financial assistance for the work by Boiron Laboratories,Lyon, France, is gratefully acknowledged.
References
1. Reed JC. Apoptosis-targeted therapies for cancer. Cancer Cell.2003;3:17e22.
2. Demaurex N, Distelhorst C. Apoptosisdthe calcium connec-tion. Science. 2003;300:65e67.
3. Hajnoczky G, Davis E, Madesh M. Calcium and apoptosis. Bio-chem Biophys Res Commun. 2003;304:445e454.
4. Wang X. The expanding role of mitochondria in apoptosis.Genes Dev. 2001;15:2922e2933.
5. Ghafourifar P, Klein SD, Schucht O, Schenk U, Pruschy M,Rocha S, et al. Ceramide induces cytochrome C release fromisolated mitochondria. Importance of mitochondrial redoxstate. J Biol Chem. 1999;274:6080e6084.
6. Starkov AA, Polster BM, Fiskum G. Regulation of hydrogenperoxide production by brain mitochondria by calcium and Bax.J Neurochem. 2002;83:220e228.
7. Das S, Das J, Samadder A, Khuda-Bukhsh AR. Dihydroxy-iso-steviol methyl ester isolated from Pulsatilla nigricans inducesapoptosis in HeLa cells: its cytoxicity and interaction with calfthymus DNA as target. Phytother Res. 2012. http://dx.doi.org/10.1002/ptr.4768. Q
8. Das J, Das S, Samadder A, Bhadra K, Khuda-Bukhsh AR. Poly(-lactide-co-glycolide) encapsulated extract of Phytolacca dec-andra demonstrates better intervention against induced lungadenocarcinoma in mice and on A549 cells. Eur J Pharm Sci.2012;47:313e324.
9. Das S, Das J, Samadder A, Boujeddaini N, Khuda-Bukhsh AR.Apigenin-induced apoptosis in A375 and A549 cells throughselective action and dysfunction of mitochondria. Exp BiolMed. 2012;237:1433e1448.
10. Famsworth NR, Akerele O, Bingel AS, Soejarto DD, Guo Z.Medicinal plants in therapy. Bull World Health Org. 1985;63:965e981.
11. Ellingwood F. The American Materia Medica, Therapeutics andPharmacognosy. Sandy, OR: Eclectic Medical Publications;1919.
12. Wang L, Bai L, Nagasawa T, Hasegawa T, Yang X, Sakai J, et al.Bioactive triterpene saponin from the roots of Phytolaccaamericana. J Nat Prod. 2008;71:35e40.
13. Fessi H, Puisieux F, Devissaquet JP, Ammoury N, Benita S.Nanocapsule formation by interfacial polymer depositionfollowing solvent displacement. Int J Pharm. 1989;55:1e4.
ntial of Nano-triterpenoid from Phytolacca decandra against A549ay, Journal of Acupuncture and Meridian Studies (2013), http://

Q13
Anticancer potential of Phytolacca decandra 11
+ MODEL
1241124212431244124512461247124812491250125112521253125412551256125712581259126012611262
1263126412651266126712681269127012711272127312741275127612771278127912801281128212831284
JAMS135_proof ■ 19 August 2013 ■ 11/11
14. Mosmann T. Rapid colorimetric assay for cellular growth andsurvival. J Immunol Methods. 1983;65:55e63.
15. Biswas R, Mandal SK, Datta S, Bhattacharyya SS, Boujeddaini N,Khuda Bukhsh AR. Thujone-rich fraction of Thuja occidentalisdemonstrates major anti-cancer potentials: evidence fromin vitro studies on A375 cells. Evid Based Compliment AlternMed. 2010. http://dx.doi.org/10.1155/2010/neq042, 2011.
16. Schafer ZT, Grassian AR, Song L, Jiang Z, Hines ZG, Irie HY,et al. Antioxidant and oncogene rescue of metabolic defectscaused by loss of matrix attachment. Nature. 2009;461:109e113.
17. Ishmukhametov RR, Pond JB, Al-Huqail A, Galkin MA, Vik SB.ATP synthesis without R210 of subunit a in the Escherichia coliATP synthase. Biochim Biophys Acta. 2008;1977:32e38.
18. Dhiman R, Kathania M, Raje M, Majumder S. Inhibition of bf1-1/A1 by SiRNA inhibits mycobacterial growth in THF-1 cellsby enhancing phagosomal acidification. Biophys Acta. 2008;1780:733e742.
19. Hsin LI, Ou CC, Wu TC, Jan MS, Wu MF, Chiu LY, et al. GMI, animmunomodulatory protein from Ganoderma microsporum,induces autophagy in non-small cell lung cancer cells. Auto-phagy. 2011;7:873e882.
Please cite this article in press as: Das J, et al., Strong Anticancer PoteAdenocarcinoma via a Ca2þ-dependent Mitochondrial Apoptotic Pathwdx.doi.org/10.1016/j.jams.2013.07.009
20. Burnette WN. “Western blotting”: electrophoretic transfer ofproteins from sodium dodecyl sulfate-polyacrylamide gels tounmodified nitrocellulose and radiographic detection withantibody and radio iodinated protein. Anal Biochem. 1981;112:195e203.
21. Bhattacharyya SS, Paul S, Khuda-Bukhsh AR. Encapsulatedplant extract (Gelsemium sempervirens) poly(lactide-co-glycoside) nanoparticles enhance cellular uptake and increasebioactivity in vitro. Exp Biol Med. 2010;235:678e688.
22. Wertz IE, Dixit VM. Characterization of calcium release acti-vated apoptosis of LNCaP prostate cancer cells. J Biol Chem.2000;275:11470e11477.
23. Kass GEN, Orrenius S. Calcium signaling and cytotoxicity. En-viron Health Perspect. 1999;107:25e35.
24. Yang H, Zhang Q, He J, Lu W. Regulation of calcium signaling inlung cancer. J Thorac Dis. 2010;2:52e56.
25. Gao LL, Li FR, Jiao P, Yao ST, Sang H, Si YH. Apoptosis of humanovarian cancer cells induced by Paris chinensis dioscin via aCa2þ-mediated mitochondrion pathway. Asian Pacific J CancerPrev. 2011;12:1361e1366.
26. Green DR. Apoptotic pathways: ten minutes to dead. Cell.2005;121:671e674.
ntial of Nano-triterpenoid from Phytolacca decandra against A549ay, Journal of Acupuncture and Meridian Studies (2013), http://


















