
Special Issue Article -...
10
Received: 9 June 2010, Revised: 15 September 2010, Accepted: 15 October 2010, Published online in Wiley Online Library: 2011 Structure and assembly–disassembly properties of wild-type transthyretin amyloid protofibrils observed with atomic force microscopy y Ricardo H. Pires a,b * , Maria J. Saraiva b,c , Ana M. Damas b,c ** and Miklo ´ s S. Z. Kellermayer a *** Transthyretin (TTR) is an important human transport protein present in the serum and the cerebrospinal fluid. Aggregation of TTR in the form of amyloid fibrils is associated with neurodegeneration, but the mechanisms of cytotoxicity are likely to stem from the presence of intermediate assembly states. Characterization of these intermediate species is therefore essential to understand the etiology and pathogenesis of TTR-related amyloidoses. In the present work we used atomic force microscopy to investigate the morphological features of wild-type (WT) TTR amyloid protofibrils that appear in the early stages of aggregation. TTR protofibrils obtained by mild acidification appeared as flexible filaments with variable length and were able to bind amyloid markers (thioflavin T and Congo red). Surface topology and contour-length distribution displayed a periodic pattern of 15 nm, suggesting that the protofibrils assemble via an end-binding oligomer fusion mechanism. The average height and periodic substructure found in protofibrils is compatible with the double-helical model of the TTR amyloid protofilament. Over time protofibrils aggregated into bundles and did not form mature amyloid-like fibrils. Unlike amyloid fibrils that are typically stable under physiological conditions, the bundles dissociated into component protofibrils with axially compacted and radially dilated structure when exposed to phosphate-buffered saline solution. Thus, WT TTR can form metastable filamentous aggregates that may represent an important transient state along the pathway towards the formation of cytotoxic TTR species. Copyright ß 2011 John Wiley & Sons, Ltd. Keywords: transthyretin; atomic force microscopy; protofibril; periodicity; amyloid; fibrillogenesis; cytotoxic oligomers INTRODUCTION Amyloidoses encompass a wide spectrum of highly debilitating disorders involving systemic pathological lesions, neurodegen- eration and several other tissue-specific dysfunctions (Chiti and Dobson, 2006). In these disorders, perturbation of the protein’s native fold results in aggregation, which ultimately leads to the formation of large deposits constituted mostly of amyloid fibrils. A related misfolding process is thought to result in the amyloidogenic conversion of transthyretin (TTR). TTR is a homotetrameric protein synthesized in the liver and the choroid plexus of the brain which is then exported to the blood serum and the cerebrospinal fluid, respectively (Fleming et al., 2009). Aggregation of TTR in the form of amyloid fibrils that deposit in extracellular space leads to a condition known as senile systemic amyloidosis in the case of wild type (WT) TTR (Westermark et al., 1990), while mutations in the TTR sequence are often associated with familial amyloidotic polyneuropathy (FAP) (Sousa and Saraiva, 2003). In the case of WT TTR and, similarly to other proteins, the in vitro amyloid fibrillogenesis is often initiated by acidifying the medium so as to mimic the lysosomal milieu (Colon and Kelly, 1992). The lysosome is a mildly acidic (pH 5) cytoplasmic vesicle whose housekeeping activity of degrading subcellular structures has been associated with amyloidogenic disorders (Nixon, 2007). (wileyonlinelibrary.com) DOI:10.1002/jmr.1112 Special Issue Article * Correspondence to: R. H. Pires, Department of Biophysics and Radiation Biology, Faculty of Medicine, Semmelweis University, Tu ˝zolto ´ u. 37-47, Buda- pest IX, H1094, Hungary. E-mail: [email protected] ** Correspondence to: A. M. Damas, IBMC-Institute for Molecular and Cell Biology; Rua do Campo Alegre, 823, 4150-180 Porto, Portugal. E-mail: [email protected] *** Correspondence to: M. S. Z. Kellermayer, Department of Biophysics and Radiation Biology, Faculty of Medicine, Semmelweis University, Tu ˝zolto ´ u. 37-47. Budapest IX, H1094, Hungary. E-mail: [email protected] a R. H. Pires, M. S. Z. Kellermayer Department of Biophysics and Radiation Biology, Faculty of Medicine, Semmelweis University, Tu ˝zolto ´ u. 37-47, Budapest IX, H1094 Hungary b A. M. Damas, R. H. Pires, M. J. Saraiva IBMC-Institute for Molecular and Cell Biology; Rua do Campo Alegre, 823, 4150-180 Porto, Portugal c A. M. Damas, M. J. Saraiva ICBAS-Instituto de Cie ˆncias Biome ´dicas de Abel Salazar, Universidade do Porto, Largo Prof. Abel Salazar, 2, 4099-003 Porto, Portugal y This article is published in Journal of Molecular Recognition as a focus on AFM on Life Sciences and Medicine, edited by Jean-Luc Pellequer and Pierre Parot (CEA Marcoule, Life Science Division, Bagnols sur Ce `ze, France). Abbreviations: AFM, atomic force microscopy; CAC, circular autocorrelation; DFT, discrete Fourier transform; FAP, familial amyloidotic polyneuropathy; FFT, fast Fourier transform; nCAC, normalized circular autocorrelation; PBS, phosphate buffered saline; pI, isoelectric point; ThT, thioflavin T; TTR, transthyretin; WT, wild type. J. Mol. Recognit. 2011; 24: 467–476 Copyright ß 2011 John Wiley & Sons, Ltd. 467
Transcript of Special Issue Article -...

Special Issue Article
Received: 9 June 2010, Revised: 15 September 2010, Accepted: 15 October 2010, Published online in Wiley Online Library: 2011
(wileyonlinelibrary.com) DOI:10.1002/jmr.1112
Structure and assembly–disassemblyproperties of wild-type transthyretin amyloidprotofibrils observed with atomic forcemicroscopyy
Ricardo H. Piresa,b*, Maria J. Saraivab,c, Ana M. Damasb,c**and Miklos S. Z. Kellermayera***
Transthyretin (TTR) is an important human transp
J. Mol. Rec
ort protein present in the serum and the cerebrospinal fluid.Aggregation of TTR in the form of amyloid fibrils is associated with neurodegeneration, but the mechanisms ofcytotoxicity are likely to stem from the presence of intermediate assembly states. Characterization of theseintermediate species is therefore essential to understand the etiology and pathogenesis of TTR-related amyloidoses.In the present work we used atomic force microscopy to investigate the morphological features of wild-type (WT) TTRamyloid protofibrils that appear in the early stages of aggregation. TTR protofibrils obtained by mild acidificationappeared as flexible filaments with variable length and were able to bind amyloid markers (thioflavin T and Congored). Surface topology and contour-length distribution displayed a periodic pattern of �15nm, suggesting that theprotofibrils assemble via an end-binding oligomer fusion mechanism. The average height and periodic substructurefound in protofibrils is compatible with the double-helical model of the TTR amyloid protofilament. Over timeprotofibrils aggregated into bundles and did not form mature amyloid-like fibrils. Unlike amyloid fibrils that aretypically stable under physiological conditions, the bundles dissociated into component protofibrils with axiallycompacted and radially dilated structure when exposed to phosphate-buffered saline solution. Thus, WT TTR can formmetastable filamentous aggregates that may represent an important transient state along the pathway towards theformation of cytotoxic TTR species. Copyright � 2011 John Wiley & Sons, Ltd.
