






Strain-Specific Role of RNAs in Prion Replication. Running...
46
1 Strain-Specific Role of RNAs in Prion Replication. 1 Running Title: RNA and Prion Replication 2 Paula Saá 1,2,# , Gian Franco Sferrazza 1,3 , Gregory Ottenberg 1,3 , Anja M. 3 Oelschlegel 1 , Kerri Dorsey 2 and Corinne I. Lasmézas 1,# 4 1 The Scripps Research Institute, Scripps Florida, Department of Infectology, 130 Scripps Way, Jupiter, 5 FL 33458, USA 6 2 American Red Cross, Biomedical Services, Transmissible Diseases Department, 15601 Crabbs Branch 7 Way, Rockville MD 20855, USA 8 3 Present address: OPKO Health Inc., Diagnostic Research, 555 Heritage Dr., Jupiter FL 33458, USA. 9 # Corresponding authors: [email protected] , [email protected] 10 Abstract word count: 250 11 Manuscript word count: 8,921 12 13 Copyright © 2012, American Society for Microbiology. All Rights Reserved. J. Virol. doi:10.1128/JVI.01286-12 JVI Accepts, published online ahead of print on 18 July 2012 on June 12, 2018 by guest http://jvi.asm.org/ Downloaded from
Transcript of Strain-Specific Role of RNAs in Prion Replication. Running...

1
Strain-Specific Role of RNAs in Prion Replication. 1 Running Title: RNA and Prion Replication 2 Paula Saá1,2,#, Gian Franco Sferrazza1,3, Gregory Ottenberg1,3, Anja M. 3 Oelschlegel1, Kerri Dorsey2 and Corinne I. Lasmézas1,# 4 1 The Scripps Research Institute, Scripps Florida, Department of Infectology, 130 Scripps Way, Jupiter, 5 FL 33458, USA 6 2 American Red Cross, Biomedical Services, Transmissible Diseases Department, 15601 Crabbs Branch 7 Way, Rockville MD 20855, USA 8 3 Present address: OPKO Health Inc., Diagnostic Research, 555 Heritage Dr., Jupiter FL 33458, USA. 9 # Corresponding authors: [email protected], [email protected] 10 Abstract word count: 250 11 Manuscript word count: 8,921 12
13
Copyright © 2012, American Society for Microbiology. All Rights Reserved.J. Virol. doi:10.1128/JVI.01286-12 JVI Accepts, published online ahead of print on 18 July 2012
on June 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

2
Abstract 14 Several lines of evidence suggest that various co-factors may be required for prion 15 replication. PrP binds to polyanions and RNAs were shown to promote the conversion 16 of PrPC into PrPSc in vitro. In the present study we investigated strain-specific 17 differences in RNA requirement during in vitro conversion and the potential role of RNA 18 as strain-specifying component of infectious prions. We found that RNase treatment 19 impairs PrPSc converting activity of 9 murine prion strains by PMCA in a strain-specific 20 fashion. While the addition of RNA restored PMCA conversion efficiency, the effect of 21 synthetic polynucleotides or DNA was strain-dependent, showing a different promiscuity 22 of prion strains in co-factor utilization. The biological properties of RML propagated by 23 PMCA under RNA-depleted conditions were compared to brain-derived and PMCA 24 material generated in the presence of RNA. Inoculation of RNA-depleted RML in Tga20 25 mice resulted in an increased incidence of a distinctive disease phenotype 26 characterized by forelimb paresis. However, this abnormal phenotype was not 27 conserved in wild-type mice or upon secondary transmission. Immunohistochemical and 28 cell panel assay analyses of mouse brains did not reveal significant differences between 29 mice injected with the different RML inocula. We conclude that replication under RNA-30 depleted conditions did not modify RML prion strain properties. Our study cannot, 31 however, exclude small variations of RML properties that would explain the abnormal 32 clinical phenotype observed. We hypothesize that RNA molecules may act as catalysts 33 of prion replication, and that variable capacity of distinct prion strains to utilize different 34 co-factors may explain strain-specific dependency upon RNA. 35 36
on June 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

3
Introduction 37 Transmissible spongiform encephalopathies (TSEs) or prion diseases are characterized 38 by the accumulation in the brain and sometimes in the lymphoid tissues (13, 27) of an 39 abnormally structured form (PrPSc) of the host cellular prion protein (PrPC ) (26). PrPSc is 40 thought to be the only (5, 9, 21, 33) or the major (7, 11, 35) constituent of the infectious 41 agent, the prion. Prions occur in the form of diverse strains exhibiting specific biological 42 and biochemical characteristics (1, 4). Different prion strains show distinct interspecies 43 transmission properties, and in particular different pathogenicity for humans (20, 37). 44 The strain phenomenon is also important from a fundamental standpoint, as strain-45 specific properties of infectious agents have hitherto been encoded by the nucleic acid 46 genome of the pathogen. In the case of prions, strain-specific properties might be 47 determined by differences in PrPSc conformation, by differences in complex 48 glycosylation or by a yet-to-be defined informational molecule associated with PrPSc. 49 The ability to convert PrP into infectious PrPSc in vitro lends support to the concept that 50 the prion protein is the major component of prions (5, 9, 21, 33). Furthermore, it has 51 been shown that RNA molecules facilitate in vitro amplification of infectious PrPSc (9, 15, 52 33). However, the exact role of RNA in the amplification process remains unknown. 53 RNA could act as mere catalysts of the PrP misfolding process. On the contrary, RNA 54 may be associated with the infectious particle and contribute to prion strain 55 characteristics. A recent study showed that the requirement of RNA for in vitro 56 amplification of PrPSc is species dependent, with hamster-derived PrPSc being largely 57 dependent on the presence of RNA in the PMCA reaction mixture, whereas mouse-58 derived PrPSc does not require RNA for in vitro amplification (10). Another study showed 59
on June 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

4
similar RNA-dependent amplification of six hamster prion strains (15). Other 60 endogenous polyanions such as DNA, heparan sulfates or lipids may come into play in 61 conditions of RNA deficiency (10). 62 In the present study we further investigated the possibility of a strain-specific 63 dependency upon RNA during in vitro PrPSc amplification, and addressed the role of 64 RNA as a strain-specifying component of infectious prions. To this purpose, we 65 conducted PMCA amplification of various mouse prions in RNA-depleted as well as 66 control reaction mixtures. We studied the role of RNA in the efficiency of the 67 amplification reaction for each of 9 strains of mouse prions. We determined RML strain 68 characteristics by bioassay after amplification in RNA-depleted versus control PMCA 69 reactions. We conclude that RNA-dependency for in vitro conversion is prion strain-70 specific, and that RML prion strain identity is maintained after in vitro amplification under 71 conditions of RNA depletion. We propose that RNA molecules may act as strain-specific 72 catalysts of prion replication. 73 Materials and Methods 74 Preparation of Tissue Homogenates 75 Healthy animals were sacrificed by CO2 inhalation and immediately perfused with 76 phosphate-buffered saline (PBS) plus 5 mM ethylenediamminetetraacetic acid (EDTA) 77 prior to harvesting the brain. The perfusion was conducted by inserting a 20G needle 78 directly into the mouse heart and manually injecting an approximate volume of 30 ml of 79 5 mM EDTA in PBS. Terminal prion infected mice were sacrificed by CO2 inhalation and 80 brains were immediately harvested. Mouse brains were either homogenized or flash 81
on June 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

5
frozen in liquid nitrogen. Ten percent brain homogenates (w/v) were prepared in pre-82 chilled conversion buffer (PBS containing 150 mM NaCl, 1.0% Triton X-100, and the 83 Complete Protease Inhibitor Cocktail tablets containing EDTA from Roche Applied 84 Science). Infected and normal brain homogenates were prepared manually using a 85 glass potter-elvehjem homogenizer. Sample heating was avoided by keeping the potter 86 on ice throughout the process. 87 Dilutions of prion infected brain homogenates (IBHs) were done in normal brain 88 homogenates (NBHs), and they are expressed in relation to the brain; for example, a 89 100-fold dilution is equivalent to a 1% brain homogenate. 90 Preparation of PMCA Substrates 91 RNase A treatment: Aliquots of 10% normal brain homogenates prepared as described 92 above, were treated with 48 µg/ml (120 U/ml, final concentration) of RNase A (MP 93 Biomedicals, Solon, OH) for 1 hour at 37 ºC. RNase Inhibitor treatment: 100 µl 94 aliquots of 10% NBH were treated with 5 µl of RNase Out (2000 U/ml) (Invitrogen, 95 Carlsbad, CA) for 1 hour at 37 ºC. Control substrates: 100 µl aliquots of 10% NBH 96 were mixed with 5 µl of conversion buffer and incubated at 37 ºC for one hour. 97 Preparation of PMCA substrates for reconstitution with exogenous RNA: 100 µl 98 aliquots of 10% NBH were treated with 0.86 µg/ml (2 U/ml) of RNase A for 1 hour at 37 99 ºC. Then, 2000 U/ml of RNase Out were added to these preparations to inhibit RNase A 100 activity and allow sample reconstitution with exogenous RNA molecules, added to a 101 final concentration of 0.12 µg/µl. Preparation of PMCA substrates for reconstitution 102 with synthetic polyanions: 100 µl aliquots of 10% NBH were treated with 8.6 µg/ml 103
on June 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

