SPINELESS-ARISTAPEDIA: A HOMEOTIC GENE …spineless-aristapedia alleles (called SS~".~,~) were...
13
Copyright 0 1982 by the Genetics Society of America SPINELESS-ARISTAPEDIA: A HOMEOTIC GENE THAT DOES NOT CONTROL THE DEVELOPMENT OF SPECIFIC COMPARTMENTS IN DROSOPHILA GARY STRUHL’ Medical Research Council Laboratory of Molecular Biology, Cambridge CB2 ZQH, England Manuscript received May 10.1982 Revised copy accepted August 16, 1982 ABSTRACT A two-step screen for isolating null mutations of the spineless-aristapedia locus has been performed, and several amorphic mutations, as well as a small deficiency, have been obtained. With the exception of the deficiency, which deletes genes required for viability on either side of the spineless-aristapedia locus, these mutations result in a transformation of only the distal antenna into distal leg, thereby indicating that the spineless-aristapedia gene is required for specifying antennal as opposed to leg development in only the distal portion of the antenna. Because this distal region does not appear to be a developmental compartment, it is probable that the spineless-aristapedia gene, unlike several other homeotic genes, is required far maintaining the correct determined state in a population of cells defined by their relative position, not by their ancestry. TUDIES of cell lineage in Drosophila and Oncopeltus have shown that the S epidermis of the adult insect is subdivided into precisely defined regions (compartments), each formed exclusively by the descendants of a group of founder cells (GARCIA-BELLIDO, RIPOLL and MORATA 1973,1976; LAWRENCE 1973). Since each founder cell gives rise to a clone, a compartment is said to be formed by a polyclone (CRICK and LAWRENCE 1975). Mutations in several homeotic genes are known that transform one or more entire compartments into others (GARCIA-BELLIDO 1975, 1977). For example, both the meso- and metathoracic segments of the fly are subdivided into anterior and posterior compartments (GARCIA-BELLIDO, RIPOLL and MORATA 1973, 1976; STEINER 1976); the bithorax mutation transforms the anterior compartment of the metathorax into the anterior compartment of the mesothorax while leaving the posterior compart- ment unaffected (LEWIS 1963; MORATA and GARCIA-BELLIDO 1976; LAWRENCE, STRUHL and MORATA 1979). Thus, the realms of action of at least some homeotic genes appear to be restricted to particular polyclones. These results led GARCIA- BELLIDO (1975) to propose that the progressive subdivision of the developing insect into polyclones involves the activation of a unique combination of selector genes in each group of founder cells. The selector genes then remain active in all of the descendent cells of the founder group and specify the particular developmental pathway the polyclone follows. I Present address Department of Biochemlstry and Molecular Biology, Harvard Unlverslty, Cambridge, MA 02138 Genetics 102: 737-749 December, 1982
Transcript of SPINELESS-ARISTAPEDIA: A HOMEOTIC GENE …spineless-aristapedia alleles (called SS~".~,~) were...

Copyright 0 1982 by the Genetics Society of America
SPINELESS-ARISTAPEDIA: A HOMEOTIC GENE THAT DOES NOT CONTROL THE DEVELOPMENT OF SPECIFIC
COMPARTMENTS IN DROSOPHILA
GARY STRUHL’
Medical Research Council Laboratory of Molecular Biology, Cambridge CB2 ZQH, England
Manuscript received May 10.1982 Revised copy accepted August 16, 1982
ABSTRACT
A two-step screen for isolating null mutations of the spineless-aristapedia locus has been performed, and several amorphic mutations, as well as a small deficiency, have been obtained. With the exception of the deficiency, which deletes genes required for viability on either side of the spineless-aristapedia locus, these mutations result in a transformation of only the distal antenna into distal leg, thereby indicating that the spineless-aristapedia gene is required for specifying antennal as opposed to leg development in only the distal portion of the antenna. Because this distal region does not appear to be a developmental compartment, it is probable that the spineless-aristapedia gene, unlike several other homeotic genes, is required far maintaining the correct determined state in a population of cells defined by their relative position, not by their ancestry.
TUDIES of cell lineage in Drosophila and Oncopeltus have shown that the S epidermis of the adult insect is subdivided into precisely defined regions (compartments), each formed exclusively by the descendants of a group of founder cells (GARCIA-BELLIDO, RIPOLL and MORATA 1973,1976; LAWRENCE 1973). Since each founder cell gives rise to a clone, a compartment is said to be formed by a polyclone (CRICK and LAWRENCE 1975). Mutations in several homeotic genes are known that transform one or more entire compartments into others (GARCIA-BELLIDO 1975, 1977). For example, both the meso- and metathoracic segments of the fly are subdivided into anterior and posterior compartments (GARCIA-BELLIDO, RIPOLL and MORATA 1973, 1976; STEINER 1976); the bithorax mutation transforms the anterior compartment of the metathorax into the anterior compartment of the mesothorax while leaving the posterior compart- ment unaffected (LEWIS 1963; MORATA and GARCIA-BELLIDO 1976; LAWRENCE, STRUHL and MORATA 1979). Thus, the realms of action of at least some homeotic genes appear to be restricted to particular polyclones. These results led GARCIA- BELLIDO (1975) to propose that the progressive subdivision of the developing insect into polyclones involves the activation of a unique combination of selector genes in each group of founder cells. The selector genes then remain active in all of the descendent cells of the founder group and specify the particular developmental pathway the polyclone follows.
