Some aspect os f the developmen otf the notochord in mouse ...longitudinal sections it was found...
33
/. Embryol. exp. Morph. Vol. 32, 1, pp. 1-33, 1974 Printed in Great Britain Some aspects of the development of the notochord in mouse embryos By A. JURAND 1 From the Institute of Animal Genetics, Edinburgh This paper is dedicated to Professor C. H. Waddington, F.R.S. SUMMARY Prenotochordal cells are derived by proliferation from the pluripotential ectoderm in the Hensen's node area, and they migrate in the cephalic direction to be incorporated into the roof of the archenteron. Later, after separation from the archenteron, the notochord probably includes some endodermal cells. Topographically it occupies an intermediate position between the ectoderm and endoderm. In this way it assumes a mesodermal-like character. After separation fiom the archenteron, the differentiation of the organ and its cells follows a cranio- caudal gradient. At the time of separation the basal lamina, originally covering the dorsal side of both the prenotochordal and endodermal cells, remains continuous between the endoderm and the notochord. Its presence from the earliest stages of morphogenesis of the notochord presumably indicates that it plays an important formative role. The cells of Hensen's node contain numerous microtubules, centrioles and cilia. In later stages fibrillogenesis in the prenotochordal and notochordal cells and excortication (ecdysis) occur. The intracellular fibrillar material in the notochordal cells may first form the basal lamina and later the perichordal sheath. In 12 to 13-day-old embryos, during excortication of the fibrillar material, the basal lamina undergoes disruption and eventually disappears. From the 10th day onwards, the outer plasma membrane shows an active endocytosis, by the formation of micropinocytic vesicles. Vacuolization occurs in the later stages of morpho- genesis and it is suggested that this is due to dilation of endoplasmic reticulum and mito- chondria. There is no evidence of a syncytial stage in the development of the notochord in mouse embryos. INTRODUCTION The notochord and its development have been extensively investigated since the nineteenth century when it was recognized as a characteristic of all vertebrate embryos. Its presence in all representatives of this subphylum supported the theory of evolution, and later three other, more primitive, groups of animals were included with vertebrates in the phylum of chordates, because they possessed notochords. In vertebrates, the notochord is present only in early stages of embryonic life; it persists relatively longer in lower vertebrates. In some of the lowest chordates such as Amphioxus, and in Cyclostomata, the notochord persists 1 Author's address: Institute of Animal Genetics, West Mains Road, Edinburgh EH9 3JN, U.K. I EMB 32
Transcript of Some aspect os f the developmen otf the notochord in mouse ...longitudinal sections it was found...

/ . Embryol. exp. Morph. Vol. 32, 1, pp. 1-33, 1974
Printed in Great Britain
Some aspects of the development of thenotochord in mouse embryos
By A. JURAND1
From the Institute of Animal Genetics, Edinburgh
This paper is dedicated to Professor C. H. Waddington, F.R.S.
SUMMARYPrenotochordal cells are derived by proliferation from the pluripotential ectoderm in the
Hensen's node area, and they migrate in the cephalic direction to be incorporated into theroof of the archenteron. Later, after separation from the archenteron, the notochord probablyincludes some endodermal cells. Topographically it occupies an intermediate position betweenthe ectoderm and endoderm. In this way it assumes a mesodermal-like character. Afterseparation fiom the archenteron, the differentiation of the organ and its cells follows a cranio-caudal gradient. At the time of separation the basal lamina, originally covering the dorsalside of both the prenotochordal and endodermal cells, remains continuous between theendoderm and the notochord. Its presence from the earliest stages of morphogenesis of thenotochord presumably indicates that it plays an important formative role.
The cells of Hensen's node contain numerous microtubules, centrioles and cilia. In laterstages fibrillogenesis in the prenotochordal and notochordal cells and excortication (ecdysis)occur. The intracellular fibrillar material in the notochordal cells may first form the basallamina and later the perichordal sheath. In 12 to 13-day-old embryos, during excortication ofthe fibrillar material, the basal lamina undergoes disruption and eventually disappears. Fromthe 10th day onwards, the outer plasma membrane shows an active endocytosis, by theformation of micropinocytic vesicles. Vacuolization occurs in the later stages of morpho-genesis and it is suggested that this is due to dilation of endoplasmic reticulum and mito-chondria.
There is no evidence of a syncytial stage in the development of the notochord in mouseembryos.
INTRODUCTION
The notochord and its development have been extensively investigated sincethe nineteenth century when it was recognized as a characteristic of all vertebrateembryos. Its presence in all representatives of this subphylum supported thetheory of evolution, and later three other, more primitive, groups of animalswere included with vertebrates in the phylum of chordates, because theypossessed notochords.
In vertebrates, the notochord is present only in early stages of embryoniclife; it persists relatively longer in lower vertebrates. In some of the lowestchordates such as Amphioxus, and in Cyclostomata, the notochord persists
1 Author's address: Institute of Animal Genetics, West Mains Road, Edinburgh EH9 3JN,U.K.
I EMB 32

2 A. JURANDthroughout life and acts as an endoskeleton. It is one of the earliest organs,being initiated during gastrulation and neurulation. Its origin, development andarchitecture are very similar in all vertebrates. Its fundamental roles are to pro-vide the cranio-caudal orientation of the embryonic axis and to act as primaryorganizer for adjacent embryonic organs. Thus it contributes to the develop-ment of the spinal cord and the vertebral column (Bruni, 1912; Weiss, 1939;Holtzer & Detwiler, 1953; Waddington, 1956; Zilliken, 1967). It is not essentialfor the formation of somites (Bellairs, 1963), although chondrogenesis of thesomite mesoderm depends partially on induction by the notochord and neuraltube (Kitchin, 1949; Theiler, 1950; Watterson, Fowler & Fowler, 1954; Cooper,1965). However, after the 3rd day of incubation, somites in chick embryosare destined to form cartilage even when isolated as chorio-allantoic grafts, soat this stage the notochord is not essential for chondrogenesis (Watterson et al.1954; Ellison, Ambrose & Easty, 1969). The inductive activity of the notochordon myogenesis in the myotomes (the chorda factor) was discovered in amphibiansby Yamada (1940) and later confirmed by Smithberg (1954) and Niazi (1969).
The rostral end of the notochord is very close to Rathke's pocket, and inPleurodeles waltlii its inductive action affects the development and growth of theanterior lobe of the pituitary gland (Eyal-Giladi, 1958).
In later stages, when the notochord is an elongated rod detached from theother axial organs, it has lost most of its inductive capacity (Waddington, 1952).Then its turgidity, due to vacuolation of its cells, makes it into a preskeletalstructure, elongating and shaping the body and providing some elasticity andstiffness (Waddington, 1956).
Notochord development has been investigated in all classes of chordates.Most of the work in vertebrates has been related to the development of thevertebral column. The development of the mammalian notochord has beeninvestigated in sheep (Carlier, 1890), pigs (Williams, 1908), mice (Selenka, 1883;Sobotta, 1911; Dawes, 1930), guinea-pigs (Huber, 1918) and, more recently, inrabbits (Leeson & Leeson, 1958).
This investigation follows the origin of the notochord in mouse embryos,examines the process of separation from the archenteron roof, and describesthe basal lamina during morphogenesis of the notochord, when it assumes itsrod-like structure. In later stages associations of the notochord with the surround-ing axial embryonic organs are examined. Electron microscopic studies of somestages were designed to follow the subcellular events of cell differentiation asso-ciated with the developmental changes seen at the light microscope level.
MATERIALS AND METHODS
The mouse embryos used belonged to the JBT/Jd strain, which was derivedfrom the outbred JC strain selected for genes a, b and bt (Jurand, 1968) and inbredfor 46 generations by brother-sister litter-mate matings with retention of

Development of mouse notochord 3
sublines. Females aged 8-10 weeks were kept with males overnight and examinedfor copulation plugs each morning. The age of the embryos was determined inday or half-day stages, the day when the copulation plug was found being con-sidered the first day of pregnancy. More precise staging is unnecessary, as thedevelopment of embryos in one litter shows variation equivalent to at least12 h (Theiler, 1972).
The embryos examined by light and electron microscopy were aged 8-16 days.After dissection under ether anaesthesia the uteri were prefixed in toto with2-5% glutaraldehyde in 0-1 N cacodylate buffer (Plumel, 1948) at 1-2 °C forabout 2 min; the embryos were then removed from the uterus and fixed for afurther 1-1 ̂ -h in the same fixative. For light microscopy, the embryos weredehydrated with graded ethanol, embedded in 56 °C paraffin wax in the usualway, and sectioned transversely and sagittally at a thickness of 6 /*m. Stainingwith Unna-Pappenheim methyl green-pyronin was particularly successful afterglutaraldehyde fixation.
For demonstration of the basal laminae at the light microscope level, someearly embryos (8-10 days old) after glutaraldehyde fixation were stained withthe allochrome procedure (Lillie, 1951).
For electron microscopy, axial organs in the trunk region were dissected, atthe time of fixation, from embryos aged 11 days or more; younger embryos wereprocessed without dissection. After glutaraldehyde fixation the material wasrinsed three times (5 min each) with 7-5% sucrose solution in 0-1 N cacodylatebuffer, and then post-fixed for 1 h with osmium tetroxide fixative (Palade, 1952)with 4-5% sucrose added. This was followed by dehydration with graded ethanol,rinsing with 1,2-epoxypropane and embedding in Araldite, using the rotaryshaker (Jurand & Ireland, 1965) to facilitate penetration of the embeddingmedium. Sectioning was transverse to the long axis, in younger embryosapproximately half-way along the head process and in older embryos at thelevel of the forelimbs. Ultra-thin sections on collodion-carbon-coated gridswere stained with 2-5 % uranyl acetate followed by lead citrate, according to themethod of Reynolds (1963), and examined in an AEI EM 6 B microscope.
RESULTS
Light microscope observations on the organogenesisof the notochord
The formation of the notochord is first detected in 8-day-old embryos, whenHensen's node appears at the cranial end of the primitive streak. Usually, onlya slight invagination in the ectoderm in the caudal part of the embryonic disc isvisible in longitudinal parasagittal sections. In more advanced embryos, even inthe same litter, there is a very short head process extending in the cephalic directionin the mid-sagittal area from Hensen's node (Fig. 1). Using transverse andlongitudinal sections it was found that mitotic figures are more numerous in