6
(20 U/ml final concentration) of RNase A for 1 hour at 37 ºC. Then, synthetic poly(A) 104 and poly(G) (Sigma, St. Louis, MO) were added to these preparations to a final 105 concentration of 0.12 µg/µl. Preparation of PMCA substrates for reconstitution with 106 dsDNA: 100 µl aliquots of 10% NBH were treated with 8.6 µg/ml (20 U/ml final 107 concentration) of RNase A for 1 hour at 37 ºC. Thereafter, 500-200 bp salmon sperm 108 DNA fragments (Trevigen, Gaithersburg, MD) were added to these preparations to a 109 final concentration of 0.12 µg/µl. 110 Prion Replication by PMCA 111 Aliquots of 10% brain homogenates from prion infected mice were serially diluted into 112 10% normal brain homogenates pre-treated as described above and loaded onto 0.2 ml 113 PCR tubes containing zirconia/silica beads (Biospec Products Inc, Bartlesville, OK). 114 Tubes were positioned on an adaptor placed on the plate holder of a microsonicator 115 (Misonix Model 3000, Farmingdale, NY) and programmed to perform cycles of 30 min 116 incubation followed by a 20 s pulse of sonication set at potency of 9. Samples were 117 incubated, without shaking, immersed in the water of the sonicator bath set at 37 ºC. 118 The PMCA reaction was stopped after 24 hours (48 cycles) by applying a 20 s 119 sonication pulse. To avoid formation of amorphous aggregates that will interfere with 120 proteinase K (PK) digestion, this treatment was preformed prior to sample freezing. 121 For serial automated PMCA (saPMCA), a fraction of the amplified material specific for 122 each strain (see below) was transferred to a new PCR tube containing fresh pre-treated 123 normal brain homogenate and beads and amplified under the conditions described 124 above. To avoid generation of amorphous aggregates and in order to have 125
on June 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

7
homogenous samples, this dilution step was performed right after each PMCA round 126 and prior to sample freezing. To monitor the amplification efficiency, 18 µl aliquots were 127 taken prior to each round of PMCA and used as controls. To achieve similar conversion 128 efficiency for all prion strains, amplified RML, ME7 and 22L were diluted 10, 5 and 8 129 times respectively into NBH to seed the next round of amplification. 130 PK digestion 131 Samples were subjected to incubation in the presence of PK (100 µg/ml) for 60 min at 132 56 ºC with constant shaking at 450 rpm. The digestion was stopped by addition of 133 NuPAGE LDS sample buffer (Invitrogen, Carlsbad, CA) and heated at 100 ºC for 10 134 min. The presence of protease-resistant PrP was revealed by Western blotting. 135 NaPTA Precipitation 136 Aliquots of PMCA generated material were NaPTA precipitated as described in (32). 137 Thermolysin digestion 138 Samples were subjected to incubation in the presence of 50 µg/ml thermolysin for 60 139 min at 70 ºC with constant shaking at 450 rpm. The digestion was stopped by addition 140 of NuPAGE LDS sample buffer (Invitrogen, Carlsbad, CA) and heated at 100 ºC for 10 141 min. The presence of protease-sensitive PrPSc was revealed by Western blotting. 142 Western Blotting 143 Proteins in NuPAGE LDS sample buffer (Invitrogen, Carlsbad, CA) were fractionated by 144 SDS-polyacrylamide gel electrophoresis, electro-blotted onto a PVDF membrane 145 (Millipore) and probed with D18 antibody, diluted 1:30,000 from a 1 mg/ml, stock 146
on June 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

8
followed by a murine HRP-conjugated anti human IgG antibody at a 1:15,000 dilution 147 (Southern Biotech, Birmingham, AL). Immunoreactive bands were visualized by 148 incubation with West Pico (Pierce, Rockford, IL) and imaged using a BioSpectrum® 149 Imaging System (UVP, Upland, CA) 150 RNA Purification and Fractionation 151 RNA samples were purified from mouse brain and liver. Total RNA as well as large and 152 small RNA preparations were purified with the mirVana™ Kit (Ambion, Carlsbad, CA) 153 following the manufacturer’s instructions. After total RNA purification, the samples were 154 treated with DNase and PK to eliminate potential DNA and protein contamination that 155 may interfere with the result interpretation. Upon conclusion of these treatments, the 156 samples were re-purified using the same kit to remove the enzymes. Thereafter, RNA 157 purification was either finished or continued for further RNA fractionation into large 158 (≥200 nt) or small (<200 nt) molecules according to the mirVana™ protocol. RNA 159 concentration and size was evaluated by RiboGreen® RNA Quantitation (Molecular 160 Probes, Eugene, OR) 1.2 % denaturing agarose electrophoresis (RNA ≥800 nt) and 161 Novex® 15% denaturing polyacrylamide TBE-Urea gels (RNA 20-800 nt) (Invitrogen, 162 Carlsbad, CA). 163 RT-PCR 164 RNA samples were purified from PMCA substrates after treatment with RNase A, 165 RNase inhibitor or left untreated with the mirVana™ Kit (Ambion, Carlsbad, CA). 166 Thereafter, quantification of the levels of specific miRNAs (as indicated in the results 167 section) rRNA and housekeeping RNAs was evaluated with the TaqMan® MicroRNA 168
on June 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

9
Assays or the TaqMan® Ribosomal RNA Control Reagents following the manufacturer 169 instructions. 170 Mouse Bioassay and Titrations 171 The use of animals was conducted according to institutional guidelines after review of 172 the protocol by the Institutional Animal Care and Use Committee. Tga20 mice 173 expressing wild-type PrPC at about 8 times the endogenous levels, or wild-type 174 C57BL/6 mice were used as an experimental model of scrapie. Animals were 4–6- 175 weeks old at the time of inoculation. Mice anesthetized by isoflurane were injected 176 intracerebrally in the prefrontal cortex area with 20 µl of the sample. They were 177 euthanized by CO2 followed by decapitation when they showed terminal signs of prion 178 disease. Brains were extracted and analyzed biochemically and 179 immunohistochemically. The right cerebral hemisphere was frozen and stored at -80 °C 180 for secondary transmission experiments and biochemical examination of PrPSc using 181 Western blotting analysis after PK digestion as described above. The left hemisphere 182 was fixed in Carnoy’s solution and processed as described below. 183 For inocula titration, groups of 8 or 6 mice were inoculated as above with limiting 184 dilutions of brain-derived or PMCA generated RML or ME7 (where indicated) in the 185 presence or absence of RNase A and RNase Inhibitor. To control the spontaneous 186 generation of prions by PMCA mice were inoculated with unseeded NBHs subjected to 187 the same rounds of PMCA as seeded reactions. Additional controls included unseeded 188 NBHs as well as brain-derived RML treated with RNase A and RNase Inhibitor. The 189
on June 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

10
lethal dose 50 (LD50) of each inocula was calculated according to the Reed & Muench 190 method (28). 191 Ethics Statement 192 The Institutional Animal Care and Use Committee (IACUC) of 193 The Scripps Research Institute, (TSRI) Scripps Florida reviewed and approved the 194 animal protocol entitled Role of RNAs in prion replication (Animal protocol number 07-195 012). The Scripps Research Institute, (TSRI) Scripps Florida maintains a 196 centralized animal care and use program registered by the USDA, assured with the 197 Office of Laboratory Animal Welfare (OLAW) and accredited by the Association for 198 Assessment and Accreditation of Laboratory Animal Care, International (AAALAC). 199 Housing and care of animals is consistent with the Public Health Service (PHS) Policy 200 on Humane Care and Use of Laboratory Animals, The Guide for the Care and Use of 201 Laboratory Animals, the Animal Welfare Act, and other applicable state and local 202 regulations. 203 Scrapie Associated Fibers (SAF) Purification 204 100 µl of PMCA and brain-derived samples were mixed with 4 µl of Benzonase (2.5 205 U/µl, stock 1:10 diluted in PBS → 50 U/ml) + 1 µl of 0.2 M MgCl2 (final conc. 1 mM), and 206 incubated for 30 min at 37 ºC. The samples were then mixed with 200 µl of buffer T7 207 (10% NaCl, 10% Sarkosyl, 1% SB 3-14, 2 mM Tris pH 7.4) and incubated for 15 min at 208 RT (25 ºC) in a Thermomixer (Bio-Rad) set at 450 rpm. Following this incubation the 209 preparations were layered over 100 µl of a sucrose cushion (10% NaCl, 10% Sucrose, 210 0.1% SB 3-14, 2 mM Tris pH7.4) and spun for 2 h at maximum speed (13,200 rpm, 211
on June 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