Keywords: transthyretin; atomic force microscopy; protofibril; periodicity; amyloid; fibrillogenesis; cytotoxic oligomers
* Correspondence to: R. H. Pires, Department of Biophysics and RadiationBiology, Faculty of Medicine, Semmelweis University, Tuzolto u. 37-47, Buda-pest IX, H1094, Hungary.E-mail: [email protected]
** Correspondence to: A. M. Damas, IBMC-Institute for Molecular and CellBiology; Rua do Campo Alegre, 823, 4150-180 Porto, Portugal.E-mail: [email protected]
*** Correspondence to: M. S. Z. Kellermayer, Department of Biophysics andRadiation Biology, Faculty of Medicine, Semmelweis University, Tuzolto u.37-47. Budapest IX, H1094, Hungary.E-mail: [email protected]
a R. H. Pires, M. S. Z. Kellermayer
Department of Biophysics and Radiation Biology, Faculty of Medicine,
Semmelweis University, Tuzolto u. 37-47, Budapest IX, H1094 Hungary
b A. M. Damas, R. H. Pires, M. J. Saraiva
IBMC-Institute for Molecular and Cell Biology; Rua do Campo Alegre, 823,
4150-180 Porto, Portugal
c A. M. Damas, M. J. Saraiva
ICBAS-Instituto de Ciencias Biomedicas de Abel Salazar, Universidade do
Porto, Largo Prof. Abel Salazar, 2, 4099-003 Porto, Portugal
y This article is published in Journal of Molecular Recognition as a focus on AFMon Life Sciences and Medicine, edited by Jean-Luc Pellequer and Pierre Parot(CEA Marcoule, Life Science Division, Bagnols sur Ceze, France).
Abbreviations: AFM, atomic force microscopy; CAC, circular autocorrelation; DFT,
discrete Fourier transform; FAP, familial amyloidotic polyneuropathy; FFT, fast Fourier
transform; nCAC, normalized circular autocorrelation; PBS, phosphate buffered
saline; pI, isoelectric point; ThT, thioflavin T; TTR, transthyretin; WT, wild type. 4
INTRODUCTION
Amyloidoses encompass a wide spectrum of highly debilitatingdisorders involving systemic pathological lesions, neurodegen-eration and several other tissue-specific dysfunctions (Chiti andDobson, 2006). In these disorders, perturbation of the protein’snative fold results in aggregation, which ultimately leads tothe formation of large deposits constituted mostly of amyloidfibrils. A related misfolding process is thought to result in theamyloidogenic conversion of transthyretin (TTR). TTR is ahomotetrameric protein synthesized in the liver and the choroidplexus of the brain which is then exported to the blood serumand the cerebrospinal fluid, respectively (Fleming et al., 2009).Aggregation of TTR in the form of amyloid fibrils that deposit inextracellular space leads to a condition known as senile systemicamyloidosis in the case of wild type (WT) TTR (Westermark et al.,1990), while mutations in the TTR sequence are often associatedwith familial amyloidotic polyneuropathy (FAP) (Sousa andSaraiva, 2003).In the case of WT TTR and, similarly to other proteins, the
in vitro amyloid fibrillogenesis is often initiated by acidifying themedium so as to mimic the lysosomal milieu (Colon and Kelly,1992). The lysosome is a mildly acidic (pH 5) cytoplasmic vesiclewhose housekeeping activity of degrading subcellular structureshas been associated with amyloidogenic disorders (Nixon, 2007).
ognit. 2011; 24: 467–476 Copyright � 2011 John Wiley & Sons, Ltd.
67

R. H. PIRES ET AL.
468
The acidic environment leads to the disassembly of TTR’squaternary structure followed by partial unfolding of themonomer that becomes prone to aggregation (Lai et al., 1996;Foss et al., 2005). The acidification process has been proposed tobe sufficient for initiating the assembly of TTR amyloid fibrils(Colon and Kelly, 1992). However, the process of fibrillogenesisdepends on various additional conditions as well, such astemperature, protein concentration, ionic strength, agitation andpH. Manipulation of some of these parameters has been shown tolead to the appearance of various intermediate aggregationstates prior to the appearance of mature amyloid fibrils (Kodaliand Wetzel, 2007). These intermediate states have been receivingincreased attention because they possess much greater cytotoxicpotential than the fibrils (Caughey and Lansbury, 2003).One of the amyloid fibrillogenic intermediate states is
the protofibril, which can be frequently found in diverseamyloidogenic protein preparations and displays high cytotoxicactivity (Caughey and Lansbury, 2003). The term ‘protofibril’ isused in the literature rather loosely, often referring to a widerange of intermediates sometimes irrespective of their mor-phology. Here we adopt the terms ‘oligomer’, ‘protofibril’,‘protofilament’, and ‘fibril’ as defined in a recent review (Kodaliand Wetzel, 2007). Accordingly, protofibrils are flexible linearaggregates approximately 500 nm in length, andmay (but usuallydo not) contain a periodic substructure. Protofibrils may be thedirect precursors of protofilaments which, via a hierarchicalassembly process, give rise to amyloid fibril structure (Khuranaet al., 2003). In fact, in the case of the Aß1–40 peptide, thepresence of protofibril-specific antibodies in the aggregationreaction effectively precludes the formation of mature amyloidfibrils, leading to the accumulation of protofibrils (Habicht et al.,2007). In contrast, in the case of ß2-microglobulin (ß2m), undercertain conditions the in vitro fibrillation reaction leads to theaccumulation of protofibrils without the formation of matureamyloid fibrils (Kad et al., 2001; Gosal et al., 2005). Furthermore,these protofibrils are unable to accelerate the assembly ofamyloid fibrils under mature fibril-forming conditions, suggestingthat they might be ‘off-pathway’ kinetic traps (Kad et al., 2001).In the case of TTR the structural features of intermediate states
is relatively unexplored despite their high cytotoxic activity(Sousa et al., 2001; Sousa et al., 2002). Detailed structural studieshave been carried out only down to the level of the TTR amyloidprotofilament for which two main models have been proposed.One of them suggests that the protofilament is composed by asingle array of monomers (Inouye et al., 1998), that laterallyassociate via a hierarchical assembly mechanism to form amyloidfibrils (Cardoso et al., 2002). Another model proposes a doublehelical structure containing an 11.55 nm repeat (Blake andSerpell, 1996; Blake et al., 1996).Since protofibrils have been proposed to direct precursors of
amyloid fibrils displaying cytotoxic activity, studies addressingtheir structure and assembly dynamics are relevant in under-standing their cytotoxic behavior as well as their role withinthe amyloid aggregation pathway. However, so far no study ofamyloid protofibrils has been undertaken with high enoughresolution to allow the direct comparison of their morphologywith structural models of the amyloid protofilament. In thecurrent study we analyzed the morphological properties of TTRamyloid protofibrils with atomic force microscopy (AFM) underliquid buffer conditions. We show that protofibrils formed bymildacidification display characteristics that are more closely relatedto the double-helical model of the amyloid protofilament. These
wileyonlinelibrary.com/journal/jmr Copyright � 2011 John Wiley
structures are able to bind thioflavin T and Congo red and appearto grow via an oligomer fusion mechanism. However, maturationof the protofibrils into amyloid-like fibrils was not seen on a timescale of several months. Rather, they showed a tendency to formunorganized bundles that undergo dissociation at neutral pH.Thus, the protofibrils described in our work possibly representa distinct structural entity whose potential cytotoxic activity—ahallmark of oligomeric amyloid species—may be modulated byenvironmental conditions that either preserve their protofibrillarstate, or induce their dissociation.