A. JURAND

Development of mouse notochord 5Hensen's node area and in the head process than in the adjacent cranial or caudalparts (Fig. 2).
In slightly older embryos the cells forming the cephalic end of the head processare elongated, and are indistinguishable from the squamous endodermal cellswith which they fuse to form the archenteron roof (Fig. 3). The most centralportion of the archenteron roof is the prenotochordal tissue forming the chordalplate, from which the notochord will develop.
In 9-day-old embryos the chordal plate is elongated cranially, and its entirelength is incorporated into and fused with the endoderm. In transverse sections,however, the primordial notochord cells differ from the endoderm cells, whichlie at the same level, by being more columnar and by their oval nuclei beingmore perpendicularly orientated. At this stage, 2 or 3 sections through the caudalend of the chordal plate usually show a continuity with the adjacent neuralgrove. This continuity is probably a remnant of the continuity between Hensen'snode and the head process. It persists for about the next 24 h (Fig. 3).
In 9^-day-old embryos the notochord starts to separate from the endodermof the archenteron roof by a process of folding-off in the dorsal direction. Thusthe notochord becomes a slender flattened rod, initially apposing the ventralsurface of the neural groove on one side and adhering dorsally to the endo-derm on the other. The endoderm consolidates to form a continuous layer afterthe notochord has separated. The participation of the basal lamina in theseparation of the notochord from the endoderm will be discussed later.
The folding-off process in mouse embryos does not lead to the formation ofthe chorda canal proper, but by electron microscopy it is possible to find arelatively small intercellular space in the centre of the notochord which mayrepresent the chorda canal (Fig. 4). Later, however, traces of a narrow discon-tinuous chorda canal are found by light microscopy (Fig. 5). For 24 h after
FIGURES 1-4
Fig. .1. Sagittal section of a 9-day-old mouse embryo. The arrow indicates theposition of Hensen's node. The anterior part of the section is not in the median planeand shows three somites and part of the foregut. x 63.Fig. 2. Higher magnification of a sagittal section of the Hensen's node area (arrow)in a 9-day-old embryo showing a group of several dividing cells. On the left ofHensen's node the head process also shows mitotic figures and is already incorpor-ated into the endoderm but in the extreme left the squamous endodermal cells arestill separate from the head process cells, x 560.Fig. 3. Transverse section through the neural groove at the level of the caudal endof the notochordal plate in 9i-day-old embryo. Note the residual connexion betweenthe neural groove and the notochordal plate, regarded here as a trace of Hensen'snode, x 420.Fig. 4. Electron micrograph of a transverse section through the notochord in a 10-day-old embryo at the time of separation from the archenteron roof. Note a sizeableintercellular space to the right of the centre which is probably a vestigial chorda canal,x 3500.

A. JURAND

Development of mouse notochord 1
separation from the archenteron roof the notochord adheres to the dorsal sideof the archenteron, and there is a short interval before it attaches to the ventralsurface of the neural tube (Fig. 6). It is in 10^-day-old embryos that the noto-chord is detaching itself from the endoderm, first at the level of the cardiacprimordium, then gradually in the caudal direction to come into contact withthe ventral surface of the neural tube. At this stage the notochord, as it detachesfrom the endoderm, bends like a wave towards the neural tube and attachesitself to the ventral surface of the latter (Fig. 9). In 11-day-old embryos the entiretrunk portion of the notochord is attached to the neural tube, and remainsattached to the endoderm only at the extreme cephalic and caudal ends (Figs. 7,21). At this and later stages all the changes in the notochord occur in a cranio-caudal gradient. In the occipital area, however, after detachment from theendoderm, the notochord remains free from the ventral side of the developingbrain; later it lies on the dorsal side of the basioccipital cartilage (Fig. 8).
From 11 days onwards the notochord is circular in transverse section, and itsdiameter is 20 jum. At the cephalic end it is slightly thinner (about 15/^rn indiameter). Eventually, in 11^-day-old embryos, the notochord leaves the endo-derm along almost all its length, and except for the hind-brain area, adheresto the ventral side of the neural tube (Fig. 22).
In 12-day-old embryos the notochord detaches also from the neural tube,apparently because mesenchyme cells penetrate between the neural tube and thenotochord (Fig. 10). This process also follows a cranio-caudal gradient. Theactivity of the mesenchyme cells detaches the notochord completely from theaxial organs and it becomes surrounded by the mesenchyme (Figs. 11, 12). Thusin a central position, parallel to and about 100 /tm ventral from the neural tube,it indicates the site of the future vertebral column. It is now a uniform cylindricalrod, 25 jum in diameter, extending from a point close to Rathke's pocket(Fig. 13) to the end of the tail.
In 13-day-old embryos, the notochord is surrounded by evenly scatteredmesenchyme cells which are denser than previously. At this stage, in thelumbo-sacral area the notochord shows 10-15 slight short flexures which are aslong as the future vertebral segments. The rest of the notochord does not show
FIGURES 5-8
Fig. 5. Longitudinal section of the notochord in a 13-day-old embryo, x 600.Fig. 6. Transverse section through a 10-day-old embryo showing the notochord (w)at the time of folding-off from the archenteron. x 400.Fig. 7. Sagittal section through a lOJr-day-old embryo showing the wave of thenotochord at the level of future forelimbs (caudal direction to the right) by whichthe notochord (/?) detaches itself from the archenteron (arch) and then attaches to theneural tube (tit). For explanation compare with Fig. 21. x 290.Fig. 8. Section through the basioccipital cartilage condensation (bcc) in a 14-day-oldembryo, showing its support for the notochord. x 170.

A. JURAND
13

Development of mouse notochord 9
such flexures. Ventral convexities are opposite the future intervertebral discs,and dorsal ones opposite the future vertebral centra (Fig. 14).
During the next 36 h the mesenchyme around the notochord undergoessegmentation and forms more distinct vertebral cartilage condensations, whilesegmental bends in the lumbo-sacral region disappear. The formation of thesecondensations also proceeds cranio-caudally.
In 14-day-old embryos the notochord first shows dilations opposite the pro-spective intervertebral discs, and corresponding constrictions at the level of thevertebral centra; thus, it has a moniliform shape (Fig. 15). Like all otherprocesses, this progresses caudally, so that in one embryo the notochord may bemoniliform from the first cervical vertebra to the proximal part of the tail, butthe rest of it is still a straight rod (Fig. 25). These dilations may be formed bydifferent pressures being exerted on the notochord by the dense cartilagecondensations of the vertebral centra and by the less-dense intervertebral con-densations (Dawes, 1930).
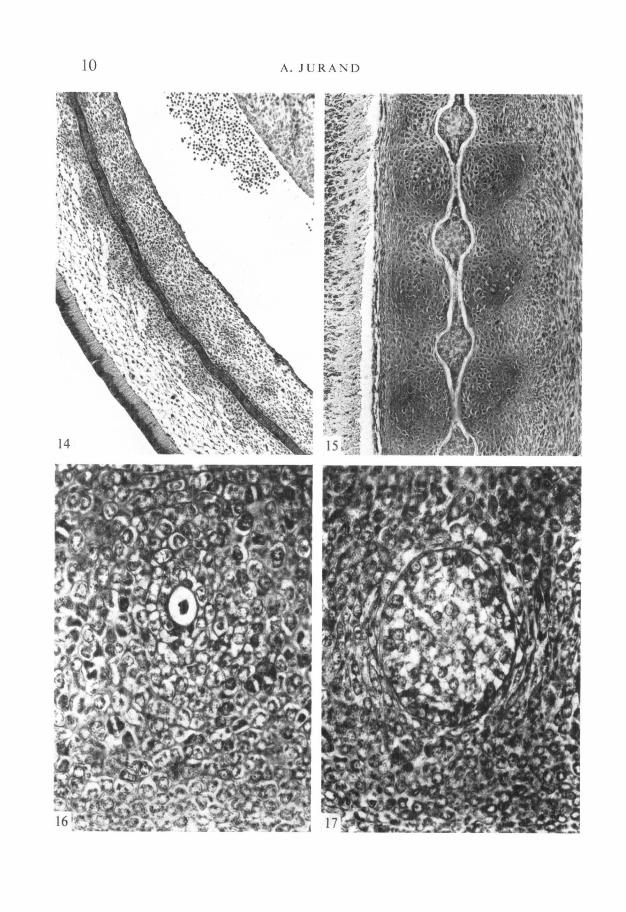
In the constricted parts the slender notochord has a well-preserved perichordalsheath, but contains within the sheath, at most, some cellular debris (Fig. 16).In the dilatations the sheath is thinner and the notochord cells show vacuoliza-tion (Fig. 17). The cells at the junction of dilatations and constrictions arenecrotic (Fig. 15).
As the more distal parts of the tail (where the vertebral cartilage condensationsare less dense) have no dilatations, their formation may depend on the pressureexerted by the vertebral cartilage condensations. This suggestion is supportedby the observation that the notochord remains cylindrical between the rostralend and the first cervical vertebra, where it is not surrounded by the segmentalcondensations but lies on the basioccipital cartilage (Fig. 8).
The mouse notochord, shortly after it separates from the archenteron roof, isa slender flattened rod with dimensions (in 10-day-old embryos) of 25 /on and10 /on for the oval profile. Caudally at this stage the notochord is also flat,having axes of 10 and 5 /an. From 11 days onwards, however, the notochord is
FIGURES 9-13
Fig. 9. Section through a 104-day-old embryo at the level of the forelimbs where thenotochord adheres to the ventral side of the neural tube, x 450.Fig. 10. Similar section to Fig. 9 through the same area in 11-day-old embryo. Notethe mesenchyme cells between the neural tube and the notochord. Compare withFig. 52. x450.Fig. 11. Similar cross-section to Figs. 9 and 10, in 11-day-old embryo, showing thenotochord detached from the neural tube, x 450.Fig. 12. Section through the notochord surrounded by the dense mesenchyme tissueat the level of the forelimbs in 12-̂ -day-old embryo, x 440.Fig. 13. Sagittal section through the rostral end of the notochord (n) located about01 mm from Rathke's pocket (rp). bcc, Basioccipital cartilage condensation, x 150.
10 A. JURAND