11
16,100 rfc) on a bench top centrifuge (eppendorf, 5415R) set at 18 ºC. The pellets were 212 resuspended by sonication in 100 µl of PBS containing 3 mg/ml of BSA. 10% NBHs 213 prepared from perfused non-inoculated C57BL/6 mice were used as negative controls. 214 Calculation of PrPSc Specific Infectivity 215 To calculate the specific infectivity per PrPSc molecule of RML generated by PMCA, 1:2 216 serial dilutions of SAF preparations obtained from PMCA and brain-derived samples 217 were analyzed by Western blot (Fig. 5A). The band intensity was quantified by 218 densitometry using a BioSpectrum® Imaging System (UVP, Upland, CA). After 219 background subtraction, the background being the signal generated by the uninfected 220 brain homogenate (Fig. 5A) we calculated the band density corresponding to 20 µl of 221 sample; that was the volume inoculated in mice. The band density of the sample dilution 222 killing 50% of the inoculated mice was calculated by multiplying the band density in 20 223 µl of sample by the LD50 (calculated from the data in Table 1). We arbitrarily defined 1 224 pixel unit as 1 molecule of PrPSc; and the specific infectivity per PrPSc molecule 225 generated by PMCA was calculated as the reciprocal of the density of the dilution killing 226 50% of the inoculated mice relative to the standard RML inoculum. 227 Immunohistochemistry 228 Brains were harvested immediately after euthanasia. The right cerebral hemisphere was 229 frozen and stored at -80 °C as described above. The left hemisphere was fixed in 230 Carnoy’s solution for 48 hours before proceeding. Brain tissues were processed using a 231 Leica ASP300 tissue processor (Leica, Buffalo Grove, IL) and embedded using a Leica 232 paraffin embedding machine. Tissues were mounted in sagittal orientations and 233
on June 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

12
sectioned at 4 microns for immunohistochemistry or 10 microns for hematoxylin and 234 eosin (H&E) staining. For immunohistochemistry, sections were labeled with PrP 235 primary antibody Bar233 (Cayman Chemicals, Ann Arbor, MI) according to (18). 236 Extended Cell Panel Assay 237 Cell lines were maintained and the Cell Panel Assay was performed as described in (3, 238 22, 25). 239 Statistics 240 The Chi-Square test was performed to determine whether the abnormal clinical 241 phenotype observed in treatment groups RML-PRNaseA and RML-PRNaseOut were 242 statistically different from each other at a 5% level. 243 A One-way ANOVA test was used to determine statistically significant differences in 244 survival times amongst the treatment (RML-BRNaseA and RML-BRNaseOut) and control 245 (RML-B) groups. Additionally, if a difference was found, a Scheffe's test was utilized to 246 identify exactly which groups were different. A Log-Rank test was also used to 247 determine if the survival curves for each group were statistically different from each 248 other. 249 SAS 9.2 (Cary, North Carolina) was used for all calculations. 250 Results 251 RNA depletion impairs in vitro prion replication in a strain-specific fashion 252
on June 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

13
To address the role of RNA molecules in prion pathogenesis, we first evaluated the 253 effect of RNA depletion by RNase A in in vitro PrPSc amplification reactions. Aliquots of 254 10% normal brain homogenates (NBHs) were treated for 1 hour with RNase A, RNase 255 inhibitor (“RNase Out”) or left untreated, as described in the methods section. The 256 efficiency of RNA removal was evaluated by Real Time quantitative PCR (RT-qPCR) 257 using specific probes for the brain-specific miRNA miR-124a, ubiquitous miRNAs like 258 let-7b and the ribosomal RNA 18S (Fig. S1). Infected brain homogenates (IBH) 259 prepared from C57BL/6 mice inoculated with the mouse prion strains RML, ME7 and 260 22L were treated with the same enzymes, added to these substrates and amplified by 261 serial automated PMCA (saPMCA). After initial efficient replication in all substrates, the 262 conversion of various prion strains over several rounds was distinctively affected in the 263 absence of RNAs (Fig. 1). While RML amplification remained unaffected for up to 9 264 saPMCA rounds, ME7 conversion ceased completely after 4 rounds in RNase A-treated 265 substrates. 22L exhibited an intermediate behavior as illustrated by decreased 266 replication efficiency after 3 rounds of saPMCA that was maintained in subsequent 267 rounds (Fig. 1). It has been reported that PrPSc becomes sensitive to PK digestion after 268 RNase A treatment suggesting that RNA is a component of PrPSc and that it confers 269 proteinase resistance (17). To investigate whether the absence of ME7 signal in our 270 PMCA reaction was due to an increased sensitivity to PK or to impaired replication in 271 the absence of RNA, we evaluated the presence of PK-sensitive PrPSc (sPrPSc) in our 272 samples. Aliquots of PMCA material generated in the presence or absence of RNA 273 were NaPTA precipitated and thermolysin digested according to previous reports (8). 274
on June 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

14
Even after this treatment no PrPSc signal could be detected in ME7 samples generated 275 in the absence of RNA molecules (data not shown). 276 These findings prompted us to investigate the degree of RNA dependency for efficient 277 in vitro amplification of several mouse prion strains. Since the three strains above are 278 characterized by different conversion efficiencies under normal conditions, with RML 279 being the most efficient followed by 22L and ME7, we wondered whether the observed 280 impaired amplification under RNA-depleted conditions correlated with replication 281 efficiency. To investigate this question, we utilized several mouse-propagated prion 282 strains characterized by different incubation times in the mouse bioassay, as well as 283 three additional prion strains generated in vitro by saPMCA (kindly provided by Dr. J. 284 Castilla). These samples were serially diluted into RNA-depleted or control NBHs and 285 aliquots of these dilutions were in vitro amplified for one PMCA round (Fig. 2A). This 286 method highlighted the differential in vitro amplification capacity of prion strains under 287 normal conditions (Fig. 2A, Control PMCA); the extent to which the seed can be diluted 288 while still allowing amplification to occur is indicative of the conversion efficiency of the 289 strain under consideration. Interestingly, RNA depletion of the conversion reaction 290 affected this parameter differently depending on the strain. These differences could also 291 be seen between strains with comparable replication capacity such as 139 A and 766 292 (Fig. 2A and 2B). Moreover, lower conversion efficiency was not necessarily associated 293 with higher RNA dependency. Strains 139A, 303 and 726 exhibited lower conversion 294 efficiency than RML and 79A, yet their in vitro conversion was less affected by RNA 295 depletion (Fig. 2A and 2B). These results indicate that RNA-dependency for in vitro 296 prion replication is strain-specific. 297
on June 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

15
Addition of exogenous RNA to RNA-depleted PMCA reactions enhances prion 298 replication 299 To demonstrate that the inhibitory effect of RNase A treatment on PMCA reactions was 300 due to RNA depletion, we reconstituted RNA-depleted substrates with brain-extracted 301 total RNA and evaluated the efficiency of ME7 conversion in these samples in 302 comparison with non-reconstituted substrates. The presence of exogenous RNA 303 enhanced ME7 amplification (Fig. 3A). Liver-derived RNA showed similar efficiency to 304 brain-derived RNA; moreover, we observed a dose-dependent reconstitution of prion 305 conversion (Fig. 3B). While 0.01 µg/µl of total RNA were not sufficient to increase prion 306 replication after RNase A treatment, addition of 0.06 µg/µl and 0.12 µg/µl enhanced 307 ME7 conversion in a dose-dependent manner (Fig. 3B panels 1, 5 and 6). We evaluated 308 the levels of RNA in the substrates after RNase A and RNase Inhibitor treatments as 309 well as after RNA reconstitution, both before and after PMCA (Fig. S2). Addition of 0.12 310 µg/ml of RNA to RNase A treated NBHs brought RNA levels back to those found in 311 untreated NBH as determined by RiboGreen Quantitation (Fig. S2 compare lanes 3 and 312 9). 313 To investigate the size of RNA involved in PrPSc replication, RNA was purified and size 314 fractionated from mouse livers. Large (≥200 nt) and small (<200 nt) RNA samples were 315 added at different concentrations to aliquots of serially diluted ME7 and RML on RNA-316 free substrates. We again observed the dose-dependent effect of RNA in PrPSc 317 conversion. Large and small RNAs restored PrPSc amplification equally well (Fig. 3B 318 compare panels 2, 3, 5 and 6). We analyzed the quality of the RNA after PMCA. A 2:1 319 ratio for 28S:18S rRNA has long been considered the benchmark for intact RNA. This 320
on June 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

