
Species Distribution Modeling for Arid Adapted Habitat ...
124
Stephen F. Austin State University Stephen F. Austin State University SFA ScholarWorks SFA ScholarWorks Electronic Theses and Dissertations Winter 12-15-2020 Species Distribution Modeling for Arid Adapted Habitat Species Distribution Modeling for Arid Adapted Habitat Specialists in Zion National Park Specialists in Zion National Park Sam Driver Stephen F Austin State University, [email protected] Chris M. Schalk Stephen F Austin State University, [email protected] Daniel Unger Stephen F Austin State University, [email protected] David Kulhavy Stephen F Austin State University, [email protected] Follow this and additional works at: https://scholarworks.sfasu.edu/etds Part of the Desert Ecology Commons, Environmental Studies Commons, Geomorphology Commons, Natural Resources and Conservation Commons, and the Other Earth Sciences Commons Tell us how this article helped you. Repository Citation Repository Citation Driver, Sam; Schalk, Chris M.; Unger, Daniel; and Kulhavy, David, "Species Distribution Modeling for Arid Adapted Habitat Specialists in Zion National Park" (2020). Electronic Theses and Dissertations. 354. https://scholarworks.sfasu.edu/etds/354 This Thesis is brought to you for free and open access by SFA ScholarWorks. It has been accepted for inclusion in Electronic Theses and Dissertations by an authorized administrator of SFA ScholarWorks. For more information, please contact [email protected].
Transcript of Species Distribution Modeling for Arid Adapted Habitat ...

Stephen F. Austin State University Stephen F. Austin State University
SFA ScholarWorks SFA ScholarWorks
Electronic Theses and Dissertations
Winter 12-15-2020
Species Distribution Modeling for Arid Adapted Habitat Species Distribution Modeling for Arid Adapted Habitat
Specialists in Zion National Park Specialists in Zion National Park
Sam Driver Stephen F Austin State University, [email protected]
Chris M. Schalk Stephen F Austin State University, [email protected]
Daniel Unger Stephen F Austin State University, [email protected]
David Kulhavy Stephen F Austin State University, [email protected]
Follow this and additional works at: https://scholarworks.sfasu.edu/etds
Part of the Desert Ecology Commons, Environmental Studies Commons, Geomorphology Commons,
Natural Resources and Conservation Commons, and the Other Earth Sciences Commons
Tell us how this article helped you.
Repository Citation Repository Citation Driver, Sam; Schalk, Chris M.; Unger, Daniel; and Kulhavy, David, "Species Distribution Modeling for Arid Adapted Habitat Specialists in Zion National Park" (2020). Electronic Theses and Dissertations. 354. https://scholarworks.sfasu.edu/etds/354
This Thesis is brought to you for free and open access by SFA ScholarWorks. It has been accepted for inclusion in Electronic Theses and Dissertations by an authorized administrator of SFA ScholarWorks. For more information, please contact [email protected].

Species Distribution Modeling for Arid Adapted Habitat Specialists in Zion Species Distribution Modeling for Arid Adapted Habitat Specialists in Zion National Park National Park
This thesis is available at SFA ScholarWorks: https://scholarworks.sfasu.edu/etds/354


SPECIES DISTRIBUTION MODELING FOR ARID ADAPTED
HABITAT SPECIALISTS IN ZION NATIONAL PARK
by
Samuel Macon Driver, B.S.
Presented to the Faculty of the Graduate School of
Stephen F. Austin State University
In Partial Fulfillment
of the Requirements
For the Degree of
Master of Science
Stephen F. Austin State University
December 2020

SPECIES DISTRIBUTION MODELING FOR ARID ADAPTED
HABITAT SPECIALISTS IN ZION NATIONAL PARK
by
Samuel Macon Driver, B.S.
APPROVED:
__________________________________________
Dr. Daniel R. Unger, Committee Chair
__________________________________________
Dr. David L. Kulhavy, Committee Member
__________________________________________
Dr. Christopher M. Schalk, Committee Member
__________________________________
Pauline M. Sampson, Ph.D.
Dean of Research and Graduate Studies

i
ABSTRACT
The Arizona toad (Anaxyrus microscaphus) and Jones’ waxy dogbane
(Cycladenia humilis var. jonesii) are habitat specialists with historical ranges in the desert
southwest and specifically, Zion National Park (ZION). The machine learning method,
MaxEnt, constructed species distribution models (SDMs) in ZION for the two study
species at 30 m and 900 m spatial resolutions using climate, topographic, and remotely
sensed data. Additionally, 900 m forecasting models were constructed to observe the
shifts in suitable habitat for the years 2050 and 2070, based off two representative
concentration pathway scenarios. Results indicate promising predictive power for both
high resolution models (30m) for C. humilis var. jonesii and A. microscaphus with area
under curve (AUC) test analysis of 0.715 and 0.810, respectively. Forecasting models
displayed decreasing suitability for A. microscaphus with both climate scenarios applied
to the model. However, C. humilis var. jonesii habitat increased with future scenarios
applied to the MaxEnt models.

ii
ACKNOWLEDGEMENTS
I’d first like to express my gratitude to Stephen F. Austin State University for
allowing me to develop, pursue, and achieve my academic goals. To all my professors in
the environmental and spatial science divisions at the SFA Forestry Department, I am
beyond grateful for all that you’ve helped me accomplish. To my thesis advisor, Dr.
Daniel Unger, thank you for equipping me with the tools to succeed in my future career
field. Thank you to my thesis committee member, Dr. David Kulhavy, for your guidance
and steadfast belief in me. To my graduate representative, Dr. Yanli Zhang, your intro
GIS class was the first I attended at SFA and your advanced GIS class happened to be my
last at SFA; thank you for keeping my intellectual curiosity up from beginning to end.
And how could I forget my thesis committee member, Dr. Christopher Schalk, thank you
for continuously checking in on my progress and staying on me to learn the ecology
behind species modeling. “Bonesaw is ready!!!”
Special thank you to Dr. Kenneth Farrish and Mary Ramos. Thank you, Shauna,
Adam, and Aven for taking me in for the summer in beautiful Zion National Park while
also planting the seed for this research project. I’ll cherish our adventures for a lifetime
and can’t wait to make more.
Above all, I will never be able to thank my family enough. You constantly believe
in my dreams and have supported me through all my life endeavors. To my brother,

iii
Daniel, thank you for cookin’ up gumbos, smoking briskets, and playing that squeezebox
till them roosters’ crow, and above all, always providing the brightest of outlooks when
the odds seemed stacked up against me. Laissez les bons temps rouler! To my mother and
father, Margaret and Paul, thank you for allowing your adult son to park and live in an
ugly RV for two years in your front yard. On a serious note, words don’t do justice for
how grateful I am to have two amazing role models to always look up to, thank you for
everything.
“The only way to get there son is one mile at a time. This road’s crooked
steep and rocky as she winds.”
– Uncle Steve Hartz

iv
TABLE OF CONTENTS
ABSTRACT ......................................................................................................................... i
ACKNOWLEDGEMENTS ................................................................................................ ii
TABLE OF CONTENTS ................................................................................................... iv
LIST OF FIGURES .......................................................................................................... vii
LIST OF TABLES ........................................................................................................... xiii
INTRODUCTION ...............................................................................................................1
OBJECTIVES ......................................................................................................................7
LITERATURE REVIEW ....................................................................................................8
Zion National Park .......................................................................................................... 8
Arizona Toad (Anaxyrus microscaphus) ....................................................................... 10
Jones’ waxy dogbane (Cycladenia humilis var. jonesii) ............................................... 12
Species Distribution Models ......................................................................................... 15
Niche Concepts ............................................................................................................. 16
Maximum Entropy (MaxEnt) ........................................................................................ 17
Sample Size ................................................................................................................ 19

v
Variable Selection ..................................................................................................... 19
Spatial Scale .............................................................................................................. 20
Spatial Resolution ...................................................................................................... 21
Thresholds ................................................................................................................. 23
Sampling Bias ............................................................................................................ 26
Forecasting ................................................................................................................ 27
METHODS ........................................................................................................................30
Study Area ..................................................................................................................... 30
Occurrence Data ............................................................................................................ 32
Data Acquisition ............................................................................................................ 32
Digital Elevation Model Acquisition ......................................................................... 32
Remote Sensing Acquisition ...................................................................................... 33
Climatic Data Acquisition ......................................................................................... 33
Environmental Variable Justification ............................................................................ 35
Variable Selection ......................................................................................................... 45
Model Parameters .......................................................................................................... 46
Spatial Scale .............................................................................................................. 46

vi
Spatial Resolution ...................................................................................................... 49
MaxEnt Calibration ................................................................................................... 49
Model Performance ....................................................................................................... 51
30-m and 900-m Current Models .............................................................................. 51
Model Forecasting..................................................................................................... 51
RESULTS ..........................................................................................................................53
Model Performance ....................................................................................................... 53
Resolution Comparison ................................................................................................. 62
Future Climate Trends ................................................................................................... 68
DISCUSSION & CONCLUSIONS ...................................................................................77
LITERATURE CITED ......................................................................................................86
VITA ................................................................................................................................104

vii
LIST OF FIGURES
Figure 1. ZION is in southwestern Utah and includes habitat for many threatened and
endangered species, including habitat for C. humilis var. jonesii and the A. microscaphus.
..............................................................................................................................................9
Figure 2. Location of the habitat range generated by the USGS for the Arizona toad
(Anaxyrus microscaphus) in the states of California, Nevada, Utah, New Mexico, and
Arizona. A portion of the habitat range is located inside of ZION (orange). The top-right
inset displays an adult Arizona toad. .................................................................................11
Figure 3. Location of the habitat range generated by the Fish and Wildlife Service for
Jones’ cycladenia (Cycladenia humilis var. jonesii) scattered throughout southeastern
Utah and northern Arizona. Habitat for C. humilis var. jonesii is fragmented and is known
to occur in only southeast Utah and far northern Arizona. The top-left inset displays a
flowering C. humilis var. jonesii........................................................................................13
Figure 4. A demonstration of a representative concentration pathway depicting the four
climate scenarios of (2.6, 4.5, 6.0, and 8.5). RCPs begin to differ from 2025-2030 and are
extrapolated to the year 2100. RCP 2.6 is considered the best-case scenario while RCP
8.5 is the worst-case climate scenario (). ...........................................................................29

viii
Figure 5. ZION is the study area for the SDMs created for A. microscaphus and C.
humilis var. jonesii. The inset in the top right displays the five national parks found
within Utah.........................................................................................................................30
Figure 6. Flow chart displaying the procedures in creating the proper parameters from
start to finish for use within the MaxEnt model for both study species. ...........................31
Figure 7. Digital elevation model for ZION at 30 m resolution. .......................................36
Figure 8. Slope derived from the original DEM for ZION at 30 m resolution. .................37
Figure 9. Aspect derived from the original DEM for ZION at 30 m resolution. ...............39
Figure 10. Terrain ruggedness index derived from the original DEM for ZION at 30 m
resolution............................................................................................................................40
Figure 11. Topographic position index derived from the original DEM for ZION at 30 m
resolution. This variable describes the valley bottom flatness in green and higher
elevation peaks in the red. ..................................................................................................42
Figure 12. Normalized difference vegetation index derived from Landsat 5 satellite
imagery for ZION at 30 m resolution. Green represents vegetation in this map and red
represents lack of vegetation. .............................................................................................43
Figure 13. Bare soil index derived from Landsat 5 satellite imagery for ZION at 30 m
resolution............................................................................................................................44
Figure 14. The model training for A. microscaphus was determined by constructing a 50
km buffer around the occurrence points obtained from GBIF.org. Of the 327 occurrence
points, 87 points remained after spatial rarefying which were used to train the MaxEnt

ix
model. Occurrence localities used inside and outside of ZION were used to train the
SDMs. ................................................................................................................................47
Figure 15. The model training extent was determined by constructing a 50 km buffer
around the occurrence points obtained from GBIF.org. Spatial rarefying was not
conducted on the occurrence points for C. humilis var. jonesii due to the low volume of
occurrence localities...........................................................................................................48
Figure 16. A. The average receiver operating curve (AUC) for A. microscaphus with the
five replicates run in MaxEnt. The red line representing the fit of the model to the
training data. The blue line represents the fit of the model to the 25% testing data. AUC
over 0.7 assumes positive predictive power for the model. B. Represents the test omission
rate and predicted area as a function of the cumulative threshold. ....................................54
Figure 17. A. The average receiver operating curve (AUC) for C. humilis var. jonesii with
the five replicates run in MaxEnt. The red line representing the fit of the model to the
training data. The blue line represents the fit of the model to the 25% testing data. AUC
over 0.7 assumes positive predictive power for the model. B. Represents the test omission
rate and predicted area as a function of the cumulative threshold. ....................................55
Figure 18. Partial dependence plots displaying the marginal response of the 12
environmental variables selected for A. microscaphus in the MaxEnt model. Each
response curve demonstrates the range of suitability for each environmental variable if
each variable were used to create a MaxEnt model independent of other variables. ........57

x
Figure 19. Partial dependence plots displaying the marginal response of the 13
environmental variable selected for Cycladenia humilis var. jonesii in the MaxEnt model.
Each response curve demonstrates the range of suitability for each environmental
variable if each variable were used to create a MaxEnt model independent of other
variables. ............................................................................................................................59
Figure 20. MaxEnt model output for 30 m resolution habitat suitability maps for A.
microscaphus in ZION. Suitability classes describe the ranking of habitat preference for
the species being modeled. Models were created using MaxEnt.......................................60
Figure 21. MaxEnt model output for 30 m resolution habitat suitability maps for C.
humilis var. jonesii in ZION. Suitability classes describe the ranking of habitat preference
for the species being modeled. Models were created using ArcMap and MaxEnt. ...........61
Figure 22. MaxEnt habitat suitability outputs for A. microscaphus with differing
resolution sizes to compare low to high suitability habitat for ZION. Highest suitability
habitat increased within the park with the higher resolution. All environmental variables
(topographic, remotely-sensed, and climate) were applied to these two outputs. .............66
Figure 23. MaxEnt habitat suitability outputs for C. humilis var. jonesii with differing
resolution sizes to compare low to high suitability habitat for ZION. Highest suitability
habitat decreased within the park with the higher resolution. All environmental variables
(topographic, remotely-sensed, and climate) were applied to these two maps. .................67
Figure 24. Forecasting SDM of A. microscaphus contrasting the suitable habitat for future
climate scenarios for representative concentration pathways that describe greenhouse gas

xi
concentration of 2.6 W/m2 and 8.5 W/m2 for the year 2050. The SDM covers the
complete training extent of the toad to better understand changes in suitable habitat (see
Figure 14). The ‘10 percentile training presence’ threshold was used to delineate suitable
habitat in this analysis. .......................................................................................................69
Figure 25. Forecasting SDM of A. microscaphus contrasting the suitable habitat for
climate scenarios for representative concentration pathways, which describe greenhouse
gas concentration of 2.6 W/m2 and 8.5 W/m2 for the year 2070 (see Figure 14). The SDM
covers the complete training extent of the toad to better understand changes in suitable
future habitat. The ‘10 percentile training presence’ calculated by the MaxEnt output was
used as the threshold to delineate suitable habitat. ............................................................70
Figure 26. Forecasting SDM of A. microscaphus for the current year (2020) and 2050
using binary distribution of suitable versus not suitable habitat. The ‘10 percentile
training presence’ threshold was calculated by MaxEnt to delineate suitable habitat in this
analysis. ..............................................................................................................................71
Figure 27. Forecasting SDM of A. microscaphus for the current year (2020) and 2070
using binary distribution of suitable versus not suitable habitat. The ‘10 percentile
training presence’ threshold was calculated by MaxEnt to delineate suitable habitat in this
analysis. ..............................................................................................................................72
Figure 28. Forecasting SDM of C. humilis var. jonesii contrasting the suitable habitat for
future climate scenarios for representative concentration pathways that describe
greenhouse gas concentration of 2.6 W/m2 and 8.5 W/m2 for the year 2050. The SDM

xii
covers the complete training extent of the plant to better understand changes in suitable
future habitat (see Figure 15). The ‘10 percentile training presence’ calculated by the
MaxEnt output was used to delineate suitable habitat in this analysis. .............................73
Figure 29. Forecasting SDM of C. humilis var. jonesii contrasting the suitable habitat for
future climate scenarios for with representative concentration pathways that describe
greenhouse gas concentration of 2.6 W/m2 and 8.5 W/m2 for the year 2070. The SDM
covers the complete training extent of the plant to better understand changes in suitable
habitat (see Figure 15). The ‘10 percentile training presence’ calculated by the MaxEnt
output was used to delineate suitable habitat in this analysis. ...........................................74
Figure 30. Forecasting SDM of C. humilis var. jonesii in ZION for the current year
(2020) and 2050 using binary distribution of suitable versus not suitable habitat. The ‘10
percentile training presence’ threshold was calculated by MaxEnt to delineate suitable
habitat in this analysis. .......................................................................................................75
Figure 31. Forecasting SDM of C. humilis var. jonesii in ZION for the current year
(2020) and 2070 using binary distribution of suitable versus not suitable habitat. The ‘10
percentile training presence’ threshold was calculated by MaxEnt to delineate suitable
habitat in this analysis. .......................................................................................................76

xiii
LIST OF TABLES
Table 1. List of the 19 bioclimatic variables taken from the WorldClim database and used
for analysis within the target species’ SDMs. Pearson’s correlation test was first used to
eliminate highly correlated variables with topographic and remotely sensed variables. ...34
Table 2. Results from Pearson’s correlation coefficient used to quantify correlation
among environmental variables. Resulting variables were used to train the model within
MaxEnt. ..............................................................................................................................45
Table 3. The MACHISPLIN package results for the climate variables downscaled to 30
m resolution and the r2 values associated with each layer. Results are based off an
ensemble approach using six algorithms. Independent variables used in the approach
include elevation, slope, aspect, and topographic wetness index. .....................................50
Table 4. Permutation importance values for each bioclimatic variable within the MaxEnt
model for the 30 m A. microscaphus SDM. The permutation value is determined by
randomly permuting the values of each independent variables against the training points.
Values are then normalized to provide percentages; higher values suggest greater
influence on the model. ......................................................................................................56
Table 5. Permutation importance values for each WorldClim variable for the MaxEnt
model for the 30 m C. humilis var. jonesii SDM. The permutation value is determined by

xiv
randomly permuting the values of each independent variables against the training points.
Values are then normalized to provide percentages; higher values suggest greater
influence on the model. ......................................................................................................58
Table 6. Differing MaxEnt outputs for both study species comparing the contrast ..........63
between 30 m and 900 m resolution. .................................................................................63
Table 7. The percent change between 30 m and 900 m SDM outputs for the study species
within five habitat suitability classes. ................................................................................64
Table 8. Permutation importance values for each bioclimatic variable within the MaxEnt
model for the 900 m A. microscaphus and C. humilis var. jonesii SDMs. The permutation
value is determined by randomly permuting the values of each independent variables
against the training points. Values are then normalized to provide percentages; higher
values suggest greater influence on the model. .................................................................65
Table 9. Area of suitable habitat for A. microscaphus within the training extent for future
climate scenarios for 2050 and 2070 with differing RCPs of 2.6 and 8.5 W/m2. ..............70
Table 10. Area of suitable habitat for A. microscaphus within ZION for current and future
climate scenarios of 2050 and 2070 with differing RCPs of 2.6 and 8.5 W/m2. ...............72
Table 11. Area of suitable habitat for C. humilis var. jonesii within the training extent for
future climate scenarios of 2050 and 2070 with differing RCPs of 2.6 and 8.5 W/m2. .....74
Table 12. Area of suitable habitat for C. humilis var. jonesii within ZION for future
climate scenarios of 2050 and 2070 with differing RCPs of 2.6 and 8.5 W/m2. ...............76