I Present address Department of Biochemlstry and Molecular Biology, Harvard Unlverslty, Cambridge, MA 02138
Genetics 102: 737-749 December, 1982

738 G. STRUHL
Under GARCIA-BELLIDO'S hypothesis, mutations in selector genes would be expected to alter the phenotype of entire compartments. Mutations of the homeotic gene spineless-aristapedia thus pose an interesting puzzle. These recessive mutations lead to a transformation of the distal portion of the antenna into the corresponding distal portion of the mesothoracic leg (BALKASCHINA 1929; LINDSLEY and GRELL 1968). However, lineage studies of the normal antenna have failed to reveal any compartmental segregation between the proximal and distal portions of the antenna (POSTLETHWAIT and SCHNEIDERMAN 1971; MORATA and LAWRENCE 1979). Thus, if the spineless-aristapedia gene is a selector gene, it is necessary to posit that the antenna is subdivided into proximal and distal compartments that have not been detected. Alternatively, it is possible that the available mutations reduce, but do not eliminate, the gene's activity and, hence, do not affect the entire region under its control.
To investigate this puzzle, a two-step screen for isolating null alleles of the spineless-aristapedia locus has been performed. In the first step, a mutation was isolated that in trans to a deficiency for the locus results in a viable and fertile adult having a moderate aristapedia phenotype. This new allele, ssaC3, was then used to screen exhaustively for new alleles of the spineless-aristapedia locus. The phenotypes of these new mutant alleles indicate that the spineless-arista- pedia gene is normally required for specifying the correct determined state in only the distal portion of the antenna. On the basis of these results, as well as previous findings, I conclude that the spineless-aristapedia gene is not involved in the development of a specific compartment and, hence, is not a selector gene.
MATERIALS A N D METHODS
Two-step screen for null olleles of the spineless-aristapedio locus: crosses and genotypes employed
Unless otherwise stated, descriptions of all of the mutations and chromosomal aberrations used in this study can be found in LINDSLEY and GRELL (1968). One chromosomal aberration, ln(3LRJP88 deserves special mention. The 89C-D breakpoint of this inversion has been described as a deficiency of a few bands in the 89C-D region that deletes the spineless-aristapedia locus (E. B. LEWIS, in LINDSLEY and GRELL 1968). However, as reported here, In(3LR)P88 is viable in trans to several different deficiencies which unquestionably delete the spineless-aristapedia locus, as well as neighboring loci on either side. In addition, In(3LR)P88/ss- flies show an extreme spineless- aristapedia phenotype but appear otherwise normal. Consequently, In(3LR)P88 behaves like a point mutation of the spineless-aristapedia gene and, hence, cannot be regarded as a certain deletion of the spineless-aristapedia locus.
Step 1: p"e" males were mutagenized with ethyl methane sulfonate (EMS) and outcrossed to p h pp ssn('' Ubx' e"/TMZ virgin females [see MORATA and LAWRENCE (1979) for a description of SS"~' ] .
p" e"/pb p" ssfZrt Uhx' e t ' progeny showing an aristapedia phenotype were outcrossed to In(3LR)Uhx1", e"/TMI, e'' flies to obtain pp ssac* e"/TMl, e'' males which were then individually tested by outcrossing to pb pp ssnC1 Uhx' e"/TMI virgin females. pp ssac* e''/TMl progeny from single successful test crosses were used to set up balanced stocks of the new alleles. Three new spineless-aristapedia alleles (called S S ~ " . ~ , ~ ) were obtained.
Step 2: st p" e" males were mutagenized with EMS and outcrossed to virgin females from a homozygous, ss"('' stock, and eggs were collected from 11 independent mass matings. st pp elt/ssnC" progeny showing an aristapedia phenotype were each outcrossed to In(3LR)Ubx1"', e'/TM2, e" flies to obtain ten (st pp ss"'*) e"/TMI, e" males which were then individually tested by outcrossing to In(3LR)P88/TMI virgin females. st p p sac* e"/TMI progeny from single successful test crosses were used to set up balanced stocks of the new alleles. A total of 12 new spineless-aristapedia

THE S S " GENE OF DROSOPHILA 739
mutations (called . . I 6 ) were obtained. Of these, ss"c5~10~11 arose from independent mass mating,
an FI mosaic in which one antenna was phenotypically wild type and the other partially transformed into leg.
ss"(6.16 arose from the same mass mating, as did ssnC7.*, S S " ~ ~ ~ ~ ~ : ' and ssRcY,12,14. , SSnC13 arose initially as
Unless otherwise stated, all crosses were performed at 25".