MATERIALS AND METHODS
Protein purification
Recombinant wild type (WT) TTR expressed in BL21 E. coli cellswas isolated and purified as described previously (Almeida et al.,1997) followed by high-affinity anion exchange chromatography(MonoQ column, GE Healthcare) equilibrated with 100mMBisTris pH 6.8. The protein eluting with approximately 150mMNaCl was subsequently concentrated and loaded onto acalibrated analytical size exclusion chromatography superdexS75 column (GE Healthcare) equilibrated with 50mM HEPES pH7.0 and 150mM NaCl. Protein eluting in a peak corresponding totetrameric TTR (55 kDa) was collected and dialyzed overnightagainst a weakly buffered solution (10mM HEPES pH 7.0).The protein was then concentrated to 10–15mg/ml. Proteinconcentration was determined with the Bradford method(BioRad). Samples were approximately 98% pure as judged byCoomassie-blue staining of SDS-PAGE gels. According to dynamiclight scattering measurements (Zetasizer Nano ZS, Malvern) thesamples were considered monodisperse. The hydrodynamicradius of the particles ranged between 6.4 and 6.8 nm, consistentwith previous observations (Hou et al., 2007).
Sample preparation
TTR protofibrils were prepared by incubating WT TTR in 50mMsodium acetate buffer at pH 3.6, 378C and at a concentration of1mg/ml for a period of 1 year. At several time points followingacidification, samples were taken and diluted before surfacedeposition to reduce molecular overcrowding. Thus, sampleswere diluted 500-fold in acetate buffer at pH 3.6 and imagedas described below. To test the stability of protofibrils at morephysiological conditions, samples in an advanced state ofaggregation were diluted 25-fold with phosphate-buffered salinesolution (PBS), followed by incubation at room temperaturefor 1min. Subsequently, the samples were diluted 20-fold inultrapure water (�pH 6.3) prior to further investigation.
Atomic force microscopy
AFM imaging was carried out by using procedures reported inprevious papers on Aß fibrils (Karsai et al., 2005; Kellermayer et al.,2005; Karsai et al., 2006; Karsai et al., 2007; Karsai et al., 2008;Kellermayer et al., 2008) with modifications. Hundred microlitersof the diluted WT TTR sample was deposited on freshly cleavedmica, incubated for 5min at room temperature; the surface wasthen rinsed with buffer. Imaging was carried out on a daily basisover a period of 2 months and then periodically for up to 1 year.AFM images of the samples were acquired with an MFP-3D AFMinstrument (Asylum Research, Santa Barbara, CA) in non-contact
& Sons, Ltd. J. Mol. Recognit. 2011; 24: 467–476

WILD-TYPE TTR PROTOFIBRIL STRUCTURE
(AC) mode. Imaging was done by scanning in liquid, either in50mM acetate buffer, pH 3.6 or in ultrapure water for sampleswhich were incubated in PBS. Imaging was performed by usinglow-noise cantilevers (Biolever, Lever A, Olympus) with a springconstant of 30 pN/nm and a resonance frequency of �9.2 kHz.The free amplitude was set to�0.3 V and the amplitude set pointto �0.2 V, and the images were recorded at a typical scanningfrequency of �0.8 Hz.
Image processing and analysis
Raw AFM images were flattened by using a linear functionfollowed by masking out particles with heights above 50 pmand applying another first-order flattening to the (unmasked)background to correct for artifacts arising from the first flatteningstep. For any given image, a distribution of pixel heights wasused to find the background ‘height’ relative to which all heightmeasurements were carried out. Images were smoothed withtwo passes of a Gaussian convolution filter with one pixelneighborhood.Protofibril length was measured by taking the tip convolution
effect into account (Figure 1). The cantilever tip was approxi-mated by a hemisphere with a radius (R) of 30 nm (manufacturer’sspecifications) and the protofibrils as cylinders with diameter (h)corresponding to the average protofibril height. Protofibril length(L) was obtained from the measured fibril length (M) as:
L ¼ M�2ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi2hR�h2
p: (1)
For small oligomers that appeared nearly spherical under theAFM, the molecular volume (V) was calculated by measuring theheight (h) and taking the full width at half height as the diameter(d) which significantly compensates for the tip convolution effectand these were used as follows (Carnally et al., 2008):
V ¼ ph
6h2 þ 3d2
4
� �(2)
The number of TTR monomers present in a given oligomerwas estimated by considering the dimensions of the native WTTTR monomer: 2� 3� 4.5 nm (Blake et al., 1978) or 27 nm3.Statistical analysis of periodicity was carried out on severalprotofibrillar axial sections which exhibited a clearly corrugatedstructure. The data were analyzed in two independent ways.In the first, the distance between consecutive local heightmaxima or shoulders was measured, based on which a histogramof periodicities was constructed. In the second, analytical app-roaches (autocorrelation and Fourier transformation) wereapplied to axial height sections of 20 protofibrils taken fromhigh-resolution (1024� 1024 pixel) images. The height valueswere treated as a discrete homogeneous periodic signal by
Figure 1. Schematics of correction for AFM tip convolution in calculat-
ing the real length of TTR protofibrils.
J. Mol. Recognit. 2011; 24: 467–476 Copyright � 2011 John Wiley & S
ignoring the slant height profile at the protofibril termini andconsidering only the harmonic component of the axial topology.Thus, for any given harmonic segment of an axial section theheight variations (z) can be approximated as
zðdÞ ¼ hþ A� sinð2p� v� d þ uÞ; (3)
where h is the average height of the protofibril, A is theheight amplitude, v is the frequency, d is the distance alongthe protofibril axis, and u is the phase. To obtain a statisticalrepresentation of the major frequency components in protofibrilperiodicity, an amplified waveform signal f(d) was obtained bysumming each i axial height section decremented by theirindividual average height as
f ðdÞ ¼Xni¼1
zi dð Þ�hi� �
¼Xni¼1
Ai � sin 2p� vi � d þ uð Þð Þ; (4)
where d2N0, 0� d� 99. To allow amplification of the periodicsignal by limiting destructive phase interference, zi(0) was set atu¼ 908 which gives zið0Þ ¼ hi þ Ai . Since the periodic pattern isexpected to repeat itself over the analyzed protofibril length(99 nm), a circular autocorrelation (CAC) was then obtained fromf(d) of protofibrils grown in pH 3.6 and from f(d) of protofibrilsobtained upon disassembly of bundles after PBS incubation.To allow direct comparison between the two data sets a powerspectral density (PSD) was obtained from a normalized CAC(nCAC). Thus, a discrete Fourier transform (DFT) of nCAC wascomputed through a fast Fourier transform (FFT) algorithmwith ahanning window function and the data analyzed betweenperiods of 2–50 nm. All analyses were performed by using IGORPro v6.06 (Wavemetrics, Oregon, USA).
Measurement of spectral properties
Thioflavin T Assay: 10mM thioflavin-T (ThT) (e441¼ 2.2�104M�1 cm�1) stock solution was prepared fresh in ThT buffer(50mM glycine/NaOH, pH 9.0). The solution was passed througha 0.22mm PVDF filter and stored in the dark and on ice. NativeWT TTR protein solution and protofibril suspension (1mg/ml ofprotein) were diluted 10-fold in ThT buffer. Emission was detectedat 482 nm with the excitation and emission slits set to 5 nm andthe spectrum recorded at room temperature on a Perkin-ElmerLS50B fluorescence spectrometer.Congo Red Redshift Assay: Measurements were carried out
as reported earlier (Bonifacio et al., 1996). Briefly, 5ml of TTRaggregates (1mg/ml of protein) were mixed with 55ml of a10mM Congo red solution prepared in 100mM Tris/HCl, pH 7.0.Spectra were recorded at room temperature in a ShimadzuUV-2401 PC spectrophotometer.