16
ratio was detected in our NBHs, RNase Inhibitor treated or reconstituted samples prior 321 to PMCA amplification; however, similar to a previous report (15) we observed 322 fractionation of large RNA species into smaller fragments (<200nt) at the end of the 48 323 cycles PMCA round (Fig. S2 compare odd and even lanes) 324
We evaluated the capacity of synthetic RNA or DNA to reconstitute mouse prion 325 amplification in RNA-depleted substrates. Taking advantage of the RNase A specificity 326 for pyrimidine nucleoside linkages, heteropolymeric preparations of polyadenylic 327 [poly(A)] or polyguanylic [poly(G)] acids were added to aliquots of RNase A-treated 328 substrates in the absence of RNase Out. Serial dilutions of RML and ME7 prions were 329 prepared in these substrates and aliquots of these samples amplified by PMCA. 330 Interestingly, conversion stimulation by poly(A) and poly(G) was different between the 331 two strains. ME7 amplification was restored to control levels by poly(A) but not poly(G) 332 (Fig. 4A). PMCA amplification of RML, on the other hand, was reconstituted by poly(A) 333 and poly(G) (Fig. 4A). These results suggested a higher co-factor selectivity of ME7 334 compared to RML. In line with this hypothesis, the addition of 200-500bp fragments of 335 DNA to RNA-depleted substrate did not reconstitute ME7 conversion, whereas it did 336 restore RML amplification (Fig. 4B). 337 RML strain identity is maintained after propagation in RNA depleted substrates 338 The unified theory of prion propagation hypothesizes that nucleic acid molecules may 339 confer specific characteristics to prion strains (35). Our data show that strains differ in 340 their nucleic acid requirements to propagate in vitro. These findings prompted us to 341 investigate whether prion amplification in RNA-depleted substrates affected their strain 342
on June 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

17
properties. To this end, we generated, by in vitro conversion, RML and ME7 343 preparations devoid of RNA and brain-derived PrPSc molecules (RML-PRNaseA and ME7-344 PRNaseA). We performed rounds of saPMCA until a 10-15 dilution of the IBH was achieved 345 according to previous calculations (5). Due to differences in replication efficiency 346 between strains, a dilution factor of 10 and 5 had to be utilized for RML and ME7 347 respectively, to obtain comparable amplification between rounds (Fig. 1), implying to 348 perform 13 and 18 rounds of saPMCA for RML and ME7 respectively to achieve a 10-15 349 dilution of the infected brain homogenate (IBH). As controls, these strains were serially 350 propagated in untreated (RML-P and ME7-P) and RNase Out-treated brain homogenate 351 used as substrates (RML-PRNaseOut and ME7-PRNaseOut). Similar to the experiment 352 depicted in Fig. 1, amplification of ME7 in the absence of RNA was lost after 5 rounds of 353 saPMCA, while RML amplification was unchanged for up to 8 rounds. Despite a 354 decrease in conversion efficiency in RNA-depleted substrates, RML conversion was 355 maintained until round 13 of saPMCA. Propagation of RML and ME7 in control 356 substrates was efficiently maintained over 13 and 18 rounds respectively (not shown). 357
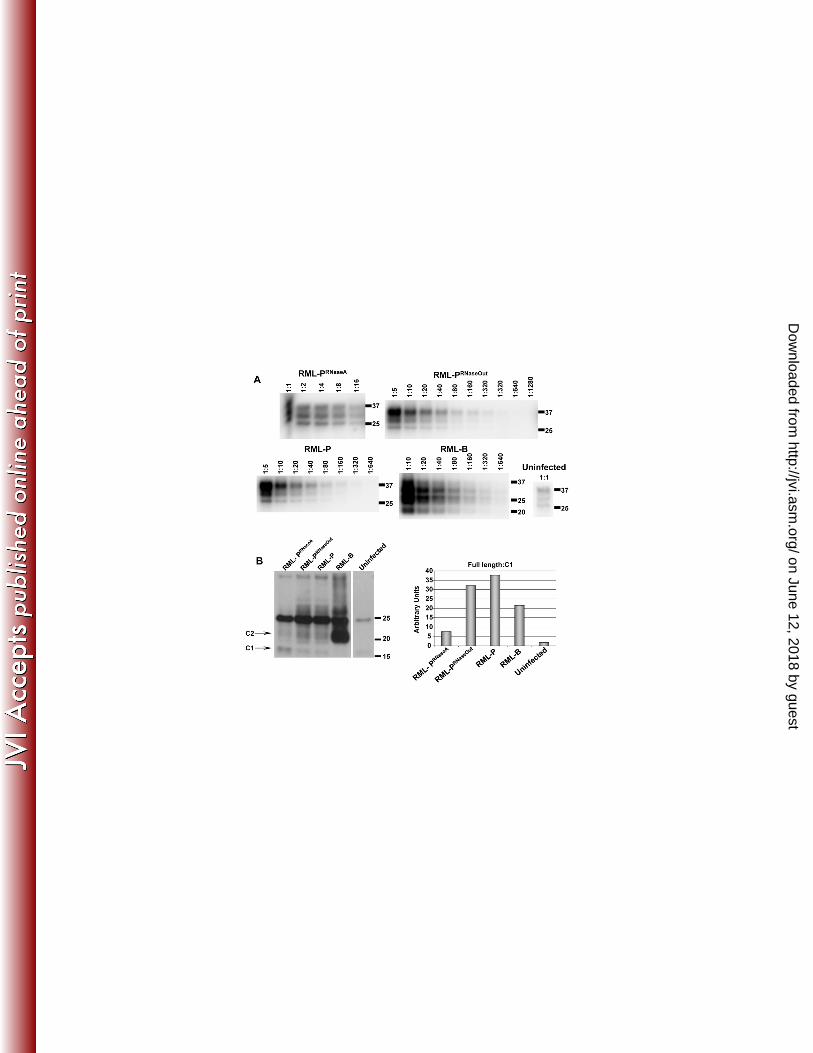
PrPSc levels of RML prions serially amplified in vitro were evaluated by Western 358 blot analysis and compared to brain-derived RML preparations (Fig. 5A). Intriguingly, 359 the PrPSc pattern in RML-PRNaseA samples was more similar to that observed in brain 360 homogenate samples than RML-P and RML-PRNaseOut samples. We performed PrP 361 deglycosylation experiments to analyze the proportion of truncated PrP species C1 362 (generated from PrPC in the absence of prion infection) and C2 (generated from PrPSc 363 during prion infection) (6, 38). C1 level was significantly higher in RML-PRNaseA than in 364 RML-PRNaseOut and RML-P samples, whereas brain-derived RML exhibited a high level 365
on June 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

18
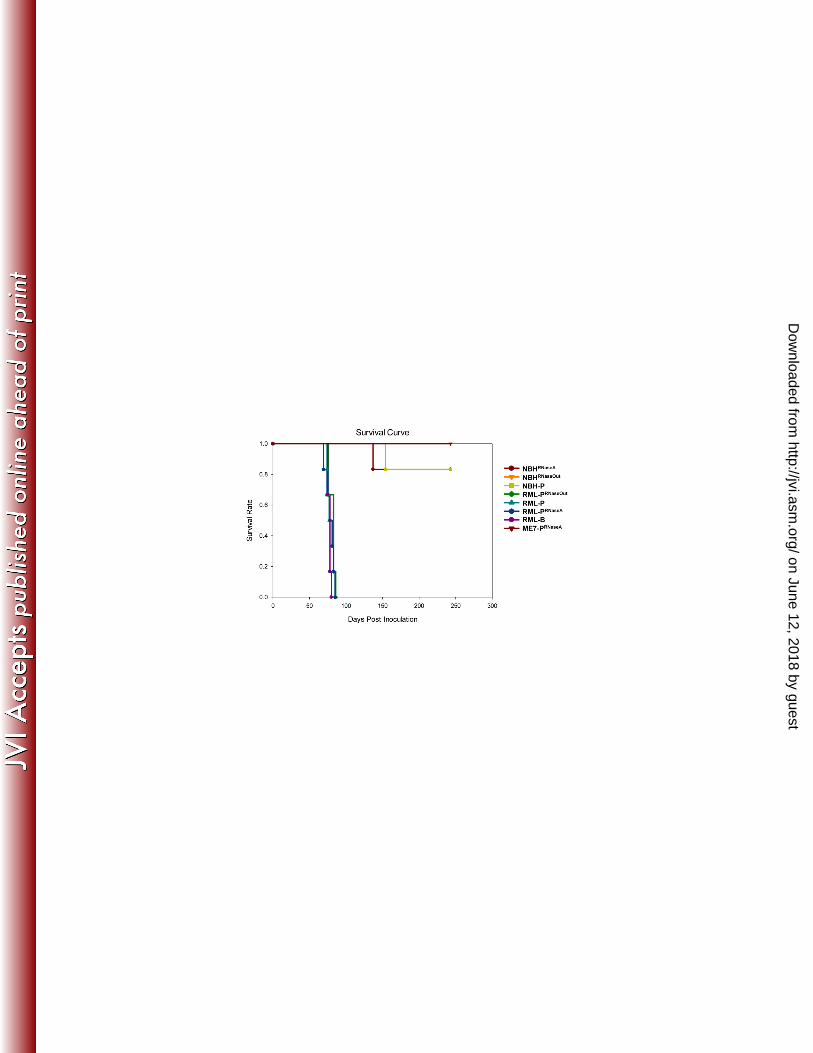
of C2 (Fig. 5B). The higher level of C1 or C2 in both preparations is likely to account for 366 the similar pattern observed in the complex glycosylated samples in Fig. 5A. Higher C2 367 levels in the infected brain homogenate than in PMCA samples is likely due to the 368 activity of endogenous PrPSc-cleaving enzymes as described previously (14, 38), 369 whereas protease inhibitors present in the PMCA reactions prevent this activity. At the 370 present time, we can only speculate about the reasons underlying the difference in the 371 proportion of full-length PrP to C1 observed between RML-PRNaseA and the other PMCA 372 samples. Absence of RNA binding to PrP might lead to a loss of steric protection or to a 373 conformational change rendering PrP slightly more sensitive to the limited proteolytic 374 cleavage that occurs despite the presence of protease inhibitors. To investigate if the 375 absence of RNA during RML conversion affected its strain-specific characteristics, 1/10 376 dilutions of RML-PRNaseA were prepared and titrated by intra-cerebral inoculation into 377 Tga20 mice. Incubation periods were not statistically different from those resulting from 378 titration of control samples performed in parallel (Fig. 6 and Table 1). As expected, none 379 of the NBH samples treated with either RNase A or RNase Out caused prion disease in 380 inoculated mice. Likewise, NBHs subjected to 18 rounds of saPMCA in the absence of 381 prion seeds were not infectious to Tga20 mice, ruling out the spontaneous generation of 382 PrPSc under these conditions (Fig. 6 and Table 1). We did not detect infectivity in any of 383 the ME7-PRNaseA samples, showing that absence of RNA abolished both in vitro PrPSc 384 conversion and amplification of infectious ME7 prions (Fig. 6). Treatment of brain-385 derived RML with RNase A or RNase Out for 1 hour at 37 ºC prior to mouse inoculation 386 had no effect in the disease incubation and clinical manifestation (Table 1, compare 387 RML-B 10-5 with RML-BRNaseA and RML-BRNaseOut 10-5. p-value 0.17 as determined by 388
on June 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