Mutagenesis procedure
Males aged 5-7 days were placed in empty tubes for 6 hr at 25' and then fed with a 0.25% solution of EMS for 24 hr as described by LEWIS and BACHER (1968). Treated males were immediately crossed to virgin females (aged 5-7 days) carrying the appropriate spineless-aristapedia test chromosome. Eggs were collected over a 6-day period, the males having been discarded on day 3. FI progeny were collected until the 14th day after the eggs were laid (care was taken to avoid crowding so that most of the FI progeny emerged by the 10th day after egg laying). The effectiveness of the mutagenesis procedure was assessed by outcrossing mutagenized males to C(l)DX, y f virgin females and comparing the male to female ratio of their progeny with that of the progeny of a control cross using unmutagenized males (see RESULTS).
Studies of mutant phenotype: viability
No distinction was made between degrees of viability. Any two spineless-aristapedia mutations were scored as viable in trans if any trans heterozygous adult progeny emerged after a test cross. In crosses between flies carrying the more extreme spineless-aristapedia mutations, the fused tarsi of the trans heterozygous progeny often became tangled in the pupal cuticle. When necessary, these progeny were pulled free of the pupal case.
In a previous study of ssn mutations, BOWNES, BOURNIAS-VARDIABASIS and SPARE (1979) reported that the SS"(" mutation (designated incorrectly by these authors as ssnCnm) is lethal both when homozygous and in trans to Df(3R)bxd'". However, I find both ssnC1/ssnC1 and ssaC1/Df(3R)bxd'"' flies to be viable. Because both I and BOWNES, BOURNIAS-VARDIABASIS and SPARE obtained the ssRC' mutation from the same source (MORATA and LAWRENCE 1979), their failure to find S S " ( ' ~ / ~ S ~ ~ ' and s~"~'/Df(3R)bxd'" flies can most probably be attributed to different culture conditions.
The aristapedia, spineless, and leg fusion phenotypes were scored under the dissecting micro- scope and, in many cases, under the compound microscope. Specimens were mounted for exami- nation under the compound microscope in a 1.75-g/ml mixture of Canada balsam in methyl salicylate (STRUHL 1981).
Complementation tests between SS"'~ and other mutations and deficiencies in the vicinity of the spineless-aristapedia locus
All possible inter se crosses were performed between flies carrying S S ~ ~ ' ~ , and the deficiencies and chromosomal rearrangements are shown in Table 1. In each case, the rearrangements or deficiencies were maintained in trans to balancer chromosomes carrying dominant and recessive mutations such that it was possible to identify trans heterozygous progeny unambiguously.
RESULTS
A screen for null alleles of the spineless-aristapedia locus A two-step procedure has been used to screen for null alleles of the spineless-
aristapedia locus. In the first step, appropriately marked males were mutagen- ked with EMS and outcrossed to females carrying an extreme mutant allele of the locus, ssaC1 (MORATA and LAWRENCE 1979). Of 1800 F1 progeny carrying the ssQc'-bearing chromosome, three showed an aristapedia phenotype. On subse- quent outcrosses, each of these three flies segregated a new spineless-aristapedia mutation. Each of these mutations ( ssaC2, ssaC3, and ssaC4) was outcrossed to stocks carrying In(3LR)P88, a third chromosome carrying an extreme spineless- aristapedia mutation (LINDSLEY and GRELL 1968; see MATERIALS AND METHODS),

740 G. STRUHL
TABLE 1
Chromosomal rearrangements and deficiencies employed
Designation Description
ln(3LR)P88
Tp(3) bx d I"'
Df(3R)bxd'"''
T(2;3)P10
Df(3R)PlO
Tp(3)sbd"'
Df(3R)s bd'"
T(1;3)P115
Df(3R)Pll5
Tp(3)Pl46
Dp(3;3)P146 Df(3R)P115
Df(3R)PIO Dp(3;3)sbdr"
Df(3R)Ubx"
Inversion with breakpoints in 61A and 89C-D. Deficient for bands in 89C-D, mutant for SS" (LEWIS, in LINDSLEY and GRELL 1968).
Transposition carrying the region between 89B6 and 89EZ to 66C. Mutant for bxd (LEWIS, in LINDSLEY and GRELL 1968).
Deficiency of 89B6 to 89E2 derived from Tp(3)bxd'". Deletes the SS",
bx and Ubx loci and is mutant for bxd (LEWIS, in LINDSLEY and GRELL 1968).
89E1-2 to 29A-C on the 2nd chromosome. Mutant for iab-2 (LEWIS 1978, 1980).
ssn, bx, Ubx, bxd and pbx loci and is mutant for iab-2 (LEWIS 1978, 1980).
Transposition carrying the region from 89B5 to the end of 89C to 91B. Mutant for sbd (LEWIS 1980).
Deficiency of 89B5 to the end of 89C derived from Tp(3)sbd'". De- letes the s~ ' ' locus and is mutant for sbd (LEWIS 1980).
Insertional translocation carrying the region between 89B and 89E5- 6 to the X chromosome (LEWIS, in MORATA and GARCIA-BELLIDO 1976).