4
RESULTS
High-resolution AFM investigation of WT TTR protofibrils wascarried out in the present work. Amyloidogenic transformation ofWT TTR was induced by acidification of the buffer solution. Withinthe first week of incubation it was possible to observe monomersand different size oligomeric structures (Figure 2A, inset) aswell as short flexible fibrillar structures of worm-like shape andnodular morphology (Figure 2A). While monomers/dimers werepresent throughout the entire incubation period (note thebackground in Figures 2C and 3A), the number of oligomers
ons, Ltd. wileyonlinelibrary.com/journal/jmr
69

Figure 2. Morphology and spectroscopic properties of WT TTR aggregates formed by incubation at pH 3.6. (A) WT TTR imaged by AFM on the 5th day
of incubation showing the presence of small protofibrils and oligomers (image 1024� 512 pixels). Inset: magnified view of oligomeric structures (arrows)
with the indication of the estimated number of TTR monomers. (B) On the 6th day of incubation protofibrils appear longer, seem more frequent thansmall oligomers, and already display some tendency to associate (image 1024� 1024 pixels). Inset: magnified view of a protofibril displaying periodic
axial topographical substructure; the green line indicates trajectory along which the profile plot was obtained (see Figure 3D). (C) After 2 weeks of
incubation protofibrils show increased tendency to associate via their termini, forming a network containing nodules from which protofibrils extend in
quasi-radial symmetry as highlighted by circles (image 1024� 2014 pixels). (D) 10 weeks of incubation lead to a formation of protofibril networks ofincreased density forming protofibril bundles (image 512� 512 pixels). (E) Fluorescence excitation spectra of Thioflavin T only (green), in the presence of
nativeWT TTR (blue), and in the presence ofWT TTR protofibrils (red). A red shift is well observable in the latter case as expected for an amyloid aggregate.
(F) UV/VIS spectra of Congo red in the absence of protein (green), in the presence of native WT TTR (blue), and upon addition of protofibrils (red). The
presence of protofibrils induces a red shift in the absorbance spectrum that is characteristic of amyloid aggregates with a shoulder at 540 nm as seen fromthe difference spectrum (black). Comparatively, the presence of native WT TTR does not induce any spectral changes as seen from difference spectrum
(gray).
wileyonlinelibrary.com/journal/jmr Copyright � 2011 John Wiley & Sons, Ltd. J. Mol. Recognit. 2011; 24: 467–476
R. H. PIRES ET AL.
470

Figure 3. Morphological properties of protofibrils before and after transient (1min) incubation in PBS. (A) Protofibril bundle imaged 8 weeks after
sample incubation in 50mM sodium acetate at pH 3.6. (B) Incubation for 1min in PBS of the sample shown in (A) leads to the disassembly of bundles into
its constituting protofibrils. Inset: enlarged view of protofibrils displaying pronounced axial periodicity; the red line indicates trajectory along which
profile plot was obtained (see Figure 3D). (C) Contour length histogram of protofibrils in assembling conditions (pH 3.6, blue histogram) and following thedisassembly of bundles (PBS, red histogram). The arrowheads indicate local maxima spaced 15 nm apart for assembling protofibrils: 35, 50, 65, 80, 95 nm,
and spaced by 12 nm for protofibrils in disassembly conditions: 14, 26, 38 and 50 nm. (D) Topographical profile plots along the long axis of protofibrils in
assembling (blue, from Figure 2B) and disassembling (red, from Figure 3B) conditions; also indicated are the peak-to-peak distances in nanometers.
WILD-TYPE TTR PROTOFIBRIL STRUCTURE
4
decreased while the fibrillar structures became increasinglyprevalent (Figure 2B).We carried out a statistical analysis of the average topogra-
phical height of the filamentous structures. Because the observedwidth is inflated by cantilever tip convolution, it is the heightdata that was correlated with the thickness of the filaments. Thehistogram of height data (Figure 4A) displayed normal distri-bution with a mean of 3.2 nm (� 0.8 nm SD, n¼ 3505, 63filaments), and a height variation amplitude of 1.0 nm (�0.3 nmSD, n¼ 607, 127 filaments). Based on the thickness of thefilamentous structures, which is smaller than that of TTR amyloidfibrils which are typically 10–12 nm (Serpell et al., 1995; Serpellet al., 2000), we identify them as protofibrils and will refer tothem as such. Analysis of protofibril contour lengths revealed awide distribution between 30 and 300 nm (Figure 3C). In thecontour-length histogram local maxima can be observed, whichare separated by approximately 15 nm (arrowheads in Figure 3C).The discrete distribution indicates that the length of theprotofibrils is an integer multiple of subunits with �15 nmdimensions. Protofibrils may therefore be considered as alinear assembly of 15-nm-long subunits. To further test for thispossibility we analyzed the height topography of the protofibrilsalong the longitudinal axis. In the axial profile plot (Figure 3D,blue curve) periodically spaced peaks and valleys can bediscerned. Statistical analysis of the peak-to-peak distance
J. Mol. Recognit. 2011; 24: 467–476 Copyright � 2011 John Wiley & S
(Figure 4B, blue histogram) shows an average distance of15 nm (�0.2 nm SD, n¼ 266, 45 protofibrils). Thus, the protofibrilsindeed seem to be linear chains of subunits with axial dimensionsof �15 nm. Given their large dimensions compared with thesize of monomers, these aggregation subunits are likely to beoligomers themselves.Already within the first week, but extending throughout
the whole observation period, protofibrils showed a tendency toassociate into higher-order aggregates. An example of such anaggregate, recorded in the second week of incubation, is shownin Figure 2C. The figure displays an elongated structure withapproximately 650 nm in length that appears to result fromthe association of several �100-nm-long protofibrils. Protofibrilsassociate mostly via their termini, forming axial aggregates withadditional protofibrils annealing in a quasi-radial symmetry andextending away from central nodules (Figure 2C, circles). Thistype of architecture increased in size and complexity as a functionof incubation time, but it was not accompanied by an increase inorder. This is seen for example on an image obtained on the 10thweek of incubation where an arrangement that is more similar toa protofibril bundle, rather than an amyloid fibril, can be seen(Figure 2D). Although the sample was imaged for up to one yearafter the start of incubation, no noteworthy changes whereobserved henceforth; isolated protofibrils became scarcer andonly large bundles (�1–4mm2) were routinely present.