1
INTRODUCTION
Since the incorporation of new statistical methods and GIS tools, the development
of predictive species distribution models (SDMs) has expanded in the field of ecology,
biogeography, and conservation biology (Raes, 2012). SDMs describe how climatic and
environmental factors relate to species occurrences in geographic space, in order to
delineate suitable habitat over local, regional, and global scales. Common applications for
SDMs include projecting species distribution for current, past, and future climates,
studying relationships between environmental parameters and species richness, mapping
invasive species habitat range, and conservation planning (Melo-Merino et al., 2020).
Of notable interest from a conservation and management standpoint, is the
construction of SDMs to understand the current and future distribution of available
habitat for species, particularly habitat specialist. Habitat specialists display a narrow
range of environmental factors and have relatively limited geographic requirements, often
constricting the species to a defined range of suitable habitats for which they are well-
adapted (Hernandez et al., 2006; Büchi and Vuilleumier, 2014). In their optimal habitat, it
is believed that specialists perform better than generalists, with a trade-off to generalists
on performance and fitness in suboptimal habitats (Levins, 1968; Lawlor and Smith
1976; Marvier et al. 2004; Jasmin and Kassen 2007). However, alterations to resource

1
gradients can lead to unfavorable impacts on specialists. Specialist species are susceptible
to anthropogenic factors, such as climate change and urbanization (McKinney and
Lockwood, 1999). Interspecific competition also contributes to specialization within a
species (Biedma et al. 2019). Generalists can alter ecosystems by outcompeting
specialists, homogenizing ecosystems, and reducing biodiversity at the community level
(Büchi and Vuilleumier, 2013). These reductions in availability and resources can
fragment the available habitat, resulting in demographic isolation, population decline,
species extirpation, and ultimately leading to biodiversity loss (Vrba, 1987; Ricketts,
2001; Büchi and Vuilleumier, 2013). Monitoring the loss of biodiversity, especially
within specialist species is important to understand the identity, abundance, and shifts in
their habitat range (Díaz et al., 2006).
Due to the effects of climate change and other factors on desert landscapes,
understanding the available habitat to specialist species is of particular importance (IPCC,
2014). Globally, desert climates are changing faster than other non-polar terrestrial
ecosystems due to climate change (IPCC, 2014). Increased effects of climate change are
projected across the desert southwest in the 21st century with increases in aridity and
temperatures, along with longer drought durations (Cayan et al., 2010; Dominguez et al.,
2010; Seager and Vecchi, 2010). Arid environments, such as the desert southwest of the
United States, provide an array of ecosystems and microclimates conducive to examine
the current and projected availability of habitats for specialist species. Across regions of
the southwest, seasonal precipitation is erratic and prolonged droughts are common,

2
leading to adverse effects on landscape and ecosystems (Notaro et al., 2011). These
abiotic factors have shifted species to become better adapted to their xeric landscape.
Plants have become drought tolerant by growing deeper tap roots, inducing seed
dormancy, or utilizing paraheliotropism to minimize sun exposure (Canadell et al., 1996;
Chávez et al., 2016). Desert anurans have adapted to diminishing water resources by
becoming fossorial, utilizing explosive breeding behaviors, accelerating metamorphosis,
and becoming restricted geographically to stable water sources (Kulkarni et al., 2011;
Schalk et al., 2015).
To better understand species habitat requirements and the effects of future climate
change scenarios on species, researchers use SDMs such as the maximum entropy
modeling method (MaxEnt) to analyze these changes (Elith et al., 2010). MaxEnt is a
machine-learning technique used in modeling the distribution of a species’ habitat using
presence-only occurrence records (Phillips et al., 2006). The maximum entropy algorithm
attempts to estimate a probability distribution of species occurrence that is closest to
uniform while maintaining its environmental constraints (Elith et al., 2010). MaxEnt has
become a popular platform for species distribution modelling because of an ease of use
interface, implementation of presence-only data, low occurrence data requirements,
future forecasting ability, and its use of environmental data from across the study area
rather than a discriminative approach (Phillips and Elith, 2013). MaxEnt is also capable
of projecting one set of environmental layers to other locations using similarly formatted
environmental layers. Projecting is often used to map species in areas of changing

3
climate, observing potential habitat for invasive species, or building models in unknown
areas for target species evaluation (Phillips, 2017). MaxEnt is capable of handling both
continuous and categorical (discrete) environmental variables within its algorithm
(Phillips and Dudík, 2008). Using both continuous and categorical environmental data,
occurrence locations of the target species are then included into the MaxEnt algorithm to
build a model that projects a species habitat range across a geographic landscape to
identify other potential locations of suitable habitat.
With changing climates and diminishing habitats for many species, forecasting
SDMs has become a powerful tool for conservation practitioners and resource managers
as changing climates impact ecological systems (Guisan et al. 2013). MaxEnt can
construct SDMs to predict the changes in the geographic distribution of a species under
different climate change scenarios. These climate change scenarios are represented by
representative concentration pathways (RCPs), which are the developments of scenario
sets containing emissions, concentrations, and land-use trajectories (Vuuren et al., 2011).
RCPs project a potential future scenario and allow SDMs, such as MaxEnt, to capture the
shifts in suitable habitat for a species. This provides an invaluable tool for proactively
monitoring and planning conservation efforts for specialist species who are at risk of
extirpation and declining habitat due to changing climates.
One of the most diverse protected landscapes in the desert southwest, Zion
National Park (ZION) provides refuge to various protected species within its boundaries.
ZION was chosen as the study area due to its diverse landscape characterized by high

4
plateaus and deep sandstone canyons carved out by the Virgin River and additional
tributaries which support many microclimates. The southern section of the park is
characterized by desert habitat while the norther portion of the park is covered with high
plateau forests (US DOI, 2013a). An abundance of specialist species inhabit the park,
including the Arizona toad (Anaxyrus microscaphus), desert tortoise (Gopherus
agassizii), Gila monster (Heloderma suspectum), and Mexican spotted owl (Strix
occidentalis lucida) (US DOI, 2009; US DOI, 2013a). These and many other species are
sensitive to environmental alterations occurring such as habitat degradation, invasive
species encroachment, changes in hydrologic regimes, and rising temperatures due to
climate change (Ryan et al., 2014). To protect sensitive habitat within the park from the
changes in habitat, ZION complies with the National Environmental Policy Act in
addition to other environmental regulations, including the Endangered Species Act and
the National Historic Preservation Act (US DOI, 2013a).
This study concentrates on the habitat range of two arid adapted habitat specialists
within ZION, the Arizona toad (Anaxyrus microscaphus) and Jones’ waxy dogbane
(Cycladenia humilis var. jonesii) (Tilley et al., 2010; Ryan et al., 2015). Both species are
endemic to the desert southwest and display morphological traits typically found in
regions of prolonged drought and extreme temperatures. Anaxyrus microscaphus is a
habitat specialist that requires slow moving streams, sandy floodplains for burrowing,
and a narrow temperatures range for breeding (Sullivan, 1992; Ryan et al., 2017). Their
habitat is currently threatened by changes in the hydrological cycle, habitat

5
modifications, forest fires, hybridization, and introduced pathogens (Sullivan and Lamb,
1988; Ryan et al., 2014). Reports have shown that on a regional scale, toads are
declining, but locally have more stable populations based upon habitat conditions
(Sullivan, 1993; Bradford et al., 2005; Ryan et al., 2017). The habitat for C. humilis var.
jonesii is highly specialized, requiring gypsiferous and saline soils that are primarily
fragmented rock surfaces with soils at least 50 cm in depth (Welsh et al., 1987; USFWS,
2008). Main threats to C. humilis var. jonesii habitat arise from shifts in climate and land
use practice (Tilley et al., 2010). Populations for C. humilis var. jonesii are currently
geographically disjunct across southeastern Utah, little is known about the taxon’s
historic range (Sipes et al., 1994; Sipes and Wolf, 1997).Suitable habitat currently
remains for both study species inside of ZION, with common sightings of A.
microscaphus along riparian zones and other ephemeral water sources (Dalh et al., 2000).
Unfortunately, there have been no official sightings of C. humilis var. jonesii within the
park. Nearest populations are to the east in Garfield County, Utah and Mohave County,
Arizona (Welsh et al., 1987; Sipes et al. 1994).
Habitat specialists are known to have restricted spatial distribution patterns which
typically leads to limited occurrences localities (Kattan, 1992; Segurado and Araújo,
2004; Elith et al., 2006). Furthermore, SDMs for habitat specialists are known to have
narrow geographic ranges but have higher SDM accuracy than those of generalist species
(Luoto et al., 2005; Elith et al., 2006). Within this study, MaxEnt is used to capture the
distribution of A. microscaphus and C. humilis var. jonesii, with differing spatial

6
resolutions providing detail into the estimation of suitable habitat for higher resolution
models. Forecasting models with MaxEnt also observed the long-term habitat shifts due
to abiotic factors within the region and ZION.

7
OBJECTIVES
The goal of this study is to develop a SDM for arid adapted habitat specialist
species within ZION using the maximum entropy modelling methods (MaxEnt).
Generating reliable SDMs will benefit environmental managers in mapping valuable
species habitat to help establish a firm ecological background to assist in understanding
complex management issues. Below are the following objectives for the study:
1. Create a species distribution model for both the Arizona toad (Anaxyrus
microscaphus) and Jones’ Waxy Dogbane (Cycladenia humilis var. jonesii) to
delineate suitable habitat range within the ZION boundaries using the MaxEnt
software to construct ecologically relevant climate, topographic, and remotely
sensed variables to maximize effectiveness of model strength;
2. Construct SDMs for the target species at 30 m and 900 m spatial resolutions
within ZION, for comparison of model strength between the two resolutions; and
3. Utilize forecasting techniques to project each species’ distribution for the years
2050 and 2070 to understand the effect of climate change on habitat suitability for
the target species based on 2.6 and 8.5 W/m2 RCP scenarios. Future habitat
scenarios will be estimated by representative concentration pathways that predict
the measurement of greenhouse gas concentration for alternative future climates.

8
LITERATURE REVIEW
Zion National Park
ZION is in southwestern Utah (Figure 1) within Washington, Iron, and Kane
counties. ZION entered the national park system in 1919 under the signing of President
Woodrow Wilson. The park has an area of 601.9 km2, with 84% designated as wilderness
(US DOI, 2013a). The park is located at the juncture of the Colorado Plateau, Mojave
Desert, and Great Basin ecoregions. The elevation ranges from 2,660 m at its highest
point (Horse Ranch Mountain) to 1,117 meters (Coal Pits Wash) at its lowest point (US
DOI, 2013a). More than 1,000 plant species inhabit ZION with approximately 78 species
of mammals, 30 reptile species, 7 amphibians, 8 fish, and 291 species of birds (NPS,
2018). The last known stable population studied in ZION was located along the Virgin
River and Oak Creek riparian zones from 1998-1999 (Dahl et al., 2000). They are
believed to still inhabit the park, though no recent studies can support this claim. There is
no known literature of C. humilis var. jonesii populations occurring inside of ZION, only
potential suitable habitat remains within the park boundaries for the plant (US DOI,
2013b).

9
Figure 1. ZION is in southwestern Utah and includes habitat for many
threatened and endangered species, including habitat for C. humilis
var. jonesii and the A. microscaphus.

10
Arizona Toad (Anaxyrus microscaphus)
Anaxyrus microscaphus was originally described by Cope (1867) as Bufo
microscaphus. The toads’ habitat range expands primarily along the Mogollon Plateau in
western New Mexico, expanding through Arizona into far southwestern Utah and eastern
Nevada along the Virgin and Colorado River basins and its tributaries (Figure 2) (Dodd,
2013; Blais et al., 2016). Aside from the Virgin and Colorado River locations, historical
occurrences for the toad have been found in the Agua Fria, Salt, Verde, Bill Williams,
and Hassayampa Rivers in Arizona and the Gila, Mimbres, and San Francisco Rivers in
New Mexico (Sullivan and Lamb, 1988; Ryan et al., 2015). In New Mexico, roughly
70% of historical sites monitored for A. microscaphus recorded no observations in past
decades, implying a decline in New Mexico populations over that time span (Ryan et al,
2017). Monitoring of A. microscaphus populations by Ryan et al. (2017) between 2013
and 2016 along the Gila and San Francisco River showed that toad populations were
stable within those years, although local populations were vulnerable to local extirpation,
mainly due to random weather events. Currently, A. microscaphus is considered a
Species of Greatest Conservation Need in New Mexico and a state ‘sensitive’ species in
Arizona, Nevada, and Utah (New Mexico DGF, 2006; Dodd, 2013).
The toad is found at elevations of 365-2700 m and typically occupies marginal
zones or terraces, preferring mixtures of dense willow clumps and open flats or flood
channels (Sweet, 1992). Toads are typically observed from February to September where
they enter torpor for winter months (Schwaner and Sullivan, 2009). During the breeding

11
Fig
ure
2. L
oca
tion o
f th
e hab
itat
ran
ge
gen
erat
ed b
y t
he
US
GS
for
the
Ari
zona
toad
(Anaxy
rus
mic
rosc
aphus)
in t
he
stat
es o
f C
alif
orn
ia, N
evad
a, U
tah, N
ew M
exic
o, an
d
Ari
zona.
A p
ort
ion o
f th
e hab
itat
ran
ge
is l
oca
ted i
nsi
de
of
ZIO
N (
ora
ng
e).
The
top
-rig
ht
inse
t dis
pla
ys
an a
dult
Ari
zona
toad
.

12
season, males begin their calling when air temperatures range anywhere from 8 to 18°C
(Sullivan, 1992). Arizona toads remain close to flowing water sources during warmer
months and seldom migrate further than 200 m, typically remaining within floodplain
habitat (Schwaner and Sullivan, 2005). Clutch size average is around 4,500 eggs per
clutch and eggs are deposited in riparian areas of streams, shallows, backwashes, and
side-pools, where they hatch anywhere from 3-6 days (Blair, 1955; Schwaner and
Sullivan, 2005). Under normal conditions, tadpoles require relatively shallow, slow
flowing streams, and avoid faster moving water (Ryan et al, 2017).
Jones’ waxy dogbane (Cycladenia humilis var. jonesii)
Jones’ waxy dogbane is found in southern Utah counties (Emery, Grand, Garfield,
and Kane Counties) and Northern Arizona (Figure 3), occurring at a narrow range of
latitudes between 36° and 39° north (USFWS, 2008). Cycladenia humilis var. jonesii can
be found at elevations ranging from 1,300-1,800 meters on side slopes or at the base of
mesas, and typically within plant communities of mixed desert scrub, juniper, or wild
buckwheat-Mormon tea receiving 6 to 9 inches of mean annual precipitation (Tilley et
al., 2010). Cycladenia humilis var. jonesii is a long-lived herbaceous perennial in the
Dogbane family and grows 10-15 cm in height (USFWS, 2008). Flowering of the plant
takes place typically in April through June and produces a pink or rose-colored, trumpet
shaped flower. Soil requirements are edaphic and most if not all plants are found in
gypsiferous and saline soils of the Cutler, Summerville, and Chinle formations (USFWS,
2008). Often, habitat has been found on (80 to 100%) rock fragments, with shallow soils

13
Fig
ure
3. L
oca
tion o
f th
e hab
itat
ran
ge
gen
erat
ed b
y t
he
Fis
h a
nd W
ildli
fe S
ervic
e fo
r
Jones
’ cy
clad
enia
(C
ycla
den
ia h
um
ilis
var
. jo
nes
ii)
scat
tere
d t
hro
ug
ho
ut
south
east
ern
Uta
h a
nd n
ort
her
n A
rizo
na.
Hab
itat
for
C. hum
ilis
var
. jo
nes
ii i
s fr
agm
ente
d a
nd i
s know
n
to o
ccur
in o
nly
south
east
Uta
h a
nd f
ar n
ort
her
n A
rizo
na.
The
top
-lef
t in
set
dis
pla
ys
a
flow
erin
g C
. hum
ilis
var.
jones
ii.