Deficiency of 898 to 89E5-6 derived from T(1;3)P115. Deletes the SS"
locus and the entire bjthorax complex (BX-C) (LEWIS, in MORATA and GARCIA-BELLIDO 1976).
Transposition carrying a portion of the 3rd chromosome containing the entire BX-C to the left arm of chromosome 3 (homozygous viable)."
Df(3R)P115 is carried by Dp(3;3)P146 derived from Tp(3)P146. De- letes the s ~ ' locus.
Df(3R)PlO is carried by Dp(3;3)sbdJo4. Deletes bx, Ubx, bxd, and pbx, and is mutant for jab-2.
Deficiency of the region from 89D1-2 to 89E1-2. Deletes part of the EX-C including bx, Ubx, bxd, pbx and iab-2 (LEWIS 1978, 1980).
Insertion translocation carrying the region between 89C1-2 and
Deficiency of 89C1-2 to 89El-2 derived from T(2;3)P10. Deletes the
Synthetic deficiency in which part of the region deleted by
Synthetic deficiency in which part of the region deleted by
Tp(3)P146 is a previously unpublished transposition kindly provided by E. B. LEWIS.
and Df(3R)bxd loo in which the spineless-aristapedia locus as well as neighboring loci on either side are completely deleted. Of the three mutations, ssaC3 showed the weakest aristapedia phenotype when in trans to either In(3LR)P88 or Df(3R)bxd". ssUC3/In(3LR)P88 and ssUC3/Df(3R)bxd" flies are fully viable and fertile; moreover, virtually all flies of this genotype express a moderate arista- pedia phenotype (e.g., Figure 1). The ssaC3 mutation was, therefore, chosen for the second step of the screening procedure-an exhaustive F1 screen for new mutations of the spineless-aristapedia locus. In this step, appropriately marked males were mutagenized with EMS and outcrossed to females homozygous for the ssaC3 mutation. Of 49,950 F1 progeny obtained, 30 showed a moderate aristapedia phenotype. Of these 30 progeny, four were sterile, 14 segregated

THE SS" GENE OF DROSOPHILA 741
FIGURE 1.-A. Phenotypically normal antennae of a fly homozygous for the ssoCI mutation. B. Antennae of a ssf""/In(3LR)P88 fly showing a characteristic moderate aristapedia phenotype. The proximal portion of the arista is swollen and carries bracted bristles Characteristic of the normal tarsus: similar bracted bristles are also found on the distal portion of the third antennal segment. C. Fully formed, aristapcdia tarsi of an ssf'/In(3LR)P88 fly. Note that thc arista and part of the third antennal segment on each side arc transformed into a well-formed distal leg carrying two claws as well as three to four tarsal segments. Note also that the thickness and length of the bristles on the proximal portions of the antenna are reduced relative to B and A. ( X l l O ) .
only ssac3 and ss+ on subsequent outcrosses, and 12 gave rise to new spineless- aristapedia mutations (called SS"~"*'*. . ." ). The mutagenesis was independently assessed by outcrossing some of the mutagenized males to compound X females. The ratio of male to female progeny in this cross (404:766) compared with the ratio of males to females observed in a control cross in which the parental males were not mutagenized (741:594) indicates an average mutation frequency of

742 G. STRUHL
approximately one lethal hit per X chromosome. Under these conditions, the frequency with which an average gene is expected to mutate to a lethal state is on the order of 1 x IO-" [e.g., O'BRIEN and MACINTYRE (1978)l.
Analysis of mutant phenotypes Males carrying the new spineless-aristapedia alleles, ssa or ssaC1 in trans to
the TM1 balancer chromosome were outcrossed both to females carrying In(3LR)P88 and to females carrying Df(3R)bxd". Progeny from all crosses to In(3LR)P88 were allowed to develop both at 17' and 25'. In addition, all possible crosses were performed between stocks carrying the 12 mutant alleles isolated in step 2 of the screening procedure (i.e., S S ~ ~ ~ ~ ~ ~ ). The results obtained may be summarized as follows.
All of the mutant alleles tested, with the exception of S S ~ ~ ' ~ , were viable (see MATERIALS AND METHODS) in trans both to In(3LR)P88 and to Df(3R)bxd" and showed an aristapedia phenotype. Moreover, the phenotype of flies carrying each of the new alleles in trans to ln(3LRjP88 did not depend appreciably on whether development had occurred at 17' or 25'. In addition, all possible trans combinations of the ssaC5,'. alleles were viable and aristapedia in phenotype.