ons, Ltd. wileyonlinelibrary.com/journal/jmr
71
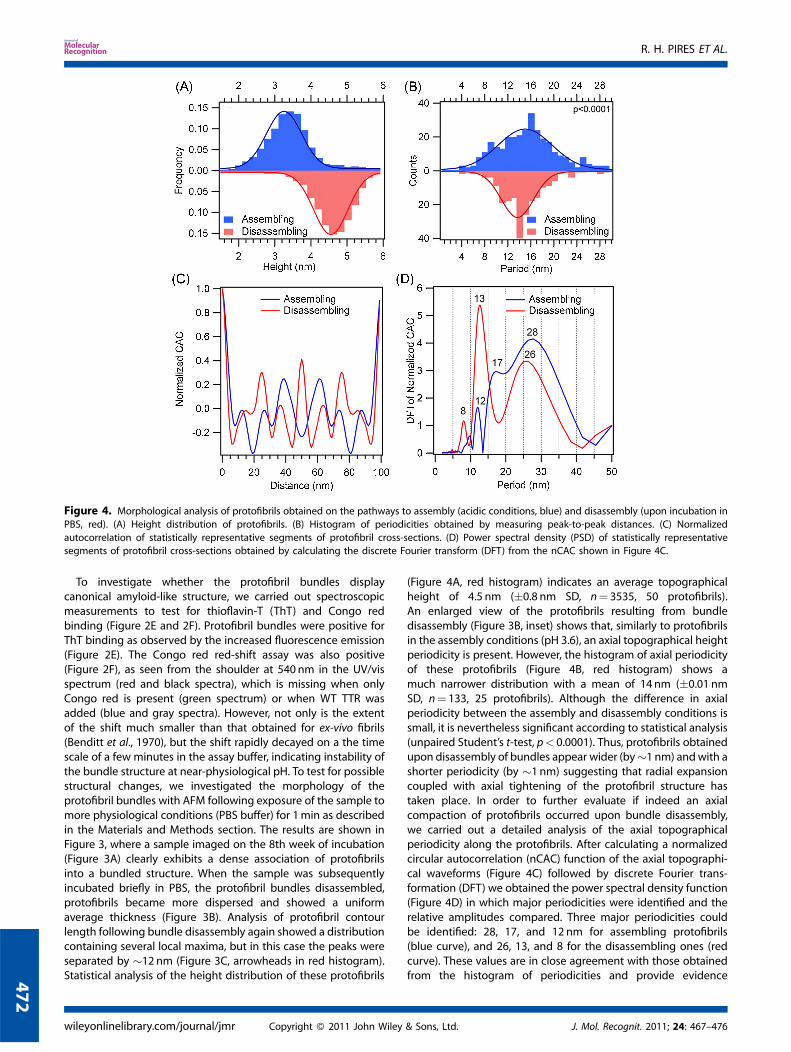
Figure 4. Morphological analysis of protofibrils obtained on the pathways to assembly (acidic conditions, blue) and disassembly (upon incubation in
PBS, red). (A) Height distribution of protofibrils. (B) Histogram of periodicities obtained by measuring peak-to-peak distances. (C) Normalized
autocorrelation of statistically representative segments of protofibril cross-sections. (D) Power spectral density (PSD) of statistically representativesegments of protofibril cross-sections obtained by calculating the discrete Fourier transform (DFT) from the nCAC shown in Figure 4C.
R. H. PIRES ET AL.
472
To investigate whether the protofibril bundles displaycanonical amyloid-like structure, we carried out spectroscopicmeasurements to test for thioflavin-T (ThT) and Congo redbinding (Figure 2E and 2F). Protofibril bundles were positive forThT binding as observed by the increased fluorescence emission(Figure 2E). The Congo red red-shift assay was also positive(Figure 2F), as seen from the shoulder at 540 nm in the UV/visspectrum (red and black spectra), which is missing when onlyCongo red is present (green spectrum) or when WT TTR wasadded (blue and gray spectra). However, not only is the extentof the shift much smaller than that obtained for ex-vivo fibrils(Benditt et al., 1970), but the shift rapidly decayed on a the timescale of a few minutes in the assay buffer, indicating instability ofthe bundle structure at near-physiological pH. To test for possiblestructural changes, we investigated the morphology of theprotofibril bundles with AFM following exposure of the sample tomore physiological conditions (PBS buffer) for 1min as describedin the Materials and Methods section. The results are shown inFigure 3, where a sample imaged on the 8th week of incubation(Figure 3A) clearly exhibits a dense association of protofibrilsinto a bundled structure. When the sample was subsequentlyincubated briefly in PBS, the protofibril bundles disassembled,protofibrils became more dispersed and showed a uniformaverage thickness (Figure 3B). Analysis of protofibril contourlength following bundle disassembly again showed a distributioncontaining several local maxima, but in this case the peaks wereseparated by �12 nm (Figure 3C, arrowheads in red histogram).Statistical analysis of the height distribution of these protofibrils
wileyonlinelibrary.com/journal/jmr Copyright � 2011 John Wiley
(Figure 4A, red histogram) indicates an average topographicalheight of 4.5 nm (�0.8 nm SD, n¼ 3535, 50 protofibrils).An enlarged view of the protofibrils resulting from bundledisassembly (Figure 3B, inset) shows that, similarly to protofibrilsin the assembly conditions (pH 3.6), an axial topographical heightperiodicity is present. However, the histogram of axial periodicityof these protofibrils (Figure 4B, red histogram) shows amuch narrower distribution with a mean of 14 nm (�0.01 nmSD, n¼ 133, 25 protofibrils). Although the difference in axialperiodicity between the assembly and disassembly conditions issmall, it is nevertheless significant according to statistical analysis(unpaired Student’s t-test, p< 0.0001). Thus, protofibrils obtainedupon disassembly of bundles appear wider (by�1 nm) andwith ashorter periodicity (by �1 nm) suggesting that radial expansioncoupled with axial tightening of the protofibril structure hastaken place. In order to further evaluate if indeed an axialcompaction of protofibrils occurred upon bundle disassembly,we carried out a detailed analysis of the axial topographicalperiodicity along the protofibrils. After calculating a normalizedcircular autocorrelation (nCAC) function of the axial topographi-cal waveforms (Figure 4C) followed by discrete Fourier trans-formation (DFT) we obtained the power spectral density function(Figure 4D) in which major periodicities were identified and therelative amplitudes compared. Three major periodicities couldbe identified: 28, 17, and 12 nm for assembling protofibrils(blue curve), and 26, 13, and 8 for the disassembling ones (redcurve). These values are in close agreement with those obtainedfrom the histogram of periodicities and provide evidence
& Sons, Ltd. J. Mol. Recognit. 2011; 24: 467–476

WILD-TYPE TTR PROTOFIBRIL STRUCTURE
that upon disassembly of bundles the resulting protofibrils arecompacted and display reduced periodicities.
4
DISCUSSION
In the present work we investigated the assembly–disassemblypathway of transthyretin amyloidogenesis with high-resolutionnon-contact atomic-force microscopic imaging under liquidconditions. The commitment of WT TTR towards the amyloido-genetic pathway was evoked by lowering the pH of the bufferto 3.6. Acidification has been shown in the past to induce theassembly of WT TTR into filamentous aggregates that exhibitmorphological, structural, and chemical properties similar tothose of tissue-formed amyloid fibrils (Bonifacio et al., 1996).In recent years it has become clear that the aggregation reactionin amyloidogenic systems is more complex than previouslythought, involving the assembly of oligomeric and protofibrilstructures with very heterogeneous morphologies. The involve-ment of these species in amyloid fibril assembly is still debated(Kodali and Wetzel, 2007). Because these pre-fibrillar speciesappear to be the key agents in amyloid cytotoxicity, it isimportant to characterize their structural properties in full detail.Here we described the appearance of nodular protofibrillar
species in the amyloidogenic aggregation of TTR (Figure 2Band 2C), with a topographical height that averaged 3.2 nmand ranged between 2.7 and 3.7 nm (1 nm height amplitude)(Figure 4A). The length repeat is consistent with the observedtopographical height periodicity (Figure 4B) and suggests thatprotofibrils grow in stepwise fashion by the fusion of �15 nmoligomers with the free end of the protofibril. Such an oligomerfusion mechanism has been observed for other types of amyloidprotofibrils (Modler et al., 2003; Modler et al., 2004; Carrottaet al., 2005; Hill et al., 2009). Similarly, we observe that the earlyprotofibrillar stages are characterized by the presence of smalloligomers (Figure 2A) that tend to disappear as protofibrils growin number. In this aggregation mechanism an oligomer is oneperiodic unit. Interestingly, in an earlier work, the pre-fibrillar statein the aggregation of WT TTR was reported to be populated by8-nm globular oligomers (Cardoso et al., 2002) the size of whichis approximately one half of the periodicity observed here.The 8 nm oligomers were found to disappear as 4–5 nm wideprotofibrils were formed, giving rise to higher order aggregatesof amyloid-like fibrils through hierarchical assembly (Cardosoet al., 2002). We did not observe the formation of amyloid fibrils.Instead, protofibrils appeared to bundle via their termini formingclusters displaying quasi-radial symmetry rather than by lateralassociation (Figure 2C). Protofibril bundles have been suggestedto form an intermediate state leading to the assembly of amyloidfibrils (Arimon et al., 2005). This does not appear to be the casewith WT TTR under the conditions used in this study, where atransition from protofibril bundles to mature fibrils was neverobserved even after 1 year of study. The samples neverthelesswere positive for ThT (Figure 2E) and Congo red (Figure 2F),which are classical amyloid markers. However, the spectral red-shiftof the Congo red absorption maximum decayed on the time-scaleof a few minutes, suggesting that disaggregation of the bundlestructure under conditions of the assay (neutral pH) may haveoccurred. This observation is in accordance with several reportsthat show that amyloid protofibrils and fibrils formed in acidicmedium can undergo depolymerization under neutral or alkalineconditions (Yamaguchi et al., 2001; Yamamoto et al., 2005).