19
One-way ANOVA). However, an atypical phenotype characterized by forelimb paresis 389 (Movie S1 and S2) was observed in groups RML-PRNaseA (12 out of 22 mice) and RML-390 PRNaseOut (2 out of 22 mice). The higher incidence of this phenotype in the RML-PRNaseA 391 group was statistically significant (p-value 0.0012 as determined by Chi-Square). 392 Despite containing lower levels of PrPSc, RML-PRNaseA inoculum produced incubation 393 times in Tga20 comparable to those of RML-PRNaseOut and RML-P inocula (Fig. 5A and 394 Table 1). Hence, we measured specific infectivity of PrPSc in samples generated with 395 and without RNAs. Our estimations indicated that PrPSc in RML-PRNaseA samples was 396 associated with 3.5 times more infectivity than that in control samples (Table 2); 397 however, the difference was too small to be attributed to the generation of a new prion 398 strain. 399 To further test the possibility of the emergence of a new RML-derived prion strain, we 400 performed intracerebral inoculations of PMCA material amplified in the presence (RML-401 P) or absence of RNA (RML-PRNaseA) into wild-type C57BL/6 mice. We evaluated the 402 infectivity of RML-PRNaseA by inoculating the same amount of PrPSc in those and in 403 PMCA control samples (RML-P) and comparing disease incubation times. We found 404 clinical end-points at 159±1 days after RML-P inoculation and 154±2 days after RML-405 PRNaseA injection, and classical RML clinical signs in 100% of the mice (Table 3), 406 suggesting that RML-PRNaseA and RML-P are the same prion strain. Secondary 407 transmissions in C57BL/6 or Tga20 mice of RML-PRNaseA propagated in C57BL/6 and 408 Tga20 mice, respectively, were characterized by typical clinical sings in all mice, and 409 incubation periods of 141±4 and 66±2 days respectively, very similar to the 144±5.5 and 410 59 days induced by brain-derived RML (Table 4 and (18)). 411
on June 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

20
To investigate if the forelimb paresis induced by RML-PRNaseA inoculation reflected a 412 change in strain neurotropism, we conducted a blind immunohistochemical analysis of 413 Tga20 and C57BL/6 mouse brains inoculated with RML-PRNaseA as well as RML-P. 414 Analysis of sections of the hippocampus, cortex, thalamus, cerebellum, medulla and 415 pons from these animals did not reveal any differences in vacuolation patterns (Fig. 7A 416 and 7B). We also failed to detect differences in PrPSc accumulation patterns suggesting 417 that RML strain was not modified by replication in the absence of RNA (Fig. 7C). In 418 addition, we examined RML properties in vitro in a Extended Cell Panel Assay (ECPA) 419 (3, 22, 25). The latter examines the capacity of a given prion strain to infect different cell 420 types in culture and its susceptibility to different pharmacological inhibitors of prion 421 replication. These responses have been shown to vary from one strain to another and 422 can even be used to distinguish between “substrains” of prions. PMCA-generated 423 material was either ultracentrifuged (100,000 x g, 1 h, 4 ºC) or diluted 2000X prior to cell 424 inoculation to reduce Triton X-100 levels contained in these preparations that are toxic 425 to cultured cells. Unfortunately, these procedures yielded preparations with very low 426 titers of infectivity that hindered us from drawing any conclusions (not shown). 427 Alternatively, we used Tga20 brain homogenates infected with RML-PRNaseA, RML-428 PRNaseOut and original RML. All RML-related inocula showed similar properties in the 429 ECPA. ME7 and 22L properties in the ECPA are shown as reference (Fig. 8). 430 Discussion 431 In the present study, we show that RNA enhances in vitro amplification of several 432 mouse prions strains: RML, 79A, ME7, 139A, 22L, 22F, 303, 766 and 726 (the latter 433 three murine prion strains were generated by PMCA, a generous gift of J. Castilla). 434
on June 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

21
Therefore, the effect of RNA as a co-factor for PMCA is not restricted to hamster prions. 435 Deleault et al. previously reported an absence of effect of RNase treatment on the 436 amplification of RML and 22L scrapie strains (10). This difference might be due to the 437 fact that we used an 80 times higher dose of RNase A and that the PMCA reaction was 438 performed in the presence of beads, a condition that was shown to enhance in vitro 439 conversion in general as well as the RNA stimulating effect on in vitro amplification by a 440 mechanism that has yet to be determined (15). 441 However, the extent of reduction of amplification efficacy by RNA depletion varied from 442 one strain to another. The effect of RNA depletion on conversion efficiency could be 443 best appreciated by the serial seed dilution approach. Amplification efficiency was 444 assessed by the number of 6-fold dilutions of the inoculum that could be performed 445 while still allowing in vitro conversion. Even two RML-derived strains, 79A and 139A 446 exhibited a two log difference between them in their response to RNA depletion. 447 Importantly, the sensitivity to RNA depletion was independent of the intrinsic, and strain-448 specific, in vitro conversion capacity. For example, RML or 79A, two strains with high in 449 vitro conversion efficiency, were more RNA sensitive than the less PMCA efficient 450 strains 139A, 303, 726 or 22F. Two strains with similar conversion efficacy, namely 766 451 and 139A, exhibited different responses to RNA depletion. The effect of RNA depletion 452 on ME7 could not be quantitatively assessed due to the low conversion efficiency of this 453 strain. Similar to previous findings with hamster prions (10, 15), we could not find a 454 mouse prion strain that is completely unresponsive to RNA depletion. Reconstitution of 455 the reaction mixtures with brain or liver RNA restored in vitro amplification. We did not 456 observe any difference between the effects of large versus small RNA populations. 457
on June 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

22
Another strain-specific phenomenon was the capacity to utilize polyanions other than 458 endogenous RNA for replication. RML amplification in RNA-depleted substrate was 459 restored by the addition of poly(A), poly(G) or DNA, whereas ME7 amplification could be 460 restored by poly(A), but not poly(G) or DNA. A previous study showed that infectious 461 PrPSc can be generated in vitro from recombinant PrP amplified in the presence of 462 poly(A) and lipids (34). Our results demonstrate a strain-specific ability to utilize a 463 variety of co-factors. It is logical to stipulate that this phenomenon underlies the strain-464 specific effect of RNA depletion with the more promiscuous strains being less affected 465 than those strictly dependent on RNA or certain RNA structures for PrPC to PrPSc 466 conversion. 467 The role of co-factors in prion strain determination, although hypothesized a long time 468 ago (35) has never been specifically addressed. We exploited the fact that RML could 469 amplify in vitro in the absence or quasi-absence of RNA to determine if RNA confers 470 strain-specific properties. We amplified ME7 and RML over 18 and 13 rounds, 471 respectively, to a level where no initial inoculum is present in the sample anymore and 472 inoculated Tga20 as well as wild-type mice with the amplification product. We cannot 473 exclude the presence of undetectable amounts of RNA in the PMCA reactions, but 474 these would be out of proportion with the amounts of newly amplified PrPSc. ME7 PMCA 475 samples from RNA-depleted PMCA conditions were devoid of detectable PrPSc and 476 infectivity, showing that loss of PrPSc was accompanied by a loss of infectivity. In vitro 477 amplified RML under standard and RNA-depleted conditions exhibited similar incubation 478 periods and histopathological features in either Tga20 or C57BL/6 mice. The similarity 479 between the two types of prions was confirmed using the ECPA that allows prion strain 480
on June 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