Development of mouse notochord 11
FIGURES 14-17
Fig. 14. Longitudinal section of the notochord in the lumbo-sacral region of a 13-day-old embryo showing about seven segmental flexures. Note vertebral cartilagecondensation at the level of dorsal bends (directed towards the lower left corner ofthe photograph) of the notochord and ventral bends at the level of intervertebraldiscs. Compare with Fig. 23. x 100.Fig. 15. Longitudinal section of the moniliform notochord in 14-day-old embryo,with segmental dilatations opposite future intervertebral discs and constrictionsenclosed in partially chondrified vertebral centra. Note necrotic areas in thenarrowing parts of the dilatations, x 150.
Fig. 16. Section through a constriction of the notochord, containing some cellulardebris, in the middle of the vertebral cartilage condensation; 15-day-old embryo.x450.Fig. 17. Section through a dilatation of the notochord in a 15-day-old embryo, withvacuolization of the notochord cells, x 450.
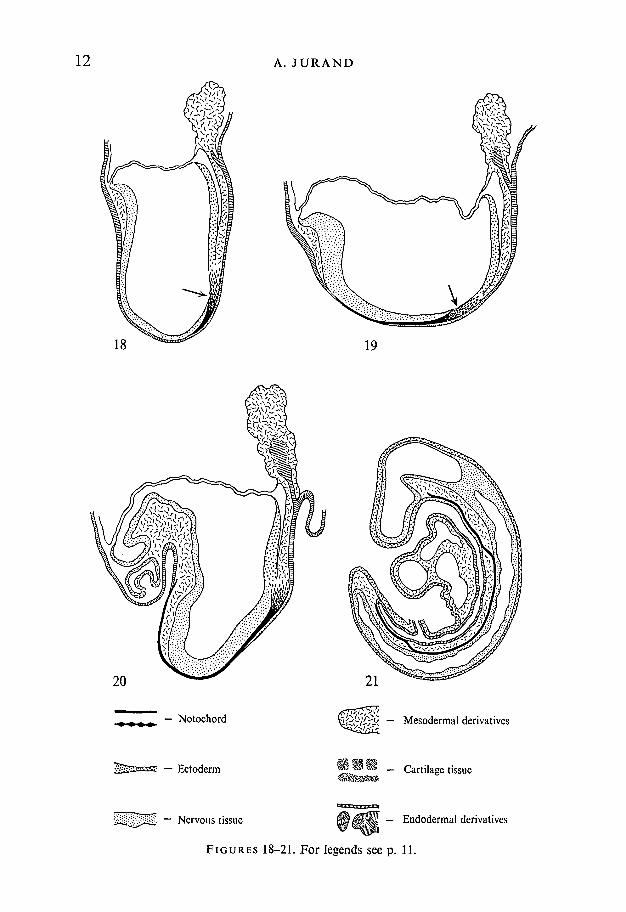
FIGURES 18-21
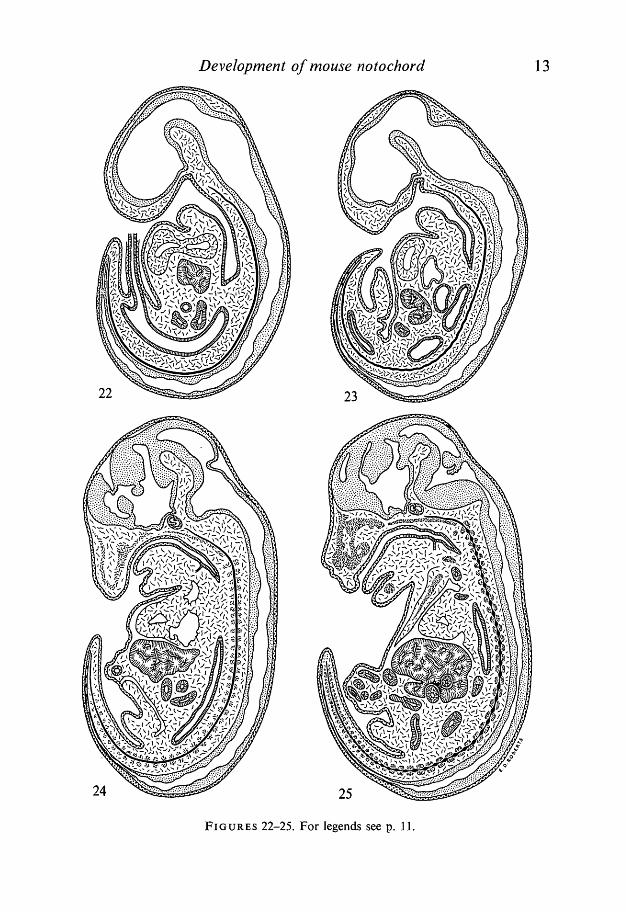
Figs. 18-25. Diagrams of mid-sagittal sections through mouse embryos showingposition of the notochord in relation to adjacent embryonic organs.Fig. .18. 8-day-old mouse embryo, in which the notochord begins to be formed fromHensen's node (arrow), and elongates cranially before incorporation into thearchenteron roof. Magnification approx. x 75.Fig. 19.8i-day-old mouse embryo with well-pronounced head fold shows less lordosisthan the previous stage. The prechordal plate reaches to the area of the heart pri-mordium. The more cranial part is incorporated into the archenteron roof. Arrowindicates the area of Hensen's node. Magnification approx. x 70.Fig. 20. 9-day-old mouse embryo at the time when the fore- and hind-gut have beenformed. The notochord runs on its entire length and remains incorporated into thearchenteron roof. Magnification approx. x 60.Fig. 21. 10-day-old mouse embryo after the closure of the neural tube and reversalof the germ layers by coiling and untwisting. The notochord is separated from theendoderm in the trunk region but remains attached to the endoderm in the cranialand caudal regions. In the trunk region it adheres to the ventral side of the neuraltube, from which it bends and forms wave-like connexions with the cranial and caudalparts. Magnification approx. x 40.
FIGURES 22-25
Fig. 22. lli-day-old mouse embryo showing the notochord adhering on its entirelength to the neural tube, except for the extreme rostral part where it is surroundedby the head mesenchyme. Magnification approx. x 25.Fig. 23. 13-day-old mouse embryo with the notochord detached from the neuraltube except for the extreme caudal end. Note that in the lumbar and sacral regionsthe notochord has about 15 characteristic segmental flexures. (See also Fig. 14.)The rostral end of the notochord reaches the vicinity of Rathke's pocket. Magnifica-tion approx. x 15.Fig. 24.14-day-old embryo showing the notochord running through the centre of thecartilage condensations of the vertebral centra. Formation of these cartilage conden-sations proceeds in the cranio-caudal direction so that in the extreme tail region theyare not fully developed. The head part of the notochord lies on the basioccipital carti-lage condensation. Magnification approx. x 12.Fig. 25. 15-day-old mouse embryo with the moniliform notochord having spindle-shaped dilatations between the adjacent vertebral centra, and narrow constrictionsin the middle of each centrum. The formation of dilatations proceeds in the cranio-caudal direction and therefore they are not present in the extreme caudal part.Magnification approx. x 9.
12 A. J U R A N D
19
20
i/i< — Mesodermal derivatives
V**m _ Cartilage tissue
— Notochord
— Ectoderm
Nervous tissue OMM'M. - Endodermal derivatives
FIGURES 18-21. For legends see p. 11.
Development of mouse notochord 13
24 25
FIGURES 22-25. For legends see p. 11.