14
less than 50 cm deep (Welsh et al., 1987). Cycladenia humilis replicates mainly by the
spreading of its rhizomes rather than by sexual reproduction, according to a study by
Sipes et al. (1994), supporting the theory of a lack of active primary pollinators to the
flower. It overwinters as a subterranean rhizome and is considered rhizomatous, meaning
it contains a long underground stem system not viewable from the ground surface.
Because C. humilis var. jonesii is a rhizomatous plant species it is made up of ramets,
which is an underground system of genetically identical individuals, the colony of ramets
makes up a genet (Sipes and Tepedino, 1996; USFWS, 2008).
Cycladenia humilis is a genus with three varieties currently recognized within the
species: C. humilis var. humilis, C. humilis var. venusta, and C. humilis var. jonesii.
Cycladenia humilis var. humilis is endemic to northern California while C. humilis var.
venusta is endemic to southern California (Hickman, 1993). Results from a study by
Brabazon (2015) supports the variation of jonesii indicates significant genetic structure,
supporting a possible delineation of jonesii as its own distinct species apart from the two
California variations. Cycladenia humilis var. jonesii was listed as a threatened species in
June of 1986 with an estimated total of 7,500 known individuals in the habitat range
during that time. As of 2008, there is believed to only be 1,100 individuals (Sipes and
Tepedino, 1996). Threats to C. humilis var. jonesii habitat are anthropogenic in nature
with disturbances including off-highway vehicle (OHV), oil and gas exploration,
livestock grazing, and the threat of rising temperatures due to climate change (Welsh et
al., 1987; Sipes et al., 1994). According to the recovery plan documented by FWS,

15
further monitoring and implementing of management plans for conservation of habitat is
currently being conducted (USFWS, 2008).
Species Distribution Models
SDM or environmental niche model (ENM) is an algorithmic method for the
modeling of a species habitat range based on the correlation between known occurrences
and the environmental conditions of occurrence localities (Elith and Leathwick, 2009). In
a Grinnellian sense, habitat modelling of an organism is adapted to tolerance zones or
niches, which are considered abiotic requirements in which a species is capable of
surviving within (Lorini and Vail, 2015). The utilization of species modelling has become
ubiquitous in many fields, especially those of analytical biology and can be used
extensively in conservation, natural resource management, ecology, evolution, and
invasive-species management (McShea, 2014; Pollock et al., 2014).
Among many types of models used in mapping species range habitat, some of the
more prevalently known statistical models fall under regression-based techniques, such
as: generalized linear model (GLM), generalized additive models (GAM), and
multivariate adaptive regression splines (Guisan et al., 2002; Elith and Leathwick, 2007).
The advancement of these particular analyses pioneered the development and growth of
innovative statistical methods and led to a renaissance of mechanistic models and
machine learning approaches. Between the years of 1992 and 2010 the increase in
published SDM related articles in ecological literature has increased from ten articles in
1992 up to 350 articles per year in 2010 (Brotons, 2014). As of 2019, the increase on

16
mendely.com has risen to 2,769 published articles on species distribution modelling.
Increasing in popularity, with the aid of highly effective computer system machine
learning techniques like those of MaxEnt, artificial neural networks (ANN), Genetic
algorithm for rule set production (GARP), boosted regression trees (BRT), random forest
(RF), support vector machines (SVM), and also ensemble models (Pearson et al., 2002;
Phillips et al., 2006; Elith et al., 2008; Evans et al., 2011; Grenouillet et al., 2011). Many
factors have contributed to the quick growth in the usage of species distribution
modelling such as the expanding accessibility in occurrence databases like that of
websites like International Union for Conservation of Nature (IUCN), iNaturalist, Global
Biodiversity Information Facility (GBIF), Biodiversity Heritage Library, Birdlife
International, or FishBase. These species occurrence databases are typically open source
websites that accumulate data by use of citizen scientists, uploading species sightings to
the website with exact coordinate location and additional detailed locality data.
Digitization of historical museum specimens has also contributed to the expanding
database collection for species occurrences.
Niche Concepts
Arauijo and Guisan (2006) proposed that one of the biggest challenges and most
overlooked elements of modelling species distribution is understanding and clarifying the
niche concept. Recognizing the differences between a fundamental niche and realized
niche is vital in comprehending the fluidity of the ever-shifting interactions with
interspecific interactions (i.e. predation, competition, mutualism). A Hutchinsonian

17
definition of a fundamental niche is the set of all conditions that allow for a species long-
term survival in the absence of competition, whereas a realized niche is a subset of the
fundamental niche that the species currently occupies with the presence of competition.
Chase and Leibold (2003) proposed a contrarian approach to defining a niche by
excluding the idea of a fundamental niche and realized niche altogether, they stated a
niche is limited by environmental factors that allows a population to reproduce at a rate
that is higher than the rate of mortality. Ambiguity on what a model represents often
results in misleading or inaccurate models. Soberón and Peterson (2005) supported the
idea that niche models provide an approximation of the species’ fundamental niche.
Conversely, other researchers have supported that models are spatial representations of
the realized niche (Guisan and Zimmerman, 2000; Pearson et al., 2002). Whether or not a
model represents a fundamental niche or a realized niche, the condition is dependent on
the parameters, variables, and algorithm representing the range in which a species
occupies.
Maximum Entropy (MaxEnt)
The principle of maximum entropy was presented by Edwin Thompson Jaynes in
1957, and since has helped expand disciplines such as thermodynamics, economics,
forensics, and ecology. MaxEnt software became available in 2004 and is a general-
purpose statistical machine-learning algorithm for making predictions from incomplete
datasets using presence-only data. MaxEnt contrasts presence data against background
samples, which are often called pseudo-absences (Phillips et al., 2009). Entropy is a

18
concept in information theory that measures the amount of information lost when the
value of a random variable is not known (Shannon, 1948). Lowering this amount of
entropy is key in developing a strong model. The more background information that we
have available the more entropy is lowered and the more uncertainty is reduced.
Increasing the data that indicates a species is present within an environment of ecological
conditions is information that will theoretically reduce the entropy within the model.
Within the MaxEnt model, entropy is measured on a grid cell (raster), the grid cell is
made up of pixels and within each pixel an occurrence point is either present or absent.
Any pixel that contains an occurrence point would be expected to demonstrate a
relatively low amount of entropy, while a pixel absent of an occurrence point would be
expected to have a high level of entropy (Phillips and Dudík, 2008). Occurrence points
are any coordinates denoting localities of where a particular species has been previously
recorded, typically using latitude and longitude. Many of these occurrence points are
derived from historical museum records or citizen science websites.
After each completion of a model, MaxEnt computes the area under the receiver
operating characteristic curve (AUC) as a tool for evaluating the predicted distribution of
species in a model. AUC was first developed for radar signal detection before being used
in medical research field and later accepted as the standard for assessing accuracy of
SDMs (Pepe, 2000; Jiménez-Valverde, 2012). Li and He (2018) proposed an approximate
guide for classifying the accuracy of AUC on scale ranging from 0-1: 0.90–1.00 =
excellent, 0.80–0.90 = good, 0.70–0.80 = fair, 0.60–0.70 = poor, and 0.50–0.60 = fail.

19
AUC values characterize the model’s ability to distinguish presence records from
background data.
Sample Size
Sample size in a species distribution model refers to the quantity of occurrence
point data collected for a species. The effects of sample size on a model are often weakly
considered in SDMs but can greatly influence the success rate of predicting suitable
species habitat (Stockwell and Peterson, 2002). Depending on the rarity of the species,
there is often a limit on occurrence data and exceptions must be implemented in
situations dealing with low occurrence records. Model performance is known to decrease
with samples sizes smaller than 15 and decrease dramatically for sample sizes smaller
than five (Pearson et al., 2006; Papeş and Gaubert, 2007). With small sample sizes,
outliers carry more weight in analyses, whereas more occurrence points help balance
outlier effects (Wisz et al., 2008). Also, uncertainty related to parameter estimates (e.g.
means, modes, medians) decrease with an increase in sample size (Crawley, 2002).
Though many model techniques are available, Hernandez et al. (2006) concluded in a
study that MaxEnt is the most capable in producing useful model results with smaller
sample sizes.
Variable Selection
Selection of environmental variables for SDMs should correspond with a deep
ideology and understanding of the species biogeography, ecology, population dynamics
and human disturbance. Careful selection of environmental variables is important in

20
producing a high quality, low bias model (Araújo and Guisan, 2006). MaxEnt, along with
many other machine learning models can use topographic, climatic, soil, and remotely
sensed variables. Yiwen et al. (2016) presents two methods for selecting environmental
variables in MaxEnt. The first method consists of selecting environmental variables based
from a priori or pre-selected ecological and biological knowledge. The second approach
utilizes a reiterative process of a stepwise removal of least contributing variables, both
approaches reduce overfitting and increase model accuracy.
In an article produced by Brown (2014) he outlines the use of a computer program
called “SDM Toolbox”, intended to work as a platform connecting both Python and
AcrGIS 10.1 (or higher). The toolbox consists of 59 scripts for use in macroecology,
landscape genetics, landscape ecology, and evolutionary studies. Among the many scripts
in the toolbox is the jackknifing tool, which measures variable importance and
systematically excludes one environmental variable at a time when running the model.
This process informs the user of variable contribution within the model while also
identifying highly correlated variables.
Spatial Scale
Spatial scale, commonly referred to as spatial extent or training range, is simply
the overall size of the study area in an SDM (Turner et. al, 1989). A common challenge
when constructing a species model is determining the appropriate extent of the study
area. Many study areas are determined by geographical or political borders, resulting in
poor model calibration leading to an incomplete range of environmental conditions. This

21
issue can lead to errors when extrapolating beyond the training range or when using
forecasting techniques for future modelling (El-Gabbas and Dormann, 2018).
A study by Williams et al. (2009) implemented a spatial scale design by
producing a 50-km buffer around occurrence points using a convex hull. Likewise,
Brown (2014) implements a convex hull buffer solution by buffering a set distance
around the occurrence points, in most cases 50-km. This helps eliminate overfitting by
reducing the spatial extent range and allows the model to select background points at only
feasible areas of dispersion (Brown, 2014).
Spatial Resolution
Spatial resolution, or grain size, is the minimum unit of a pixel or cell size within
a spatial grid. Studies suggest that consideration of pixel size and study extent can greatly
influence SDM performance (Martes and Jetz, 2018; Morgan and Guénard, 2019).
Natural environments are made up of geologic, climatic, topographic, and biological
processes with varying characteristics and spatial scales. Within each of these
environmental factors, species respond differently as spatial scales range from small
(local) to large (global) (Morgan and Guénard, 2019).
As computing power and high spatial resolution imagery become more powerful,
model performance and increasing model accuracy has proceeded. As is common with
SDMs, higher computational power for finer grain size resolution is often unnecessary
when modeling at larger extents. Coarser scaled models require less computational power
but can pose issues with overestimation of species models when mapping out species

22
distribution for local scaled habitats. Understanding and considering overestimation of
SDMs is important because a species’ actual distribution and geographic range may be
distorted at coarser scales (Jetz et al., 2007).
Advantages arise when modelling for local scale with higher grain resolution
rather than coarse-resolution models. Finer grain size enhances the details of the
landscape by sharpening the features and making the landscape more prominent and
distinguishable (Gottschalk et al., 2011). Spatial resolutions ranging from 10-100 m can
capture species distributions of features not visible at lower resolutions (1,000-10,000 m)
(Morgan and Guénard, 2019). In a study conducted by Nezer et al. (2017) on the grain
size effects of species distribution models of the Asiatic wild ass (Equus hemionus), high
resolution mapping allowed for detection of four habitat components essential to the wild
ass: potential movement corridors, isolated habitat patches, important topographic
features, and anthropogenic effect on distribution. The study demonstrated that
environmental variables such as slope and vegetation were nearly meaningless when
approaching 1 km resolution and that consideration must be considered for environmental
variables selection with respect to study extent (Nezer et al., 2017). In summary, fine-
scale distribution models are preferred for management and conservation planning when
modeling species at local scales (Hess et al., 2006).
Downscaling approaches for climate grids have only recently been introduced and
accepted in climate grid construction (Wang et al., 2011; Meineri and Hylander, 2017;
Morgan and Guénard, 2019). There are two known forms of downscaling: statistical and

23
dynamical. Dynamical downscaling utilizes regional climate models to extrapolate global
climate models to a regional or local resolution (Tang et al., 2016). Statistical
downscaling uses statistical relationships to predict regional or local climate grids from
low resolution variables (Benestat, 2004). The Worldclim climate grids, for example, is a
very well-known statistically downscaled database for climate surfaces that implements
thin-plate splines with covariates that include elevation, distance to the coast, minimum
and maximum land surface temperature, and cloud cover (Fick and Hijmans, 2017).
A study by Meineri and Hylander (2017) challenged the viewpoint that climate
station data are inadequate for producing downscaled climate data with justifiable results.
The study used data from climate stations, rather than weather data loggers, to build high
resolution climate grids over a large extent. Linear models regressing the temperature
against topographic variables were constructed, with thin-plate spline interpolation on the
regression residuals. Topographic variables of 30 m resolution were used which included
latitude, altitude, solar radiation, aspect, relative elevation, distance to sea and water
body, and topographic wetness index.
Thresholds
Primarily, the output for a typical SDM is a raster that displays the probability of
a species occurring in an area based on an algorithm with input data including both
environmental variables and species location datasets. This representation transforms
continuous results into a binary format and displays classes such as suitable, unsuitable,
or marginally suitable. Binary model results are often required when assessing ecological

24
issues such as climate change impacts, invasive species impacts, reintroduction sites
identification, and conservation planning. Selection of the threshold parameters greatly
influence model outcome and thoughtful consideration should be given in determining
the preferred requirements. Mismanagement of threshold selection can lead to overfitting
or underfitting of a model. Overfitting occurs when a model fits the calibration data too
closely in environmental or geographic space, whereas an underfit model fails to provide
adequate discrimination. Both overfitting and underfitting models lead to complications
when transferring the model to another region due to a lack of generality, this is known as
transferability (Radosavljevic and Anderson, 2014).
The simplest technique for displaying habitat suitability was presented by Phillips
and Dudík (2008); in order for an area to be considered suitable, the pixel value
encompassing areas of suitability must contain a probability greater than 0.5 as ‘present’
and all areas below 0.5 as ‘absent’. This leads to a clear distinction in determining the
rate of sensitivity and specificity, where sensitivity is the percent of ‘true’ presences
correctly classified as present in the model and specificity is the percent of ‘true’
absences labeled absent. Although this approach seems straight forward, it has been
drawn into question based on the ratio of presences to absences in that models are seldom
equal, providing bias when selecting arbitrary values such as 0.5 (Liu et al., 2005)
The lowest presence threshold was used by Philips et al. (2006), which
implements the minimum predicted value for the training sites as the threshold. This
technique of threshold selection is extremely sensitive to low sample sizes and should

25
only be used when using presence-only data. Once the threshold has been applied, model
performance can be evaluated using the extrinsic omission rate, which is a percentage of
test localities that fall into a pixel not predicted as suitable, and the proportional predicted
area, which is a percentage of the pixels that are predicted as suitable for the species
(Phillips et al., 2006) . Low omission rates are typically preferred for an above average
model (Anderson et al., 2003)
Liu et al. (2005) produced one of the most well-known threshold selection
methods for presence/absence data, referred to as maximizing the sum of sensitivity and
specificity (maxSSS). This method is supported as valid in use with presence-only data
when pseudo-absences are used instead of true absence data. This form of threshold
selection considers three criteria (objectivity, equality, and discriminability). Liu et al.
(2005) mathematically determined that maxSSS produced higher sensitivity, higher true
skill statistic, and higher kappa while also supporting that maxSSS produces the same
threshold using either presence/absence or presence-only data. Among other threshold
selection methods tested against maxSSS include: 1) training data prevalence (trainPrev),
2) mean predicted value (meanPred), 3) mid-point between the average predicted values
(midpoint), 4) maximizing kappa (max kappa), 5) maximizing overall accuracy (max
OA), 6) maximizing the F measure (max F), 7) minimizing the difference between
sensitivity and specificity (min DSS), 8) receiver operating characteristics (ROC), 9)
minimizing the distance between the precision-recall curve and the point (min D11) and
12) the predicted and observed prevalence equalization (equalPrev) (Liu et al., 2013). As

26
is the case with calculating sensitivity and specificity in a four-cell confusion matrix, the
same technique is used when applying to SDMs. Presence-only data uses computer
generated random points (pseudo-absences) rather than surveyed absence data. True
presences and false absences are calculated the same as with presence/absences data, and
the ‘true absences’ and ‘false absences’ are calculated using pseudo-absences (Liu et al.,
2015). MaxSSS is capable of being produced in both MaxEnt and open-source R
software.
Sampling Bias
Accuracy and validity of any species model is dependent upon the quality of the
input data. Sampling bias artificially increases spatial autocorrelation of the localities and
can lead to a model overfitting locality data in geographic space. Yackulic et al. (2013)
found that 87% of MaxEnt models used occurrence data likely influenced by sample
selection bias. MaxEnt models are commonly constructed on occurrence data that are
spatially biased towards easily accessed or better-surveyed areas, such as roads,
populated areas, or common water features (Reddy and Dávalos, 2003; Phillips et al.,
2009; Ruiz-Gutierrez and Zipkin, 2011). Consequently, it is of utmost importance to be
aware of inaccurate data due to the ramifications of incorrect models that in turn lead to
inappropriate management decisions (Phillips et al., 2009). Beck et al. (2014) detailed
that reducing spatial bias, at the loss of reduced input data, increases the predictive
species models to a degree. Fortunately, sampling bias can be reduced by spatially
filtering the occurrence dataset to reduce the degree of overfitting in a model. This

27
process considers the clustering of occurrence points within a particular radius and
randomly removes the localities, reducing the overall occurrences but in return,
improving model accuracy (Boria et al., 2014).
Forecasting
Forecasting has become a powerful tool for conservation practitioners and
resource managers as climate change impacts ecological systems. Resource managers
must constantly adapt to species shifting their distribution ranges in response to changing
temperature and precipitation. Deciphering how a species will respond to patterns of
land-use change allows land managers to design landscapes to better accommodate both
human and non-human resource needs. Many species respond to rising temperatures by
moving upward in elevation or poleward in latitude (Parmesan et al., 1999; Lenoir et al.,
2008). Over the past century, global average temperatures have risen 0.6 °C with
projections to rise between 1.1 and 6.4 °C in the next 100 years (IPCC 2014). Climate
change has become an extremely impactful ecological manipulator as it drives alterations
in hydrology, fire regimes, pathogen distribution, and distribution and cultures of human
populations (Lawler et al., 2011).
Often referred to as climate-envelope models, these forecasting models can
provide insight into future climate scenarios by projecting habitat suitability based on
potential changes in environmental conditions. These environmental conditions are
commonly composed of measured habitat attributes such as the structure of vegetation,
landscape patterns, soil type, and topography (Lawler et al., 2011). A study developed by

28
Hijmans et al. (2005) produced 1 km2 spatially interpolated climate data using thin-plate
smoothing spline algorithm to compile monthly averages using weather data from the
years (1950-2000). The data included in the forecast models include latitude, longitude,
and elevation variables to construct climate surfaces for monthly minimum, maximum,
and average temperature and precipitation. These climate surfaces are regularly used in
forecasting for species distribution and are available for download at
http://www.worldclim.org.
Future climate models are based on global climate model (GCMs), which use
representative concentration pathways (RCPs), an RCP is a call to the scientific
community to the request by the Intergovernmental Panel on Climate Change (IPCC) to
develop a set of scenarios to facilitate the future of climate change (IPCC, 2007). An
RCP is based on simulations from a set of integrated assessment models that provide
scenarios on concentrations and emissions of greenhouse gases, emissions of aerosols,
and associated land cover change scenarios (Arora et al., 2011). Based on Moss et al.
(2008) process on RCP design criteria, the following must be contained in the design: 1)
the RCP should be based on literature and contain an internally consistent description of
the future; 2) the RCP should provide information on all components of radiative forcing
in a geographically explicit way; 3) the RCP should have smooth transition between
analyses of historical and future periods; and 4) the RCPs should cover the time period up
to 2100. RCPs are based off four emission scenarios (Figure 4), a very low forcing level
(RCP 2.6), two medium stabilization scenarios (RCP 4.5 and 6), and (RCP 8.5). RCP

29
measures of units are based on watts per square meter (W/m2), that is, the sum of all
contributing emission sources (Vuuren et al., 2011).
A common RCP chosen for forecasting models is the Community Climate System
Model (CCSM4). This RCP was made available to public use in April 2010 and is a used
by a community of scientists, national laboratories, universities, and other institutions.
CCSM4 is a general circulation model consisting of atmosphere, land, ocean, and sea
components that are linked by state information and fluxes between components (Gent et
al., 2011). CCSM4 bioclimatic layers can be retrieved from the WorldClim website for
the years 2050 and 2070 with the RCPs of (2.6, 4.5, 6.0, and 8.5).
Figure 4. A demonstration of a representative concentration pathway
depicting the four climate scenarios of (2.6, 4.5, 6.0, and 8.5). RCPs
begin to differ from 2025-2030 and are extrapolated to the year 2100.
RCP 2.6 is considered the best-case scenario while RCP 8.5 is the worst-
case climate scenario.