J. Mol. Recognit. 2011; 24: 467–476 Copyright � 2011 John Wiley & S
The possibility of instability of TTR amyloid protofibril bundlesat neutral pH conditions prompted us to undertake a mor-phological study. As seen from the AFM images (Figure 3Aand 3B), by incubation of protofibrils for 1min in PBS resultedin the disassembly of bundles into individual protofibrils. Inaddition, small oligomers and monomers which populated thebackground when imaging was performed at acidic buffer are nolonger present, suggesting that, since TTR is an acidic proteinwith pI �5 (Connors et al., 1998), increasing the pH might affectthe protein surface charge to an extent that inhibits theiradsorption to the surface. The protofibrils thus obtained revealslightly altered structural features. The changes are characterizedby radial expansion, with the average height of protofibrilsincreasing from 3.2 to 4.5 nm after PBS exposure (Figure 4A),which can be partly explained by differences in electrostaticinteractions arising from the differences in pH and ionic strengthin the two samples (Muller and Engel, 1997). However, we alsoobserved that upon disassembly an axial compaction alsotook place, as seen from the decrease in axial periodicity from 15to 14 nm (Figure 4B). Although this difference is relatively small,it is statistically significant (p< 0.0001). In addition, while thecontour-length histogram of assembling protofibrils displays amultimodal distribution with local maxima spaced at integermultiples of �15 nm (Figure 3C, blue histogram), upondisassembly of bundles each mode of the distribution becomesseparated by �12 nm (Figure 3C, red histogram). Both resultssuggest that, upon disassembly a structural transition takes placethat induces a small but significant axial compaction of theprotofibrils. A similar observation can be made from the analysisof the power spectral density (PSD) of the statistical representa-tion of protofibrils (Figure 4D). The identified periodicities, 12, 17,and 28 nm (on assembly) and 8, 13, and 26 nm (on disassembly),are comparable to the values obtained from the statisticalmeasurement of peak-to-peak distances (Figure 4B). Since thetwo types of analyses are fundamentally different is not possibleto directly compare the magnitude of the power in a PSDcurve with the magnitude in a periodicity histogram (counts/frequency). However, the Fourier transform analysis of waveformsis an extremely sensitive approach to study periodic structures,allowing the identification of periods that could otherwisebe unnoticeable in a histogram derived from peak-to-peakdistances. While the histograms show a sensible shortening ofperiodicities, these changes are far more noticeable in the PSDanalysis where periods identified on protofibril disassembly aregenerally shorter than those in the assembly route (Figure 4D).On the assembly and disassembly PSD curves, only a centralperiod at 12–13 nm appears to be a common feature, albeit withdifferent power. Upon disassembly, the 17 nm periodicity presentin protofibrils during assembly is no longer apparent, and thischange is accompanied by a noticeable increase in the power ofthe 12–13 nm period. This shortening in periodicity by�4.5 nm iswithin the dimensions of a single TTR monomer (Blake et al.,1978). Thus, as suggested by the periodicity histogram, transientincubation of protofibrils in PBS leads to a tightening of theprotofibril structure, a process that might be coupled to theobserved radial expansion. Additional periodicities were alsoidentified by the PSD analysis that are weakly represented inthe periodicity histogram, notably the 28 nm as well as the 8and 26 nm periods found in assembling and disassemblingprotofibrils, respectively. The shorter periods are likely to beassociated with very small height variations as indicated by theirsmaller power in the PSD curves, and thus may have been
ons, Ltd. wileyonlinelibrary.com/journal/jmr
73

R. H. PIRES ET AL.
474
occasionally overlooked during peak-to-peak distance measure-ments. On the other extreme are the large periods of 26and 28 nm which are relatively large when compared to themeasured peak-to-peak distances, and may reflect a more globalmorphological feature of the protofibril structure on binding tothe surface. Altogether, the morphological changes observedlikely stem from molecular rearrangements at the level ofindividual monomers, underlying the structural dynamism thatis present within the TTR protofibril that could lead to thedisassembly of the protofibril bundles.Since it has been proposed that protofibrils could be the direct
precursors of the protofilament, it is important to compare ourmorphological data obtained here with the existing structuralmodels for the TTR amyloid protofilament. Analysis of electronmicrographs of ex vivo TTR amyloid fibril cross sections has revealedthat the protofilament is 4–5nm in diameter (Serpell et al., 1995).For the in vitro assembled TTR amyloid-like fibrils, protofibrilsmeasuring 4nm were also found to be the fundamental polymericunit (Cardoso et al., 2002). In this latter work, mass-per-lengthmeasurements in STEM data suggest that the unitary filamentousstructure is a linear array of monomers. This is in agreement with apreviousmodel for the TTRprotofilament based on X-ray diffractiondata obtained on ex vivo fibrils (Inouye et al., 1998). In the report byCardoso and others, no periodic substructure was resolved forprotofibrils (Cardoso et al., 2002). However, molecular dynamicssimulations for a TTR protofilament modeled as a series ofmonomers that form two extended b-sheets have proposed theexistence of a helical structure with a 48 b-strand repeat (Correiaet al., 2006), or a 23.04nm periodic unit considering 0.48 nm as theinter-strand distance. There is, however, another model for the TTRprotofilament also based on X-ray diffraction of ex vivo amyloidfibrils, which is described as double-helical structure containing a11.55 nm repeat andmeasuring 5 nm in height (Blake and Serpell,1996; Sunde et al., 1997).While the height of the protofibrils is in good agreement with
the diameter of the TTR protofilament, the periodicity of12–13 nm detected on the PSD analysis is very close to the11.55 nm repeat of the double-helical model, with a difference of�1 nm that is within the error of the measurements of our AFMimages in the XY-plane (1024� 1024 pixels for 1� 1mm images).Assuming an overall helical conformation for the protofibrilsdescribed here, as it has been proposed for protofibrils fromlithostathine (Gregoire et al., 2001) the 15–17 nm periodicityfound in protofibrils in assembly conditions, may result from amore relaxed conformation than that of the protofilaments whichare likely to have their structure more constrained by theremaining fibril structure. In addition, transient incubation of thesame protofibrils in near-physiological conditions of pH and ionicstrength, lead to a structural transition where the height andperiodicities match quite remarkably the dimensions proposedfor the double-helical model of the protofilament.Despite the morphological similarities between the TTR
protofibrils and the double-helical model of the protofilament,the protofibrils did not assemble into higher order aggregatesthat would resemble mature amyloid fibrils as seen from ex vivo
wileyonlinelibrary.com/journal/jmr Copyright � 2011 John Wiley
preparations. Thus, protofibrils, despite some morphologicalsimilarity to current protofilament models, may have a verydistinct structural organization that effectively precludes theirhierarchical assembly into amyloid fibrils and may therefore beconsidered ‘off-pathway’ products. However, it may also happenthat the protofibrils obtained here simply require an additionalfactor that promotes their self-assembly into amyloid fibrils.For example, ex vivo TTR amyloid fibrils are often found to containfragments of the protein that appear to influence fibrillarmorphology (Bergstrom et al., 2005). In addition, it is known thatthe in vitro and in vivo assembly can be stimulated by a variety ofcellular components, including proteoglycans, glucosaminogly-cans, lipids, or collagen (Relini et al., 2006; Relini et al., 2008; Naikiand Nagai, 2009). Inclusion of these elements in future experimentswill yield further insights into the mechanisms of TTR amyloidfibrillogenesis. Since the acidification of TTR induces the formationof cytotoxic species before the pre-fibrillar state, it is very likely thatthe protofibrils described here have a cytotoxic activity. The factthat theymanifest instability at physiological conditions is thereforelikely to be of relevance for the etiology of TTR-related amyloidoses.