14 A. JURAND
28

Development of mouse notochord 15
cylindrical (20/*m in diameter) opposite the forelimbs, where it is adheringclosely to the neural tube. In the head it is slightly thinner (15 /an.) and apposedneither to the brain nor to the foregut. During the next 2 days its diameterincreases to reach 35 /«n at the level of the forelimbs in 13-day-old embryos. In14-day-old embryos the first intervertebral dilatations, appearing in the cervicaland thoracic area, are 100/*m in diameter. At this stage the intravertebralsegments of the notochord have their diameter reduced to 20 jum.
Figs. 18-25 summarize the changes seen by light microscopy during the de-velopment of the notochord.
Electron microscope observations
The ultrastructure of the cells of 8-day-old embryos in the area of Hensen'snode and of the early head process is typical for undifferentiated pluripotentialcells. Their large oval nuclei contain uniformly granular chromatin and electron-translucent nuclear matrix. The nucleoli (usually one, but occasionally twoper section) have a typical granulo-fibrous ultrastructure. The electron-trans-lucent cytoplasm contains only a few small mitochondria (0-5 jum. in diameter).Free endoplasmic reticulum cisternae are very scarce, although some profilesare often attached to, and continuous with, the outer nuclear membrane (Figs.26, 27). This suggests that endoplasmic reticulum is formed in such, earlyembryonic cells by the outer nuclear membrane. Ribosomes, mainly in the formof polysomes, are sparsely scattered throughout the cytoplasm and attached tothe existing endoplasmic reticulum. Golgi apparatus is only occasionally found;it consists of small stacks of 2-3 flattened saccules surrounded by clusters ofmany small vesicles (Fig. 28). Some of these vesicles may align to form flattenedsaccules.
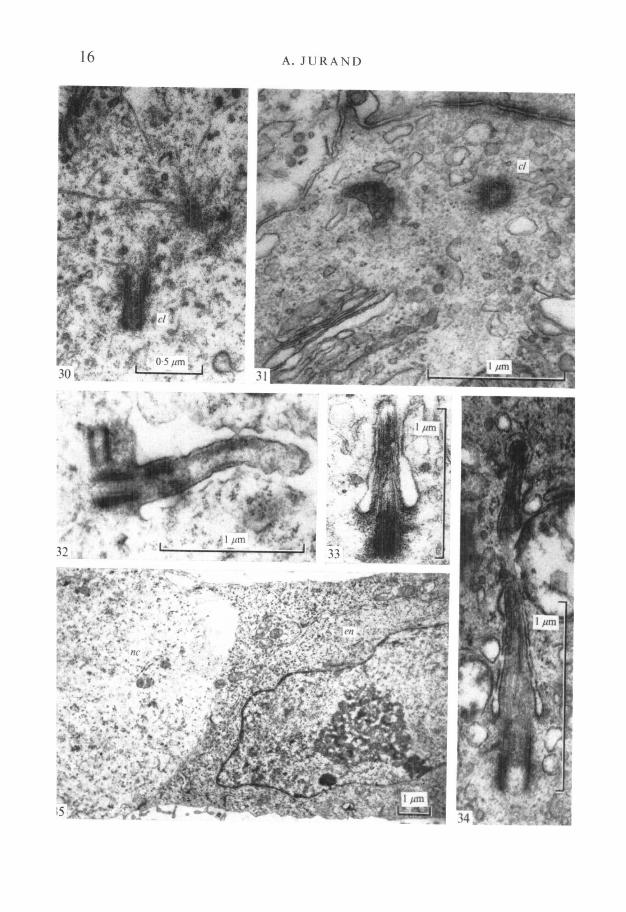
Microtubules about 20 nm in diameter are scattered randomly in the Golgiarea of Hensen's node cells (Fig. 29), but they are concentrated near the cen-trioles which are common in these cells (Fig. 30). The centrioles may be free inthe cytoplasm (Fig. 31) or they may act as basal bodies for the occasional cilia
FIGURES 26-29
Fig. 26. Electron micrograph of a cell from the area of Hensen's node. Note scarcityof endoplasmic reticular profiles and mitochondria, but there is one Golgi area (g)in an early stage of development, x 9000.Fig. 27. Part of prenotochordal cell showing an extensive profile of the endoplasmicreticulum continuous with the outer nuclear membrane. Note mitochondria (m)close to the profile, x 16000.Fig. 28. Part of a prenotochordal cell (9-day-old embryo) from area of Hensen's nodeshowing the initial development of a Golgi apparatus. Note numerous small vesicularprofiles and scarcity of flat parallel saccules (dictyosomes). In the upper right cornerthere are two intercellular junctions, x 25000.Fig. 29. Early Golgi apparatus area in a prenotochordal cell in 9^-day-old embryo,showing microtubules scattered between small vesicular bodies, x 30000.
16 A. JURAND
wmm

Development of mouse notochord 17
which are seen at this stage (Fig. 32). Cilia are also found later (in 9i-day-oldembryos) in the prenotochordal cells in the archenteron roof; they even occurin 10-day-old embryos when separation from the archenteron roof occurs, andoccasionally in embryos up to 14 days old, when they are squeezed between thenotochordal cells (Figs. 33, 34).
In 8j- to 9-day-old embryos, when the prenotochordal cells fuse with, thearchenteron roof, their structure differs from the adjacent endodermal cells. Thecytoplasm of the prenotochordal cells is less electron-dense than that of theendodermal cells, due to fewer free ribosomes and polysomes. The prenoto-chordal cells also contain less endoplasmic reticulum cisternae, which indicatesthat they are less differentiated than the endodermal cells. The endoplasmicreticulum may still be continuous with the outer nuclear membrane. The dorsalsurface of both these kinds of cells is covered by a common basal lamina whichis about 15 nm thick and not fully developed (Fig. 35).
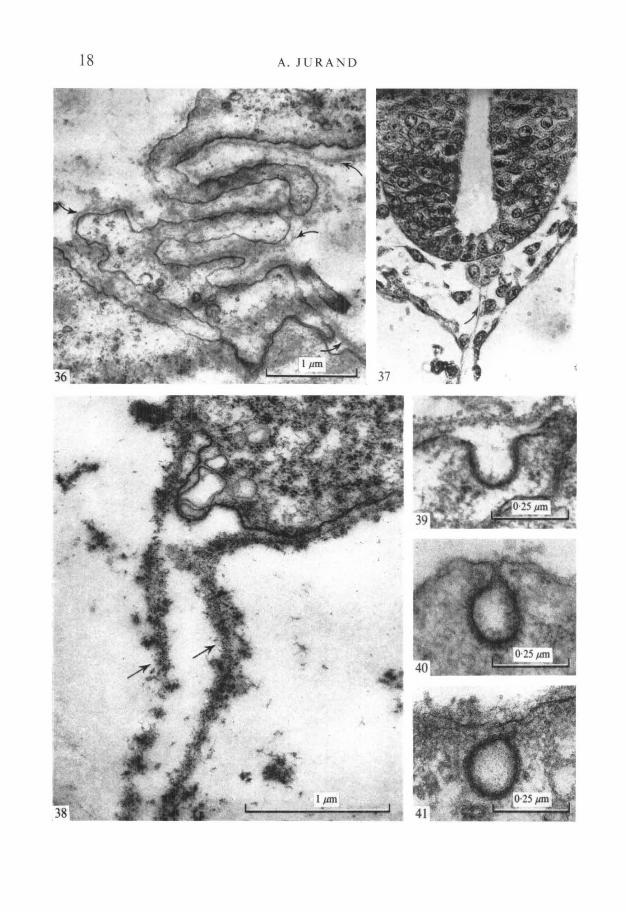
During separation from the archenteron roof in 9^-day-old embryos thenotochord is covered by a more definite 25 nm thick basal lamina. When separa-tion is ending (in some 10-day-old embryos) a complex junction is seen betweenventral cells of the notochord and the underlying endodermal cells. Thisjunction is formed by interdigitating microvilli of notochordal and endodermalcells, and is covered by a continuous basal lamina (Fig. 36); this transientconnexion between the notochord and the endoderm disappears when separationis complete.
In older embryos of this stage (10-day-old embryos) when the notochordappears separate from the endoderm, it is still covered by the basal lamina, whichremains temporarily continuous with the basal lamina of the endoderm. Whennotochordal separation appears complete by light microscopy in preparationsstained with methyl green-pyronin, its basal lamina can still be seen as a free
FIGURES 30-35
Fig. 30. Microtubules associated with a pair of centrioles (cl) in a prenotochordalcell in 9-day-old embryo. Note a micropinocytic vesicle in the bottom right corner,x 37000.
Fig. 31. Notochord cell in 10-day-old embryo showing a transverse section of acentriole (cl) in the upper part of the micrograph. Above it note three intercellularjunctions, x 38000.Fig. 32. Longitudinal section through a cilium of cell of Hensen's node (9-day-oldembryo). At right angles to the basal body is another centriole. x 40000.Fig. 33. A cilium in a notochordal cell in 10^-day-old embryo. Note that thepericiliary space is narrower, x 40000.Fig. 34. A cilium squeezed between adjacent notochordal cells in a 14-day-oldembryo. The periciliary space is almost absent, x 50000.Fig. 35. The electron-dense endoderm cell (en) with the paler prenotochordal cell(nc) at the time of incorporation of the chordal plate into the archenteron roof,x 9000.
2 EMB 32
18 A. JURAND