23
discrimination in cultured cells by exploiting the differential capacity of prion strains to 481 replicate in different cell types and pharmacological conditions (3, 22, 25). Interestingly, 482 we observed an unusual clinical symptom (forelimb paresis) with statistically higher 483 incidence in the group of mice inoculated with RML amplified under RNA-depleted 484 conditions than in the standard PMCA group or the group that received RML brain 485 homogenate. This observation might be an epiphenomenon or might indicate a slight 486 modification of the RML strain characteristics too small to be considered a strain change 487 and be detected by the tools used herein. Recent studies show that prion strains are not 488 homogenous but are populated by “substrains” of prions detectable in vitro. A shift from 489 the main “substrain” to another may account for a transient change of phenotype of a 490 given strain under particular replication conditions (36). Since our PMCA-generated 491 material could not be directly evaluated in the ECPA we cannot rule out the possibility 492 that a “substrain” of RML had been selected in vitro under RNA-depleted conditions and 493 reverted to the original strain upon transmission to mice (23). Nonetheless, our results 494 strongly suggest that RNA is not essential to maintain strain-specific characteristics of 495 prions. 496 Our study suggests that RNA molecules behave like catalysts of in vitro amplification of 497 prions, the extent of the catalytic effect being strain-dependent, and that RNA molecules 498 are not required to specify strain-specific information. Purified PrPSc amplified with 499 recombinant PrP in the absence of addition of mammalian cofactor leads to the 500 formation of infectivity at a relatively low level, as judged by some incomplete 501 transmission rates and variability of survival times (19), which is in good agreement with 502 the concept that RNAs constitute prion replication catalysts. As poly(A) (this study and 503
on June 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

24
(34)) and, for one of two tested strains, poly(G) can reconstitute PMCA efficacy, it will 504 be of high interest to evaluate strain characteristics of prions amplified under conditions 505 of such synthetic catalysts. 506 The alternative hypothesis to that stipulating that a PrP associated co-factor is strain-507 defining is that strain characteristics are supported by differences in PrPSc conformation. 508 Direct experimental evidence of strain-specific differences in PrPSc conformation has 509 been provided (29, 30). Different PrPSc conformations are likely to result in variable PrPC 510 seeding efficiency leading to different PrPSc amplification kinetics in vitro and replication 511 efficiency in vivo, and variable reliance on catalytic activity. Some PrPSc conformations 512 may require highly specific and efficient RNA catalysts to overcome the energy barrier 513 to PrPC conversion, while other PrPSc conformations might be less demanding in the 514 type of polyanionic catalyst required, meaning that “weaker” catalysts might be sufficient 515 for them to trigger PrPC to PrPSc conversion. 516 The question arises as to the cellular source of RNA able to catalyze PrPC to PrPSc 517 conversion. RNA molecules, in particular non-coding RNAs, are present in the 518 extracellular environment and body fluids either associated to proteins (90%) or within 519 multivesicular bodies (MVB) like exosomes (12, 40). Extracellular RNA may interact with 520 PrP present in the outer leaflet of the plasma membrane and catalyze the conversion of 521 PrPC into PrPSc either in cholesterol-rich microdomains (16, 31, 39) or along the 522 endosomal pathway, (2, 24) two sites hypothesized to sustain prion replication. 523 Acknowledgements 524
on June 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

25
We thank Dr. Joaquín Castilla for the gift of the PMCA-generated murine prion strains 525 and for bringing to our attention the use of beads for PMCA reactions. We gratefully 526 acknowledge the expert help of the Animal Resource Center personnel at Scripps 527 Florida. We thank Dr. Weissmann for his insight and advice on the project and for 528 critical reading of the manuscript. We are grateful to Dr. Cervenakova for her comments 529 on the manuscript. 530 531 References 532 1. Bessen, R. A., and R. F. Marsh. 1992. Biochemical and physical properties of 533
the prion protein from two strains of the transmissible mink encephalopathy 534 agent. J. Virol. 66:2096-2101. 535
2. Borchelt, D. R., A. Taraboulos, and S. B. Prusiner. 1992. Evidence for 536 Synthesis of Scrapie Prion Proteins in the Endocytic Pathway. J Biol Chem 537 267:16188-16199. 538
3. Browning, S., C. A. Baker, E. Smith, S. P. Mahal, M. E. Herva, C. A. 539 Demczyk, J. Li, and C. Weissmann. 2011. Abrogation of complex glycosylation 540 by swainsonine results in strain- and cell-specific inhibition of prion replication. J 541 Biol Chem 286:40962-40973. 542
4. Bruce, M. E. 1993. Scrapie Strain Variation and Mutation. Br Med Bull 49:822-543 838. 544
5. Castilla, J., P. Saa, C. Hetz, and C. Soto. 2005. In vitro generation of infectious 545 scrapie prions. Cell 121:195-206. 546
on June 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

26
6. Chen, S. G., D. B. Teplow, P. Parchi, J. K. Teller, P. Gambetti, and L. Autilio-547 Gambetti. 1995. Truncated forms of the human prion protein in normal brain and 548 in prion diseases. J Biol Chem 270:1913-19180. 549
7. Couzin, J. 2004. Biomedicine. An end to the prion debate? Don't count on it. 550 Science 305:589. 551
8. Cronier, S., N. Gros, M. H. Tattum, G. S. Jackson, A. R. Clarke, J. Collinge, 552 and J. D. Wadsworth. 2008. Detection and characterization of proteinase K-553 sensitive disease-related prion protein with thermolysin. Biochem J 416:297-305. 554
9. Deleault, N. R., B. T. Harris, J. R. Rees, and S. Supattapone. 2007. Formation 555 of native prions from minimal components in vitro. Proc Natl Acad Sci U S A 556 104:9741-9746. 557
10. Deleault, N. R., R. Kascsak, J. C. Geoghegan, and S. Supattapone. 2010. 558 Species-dependent differences in cofactor utilization for formation of the 559 protease-resistant prion protein in vitro. Biochemistry 49:3928-3934. 560
11. Dickinson, A. G., G. W. Outram, D. M. Taylor, and J. D. Foster. 1989. Further 561 evidence that scrapie agent has an independant genome., p. 446-459. In L. A. 562 Court, D. Dormont, P. Brown, and D. T. Kingsbury (ed.), Unconventional virus 563 diseases of the central nervous system (Paris 2-6 december 1986). CEA 564 Diffusion, Fontenay-aux Roses, France. 565
12. Dinger, M. E., T. R. Mercer, and J. S. Mattick. 2008. RNAs as extracellular 566 signaling molecules. Journal of molecular endocrinology 40:151-159. 567
13. Doi, S., M. Ito, M. Shinagawa, G. Sato, M. Isomura, and H. Goto. 1988. 568 Western blot detection of scrapie-associated fibril protein in tissues outside the 569
on June 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

27
central nervous system from preclinical scrapie-infected mice. J Gen Virol 570 69:955-960. 571
14. Dron, M., M. Moudjou, J. Chapuis, M. K. Salamat, J. Bernard, S. Cronier, C. 572 Langevin, and H. Laude. 2010. Endogenous proteolytic cleavage of disease-573 associated prion protein to produce C2 fragments is strongly cell- and tissue-574 dependent. J Biol Chem 285:10252-10264. 575
15. Gonzalez-Montalban, N., N. Makarava, R. Savtchenko, and I. V. Baskakov. 576 2011. Relationship between conformational stability and amplification efficiency 577 of prions. Biochemistry 50:7933-7940. 578
16. Goold, R., S. Rabbanian, L. Sutton, R. Andre, P. Arora, J. Moonga, A. R. 579 Clarke, G. Schiavo, P. Jat, J. Collinge, and S. J. Tabrizi. 2011. Rapid cell-580 surface prion protein conversion revealed using a novel cell system. Nat 581 Commun 2:281. 582
17. Jeong, B. H., N. H. Kim, J. K. Jin, J. K. Choi, Y. J. Lee, J. I. Kim, E. K. Choi, 583 R. I. Carp, and Y. S. Kim. 2009. Reduction of prion infectivity and levels of 584 scrapie prion protein by lithium aluminum hydride: implications for RNA in prion 585 diseases. J Neuropathol Exp Neurol 68:870-879. 586
18. Karapetyan, Y. E., P. Saa, S. P. Mahal, G. F. Sferrazza, A. Sherman, N. Sales, 587 C. Weissmann, and C. I. Lasmezas. 2009. Prion strain discrimination based on 588 rapid in vivo amplification and analysis by the cell panel assay. PLoS ONE 589 4:e5730. 590
19. Kim, J. I., I. Cali, K. Surewicz, Q. Kong, G. J. Raymond, R. Atarashi, B. Race, 591 L. Qing, P. Gambetti, B. Caughey, and W. K. Surewicz. 2010. Mammalian 592
on June 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