CONCLUSIONS
We have described the morphological features of transthyretinprotofibrillar amyloid intermediates by using atomic forcemicroscopy under liquid conditions. The protofibrils, obtainedby mild acidification of the sample, display an axial periodicityrelated possibly to the oligomeric fusion mechanism of theirassembly. The protofibrils aggregate into irregular bundles on atime-scale of several weeks, but disperse into axially compactedand radially expanded structures upon neutralizing the pH. Theperiodicity observed here in protofibrils agrees with thepredictions of the double-helical model of TTR assembly. Despitedisplaying the hallmarks of amyloid features, protofibrils failed toaggregate into mature fibrils even during extended time periods.Thus, the protofibrils seen here likely represent a kineticallytrapped structural variant, for which a precise structuraldescription awaits further high-resolution investigations. Theaxial compaction and radial expansion indicate that theprotofibrils are nevertheless structurally dynamic, a fact thatmay be relevant for their potentially cytotoxic activity.
Acknowledgements
The authors are thankful to Andras Kaposi for insightful com-ments and suggestions. This work was supported by grantsfrom the Hungarian Science Foundation (OTKA K73256), theHungarian National Office of Research and Technology(NANOAMI KFKT-1– 2006–0021, OMFB-380/2006), and the Hun-garian Medical Research Council (ETT-229/09) to MSZK; andProject GRICES&FCT – Proj. 4.1.1-Hungary, Portugal and Projectn. 037525 EURAMY (FP6-LIFESCIHEALTH-6) from EU to AMD. RHPacknowledges the award of a short-term travel grant by theCalouste Gulbenkian Foundation of Portugal.
REFERENCES
Almeida MR, Damas AM, Lans MC, Brouwer A, Saraiva MJ. 1997. Thyroxinebinding to transthyretin Met 119. Comparative studies of differentheterozygotic carriers and structural analysis. Endocrine 6: 309–315.
Arimon M, Diez-Perez I, Kogan MJ, Durany N, Giralt E, Sanz F, Fernandez-Busquets X. 2005. Fine structure study of Abeta 1-42 fibrillogenesiswith atomic force microscopy. Faseb J. 19: 1344–1346.
& Sons, Ltd. J. Mol. Recognit. 2011; 24: 467–476

WILD-TYPE TTR PROTOFIBRIL STRUCTURE
Benditt EP, Eriksen N, Berglund C. 1970. Congo red dichroism withdispersed amyloid fibrils, an extrinsic cotton effect. Proc. Natl Acad.Sci. USA 66: 1044–1051.
Bergstrom J, Gustavsson A, Hellman U, Sletten K, Murphy CL, Weiss DT,Solomon A, Olofsson BO, Westermark P. 2005. Amyloid depositsin transthyretin-derived amyloidosis: cleaved transthyretin isassociated with distinct amyloid morphology. J. Pathol. 206:224–232.
Blake C, Serpell L. 1996. Synchrotron X-ray studies suggest that the core ofthe transthyretin amyloid fibril is a continuous beta-sheet helix.Structure 4: 989–998.
Blake CC, Geisow MJ, Oatley SJ, Rerat B, Rerat C. 1978. Structureof prealbumin: secondary, tertiary and quaternary interactionsdetermined by Fourier refinement at 1.8 A. J. Mol. Biol. 121:339–356.
Blake CC, Serpell LC, Sunde M, Sandgren O, Lundgren E. 1996. A molecularmodel of the amyloid fibril. Ciba Found Symp. 199(6-15): discussion15-21, 16–40.
Bonifacio MJ, Sakaki Y, Saraiva MJ. 1996. ‘In vitro’ amyloid fibrilformation from transthyretin: the influence of ions and theamyloidogenicity of TTR variants. Biochim. Biophys. Acta 1316:35–42.
Cardoso I, Goldsbury CS, Muller SA, Olivieri V, Wirtz S, Damas AM,Aebi U, Saraiva MJ. 2002. Transthyretin fibrillogenesis entailsthe assembly of monomers: a molecular model for in vitroassembled transthyretin amyloid-like fibrils. J. Mol. Biol. 317:683–695.
Carnally SM, Dev HS, Stewart AP, Barrera NP, Van Bemmelen MX, Schild L,Henderson RM, Edwardson JM. 2008. Direct visualization of thetrimeric structure of the ASIC1a channel, using AFM imaging.Biochem. Biophys. Res. Commun. 372: 752–755.
Carrotta R, Manno M, Bulone D, Martorana V, San BiagioPL., 2005.Protofibril formation of amyloid beta-protein at low pH via anon-cooperative elongation mechanism. J. Biol. Chem. 280: 30001–30008.
Caughey B, Lansbury PT. 2003. Protofibrils, pores, fibrils, and neurode-generation: separating the responsible protein aggregates from theinnocent bystanders. Annu. Rev. Neurosci. 26: 267–298.
Chiti F, Dobson CM. 2006. Protein misfolding, functional amyloid, andhuman disease. Annu. Rev. Biochem. 75: 333–366.
Colon W, Kelly JW. 1992. Partial denaturation of transthyretin issufficient for amyloid fibril formation in vitro. Biochemistry 31:8654–8660.
Connors LH, Ericsson T, Skare J, Jones LA, Lewis WD, Skinner M. 1998. Asimple screening test for variant transthyretins associated withfamilial transthyretin amyloidosis using isoelectric focusing. Biochim.Biophys. Acta 1407: 185–192.
Correia BE, Loureiro-Ferreira N, Rodrigues JR, Brito RM. 2006. Astructural model of an amyloid protofilament of transthyretin.Protein Sci. 15: 28–32.
Fleming CE, Nunes AF, Sousa MM. 2009. Transthyretin: more than meetsthe eye. Prog. Neurobiol. 89: 266–276.
Foss TR, Wiseman RL, Kelly JW. 2005. The pathway by which thetetrameric protein transthyretin dissociates. Biochemistry 44:15525–15533.
Gosal WS, Morten IJ, Hewitt EW, Smith DA, Thomson NH, Radford SE. 2005.Competing pathways determine fibril morphology in the self-assembly of beta2-microglobulin into amyloid. J. Mol. Biol. 351:850–864.
Gregoire C, Marco S, Thimonier J, Duplan L, Laurine E, Chauvin JP, MichelB, Peyrot V, Verdier JM. 2001. Three-dimensional structure of thelithostathine protofibril, a protein involved in Alzheimer’s disease.Embo J. 20: 3313–3321.
Habicht G, Haupt C, Friedrich RP, Hortschansky P, Sachse C, MeinhardtJ, Wieligmann K, Gellermann GP, Brodhun M, Gotz J, HalbhuberKJ, Rocken C, Horn U, Fandrich M. 2007. Directed selection of aconformational antibody domain that prevents mature amyloid fibrilformation by stabilizing Abeta protofibrils. Proc. Natl Acad. Sci. USA104: 19232–19237.