Development of mouse notochord 19
double sheet running to the endoderm, when viewed by light microscopy afterallochrome staining (Lillie, 1951) or by electron microscopy. The connectingdouble sheet of the basal lamina lies free in the intercellular space between thenotochord and the archenteron roof (Figs. 37, 38).
In 10- and 11-day-old embryos the plasma membranes of the notochordalcells, covered by basal lamina, show active endocytosis, forming micropinocytic,internally coated invaginations and vesicles (Figs. 39-41).
Diagrams of four consecutive developmental stages showing the role of thebasal lamina in the process of separation of the notochord from the endodermare shown in Figs. 42-45.
In 9-j- and 10-day-old embryos the notochordal cells contain numerousGolgi groups which are of a more dictyosomal type than the Golgi apparatus ofthe cells of Hensen's node; they consist of stacks of 3-5 flat saccules surroundedby fewer vesicles. These Golgi groups are more mature than Golgi apparatusin the cells of Hensen's node (Figs. 28, 46).
Bundles of endoplasmic filamentous material consisting of microfibrillarfilaments about 4 nm in diameter always surround the Golgi apparatus at thisstage (Fig. 47). These filaments are in the perinuclear area and elsewhere, notablyclose to the outer cell membranes. In this region there are signs of excortication(ecdysis), with the filaments penetrating the plasma membrane and beingincorporated into the developing basal lamina. Where excortication occurs theplasma membrane appears disrupted (Fig. 48). Single, elongated and slightlydilated cisternae of endoplasmic reticulum are scattered in the cytoplasm.Mitochondria about 0-3 /*m in diameter are present, about 6 per section per cell.
FIGURES 36-41
Fig. 36. Electron micrograph showing the temporary complex junction between thenotochord (above) and endoderm formed by microvilli-like extensions of theopposing cells. Note that the whole junction is enveloped by a continuous basallamina (arrows), x 25000.Fig. 37. Light micrograph showing transverse section through the notochordadhering to the neural tube in a 10-day-old embryo. Note two sheets of the basallamina (arrow) attached to the notochord and running through the intercellularspace towards the archenteron roof. Stained with the allochrome procedure, x 500.Fig. 38. Electron micrograph showing two sheets of the basal lamina (arrows)suspended in the intercellular space and continuous with the basal lamina on thenotochordal cell, x 40000.Figs. 39-41. Micropinocytic coated vesicle as an invagination of the external cellmembrane of a notochordal cell in 11-day-old embryo. Note that the basal laminapasses over the invagination. x 85000.Fig. 40. Micropinocytic coated vesicle in an advanced stage of formation by pinch-ing off the cell membrane; 10^-day-old embryo, x 85000.Fig. 41. Micropinocytic vesicle inside the notochord cell after detachment from thecell membrane; 10-day-old embryo, x 85000.

20 A. JURAND
Figs. 42-45. Diagrams of sections through the notochord and adjacent embryonicorgans at a level just caudal from the forelimb fields, showing participation of thebasal lamina in formation of the notochord in four consecutive stages of develop-ment (based on light and electron microscopy).Fig. 42. Chordal plate in 9-day-old mouse embryo at the time of incorporation intothe archenteron roof. Its basal lamina is continuous with that of the endoderm, whilethe neural plate has its own basal lamina. Magnification approx. x 200.Fig. 43. In the 9^-day-old embryo, when the notochord folds off from the archen-teron roof its basal lamina is still continuous with that of the endoderm. Adhesionto the neural groove is reinforced by a plate which is common to both neural grooveand notochord and is somewhat thicker and more compact than the normal basallamina. Magnification approx. x 300.Fig. 44. The notochord in a 10^-day-old embryo is attached to the neural groove, asin Fig. 43, by means of the basal lamina-like structure common to both organs(see also Fig. 5.1). Ventrally, the basal lamina of the notochord is connected to thebasal lamina of the endoderm by a free double sheet suspended in the intercellularspace. Magnification approx. x 300.
Fig. 45. The notochord in a 12^-day-old embryo, after detachment from the neuraltube, is surrounded by uniformly spaced mesenchymal cells. The neural tube, noto-chord and endoderm are covered by separate basal laminae. Magnification approx.x300.
Occasionally, after separation from the roof of the archenteron, there arenoticeable differences in the electron density of the cytoplasm between adjacentcells of the notochord. The more electron-dense cells contain more Golgi groups,endoplasmic reticulum cisternae, ribosomes and fibrous material than themajority of adjacent electron-translucent cells (Fig. 49). When they are seen in

Development of mouse notochord 21
the notochord separated from, the archenteron roof, the few electron-dense cellsseem to be endodermal cells which have been derived from the archenteron,accompanying the more numerous but less electron-dense prenotochordal cells.
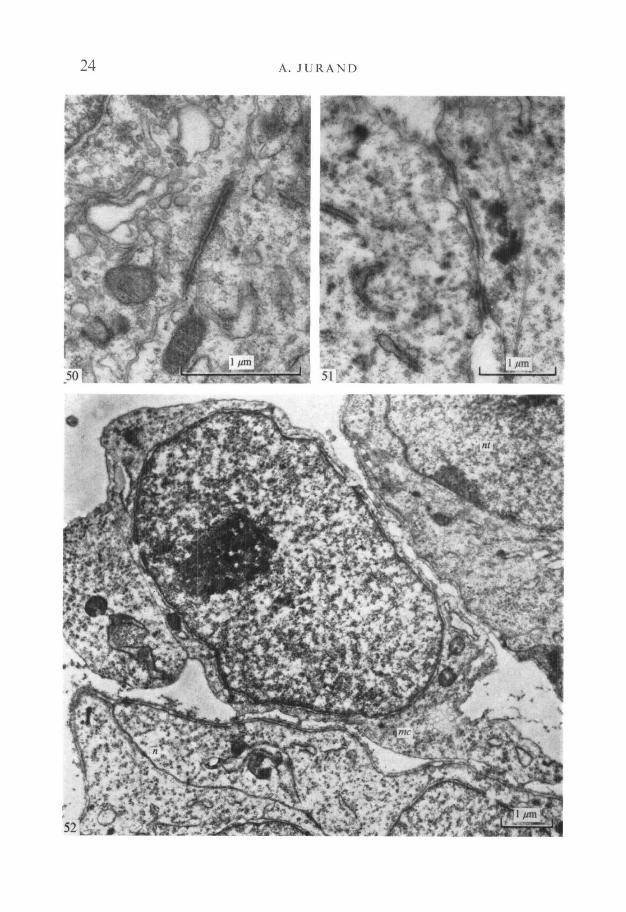
Junctional complexes are present between prenotochordal cells in the headprocess during incorporation into the archenteron roof, and particularly betweenthe notochordal cells soon after separation from the endoderm. These occludingor adhering junctions are not always near the outer free cell margin (Fig. 50).At these stages the cell junctions are not developed sufficiently to be called desmo-somes, but they include discrete cytoplasmic material subjacent to the apposedcell membranes.
After separating from the archenteron the notochord remains in contactwith the neural tube, and adhesion between them is reinforced by 45 nm-thicksingle plates of basal lamina-like material. These plates are thicker and moreelectron-dense than the normal basal laminae of the neural tube and the notochord(Fig. 51).
In 11^-day-old embryos the notochord begins to separate from the neuraltube, apparently assisted by migrating mesenchymal cells, which project pseudo-podia between the notochord and neural tube. These mesenchymal cells can beeasily recognized by the absence of a basal lamina (Fig. 52).
In 11-day and older embryos the intracytoplasmic filaments are thicker (5 nmin diameter) and lie under the external membranes, as if extruded through theplasma membrane and through the previously deposited basal lamina. They mayform the perichordal sheath. During extrusion, the basal lamina is disrupted,becomes incomplete and eventually, on the 13th and 14th day, disappears fromthe surface (Figs. 53, 54).
In 14-day-old embryos notochordal cell cytoplasm begins to vacuolize by theswelling and disruption of both endoplasmic reticulum cisternae and mito-chondria. Vesicles associated with the Golgi complexes are remarkably stableand the surrounding mesenchyme cells and the adjacent neural tube cells showno swelling or dilatations of these organelles (Figs. 55, 56).
Along the cranio-caudal axis in the notochord the vacuoles derived from endo-plasmic reticulum cisternae precede those from disrupted mitochondria. Itappears that in the trunk region, where the cells are more differentiated, thereare many more vacuoles due to swollen mitochondria; near its caudal end, wherethe process is less advanced, there are more vacuoles derived from the endo-plasmic reticulum cisternae, and the mitochondria are normal.
DISCUSSION
The present observations on the development of the notochord can be dividedinto two groups - those on its origin and morphogenesis, and those on the cyto-logical events associated with morphogenesis.
The problem of the origin of the notochord, i.e. from which germ layer it