Flies carrying any one mutant allele in trans to Df(3R)bxdlW showed little phenotypic variation; also, they appeared similar, if not identical, to flies carrying the same mutant allele in trans to In(3LR)P88. However, the character- istic mutant phenotypes caused by the different mutant alleles were not all identical. The ssuC:3 allele produced the weakest phenotype partially transform- ing the distal antenna into a distal tarsus but leaving the fly otherwise unaffected (Figure 1). The most extreme phenotype was exhibited by the S S ~ ~ ~ . ~ ~ ~ ~ ~ * ~ ~ , ~ ~ ~ ~ ~
alleles. When in trans to either In(3LR)P88 or Df(3R)bxd1", or in trans to each other, these alleles produced a complete transformation of the distal antenna to distal tarsus, an extreme and consistent spineless phenotype on bristles all over the fly, and fusions of most of the tarsal segments of each of the three legs and also of the transformed aristapedia tarsus (Figures 2 and 3). Alleles that pro- duced intermediate phenotypes (e.g., ssa and ssaC3) affected the extents of the aristapedia, spineless and leg fusion phenotypes in a coordinate fashion (e.g., Figures 1-3).
Thus, all of the new mutations, whether in trans to In(3LR)P88 or in trans to each other, were viable and resulted in a homeotic transformation of at most the distal portion of the antenna into a distal tarsus. In addition, all but one of the new mutations were viable in trans to Df(3R)bxd"'O.
Analysis of the ssdC1:' mutation Unlike all of the other spineless-aristapedia mutations obtained in this study,
the ssuC'5 mutation is lethal in trans to Df(3R)bxd'"". Lethality could result if the mutation were a deficiency that deletes not only the spineless-aristapedia SSaCIS
locus, but also neighboring genes required for viability. To test this possibility, all possible inter se crosses were performed between the S S ~ ~ ~ ~ mutation and several deficiencies and chromosomal rearrangements in the vicinity of the spineless-aristapedia locus (See MATERlALS AND METHODS, Table 1). The inter-
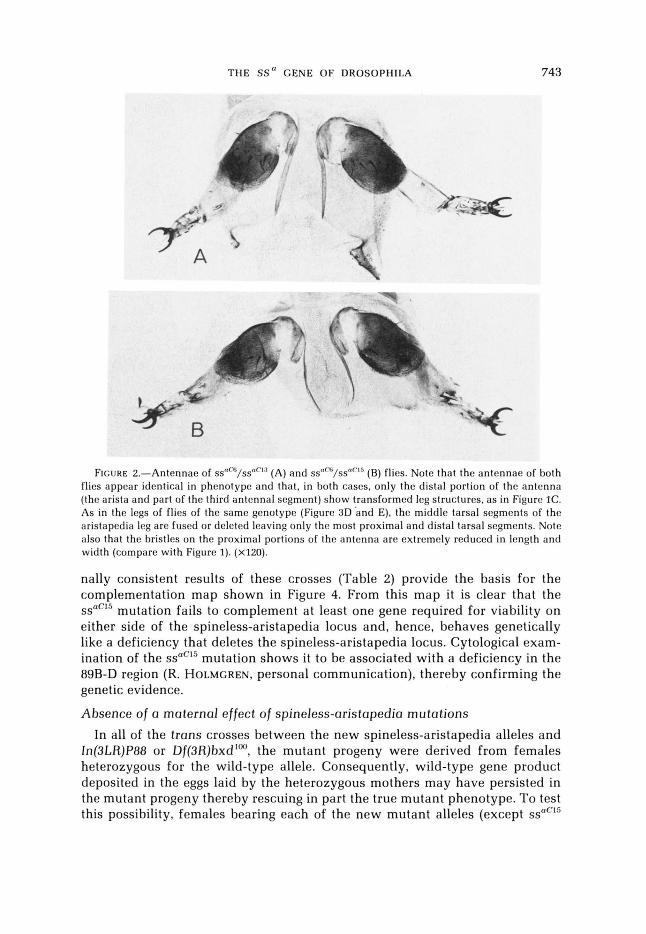
THE SS" GENE OF DROSOPHILA 743
FIGURE 2.-Antennae of S S " ~ / S S " ~ ' ~ (A) and S S " ~ / S S " ~ ' ~ (B) flies. Note that the antennae of both flies appear identical in phenotype and that, in both cases, only the distal portion of the antenna (the arista and part of the third antennal segment) show transformed leg structures, a s in Figure IC. As in the legs of flies of the same genotypc (Figure 3D and E), the middle tarsal segments of the aristapedia leg are fused or deleted leaving only the most proximal and distal tarsal segments. Note also that the bristles on the proximal portions of the antenna are extremely reduced in length and width (compare with Figure 1). ( ~ 1 2 0 ) .
nally consistent results of these crosses (Table 2) provide the basis for the complementation map shown in Figure 4. From this map it is clear that the
mutation fails to complement at least one gene required for viability on either side of the spineless-aristapedia locus and, hence, behaves genetically like a deficiency that deletes the spineless-aristapedia locus. Cytological exam- ination of the s~~~~~ mutation shows it to be associated with a deficiency in the 89B-D region (R. HOLMGREN, personal communication), thereby confirming the genetic evidence.