30
METHODS
Study Area
The primary study area was focused in Zion National Park, located in
southwestern Utah. ZION has an area of 601.81 km2 within the boundaries of the park
(Figure 5). All proceeding MaxEnt models, excluding the forecasting models, were used
to project SDMs into the ZION boundary. A workflow for data collection was
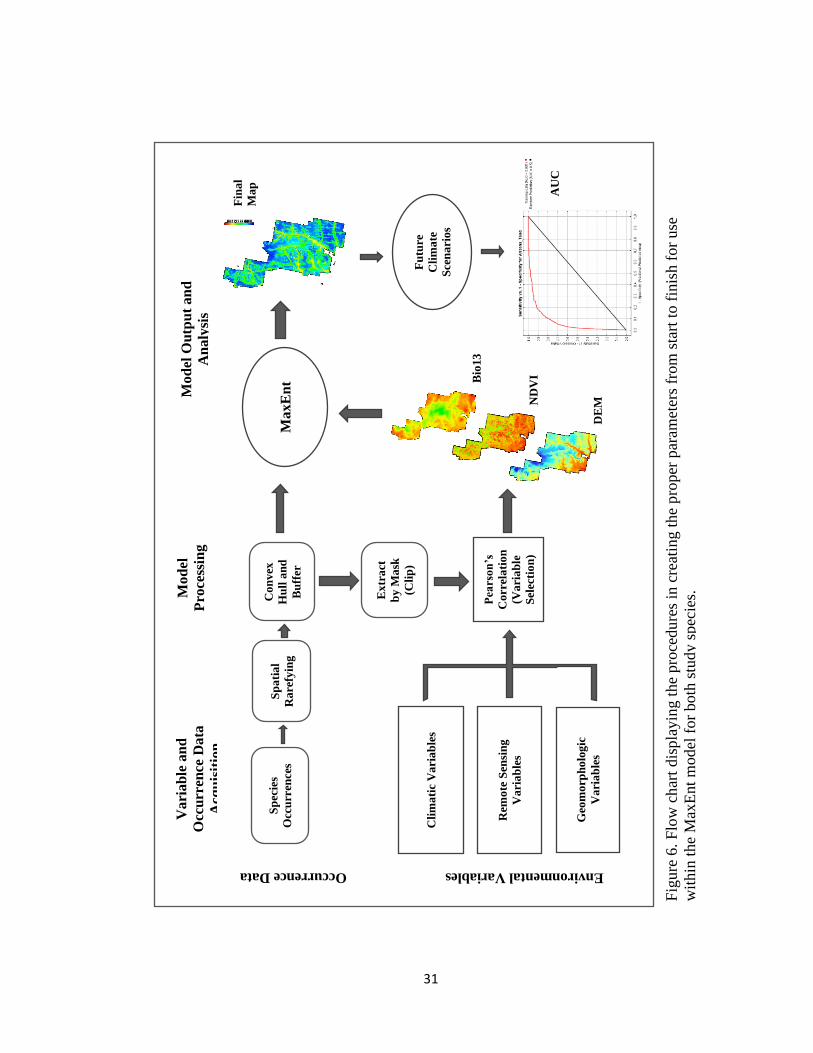
constructed to display the steps taken before model execution and analysis (Figure 6).
Figure 5. ZION is the study area for the
SDMs created for A. microscaphus and C.
humilis var. jonesii. The inset in the top right
displays the five national parks found within
Utah.
31
Va
ria
ble
an
d
Occ
urr
ence
Da
ta
Acq
uis
itio
n
Mod
el
Pro
cess
ing
Mod
el O
utp
ut
an
d
An
aly
sis
Rem
ote
Sen
sin
g
Va
ria
ble
s
Cli
ma
tic
Va
ria
ble
s
Geo
mo
rph
olo
gic
Va
ria
ble
s
Pea
rso
n’s
Co
rrel
ati
on
(Va
ria
ble
Sel
ecti
on
)
Sp
ecie
s
Occ
urr
ence
s S
pa
tia
l
Ra
refy
ing
Co
nv
ex
Hu
ll a
nd
Bu
ffer
Bio
13
ND
VI
DE
M
MaxE
nt
Environmental Variables
Fin
al
Ma
p
Occurrence Data
AU
C
Fu
ture
Cli
ma
te
Sce
na
rio
s
Ex
tra
ct
by
Ma
sk
(Cli
p)
Fig
ure
6.
Flo
w c
har
t dis
pla
yin
g t
he
pro
cedu
res
in c
reat
ing t
he
pro
per
par
amet
ers
from
sta
rt t
o f
inis
h f
or
use
wit
hin
the
Max
Ent
model
for
both
stu
dy
spec
ies.

32
Occurrence Data
Occurrence data were obtained using the Global Biodiversity Information Facility
(GBIF) (https://www.gbif.org/), the largest online open source provider of distribution
records. To prepare the coordinate data for the model, the occurrence points were
downloaded to an Excel sheet and were subjected to data cleaning (i.e. duplicate removal
and extreme outlier removal). After acquiring occurrence data for A. microscaphus, a
spatial filtering process was executed to reduce spatial autocorrelation. Filtering was not
done for A. humilis due to the limited amount of occurrence points available for
modeling, a total of 16 localities (Pearson et al., 2006; Papeş and Gaubert, 2007). Spatial
filter was completed by removing localities within a 30 km radius of one another. The
spatial filtering step was performed using the SDMtoolbox with the tool ‘Spatially Rarefy
Occurrence Data’ (Brown, 2014). Of the 327 occurrence points for A. microscaphus, 87
rarified occurrence localities were used in analyses.
Data Acquisition
Digital Elevation Model Acquisition
Digital elevation model (DEM) rasters were obtained from the NASA Earthdata
website (https://earthdata.nasa.gov/) and mosaiced together using ESRI ArcMap 10.6.1 to
form a master DEM. The master DEM was responsible for creating topographic
environmental variables for the model. North America Albers Equal Area Conic was
chosen as the projected coordinate system for creating environmental variables for both
study species, due to the regional scale of the model training area extent.

33
Remote Sensing Acquisition
Data acquisition for the remote sensing aspect of the environmental variables
were acquired using the USGS website (https://earthexplorer.usgs.gov/). The scenes
collected were atmospherically corrected by ordering the level two Landsat satellite
imagery. Landsat scenes were chosen and date back to a time frame that correlates with
average temperature, which coincides positively during breeding seasons for A.
microscaphus and flowering months for C. humilis var. jonesii. Imagery for A.
microscaphus used Landsat 7 imagery with <5% cloud cover with dates ranging in May
2002. Remote sensing covariates for both target species used normalized difference
vegetation index (NDVI), for characterizing various aspects of vegetation growth, and
bare soil index (BSI), to characterize areas of bare soil. C. humilis var. jonesii used
Landsat 5 imagery with <5% cloud cover and included scenes taken from May of 2009.
Climatic Data Acquisition
Climate data rasters were downloaded from the WorldClim website
https://www.worldclim.org/data/index.html at a 30 second resolution, roughly 1 km2 at
the equator, but varying resolution in the desert southwest at ~ 800 km2. WorldClim
climate data were extracted from global weather stations ranging from (1970-2000). The
19 variables (Table 1) represent annual trends and included the following: annual mean
temperature (BIO1), mean diurnal range (BIO2), isothermality (BIO3), temperature
seasonality (BIO4), max temperature of warmest month (BIO5), min temperature of
coldest month (BIO6), temperature annual range (BIO7), mean temperature of wettest

34
quarter (BIO8), mean temperature of driest quarter (BIO9), mean temperature of warmest
quarter (BIO10), mean temperature of coldest quarter (BIO11), annual precipitation
(BIO12), precipitation of wettest month (BIO13), precipitation of driest month (BIO14),
precipitation seasonality (BIO15), precipitation of wettest quarter (BIO16), precipitation
of driest quarter (BIO18), and precipitation of coldest quarter (BIO19).
Table 1. List of the 19 bioclimatic variables taken from the WorldClim database and used
for analysis within the target species’ SDMs. Pearson’s correlation test was first used to
eliminate highly correlated variables with topographic and remotely sensed variables.
BIO1 Annual Mean Temperature
BIO2 Mean Diurnal Range
BIO3 Isothermality
BIO4 Temperature Seasonality
BIO5 Max Temperature of Warmest Month
BIO6 Min Temperature of Coldest Month
BIO7 Temperature Annual Range
BIO8 Mean Temperature of Wettest Quarter
BIO9 Mean Temperature of Driest Quarter
BIO10 Mean Temperature of Warmest Quarter
BIO11 Mean Temperature of Coldest Quarter
BIO12 Annual Precipitation
BIO13 Precipitation of Wettest Month
BIO14 Precipitation of Driest Month
BIO15 Precipitation Seasonality
BIO16 Precipitation of Wettest Quarter
BIO17 Precipitation of Driest Quarter
BIO18 Precipitation of Warmest Quarter
BIO19 Precipitation of Coldest Quarter

35
Environmental Variable Justification
Digital Elevation Model
The digital elevation model was chosen to represent the elevation for each species
preferred habitat (Figure 7). All proceeding variables were created using ArcMap 10.6.1.
Slope
The slope and steepness of a region significantly influences runoff, especially in
mountainous areas like ZION. Cycaldenia humilis var. jonesii is known to reside on
barren gypsiferous clay hills that form sides and lower slopes (USFWS, 1986).
Alternatively, A. microscaphus is commonly found in areas with little to no slope,
although the toad prefers breeding and egg deposition in lightly flowing water (Ryan et
al., 2017). Slope (Figure 8) was constructed using the master DEM by calculating the
maximum rate of change from the target cell and the eight surrounding neighbors in the
raster.

36
Figure 7. Digital elevation model for ZION at 30 m resolution.

37
Figure 8. Slope derived from the original DEM for ZION at 30 m resolution.

38
Aspect
Anaxyrus microscaphus prefers habitat in valley bottom or areas with high canyon
walls surrounding streams and rivers. This habitat could be influenced by solar exposure
and a more southern facing valley could present preferred habitat for the toad. C. humilis
var. jonesii prefers areas with moderate slopes, although there is no literature on preferred
directional facing slopes. Surface temperatures between north- and south-facing slopes
can vary by 20°C, which is equivalent to 2000 km change in latitude (Scherrer and
Körner, 2010). Aspect (Figure 9) measures the direction the downhill slope faces and was
constructed using the master DEM as the input data.
Terrain Ruggedness Index
Terrain Ruggedness Index (TRI) expresses the difference in elevation from the
center cell and the eight cells directly surrounding it. The differences are then squared
and averaged, the square root of this average results in the TRI for that cell (Riley et al.,
1999). TRI was calculated and built (Figure 10) using the Vector Ruggedness Measure
tool developed by Sappington et al. (2007).

39
Figure 9. Aspect derived from the original DEM for ZION at 30 m
resolution.

40
Figure 10. Terrain ruggedness index derived from the original DEM for ZION
at 30 m resolution.

41
Topographic Position Index
Topographic position index was chosen for A. microscaphus to represent valley
bottoms which typically display a high probability of habitat suitability due to the
likelihood of alluvial accumulation and stream presence. TPI was calculated by the using
difference between a cell elevation value and the average elevation of the surrounding
neighborhood of the cell. TPI variables were created (Figure 11) using the Land Facet
Corridor Analysis version 1.2.605 toolbox (Jenness, 2006).
Normalized Difference Vegetation Index
NDVI was chosen because it quantifies vegetation along riparian areas where A.
microscaphus spends the majority of their life cycle near (Sweet, 1992). Additionally, C.
humilis var. jonesii has been observed in various plant communities (Tilley et al., 2010).).
NDVI (Figure 12) was calculated by obtaining temporally relevant Landsat imagery and
using the near-infrared and red color bands to generate an image displaying vegetation
abudnance.
Bare Soil Index
Bare soil index (BSI) was selected to quantify the localities inhabiting terrain
displaying a lack of vegetation. BSI relies on the short-wave infrared and red spectral
bands to quantify soil mineral composition while the blue and near infrared bands display
vegetative density. BSI environmental variables (Figure 13) were created using the
‘Raster Calculator’.

42
Figure 11. Topographic position index derived from the original DEM for
ZION at 30 m resolution. This variable describes the valley bottom flatness in
green and higher elevation peaks in the red.

43
Figure 12. Normalized difference vegetation index derived from Landsat 5
satellite imagery for ZION at 30 m resolution. Green represents vegetation in
this map and red represents lack of vegetation.

44
Figure 13. Bare soil index derived from Landsat 5 satellite imagery for
ZION at 30 m resolution.

45
Variable Selection
Environmental variables for both A. microscaphus and C. humilis var. jonesii
were constructed using the a priori process followed by removal of highly correlated
variables using Pearson’s correlation coefficient. To adjust for multicollinearity,
covariates displaying a high correlation above 0.9 were excluded from the model (Table
2). Within SDM Toolbox v2.4 in ArcMap, all the topographic, remotely sensed, and
climate layers were inserted into the ‘Remove Highly Correlated Variables’ in ASCII file
format. To reduce computational issues, the rasters were first resampled to a coarser
scale.
Table 2. Results from Pearson’s correlation coefficient used to quantify correlation
among environmental variables. Resulting variables were used to train the model within
MaxEnt.
Anaxyrus microscaphus Cycladenia humilis var. jonesii
BIO1 Annual Mean Temperature BIO1 Annual Mean Temperature
BIO2 Mean Diurnal Range BIO4 Temperature Seasonality
BIO3 Isothermality BIO6 Min Temperature of Coldest Month
BIO15 Precipitation Seasonality BIO9 Mean Temperature of Driest Quarter
BIO19 Precipitation of Coldest Quarter BIO13 Precipitation of Wettest Month
Digital Elevation Model BIO15 Precipitation Seasonality
Slope BIO17 Precipitation of Driest Quarter
Aspect Digital Elevation Model
Topographic Position Index Slope
Normalized Difference Vegetation Index Aspect
Bare Soil Index Normalized Difference Vegetation Index
Bare Soil Index

46
Model Parameters
Spatial Scale
The training extent for both species was determined by constructing a convex hull
fitted to the spatially filtered occurrence points followed by a 50 km buffer around the
hull. The hull is a perimeter that is fitted around the most outside group of points. The
convex hull was generated in ArcMap 10.6.1 using the ‘convex hull’ tool followed by a
buffer around the hull. The 50 km buffer allowed adequate background data to be
sampled outside the known habitat of each species (Figures 14 & 15). The hull was then
used to clip proceeding environmental variable rasters using the same extent, coordinate
system, pixel count, and resolution. Although the training extent is outside of ZION, it
was still used to train the model, in which case MaxEnt then projected the species
distribution for both species into ZION using the training extent.

47
Fig
ure
14. T
he
model
tra
inin
g f
or
A. m
icro
scaphu
s w
as d
eter
min
ed b
y c
on
stru
ctin
g a
50 k
m b
uff
er a
round
the
occ
urr
ence
poin
ts o
bta
ined
fro
m G
BIF
.org
. O
f th
e 327 o
ccurr
ence
poin
ts, 87 p
oin
ts r
emai
ned
aft
er
spat
ial
rare
fyin
g w
hic
h w
ere
use
d t
o t
rain
the
Max
En
t m
odel
. O
ccurr
ence
loca
liti
es u
sed i
nsi
de
and o
uts
ide
of
ZIO
N w
ere
use
d t
o t
rain
the
SD
Ms.

48
Fig
ure
15. T
he
model
tra
inin
g e
xte
nt
was
det
erm
ined
by c
onst
ruct
ing a
50 k
m b
uff
er a
round t
he
occ
urr
ence
poin
ts o
bta
ined
fro
m G
BIF
.org
. S
pat
ial
rare
fyin
g w
as n
ot
cond
uct
ed o
n t
he
occ
urr
ence
poin
ts f
or
C. hum
ilis
var
. jo
nes
ii d
ue
to t
he
low
volu
me
of
occ
urr
ence
loca
liti
es.

49
Spatial Resolution
The resolution for the environmental variable rasters were constructed using both
30 m and 900- m spatial resolutions. Topographic and remotely sensed covariates had a
predetermined spatial resolution of 30 m grain size and do not need further modifications
to meet grain size requirements. To create the 900 m topographic and remotely sensed
environmental variables, the 30 m resolution environmental variables were upscaled in
ArcMap to 900 m using cubic convolution.
In order to downscale climate data, the ‘R’ package MACHISPLIN was used to
interpolate Worldclim climate grids to 30 m spatial resolution through a machine learning
ensemble approach that used six algorithms: boosted regression trees, neural networks,
generalized additive model, multivariate adaptive regression splines, support vector
machines, and random forests (Hutchinson and Xu, 2013). The ensemble model
approach applied climate-forcing covariates of DEM, slope, aspect, and topographic
wetness index. Thin plate spline geographic interpolation was then used with the
residuals for smoothing of the climate rasters. Final r2 values and weighted model
algorithms used in the ensemble model were then displayed as an output from the
MACHISPLIN package (Table 3).
MaxEnt Calibration
Preconditioned settings were applied to MaxEnt to ‘cross-validate’ all replicates
for C. humilis var. jonesii, meaning the 16 occurrence points were divided into
subsections and each iteration used different occurrence points per iteration to avoid

50
duplicates (Phillips, 2017). The ‘bootstrap’ method was chosen for A. microscaphus to
account for larger sample size. MaxEnt used 25% of localities for testing (Phillips, 2017).
MaxEnt used cloglog to provide output species maps that ranged from zero to one, with
zero being least suitable and one being most suitable. A total of five iterations were used
to build each model, with the average of the five iterations being the final output.
Variable 01 02 03 04 05
Best Model br b b mrv b
Weight 76.3:23.7 100 100 25.3:60.2:14.5 100
R2 0.996 0.888 0.870 0.986 0.992
Variable 06 07 08 09 10
Best Model b b br bmrv br
Weight 100 100 69.4:30.6 52.4:17.8:168.8:13.1 80:20
R2 0.966 0.920 0.926 0.996 0.888
Variable 11 12 13 14 15
Best Model br mrv b bmr b
Weight 80.5:19.5 33.5:42.9:23.6 100 61.3:21.5:17.2 100
R2 0.870 0.986 0.992 0.966 0.920
Variable 16 17 18 19
Best Model b bv bm b
Weight 100 88.4:11.6 79.4:20.6 100
R2 0.926 0.996 0.888 0.870
Table 3. The MACHISPLIN package results for the climate variables downscaled to
30 m resolution and the r2 values associated with each layer. Results are based off an
ensemble approach using six algorithms. Independent variables used in the approach
include elevation, slope, aspect, and topographic wetness index.
*Letters depict the model algorithms: b = boosted regression trees, g = generalized additive
model, m = multivariate adaptive regression splines, v = support vector machines, r =
random forests, n = neural networks

51
Model Performance
30-m and 900-m Current Models
Habitat suitability rasters were divided into 5 classes by reclassifying the pixel
values in ArcMap and then assigning each pixel group to its suitability class. Suitability
classes ranged from 0-100%, with 0-20% being the least likely habitat suitability and 80-
100% being the most likely suitable habitat. Output rasters for the 30 m suitability maps
calculated the area of ZION at 601.81 km2 while 900 m maps calculated at 604.26 km2.
The shapefile used to extract the boundaries of ZION had an area of 601.81 km2 so no
discrepancies should have resulted due to inadequacies in measurement tools. Differing
cell resolution in rasters will result in both outputs being slightly dissimilar due to the clip
of the raster not having similar spatial extent and cell size. Many of the occurrence points
occur outside of ZION, therefore, the projection feature in MaxEnt interface was used to
map the habitat distribution for the target species within the park boundaries. Response
curves and percent permutation for each variable were also chosen as an output in the
MaxEnt settings to display model analyses. MaxEnt outputs for both A. microscaphus
and C. humilis var. jonesii were calculated statistically by using the area under the
receiver operating characteristic curve (AUC) to quantify strength of model.
Model Forecasting
Future climate scenario SDMs were based on representative concentration
pathways (RPCs) involving two emission scenarios, 2.6 W/m2 and 8.5 W/m2, for the
years 2050 and 2070. The Community Climate System Model version 4 (CCSM4) is the

52
climate model used for forecasting, it is composed of four models simultaneously
simulating the earth’s atmosphere, ocean, land surface, and sea-ice. The forecasting
models used the same environmental variables as previously mentioned (See Table 2) but
removed remotely sensed variables (NDVI and BSI). Remotely sensed variables were
excluded from both models due to uncertainty in vegetation and bare soil abundance for
future climates. Spatial resolution for the future climate data will be 30 second (900 m2)
spatial resolution, the finest resolution available within the WorldClim database. ZION
and the MaxEnt training extent (see Figures 14 & 15) were modeled for both species to
observe larger shifts in habitat suitability. The extent for A. microscaphus has a training
extent of 233,078 km2. Cycladenia humilis var. jonesii has a training extent area of
35,923 km2. In addition, ZION boundary forecasting models were also created for future
SDMs. To keep analysis consistent, the ‘10 percentile training presence’ was used as the
threshold to delimit suitable habitat against unsuitable habitat for each model (Escalante
et al., 2013). This threshold excludes all regions with habitat suitability lower than the
suitability values for the lowest 10% or occurrence records MaxEnt was used to compare
the differences between current day suitable habitat and the 2.6 and 8.5 W/m2 scenarios.