22 A. JURAND
48
f .'At
W -%|

Development of mouse notochord 23
originates, remains unsettled. This may not be important since, at least underexperimental conditions, there may be no profound and permanent physiologicaldifferences between the three germ layers (Waddington, 1956). However, theorigin of the notochord is often discussed in textbooks of zoology and embryologyand the controversy persists. Recent original publications resemble the reviewof Kingsbury (1920), who pointed to the conflict in this field. The discrepanciesarise not from any lack of information about the morphogenesis of the notochord,but rather from semantic differences in the terms used to describe its origin.
It is agreed that the notochord proliferates from the dorsal lip of the blasto-pore in lower vertebrates and from the cranial end of the primitive streak, i.e.Hensen's node, in higher vertebrates. It should thus be regarded as a mesodermalstructure, as it proliferates from Hensen's node at the time when the mesodermis being formed by the primitive streak. Hensen's node and the rest of theprimitive streak are thought by many to be mesodermal (Kingsbury, 1920;Romer, 1949; Rand, 1950; Nelsen, 1953; Clara, 1955; Patten, 1958; Saunders,1968). This is why the rudimentary notochord at the time of neurulation isoften called the chorda mesoblast (Ebert & Sussex, 1970). However, the noto-chord is not derived from preformed mesoderm; it develops independentlyalthough simultaneously with the mesoderm from the cephalic end of the primi-tive streak.
The view that the ectodermal position of the cells in the blastopore lip and inHensen's node indicates that the notochord is of ectodermal origin seems to beequally justifiable (Cerfontaine, 1906; Huber, 1918; Walter, 1928; Nieuwkoop&Ubbels, 1972).
An endodermal origin is often suggested, because shortly after its formation,the notochord is closely associated with the endoderm as it fuses to form theroof of the archenteron (Sobotta, 1911; Kerr, 1919; Boenig, 1950; Green, 1966;Rugh, 1968). Endodermal origin is not justified, however, as the notochord isonly temporarily incorporated into the roof of the archenteron as the chordalplate. Nevertheless it is possible that, after incorporation into the archenteron,
FIGURES 46-49
Fig. 46. Electron micrograph showing mature Golgi groups in a notochordal cellof a 10-day-old embryo, x 21000.Fig. 47. 10-day-old notochordal cell with microfibrillar material (/) in the vicinityof the Golgi apparatus, x 40000.Fig. 48. Plasma membrane of a notochordal cell in 10-day-old embryo during for-mation of the basal lamina by excortication (ecdysis). Note that the appearance ofthe fibrous material of the basal lamina is virtually identical to that of the fibrousmaterial underneath the cell membrane, x 30000.Fig. 49. Electron micrograph showing the difference between the ultrastructure of anotochordal cell cytoplasm (upper part of the micrograph) and that of a moredifferentiated electron-dense cell, x 16000.
24 A. JURAND
52

Development of mouse notochord 25
some endodermal cells take part in the histogenesis of the notochord when itseparates from the archenteron.
Also, it has been stated that in lower chordates (particularly in Amphioxus) thenotochord is of endodermal origin, while in birds and mammals it is of ecto-dermal origin (Walter, 1928; Shumway, 1942). According to Huettner (1941)the notochord is of ectodermal origin in Amphioxus and of mesodermal origin inAmphibia. Such ideas would mean that notochords in different chordates arenot homologous organs.
To avoid this confusion the blastopore and Hensen's node should be regardedas undifferentiated pluripotential centres. The notochord proliferates from thesecentres in the same or closely adjacent area as that which gives rise to themesoderm. Hence the notochord is mesodermal in nature but originates fromectodermal cells in the blastopore lip or Hensen's node. However, any distinctionbetween ectoderm and mesoderm in the area of the primitive streak is meaning-less, as here the mesoderm is produced by the ectoderm (Huber, 1918; Griine-berg, 1963).
The mesodermal nature of the notochord is reflected in its intermediateposition between the ectoderm and endoderm, and in its later endoskeletalfunction, like that of the mesodermal skeletons in all vertebrates. This was, infact, the opinion of Kingsbury (1920), and the only amendment to this opinionis that some endodermal cells may contribute to notochord histogenesis bybeing retained with the presumptive notochordal cells when the notochordseparates from the endoderm of the archenteron roof (Rand, 1950).
This view is confirmed by the observation that the prenotochordal cells of thechordal plate show ultrastructural differences in comparison with endodermalcells. The cytoplasm of the prenotochordal cells is less electron-dense, and thecells are less differentiated than endodermal cells. Later, even after separationof the notochord, these two types of cell are occasionally found side by side,but fewer cells have a less electron-translucent cytoplasm. It is possible, therefore,that these cells are derived from endoderm cells retained with prenotochordalcells at the time of separation.
In the mouse embryo Hensen's node is not prominent (Sobotta, 1911). In
FIGURES 50-52
Fig. 50. A zonula adherens between two notochordal cells, not at the outer surfaceof the notochord, in a 10-day-old embryo, x 33000.Fig. 51. The area of adhesion between the notochord (on the left) and the ventral sideof the neural tube (on the right) is shown. Note between the two organs singleelectron-dense plates about 45 nm thick, formed by their mutual basal laminae.This structure is shown diagrammatically in Figs. 43 and 44. x 21000.Fig. 52. A mesenchymal cell (me) lies between the neural tube (nt) and the noto-chord (n). Note that both the neural tube and the notochord are covered by basallaminae but the mesenchyme cell is not; lO^-day-old embryo, x 14000.


Development of mouse notochord 27
guinea-pig it is more easily noticeable and has been referred to as Hensen's orthe primitive node (Huber, 1918). In our material it appears as a small mid-sagittal depression in the caudal area of the presomitic embryonic shield. Inlater stages, in embryos with 1-3 somites, chorda plate in this restricted area iscontinuous with the bottom of the neural groove (Fig. 3; see also Green, 1966).This continuity probably represents the last trace of Hensen's node.
The separation of the mouse notochord from the archenteron roof takesplace as described by Huber (1918) for guinea-pig embryos. Firstly, the chordaplate is undergrown by a continuous one-cell-thick endodermal layer, and at thesame time, the chorda plate folds dorsally to form an inverted trough whichwill give rise to the residual chorda canal. In early mouse embryos, however, thechorda canal is an intercellular space, only 1-2 jam in diameter; later it is possibleto detect it as a discontinuous canal.
The light-microscopic changes in shape and dimensions during the first daysof development reported here agree with those published by Dawes (1930), ifhis crown-rump lengths are converted to age by using the tables of Griineberg(1943). Dawes (1930) also described, in embryos 4 mm long (i.e. 10- to 10|-day-old), segmental flexures (waves) of the mouse notochord (except in the cranio-cervical and occipital regions) which persist until the intervertebral dilatationsand intravertebral constrictions appear in embryos 8 mm long (i.e. 13-day-old).These flexures were originally described in eight other mammals by Minot (1907).There is some disagreement between these two authors, because Minot saw thedorsal bends opposite the future intervertebral discs and Dawes claimed that inmice the situation is reversed. However, Minot did notice in pig embryos thatthe bends in the cervical segments shift cranially towards the levels of thevertebral centra.
Our material supports Dawes' statement; the ventral bends correspond to thediscs and the dorsal ones to the vertebral centra. Furthermore, the segmentalflexures appear only in 13- and 13^-day-old embryos-that is, 24-36 h afterdetachment of the notochord from the ventral surface of the neural tube; only
FIGURES 53-56
Fig. 53. Peripheral notochordal cell membrane in lH-day-old embryo, showingdiscontinuity of the basal lamina during excortication. Note intracytoplasmicfibrillar material (/) consisting of microfibrils 5 nm in diameter, x 45000.Fig. 54. Continuity between the intracytoplasmic fibrillar material and the fibrousextensions outside the notochord as the perichordal sheath forms. Note at the pointsof excortication (ecdysis) the cell membrane is discontinuous. 14-day-old embryo,x 112000.Fig. 55. Part of the cytoplasm of a notochordal cell in 15-day-old embryo showingdilatations of the endoplasmic reticulum. x 21000.Fig. 56. Swollen mitochondria in 14-day-old notochordal cell. Note that Golgisaccules are unaffected, x 27000.