Absence of a maternal effect of spineless-aristapedia mutations In all of the trans crosses between the new spineless-aristapedia alleles and
In(3LR)P88 or Df(3R)bxdIcm, the mutant progeny were derived from females heterozygous for the wild-type allele. Consequently, wild-type gene product deposited in the eggs laid by the heterozygous mothers may have persisted in the mutant progeny thereby rescuing in part the true mutant phenotype. To test this possibility, females bearing each of the new mutant alleles (except ssaCtS
SSaC15

744 G. STRUHL
FIGURE 3.-Second legs of S S " ~ / S S ~ ' ~ (A) and ssnm3/In(3LR)P88 (B) flies are phenotypically normal. However, legs of ssn/In(3LR)P88 (C) flies show an apparent fusion of the basitarsis with the remaining tarsal segments, and in legs of both S S " ~ / S S " ~ ' ~ (D) and S S ~ ~ / S S " ( " ~ (E) flies. all of the tarsal segments between the basitarsus and the most distal tarsal segment carrying the claws appear to be deleted or fused. Arrows point to the junctions between the most proximal tarsal segment (the basitarsus) and the tibia. (X150).
and S S " " ~ ) in trans to In(3LR)P88 were fertilized by In(3LR)P88/TMz males. In every case, the phenotype of mutant s~"~*/In(3LR)P88 progeny was indistin- guishable from that of the mutant mothers. Moreover, the ss+ (TMI) progeny developed normally. Thus, mutations of the spineless-aristapedia locus do not show a maternal effect.
DISCUSSION
My main purpose has been to test whether the spineless-aristapedia gene has the properties expected of a selector gene. The experimental approach has been to obtain new mutations of the locus under conditions favorable for isolating null alleles and then to study the phenotypes of these mutations during both oogenesis and zygotic development. It is, therefore, necessary to assess whether the screening procedure has produced null mutations and, if so, whether they behave like null mutations of a selector gene.

THE S S a GENE OF DROSOPHILA 745
TABLE 2
Complementation analysis of the ssaCl5 mutation using deficiencies and chromosomal rearrangements in or near the spineless-oristapedia locus"
SSaC5-14,16
In(3LR)P88 Tp(3)sbd" Df(3R)sbdjW Tp(3) bxd" Df(3R) bxd" T(1;3)P115 Df(3RlP115 Dp(3;3)P146 Df(3R)P115 T(2;3)P10 Df(3R)PlO Df(3R)PlO Dp(3;3)sbd" Df(3R) Ubx"
s s S S L V V L L S L L V V V V L L S V L L L L V V L V L L L S V L L L L L L S V L V L L L L v v v v v v v L v L L S V L L L V L L L L L V V V L L V L V L L L V V V V L L V L V L L L L
" See Table 1 for descriptions of the deficiencies and chromosomal rearrangements used. V = trans heterozygous progeny are viable: S = trans heterozygous progeny are viable and show an extreme spineless-aristapedia phenotype (e.g., Figures 2 and 3); L = trans heterozygous progeny are lethal (no distinction was made between degrees of viability: trans heterozygous progeny were scored as viable if any adult progeny emerged after a test cross and as lethal if no adult progeny emerged).
In Drosophila, the only mutations that can be said with certainty to be null alleles of a given gene are mutations that can be shown biochemically to result in complete absence of the gene or its product or deficiencies that delete not only the locus of interest but also well-defined loci on either side. At present, neither the spineless-aristapedia gene nor its product can be assayed biochem- ically. Consequently, only deficiencies that are sufficiently large to delete genes on either side of the spineless-aristapedia locus qualify as unambiguous null mutations. Unfortunately, deficiencies that delete more than a few genes are generally cell lethal when homozygous (GARCIA-BELLIDO and RIPOLL 1978) and, hence, of little value in studying the null phenotype. The two-step screening procedure used in this study has allowed an exhaustive screen for spineless- aristapedia mutations under conditions in which a null allele of the locus (e.g., Df(3R)bxd", which is known to be deficient for the spineless-aristapedia locus (LEWIS, in LINDSLEY and GRELL 1968)) would be viable, fertile and detectable. Thus, assuming that the spineless-aristapedia gene is mutable to a null state by the action of the mutagen EMS, one might expect the collection of mutations obtained to include one, if not several, null alleles of the locus. Several of the new spineless-aristapedia mutations appear to have an identical phenotype whether in trans to each other or in trans to either In(3LRIP88 or Df(3R)bxd'"

746 G . STRUHL
ln(3LRIP88 0
ss aC15 - - - - - - - .-. Df(3R)sbd' O4 0 *
Df(3R)PIO Dp(3;3)sbd 0 0
DD(3;3)P146 Df(3R)PI 15 - Df(3R)PI 15 0
Df(3R)bxd O0 0 0
Df(3R)PlO 0 0
Df(3R)Ubx' O9
+ sbd
+ ssa
- ax-c
FIGURE 4.-Summary of the results of all possible inter se crosses performed between SS"~" and deficiencies and chromosomal rearrangements in the vicinity (see MATERIALS A N D METHODS and Tables 1 and 2). Open and closed circles signify breakpoints that are viable and lethal, respectively, when in trans to deficiencies for that region (e.g., In(3LR)P88 was viable in trans to all deficiencies tested, whereas T(1;3)P115 was lethal in trans to Df(3R)~bd"'~, Df(3R)bxd"" and SS"'~~, indicating that the breakpoint of T(1;3)P115 closest to the centromere of chromosome 3 is associated with a recessive lethal mutation). Any two deficiencies drawn as overlapping (e.g., Df(3R)PZO D~(3;3)sbd'"~ and ssnCI5) were lethal in trans, whereas nonoverlapping deficiencies (e.g., Df(3R)Ubx IO9 and SS""~)
were viable in trans. The region that is deleted by ssoc15 is indicated by a solid line; dotted lines indicate the possible extent of this deficiency on either side. The positions of stubbloid (sbd), spineless-aristapedia (ss") and the bithorax-complex (BX-C) are indicated by arrows or a bracket.