Hill SE, Robinson J, Matthews G, Muschol M. 2009. Amyloid protofibrils oflysozyme nucleate and grow via oligomer fusion. Biophys. J. 96:3781–3790.
Hou X, Parkington HC, Coleman HA, Mechler A, Martin LL,Aguilar MI, Small DH. 2007. Transthyretin oligomers induce
J. Mol. Recognit. 2011; 24: 467–476 Copyright � 2011 John Wiley & S
calcium influx via voltage-gated calcium channels. J. Neurochem.100: 446–457.
Inouye H, Domingues FS, Damas AM, Saraiva MJ, Lundgren E, SandgrenO, Kirschner DA. 1998. Analysis of x-ray diffraction patterns fromamyloid of biopsied vitreous humor and kidney of transthyretin (TTR)Met30 familial amyloidotic polyneuropathy (FAP) patients: axiallyarrayed TTR monomers constitute the protofilament. Amyloid 5:163–174.
Kad NM, Thomson NH, Smith DP, Smith DA, Radford SE. 2001. Beta(2)-microglobulin and its deamidated variant, N17D form amyloidfibrils with a range of morphologies in vitro. J. Mol. Biol. 313:559–571.
Karsai A, Grama L, Murvai U, Soos K, Penke B, Kellermayer MSZ. 2007.Potassium-dependent oriented growth of amyloid ß 25-35 fibrils onmica. Nanotechnology 18: 345102.
Karsai A, Martonfalvi Z, Nagy A, Grama L, Penke B, Kellermayer MSZ. 2006.Mechanical manipulation of Alzheimer’s amyloid ß 1-42 fibrils. J.Struct. Biol. 155: 316–326.
Karsai A, Murvai U, Soos K, Penke B, Kellermayer MS. 2008. Orientedepitaxial growth of amyloid fibrils of the N27C mutant beta 25-35peptide. Eur. Biophys. J. 37: 1133–1137.
Karsai A, Nagy A, Kengyel A, Martonfalvi Z, Grama L, Penke B, KellermayerMSZ. 2005. Effect of lysine-28 side chain acetylation on the nano-mechanical behavior of Alzheimer amyloid ß25-3 fibrils. J. Chem. Info.Mod. 45: 1641–1646.
Kellermayer MS, Grama L, Karsai A, Nagy A, Kahn A, Datki ZL, Penke B.2005. Reversible mechanical unzipping of amyloid beta-fibrils. J. Biol.Chem. 280: 8464–8470.
Kellermayer MS, Karsai A, Benke M, Soos K, Penke B. 2008. Stepwisedynamics of epitaxially growing single amyloid fibrils. Proc. Natl Acad.Sci. USA 105: 141–144.
Khurana R, Ionescu-Zanetti C, Pope M, Li J, Nielson L, Ramirez-Alvarado M,Regan L, Fink AL, Carter SA. 2003. A general model for amyloid fibrilassembly based on morphological studies using atomic force micro-scopy. Biophys. J. 85: 1135–1144.
Kodali R, Wetzel R. 2007. Polymorphism in the intermediates and productsof amyloid assembly. Curr. Opin. Struct. Biol. 17: 48–57.
Lai Z, ColonW, Kelly JW. 1996. The acid-mediated denaturation pathway oftransthyretin yields a conformational intermediate that can self-assemble into amyloid. Biochemistry 35: 6470–6482.
Modler AJ, Fabian H, Sokolowski F, Lutsch G, Gast K, Damaschun G. 2004.Polymerization of proteins into amyloid protofibrils shares commoncritical oligomeric states but differs in the mechanisms of theirformation. Amyloid 11: 215–231.
Modler AJ, Gast K, Lutsch G, Damaschun G. 2003. Assembly of amyloidprotofibrils via critical oligomers–a novel pathway of amyloidformation. J. Mol. Biol. 325: 135–148.
Muller DJ, Engel A. 1997. The height of biomolecules measured withthe atomic force microscope depends on electrostatic interactions.Biophys. J. 73: 1633–1644.
Naiki H, Nagai Y. 2009. Molecular pathogenesis of protein misfoldingdiseases: pathological molecular environments versus quality controlsystems against misfolded proteins. J. Biochem. 146: 751–756.
Nixon RA. 2007. Autophagy, amyloidogenesis and Alzheimer disease. J.Cell Sci. 120: 4081–4091.
Relini A, Canale C, De Stefano S, Rolandi R, Giorgetti S, Stoppini M, Rossi A,Fogolari F, Corazza A, Esposito G, Gliozzi A, Bellotti V. 2006. Collagenplays an active role in the aggregation of beta2-microglobulin underphysiopathological conditions of dialysis-related amyloidosis. J. Biol.Chem. 281: 16521–16529.
Relini A, De Stefano S, Torrassa S, Cavalleri O, Rolandi R, Gliozzi A, GiorgettiS, Raimondi S, Marchese L, Verga L, Rossi A, Stoppini M, Bellotti V.2008. Heparin strongly enhances the formation of beta2-microglobulin amyloid fibrils in the presence of type I collagen. J.Biol. Chem. 283: 4912–4920.
Serpell LC, Sunde M, Benson MD, Tennent GA, Pepys MB, Fraser PE. 2000.The protofilament substructure of amyloid fibrils. J. Mol. Biol. 300:1033–1039.
Serpell LC, Sunde M, Fraser PE, Luther PK, Morris EP, Sangren O, LundgrenE, Blake CC. 1995. Examination of the structure of the transthyretinamyloid fibril by image reconstruction from electron micrographs. J.Mol. Biol. 254: 113–118.
Sousa MM, Cardoso I, Fernandes R, Guimaraes A, Saraiva MJ. 2001.Deposition of transthyretin in early stages of familial amyloidotic
ons, Ltd. wileyonlinelibrary.com/journal/jmr
475

R. H. PIRES ET AL.
476
polyneuropathy: evidence for toxicity of nonfibrillar aggregates. Am.J. Pathol. 159: 1993–2000.
Sousa MM, Fernandes R, Palha JA, Taboada A, Vieira P, Saraiva MJ. 2002.Evidence for early cytotoxic aggregates in transgenic mice for humantransthyretin Leu55Pro. Am. J. Pathol. 161: 1935–1948 .
Sousa MM, Saraiva MJ. 2003. Neurodegeneration in familial amyloidpolyneuropathy: from pathology to molecular signaling. Prog. Neu-robiol. 71: 385–400.
Sunde M, Serpell LC, Bartlam M, Fraser PE, Pepys MB, Blake CC. 1997.Common core structure of amyloid fibrils by synchrotron X-raydiffraction. J. Mol. Biol. 273: 729–739.
wileyonlinelibrary.com/journal/jmr Copyright � 2011 John Wiley
Westermark P, Sletten K, Johansson B, Cornwell GG 3rd. 1990. Fibril insenile systemic amyloidosis is derived from normal transthyretin.Proc. Natl Acad. Sci. USA 87: 2843–2845.
Yamaguchi I, Hasegawa K, Takahashi N, Gejyo F, Naiki H. 2001. Apolipo-protein E inhibits the depolymerization of beta 2-microglobulin-related amyloid fibrils at a neutral pH. Biochemistry 40:8499–8507.
Yamamoto S, Hasegawa K, Yamaguchi I, Goto Y, Gejyo F, Naiki H. 2005.Kinetic analysis of the polymerization and depolymerization ofbeta(2)-microglobulin-related amyloid fibrils in vitro. Biochim. Bio-phys. Acta 1753: 34–43.
& Sons, Ltd. J. Mol. Recognit. 2011; 24: 467–476


