53
RESULTS
Model Performance
Anaxyrus microscaphus was recorded at 0.853 for training data AUC and 0.810
for test data AUC (Figure 16). Cycladenia humilis var. jonesii returned an AUC value of
0.796 for training data and 0.715 for testing data (Figure 17). Both models displayed
good predictive power with AUC for test data ranging above the 0.7 threshold and much
higher than 0.5, which represents a model that is no better than random. Percent
contribution to the model, based on permutation importance, showed that annual mean
temperature, elevation, and isothermality were the most contributing environmental
variables for A. microscaphus (Table 4 & Figure 18). Precipitation seasonality, NDVI,
and isothermality were the leading contributing environmental variables for C. humilis
var. jonesii (Table 5 & Figure 19). Maps displaying low against high habitat suitability
were created for both target species within ZION (Figures 20 & 21).

54
Figure 16. A. The average receiver operating curve (AUC) for A. microscaphus
with the five replicates run in MaxEnt. The red line representing the fit of the
model to the training data. The blue line represents the fit of the model to the
25% testing data. AUC over 0.7 assumes positive predictive power for the model.
B. Represents the test omission rate and predicted area as a function of the
cumulative threshold.
A.
B.

55
B.
A.
Figure 17. A. The average receiver operating curve (AUC) for C. humilis
var. jonesii with the five replicates run in MaxEnt. The red line representing
the fit of the model to the training data. The blue line represents the fit of the
model to the 25% testing data. AUC over 0.7 assumes positive predictive
power for the model. B. Represents the test omission rate and predicted area
as a function of the cumulative threshold.

56
Table 4. Permutation importance values for each bioclimatic variable within the MaxEnt
model for the 30 m A. microscaphus SDM. The permutation value is determined by
randomly permuting the values of each independent variables against the training points.
Values are then normalized to provide percentages; higher values suggest greater
influence on the model.
Variable Permutation Importance (%)
Annual Mean Temperature 22.9
Elevation 22.7
Isothermality 11.5
Normalized Difference Vegetation Index 11.1
Mean Diurnal Range 11
Precipitation Seasonality 5.7
Topographic Position Index 3.5
Slope 3.2
Precipitation of Coldest Quarter 2.7
Ruggedness 2.4
Aspect 2.1
Bare Soil Index 1.2

57
Figure 18. Partial dependence plots displaying the marginal response of the 12
environmental variables selected for A. microscaphus in the MaxEnt model. Each
response curve demonstrates the range of suitability for each environmental variable
if each variable were used to create a MaxEnt model independent of other variables.

58
Table 5. Permutation importance values for each WorldClim variable for the MaxEnt
model for the 30 m C. humilis var. jonesii SDM. The permutation value is determined by
randomly permuting the values of each independent variables against the training points.
Values are then normalized to provide percentages; higher values suggest greater
influence on the model.
Variable Permutation Importance (%)
Precipitation Seasonality 47.2
Normalized Difference Vegetation Index 34.2
Minimum Temp of Coldest Quarter 8.9
Elevation 8.5
Slope .4
Ruggedness .3
Temperature Seasonality 0
Mean Temperature of Driest Quarter 0
Precipitation of Wettest Month 0
Aspect 0
Bare Soil Index 0
Precipitation of Driest Quarter 0
Annual Mean Temperature 0

59
Figure 19. Partial dependence plots displaying the marginal response of the 13
environmental variable selected for Cycladenia humilis var. jonesii in the MaxEnt
model. Each response curve demonstrates the range of suitability for each
environmental variable if each variable were used to create a MaxEnt model
independent of other variables.

60
Fig
ure
20. M
axE
nt
model
outp
ut
for
30 m
res
olu
tion h
abit
at s
uit
abil
ity
map
s fo
r A
.
mic
rosc
aphus
in Z
ION
. S
uit
abil
ity c
lass
es d
escr
ibe
the
rankin
g o
f h
abit
at p
refe
rence
for
the
spec
ies
bei
ng m
odel
ed. M
odel
s w
ere
crea
ted u
sing M
axE
nt.

61
Fig
ure
21. M
axE
nt
model
outp
ut
for
30 m
res
olu
tion h
abit
at s
uit
abil
ity m
aps
for
C.
hum
ilis
var
. jo
nes
ii i
n Z
ION
. S
uit
abil
ity c
lass
es d
escr
ibe
the
ran
kin
g o
f h
abit
at
pre
fere
nce
for
the
spec
ies
bei
ng m
odel
ed. M
odel
s w
ere
crea
ted u
sing A
rcM
ap a
nd
Max
Ent.

62
Resolution Comparison
The test AUC for A. microscaphus recorded an output of 0.815, while C. humilis
var. jonesii recorded an output of 0.760. Both AUC outputs for the 900 m SDMs
provided higher model prediction power than the 30 m SDMs (Table 6). Most suitable
habitat increased from 900 m to 30 m for A. microscaphus, while the habitat decreased
from 900 m to 30 m spatial resolution for C. humilis var. jonesii. The percent change for
A. microscaphus was a 102% increase in the most suitable habitat range from 900 m to
30 m resolution. Percent change for C. humilis var. jonesii decreased by 68.7% for the
most suitable habitat range (Table 7). Leading variable contribution for the 900 m A.
microscaphus SDM was mean diurnal range, isothermality, and topographic position
index. The most contributing variables for C. humilis var. jonesii were NDVI,
precipitation seasonality, and aspect (Table 8). Maps displaying varying resolution sizes
for both target species were created within ZION (Figures 22 & 23).

63
Table 6. Differing MaxEnt outputs for both study species comparing the contrast
between 30 m and 900 m resolution.
Species Resolution Test
AUC
Suitability
Class
(km2) Hectare Percent
Anaxyrus
microscaphus
30m
.810
1-20% 174.28 17,428 28.96
20.1-40% 144.90 14,489 24.08
40.1-60% 123.21 12,321 20.47
60.1-80% 98.88 9,888 16.43
80.1-100% 60.54 6,054 10.06
900m
.815
1-20% 283.5 28,350 46.92
20.1-40% 127.17 12,717 21.05
40.1-60% 97.2 9,720 16.09
60.1-80% 66.42 6,642 10.99
80.1-100% 29.97 2,997 4.96
Cycladenia
humilis var.
jonesii
30m
.715
1-20% 343.62 34,362 57.10
20.1-40% 137.15 13,715 22.79
40.1-60% 76.88 7,688 12.77
60.1-80% 42.13 4,213 7.00
80.1-100% 2.03 203 .33
900m
.760
1-20% 469 46,899 77.61
20.1-40% 65.61 6,561 10.86
40.1-60% 36.45 3,645 6.03
60.1-80% 26.73 2,673 4.42
80.1-100% 6.48 648 1.07
.

64
Tab
le 7
. T
he
per
cent
chan
ge
bet
wee
n 3
0 m
and 9
00 m
SD
M o
utp
uts
for
the
study
spec
ies
wit
hin
fiv
e
hab
itat
suit
abil
ity c
lass
es.
Sp
ecie
s R
esolu
tion
S
uit
ab
ilit
y C
lass
Anaxy
rus
mic
rosc
aphus
1-2
0%
20.1
-40%
40
.1-6
0%
60.1
-80%
80.1
-100%
90
0m
283.5
127.1
7
97.2
66.4
2
29.9
7
30m
174.2
8
144.9
0
123.2
1
98.8
8
60.5
4
%
Chan
ge
-38.8
+
13.9
+
26.8
+
48.9
+
102.0
Cyc
laden
ia h
um
ilis
var
. jo
nes
ii
90
0m
469
65.6
1
36.4
5
26.7
3
6.4
8
30m
343.6
2
137.1
5
76.8
8
42.1
3
2.0
3
%
Chan
ge
-26.7
+
109.0
3
+110
.92
+57.6
-6
8.7

65
Table 8. Permutation importance values for each bioclimatic variable within the MaxEnt
model for the 900 m A. microscaphus and C. humilis var. jonesii SDMs. The permutation
value is determined by randomly permuting the values of each independent variables
against the training points. Values are then normalized to provide percentages; higher
values suggest greater influence on the model.
Anaxyrus microscaphus Variables Permutation Importance (%)
Mean Diurnal Range 22.1
Isothermality 17.2
Topographic Position Index 15.9
Normalized Difference Vegetation Index 10.6
Elevation 10.5
Annual Mean Temperature 6.6
Ruggedness 4.8
Aspect 3.6
Precipitation Seasonality 2.8
Bare Soil Index 2.4
Precipitation of Coldest Quarter 2.4
Slope 1.1
Cycladenia humilis var. jonesii Variables Permutation Importance (%)
Normalized Difference Vegetation Index 35.7
Precipitation Seasonality 25.1
Aspect 15.5
Slope 10.3
Minimum Temp of Coldest Month 7.7
Elevation 2.7
Ruggedness 1.9
Annual Mean Temperature 0.6
Precipitation of Wettest Month 0.3
Bare Soil Index 0.1
Temperature Seasonality 0.0
Mean Temperature of Driest Quarter 0.0
Precipitation of Driest Quarter 0.0

66
Fig
ure
22. M
axE
nt
hab
itat
suit
abil
ity o
utp
uts
for
A. m
icro
sca
phus
wit
h d
iffe
ring
reso
luti
on s
izes
to c
om
par
e lo
w t
o h
igh s
uit
abil
ity h
abit
at f
or
ZIO
N. H
ighes
t
suit
abil
ity h
abit
at i
ncr
ease
d w
ithin
the
par
k w
ith t
he
hig
her
res
olu
tion. A
ll
envir
onm
enta
l var
iable
s (t
opogra
phic
, re
mote
ly-s
ense
d, an
d c
lim
ate)
wer
e ap
pli
ed t
o
thes
e tw
o o
utp
uts
.

67
Fig
ure
23. M
axE
nt
hab
itat
suit
abil
ity o
utp
uts
for
C. hum
ilis
var
. jo
nes
ii w
ith d
iffe
ring
reso
luti
on s
izes
to c
om
par
e lo
w t
o h
igh s
uit
abil
ity h
abit
at f
or
ZIO
N. H
ighes
t su
itab
ilit
y
hab
itat
dec
reas
ed w
ithin
the
par
k w
ith t
he
hig
her
res
olu
tion. A
ll e
nvir
onm
enta
l var
iable
s
(topogra
phic
, re
mote
ly-s
ense
d, an
d c
lim
ate)
wer
e ap
pli
ed t
o t
hes
e tw
o m
aps.

68
Future Climate Trends
Based on the forecasting models for A. microscaphus, habitat suitability for the
training extent maps diminish as projections into the future for both 2.6 and 8.5 W/m2
scenarios. The 2070 8.5 W/m2 scenario projects a 5.14% suitable habitat compared to the
current day projection of 42.63% (Figures 24 & 25) (Table 9). Additionally, the
forecasting maps for ZION also display a reduction of suitable habitat within the park for
future climate scenarios (Figures 26 & 27) (Table 10). Conversely, the training extent
forecasting models for C. humilis var. jonesii projects minimal shifts in suitability for
both RCPs in the years 2050 and 2070 compared to the 2020 SDMs (Figures 28 & 29)
(Table 11). The C. humilis var. jonesii forecasting maps for ZION produced suitability
maps that showed an increase in potential habitat for the 2050 2.6 W/m2 RCP but a
decrease for the 2050 8.5 W/m2 RCP (Figure 30). Both 2070 RCPs displayed an increase
in similar suitable habitat for 2.6 and 8.5 W/m2 scenarios (Figures 31) (Table 12).

69
Figure 24. Forecasting SDM of A. microscaphus contrasting the suitable
habitat for future climate scenarios for representative concentration pathways
that describe greenhouse gas concentration of 2.6 W/m2 and 8.5 W/m2 for the
year 2050. The SDM covers the complete training extent of the toad to better
understand changes in suitable habitat (see Figure 14). The ‘10 percentile
training presence’ threshold was used to delineate suitable habitat in this
analysis.

70
Table 9. Area of suitable habitat for A. microscaphus within the training extent for future
climate scenarios for 2050 and 2070 with differing RCPs of 2.6 and 8.5 W/m2.
Species Year RCP (km2) suitability % suitability
Anaxyrus
microscaphus
Current (2020) Current 99,370 42.63%
2050 2.6 W/m2 15,125 6.48%
8.5 W/m2 17,702 7.59%
2070 2.6 W/m2 17,942 7.70%
8.5 W/m2 11,976 5.14%
Figure 25. Forecasting SDM of A. microscaphus contrasting the suitable
habitat for climate scenarios for representative concentration pathways,
which describe greenhouse gas concentration of 2.6 W/m2 and 8.5 W/m2 for
the year 2070 (see Figure 14). The SDM covers the complete training
extent of the toad to better understand changes in suitable future habitat.
The ‘10 percentile training presence’ calculated by the MaxEnt output was
used as the threshold to delineate suitable habitat.

71
Figure 26. Forecasting SDM of A. microscaphus for the current year (2020) and
2050 using binary distribution of suitable versus not suitable habitat. The ‘10
percentile training presence’ threshold was calculated by MaxEnt to delineate
suitable habitat in this analysis.

72
Table 10. Area of suitable habitat for A. microscaphus within ZION for current and future
climate scenarios of 2050 and 2070 with differing RCPs of 2.6 and 8.5 W/m2.
Species Year RCP (km2) suitability % suitability
Anaxyrus
microscaphus
Current (2020) Current 343.4 57.22
2050 2.6 W/m2 11.34 1.8
8.5 W/m2 21.06 3.50
2070 2.6 W/m2 32.4 5.4
8.5 W/m2 2.43 0.4
Figure 27. Forecasting SDM of A. microscaphus for the current year (2020) and
2070 using binary distribution of suitable versus not suitable habitat. The ‘10
percentile training presence’ threshold was calculated by MaxEnt to delineate
suitable habitat in this analysis.

73
Figure 28. Forecasting SDM of C. humilis var. jonesii contrasting the suitable
habitat for future climate scenarios for representative concentration pathways
that describe greenhouse gas concentration of 2.6 W/m2 and 8.5 W/m2 for the
year 2050. The SDM covers the complete training extent of the plant to better
understand changes in suitable future habitat (see Figure 15). The ‘10 percentile
training presence’ calculated by the MaxEnt output was used to delineate
suitable habitat in this analysis.

74
Table 11. Area of suitable habitat for C. humilis var. jonesii within the training extent for
future climate scenarios of 2050 and 2070 with differing RCPs of 2.6 and 8.5 W/m2.
Species Year RCP (km2) suitability % suitability
Cycladenia
humilis var.
jonesii
Current (2020) Current 20,516 57.10%
2050 2.6 W/m2 25,347 70.55%
8.5 W/m2 31,371 87.32%
2070 2.6 W/m2 19,947 55.52%
8.5 W/m2 21,565 60.02%
Figure 29. Forecasting SDM of C. humilis var. jonesii contrasting the suitable
habitat for future climate scenarios for with representative concentration pathways
that describe greenhouse gas concentration of 2.6 W/m2 and 8.5 W/m2 for the year
2070. The SDM covers the complete training extent of the plant to better understand
changes in suitable habitat (see Figure 15). The ‘10 percentile training presence’
calculated by the MaxEnt output was used to delineate suitable habitat in this
analysis.

75
Figure 30. Forecasting SDM of C. humilis var. jonesii in ZION for the current
year (2020) and 2050 using binary distribution of suitable versus not suitable
habitat. The ‘10 percentile training presence’ threshold was calculated by
MaxEnt to delineate suitable habitat in this analysis.

76
Table 12. Area of suitable habitat for C. humilis var. jonesii within ZION for future
climate scenarios of 2050 and 2070 with differing RCPs of 2.6 and 8.5 W/m2.
Species Year RCP (km2) suitability % suitability
Cycladenia
humilis var.
jonesii
Current (2020) Current 220.32 36.46
2050 2.6 W/m2 546.75 90.48
8.5 W/m2 42.12 6.97
2070 2.6 W/m2 557.28 92.22
8.5 W/m2 490.86 81.23
Figure 31. Forecasting SDM of C. humilis var. jonesii in ZION for the current
year (2020) and 2070 using binary distribution of suitable versus not suitable
habitat. The ‘10 percentile training presence’ threshold was calculated by
MaxEnt to delineate suitable habitat in this analysis.