28
prions generated from bacterially expressed prion protein in the absence of any 593 mammalian cofactors. J Biol Chem 285:14083-14087. 594
20. Lasmézas, C. I., J.-G. Fournier, V. Nouvel, H. Boe, D. Marcé, F. Lamoury, N. 595 Kopp, J.-J. Hauw, J. Ironside, M. Bruce, D. Dormont, and J.-P. Deslys. 2001. 596 Adaptation of the bovine spongiform encephalopathy agent to primates and 597 comparison with Creutzfeldt-Jakob disease: Implications for human health. Proc 598 Natl Acad Sci USA 98:4142-4147. 599
21. Legname, G., I. V. Baskakov, H. O. Nguyen, D. Riesner, F. E. Cohen, S. J. 600 DeArmond, and S. B. Prusiner. 2004. Synthetic mammalian prions. Science 601 305:673-676. 602
22. Mahal, S. P., C. A. Baker, C. A. Demczyk, E. W. Smith, C. Julius, and C. 603 Weissmann. 2007. Prion strain discrimination in cell culture: the cell panel 604 assay. Proc Natl Acad Sci U S A 104:20908-20913. 605
23. Mahal, S. P., S. Browning, J. Li, I. Suponitsky-Kroyter, and C. Weissmann. 606 2010. Transfer of a prion strain to different hosts leads to emergence of strain 607 variants. Proc Natl Acad Sci U S A 107:22653-22658. 608
24. Marijanovic, Z., A. Caputo, V. Campana, and C. Zurzolo. 2009. Identification 609 of an intracellular site of prion conversion. PLoS Pathog 5:e1000426. 610
25. Oelschlegel, A. M., M. Fallahi, S. Ortiz-Umpierre, and C. Weissmann. 2012. 611 The Extended Cell Panel Assay Characterizes the Relationship of Prion Strains 612 RML, 79A, and 139A and Reveals Conversion of 139A to 79A-Like Prions in Cell 613 Culture. J Virol 86:5297-5303. 614
on June 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

29
26. Pan, K. M., M. Baldwin, J. Nguyen, M. Gasset, A. Serban, D. Groth, I. 615 Mehlhorn, Z. Huang, R. J. Fletterick, F. E. Cohen, and S. B. Prusiner. 1993. 616 Conversion of alpha-helices into beta-sheets features in the formation of the 617 scrapie prion proteins. Proc. Natl. Acad. Sci. USA 90:10962-10966. 618
27. Prusiner, S. B. 1982. Novel proteinaceous infectious particles cause scrapie. 619 Science 216:136-144. 620
28. Reed, L. J., and H. Muench. 1938. A simple method of estimating fifty percent 621 endpoints. Amer J Hygiene 27:493-496. 622
29. Safar, J., H. Wille, V. Itri, D. Groth, H. Serban, M. Torchia, F. E. Cohen, and 623 S. B. Prusiner. 1998. Eight prion strains have PrP(Sc) molecules with different 624 conformations. Nat Med 4:1157-1165. 625
30. Smirnovas, V., G. S. Baron, D. K. Offerdahl, G. J. Raymond, B. Caughey, 626 and W. K. Surewicz. 2011. Structural organization of brain-derived mammalian 627 prions examined by hydrogen-deuterium exchange. Nature structural & 628 molecular biology 18:504-506. 629
31. Taraboulos, A., M. Scott, A. Semenov, D. Avrahami, L. Laszlo, S. B. 630 Prusiner, and D. Avraham. 1995. Cholesterol depletion and modification of 631 COOH-terminal targeting sequence of the prion protein inhibit formation of the 632 scrapie isoform. J Cell Biol 129:121-132. 633
32. Wadsworth, J. D., S. Joiner, A. F. Hill, T. A. Campbell, M. Desbruslais, P. J. 634 Luthert, and J. Collinge. 2001. Tissue distribution of protease resistant prion 635 protein in variant Creutzfeldt-Jakob disease using a highly sensitive 636 immunoblotting assay. Lancet 358:171-180. 637
on June 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

30
33. Wang, F., X. Wang, C. G. Yuan, and J. Ma. 2010. Generating a prion with 638 bacterially expressed recombinant prion protein. Science 327:1132-1135. 639
34. Wang, F., Z. Zhang, X. Wang, J. Li, L. Zha, C. G. Yuan, C. Weissmann, and J. 640 Ma. 2012. Genetic informational RNA is not required for recombinant prion 641 infectivity. J Virol 86:1874-1876. 642
35. Weissmann, C. 1991. A ‘unified theory’ of prion propagation. Nature 352:679-643 683. 644
36. Weissmann, C., J. Li, S. P. Mahal, and S. Browning. 2011. Prions on the 645 move. EMBO reports 12:1109-1117. 646
37. Will, R. G., J. W. Ironside, M. Zeidler, S. N. Cousens, K. Estibeiro, A. 647 Alperovitch, S. Poser, M. Pocchiari, A. Hofman, and P. G. Smith. 1996. A 648 new variant of Creutfeldt-Jakob disease in the UK. Lancet 347:921-925. 649
38. Yadavalli, R., R. P. Guttmann, T. Seward, A. P. Centers, R. A. Williamson, 650 and G. C. Telling. 2004. Calpain-dependent endoproteolytic cleavage of PrPSc 651 modulates scrapie prion propagation. J Biol Chem 279:21948-21956. 652
39. Ying, Y. S., R. G. Anderson, and K. G. Rothberg. 1992. Each caveola contains 653 multiple glycosyl-phosphatidylinositol-anchored membrane proteins. Cold Spring 654 Harbor symposia on quantitative biology 57:593-604. 655
40. Zhu, H., and G. C. Fan. 2011. Extracellular/circulating microRNAs and their 656 potential role in cardiovascular disease. American journal of cardiovascular 657 disease 1:138-149. 658
659 Figure Legends 660
on June 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

31
Figure 1 RNA depletion by RNase A impairs in vitro conversion in a strain-661 dependent fashion. A) Serial replication of mouse prion strains is affected by 662 treatment with RNase A. 10% uninfected brain homogenates (substrate) treated with 663 RNase A (A), RNase Inhibitor (“RNase Out”) (O) or left untreated (C) were inoculated 664 with 10% brain homogenates of mice infected with RML, ME7 and 22L (inocula) 665 subjected to the same RNase treatments. These preparations were amplified by several 666 rounds of PMCA. After each round, aliquots were PK digested and immunoblotted with 667 the anti-PrP monoclonal antibody D18. After RNase A treatment, the ME7 PrPSc signal 668 was significantly reduced from round 2 and disappeared after 4 rounds. Amplification of 669 22L prions decreased after 3 rounds in the presence of RNase A. RML was unaffected 670 until round 9. The experiment was done 5 more times until rounds 3, 5, 8 or 10, with 671 comparable results. Molecular weight markers in kilo Dalton (kDa) are indicated on the 672 right side of the western blot panels. 673 Figure 2 The requirement of RNA molecules for in vitro amplification is highly 674 strain-dependent. Brain derived RML, 79A, ME7, 139A, 22L and 22F as well as three 675 in vitro generated prion strains: 303, 766 and 726 were propagated in RNase A treated 676 substrates to evaluate RNA dependency. The calculated reduction of PMCA efficiency 677 under RNA-deficient conditions compared to standard conditions is indicated under 678 each group of western blots. Except for 22F and 726, each strain was tested 2 to 4 679 times; representative blots are shown. “No PMCA” panels show the levels of PrPSc prior 680 to PMCA conversion. PrPSc serial dilutions: 2x10-3, 3.3x10-4, 5.6x10-5, 9.3x10-6, 1.5x10-6, 681 2.6x10-7, 4.3x10-8, 7.1x10-9.C) The relative conversion efficiencies of individual prion 682 strains in normal (open bars) and RNase A treated brain homogenates (black bars) 683
on June 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