28 A. JURAND
10-15 such flexures are found in the lumbo-sacral region and they do not persistin 14-day-old embryos. They disappear shortly before this stage, when chondrifi-cation of the vertebral column begins, and are replaced by intervertebral dilata-tions and intravertebral constrictions.
Cell differentiation and morphogenesis of the mouse notochord follow thesame cranio-caudal gradient as described in the urodele notochord (Waddington& Perry, 1962). This phenomenon is evident in changes such as the separation ofthe notochord from the endoderm (Willis, 1958), its detachment from its transientadhesion to the neural tube, and the formation of the intervertebral dilatationswhich make it moniloform. Similarly, at the cellular level, vacuolization of thenotochordal cells follows a cranio-caudal gradient. It is noticeable that the con-densations of cartilage forming the vetebrae also follow the same gradient. Thisis possible because the chondrogenesis-inducing capacity of the notochorddepends on its state of cytodifferentiation.
The cells of Hensen's node appear undifferentiated, in that their cytoplasm iselectron-translucent, ribosomes are scarce and endoplasmic reticular cisternaeare being actively formed. The occasional Golgi apparatus resembles thosedescribed in other embryonic cells (Ward, 1965; Ward & Ward, 1968; Morre,Mollenhauer & Bracker, 1971).
Another characteristic of the cells of Hensen's node and the head process isthe presence of microtubules distributed in the cytoplasm and associated withcentrioles. Microtubules may act as agents of cell elongation (Schroeder, 1971;Burnside, 1971; Karfunkel, 1972), or cell motility (Loewy & Siekievitz, 1970;Novikoff & Holtzman, 1970; De Robertis, Nowinski & Saez, 1970; Toner &Carr, 1971). It is known from carbon marking experiments that cells migratefrom Hensen's node towards the head process and, in doing so, they elongate.Thus the microtubules may be instrumental in cell elongation and they mightcontribute to cell motility.
The presence of centrioles in Hensen's node and early head process cells seemsto be connected with the presence of cilia, for which centrioles serve as basalbodies (kinetosomes). Cilia themselves are a characteristic of early embryoniccells such as those in the neural plate (Sotelo & Trujillo-Cenoz, 1959), in theneural tube (Duncan, 1957) or in the limb-bud apical ectodermal ridge (Jurand,1965).
In 9^-day-old embryos and in later stages, fibrillogenesis is a conspicuousfeature in the notochordal cell cytoplasm. Bundles of microfibrils are presentin the perinuclear cytoplasm and near the Golgi apparatus, as well as under theperipheral plasma membrane. Similar fibrous material is present in chick noto-chordal cells at various developmental stages (Jurand, 1962). The capacity of thenotochordal cells for fibrillogenesis was clearly demonstrated by Carlson (1973).
In early stages the microfibrillar material may form the basal lamina, and inlater stages it is used for the formation of the perichordal sheath. At both timesit is probably extruded from the cells by excortication (ecdysis or delamination)

Development of mouse notochord 29
through the plasma membrane. In 11-day-old embryos excortication of thickerfibrils disrupts the basal lamina and eventually removes it from the surface ofthe notochord. Excortication as an extrusion mechanism for fibrillar endoplasmicproducts was originally described in developing cartilage by Godman & Porter(1960).
Notochordal cells in mouse embryos aged 11 days show active endocytosisby the plasma membrane, which forms pinocytic coated invaginations, andsubsequently, coated vesicles which are present in the cytoplasm. These vesiclesare identical with those formed by pinocytotic activity in erythroblasts and endo-thelial cells, as described by Fawcett (1965). An analogous observation has beenmade in chick notochord, but there, in older embryos, it coincided with thebeginning of vacuolization of the notochordal cells (Jurand, 1962). In both ani-mals this process may be active because the notochord remains avascular, andso its water requirements can be met (Fox, 1973).
Frequent connexions of the endoplasmic reticular cisternae with the outernuclear membrane, particularly in early stages of cytodifferentiation, is a featureof mouse notochordal cells as it is in newts (Waddington & Perry, 1962) and inchick embryos (Jurand, 1962). Threadgold (1967) in his monograph confirmsthese observations.
Enclosure of the notochord by its own basal lamina during separation fromthe archenteron enables the notochord to become a separate anatomical entity.It probably protects the notochordal cells, helps to form a coherent organ andprevents the separated notochord from adhering to neighbouring organs inthe vicinity. The notochordal basal lamina precedes the perichordal sheath inits protective role. Also, the basal lamina of the notochord and of the neuraltube provides a transient anchorage for the notochord when it is adhering to theventral side of the neural tube. These two basal laminae form, at the points ofadhesion, cell junctions in the form of thicker and more electron-dense singleplates, common to both organs, which provide an attachment. These plates mayhelp the notochord to modify mechanically the morphogenesis of the neuraltube (Schroeder, 1971). In particular, this could explain the role of the notochordas an 'elongator' of the neural tube (Waddington & Perry, 1962).
The protective role of the notochordal basal lamina may be confirmed by itsabnormality or absence in mice with Danforth's short-tail semi-dominant gene(Sd) (see Griineberg, 1958a). Similarly, in short-tail or Brachyury {T/- and T/T)and in taillessness or Anury (T/t) it was postulated by Gruneberg (19586) thatthe notochord in these mutants has abnormal surface properties. As the noto-chord does not separate readily in these cases after its normal transient adhesionto the neural tube, this might be due to abnormality of the basal lamina, alteringthe surface properties of the notochord. It would be interesting to investigatethe relevant stages for the quality of basal lamina, the presence of microfibrils inthe notochordal cells, and also the frequency of junctional complexes betweennotochordal cells.

30 A. JURAND
The terminology of the basal lamina has been reviewed by Goel & Jurand(1969). In the case of the notochord, the term 'basal lamina' could be replacedby the term'external lamina' (Toner & Carr, 1971), since it is related essentiallyto a non-epithelial organ, but it is used because the notochord shows an epi-thelial arrangement of its cells (Williams, 1908; Leeson & Leeson, 1958). Infact, older terms such as ' Chordaepithel' (Gegenbaur, 1864) and 'epithelio-morphe Schicht' (Grassi, 1883) have been used to describe a non-vacuolatedouter cell layer of the notochord around a vacuolated core.
Waddington & Perry (1962) have shown the significance of the generaladhesiveness of the notochord cells during early morphogenesis. Unlike thesituations in urodele and chick notochords, in the early (9- and 10-day-old)mouse notochord the notochordal cells show infrequent and incomplete inter-cellular junctional complexes which are not always near the surface of the organ.Shortly after separation, however, the cells adhere to each other more firmly,and cause the initially flattened notochord to become an almost perfect cylinder.It is probable that, in the mouse notochord, both the general adherence of thecells and their junctional complexes play a role in morphogenesis.
The older view, based on light microscopy, that the notochord is initiallycellular, then becomes syncytial and finally becomes cellular again (Williams,1908; Dawes, 1930) cannot be maintained. Electron microscopy gives no evi-dence of a syncytial stage in mouse embryos of the present study nor in rabbit(Leeson & Leeson, 1958), amphibian (Waddington & Perry, 1962) or chickembryos (Jurand, 1962).
The author wishes to thank Professor Hans Griineberg, F.R.S., University College London,for discussing the subject and reading the manuscript and Miss Helen Tait for technicalassistance in the histological part of the work.
Separate gratitude is to be expressed to our artist Mr E. D. Roberts for the skilful andelaborate reconstructions drawn from histological preparations and for mounting themicrographs. Thanks are also due to Mr R. K. Barrowdale and Mr Frank M. Johnston forthe photographic work.
This work was supported by a grant from the Distillers' Company to the University ofEdinburgh.
REFERENCES
BELLAIRS, R. (1963). The development of somites in the chick embryo. J. Embryo!, exp. Morph.11, 697-714.
BOENIG, H. (1950). Leitfaden der Entwicklungsgeschichte des Menschen. Leipzig: Georg Thieme.BRUNr, A. (1912). Uber die evolutiven und involutiven Vorgange der Chorda dorsalis in der
Wirbelsaule mit besonderer Buriicksichtigung der Amnioten. Anat. Hefte 45, 308-470.BURNSIDE, B. (1971). Microtubules and microfilaments in newt neurulation. Devi Biol. 26,
416-441.CARLIER, K. W. (1890). The fate of the notochord and development of the intervertebral
disc in the sheep with observations on the adult disc in these animals. /. Anat. 24, 578-585.CARLSON, E. C. (1973). Periodic fibrillar material in membrane-bound bodies in notochordal
epithelium of the early chick embryo. / . Ultrastruct. Res. 42, 287-297.CERFONTAINE, P. (1906). Recherches sur le developpement de PAmphioxus. Arc/is Biol., Paris
22, 229-418.