77
DISCUSSION & CONCLUSIONS
Results from this study displayed promising predictive power for the 30 m SDMs
for A. microscaphus and C. humilis var. jonesii. The most contributing variables for both
models varied across all three classes (topographic, remotely sensed, and climatic) and
displayed little preference to one variable class type over the other. For the second
objective, I observed comparable results for model performance based on AUC and
contrasting results with respect to habitat suitability within ZION for the high versus low
spatial resolutions. The SDM showed higher habitat suitability for A. microscaphus at
finer spatial resolution, while the SDM displayed lower habitat suitability for C. humilis
var. jonesii at finer spatial resolution. Forecasting for the third objective showed
decreasing suitable habitat for A. microscaphus for future climate scenarios but an
increase in suitable habitat for C. humilis var. jonesii in future climate scenarios.
The results from the 30 m SDM models display similar findings with other studies
regarding acceptable predictive power for high spatial resolution modeling of specialist
species. Prior studies have displayed results that support higher predictive accuracy when
modeling for specialist species opposed to generalist species, even when using fewer
occurrence localities (Hernandez et al., 2006; Evangelista et al., 2008). Connor et al.
(2017) found that SDMs for species in heterogenous landscapes perform better compared
to homogenous landscapes. This likely occurs due to the SDMs ability to differentiate

78
extreme shifts in the heterogeneous landscape, allowing the model to delineate areas of
high versus low suitability. This corresponds positively with A. microscaphus habitat,
which often is in ravines and streams adjacent to steep cliffs hundreds of meters high.
Similarly, C. humilis var. jonesii habitat is fragmented in areas that are dispersed in
densely clustered colonies of ramets (US FWS 2008). These ramets are dispersed in
microhabitats, or microrefugia, which are climatically unique pockets of suitable habitat.
These microrefugia can often go undetected by SDMs when using low resolution
environmental variables (Dobrowski, 2010). Furthermore, this study demonstrates that
downscaling existing lower spatial resolution climate data can produce meaningful SDMs
that display local scale species habitat distribution. This supports the studies that
proposed downscaling climate data as a reliable method for mapping species distribution
at a local scale (Franklin et al., 2013; Slavish et al., 2014; Meineri and Hylander, 2017).
Environmental variable contributions produced by MaxEnt can be an integral
component to understanding the ecology that allows a species to persist within a set of
abiotic conditions. NDVI displayed high variable contribution, likely due to the toad’s
habitat preference of relatively higher vegetated riparian habitat in arid environments
where water and vegetation are sometimes scarce (Sweet, 1992). I also found that the
contributing variables for A. microscaphus support the inferred distributional patterns
which suggests that temperature sets the range limit for several amphibians (Schall and
Pianka 1978; Duellman and Sweet 1999). Three of the top five contributing variables for
the A. microscaphus SDM were derived from temperature. Annual mean temperature was

79
the most contributing environmental variable, along with isothermality the third most
contributing variable, and mean diurnal range the fifth most contributing variable (Table
4). Moreover, Préau et al. (2018) reported that minimum temperature was the top two
most contributing variable for three different European toads. Within that study, two
different modeling approaches were used with high spatial resolution climate and
topographic variables. Two SDM methods resulted in differing habitat suitability for each
study species, thus leading to the recommendation of producing two or more modeling
methods for local scale amphibians.
I observed increased variable contribution from both precipitation and NDVI for
C. humilis var. jonesii. NDVI likely demonstrated high variable contribution due to the
plants habitat preference to exist in communities of desert scrub and juniper (Tilley et al.,
2010). The use of NDVI in this SDM allowed the model to capture the vegetation within
areas and delineate high habitat suitability versus low habitat suitability. Likewise,
precipitation was a main variable contributor in the plant SDM. Precipitation is a strong
environmental indicator in many SDM plant studies; however, the absence of soil
variables likely reduced the model performance for the plant (Woodward and Williams,
1987; Syfert et al., 2013; Yang et al., 2013). A study by Hageer et al. (2017) suggests that
predictive power of SDMs perform better when models are calibrated with both climate
and soil data. Soil data was limited within ZION for my study, therefore, I completely
excluded soil from the SDM. The bare soil index variable was used as a proxy for soil,

80
but the remotely sensed data could not replicate the chemical and physical properties
within soil needed to create a highly contributing environmental variable.
The AUC for both SDMs increased with increasing grain size. This finding
contradicts previous studies which suggest, depending on the species, AUC will increase
as the grain size becomes finer (Gottschalk et al. 2011; Scales et al. 2017; Connor et al.,
2018). Guisan et al. (2007) found that an increase in grain size of the environmental
variables reduced the predictive power of some SDMs for certain species but improved
the SDMs for others. Additionally, Pradervand et al. (2014) conducted a study to
compare the predictive power of 239 mountainous plant species at six different spatial
resolutions (2, 5, 10, 25, 50, and 100 m) with three different types of SDMs. The study
found that variations in predictive accuracy of the models AUCs displayed little change
between the six spatial resolutions. Notably, I found that the AUC for both species
increased slightly from 30 m to 900 m spatial resolution (Table 6). These observations in
model performance are likely due to environmental variable contribution at different
scales. This supports findings that environmental variables can have different meanings
and respond to different resolutions when analyzing SDM predictive power (Lassueur et
al. 2006; Guisan et al, 2007; Pradervand et al. 2014). For C. humilis var. jonesii, the
highest suitable habitat increased as the grain size increased. This partially could be due
to the plant habitat being found at higher elevations within the park where the landscape
has increased homogeneity. Additionally, it could be due to the habitat features of the
plant being less distinguishable. Unlike the toad which has contrasting habitat features

81
(i.e. elevation, vegetation, temperature), the plant has fewer distinct features, possibly
making it difficult for high spatial resolution SDMs to capture the geographic landscape.
After analyzing two spatial resolutions (30 m and 900 m), further model evaluation with
multiple resolution types is recommended to produce higher accuracy models, especially
when species modeling for conservation and land management purposes.
Understanding and expanding existing methods for mapping habitat distribution
of microhabitat and microrefugia is imperative in understanding how species populations
shift during periods of unfavorable climate. Typically, populations migrate in latitude or
altitude during warming or cooling climates (Jump et al., 2009; Hampe and Jump 2011).
In topographically heterogeneous landscapes, species are also capable of persisting in
microrefugia, which may allow the species to persist until conditions outside
microrefugia allow establishment (Scherrer and Körner, 2010; Auffret et al., 2015).
However, amphibians often lack the mobility and dispersal ability needed to adapt to
local climate stresses (Halpin, 1997). Amphibian abundance within the environment is
linked to the interactions between temperature, precipitation, and vegetation. Where
vegetation produces microhabitat that can mitigate climate impacts (Seebacher and
Alford, 2002). These climatic factors greatly influence the distribution of amphibians
through changes in their phenology. A study by Blaustein et al. (2001) observed
fluctuations in temperature on a global scale, which altered the timing of breeding,
hibernation periods, and the ability to find food for some species. Over a 17-year period,
a gradual increase (0.11-0.24°C per year) in average maximum temperature between

82
March and April led to earlier breeding in two anurans and three salamanders. In the last
five years of the study, the anurans were breeding 2-3 weeks earlier than the first five
years of the study and 5-7 weeks earlier for the salamanders compared to the first five
years. However, other amphibian species breeding activity within the study remained
constant, despite rising temperatures over this timespan. I observed a reduction in
available suitable habitat distribution in future climate scenarios for A. microscaphus.
Contributing response curves for climate variables of the toad (Figure 18) show that as
the annual mean temperature increases above 10°C habitat sustainability begins to
decrease. Mean diurnal range, the difference between daily maximum and minimum
temperature, begins to decrease rapidly below 17°C for toad suitability. The same can be
said for precipitation seasonality, which begins to reduce rapidly in suitability below 30
mm. The future global climate model used in this study, CCSM4, demonstrates RCPs that
predict future temperature increases and future precipitation decreases up to the year
2100 for 3 climate scenarios (4.5, 6.0. 8.5 W/m2) and a stabilization for RCP 2.6 W/m2
globally (Meehl et al., 2012). Likewise, the International Panel on Climate Change
projects the earth’s surface in the Southwest to rise close to 1.7°C until 2100, with lower
precipitation in the southern portion of the Southwest region and little change or a slight
increase in precipitation in the northern portion (Garfin et al., 2013). Additionally,
snowpack will decrease from February to May up to the year 2100 in addition to
reductions in runoff and streamflow from the middle to the end of the twenty-first century
(Garfin et al., 2013; IPCC, 2014).

83
Plant population response to changing climates varies from species to species,
although two patterns appear consistent, particularly amongst arid species. First,
precipitation is positively correlated with plant reproduction; and second, drought years
often result in complete reproductive failure (Fox et al. 2006; Evans et al. 2007; Levine et
al., 2008). With C. humilis var. jonesii forecasting models, I was able to observe
significant increase in suitable habitat distribution for future climate scenarios of 2050
and 2070 for both the training extent and ZION maps. However, a decrease in suitable
habitat occurred for the 2050 8.5 W/m2 forecasting model. Response curves of the plant
(Figure 19) show that increasing temperature seasonality favors plant suitability.
Temperature seasonality is the measure of temperature change over the year, implying
that C. humilis var. jonesii thrives in areas of higher temperature variation over the course
of the year (USGS, 2012). Precipitation seasonality displays a decrease in suitability for
the plant as it increases, indicating future habitat scenarios project a decrease in this
variable based on the ubiquity of this species for both 2070 scenarios and the 2050 2.6
W/m2 scenarios. Precipitation seasonality is the measure of variation in monthly
precipitation totals over the course of a year (USGS, 2012). This finding is not consistent
when observing IPCC projections that detail seasonally erratic and localized precipitation
behavior for the desert southwest region for future years. In essence, the CCSM4 global
climate model used in the forecasting SDMs displays C. humilis var. jonesii flourishing
in most future scenarios. However, other climate models display high variability for
future years, especially when observing precipitation, which could potentially display no

84
change or reduction of suitable habitat for the plant (Garfin et al., 2013). Understanding
monthly climate variables opposed to yearly climate variables is especially important to
plants because of their adaptability to erratic precipitation events in arid environments.
These finding correspond with species modeling for plants and trees, in particular
forecasting models, where a study has shown monthly climate variables rather than
yearly variables improved SDM predictions of tree species (Zimmerman et al, 2009).
For higher performing SDM outputs, considerations to modify environmental
variables and model calibration are suggested. First, the incorporation of soil data into the
model would likely improve model predictive power. Incorporating Gridded National
Soil Survey Geographic Database (GNATSGO) data, a consolidation of STATSGO2,
SSURGO, and raster soil survey data would provide chemical and physical soil
properties for model construction. The database is integrated with ArcGIS and a
comprehensive effort created in 2019 to provide soil data up to 10 m in spatial resolution
for more than 90 percent of the United States and island territories. Unfortunately, most
likely due to the extreme changes in geography in ZION, an incomplete soil map remains
for that area, making species modeling inadequate. Secondly, forecasting models within
this study used a single global climate model, providing limited insight into future habitat
changes for the study species. Inclusion of multiple GCMs and pathways using ensemble
modeling will lead to a better understanding of future habitat based on an assortment of
scenarios. Use of an ensemble platform, such as BIOMOD, will allow modelers to
incorporate multiple model algorithms, GCMs, and RCPs to discover the optimum

85
grouping of variables and produce higher predictive models. BIOMOD is open source
and implemented in R and allows assessing of species temporal turnover, response
curves, and tests species interactions with environmental variables (Thuiller et al., 2009).
Thirdly, expanding on the science and production of downscaling climate data will
reduce downscaling inaccuracies and provide higher precision detection of microhabitats.
Only recently have methods and procedures used to downscale climate data begun to
expand the literature (Wang et al., 2011; Meineri and Hylander, 2017; Morgan and
Guénard, 2019). Lastly, SDM construction and analysis is only the first step to properly
map the habitat distribution for species. Further analysis by ground truthing and long-
term species monitoring will strengthen model reliability and allow SDM construction to
expand the ecological knowledge of study species (Rebelo and Jones, 2010).
Given the findings from the SDMs in this study, MaxEnt is a capable algorithm
and platform for mapping the distribution of species using topographical, remotely
sensed, and climate data. MaxEnt was capable of identifying suitable habitat for both
study species within ZION. The use of presence-only data, along with downscaling of
climate data to a finer resolution allows a better understanding of the ecological
interaction with species and their abiotic environment. With the use of SDMs,
conservation practitioners and land managers can work collaboratively to build and
interpret model results, leading to better conservational efforts for current and future
species distribution mapping.

86
LITERATURE CITED
Anderson, R. P., D. Lew, and A. T. Peterson. 2003. Evaluating Predictive Models of
Species’ Distributions: Criteria for Selecting Optimal Models. Ecological
Modelling. 162(3): 211–32. https://doi.org/10.1016/S0304-3800(02)00349-6.
Araújo, M. B. and A. Guisan. 2006. Five (or so) Challenges for Species Distribution
Modelling. Journal of Biogeography. 33(10): 1677–1688.
https://www.jstor.org/stable/pdf/3838509.pdf.
Arora, V. K., J. F. Scinocca, G. J. Boer, J. R. Christian, K. L. Denman, G. M. Flato, V. V.
Kharin, W. G. Lee, and W. J. Merryfield. 2011. Carbon Emission Limits Required
to Satisfy Future Representative Concentration Pathways of Greenhouse Gases.
Geophysical Research Letters. 38(5): 1-6. https://doi.org/10.1029/2010GL046270.
Auffret, A. G., J. Plue, and S. A. O. Cousins. 2015. The Spatial and Temporal
Components of Functional Connectivity in Fragmented Landscapes. AMBIO.
44(Suppl. 1): S51–S59. DOI 10.1007/s13280-014-0588-6.
Beck, J., M. Böller, A. Erhardt, and W. Schwanghart. 2014. Spatial Bias in the GBIF
Database and its Effect on Modeling Species’ Geographic Distributions.
Ecological Informatics. 19: 10–15. https://doi.org/10.1016/j.ecoinf.2013.11.002.
Benestad, R. E. 2004. Empirical-Statistical Downscaling in Climate Modeling. Eos,
Transactions American Geophysical Union. 85(42): 417–22.
https://doi.org/10.1029/2004EO420002.
Biedma, L., J. Calzada, J. A. Godoy, and J. Román. 2019. Local Habitat Specialization as
an Evolutionary Response to Interspecific Competition between Two
Sympatric Shrews. Journal of Mammalogy. 101(1): 80–91.
doi.org/10.1093/jmammal/gyz203.
Blais, B., M. J. Ryan, G. Gustafson, and J.T. Giermakowski. 2016. Natural History
Notes. Herpetological Review. 47(3): 436-438.
https://ssarherps.org/herpetological-review-pdfs/.

87
Blair, A. P. 1955. Distribution, Variation, and Hybridization in a Relict Toad (Bufo
microscaphus) in Southwestern Utah. American Museum Novitates. 1722: 1-38.
Blaustein, A. R., L. K. Belden, D. H. Olson, D. M. Green, T. L. Root, and J. M.
Kiesecker. 2001. Amphibian Breeding and Climate Change. Conservation
Biology. 15(6): 1804–1809. https://doi.org/10.1046/j.1523-1739.2001.00307.x
Boria, R. A., L. E. Olson, S. M. Goodman, and R. P. Anderson. 2014. Spatial Filtering to
Reduce Sampling Bias can Improve the Performance of Ecological Niche Models.
Ecological Modelling. 275: 73–77. https://doi.org/10.1016/j.ecolmodel.2013.12.
012.Brabazon, H. K. 2015. Delimiting Species and Varieties of Cycladenia
humilis (Apocynaceae). All Student Publications. pp. 172.
https://scholarsarchive.byu.edu/cgi/viewcontent.cgi?article=1178&context=
studentpub.
Brabazon, H. K. 2015. Delimiting Species and Varieties of Cycladenia Humilis
(Apocynaceae). All Student Publications. 172. https://scholarsarchive
.byu.edu/cgi/viewcontent.cgi?article=1178&context=studentpub.
Bradford, D. F., J. R. Jaeger, and S. A. Shanahan. 2005. Distributional Changes and
Population Status of Amphibians in the Eastern Mojave Desert. Western North
American Naturalist. 65(4): 462-472. https://scholarsarchive.byu.edu/cgi/
viewcontent.cgi?article=1828&context=wnan.
Brotons, L. 2014. Species Distribution Models and Impact Factor Growth in
Environmental Journals: Methodological Fashion or the Attraction of Global
Change Science. PLOS ONE. 9(11). doi:10.1371/journal.pone.0111996.
Brown, J. L. 2014. SDMtoolbox: A Python-based GIS Toolkit for Landscape Genetic,
Biogeographic, and Species Distribution Model Analyses. Methods in Ecology
and Evolution. 5(7): 694–700. https://doi.org/10.1111/2041-210X.12200.
Büchi, L and S. Vuilleumier, 2014. Coexistence of Specialist and Generalist Species is
Shaped by Dispersal and Environmental Factors. The American Naturalist.
183(5): 612-624. https://www.jstor.org/stable/pdf/10.1086/675756.pdf.
Canadell, J., R. B. Jackson, J. R. Ehleringer, H. A. Mooney, O. E. Sala, and E. D.
Schulze. 1996. Maximum Rooting Depth of Vegetation Types at the Global Scale.
Oecologia. 108(4): 583–595. https://www.jstor.org/stable/pdf/4221458.pdf.

88
Cayan, D. R., T. Das, D. W. Pierce, T. P. Barnett, M. Tyree, and A. Gershunov. 2010.
Future Dryness in the Southwest US and the Hydrology of the Early 21st Century
Drought. Proceedings of the National Academy of Sciences USA. 107: 21271–
21276. https://www.pnas.org/content/107/50/21271.
Chase, J. M., M. A. Leibold. 2003. Ecological Niches. The University of Chicago Press.
Chávez, R. O., J.G.P.W. Clevers, M. Decuyper, S. De Bruin, and M. Herold. 2016. 50
years of Groundwater Extraction in the Pampa del Tamarugal Basin: Can
Prosopis Tamarugo Trees Survive in the Hyper-arid Atacama Desert (Northern
Chile)? Journal of Arid Environments. 124: 293-303.
Crawley, M. 2002. Statistical Computing: An Introduction to Data Analysis Using S-
plus. Chichester, U.K: John Wiley & Sons.
Connor, T., V. Hull, A. Viña, A. Shortridge, Y. Tang, J. Zhang, F. Wang, and J. Liu.
2018. Effects of Grain Size and Niche Breadth on Species Distribution Modeling.
Ecography 41(8): 1270–82. https://doi.org/10.1111/ecog.03416.
Cope, E. D. 1867 “1866”. On the Reptilia and Batrachia of the Sonoran Province of the
Nearctic Region. Proceedings of the Academy of Natural Sciences of
Philadelphia. 18: 300–314.
Dahl, A., M. P. Donovan, and T. D. Schwaner. 2000. Egg Mass Deposition by Arizona
Toads, Bufo microscaphus, Along a Narrow Canyon Stream. Western North
American Naturalist. 60: 456-458.https://scholarsarchive.byu.edu/cgi/viewcontent
.cgi?article=1226&context=wnan.
Díaz, S., J. Fargione, F. S. Chapin III, and D. Tilman. (2006). Biodiversity Loss
Threatens Human Well-Being. PLOS Biology, 4(8), e277.
https://doi.org/10.1371/journal.pbio.0040277.
Dobrowski, S. Z. 2011. A Climatic Basis for Microrefugia: The Influence of Terrain on
Climate: A Climatic Basis for Microrefugia. Global Change Biology 17(2): 1022–
35. https://doi.org/10.1111/j.1365-2486.2010.02263.x.
Dodd, C. K. 2013. Frogs of the United States and Canada. John Hopkins University
Press. 127-132. https://ebookcentral.proquest.com/lib/sfasu-
ebooks/reader.action?docID=3318708#.