32
were calculated from the blots shown in Figure 2A relative to the PrPSc signal observed 684 before PMCA amplification. These results indicate that the sensitivity to RNA depletion 685 is independent of the intrinsic conversion capacity of each strain. 686 Figure 3 Addition of RNA enhances prion in vitro amplification. A) Total RNA 687 enhances ME7 conversion efficiency. 10% normal brain homogenates (NBHs) were 688 treated with RNase A or RNase Out. After treatment, RNase A was inhibited with 689 RNase Out (panels 2 and 3) and aliquots of this mixture where either left untreated 690 (panel 2) or reconstituted by addition of brain-extracted total RNA (panel 3). Serial 691 dilutions of ME7 were performed in these substrates and amplified by PMCA. Panel 1 692 shows the levels of ME7 PrPSc in the dilution series prior to PMCA amplification (10-3, 693 2.5x10-4, 6.3x10-5, 1.6x10-5). The experiment was done 5 times with similar results. B) 694 Dose-dependent reconstitution of ME7 conversion by liver RNA independent of 695 RNA size. Total liver RNA was fractionated into large and small RNA fractions. 10% 696 NBHs were treated with RNase A (panels 1-3, 5 and 6). After treatment, RNase A was 697 inhibited by addition of RNase Inhibitor (panels 1-3 and 6). Increasing concentrations of 698 total, large and small RNA fractions were added to RNA-depleted substrates (panels 1, 699 2 and 3 respectively). All substrates were seeded with various quantities of ME7 prions 700 and amplified by PMCA (panels 1-3, 5 and 6). Panel 4 shows the levels of ME7 PrPSc in 701 the dilution series prior to PMCA amplification (10-4, 2.5x10-5, 6.3x10-6, 1.6x10-6). 702 Molecular weight markers in kilo Dalton (kDa) are shown as reference. –PK lane: 703 Undigested normal brain homogenate used as standard. The experiment was done 2 704 times with similar results. 705
on June 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

33
Figure 4 Strain-specific restoration of PMCA conversion efficiency with different 706 polyanions. A) RML and ME7 conversion efficiencies are restored by adding 707 synthetic poly(A) to the PMCA reaction under RNA-depleted conditions, but only 708 RML conversion is restored by poly(G). RNase A treated substrates were 709 reconstituted with the synthetic polyanions poly(A) (panels 4 and 9) and poly(G) (panels 710 5 and 10) and amplified by PMCA. Panels 1 and 6 show the levels of ME7 and RML 711 PrPSc before conversion. The experiment was done 7 times for RML, 5 times for ME7, 712 with similar results. Molecular weight markers in kilo Dalton (kDa) are shown as 713 reference. The asterisk in panel 2 indicates a hand drawn molecular weight marker. B) 714 RML, but not ME7, conversion efficiency is restored by adding DNA to the PMCA 715 reaction under RNA-depleted conditions. RNA depleted substrates (panels 2 and 7) 716 were reconstituted with salmon sperm DNA (panels 3 and 8). In parallel experiments, 717 DNA was added to normal PMCA substrates (panels 5 and 10) and the amplification 718 yield compared to control PMCA reactions (panels 4 and 9). These preparations were 719 seeded with decreasing concentrations of ME7 and RML prions and amplified by 720 PMCA. The experiment was done once with duplicate samples (one is shown) for RML, 721 twice for ME7. ME7 and RML dilutions are shown in panels 1 and 6 respectively. 722 Molecular weight markers in kilo Dalton (kDa) are shown as reference. 723 Figure 5 Analysis of RML strain characteristics amplified in RNA-depleted 724 reactions. A) Comparison of electrophoretic mobility and quantitation of PrPSc 725 levels in PMCA generated material (round 13) prior to mouse inoculation. Scrapie 726 associated fibers (SAFs) were purified from PMCA samples and brain-derived RML. 727 Serial dilutions of each sample were performed prior to Western blotting to determine 728
on June 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

34
the levels of PrPSc inoculated into mice. Molecular weight markers in kilo Dalton (kDa) 729 are shown as reference. The uninfected brain homogenate (uninfected) is shown as 730 control of the amount of PrPC that is unspecifically precipitated during SAF purification. 731 B) Difference in RML-PRNaseA electrophoretic pattern results from a higher 732 proportion of the PrP C1 fragment. SAF preparations used in A) were deglycosylated 733 and analyzed by Western blot to evaluate the levels of full length and truncated forms of 734 PrP. Densitometric calculations of full length and truncated PrP, shown in the adjacent 735 graph, demonstrate higher levels of C1 in RML-PRNaseA. 736 Figure 6 Survival curves. Representation of the survival times resulting from the 737 inoculation of PMCA material generated in the presence or absence of RNAs. 1 mouse 738 in each group NBHRNaseA and NBH-P died of intercurrent disease. Due to overlapping 739 curves; some data points cannot be observed in the graph. 740 NBHRNaseA: NBH treated with RNase A, NBHRNaseOut: NBH treated with RNase Out, 741 NBH-P: NBH subjected to 18 rounds of PMCA, RML-PRNaseA: RML amplified by PMCA 742 in the presence of RNase A, RML-PRNaseOut: RML amplified by PMCA in the presence of 743 RNase Out, RML-P: RML amplified by PMCA under standard conditions, RML-B: brain-744 derived RML, ME7-PRNaseA: ME7 amplified by PMCA in the presence of RNase A. 745 Figure 7 Comparison of the histopathological features in the brains of mice 746 inoculated with RML prions amplified in vitro in the presence or absence of 747 RNAs. A-B) Hematoxylin-eosin staining. The profile of spongiform degeneration is 748 similar in hematoxylin-eosin stained sagittal sections prepared from C57BL6 mouse 749 brains inoculated with equivalent amounts of RML PrPSc generated by PMCA in the 750
on June 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

35
absence (A) or presence of RNAs (B). C) PrPSc Staining. The PrPSc accumulation 751 pattern in Tga20 mouse brains inoculated with RML PrPSc generated by PMCA in the 752 absence (left panel) or presence (right panel) of RNA was visualized by staining with the 753 anti-PrP monoclonal antibody Bar233 as described in the methods section. 754 Figure 8 Extended Cell Panel Assay. A) PMCA generated material was passaged into 755 Tga20 mice and used to infect PK1, LD9 and R332H11 cell lines subjected to several 756 pharmacological treatments affecting prion replication in a strain-specific fashion. 757 Amplification of RML in the absence of RNA did not change strain properties in the 758 CPA. B) CPA profiles of brain-derived RML (RML-B) ME7 (ME7-B) and 22L (22L-B) 759 strains are shown in comparison. A) and B) represent two different experiments. Cell 760 lines: PK1 (blue), LD9 (purple), R332H11 (green). Prion inhibitors: cur: curcumin (pink, 761 dashed line), swa: swainsonine (light blue, dashed line). The horizontal dotted line 762 indicates the number of positive cells (“spots”) used to calculate the response index (RI) 763 of each cell line to a particular prion strain and pharmacological treatment (shown in 764 Table S1). 765 766 767 768 769 770 771
on June 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

36
Table 1. Survival times of Tga20 mice intracerebrally inoculated with PMCA-772 generated prions 773
774 NBHRNaseA and NBHRNaseOut: NBH treated with RNase A and RNase Out respectively. 775 NBH-P: NBH subjected to 18 rounds of PMCA in the absence of seed. RML-P: PMCA-776 generated RML. RML-PRNaseOut: PMCA-generated RML in the presence of RNase Out. 777 RML-PRNaseA: PMCA-generated RML in the presence of RNase A. RML-B: brain-derived 778 RML. RML-BRNaseA and RML-BRNaseOut: brain-derived RML treated with RNase A and Out 779 respectively. * Dilutions of PMCA-generated inocula are relative to the PMCA sample. 780 Dilutions of brain-derived inocula are relative to the brain. ** dpi: days post inoculation 781
on June 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

37
expressed as Average ± Standard error of the mean. *** The high Standard error of the 782 mean in this group is due to the presence of an outlier mouse with a prolonged survival 783 time of 216 days post inoculation. 784 Table 2. Specific infectivity of PMCA-derived PrPSc generated in the presence or 785 absence of RNA 786
787 (*) Inoculum titer per milliliter of PMCA-generated material (**) Inoculum titer per gram 788 of brain. (†) Specific infectivity of PrPSc molecules present in the inoculum relative to 789 brain-derived RML (RML-B). RML-PRNaseA: PMCA-generated RML in RNA depleted 790 conditions. RML-PRNaseOut: PMCA-generated RML in the presence of RNase Out. 791 Table 3. Survival times of C57BL/6 mice intracerebrally inoculated with RML-P 792 and RML-PRNaseA 793
794 Densitometric calculations of the Western blots shown in Fig. 5A indicated that RML-P 795 contains 3 times more PrPSc than RML-PRNaseA; therefore to inject equal amounts of 796 PrPSc we inoculated a 10-1 dilution of RML-PRNaseA and a 3x10-2 dilution of RML-P. * dpi: 797 days post inoculation expressed as Average ± Standard error of the mean. RML-798
on June 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

38
PRNaseA: PMCA-generated RML in RNA depleted conditions. RML-P: PMCA-generated 799 RML. 800 Table 4. Survival times after secondary transmission of RML-PRNaseA in C57BL/6 801 and Tga20 mice 802
803 1% Brain homogenates of RML-PRNaseA-infected C57BL/6 and Tga20 mice were 804 inoculated intracerebrally in C57BL/6 and Tga20 mice, respectively. * dpi: days post 805 inoculation expressed as Average ± Standard error of the mean. RML-PRNaseA: PMCA-806 generated RML in RNA depleted conditions. 807
on June 12, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from


