(e.g., Figure 2). Moreover, this phenotype is the most extreme phenotype in terms of the extent to which the normal pattern is altered. These extreme mutant alleles, therefore, behave as amorphic mutations of the locus and, hence, are good candidates for being null alleles:Indeed, one of the amorphic mutations obtained, ssaC15, is a cytologically visible deficiency that deletes not only the spineless-aristapedia locus, but also genes required for viability on either side. Thus, the screening procedure allowed the recovery of at least one definitive null allele.
Assuming the most extreme spineless-aristapedia mutations to be null alleles, one may ask what the null phenotype tells us about the normal role of the wild- type gene. There are three aspects of the mutant phenotype: (1) the spineless phenotype, in which bristles all over the body of the fly are greatly reduced in size, (2) the aristapedia phenotype, in which the distal portion of the antenna including the arista and part of the third antennal segment is transformed into a distal leg, and (3) the leg fusion phenotype, in which the tarsal segments of all three legs as well as the transformed aristapedia leg are fused. This pleiotropic phenotype can be interpreted in two ways. First, the spineless-aristapedia locus might contain only one gene function which when mutated produces the pleiotropic phenotype. Second, the locus might contain several gene functions, and mutations in the aristapedia function would prevent the normal expression

THE S S a GENE O F DROSOPHILA 747
of the remaining functions. The possibility that the locus is complex is supported by the existence of mutations such as spineless, which causes an extreme spineless phenotype but has no effect on the development of the legs or antennae, and spineless-variegated, which causes an aristapedia phenotype, but variegates for the spineless phenotype (see LINDSLEY and GRELL 1968).
Clearly, if there is only one genetic function in the locus, then the extent of the mutant spineless phenotype indicates that this function must be active in all tissues forming bristles of the adult fly. Because an essential property of a selector gene is that it is active in some but not other polyclones to specify a particular pathway of development, this interpretation of the locus precludes its being a selector gene. However, if the locus contains a specific aristapedia function that cannot be mutated independently from other gene functions, then one may still ask whether or not this function has the properties expected of a selector gene. The most extreme spineless-aristapedia mutations transform only the distal portion of the third antennal segment into a distal tarsus. However, cell lineage studies of the eye-antenna indicate that a lineage restriction does not separate the cells forming this distal region from the remaining cells of the antennal disk at least until the middle of the third larval instar (POSTLETHWAIT and SCHNEIDERMAN 1969; MORATA and LAWRENCE 1979). Thus, if the boundary between the transformed and untransformed regions of the antenna corre- sponds to a lineage restriction, it must be one that is formed late in the third larval instar, just prior to the end of proliferative growth. If this is so, the aristapedia gene would not be expected to function prior to the segregation of the antenna into these proximal and distal compartments. However, three lines of evidence suggest that the aristapedia gene is normally functioning at least as early as the middle of the third larval instar. First, the eye-antennal disks of homozygous ssa larvae begin to show an aberrant phenotype approximately 24 hr after the beginning of the third larval instar (VOGT 1946), which normally lasts 72 hr. Second, studies of temperature-sensitive spineless-aristapedia mu- tations have shown that the phenocritical period occurs during the first half of the third instar (VILLEE 1943; GRIGLIATTI and SUZUKI 1971). Third, studies of somatic clones that are homozygous for spineless-aristapedia mutations in otherwise heterozygous flies have shown that clones generated after the mid- third larval instar develop normally (POSTLETHWAIT and GIRTON 1974). All of these results indicate that the gene is active during and possibly before the middle of the third larval instar. Consequently, the aristapedia gene function appears to be required in the distal portion of the antennal disk at a time when no lineage segregation separates antennal imaginal cells into proximal and distal polyclones.
Thus, the phenotype of amorphic alleles of the spineless-aristapedia locus argues strongly that the locus does not contain a selector gene responsible for specifying an antennal vs. leg developmental pathway. Instead, it appears that the gene is normally required for maintaining the correct determined state in a subset of antennal cells related by position, but not by cell ancestry. The spineless-aristapedia gene, therefore, provides an example of a homeotic gene that does not appear to control the development of specific compartments.