Development of mouse notochord 31CLARA, M. (1955). Entwickhwgsgeschichte des Menschen. Leipzig: Georg Thieme.COOPER, G. W. (1965). Induction of somite chondrogenesis by cartilage and notochord. A
correlation between inductive activity and specific stages of cytodifferentiation. Devi BioL12, 185-212.
DAWES, B. (1930). The development of the vertebral column in mammals, as illustrated by itsdevelopment in Mus musculus. Phil. Trans. R. Soc. B 218, 115-170.
DE ROBERTIS, E. D. P., NOWINSKI, W. W. & SAEZ, F. A. (1970). Cell Biology. Philadelphia:W. P. Saunders.
DUNCAN, D. (1957). Electron microscope study of the embryonic neural tube and notochord.Tex. Rep. Biol. Med. 15, 367-377.
EBERT, J. D. & SUSSEX, I. M. (1970). Interacting Systems in Development. New York: Holt,Rinehart and Winston.
ELLISON, M. L., AMBROSE, E. J. & EASTY, G. C. (1969). Chondrogenesis in chick embryosomites //; vitro. J. Embryol. exp. Morph. 21, 331-340.
EYAL-GILADI, H. (1958). The notochord as inductor of the orohypophysis in Urodeles(Pleurodeles waltlii). Proc. K. ned. Akad. Wet. 61, 224-234.
FAWCETT, D. W. (1965). Surface specializations of absorbing cells. / . Histochem. Cytochem.13,75-91.
Fox, H. (1973). Degeneration of the tail notochord of Rana temporaria at metamorphicclimax. Examination by electron microscopy. Z. Zellforsch. mikrosk. Anat. 128, 371 —386.
GEGENBAUR, C. (1864). Untersuchungen zur vergleichenden Anatomie der Wirbelsdule beiAmphibien und Reptilien. Leipzig: W. Engelmann.
GODMAN, G. C. & PORTER, K. R. (1960). Chondrogenesis, studied with the electron micro-scope. /. biophys. biochem. Cytol. 8, 716-760.
GOEL, S. C. & JURAND, A. (1969). The structures at the epithelio-connective tissue junctions:A comparison of light and electron microscopic observations. Proc. R. Soc. Edinb. B 71,1-13.
GRASSF, G. B. (1883). Developpement de la colonne vertebrale chez les poissons osseuses.Archs ital. Biol. 4, 251-268.
GREEN, E. L. (1966). Biology of the Laboratory Mouse, 2nd edn. New York: McGraw-Hill.GRUNEBERG, H. (1943). The development of some external features in mouse embryos.
/ . Hered. 34, 88-92.GRUNEBERG, H. (1958a). Genetical studies on the skeleton of the mouse. XXII. The develop-
ment of the Danforth's short-tail. J. Embryol. exp. Morph. 6, 124-148.GRUNEBERG, H. (19586). Genetical studies on the skeleton of the mouse. XXIII. The develop-
ment of Brachyury and Anury. /. Embryol. exp. Morph. 6, 424-443.GRUNEBERG, H. (1963). The Pathology of Development. A study of Inherited Skeletal Disorders
in Animals. Oxford: Blackwell.HOLTZER, H. & DETWILER, S. (1953). An experimental analysis of the development of the
spinal column. III. Induction of skeletogenous cells. /. exp. Zool. 123, 335-370.HUBER, G. C. (1918). On the anlage and morphogenesis of the chorda dorsalis in Mammalia,
in particular in the guinea pig (Cavia cobaya). Anat. Rec. 14, 217-264.HUETTNER, A. F. (1941). Comparative Embryology of the Vertebrates. New York: Macmillan.JURAND, A. (1962). The development of the notochord in chick embryos. / . Embryol. exp.
Morph. 10, 602-621.JURAND, A. (1965). Ultrastructural aspects of early development of the forelimb buds in the
chick and the mouse. Proc. R. Soc. B 162, 387^05.JURAND, A. (1968). Mouse News Lett. 38, 20.JURAND, A. & IRELAND, M. J. (1965). A slow rotary shaker for embedding in viscious media.
Stain Technol. 40, 233-234.KARFUNKEL, P. (1972). The activity of microtubules and microfilaments in neurulation in the
chick. / . exp. Zool. 181, 289-302.KERR, J. G. (1919). Textbook of Embryology. Vol. II. Vertebrata with the exception of Mam-
malia. London: Macmillan.

32 A. JURAND
KINGSBURY, B. F. (J920). The developmental origin of the notochord. Science, N.Y. 51,190-193.
KITCHIN, I. C. (1949). The effects of notochordectomy in Amblystoma mexicanum. J. exp.Zool. 112, 393-415.
LEESON, T. S. & LEESON, C. R. (1958). Observations on the histochemistry and fine structureof the notochord in rabbit embryos. / . Anat. 92, 278-285.
LILLIE, R. D. (1951). The allochrome procedure. Am. J. din. Path. 21, 484-488.LOEWY, A. G. & SIEKIEVITZ, P. (1970). Cell Structure and Function. New York: Holt, Rine-
hart and Winston.MINOT, C. S. (1907). The segmental flexures of the notochord. Anat. Rec. 1, 42-50.MORRE, D. J., MOLLENHAUER, H. H. & BRACKER, C. E. (1971). Origin and continuity of
Golgi apparatus. In Origin and Continuity of Cell Organelles (ed. J. Reinert & H. Ursprung),pp. 82-126. Berlin: Springer-Verlag.
NELSEN, O. E. (1953). Comparative Embryology of the Vertebrates. New York: Blakiston.NrAzr, I. A. (1969). Differentiation capacities of the prospective tail somite region of the
neural plate in the embryos of Ambystoma mexicanum. J. Embryol. exp. Morph. 22,1-14.
NiEuwKOOP, P. D. & UBBELS, G. A. (1972). The formation of the mesoderm in urodelanamphibians. IV. Qualitative evidence for the purely 'ectodermal' origin of the entiremesoderm and of the pharyngeal endoderm. Wilhelm Roux Arch. EntwMech. Org. 169,185-199.
NOVIKOFF, A. B. & HOLTZMAN, E. (1970). Cells and Organelles. New York: Holt Rinehartand Winston.
PALADE, G. E. (1952). A study of fixation for electron microscopy. / . exp. Med. 95, 285-298.PATTEN, B. M. (1958). Foundations of Embryology. New York: McGraw-Hill.PLUMEL, M. (1948). Tampon au cacodylate de sodium. Bull. Soc. Chim. biol. 30, 129-130.RAND, H. W. (1950). The Chordates. London: H. K. Lewis.REYNOLDS, E. S. (1963). The use of lead citrate at high pH as an electron-opaque stain in
electron microscopy. / . Cell Biol. 17, 208-213.ROMER, A. S. (1949). The Vertebrate Body. Philadelphia: W. B. Saunders.RUGH, R. (1968). The Mouse, Its Reproduction and Development. Minneapolis, Minn.:
Burgess.SAUNDERS, J. W. Jr. (1968). Animal Morphogenesis. New York: Macmillan.SCHROEDER, T. E. (1971). Mechanism of morphogenesis: The embryonic neural tube. Int. J.
Neurosci. 2, 183-198.SELENKA, E. (1883). Studien iiber Entwicklungsgeschichte der Tiere. I. Heft. Keimblatter und
Primitivorgane der Maus. Wiesbaden: Kreidel.SHUMWAY, W. (1942). Introduction to Vertebrate Embryology. New York: John Wiley.SMITHBERG, M. (1954). The origin and development of the tail in the frog Rana pipiens.
J. exp. Zool. 127, 397-426.SOBOTTA, J. (1911). Die Entwicklung des Eies der Maus von ersten Auftreten des Mesoderms
an bis zur Ausbildung der Embryonalanlage und dem Auftreten der Allantois. I. Teil. DieKeimblase. Arch, mikrosk. Anat. EntwMech. 78, 271-352.
SOTELO, J. R. & TRUJILLO-CENOZ, O. (1958). Electron microscope study on the developmentof ciliary components of the neural epithelium of the chick. Z. Zellforsch. mikrosk. Anat.49, 1-12.
THEILER, K. (1950). Die Auswirkung von partiellen Chordadefekten bei Triton alpestris.Beitrag zur Entwicklungsmechanik der Wirbelsaule. Willhelm Roux Arch. EntwMech. Org.144, 476-490.
THEILER, K. (1972). The House Mouse. Development and Normal Stages from Fertilization to4 weeks of Age. Berlin: Springer-Verlag.
THREADGOLD, L. T. (1967). The Ultrastructure of the Animal Cell. Oxford: Pergamon Press.TONER, P. G. &CARR, K. E. (1971). Cell Structure. Edinburgh, London: Churchill Livingstone.WADDINGTON, C. H. (1952). The Epigenetics of Birds. London: Cambridge University Press.WADDINGTON, C. H. (1956). Principles of Embryology. London: George Allen & Unwin.WADDINGTON, C. H. & PERRY, M. M. (1962). The ultrastructure of the developing urodele
notochord. Proc. R. Soc. B 156, 459-482.

Development of mouse notochord 33WALTER, H. E. (1928). Biology of the Vertebrates. New York: Macmillan.WARD, R. T. (1965). Formation of Golgi bodies during maturation of oocytes in Rana
pipiens. Anat. Rec. 151, 430.WARD, R. T. & WARD, E. (1968). The multiplication of Golgi bodies in the oocytes of Rana
pipiens. J. Microscopie 7, 1007-1020.WATTERSON, R. L., FOWLER, I. & FOWLER, B. J. (1954). The role of the neural tube and noto-
chord in development of the axial skeleton of the chick. Am. J. Anat. 95, 337-382.WEISS, P. (1939). Principles of Development. A Text in Experimental Embryology. New York:
Henry Holt.WILLIAMS, L. W. (1908). The later development of the notochord in mammals. Am. J. Anat. 8,
251-284.WILLIS, R. A. (1958). The Borderline of Embryology and Pathology. London: Butterworth.YAMADA, T. (1940). Beinflussung der Differenzierungsleistung des isolierten Mesoderms von
Molchkeimen durch zugefiigtes Chorda- und Neuralmaterial. Folia anat. jap. 19, 131-196.ZILLIKEN, F. (1967). Notochord induced cartilage formation in chick somites. Intact tissue
versus extracts. Exp. Biol. Med. 1, 199-212.
(Received 2 July 1973, revised 4 March 1974)
E M B 32