89
Dominguez, F., J. Cañon, and J. Valdes. 2010. IPCC-AR4 Climate Simulations for the
Southwestern US: The Importance of Future ENSO Projections. Climatic Change
99: 499– 514. DOI 10.1007/s10584-009-9672-5.
Drake, J. M., C. Randin, and A. Guisan. 2006. Modelling Ecological Niches with Support
Vector Machines. Journal of Applied Ecology. 43(3): 424–432.
https://doi.org/10.1111/j.1365-2664.2006.01141.x.
Duellman, W.E. and Sweet, S.S. 1999. Distribution Patterns of Amphibians in the
Nearctic Region of North America. Patterns of Distribution of Amphibians: A
Global Perspective. pp. 31–10. Johns Hopkins University Press, Baltimore, MD.
El‐Gabbas, A. and C. F. Dormann. 2018. Wrong, but Useful: Regional Species
Distribution Models may not be Improved by Range‐wide Data Under Biased
Sampling. Ecology and Evolution. 8(4): 2196–2206.
https://doi.org/10.1002/ece3.3834.
Elith, J., C. H. Graham, R. P. Anderson, M. Dudík, S. Ferrier, A. Guisan, R. J. Hijmans,
F. Huettmann, J. R. Leathwick, A. Lehmann, J. Li, L. G. Lohmann, B. A.
Loiselle, G. Manion, C. Moritz, M. Nakamura, Y. Nakazawa, J. M. Overton, A.
T. Peterson, S. J. Phillips, K. Richardson, R. Scachetti-pereira, R. E. Schapire, J.
Soberón, S. Williams, M. S. Wisz, and N. E. Zimmermann. 2006. Novel Methods
Improve Prediction of Species’ Distributions from Occurrence Data. Ecography.
29: 129–151. https://onlinelibrary.wiley.com/doi/pdf/10.1111/j.2006.0906-
7590.04596.x.
Elith, J. and J. Leathwick. 2007. Predicting Species Distributions from Museum and
Herbarium Records Using Multiresponse Models Fitted with Multivariate
Adaptive Regression Splines. Diversity and Distributions. 13(3): 265–275.
https://doi.org/10.1111/j.1472-4642.2007.00340.x.
Elith, J., J. R. Leathwick, and T. Hastie. 2008. A Working Guide to Boosted Regression
Trees. Journal of Animal Ecology. 77(4): 802–813.
https://besjournals.onlinelibrary.wiley.com/doi/pdf/10.1111/j.1365-
2656.2008.01390.x.
Elith J. and J. R. Leathwick. 2009. Species Distribution Models: Ecological Explanation
and Prediction Across Space and Time. Annual Review Ecology, Evolution, and
Systematics. 40: 667-697. https://www.annualreviews.org/doi/pdf/10.1146/
annurev.ecolsys.110308.120159.

90
Elith, J., S. J. Phillips, T. Hastie, M. Dudík, Y. E. Chee, and C. J. Yates. 2010. A
Statistical Explanation of MaxEnt for Ecologists. Diversity and Distributions.
17(1): 43–57. https://doi.org/10.1111/j.1472-4642.2010.00725.x.
Escalante, T., G. Rodríguez-Tapia, M. Linaje, P. Illoldi-Rangel, and R. González-López.
2013. Identification of Areas of Endemism from Species Distribution Models:
Threshold Selection and Nearctic Mammals. Revista Especializada en Ciencias
Químico-Biológicas. 16(1):5-17. https://www.medigraphic.com/pdfs/revespcie
quibio/cqb-2013/cqb131a.pdf.
Evangelista, P. H., S. Kumar, T. J. Stohlgren, C. S. Jarnevich, A. W. Crall, J. B. Norman
III, and D.T. Barnett. 2008. Modelling Invasion for a Habitat Generalist and a
Specialist Plant Species. Diversity and Distributions. 14(5): 808–17.
https://doi.org/10.1111/j.1472-4642.2008.00486.x.
Evans, J. S., M. A. Murphy, Z. A. Holden, and S. A. Cushman. 2011. Modeling Species
Distribution and Change Using Random Forest. Predictive Species and Habitat
Modeling in Landscape Ecology. 139–159. https://doi.org/10.1007/978-1-4419-
7390-0_8.
Evans, M. E. K., R. Ferriere, M. J. Kane, and D. L. Venable. 2007. Bet Hedging via Seed
Banking in Desert Evening Primproses (Oenothera, Onagraceae): Demographic
Evidence from Natural Populations. American Naturalist. 169(2): 184–194.
doi:10.1086/510599.
Fick, S. E. and R. J. Hijmans. 2017. WorldClim 2: New 1km Spatial Resolution Climate
Surfaces for Global Land Areas. International Journal of Climatology. 37(12):
4302-4315. https://www.worldclim.org/data/worldclim21.html.
Fox, L. R., H. N. Steele, K. D. Holl, and M. H. Fusari. 2006. Contrasting Demographies
and Persistence of Rare Annual Plants in Highly variable Environments. Plant
Ecology. 183: 157 –170. doi:10.1007/s11258-005-9014-2.
Franklin, J., F. W. Davis, M. Ikegami, A. D. Syphard, L. E. Flint, A. L. Flint, and L.
Hannah. 2013. Modeling Plant Species Distributions under Future Climates: How
Fine Scale Do Climate Projections Need to Be? Global Change Biology. 19(2):
473–83. https://doi.org/10.1111/gcb.12051.

91
Garfin, G., A. Jardine, R. Merideth, M. Black, and S. LeRoy. 2013. Assessment of
Climate Change in the Southwest United States: A Report Prepared for the
National Climate Assessment. A report by the Southwest Climate Alliance.
Washington, DC: Island Press.
Gent, P., G. Danabasoglu, L. Donner, M. Holland, E. Hunke, S. Jayne, and D. Lawrence.
2011. The Community Climate System Model Version 4. Journal of Climate.
(24): 4973–91. https://doi.org/10.1175/2011JCLI4083.1.
Gottschalk, T. K., B. Aue, S. Hotes, and K. Ekschmitt. 2011. Influence of Grain Size on
Species–habitat Models. Ecological Modelling. 222(18): 3403–3412.
https://doi.org/10.1016/j.ecolmodel.2011.07.008.
Grenouillet, G., L. Buisson, N. Casajus, and S. Lek. 2011. Ensemble Modelling of
Species Distribution: The Effects of Geographical and Environmental Ranges.
Ecography. 34(1): 9–17. https://doi.org/10.1111/j.1600-0587.2010.06152.x.
Guisan, A. and N. E. Zimmermann. 2000. Predictive Habitat Distribution Models in
Ecology.Ecological Modelling. 135(2–3): 147–186.
https://doi.org/10.1016/S0304-3800(00)00354-9.
Guisan, A., T. C. Edwards, and T. Hastie. 2002. Generalized Linear and Generalized
Additive Models in Studies of Species Distributions: Setting the Scene.
Ecological Modelling. 157: 89-100. https://www.sciencedirect.com/science/
article/pii/S0304380002002041.
Guisan, A., C. H. Graham, J. Elith, F. Huettmann. 2007. Sensitivity of Predictive Species
Distribution Models to Change in Grain Size. Diversity and Distributions. 13(3).
https://doi.org/10.1111/j.1472-4642.2007.00342.x
Guisan, A., R. Tingley, J. B. Baumgartner, I. Naujokaitis‐Lewis, P. R. Sutcliffe, A. I. T.
Tulloch, T. J. Regan, L. Brotons, E. McDonald‐Madden, C. Mantyka‐Pringle, T.
G. Martin, J. R. Rhodes, R. Maggini, S. A. Setterfield, J. Elith, M. W. Schwartz,
B. A. Wintle, O. Broennimann, M. Austin, and Y. M. Buckley. 2013. Predicting
Species Distributions for Conservation Decisions. Ecology Letters. 16(12): 1424–
1435. https://doi.org/10.1111/ele.12189.
Hageer, Y. M. Esperón-Rodríguez, J. B. Baumgartner, and L. J. Beaumont. 2017.
Climate, Soil or Both? Which Variables are Better Predictors of the Distributions
of Australian Shrub Species? PeerJ. DOI 10.7717/peerj.3446.

92
Halpin, P. N. 1997. Global Climate Change and Natural Area Protection: Management
Responses and Research Directions. Ecological Applications. 7: 828-843.
doi.org/10.1890/1051-0761(1997)007[0828:GCCANA]2.0.CO;2.
Hernandez, P. A., C. H. Graham, L. L. Master, and D. L. Albert. 2006. The Effect of
Sample Size and Species Characteristics on Performance of Different Species
Distribution Modeling Methods. Ecography. 29(5): 773–85.
https://doi.org/10.1111/j.0906-7590.2006.04700.x.
Hess, G. R., R. A. Bartel, A. K. Leidner, K. M. Rosenfeld, M. J. Rubino, S. B. Snider,
and T. H. Ricketts. 2006. Effectiveness of Biodiversity Indicators Varies with
Extent, Grain, and Region. Biological Conservation. 132: 448–457.
https://www.sciencedirect.com/science/article/pii/S0006320706001960.
Hickman, J. C. 1993. The Jepson Manual Higher Plants of California. University of
California Press. pp. 1424.
Hijmans, R. J., S. E. Cameron, J. L. Parra, P. G. Jones, and A. Jarvis. 2005. Very High-
Resolution Interpolated Climate Surfaces for Global Land Areas. International
Journal of Climatology. 25(15): 1965–1978. https://doi.org/10.1002/joc.1276.
Hutchinson, M. F. and T. Xu. 2013. ANUSPLIN Version 4.4 User Guide. Fenner School
of Environment and Society. Australian National University.
http://fennerschool.anu.edu.au/files/anusplin44.pdf.
Intergovernmental Panel on Climate Change (IPCC). 2007. Report of the 26th Session of
the IPCC. Bangkok. April 30–May 4 2007. Intergovernmental Panel on Climate
Change, Geneva, Switzerland.
Intergovernmental Panel on Climate Change (IPCC). 2014. Climate Change 2014:
Impacts, Adaptation, and Vulnerability. Part B: Regional Aspects. Contribution of
Working Group II to the Fifth Assessment Report of the Intergovernmental Panel
on Climate Change. Cambridge University Press, New York, New York, USA.
Jasmin, J. N., and R. Kassen. 2007. On the Experimental Evolution of Specialization and
Diversity in Heterogeneous Environments. Ecology Letters. 10: 272–281.
https://doi.org/10.1111/j.1461-0248.2007.01021.x
Jenness, J. 2006. Topographic Position Index Extension for ArcView 3.x. Jenness
Enterprises. http://www.jennessent.com/arcview/tpi.html.

93
Jetz, W., C. H. Sekercioglu, and J. E. M. Watson. 2007. Ecological Correlates and
Conservation Implications of Overestimating Species Geographic Ranges.
Conservation Biology. 22(1): 110–19. https://doi.org/10.1111/j.1523-
1739.2007.00847.x.
Jiménez‐Valverde, A. 2012. Insights into the Area under the Receiver Operating
Characteristic Curve (AUC) as a Discrimination Measure in Species Distribution
Modelling. Global Ecology and Biogeography. 21(4): 498–507.
https://doi.org/10.1111/j.1466-8238.2011.00683.x.
Kattan, G. H. 1992. Rarity and Vulnerability: The Birds of the Cordillera Central of
Colombia. Conservation Biology. 6(1): 64-70.
https://www.jstor.org/stable/pdf/2385851.pdf.
Kulkarni, S. S., I. Gomez‐Mestre, C. L. Moskalik, B. L. Storz, and D. R. Buchholz. 2011.
Evolutionary Reduction of Developmental Plasticity in Desert Spadefoot Toads.
Journal of Evolutionary Biology. 24(11): 2445–2455.
https://doi.org/10.1111/j.1420-9101.2011.02370.x.
Lassueur T., J. S, C. F. Randin. 2006. Very High-Resolution Digital Elevation Models:
Do they Improve Models of Plant Species Distribution? Ecological Modelling.
198(1–2): 139–153.
https://www.sciencedirect.com/science/article/abs/pii/S0304380006001712.
Lawlor, L. R., and J. M. Smith. 1976. Co-evolution and Stability of Competing Species.
American Naturalist. 110(971): 79–99.
https://www.jstor.org/stable/pdf/2459878.pdf.
Lawler, J. J., Y. F. Wiersma, and F. Huettmann. 2011. Using Species Distribution Models
for Conservation Planning and Ecological Forecasting. Predictive Species and
Habitat Modeling in Landscape Ecology. Springer New York. pp. 271–290.
https://doi.org/10.1007/978-1-4419-7390-0_14.
Lenoir, J., J. C. Gegout, P. A. Marquet, P. de Ruffray, and H. Brisse. 2008. A Significant
Upward Shift in Plant Species Optimum Elevation During the 20th Century.
Science.320: 1768-1771. https://science.sciencemag.org/content/320/5884/1768.
Levine, J. M., A. K. McEachern, and C. Cowan. 2008. Rainfall Effects on Rare Annual
Plants. Journal of Ecology. 96: 795–806. doi:10.1111/j.1365-2745.2008.01375x.

94
Levins, R. 1968. Evolution in Changing Environments. Princeton University Press.
Princeton, NJ.
Li, F. and H. He. 2018. Assessing the Accuracy of Diagnostic Tests. Shanghai Archives
of Psychiatry. 30(3): 207–12. https://doi.org/10.11919/j.issn.1002-0829.218052.
Liu, C., P. M. Berry, T. P. Dawson, and R. G. Pearson. 2005. Selecting Thresholds of
Occurrence in the Prediction of Species Distributions. Ecography 28(3): 385–93.
https://doi.org/10.1111/j.0906-7590.2005.03957.x.
Liu, C., M. White, and G. Newell. 2013. Selecting Thresholds for the Prediction of
Species Occurrence with Presence-only Data. Journal of Biogeography. 40(4):
778–789. https://www.jstor.org/stable/pdf/23463638.pdf.
Liu, C., G. Newell, and M. White. 2015. On the Selection of Thresholds for Predicting
Species Occurrence with Presence‐only Data. Ecology and Evolution. 6(1): 337–
48. https://doi.org/10.1002/ece3.1878.
Lorini, M. L. and M. M. Vale. 2015. Publication Trends in Species Distribution Modeling
and the Pioneer Contribution of Dr. Rui Cerqueira to Ecological Biogeography
and Distribution Modeling in Brazil. Oecologia Australis. 19(01): 16–31.
https://doi.org/10.4257/oeco.2015.1901.02.
Luoto, M., J. Pöyry, R. K. Heikkinen and K. Saarinen. 2005. Uncertainty of Bioclimate
Envelope Models Based on the Geographical Distribution of Species. Global
Ecology and Biogeography. 14(6): 575-584.
https://www.jstor.org/stable/pdf/3697674.pdf.
Martes, K. and W. Jetz. 2018. Disentangling Scale Dependencies in Species
Environmental Niches and Distributions. Ecography. 41: 1604–1615.
https://onlinelibrary.wiley.com/doi/pdf/10.1111/ecog.02871.
Marvier, M., P. Kareiva, and M. G. Neubert. 2004. Habitat Destruction, Fragmentation,
and Disturbance Promote Invasion by Habitat Generalists in a Multispecies
Metapopulation. Risk Analysis 24(4): 869– 878.
https://onlinelibrary.wiley.com/doi/abs/10.1111/j.0272-4332.2004.00485.x
McKinney, M. L., and L. J. Lockwood. 1999. Biotic Homogenization: A Few Winners
Replacing Many Losers in the Next Mass Extinction. Trends in Ecology &
Evolution. 14(11): 450–453. https://doi.org/10.1016/S0169-5347(99)01679-1.

95
McShea, W. 2014. What are the Roles of Species Distribution Modeling in Conservation
Planning? Environmental Conservation. 41(2): 93-96.
https://www.researchgate.net/publication/278188304_What_are_the_roles_of_spe
cies_di stribution_models_in_conservation_planning.
Meehl, G. A., W. M. Washington, J. M. Arblaster, A. Hu, H. Teng, C. Tebaldi, B. N.
Sanderson. 2012. Climate System Response to External Forcings and Climate
Change Projections in CCSM4. Journal of Climate 25(11): 3661–3683.
https://doi.org/10.1175/JCLI-D-11-00240.1.
Meineri, E. and K. Hylander. 2017. Fine-grain, Large-domain Climate Models Based on
Climate Station and Comprehensive Topographic Information Improve
Microrefugia Detection. Ecography. 40 (8): 1003–1013.
https://doi.org/10.1111/ecog.02494.
Melo-Merino, S. M., H. Reyes-Bonilla, and A. Lira-Noriega. 2020. Ecological Niche
Models and Species Distribution Models in Marine Environments: A Literature
Review and Spatial Analysis of Evidence. Ecological Modelling. 415.
https://doi.org/10.1016/j.ecolmodel.2019.108837.
Morgan, B. and B Guénard. 2019. New 30 m Resolution Hong Kong Climate,
Vegetation, and Topography Rasters Indicate Greater Spatial Variation than
Global Grids within an Urban Mosaic. Earth System Science Data. 11(3): 1083–
1098. https://doi.org/10.5194/essd-11-1083-2019.
Moss, R., M. Babiker, S. Brinkman, E. Cal NPS o, T. Carter, J. Edmonds, I. Elgizouli, S.
Emori, and K. A. Hibbard. 2008. Towards new scenarios for analysis of
emissions, climate change, impacts, and response strategies. IPCC Expert
Meeting Report on New Scenarios. Intergovernmental Panel on Climate Change,
Noordwijkerhout.
National Park Service (NPS). 2013. Foundation Document: Zion National Park.
https://www.nps.gov/zion/learn/management/upload/ZION_Foundation_Docume
nt_SP- 2.pdf.
National Park Service (NPS). 2018. Nezer & Science. https://www.nps.gov/zion/learn/
nature/index.htm.
New Mexico Department of Game and Fish. 2006. Comprehensive Wildlife Conservation
Strategy for New Mexico. New Mexico Department of Game and Fish. Santa Fe,
New Mexico. pg. 526.