748 G . STRUHL
I thank P. A. LAWRENCE for his excellent advice and persistent encouragement, R. HOLMCREN for assessing the cytology of the S S ~ ' ' ' ~ mutation and E. B. LEWIS and G. MORATA for generously providing stocks and information. I also thank the United States National Science Foundation, the Nuffield Foundation, the Medical Research Council of Great Britain and the Thomas C. Ushcr Fund for financial support.
LITERATURE CITED
BALKASCHINA, E. J , , 1929
BOWNES, M., N. BOURNIAS-VARDIABASIS and W. J , SPARE, 1979
CRICK, F. H. C. and P. A. LAWRENCE, 1975
Ein Fall der Erbhomoosis (die Genovariation "aristapedia") bei Drosophila
Genetic analysis of the spineless-
Compartments and polyclones in insect development.
Genetic control of wing disc development in Drosophila. Ciba Found.
melanogaster. Wilhelm Roux' Arch. Entw-Mech. Org. 115: 448-463.
aristapedia homeotic mutants of Drosophila melanogaster. Mol. Gen. Genet. 174: 67-74.
Science 189: 340-347.
GARCIA-BELLIDO, A., 1975 Symp. 2 9 161-182.
GARCIA-BELLIDO, A., 1977
GARCIA-BELLIDO, A. and P. RIPOLL, 1978
Homeotic and atavic mutations in insects. Am. Zool. 17 613-629.
The number of genes in Drosophila melanogaster. Nature
Developmental compartmentalization of the
Developmental compartmentalization in the
Temperature sensitive mutations in DrosophiJa melanogas-
A clonal analysis of segment development in Oncopeltus (Hemiptera). J .
Bristle patterns and compartment boundaries
273: 399-400.
GARCIA-BELLIDO, A., P. RIPOLL and G. MORATA, 1973
GARCIA-BELLIDO, A., P. RIPOLL and G. MORATA, 1976
GRIGLIATTI, T. and D. T. SUZUKI, 1971
LAWRENCE, P. A., 1973
LAWRENCE, P. A., G. STRUHL and G. MORATA, 1979
LEWIS, E. B., 1963
LEWIS, E. B., 1978
LEWIS, E. B., 1980
LEWIS, E. B. and F. BACHER, 1968 Methods of feeding ethyl methanesulfonate (EMS) to Drosophila
LINDSLEY, D. L. and E. L. GRELL, 1968 Genetic variations of Drosophila melanogaster. Carnegie
MORATA, G. and A. GARCIA-BELLIDO, 1976 Developmental analysis of some mutants of the bithorax system of Drosophila. Wilhelm Roux' Arch. 179: 125-143.
MORATA, G. and P. A. LAWRENCE, 1979 Development of the eye-antenna imaginal disc of Dro- sophila. Dev. Biol. 70: 355-371.
O'BRIEN, S. J. and R. J. MACINTYRE, 1978 Genetics and biochemistry of enzymes and specific proteins of Drosophila. pp. 395-551. Io: The Genetics and Biology of Drosophila, Vol. Za, Academic Press, New York.
Development in genetic mosaics of aristapedia, a homoeotic mutant of Drosophila melanogaster. Genetics 76: 767-774.
A clonal analysis of develapment in Dro- sophila melanogaster: morphogenesis, determination, and growth in the wild-type antenna. Dev. Biol. 24: 477-519.
wing disk of Drosophila. Nature (New Biol.) 245: 251-253.
dorsal mesothoracic disc of Drosophila. Dev. Biol. 48: 132-147.
ter. VIII. The homeotic mutant, SS""~. Proc. Natl. Acad. Sci. USA 68: 1307-1311.
Embryol. Exp. Morphol. 30: 681-699.
in the tarsi of Drosophila. J. Embryol. Exp. Morphol. 51: 195-208.
Genes and developmental pathways. Am. Zool. 3: 33-56.
A gene complex controlling segmentation in Drosophila. Nature 276 565-570.
New mutants: report of E. B. Lewis. Drosophila Inform. Serv. 55: 207-208.
males. Drosophila Inform Serv. 43: 193.
Inst. Wash. Publ. 627.
POSTLETHWAIT, J. H. and J. R. GIRTON, 1974
POSTLETHWAIT, J. H. and H. A. SCHNEIDERMAN, 1971

THE SSa GENE OF DROSOPHILA 749
STEINER, E., 1976 Establishment of compartments in the developing leg imaginal discs of Drosoph-
STRUHL, G., 1981 Anterior and posterior compartments in the proboscis of Drosophila. Dev. Biol.
VILLEE, C., 1943 Phenogenetic studies of the homoeotic mutants of Drosophila melanogaster. 1.
VOGT, M., 1946 Zur labilen Determination der Imaginalscheiben von Drosophila. 11. Die Umwan-
Corresponding editor: R. K. HERMAN
ila melanogaster. Wilhelm Roux’ Arch. 180: 9-30.
84: 372-385.
The effects of temperature on the expression of aristapedia. 1. Exp. Zool. 93: 75-98.
dlung prasumptiven Fuhlcrgewebes in Beingewebe. Biol. Zentralb. 65: 239-254.