96
Nezer, O., S. Bar-David, T. Gueta, and Y. Carmel. 2017. High-resolution Species-
distribution Model Based on Systematic Sampling and Indirect Observations.
Biodiversity and Conservation. 26(2): 421–437. https://doi.org/10.1007/s10531-
016-1251-2.
Notaro, M. and A. Zarrin. 2011. Sensitivity of the North American Monsoon to
Antecedent Rocky Mountain Snowpack. Geophysical Research Letters. 38:
L17403. https://agupubs.onlinelibrary.wiley.com/doi/pdf/10.1029/2011GL048803
Papeş, M. and P. Gaubert. 2007. Modelling Ecological Niches from Low Numbers of
Occurrences: Assessment of the Conservation Status of Poorly Known Viverrids
(Mammalia, Carnivora) Across Two Continents. Diversity and Distributions.
13(6): 890–902. https://doi.org/10.1111/j.1472-4642.2007.00392.x.
Parmesan C., N. Ryrholm, C. Stefanescu, J. K. Hill, C. D. Thomas, H. Descimon, B.
Huntley, L. Kaila, J. Kullberg, T. Tammaru, W. J. Tennent, J. A. Thomas, and M.
Warren. 1999. Poleward Shifts in Geographical Ranges of Butterfly Species
Associated with Regional Warming. Nature. 399: 579-583.
https://www.nature.com/articles/21181.
Pearson, R. G., T. P. Dawson, P. M. Berry, and P. A. Harrison. 2002. SPECIES: A
Spatial Evaluation of Climate Impact on the Envelope of Species. Ecological
Modelling. 154(3): 289–300. https://doi.org/10.1016/S0304-3800(02)00056-X.
Pearson, R. G., C. J. Raxworthy, M. Nakamura, and P. A. Townsend. 2006. Predicting
Species Distributions from Small Numbers of Occurrence Records: A Test Case
Using Cryptic Geckos in Madagascar. Journal of Biogeography. 34(1): 102–117.
https://doi.org/10.1111/j.1365-2699.2006.01594.x.
Pepe, M.S. 2000. Receiver Operating Characteristic Methodology. Journal of the
American Statistical Association. 95(449): 308–11.
https://doi.org/10.1080/01621459.2000.10473930.
Phillips, S. J., R. P. Anderson, and R. E. Schapire. 2006. Maximum Entropy Modeling of
Species Geographic Distributions. Ecological Modelling, 190(3): 231–259.
https://doi.org/10.1016/j.ecolmodel.2005.03.026.
Phillips, S.J. and M. Dudik. 2008. Modeling of Species Distributions with MaxEnt: New
Extensions and a Comprehensive Evaluation. Ecography. 31(2): 161–75.
https://doi.org/10.1111/j.0906-7590.2008.5203.x.

97
Phillips, S. J., M. Dudík, J. Elith, C. H. Graham, A. Lehmann, J. Leathwick, and S.
Ferrier. 2009. Sample Selection Bias and Presence-only Distribution Models:
Implications for Background and Pseudo-absence Data. Ecological Applications.
19(1): 181–97. https://doi.org/10.1890/07-2153.1.
Phillips, J. S. and J. Elith. 2013. On Estimating Probability of Presence from Use-
availability or Presence-background Data. Ecology. 94(6): 1409-1419.
https://pubmed.ncbi.nlm.nih.gov/23923504/.
Phillips, S. J. 2017. A Brief Tutorial on MaxEnt. https://biodiversityinformatics.amnh.org
/open_source/maxent/Maxent_tutorial2017.pdf.
Pollock, L. J., R. Tingley, W. K. Morris, N. Golding, R. B. O’Hara, K. M. Parris, P.
A.Vesk, and M. A. McCarthy. 2014. Understanding Co-occurrence by Modelling
Species Simultaneously with a Joint Species Distribution Model (JSDM).
Methods in Ecology and Evolution. 5(5): 397–406. https://doi.org/10.1111/2041-
210X.12180.
Préau, C., A. Troche, R. Bertrand., and F. Isselin-Nondedeu. 2018. Modeling Potential
Distributions of Three European Amphibian Species Comparing ENFA and
MaxEnt. Herpetological Conservation and Biology. 13(1):91–104. LINK.
Radosavljevic, A. and R.P. Anderson. 2014. Making Better MaxEnt Models of Species
Distributions: Complexity, Overfitting and Evaluation. Journal of Biogeography.
41(4): 629–643. https://onlinelibrary.wiley.com/doi/abs/10.1111/jbi.12227.
Raes, N. 2012. Partial Versus Full Species Distribution Models. Natureza &
Conservação. 10(2): 127–38. https://doi.org/10.4322/natcon.2012.020.
Rebelo, H. and G. Jones. 2010. Ground Validation of Presence-only Modelling with Rare
Species: A case study on Barbastelles Barbastella barbastellus (Chiroptera:
Vespertilionidae). Journal of Applied Ecology. 47: 410-420. doi: 10.1111/j.1365-
2664.2009.01765.x.
Reddy. S. and L. M. Dávalos. 2003. Geographical Sampling Bias and its Implications for
Conservation Priorities in Africa. Journal of Biogeography. 30(11): 1719-1727.
https://onlinelibrary.wiley.com/doi/10.1046/j.1365-2699.2003.00946.x.
Ricketts, T. H. 2001. The Matrix Matters: Effective Isolation in Fragmented Landscapes.
American Naturalist. 1(158): 87-99.
https://www.jstor.org/stable/pdf/10.1086/320863.pdf.

98
Riley, S. J., S. D. DeGloria, and R. Ellis. 1999. A Terrain Ruggedness Index that
Quantifies Topographic Heterogeneity. Intermountain Journal of Sciences. 5(1-4):
23-27. http://download.osgeo.org/qgis/doc/reference-
docs/Terrain_Ruggedness_Index.pdf.
Ruiz-Gutiérrez, V. and E. F. Zipkin. 2011. Detection Biases Yield Misleading Patterns of
Species Persistence and Colonization in Fragmented Landscapes. Ecosphere. 2(5):
1-14. https://esajournals.onlinelibrary.wiley.com/doi/full/10.1890/ES10-00207.1.
Ryan, M. J., I. M., Latella, and J. T. Giermakowski. 2014. Final Report: Current Status
of the Arizona Toad (Anaxyrus microscaphus) in New Mexico: Identification and
Evaluation of Potential Threats to its Persistence. New Mexico Department of
Game and Fish.
Ryan, M. J., I. M. Latella, J. T. Giermakowski, and H. L. Snell. 2015. Final Report:
Status of the Arizona Toad (Anaxyrus microscaphus) in New Mexico.
https://doi.org/10.25844/PRK4-NR28.
Ryan, M. J., J. T. Giermakowski, I. M. Latella, and H. L. Snell. 2017. Final Report:
Status of the Arizona Toad (Anaxyrus microscaphus) in New Mexico. Submitted
to New Mexico Department of Game and Fish. Available at the University of
New Mexico’s Digital Repository, https://digitalrepository.unm.edu/.
Sappington, J. M., K. M. Longshore, and D. B. Thompson. 2007. Quantifying Landscape
Ruggedness for Animal Habitat Analysis: A Case Study Using Bighorn Sheep in
the Mojave Desert. Journal of Wildlife Management. 71(5): 1419–26.
https://doi.org/10.2193/2005-723.
Scales, K. L., E. L. Hazen, M. G. Jacox, C. A. Edwards, A. M. Boustany, M. J. Oliver,
and S. J. Bograd. 2016. Scale of Inference: On the Sensitivity of Habitat Models
for Wide‐ranging Marine Predators to the Resolution of Environmental Data.
Ecography. 40(1). https://doi.org/10.1111/ecog.02272.
Schalk, C. M., C. G. Moñtana, L. Springer. 2015. Morphological Diversity and
Community Organization of Desert Anurans. Journal of Arid Environments. 122:
132-140. https://www.sciencedirect.com/science/article/abs/pii/S01401963
15300069?via%3Dihub.
Schall, J.J. and Pianka, E.R. 1978. Geographic Trends in Number of Species. Science.
201(4357): 679–686. https://science.sciencemag.org/content/201/4357/679

99
Scherrer, D. and C. Körner. 2010. Infra-red Thermometry of Alpine Landscapes
Challenges Climatic Warming Projections. Global Change Biology. 16: 2602-
2613. https://onlinelibrary.wiley.com/doi/abs/10.1111/j.1365-2486.2009.02122.x.
Schwaner, T. D., and B.K. Sullivan. 2005. Bufo microscaphus Cope, 1867 “1866”:
Arizona Toad. Pp. 422–424 In Amphibian Declines: The Conservation Status of
United States Species. University of California Press, Berkeley, California, USA.
Schwaner, T. D. and B. K. Sullivan. 2009. Fifty Years of Hybridization: Introgression
Between the Arizona Toad (Bufo microscaphus) and Woodhouse’s Toad (B.
Woodhousii) Along Beaver Dam Wash in Utah. Herpetological Conservation and
Biology. 9. http://www.herpconbio.org/Volume_4/Issue_2/ Schwaner
Sullivan_2009.pdf.
Seager, R. and G. A. Vecchi. 2010. Greenhouse Warming and the 21st Century
Hydroclimate of Southwestern North America. Proceedings of the National
Academy of Sciences USA . 107: 21277–21282.
https://www.pnas.org/content/107/50/21277.
Seebacher, F. and R. Alford. 2002. Shelter Microhabitats Determine Body Temperature
and Dehydration Rates of a Terrestrial Amphibian (Bufo Marinus). Journal of
Herpetology. 36: 69–75. https://doi.org/10.1670/00221511(2002)036[0069:
smdbta]2.0.co;2.
Segurado. P. and M. B. Araújo. 2004. An Evaluation of Methods for Modelling Species
Distribution. Journal of Biogeography. 31(10): 1555-1568.
https://www.jstor.org/stable/pdf/3554758.pdf.
Shannon, C. 1948. A Mathematical Theory of Communication. The Bell System
Technical Journal. 27(3): 379-423. https://pure.mpg.de/rest/items/item_23831
62_7/component/file_2456978/content.
Sipes, S. D., P. G. Wolf, V. J. Tepedino, and J. Boettinger. 1994. Population Genetics and
Ecology of Jones cycladenia. Bureau of Land Management Cost Share Program.
Utah State Office.
Sipes, S. D. and V. J. Tepedino. 1996. Pollinator Lost? Reproduction by the Enigmatic
Jones cycladenia, Cycladenia humilis var. jonesii (Apocynaceae) in Southwestern
Rare and Endangered Plants. Flagstaff, Arizona. Gen. Tech. Rep. RM-GTR-283.
p. 328.

100
Sipes, S. D. and P. G. Wolf. 1997. Clonal Structure and Patterns of Allozyme Diversity in
the Rare Endemic Cycladenia humilis var. jonesii (apocynaceae). American
Journal of Botany. 84(3): 401-409.
https://bsapubs.onlinelibrary.wiley.com/doi/pdf/10.2307/2446013.
Slavich, E., D. I. Warton, M. B. Ashcroft, J. R. Gollan, and D. Ramp. 2014. Topoclimate
versus Macroclimate: How Does Climate Mapping Methodology Affect Species
Distribution Models and Climate Change Projections? Diversity and Distributions
20(8): 952–63. https://doi.org/10.1111/ddi.12216.
Soberón, J. and A. T. Peterson. 2005. Interpretation of Models of Fundamental
Ecological Niches and Species’ Distributional Areas. Biodiversity Informatics. 2:
1-10. https://www.researchgate.net/publication/26539066.
Stockwell, D. R. B. and A. T. Peterson. 2002. Effects of Sample Size on Accuracy of
Species Distribution Models. Ecological Modelling. 148(1): 1–13.
https://doi.org/10.1016/S0304-3800(01)00388-X.
Sullivan, B. K. and T. Lamb. 1988. Hybridization Between the Toads Bufo microscaphus
and Bufo woodhousii in Arizona: Variation in Release Calls and Allozymes.
Herpetologica. 44(3): 325–333. https://www.jstor.org/stable/3892347.
Sullivan, B. K. 1992. Calling Behavior of Southwestern Toad (Bufo microscaphus).
Herpetologica. 48(4): 383-389. https://www.jstor.org/stable/pdf/3892856.pdf
Sullivan, B. K. 1993. Distribution of the Southwestern Toad (Bufo microscaphus) in
Arizona. Great Basin Naturalist. 53(4): 402-406.
https://scholarsarchive.byu.edu/cgi/viewcontent.cgi?article=2794&context=gbn.
Sweet, S. S. 1992. Initial Report on the Ecology and Status of the Arroyo Toad (Anaxyrus
Microscaphus Californicus) on the Los Padres National Forest of Southern
California, with Management Recommendations. Contract Report to USDA,
Forest Service, Los Padres National Forest, Goleta, California. pp. 198.
Syfert, M. M., M. J. Smith, and D. A. Coomes. 2013. The Effects of Sampling Bias and
Model Complexity on the Predictive Performance of MaxEnt Species Distribution
Models. PLOS ONE 8. 2: e55158. https://doi.org/10.1371/journal.pone.0055158.

101
Tang, J., X., Niu, S., Wang, H., Gao, X., Wang, and J. Wu. 2016. Statistical Downscaling
and Dynamical Downscaling of Regional Climate in China: Present Climate
Evaluations and Future Climate Projections. Journal of Geophysical Research:
Atmospheres. 121(5): 2110–29. https://doi.org/10.1002/2015JD023977.
Tilley, D., L. St. John, and D. Ogle. 2010. Plant Guide for Jones’ Waxy Dogbane
(Cycladenia humilis var. jonesii). USDA-Natural Resources Conservation
Service. https://plants.usda.gov/plantguide/pdf/pg_cyhuj.pdf.
Turner, M. G., R. V. O. R. H. Gardner, R. V. O’Neill, R. H. Gardner, and B. T. Milne.
1989. Effects of Changing Spatial Scale on the Analysis of Landscape Pattern.
Landscape Ecology. 3: 153-162. https://link.springer.com/article/10.1007
/BF00131534.
US Department of the Interior (US DOI). National Park Service (NPS). 2001. Zion
National Park: General Management Plan.
https://www.nps.gov/zion/learn/management/upload/zion_gmp.pdf
US Department of the Interior (US DOI). National Park Service (NPS). 2009. Zion
National Park. Reptiles and Amphibians.
https://www.nps.gov/zion/learn/nature/upload/Reptiles&Amph2009.pdf
US Department of the Interior (US DOI). National Park Service (NPS). 2013a.
Foundation Document. Zion National Park. Utah.
http://npshistory.com/publications/foundation-documents/zion-fd-2013.pdf.
US Department of the Interior (US DOI). National Park Service (NPS). 2013b. Virgin
River Comprehensive Management Plan/Environmental Assessment. Zion
National Park and Bureau of Land Management.
https://www.rivers.gov/documents/plans/virgin-plan.pdf
US Department of the Interior (US DOI). National Park Service (NPS). 2014. The Basics
of Compliance with the National Environmental Policy Act and Section 106 of
the National Historic Preservation Act: Instructional Manual, Version 1. Denver,
Colorado: NPS Intermountain Region.
US Fish and Wildlife Service (US FWS). 1986. Rule to Determine Cycladenia
humilis var. jonesii (Jones’ cycladenia) to be a Threatened Species. Federal
Register 51(86): 16526-16530.

102
US Fish and Wildlife Service (US FWS). 2008. Recovery Outline for the Jones’
cycladenia (Cycladenia humilis var. jonesii). https://www.fws.gov/mountain-
prairie/es/species/plants/jonescycladenia/RecoveryOutlineDecember2008.pdf.
Vuuren, D. P. V, J. Edmonds, M. Kainuma, K. Riahi, A. Thomson, K. Hibbard, and G. C.
Hurtt. 2011. The Representative Concentration Pathways: An Overview. Climatic
Change. 109(1). https://doi.org/10.1007/s10584-011-0148-z.
Vrba, E. S. 1987. Ecology in Relation to Speciation Rates: Some Case Histories of
Miocene-Recent Mammal Clades. Evolutionary Ecology. 1: 283–300.
https://link.springer.com/content/pdf/10.1007/BF02071554.pdf.
Wang. T., A. Hamann, D. L. Spittlehouse, T. Q. Murdock. 2011. ClimateWNA—High-
Resolution Spatial Climate Data for Western North America. Journal of Applied
Meteorology and Climatology. 51: 16-29. DOI: 10.1175/JAMC-D-11-043.1.
Welsh, S. L., N. D. Atwood, L. C. Higgins, and S. Goodrich. 1987. A Utah Flora. Great
Basin Naturalist Memoirs No. 9. Brigham Young University. Provo, UT.
Williams, J. N., C. Seo, J. Thorne, J. K. Nelson, S. Erwin, J. M. O’Brien, and M. W.
Schwartz. 2009. Using Species Distribution Models to Predict New Occurrences
for Rare Plants. Diversity and Distributions. 15(4): 565–576.
https://doi.org/10.1111/j.1472-4642.2009.00567.x.
Wisz, M. S., R.J. Hijmans, J. Li, A.T. Peterson, C. H. Graham, and A. Guisan. 2008.
Effects of Sample Size on the Performance of Species Distribution Models.
Diversity and Distributions. 14(5): 763–773. https://doi.org/10.1111/j.1472-
4642.2008.00482.x.
Woodward, F. I. and B. G. Williams. 1987. Theory and Models in Vegetation Science.
Vegetatio. 69(1&3): 189-197. https://www.jstor.org/stable/20038116.
Yang, X., S. Kushwaha, S. Saran, J. Xu, and P. Roy. 2013. MaxEnt Modeling for
Predicting the Potential Distribution of Medicinal Plant, Justicia Adhatoda L. in
Lesser Himalayan Foothills. Ecological Engineering 5: 83–87.
https://doi.org/10.1016/j.ecoleng.2012.12.004.
Yackulic, C. B., R. Chandler, E. F. Zipkin, J. A. Royle, J. D. Nichols, E. H. C. Grant, and
S. Veran. 2013. Presence-only Modelling Using MaxEnt: When can we Trust
the Inferences? Methods in Ecology and Evolution. 4(3): 236–243.
https://doi.org/10.1111/2041-210x.12004.

103
Yiwen, Z., L. B. Wei, and D. C. J. Yeo. 2016. Novel Methods to Select Environmental
Variables in MaxEnt: A Case Study Using Invasive Crayfish. Ecological
Modelling. 341: 5–13. https://doi.org/10.1016/j.ecolmodel.2016.09.019.
Zimmerman, N. E., N. G. Yoccoz, T. C. Edwards, E. S. Meier, W. Thuiller, A. Guisan,
D. R. Schmatz, and P. B. Pearman. 2009. Climatic Extremes Improve Predictions
of Tree Species. PNAS. 106(2): 19726-19728.
https://www.pnas.org/content/pnas/106/Supplement_2/19723.full.pdf

104
VITA
Sam Driver attended Woden High School, Woden Texas from 2004 to 2007. In
the fall of 2007, he began his undergraduate degree at Texas State University in San
Marcos, Texas, finishing the degree of Bachelor of Science in Biology with a minor in
Chemistry in 2012. In January 2018, he entered the Graduate School of Stephen F. Austin
State University. During the summer of 2019, the U.S. National Park Service at Zion
National Park employed him as an intern working under the Environmental Protection
Specialist. During the spring of 2020 he was employed as the GIS intern for Conservation
Equity Partners in Nacogdoches, Texas. He completed his graduate studies from Stephen
F Austin in the fall of 2020 with the degree of Master of Science in Environmental
Science with a minor in Spatial Science.
This thesis was typed by Samuel M. Driver


















